Back to Journals » Cancer Management and Research » Volume 14
GNPNAT1 Predicts Poor Prognosis and Cancer Development in Non-Small Cell Lung Cancer
Received 24 March 2022
Accepted for publication 3 August 2022
Published 10 August 2022 Volume 2022:14 Pages 2419—2428
DOI https://doi.org/10.2147/CMAR.S367857
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Yong Feng, Na Li, Yi Ren
Department of Thoracic Surgery, Shenyang Chest Hospital, the Tenth People’s Hospital of Shenyang, Shenyang, People’s Republic of China
Correspondence: Yi Ren, Shenyang Chest Hospital, the Tenth People’s Hospital of Shenyang, 11 Beihai Street, Shenyang, 110044, People’s Republic of China, Tel +8615942068710, Email [email protected]
Background: Glucosamine-phosphate N-acetyltransferase 1 (GNPNAT1) is a key enzyme in the biosynthetic pathway of uridine diphosphate-N-acetylglucosamine and is upregulated in multiple malignancies. However, its function in cancer biology remains unclear.
Methods: Using TCGA dataset, this study analysed GNPNAT1 expression in non-small cell lung cancer (NSCLC) and assessed the correlation between GNPNAT1 and NSCLC patient prognosis. MTT and transwell assays were performed to determine the effect of GNPNAT1 on the growth and metastatic ability of lung cancer cells. GNPNAT1 expression was detected using immunohistochemistry in 78 NSCLC patients, and we analysed the correlation among clinicopathological parameters, overall survival (OS) and GNPNAT1 levels. Transcription factors that potentially regulate GNPNAT1 were explored using database analysis. RNF2 expression was verified using immunohistochemistry in NSCLC tissues.
Results: The results indicated that GNPNAT1 was upregulated in NSCLC, and patients with high GNPNAT1 levels had a poor prognosis. GNPNAT1 overexpression promoted the proliferative and metastatic ability of lung cancer cells, whereas GNPNAT1 knockdown showed the opposite effect. GNPNAT1 expression was upregulated in NSCLC tissues compared to matched normal tissues as assessed by immunohistochemistry. Moreover, GNPNAT1 levels were positively correlated with histological type and pathological stage. The negative correlation between GNPNAT1 levels and OS was confirmed in 78 NSCLC patients. Aberrant RNF2 partly contributed to the upregulation of GNPNAT1 expression in NSCLC.
Conclusion: These findings suggested that GNPNAT1 was upregulated and played an important role in NSCLC. GNPNAT1 is expected to represent an effective prognostic biomarker for NSCLC patients.
Keywords: non-small cell lung cancer, glucosamine-phosphate N-acetyltransferase 1, overall survival, proliferation, metastasis
Introduction
Lung cancer is a commonly diagnosed cancer worldwide with an estimated 1.8 million new lung cancer cases diagnosed each year based on global cancer statistics.1 Approximately 85% of lung cancers are classified as non-small cell lung cancer (NSCLC), which is mainly composed of lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC).2 Over the past few decades, substantial progress has been made in the screening, diagnosis and treatment of lung cancer.3 However, this progress has been far from sufficient to reduce the mortality of NSCLC. Understanding the pathogenesis of lung cancer is necessary to identify more effective screening strategies and therapies.
Glucosamine-phosphate N-acetyltransferase 1 (GNPNAT1) is a key enzyme in the biosynthetic pathway of uridine diphosphate-N-acetylglucosamine (UDP-GlcNAc), which plays a key role in the N-linked glycosylation of proteins and the generation of glycosyl phosphatidylinositol linkers.4 Inactivation of murine GNPNAT1 by homologous recombination reduces the amount of UDP-GlcNAc, therefore resulting embryonic death and the proliferation defective cells.5,6 In Saccharomyces cerevisiae, GNPNAT1 is necessary for cell survival.7 GNPNAT1 can be detected in most tissues in humans. The correlation between GNPNAT1 and cancer was first reported in prostate cancer.8 In recent years, several studies have been published sequentially and indicated that GNPNAT1 upregulation predicts poor prognosis in LUAD patients.9–12 Zhao et al found that Abraxane was a promising drug to treat NSCLC, and the application of Abraxane resulted in significant downregulation of GNPNAT1 in lung cancer cells.13 GNPNAT1 might play an important role in NSCLC biological processes and represent an underlying biomarker for prognosis.
In this study, we assessed the expression and prognostic value of GNPNAT1 in NSCLC patients, detected the possible function of GNPNAT1 in NSCLC development and analysed the underlying mechanism for GNPNAT1 upregulation in NSCLC. These results potentially uncover a valuable target or biomarker for NSCLC.
Materials and Methods
Dataset and Processing
The NSCLC cases in this study included LUAD and LUSC. GNPNAT1 mRNA expression data and prognostic information were obtained from TCGA and GEO databases. The datasets used in this study provided clinical stage information. These datasets were used to analyse the differential expression of GNPNAT1 between NSCLC and normal tissues and the correlation between GNPNAT1 levels and overall survival (OS). The transcription factors (TFs) regulating GNPNAT1 were screened using The Signalling Pathways Project (http://www.signallingpathways.org/ominer/query.jsf). We utilized TCGA database to analyse the correlation between GNPNAT1 and TFs in four datasets (LUSC tumours, LUSC tumour and lung tissues, LUAD tumours, LUAD tumour and lung tissues). Then, we obtained the intersection of the ten TFs with the highest correlation with GNPNAT1 in four datasets and finally identified four TFs that appeared in four datasets simultaneously. RNF2 mRNA levels in LUAD and LUSC were analysed using GEPIA (http://gepia.cancer-pku.cn/).
Patients
This study included 35 LUAD patients and 43 LUSC patients who received surgical treatment between May 2016 and August 2020 at Shenyang Chest Hospital. The cancer tissues and matched normal lung tissues were obtained from surgical excision. The mean age of the patients was 61.91 ± 9.33 years. All tissues were conventionally fixed with formaldehyde and embedded in paraffin. The diagnoses for all samples were confirmed by two pathologists. No patient received other therapy before surgery. All patients submitted their informed consent before enrolment. This study was approved by the Ethics Committee of Shenyang Chest Hospital. Our study complied with the Declaration of Helsinki.
Cell Construction
The ORF of GNPNAT1 was cloned into the pLVX-Puro vector (primers for GNPNAT1 overexpression: F-gaattcGCCTCTACGGACCTTACT, R-tctagaATTTATGGAGGTCACTCG). The shRNA sequence targeting GNPNAT1 was inserted into PLKO (primers for GNPNAT1 knockdown: F-CCGGGAGTCAGAATACAGCTACATTCTCGAGAATGTAGCTGTATTCTGACTCTTTTTG, R-AATTCAAAAAGAGTCAGAATACAGCTACATTCTCGAGAATGTAGCTGTATTCTGACTC). Scramble sequences were inserted into vectors to function as controls, including PLVX or negative controls (NCs). These vectors were co-transfected into 293TN cells with packing vectors to obtain pseudolentiviral particles, which were used to infect A549 (lung adenocarcinoma cells) and NCI-H266 (lung squamous carcinoma cells) cells. Cells were purified with 2 μg/mL puromycin (Beyotime, Shanghai, China). All the cells were cultured in RPMI-1640 (GIBCO, USA) supplemented with 10% foetal bovine serum (FBS; Invitrogen, Carlsbad, CA, USA), 100 μg/mL streptomycin (Beyotime), and 100 IU/mL penicillin (Beyotime) at 37 °C in a humidified atmosphere containing 5% CO2.
Real-Time PCR
Total RNA was extracted from lung cancer cells using TRIzol® (Invitrogen) and then reverse transcribed into cDNA using a reverse transcription kit (Trans Gen Biotech Co, Beijing, China). GNPNAT1 was quantified using the SYBR Green PCR Mix (Bioresearcher, Beijing, China). The reaction included 38 cycles, and the cycle included denaturation for 10s at 95 °C and annealing and prolongation for 30s at 60 °C. GAPDH served as the endogenous control. Each sample was tested in triplicate. All assays were replicated thrice. The following primers with sequences were employed: GNPNAT1, F-ACCCAAGTCTACTCAAAG and R-TCAGCAGTACAAAGAGGC; GAPDH, F-ATGGGGAAGGTGAAGGTCG and R-GGGGTCATTGATGGCAACAATA.
MTT Assay
Cells were seeded into 96-well plates and cultured for 4, 24, 48 and 72 h. Then, 20 μL of MTT solution (Beyotime) was added to the well, and the samples were incubated for 4 h. The absorbance at 492 nm was measured using a microplate reader. The relative proliferative capacity was reflected based on the OD value. The OD value obtained at 4 h after inoculation represented the number of cells at inoculation.
Transwell Assay
A total of 3×104 cells in 500 μL serum-free medium were seeded into the upper chamber coated with or without Matrigel (Corning Life Sciences, Tewksbury, MA, USA). Then, 700 μL medium containing 10% FBS was added to the lower chamber. After culturing for 24 h, the chamber was placed into a 0.5% crystal violet solution (dissolved in methanol) to fix and stain the cells. The cells on the top side of the upper chamber were removed with a cotton swab, whereas the cells on the opposite side of the insert were counted using an inverted light microscope. Three high-power fields were selected to calculate the average number of cells to reflect the cell migration and invasion abilities.
Immunohistochemistry (IHC)
Briefly, 5-µm-thick sections were cut from formalin-fixed and paraffin-embedded specimens. After deparaffinization and rehydration, the paraffin sections were treated with 3% H2O2 to block endogenous peroxidase at RT for 15 mins. Antigen retrieval was performed through microwave treatment (citrate buffer, pH 6.0). The nonspecific reaction was blocked with 5% bovine serum albumin (ZSGB Bio, Beijing, China) at RT for 15 mins. The sections were then incubated with rabbit polyclonal primary antibody against GNPNAT1 (Proteintech, Wuhan, China, catalogue number: 16282-1-AP, 1:200 dilution) and rabbit polyclonal primary antibody against RNF2 (Affinity Biosciences, Liyang, China, catalogue number: DF7403, 1:100 dilution) at 4 °C overnight. A streptavidin-biotin peroxidase kit (ZSGB Bio) and DAB (ZSGB Bio) were used to visualise the antigen-antibody reaction.
The final immunohistochemical score for each sample was calculated using the following formula: intensity score × the extent score. Staining intensity was quantified as follows: negative (0), weak (1), moderate (2), or strong (3). The staining extent was scored based on the quantity of positive cells: none (0), < 25% (1), 25–50% (2), 50–75% (3), or > 75% (4).14 Two pathologists evaluated the results in a blinded manner. The diverging scores were reviewed, and a consensus was reached between the two pathologists.
Statistical Analysis
The difference between two groups was analysed by a two-sided Student’s t test. One-way ANOVA was used to analyse the correlation between GNPNAT1 and clinical pathological parameters by screening logical independent factors. Then, multivariate linear stepwise regression was used to assess the association between GNPNAT1 level and meaningful independent factors. The effect of GNPNAT1 on overall survival (OS) was analysed using the Kaplan–Meier method through the Log rank test. The median relative mRNA level or IHC score was used as the cut-off value for high and low GNPNAT1 mRNA group classification in different datasets. The correlation between GNPNAT1 and TFs was analysed using the Pearson correlation coefficient. All the data are presented as the mean or median. P < 0.05 indicated statistical significance.
Results
GNPNAT1 mRNA Was Upregulated in NSCLC and Indicated a Poor Prognosis
We downloaded the GNPNAT1 expression datasets of NSCLC, LUAD and LUSC from the GEO and TCGA databases to compare the differential expression of GNPNAT1 mRNA between tumour and matched normal tissues. GNPNAT1 expression was significantly upregulated in NSCLC as well as in LUSC and LUAD compared to normal tissues (Figure 1A and C). GNPNAT1 mRNA levels increased with pathological stage from I to III/IV in NSCLC, LUSC and LUAD (Figure 1B and D). We used three GEO datasets to analyse the correlation between the GNPNAT1 level and the prognosis of patients with lung cancer. The results indicated that NSCLC, LUSC and LUAD patients with higher levels of GNPNAT1 had a shorter OS than those with lower levels of GNPNAT1 (Figure 1E). According to TCGA data analysis results, NSCLC patients with higher GNPNAT1 levels had a poor prognosis (Figure 1F). However, GNPNAT1 was a useful prognostic marker for LUAD patients but not for LUSC patients when the NSCLC patients were classified into LUAD and LUSC groups (Figure 1F). These results indicated that GNPNAT1 was upregulated in NSCLC tissues and might represent a biomarker to evaluate the prognosis of NSCLC patients, especially LUAD patients.
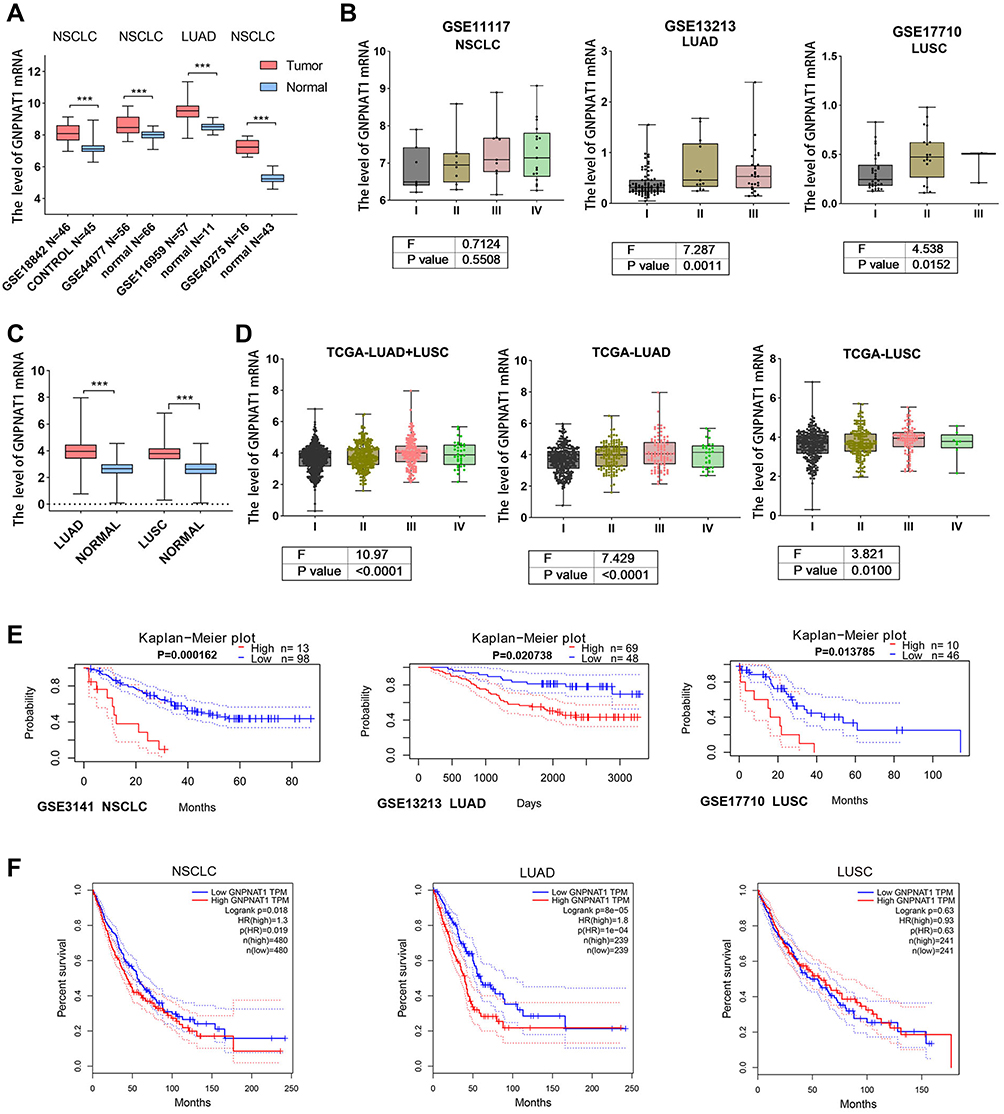 |
Figure 1 Dataset analysis. (A and C). The mRNA level of GNPNAT1 was compared between cancer and normal tissues using the GEO dataset (A) and TCGA dataset (C). B and D. GNPNAT1 mRNA levels were analysed in cancer patients with different pathological stages in the GEO dataset (B) and TCGA dataset (D). (E and F) Patients were divided into high and low groups according to the expression of GNPNAT1 in lung cancer, and then OS was compared between the two groups using the GEO dataset (E) and TCGA dataset (F). *** P<0.001. |
GNPNAT1 Improved the Proliferative and Metastatic Ability of Lung Cancer Cells
GNPNAT1 mRNA was detected by real-time PCR to verify the upregulation or knockdown of GNPNAT1 in A549 cells and NCI-H226 cells (Figure 2A). GNPNAT1 overexpression promoted cell proliferation, whereas knockdown of GNPNAT1 inhibited cell proliferation (Figure 2B). The abilities of GNPNAT1-overexpressing cells to migrate and invade were significantly improved, whereas these abilities were significantly decreased after GNPNAT1 knockdown in A549 (Figure 2C and D) and NCI-226 (Figure 2E and F) cells. The results showed that GNPNAT1 promotes the evolution of lung cancer cells.
 |
Figure 2 GNPNAT1 contributed to the evolution of lung cancer cells. (A) The mRNA level of GNPNAT1 was detected by real-time PCR. (B) The effect of GNPNAT1 on the proliferation of lung cancer cells was detected by MTT assay. (C and D) Transwell assays were used to detect the effect of GNPNAT1 on the migration (C) and invasion (D) of A549. (E and F) Transwell assays were used to detect the effect of GNPNAT1 on the migration (E) and invasion (F) of NCI-H226. |
GNPNAT1 Expression Was Increased in NSCLC Tissues and Correlated with Clinical Pathological Parameters
We detected GNPNAT1 expression in 78 NSCLC tissues (composed of 35 LUAD and 43 LUSC) and matched normal tissues by immunohistochemistry (Supplementary Figure 1). As shown in Figure 3A and B, GNPNAT1 was significantly upregulated in both LUAD and LUSC tissues compared to normal tissues. GNPNAT1 was localized in the cytoplasm (Figure 3A). GNPNAT1 was also expressed in normal tissues, but very weak expression was observed. Moreover, GNPNAT1 expression was increased in LUAD compared with LUSC (Figure 3B).
 |
Figure 3 Immunohistochemistry assay. (A) GNPNAT1 was upregulated in cancer tissues compared to normal tissues and was more highly expressed in LUAD than in LUSC. The image on the right is the enlarged image in the box. (B) The expression of GNPNAT1 was quantified as a bar chart. (C) The patients with low levels of GNPNAT1 (IHC score < 5) had a better OS than those with high levels of GNPNAT1 (IHC score ≥ 5). |
We analysed the correlation between GNPNAT1 and clinical pathological parameters in 78 NSCLC patients. The results indicated that age, histological classification, and TNM stage were correlated with GNPNAT1 expression (Table 1). Multivariate regression analysis confirmed that histological classification and TNM stage were two independent factors for GNPNAT1 expression, whereas age was excluded (Table 2). The medium GNPNAT1 staining score was higher in LUAD patients and TNM III–IV patients. Additionally, we analysed the correlation between GNPNAT1 level and prognosis. The results indicated that the patients with low GNPNAT1 levels (IHC score <5) had a better OS than the patients with high GNPNAT1 levels (IHC score ≥5), although the P value was marginal (Figure 3C).
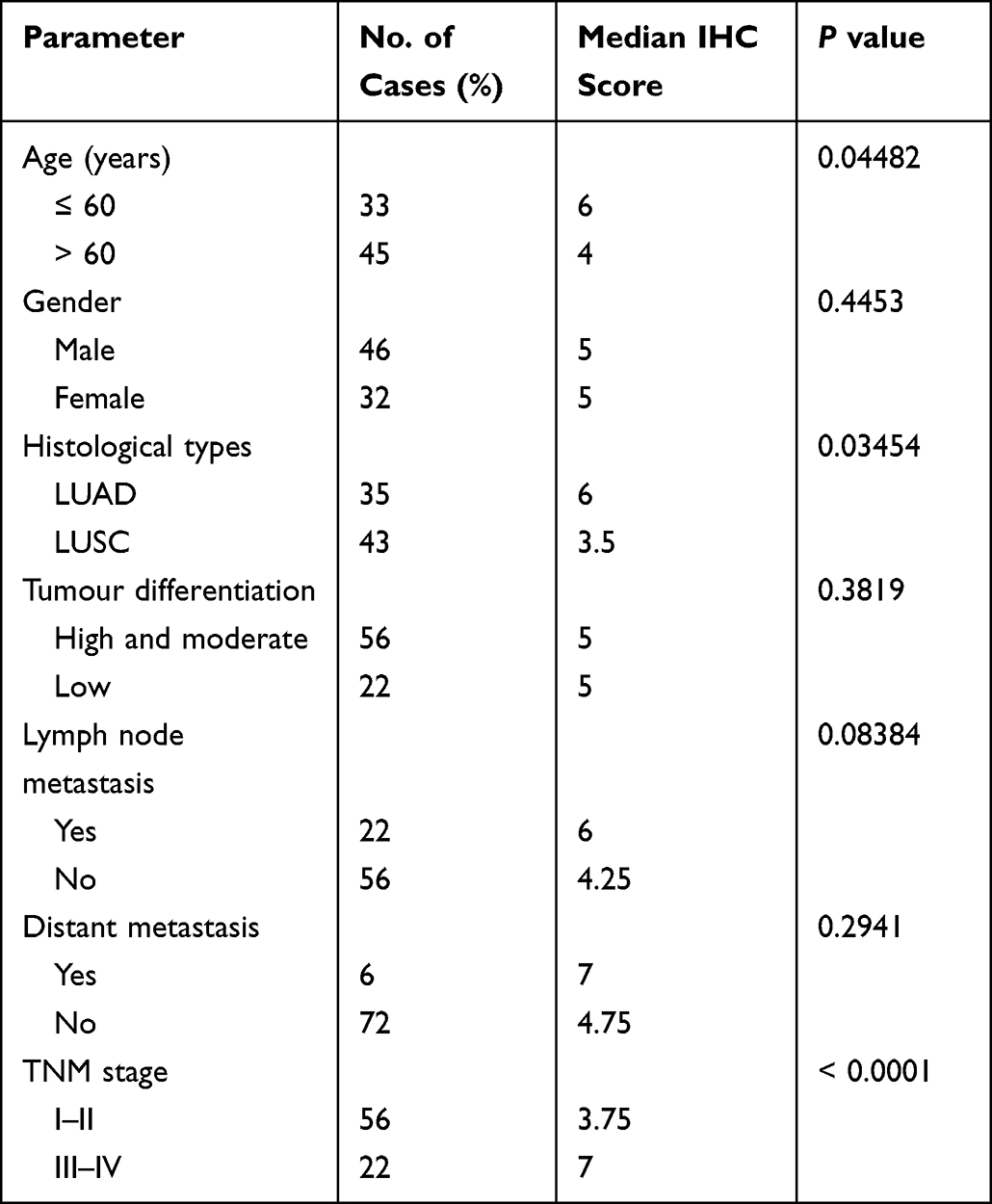 |
Table 1 Relationship Between GNPNAT1 Expression and Clinicopathological Parameters in NSCLC |
 |
Table 2 Multivariate Analysis of Factors Related to GNPNAT1 Expression |
These results suggested that GNPNAT1 had a role in normal tissues. However, its expression was significantly increased in cancer tissues. GNPNAT1 might have an important role during the development of NSCLC.
GNPNAT1 Upregulation Was Associated with Transcriptional Regulation
To explore the mechanism of GNPNAT1 upregulation, we first predicted the underlying TFs targeting GNPNAT1. A total of 504 TFs were found to have the potential to regulate GNPNAT1 transcription. We analysed the correlation between GNPNAT1 with TFs in different datasets, including LUSC tumours, LUSC tumour and lung tissues, LUAD tumours, LUAD tumour and lung tissues. We took the intersection of the ten TFs with the highest correlation with GNPNAT1 in four datasets and finaly obtained four TFs (Figure 4A). Moreover, RNF2, RAD21, YY1 and ZNF143 were positively correlated with GNPNAT1 in NSCLC consisting of LUAD and LUSC tissues (Figure 4B). Of them, RNF2 mRNA was upregulated in LUAD and LUSC (Figure 4C). We detected RNF2 expression in lung cancer tissues (Supplementary Figure 1) and found that RNF2 located in both cytoplasm and nucleus (Figure 4D). Considering that RNF2 mainly acts as a transcription factor, we focused on the expression of nucleic RNF2. The result indicated that nucleic RNF2 was not statistically different between LUAD and LUSC (Figure 4E). Moreover, RNF2 was positively correlated with GNPNAT1 in 78 cases of NSCLC (Figure 4F). In most cases, GNPNAT1 expression was higher in tissues with higher RNF2 expression, and lower in tissues with lower RNF2 expression. Thus, the upregulation of some TFs in NSCLC might represent one of the mechanisms for elevated GNPNAT1 expression.
 |
Figure 4 Aberrant TFs contributed to the upregulation of GNPNAT1 in NSCLC. (A) RNF2, ZNF143, RAD21, and YY1 were the intersecting TFs from the top ten TFs most correlated with GNPNAT1 in the four groups. (B) The four TFs were highly positively correlated with GNPNAT1 in LUAD and LUSC tissues. (C) RNF2 mRNA was increased in LUAD and LUSC tissues. (D) IHC assays indicated that RNF2 located in both cytoplasm and nucleus of lung cancer cells. (E) The nucleic RNF2 was not statistically different between LUAD and LUSC. (F) The nucleic RNF2 was positively correlated with GNPNAT1 in NSCLC. * P<0.05. |
Discussion
The glucosamine-6-phosphare acetyltransferase GNPNAT1 catalyses the reaction of glucosamine 6-phosphate with acetyl-CoA to form the precursor of UDP-GlcNAc, which plays a key role in the N-linked glycosylation of proteins.5,15 As previously reported, N-linked glycosylation is a posttranslational modification occurring in many eukaryotic secreted and surface-bound proteins that impacts the function of these proteins.16 UDP-GlcNAc is also involved in the generation of glycosyl phosphatidylinositol linkers, through which membrane proteins anchor to the cell.17 Thus, GNPNAT1 might function in multiple physiological and pathological processes by affecting UDP-GlcNAc formation.
The critical function of GNPNAT1 has been verified in different species. Toshiyuki et al found that a GNPNAT1-deletion mutant resulted in the reduced virulence of Candida albicans.7 The murine GlcNAc EMeg32 was first cloned in 2000.5 Inactivation of EMeg32 downregulated UDP-GlcNAc levels and subsequently led to embryonic death and reduced proliferation in embryonic stem cells and embryonic fibroblast cells.6 Recently, a study found that GNPNAT1 was necessary for growth plate chondrogenesis in humans.18 Knockdown of GNPNAT1 decreased cellular proliferation and the expression of chondrocyte differentiation markers.18
To date, a few studies have explored the function of GNPNAT1 in cancer. According to the results of Akash et al, GNPNAT1 was significantly elevated at the transcript and translational levels in prostate cancer compared with benign tissues, whereas lower levels were noted sites of lymph node metastasis and in castration-resistant prostate cancer (CRPC) tissues compared with localized prostate cancer.8 Loss of GNPNAT1 in CRPC-like cells resulted in increased proliferation and aggressiveness by activating the PI3K-AKT pathway.8 GNPNAT1 along with other metabolism-related genes correlate with the poor prognosis of LUAD patients, and an independent prognostic model was established based on these genes.19,20 GNPNAT1 overexpression potentially correlated with DNA copy amplification, low DNA methylation and loss of miRNA.10 Here, we showed that the upregulation of some TFs was also responsible for increased GNPNAT1 expression. The results of GNPNAT1 gene coexpression network analysis indicated that GNPNAT1 might participate in cell cycle transition, cancer development and immune infiltration in LUAD.11 The other two studies also reported a correlation between GNPNAT1 and immune infiltration, indicating the potential function of GNPNAT1 in immunomodulation and immunotherapy.9,21
In this study, we analysed GNPNAT1 expression in NSCLC. The results showed that GNPNAT1 was upregulated in NSCLC and associated with the poor prognosis of NSCLC patients. Similar results were obtained when we divided NSCLC patients into LUAD and LUSC subcategories. Several recent studies also reached consistent conclusions about GNPNAT1 in LUAD.9–12 However, few studies have reported the function of GNPNAT1 in NSCLC or LUSC. In addition, GNPNAT1 represents the key molecule facilitating the enhanced therapeutic effects of Abraxane on NSCLC.13 Abraxane downregulated the expression of GNPNAT1 and led to impaired proliferation in NSCLC cell lines.13 In this study, we identified the expression of GNPNAT1 protein in NSCLC tissues, detected its function in lung cancer cells, and consequently provided experimental evidence for GNPNAT1 function. Moreover, we found that GNPNAT1 was more valuable as a prognostic marker in LUAD patients. It is possible that GNPNAT1 expression was higher in LUAD patients than in LUSC patients according to the immunohistochemistry results. Our study revealed the potential function of GNPNAT1 in NSCLC. However, the mechanism of GNPNAT1 has not been clearly defined, which limits the clinical application of GNPNAT1. The value of GNPNAT1 as a prognostic biomarker needs to be confirmed in more patients. In the future, we will explore the mechanism by which GNPNAT1 exerts its effect on NSCLC and its potential in clinical prognosis assessment and treatment.
Conclusion
This study found that GNPNAT1 was significantly upregulated in NSCLC compared with normal lung tissues, and the effect was partly attributed to the aberrant TFs. Elevated GNPNAT1 levels are associated with the tumour histological type, stage, and poor prognosis of patients. GNPNAT1 might play an important role in NSCLC development and represent an underlying therapeutic target. More studies are needed to uncover the function of GNPNAT1 in cancer.
Abbreviations
GNPNAT1, glucosamine-phosphate N-acetyltransferase 1; NSCLC, non-small cell lung cancer; LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; UDP-GlcNAc, uridine diphosphate-N-acetylglucosamine; OS, overall survival.
Ethical Statement
All patients submitted their informed consent before enrolment. This study was approved by the Ethics Committee of Shenyang Chest Hospital.
Acknowledgments
Not available.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108.
2. Molina JR, Yang P, Cassivi SD, Schild SE, Adjei AA. Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc. 2008;83:584–594.
3. Wakelee H, Kelly K, Edelman MJ. 50 Years of progress in the systemic therapy of non-small cell lung cancer. Am Soc Clin Oncol Educ Book. 2014;1:177–189.
4. Wang J, Zhou YF, Li LF, Liang YH, Su XD. Purification, crystallization and preliminary X-ray analysis of the glucosamine-6-phosphate N-acetyltransferase from human liver. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006;62:1097–1099.
5. Boehmelt G, Fialka I, Brothers G, et al. Cloning and characterization of the murine glucosamine-6-phosphate acetyltransferase EMeg32. Differential expression and intracellular membrane association. J Biol Chem. 2000;275:12821–12832.
6. Boehmelt G, Wakeham A, Elia A, et al. Decreased UDP-GlcNAc levels abrogate proliferation control in EMeg32-deficient cells. EMBO J. 2000;19:5092–5104.
7. Mio T, Kokado M, Arisawa M, Yamada-Okabe H. Reduced virulence of Candida albicans mutants lacking the GNA1 gene encoding glucosamine-6-phosphate acetyltransferase. Microbiology. 2000;146(Pt 7):1753–1758.
8. Kaushik AK, Shojaie A, Panzitt K, et al. Inhibition of the hexosamine biosynthetic pathway promotes castration-resistant prostate cancer. Nat Commun. 2016;7:11612.
9. Zheng X, Li Y, Ma C, et al. Independent Prognostic Potential of GNPNAT1 in Lung Adenocarcinoma. Biomed Res Int. 2020;2020:8851437.
10. Liu W, Jiang K, Wang J, Mei T, Zhao M, Huang D. Upregulation of GNPNAT1 Predicts Poor Prognosis and Correlates With Immune Infiltration in Lung Adenocarcinoma. Front Mol Biosci. 2021;8:605754.
11. Zhang S, Zhang H, Li H, Guo J, Wang J, Zhang L. Potential role of glucosamine-phosphate N-acetyltransferase 1 in the development of lung adenocarcinoma. Aging. 2021;13:7430–7453.
12. Zhu P, Gu S, Huang H, et al. Upregulation of glucosamine-phosphate N-acetyltransferase 1 is a promising diagnostic and predictive indicator for poor survival in patients with lung adenocarcinoma. Oncol Lett. 2021;21:488.
13. Zhao M, Li H, Ma Y, et al. Nanoparticle Abraxane possesses impaired proliferation in A549 cells due to the underexpression of glucosamine 6-phosphate N-acetyltransferase 1 (GNPNAT1/GNA1). Int J Nanomedicine. 2017;12:1685–1697.
14. Wu DI, Liu L, Ren C, et al. Epithelial-mesenchymal interconversions and the regulatory function of the ZEB family during the development and progression of ovarian cancer. Oncol Lett. 2016;11:1463–1468.
15. Brockhausen I, Nair DG, Chen M, et al. Human acetyl-CoA: glucosamine-6-phosphateN-acetyltransferase 1 has a relaxed donor specificity and transfers acyl groups up to four carbons in length. Biochem Cell Biol. 2016;94:197–204.
16. Helenius A, Aebi M. Roles of N-linked glycans in the endoplasmic reticulum. Annu Rev Biochem. 2004;73:1019–1049.
17. Kostova Z, Rancour DM, Menon AK, Orlean P. Photoaffinity labelling with P3-(4-azidoanilido)uridine 5’-triphosphate identifies gpi3p as the UDP-GlcNAc-binding subunit of the enzyme that catalyses formation of GlcNAc-phosphatidylinositol, the first glycolipid intermediate in glycosylphosphatidylinositol synthesis. Biochem J. 2000;350(Pt 3):815–822.
18. Ain NU, Baroncelli M, Costantini A, et al. Novel form of rhizomelic skeletal dysplasia associated with a homozygous variant in GNPNAT1. J Med Genet. 2020;1:854.
19. Wang Z, Embaye KS, Yang Q, et al. Establishment and validation of a prognostic signature for lung adenocarcinoma based on metabolism-related genes. Cancer Cell Int. 2021;21:219.
20. Zhang J, Zhang J, Yuan C, et al. Establishment of the Prognostic Index Reflecting Tumor Immune Microenvironment of Lung Adenocarcinoma Based on Metabolism-Related Genes. J Cancer. 2020;11:7101–7115.
21. Yu X, Zhang X, Zhang Y. Identification of a 5-Gene Metabolic Signature for Predicting Prognosis Based on an Integrated Analysis of Tumor Microenvironment in Lung Adenocarcinoma. J Oncol. 2020;2020:5310793.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.