Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
Gut Microbiota Regulates Depression-Like Behavior in Rats Through the Neuroendocrine-Immune-Mitochondrial Pathway
Authors Liu S, Guo R, Liu F, Yuan Q, Yu Y ![]() , Ren F
, Ren F
Received 27 December 2019
Accepted for publication 1 March 2020
Published 31 March 2020 Volume 2020:16 Pages 859—869
DOI https://doi.org/10.2147/NDT.S243551
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jun Chen
Shuhan Liu,1 Rongjuan Guo,2 Fei Liu,3 Qingjie Yuan,4 Yao Yu,1 Feifei Ren1
1Second Clinical Medical College, Beijing University of Chinese Medicine, Beijing 100029, People’s Republic of China; 2Department of Neurology, Dongfang Hospital Beijing University of Chinese Medicine, Beijing 100078, People’s Republic of China; 3College of Acupuncture and Massage, Changchun University of Chinese Medicine, Changchun 130117, People’s Republic of China; 4Department of Traditional Chinese Medicine, Tsinghua University Yuquan Hospital, Beijing 100040, People’s Republic of China
Correspondence: Rongjuan Guo
Department of Neurology, Dongfang Hospital Beijing University of Chinese Medicine, No. 6 Fangxingyuan 1st Block, Fengtai District, Beijing City, People’s Republic of China
Tel +86 1 861 137 7633
Fax +86 1064286498
Email [email protected]
Purpose: Gut microbiota affects various physiological functions in the host and has crucial effects on the nervous system. There is increasing evidence of a correlation between gut microbiota and depression; however, the mechanisms underlying the regulation of depression-like behavior by gut microbiota remain unclear. In this study, we assessed the regulatory mechanism of gut microbiota on depression-like behavior in rats.
Methods: We transplanted fecal microbiota obtained from patients with depression and healthy individuals into germ-free (GF) rats (n=18) through fecal microbiota transplantation technology. Next, we assessed the affective behavior in the rats using the forced swimming test and a sucrose preference test. We used enzyme-linked immunosorbent assay (ELISA) to determine the hippocampal levels of 5-hydroxytryptamine (5-HT), dopamine (DA), and noradrenaline (NE) and the serum levels of corticosterone (CORT), adrenocorticotropic hormone (ACTH), corticotropin-releasing hormone (CRH), tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), interleukin-6 (IL-6), interleukin-1 (IL-1), interleukin-1 (IL-4), and interleukin-1 (IL-10). The mitochondrial morphology of small intestinal epithelial cells was observed through transmission electron microscopy.
Results: Rats that received fecal microbiota from patients with depression (depression microbiota) exhibited depression-like behavior. They presented decreased levels of hippocampal neurotransmitters, serum CORT levels, and anti-inflammatory cytokine levels, as well as increased ACTH, CRH, and serum levels of multiple pro-inflammatory cytokines. Observation of the mitochondria ultrastructure showed damaged mitochondria in the intestinal epithelial cells, significant endoplasmic reticulum expansion, and border aggregation of nuclear chromatin.
Conclusion: Our findings suggested that the depression-like behaviors induced by the depression microbiota through the neuroendocrine-immune-mitochondrial pathway, which were associated with neuroendocrine disorders, inflammatory responses, and mitochondrial damage.
Keywords: gut microbiota, depression, neurotransmitter, HPA axis, immune, mitochondrial
Introduction
A study on the worldwide disease burden reported that depression is the fifth leading cause of death and that it accounted for 40.5% of the disability rate of mental illness.1 The hypothesis on the complex network relationship between the gut microbiota and the host suggests a close association of depression with gut microbiota. The dysfunction of the brain-gut-microorganism axis is the main pathological basis of depression and could directly induce and influence depression factors. Gut microbiota regulation could be an effective method for depression prevention and treatment.2
There is a symbiotic relationship between humans and microorganisms. There are approximately 1014 microorganisms colonizing the gastrointestinal tract, which is 10 times the number of human cells, and most of them are bacteria.3 Gut microbiota exert a prominent regulatory effect on host brain development and behavior.4–6 Previous pathological and physiological studies on patients with functional depression have reported that the depression state in these patients is closely related to gut microbiota imbalance.7
Clinical studies have reported a significant reduction in the diversity and abundance of gut microbiota in patients with depression and that it is correlated with symptom severity.8 Probiotics can significantly reduce depression and anxiety and improve cognition and metabolism.9–11 Animal studies often use chronic stress to induce depression in experimental animals. Stress not only affects psychology and stress system but also destroys gut microbiota.12,13 Bifidobacteria can alleviate depression symptoms in rats through gut microbiota regulation.
Recent studies have reported a complex network relationship between the brain and gut microbiota, which is closely related to depression. Consequently, a new conceptual model of the “brain-gut-microbial axis” has been proposed.14–16 This model is defined as a two-way regulatory pathway that integrates the central nervous system, neuroendocrine system, neuroimmune system, autonomic nervous system, intestinal nervous system, and gut microbiota. Moreover, it plays an essential role in the assessment of cognitive mental diseases.17 Many studies have reported that patients with gastrointestinal disorders, including irritable bowel syndrome and inflammatory bowel disease, have accompanying emotional disorders, indicating the importance of the gut-brain axis in emotional disorder pathophysiology. Further, GF animals have become a new animal model type established through aseptic reproduction and aseptic feeding. They lack any microorganisms in their body and allow assessment of the effect of gut microbiota on the host. GF animals are usually kept in a 100-level purified environment since birth. Their drinking water, litter, feed, and all items in direct contact with the animals are strictly sterilized before use and are regularly tested, which effectively prevents post-birth colonization of gut microbiota.18,19 GF animals have gained increasing attention since they can be directly used to assess the effect of gut microbiota on various physiological aspects. A previous study transplanted fecal microbiota from patients with depression into GF mice and reported that they exhibited depression-like behavior, which suggests that specific microbiota could induce depression symptoms.20 However, the mechanism through which gut microbiota induces depression remains unclear.
In this study, we transplanted the gut microbiota from patients with depression into GF rats. Subsequently, we assessed changes in the behavior, hippocampal neurotransmitter levels, Hypothalamic-Pituitary-Adrenocortical (HPA) axis, inflammatory cytokine levels, and mitochondrial structure in the rats to explore the underlying mechanisms by which the gut microbiota influences host behavior.
Materials and Methods
Animals and Group Divisions
We obtained eight-week-old male GF rats (n = 18) were provided by the Institute of Experimental Animals, the Chinese Academy of Medical Sciences (Beijing, China). We randomly divided the rats into three groups of 6 rats each; namely, the depression microbiota group, the healthy microbiota group, and the blank control group. All conditions were kept sterile and were verified that they met the Chinese Laboratory Animal Microbiological Standards and Monitoring (GB 14922.2–2011) by testing the feces and skin of the GF rats. The rats were fed in an aseptic flexible-plastic isolator and housed one per cage under the following conditions: a 12 h light/dark cycle (lights on from 07:30–19:30), constant temperature of 21–22 °C, and relative humidity of 50 ± 5%. All the rats received the same formula of autoclaved standard rat feed and water ad libitum. This study was approved by the Ethics Committee of the Dongfang Hospital Beijing University of Chinese Medicine and conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 80–23).
Fecal Microbiota Transplantation (FMT)
According to the inclusion criteria and exclusion criteria of depression and healthy males, we obtained fecal samples from male patients with severe clinical depression and healthy males and used them to establish the depression microbiota and healthy microbiota groups, respectively. Inclusion criteria of depression patients: DSM-5 as a diagnostic standard of major depressive episode in all patients with depression; Hamilton Depression Scale (HAMD) evaluation: total score of 24 items of HAMD > 24 points; no antidepressants were taken in the first episode of depression; no antibiotics or prebiotics were taken; the age is between 18 and 60. Exclusion criteria of depression patients: limited or previous severe mental illness; with history of severe brain injury and brain organic diseases; with heart, liver, kidney diseases, diabetes disease or other serious physical disease; routine laboratory examination (urine routine, liver function, blood routine) was abnormal. Inclusion of healthy controls: no organic disease or mental disease; routine laboratory tests (blood, urine, feces, liver function, blood lipid) were normal; no history of drug abuse; no antibiotics or prebiotics. The feces were processed to form a fecal microbiota solution in a sterile environment. The researcher transferred each rat to a special cage and infused fecal microbiota solution into each rat’s stomach by gavage. According to the dosage of human fecal microbiota solution (1.67g/kg/d) and the average rat weight of 200 g, each rat was infused with a 1.5 mL fresh microbiota solution once a day for 2 weeks and was observed for 30 mins after gavage administration.21
The rats were placed back into the original cage after observing that there was no abnormality. Fresh fecal microbiota solution was introduced into the various isolators through an aseptic transfer window. Further, all colonization operations were carried out in the isolators to ensure strict asepsis during implementation. Other conditions were similar for rats in both groups. After 4 weeks, we conducted behavioral tests and collected tissue samples.
Forced Swimming Test
The rats were successively placed in a plexiglass cylinder (50 cm, height; 30 cm, diameter) filled with water (25 ± 1 °C) to 35 cm. The test lasted for 6 min and we recorded the time spent by the rat in an immobile state in the last 5 min. The criterion for immobility is that the rat remains motionless and afloat or makes slight movements to maintain its head above the water. Subsequently, the recorded immobility time was used to evaluate the depression-like behavior of each rat.
Sucrose Preference Test
We conducted this test at the end of 1, 2, 3 and 4 weeks before FMT. As previously described, all the rats were trained to adapt to drinking sucrose water and each rat cage had access to two bottles of 1% sucrose water.22 After adapting for one day, one of the two bottles of sucrose water was replaced with a bottle of pure water; subsequently, one day was allowed to pass for adaptation. The formal experiment began after 24 hrs of fasting. First, we recorded the volume of 1% sucrose water and pure water in the respective bottle as W1 and W2, respectively. After 1 hr, both bottles were removed from each cage and the remaining volume of sucrose water and pure water were recorded as W3 and W4, respectively. Subsequently, we calculated the saccharine preference index as follows: (W1 - W3)/(W1 + W2 - W3 - W4) × 100%. All procedures in this test were conducted in isolators under strictly aseptic conditions. We autoclaved the sucrose water and pure water and transferred them to each isolator through the corresponding aseptic transfer window.
Sampling and Index Detection
Orbital Blood
After anesthesia with 10% chloral hydrate, we collected blood samples from the rats’ orbits. The blood samples were stored in the blood collection tubes, left at room temperature for 30 mins, and centrifuged (3000 r/min, 10 min, 4 ° C) to obtain serum. The serum was divided into 1 mL clean EP tubes and stored in a −80 °C refrigerator for subsequent analysis. According to the manufacturer’s instructions, we used ELISA to determine the serum levels of TNF-α, IFN-γ, IL-6, IL-1, IL-4, IL-10, CORT, ACTH, and CRH in the three rat models. The ELISA kits are from American R & D Company, and the instrument is American Thermo Multisk Type 3 Microplate Reader.
Tissue Sampling
The rats were randomly sacrificed by neck displacement. The brain was quickly removed from the skull and the hippocampus rapidly peeled off on an ice plate and stored in a −80 °C refrigerator for cryopreservation. To measure the neurotransmitter levels, we retrieved the tissue from the refrigerator, accurately weighed it, and added normal saline at 9 times its volume based on the mass (g): volume (mL) proportion of 1:9. Next, we obtained tissue homogenate through centrifugation (3000 R/min, 4 °C, 10 min), extracted the supernatant, and analyzed the hippocampal levels of 5-HT, DA, and NE through ELISA according to the manufacturer’s instructions. Small intestine tissue was extracted on ice with a size of 1 mm × 1 mm × 3 mm. Thereafter, we post-fixed it with osmic acid after glutaraldehyde fixation. After subsequent gradient acetone dehydration, embedding, ultra-thin slice production, and staining, we observed changes in the mitochondria ultrastructure in small intestine epithelial cells under a transmission electron microscope.
Statistical Analysis
Statistical analysis was performed using SPSS version 16.0 (IBM-SPSS Inc. Chicago, IL, USA). The results were expressed as mean ± standard deviation. For normally distributed data with variance homogeneity, we adopted the single-factor analysis of variance and the least significant difference. Otherwise, the nonparametric test and Kruskal Wallis test are used. Statistically significant differences were determined at p < 0.05.
Results
Rats in the Depression Microbiota Group Exhibited Depression-Like Behaviors
We conducted behavioral assessments of the rats using the forced swimming test and the sucrose preference test. As shown in Figure 1A, the immobility time in the forced swimming test was significantly higher in the depression microbiota group (p < 0.001) than that in the blank control and healthy microbiota groups. Moreover, in the sucrose preference test (Figure 1B), the saccharine preference index curve in the depression microbiota group was significantly decreased and showed a downward trend (p < 0.001), changes in the saccharine preference index curves in the healthy microbiota and blank control groups were similar and relatively gentle. From the first week to the forth week, the saccharine preference index was significantly lower in the depression microbiota group (p< 0.001) than that in the blank control and healthy microbiota groups (Figure 1C). Contrastingly, As aforementioned, this indicated that depression microbiota induced depression-like behavior.
 |
Figure 1 Rats in the depression microbiota group exhibited depressive-like behaviors. (A) Immobility time in the three groups in the FST. ***p < 0.001 vs control, ###p < 0.001 vs healthy. (B) Saccharine preference index curves in the sucrose preference test. (C) Saccharine preference index in the three groups in the sucrose preference test. ***p < 0.001 vs control, ###p < 0.001 vs healthy. Data are represented as mean ± SEM. |
Depression Microbiota Reduces the Hippocampal Neurotransmitter Levels
To determine the effect of gut microbiota on hippocampal neurotransmitter levels, we measured the hippocampal levels of 5-HT, NE, and DA using ELISA and the results are presented in Figure 2. Compared with the healthy microbiota group, the 5-HT, NE, and DA levels were significantly reduced in both the depression microbiota and blank control groups (p < 0.01). Compared to the blank control group, the levels of 5-HT, DA, and NE were significantly reduced in the depression microbiota group (p < 0.05). These results indicated that the depression microbiota reduced the hippocampal neurotransmitter levels. Further, due to the lack of gut microbiota, the hippocampal neurotransmitter levels in the GF rats were low and were improved through healthy microbiota transplantation.
 |
Figure 2 Depression microbiota reduced the hippocampal neurotransmitter levels. The 5-HT, NE, and DA levels in the depression microbiota, healthy microbiota, and blank control groups are shown in the respective three sub-figures. (A) Regarding the 5-HT levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy, Δp < 0.05, vs Control. (B) Regarding the NE levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy, ΔΔp < 0.01, vs Control. (C) Regarding the DA levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy, Δp < 0.05, vs Control. Data are represented as mean ± SEM. |
Depression Microbiota Promotes HPA Axis Hyperfunction
To determine the effect of gut microbiota on the HPA axis hyperfunction, we measured the serum levels of CORT, ACTH, and CRH using ELISA and Figure 3 presents the results. Compared with the healthy microbiota group, there was a significant increase in the ACTH, CORT, and CRH levels in the depression microbiota group (p < 0.001). Further, compared with the blank control group, there was a significant increase in the CORT, ACTH, and CRH levels in the depression microbiota group (p < 0.001). These results indicated that depression microbiota could promote HPA axis hyperfunction.
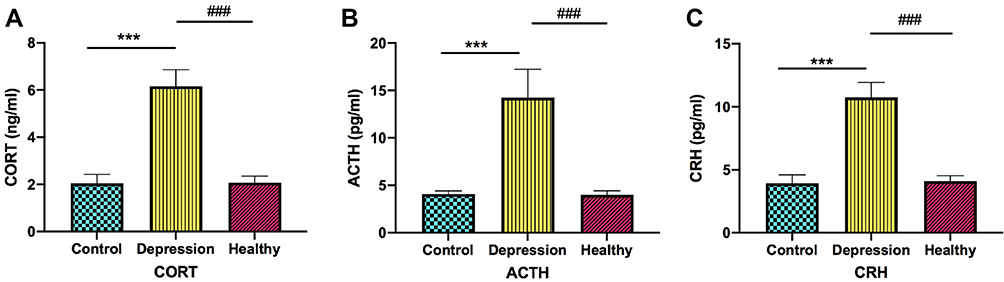 |
Figure 3 Depression microbiota promotes the HPA axis hyperfunction. The CORE, ACTH and CRH in the depression microbiota, healthy microbiota and blank control groups are shown in the respective three sub-figures. (A) Regarding the CORT levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (B) Regarding the ACTH levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (C) Regarding the CRH levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. Data are represented as mean ± SEM. |
Depression Microbiota Increases Serum Pro-Inflammatory Cytokine Levels and Decreases Anti-Inflammatory Cytokine Levels
To determine changes in serum levels of inflammatory cytokines induced by depression microbiota, we measured the serum levels of pro-inflammatory cytokines (TNF-α, IL-6, IL-1 and IFN-γ) and anti-inflammatory cytokines (IL-4 and IL-10) using ELISA. Compared with the blank control and healthy microbiota groups, there was a significant increase in the levels of TNF-α, IL-6, IL-1 and IFN-γ in the depression microbiota group (p < 0.001), as shown in Figure 4A–D. Further, there was a significant decrease in the IL-4 and IL-10 levels in the depression microbiota group (p < 0.001) in Figure 4E–F.
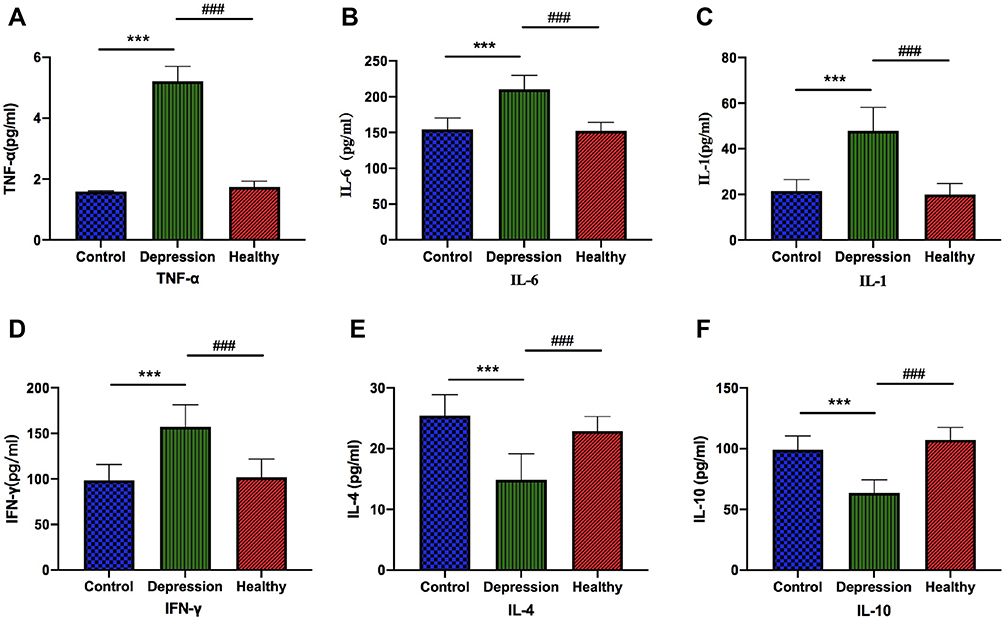 |
Figure 4 Depression microbiota increases serum pro-inflammatory cytokines and decreases anti-inflammatory cytokines. The TNF-α, IFN-γ, IL-6, IL-1, IL-4 and IL-10 in the depression microbiota, healthy microbiota and blank control groups are shown in the sub-figures. (A) Regarding the TNF-α levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (B) Regarding the IL-6 levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (C) Regarding the IL-1 levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (D) Regarding the IFN-γ levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (E) Regarding the IL-4 levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. (F) Regarding the IL-10 levels, ***p < 0.001, vs Control, ###p < 0.001, vs Healthy. Data are represented as mean ± SEM. |
Depression Microbiota Induces Mitochondrial Ultrastructural Damage in Small Intestinal Epithelial Cells
We observed the mitochondria ultrastructure in small intestine epithelial cells using the transmission electron microscope (Figure 5). We found that many mitochondria in the depression microbiota group were oval or round whole others were swollen, sparse, and even vacuolated. Numerous small vacuoles were observed in the cytoplasm and the nucleus chromatin exhibited the edge-set phenomenon. In the blank control group, the mitochondria of small intestinal epithelial cells were oval with a large number, dense distribution, intact outer membrane, clear internal cristae, uniform matrix density, and rich endoplasmic reticulum. In the healthy microbiota group, the basic morphology of mitochondria was similar to that of the blank control group; moreover, lysosomes were observed.
 |
Figure 5 Depression microbiota induces mitochondrial ultrastructural damage in small intestinal epithelial cells. Mitochondria ultrastructure, swelling, and vacuolation were observed in the depression group. |
Discussion
Depression involves interacting between social, psychological, and biological factors; further, it has complicated pathogenesis involving many systems and interactions, including neuroendocrine-immune-mitochondrial pathway. Neuroendocrine studies on depression have focused on decreased neurotransmitter levels and hypothalamic-pituitary-adrenal dysfunction. The monoamine neurotransmitter hypothesis is the most widely studied and recognized hypothesis on depression pathogenesis. It suggests that the biological basis of depression involves decreased monoamine neurotransmitter levels or dysfunction in synaptic spaces. The main neurotransmitters involved in the monoamine hypothesis include 5-HT, NE and DA. Currently, most first-line antidepressants are based on the monoamine hypothesis. The HPA axis is involved in the stress response regulation. Long-term stress exposure to the body induces excessive secretion of corticotropin-releasing factor (CRF), which decreases the glucocorticoid receptor (GR) function. This leads to the failure of negative feedback regulation of the HPA axis and continuous hyperfunction. HPA axis hyperactivity is an important depression cause; therefore, CRF receptor antagonists or GR antagonists have antidepressant effects. Clinical studies have reported significantly higher levels of IL-1 β, TNF - α, and IL-6 in the peripheral blood and cerebrospinal fluid of patients with depression compared to those in healthy individuals. Animal studies have shown that peripheral or central administration of IL-1 β, IL-6, and lipopolysaccharides could induce depression-like behaviors. Depression drugs can improve inflammation-induced depression-like behavior. Blocking cytokine production or removing their receptors could also have an antidepressant effect.23–25 Mitochondria play an important role in maintaining the normal function of cells. Abnormal mitochondria can lead to cell dysfunction, which leads to a series of functional disorders of the body and shows a variety of disease states. The results showed that the lifetime prevalence of depression in patients with mitochondrial diseases was 54%, significantly higher than that in the general population. Compared with patients with only mitochondrial diseases, patients with comorbid mental diseases are older, have higher hospitalization rate, worse physical condition and worse quality of life.26
In this study, we used the FMT method to transplant depression and healthy microbiota into the intestinal tract of GF rats to assess the effects of gut microbiota on behavior, neuroendocrine system, immune system, and mitochondria in rats. In the forced swimming test and sucrose preference test, compared with the healthy microbiota and blank control groups, the depression microbiota group showed a significant increase in the immobility time and a decrease in the saccharine preference index. This suggests that depression microbiota could induce depression-like behavior in rats. Further, it shows an alteration in the gut microbiota of patients with depression, which could be emulated through FMT of the gut microbiota to GF rats.
Neurotransmitters are messengers in the communication between nerve cells that have a crucial role in an individual’s psychology and behavior. Depression is closely related to neurotransmitter imbalance.27,28 The hypothesis of monoamine neurotransmitter deficiency suggests that positive emotions, eg, happiness, are strongly related to monoamine neurotransmitters. Depression is caused by reduced monoamine neurotransmitters in the brain and increasing these neurotransmitters could have an antidepressant effect.29 As a vital monoamine neurotransmitter in the brain, 5-HT is mainly involved in adjusting emotion, body temperature, food intake, cognition, sleep, and other functions.30–32 The levels of 5-HT and its receptor have been reported to be decreased in the brain and cerebrospinal fluid of suicide patients with depression; moreover, a positive correlation of decreased 5-HT levels with symptom severity has been reported.33 DA is not only a catecholamine transmitter but also the most critical transmitter in the reward system. It is mainly involved in the regulation of emotion, exercise, food intake, and weight.34 In the brain of depression animal models, there is dopamine neurotransmitter exhaustion and administration of dopamine antagonists and agonists worsens and improves the symptoms, respectively.35,36 The NE function is controlling attention by reflecting external stimuli and NE system dysfunction could cause depression. Patients with depression mainly have central nervous system lesions in the hippocampus, prefrontal cortex, striatum, etc.37 As an essential part of the human and other vertebrate brains, the hippocampus is involved in important functions, including learning, memory, and spatial localization. Studies have reported that stress can induce disorders in the hippocampal 5-HT, DA, and other neurotransmitters disorders in depression rat models.38 Moreover, severe depression can be induced by childhood stress with reduced hippocampal volume.39 In this study, we found decreased 5-HT, NE, and DA levels in the blank control and depression microbiota groups compared to those in the healthy microbiota group. This suggests that gut microbiota could affect hippocampal neurotransmitter levels and that lack of intestinal microorganisms reduces hippocampal neurotransmitter levels. Further, the findings showed that gut microbiota induces depression-like behavior in rats by decreasing hippocampal neurotransmitter levels.
HPA axis hyperactivity is a hypothesis that has been accepted in depression research, which mainly manifests as excessive CORT secretion.40 It was found that > 50% of patients with severe depression have maladjusted negative feedback regulation of the HPA axis with progressively increasing circulating CORT and ACTH levels; further, many of the patients even develop hypercortisonemia.41,42 Animal studies have reported excessive CORT and ACTH secretion by the HPA axis of GF rats under restraint stress, which is greatly improved by transplanting normal gut microbiota.43 In this study, we found significantly increased serum levels of CORT, ACTH, and CRH in the depression microbiota group compared to those in the healthy microbiota and blank control groups. This suggests that gut microbiota could affect the endocrine system and further induce depression-like behavior in rats by causing abnormalities in the HPA axis.
Inflammation is among the main pathological characteristics of depression. Patients with depression tend to develop immune disorders and chronic inflammation.44 Inflammatory cytokines are involved in the inflammatory response and could be either pro- or anti-inflammatory cytokines depending on their roles. According to the cytokine hypothesis, the pro-inflammatory cytokine level is negatively associated with the anti-inflammatory cytokine level in patients with depression; consequently, the overall immune response is directed towards inflammation.45 Clinical studies have reported significantly increased levels of IL-1, TNF-α, and other cytokines in patients with depression compared to those in healthy people. Further, application of antidepressants has been shown to decrease the expression of central pro-inflammatory factors, including IL-1β, IL-6, IFN-γ, TNF-α and increase the expression of anti-inflammatory factors, including IL-4, IL-8, and IL-10.46,47 Gut microbiota are essential for immune development in the host, which directly affects the role of peripheral immune cells in the central nervous system.48 High levels of pro-inflammatory cytokines contribute toward inhibition of the negative feedback function of the HPA axis, improving blood-brain barrier permeability, reducing 5-HT synthesis, and Glu energy system disturbance, which ultimately leads to depression.49,50 In this study, compared to the healthy microbiota and blank control groups, the depression microbiota group showed a significant increase in serum pro-inflammatory cytokines, including TNF-α, IFN-γ, IL-6, and IL-1, as well as a significant decrease in the serum anti-inflammatory cytokines, including IL-4 and IL-10. This suggests that gut microbiota could lead to an inflammation response, which could be among the factors inducing depression-like behavior.
The mitochondrion is the “power plant” of life activities in the cells and is widely found in all eukaryotic cells. Energy metabolism in mitochondria is critical for maintaining normal cell function. Mitochondrial damage often results in multi-organ and multi-system lesions, especially in organs with a high metabolism and energy demand. It is highly associated with depression characteristics regarding both mental symptoms and multi-system physical symptoms. In this study, we found that the mitochondria cristae of small intestinal epithelial cells in the depression microbiota group were swollen, sparse, and even vacuolated. There was obvious endoplasmic reticulum dilation and numerous small vacuoles in the cytoplasm. The nuclear chromatin exhibited the edge-set phenomenon. This could be attributed to mitochondrial functional damage, insufficient energy, excessive water accumulation in organelles, and inflammatory stimulation caused by depression microbiota. This suggests that depression microbiota can induce depression-like behavior in rats by impairing mitochondrial morphology and function.
In this study, we proved that depression microbiota could induce depression-like behavior in rats. By detecting the indicators of the neuroendocrine-immune-mitochondrial pathway in rats, we found that depression microbiota could result in the low levels of hippocampal neurotransmitters, the HPA axis hyperfunction, the Inflammation and the damaged structure of mitochondria.
There are several limitations in our study. First, we did not carry out the evaluation test of the effect of fecal microbiota transplantation on the gut microbiota of experimental rats, in order to further explore the correlation between the changes of gut microbiota and the generation of depression-like behavior in rats. Therefore, we will further verify it in the future research. Second, we did not evaluate the effect of antibiotic interference on gut microbiota and depression-like behavior in rats, so we will set up antibiotic interference control group to evaluate the effect of antibiotics on gut microbiota and behavior in future research.
Conclusion
In this study, we found that depression microbiota leads to depression-like behavior in rats determined through the FST and sucrose preference test. Assessment of indicators in the neuroendocrine-immune-mitochondrial pathway showed that depression microbiota significantly decreased hippocampal neurotransmitter levels; further, it induced abnormalities in the HPA axis, an inflammatory reaction, and mitochondrial dysfunction. All these results emphasized the effect of intestinal microorganisms on the neuroendocrine-immune-mitochondrial pathway.
Data Sharing Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Ethics Approval and Consent to Participate
This study was conducted after obtaining approval of the Animal Care and Use Committee of Dongfang Hospital Beijing University of Chinese Medicine, and conformed to the National Institutes of Health Guide for Care and Use of Laboratory Animals.
Acknowledgment
This work was supported by the National Natural Science Fund of China (81874422). We offer special acknowledgement to technician Rongjuan Guo and Fei Liu for their selfless technical support, encouragement, and guidance during this work.
Author Contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
Rongjuan Guo reports grants from National Natural Science Foundation of China, during the conduct of the study. The authors report no other conflicts of interest in this work.
References
1. Kohler CA, Freitas TH, Maes M, et al. Peripheral cytokine and chemokine alterations in depression: a meta-analysis of 82 studies. Acta Psychiatr Scand. 2017;135:373–387. doi:10.1111/acps.12698
2. Yarandi SS, Peterson DA, Treisman GJ, Moran TH, Pasricha PJ. Modulatory effects of gut microbiota on the central nervous system: how gut could play a role in neuropsychiatric health and diseases. J Neurogastroenterol Motil. 2016;22:201–212. doi:10.5056/jnm15146
3. Lozupone CA, Stombaugh JI, Gordon JI, et al. Diversity, stability and resilience of the human gut microbiota. Nature. 2012;489:220–230. doi:10.1038/nature11550
4. Collins SM, Surette M, Bercik P. The interplay between the intestinal microbiota and the brain. Nat Rev Microbiol. 2012;10:735–742. doi:10.1038/nrmicro2876
5. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behavior. Nat Rev Neurosci. 2012;13:70M2. doi:10.1038/nrn3346
6. Foster JA, McVey Neufeld KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36:305–312. doi:10.1016/j.tins.2013.01.005
7. Petra AI, Panagiotidou S, Hatziagelaki E, Stewart JM, Conti P, Theoharides TC.. Gut- Microbiota- Brain axis and its effect on neuropsychiatric disorders with suspected immune dysregulation. Clin Ther. 2015;37:984–995. doi:10.1016/j.clinthera.2015.04.002
8. Jiang HY, Ling ZX, Zhang YH, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav Immun. 2015;48:186–194. doi:10.1016/j.bbi.2015.03.016
9. Akkasheh G, Kashani-poor Z, Tajabadi-ebranimi M, et al. Clinical and metabolic response to probiotic administration in patients with major depressive disorder: a randomized, double-blind, placebo-controlled trial. Nutrition. 2016;32:315–320. doi:10.1016/j.nut.2015.09.003
10. Wallace CJK, Milev R. The effects of probiotics on depressive symptoms in humans: a systematic review. Ann Gener Psychiat. 2017;16:14–24. doi:10.1186/s12991-017-0138-2
11. Pirbaglou M, Katz J, De Souza RJ, Stearns JC, Motamed M, Ritvo P. Probiotic supplementation can positively affect anxiety and depressive symptoms: a systematic review of randomized controlled trials. Nutri Res. 2016;36:889–898. doi:10.1016/j.nutres.2016.06.009
12. Galley JD, Nelson MC, Yu ZT, et al. Exposure to a social stressor disrupts the community structure of the colonic mucosa-associated microbiota. BMC Microbiol. 2014;14(1):189.
13. I A M, J E G, Ren T, et al. Microbiota alteration is associated with the development of stress-induced despair behavior. Sci Rep. 2017;7:43859. doi:10.1038/srep43859
14. Cryan JF, O’mahony SM. The microbiome-gut-brain axis: from bowel to behavior. Neurogastroenterol Motil. 2011;23:187–192. doi:10.1111/j.1365-2982.2010.01664.x
15. Rhee SH, Pothoulakis C, Mayer EA. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat Rev Gastroenterol Hepatol. 2009;6:306–314. doi:10.1111/j.1365-2982.2010.01664.x
16. Mayer EA. Gut feelings: the emerging biology of gut-brain communication. Nat Rev Neurosci. 2011;2:453–466. doi:10.1038/nrn3071
17. Bienenstock J, Collins S. 99th Dahlem conference on infection, inflammation and chronic inflammatory disorders: psychoneuroimmunology and the intestinal microbiota: clinical observations and basic mechanisms. Clin Exp Immunol. 2010;160:85–91. doi:10.1111/j.1365-2249.2010.04124.x
18. Gootenberg DB, Turnbaugh PJ. COMPANION ANIMALS SYMPOSIUM: humanized animal models of the microbiome. J Anim Sci. 2011;89:1531–1537. doi:10.2527/jas.2010-3371
19. Tlaskalovahogenova H, Stepankova R, Kozakova H, et al. The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: contribution of germfree and gnotobiotic animal models of human diseases. Cell Mol Immunol. 2011;8:110. doi:10.1038/cmi
20. Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21:786–796. doi:10.1038/mp.2016.44
21. Gough E, Shaikh H, Manges AR. Systematic review of intestinal microbiota transplantation (fecal bacteriotherapy) for recurrent clostridium difficile infection. Clin Infect Dis. 2011;53:994–1002. doi:10.1093/cid/cir632
22. Ye YL, Zhong K, Liu DD, et al. Huanglian-Jie-Du-Tang extract ameliorates depression-like behaviors through BDNF-TrkB-CREB pathway in rats with chronic unpredictable stress. Evid Based Complement Alternat. 2017;Med.2017:7903918. doi:10.1155/2017/7903918
23. Patel A. Review: the role of inflammation in depression. Psychiatr Danub. 2013;25(Suppl 2):S216–S223.
24. Noto C, Rizzo LB, Mansur RB, McIntyre RS, Maes M, Brietzke E. Targeting the inflammatory pathway as a therapeutic tool for major depression. Neuroimmunomodulation. 2014;21:131–139. doi:10.1159/000356549
25. Martin C, Tansey KE, Schalkwyk LC, Powell TR. The inflammatory cytokines: molecular biomarkers for major depressive disorder? Biomark Med. 2015;9:169–180. doi:10.2217/bmm.14.29
26. Fattal O, Link J, Quinn K, Cohen BH, Franco K. Psychiatric comorbidity in 36 adults with mitochondrial cytopathies. CNS Spectr. 2007;12:429–438. doi:10.1017/s1092852900015303
27. Hamon M, Blier P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog Neuropsychopharm Biol Psychiat. 2013;45:54–63. doi:10.1016/j.pnpbp.2013.04.009
28. Lener MS, Niciu MJ, Ballard ED, Park M, Park LT, Nugent AC, Zarate CA
29. Hirschfeld R. History and evolution of the monoamine hypothesis of depression. J Clin Psychiat. 2000;61:4–6.
30. Nichols DE, Nichols CD. Serotonin receptors. Chem Rev. 2008;108:1614–1641. doi:10.1021/cr078224o
31. Berger M, Gray JA, Roth BL. The expanded biology of serotonin. Annu Rev Med. 2009;60:355–366. doi:10.1146/annurev.med.60.042307.110802
32. Feijófde M, Bertoluci MC, Reis C. Serotonin and hypothalamic control of hunger: a review. Rev Assoc Med Bras. 2011;57:74–77. doi:10.1016/S0104-4230(11)70020-3
33. Daszuta A, Ban M
34. Yen HH, Roseberry AG. Decreased consumption of rewarding sucrose solutions after injection of melanocortins into the ventral tegmental area of rats. Psychopharmacology. 2015;232:285–294. doi:10.1007/s00213-014-3663-6
35. Rominger A, Cumming P, Brendel M, et al. Altered serotonin and dopamine transporter availabilities in brain of depressed patients upon treatment with escitalopram: a 131I β-CIT SPECT study. Eur Neuropsychopharmacol. 2015;25:873–881. doi:10.1016/j.euroneuro.2014.12.010
36. Zhong P, Liu Y, Hu Y, Wang T, Zhao YP, Liu QS. BDNF interacts with endocannabinoids to regulate cocaine-induced synaptic plasticity in mouse midbrain dopamine neurons. J Neurosci. 2015;35:4469–4481. doi:10.1523/JNEUROSCI.2924-14.2015
37. Maletic V, Robinson M, Oakes T, Iyengar S, Ball SG, Russell J. Neurobiology of depression: an integrated view of key findings. Int J Clin Pract. 2007;61:2030–2040. doi:10.1111/j.1742-1241.2007.01602.x
38. Arnsten AF. Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci. 2009;10:410–422. doi:10.1038/nrn2648
39. Frodl T, Reinhold E, Koutsouleris N, Reiser M, Meisenzahl EM. Interaction of childhood stress with hippocampus and prefrontal cortex volume reduction in major depression. J Psychiatr Res. 2010;44:799–807. doi:10.1016/j.jpsychires.2010.01.006
40. Stetler C, Miller GE. Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosom Med. 2011;73:114–126. doi:10.1097/PSY.0b013e31820ad12b
41. Rook GA, Lowry CA. The hygiene hypothesis and psychiatric disorders. Trends Immunol. 2008;29:150–158. doi:10.1016/j.it.2008.01.002
42. Barden N. Implication of the hypothalamic-pituitary-adrenal axis in the physiopathology of depression. J Psychiat Neurosci. 2004;29:185–193.
43. Ruddick JP, Evans AK, Nutt DJ, Lightman SL, Rook GA, Lowry CA. Tryptophan metabolism in the central nervous system: medical implications. Expert Rev Mol Med. 2006;8:1–27. doi:10.1017/S1462399406000068
44. Lindqvist D, Dhabhar FS, James SJ, et al. Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology. 2017;76:197–205. doi:10.1016/j.psyneuen
45. Schiepers OJ, Wichers MC, Maes M. Cytokines and major depression. Prog Neuropsychopharm Biol Psychiat. 2005;29:201–217. doi:10.1016/j.pnpbp.2004.11.003
46. Catena-dell’osso M, Bellantuono C, Consoli G, Baroni S, Rotella F, Marazziti D. Inflammatory and neurodegenerative pathways in depression: a new avenue for antidepressant development? Curr Med Chem. 2011;18:245–255. doi:10.2174/092986711794088353
47. Tsao CW, Lin YS, Chen CC, Bai CH, Wu SR. Cytokines and serotonin transporter in patients with major depression. Prog Neuro Psychopharmacol Biol Psychiatry. 2006;30:899–905. doi:10.1016/j.pnpbp.2006.01.029
48. Berer K, Krishnamoorthy G. Commensal gut flora and brain autoimmunity: a love or hate affair? Acta Neuropathol. 2012;123:639–651. doi:10.1007/s00401-012-0949-9
49. Maes M, Leonard BE, Myint AM, et al. The new “5-HT” hypothesis of depression: cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog Neuropsychopharm Biol Psychiat. 2011;35:702–721. doi:10.1016/j.pnpbp.2010.12.017
50. Haroon E, Miller AH. Inflammation effects on brain glutamate in depression: mechanistic considerations and treatment implications. Curr Top Behav Neurosci. 2017;31:173–198. doi:10.1007/7854_2016_40
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.