Back to Journals » Nature and Science of Sleep » Volume 14
Genome-Wide Association Study and Genetic Correlation Scan Provide Insights into Its Genetic Architecture of Sleep Health Score in the UK Biobank Cohort
Authors Yao Y, Jia Y, Wen Y, Cheng B, Cheng S, Liu L, Yang X, Meng P, Chen Y, Li C, Zhang J, Zhang Z, Pan C, Zhang H, Wu C, Wang X, Ning Y, Wang S, Zhang F
Received 28 June 2021
Accepted for publication 19 December 2021
Published 6 January 2022 Volume 2022:14 Pages 1—12
DOI https://doi.org/10.2147/NSS.S326818
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Yao Yao,* Yumeng Jia,* Yan Wen, Bolun Cheng, Shiqiang Cheng, Li Liu, Xuena Yang, Peilin Meng, Yujing Chen, Chun’e Li, Jingxi Zhang, Zhen Zhang, Chuyu Pan, Huijie Zhang, Cuiyan Wu, Xi Wang, Yujie Ning, Sen Wang, Feng Zhang
Key Laboratory of Trace Elements and Endemic Diseases, National Health and Family Planning Commission, School of Public Health, Health Science Center, Xi’an Jiaotong University, Xi’an, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Zhang
Key Laboratory of Trace Elements and Endemic Diseases, National Health and Family Planning Commission, School of Public Health, Health Science Center, Xi’an Jiaotong University, Xi’an, 710061, People’s Republic of China
Email [email protected]
Purpose: Most previous genetic studies of sleep behaviors were conducted individually, without comprehensive consideration of the complexity of various sleep behaviors. Our aim is to identify the genetic architecture and potential biomarker of the sleep health score, which more powerfully represents overall sleep traits.
Patients and Methods: We conducted a genome-wide association study (GWAS) of sleep health score (overall assessment of sleep duration, snoring, insomnia, chronotype, and daytime dozing) using 336,463 participants from the UK Biobank. Proteome-wide association study (PWAS) and transcriptome-wide association study (TWAS) were then performed to identify candidate genes at the protein and mRNA level, respectively. We finally used linkage disequilibrium score regression (LDSC) to estimate the genetic correlations between sleep health score and other functionally relevance traits.
Results: GWAS identified multiple variants near known candidate genes associated with sleep health score, such as MEIS1, FBXL13, MED20 and SMAD5. HDHD2 (PPWAS = 0.0146) and GFAP (PPWAS = 0.0236) were identified associated with sleep health score by PWAS. TWAS identified ORC4 (PTWAS = 0.0212) and ZNF732 (PTWAS = 0.0349) considering mRNA expression level. LDSC found significant genetic correlations of sleep health score with 3 sleep behaviors (including insomnia, snoring, dozing), 4 psychiatry disorders (major depressive disorder, attention deficit/hyperactivity disorder, schizophrenia, autism spectrum disorder), and 9 plasma protein (such as Stabilin-1, Stromelysin-2, Cytochrome c) (all LDSC PLDSC < 0.05).
Conclusion: Our results advance the comprehensive understanding of the aetiology and genetic architecture of the sleep health score, refine the understanding of the relationship of sleep health score with other traits and diseases, and may serve as potential targets for future mechanistic studies of sleep phenotype.
Keywords: sleep, sleep health score, genome-wide association study, genetics, complex-traits
Introduction
Sleep is a complex neurological and physiological state. It is defined as a natural and reversible state of reduced responsiveness to external stimuli and relative inactivity, accompanied by a loss of consciousness.1 Sleep disorders can be classified as seven major categories: insomnia disorders, sleep-related breathing disorders, central disorders of hypersomnolence, circadian rhythm sleep-wake disorders, sleep-related movement disorders, parasomnias, and other sleep disorders.2 Emerging evidence suggested that sleep disorders were associated with various cardiovascular and neurodegenerative disorders.3 Besides, several unhealthy sleep behaviors, such as short or long sleep, evening-type chronotype preference, insomnia, snoring, and daytime dozing were associated with increased risk of psychiatric disorders.4–6 For example, from the perspective of genetics, several sleep phenotypes such as insomnia, snoring, and daytime dozing shared genes with attention-deficit/hyperactivity disorder (ADHD);4 a Mendelian randomization study conducted by Daghlas et al7 provided robust evidence of the protective association between earlier diurnal preference and major depressive disorder (MDD). From a neurobiological point of view, MDD is associated with exaggerated rapid eye movement (REM) sleep qualities, and activity in emotion-related regions increases significantly during REM sleep.8,9
Individual differences in sleep time and quality are affected by multiple variables, such as sex, age, psychological and physiological conditions, skin temperature and sweating during sleep, culture and environmental factors.10 Besides, genetic variation also constitutes an important factor to individual differences in sleep preference. Twin and family studies estimated the heritability of sleep phenotypes: self-reported napping and daytime sleep duration were 65% and 61%,11 sleep duration and sleep quality were 46% and 44%,12 chronotype preference was 37% for morningness-eveningness questionnaire (MEQ),13 insomnia was 39%.14 Single nucleotide polymorphism (SNP)-based heritability estimated at 10% for self-reported snoring and at 11.9% for daytime napping.15,16 This also was supported by a number of candidate gene studies as well as genome-wide association studies (GWAS).15–19 For example, a large-scale GWAS estimated from accelerometer data identified 47 genetic associations at P < 5 × 10−8 across seven traits representing sleep duration, quality and timing, suggesting a polygenic model of inheritance.17 The largest GWAS published to date of self-reported daytime napping identified 123 loci, and explained 1.1% of the variance in daytime napping.16 In addition, the circadian clock drives daily physiology, such as the sleep-wake cycle, by regulating transcription, protein abundance, and function. Insufficient sleep affects the human brain, liver and blood transcriptome and disrupts its circadian regulation.20,21 In brain tissue, almost 70% of transcripts have been reported to be changed in abundance during daily circadian cycles. Transcripts and proteins related to synaptic signaling accumulated preceded the active phase, whereas accumulations of messenger RNAs and protein associated with metabolism and translation have been reported before the resting phase.21 This was supported by previous transcriptome-wide association study (TWAS). A previous TWAS in up to 446,118 adults of European ancestry from the UK Biobank, identified 38 genes for which sleep duration SNPs influenced gene expression in 11 tissues, including nine brain regions, pituitary, whole blood.22 Another TWAS in one or more of the 14 tissue identified 24 genes for which insomnia SNPs influenced gene expression.23
A GWAS is commonly used to identify variants at genomic loci that are associated with complex traits. More recently, GWASs have made a major contribution to our understanding of the genetic architecture of sleep-related traits. Forty-two genome-wide significant loci for snoring were discovered in GWAS with an SNP-based heritability estimate of ~10% on the liability scale.15 In addition, 351 genomic risk loci associated with chronotype,18 123 loci associated with self-reported daytime napping16 and 202 genomic loci associated with insomnia19 were also identified by GWAS. Notably, sleep behaviors are typically related and may affect each other. A cross-sectional study reported that participants who reporting napping or frequent snoring were less likely to have insomnia symptoms.24 Chronotype was also associated with sleep duration as reported previously.25 Previous studies used various questionnaires to assess sleep quality, for example, the Pittsburgh Sleep Quality Index (PSQI), which obtains information about: subjective sleep quality, sleep latency, sleep duration, habitual sleep efficiency, sleep disturbances, use of sleeping medication, and daytime dysfunction.26,27 However, the majority of previous genetic studies did not consider the complex nature of the overall sleep behaviors and focused on individual trait. Here, we leverage data from the UK Biobank to identify the genetic architecture of the sleep health score, which is a combination of major sleep behaviors.
GWAS could identify genetic markers associated with sleep health score variation. In contrast, a proteome-wide association study (PWAS) is a new method for detecting gene-phenotype associations mediated by protein function alterations.28 Proteins are the final products of gene expression and the main functional components of cells, and thus are directly responsible for phenotype. PWAS could identify loci that confer sleep health score through their effects on brain protein and provide new insights into its mechanism. Contrary to GWAS, PWAS could reduce burden of multiple testing correction and detect a signal that is too weak and spread to appear in GWAS, which is crucial in the case of rare variants.28 TWAS is another method to identify gene-trait associations at the level of gene expression by integrating GWAS and gene expression datasets.29 These two methods could prioritize the most likely variants/genes causally affecting the phenotype more accurately.28,29
In this work, we leveraged the UK Biobank dataset to define the genetic architecture of the sleep health score. To gain insight into how these loci contribute to sleep health score mechanism, we integrated sleep health score GWAS results with human brain proteomes to perform a PWAS, and with expression data from brain to perform a TWAS, respectively. To better reveal the underlying connections across sleep-related human complex traits, we also employed linkage disequilibrium score regression (LDSC) to estimate genetic correlations between sleep health score and a variety of traits and diseases for which published large-scale genome-wide summary statistics were available. Previous studies on sleep health score assessed the relationship between sleep health score and the risk of cardiovascular disease30 or type 2 diabetes.31,32 Our findings are a step forward in identifying the genetic factors that contribute to sleep health score and add novel insights into the association of sleep health score with diseases.
Materials and Methods
Ethical Approval
The UK Biobank study has ethical approval derived from Northwest Multi-Center Research Ethics Committee (reference 11/NW/0382). All participants provided written informed consent to participate. Information about ethics oversight can be found at https://www.ukbiobank.ac.uk/ethics/.
UK Biobank and Definition of the Sleep Health Score
The genetic and phenotypic data of this study were obtained from the UK Biobank under application 46,478 (http://www.ukbiobank.ac.uk/about-biobank-uk/). UK Biobank is a large-scale cohort study on half a million participants aged 40–69 years recruited in 2006–2010.33 Information on participants’ health-related aspects was collected through touchscreen questionnaires, brief verbal interviews and a range of physical measurements. Blood samples were collected for genotyping. Genotyping, quality control (QC) and imputation were conducted by the UK Biobank and were described previously.33
In the present study, we included 336,463 unrelated participants (185,165 females, 56.98 ± 7.93 years) with complete genotypes, self-reported sleep behaviors and covariate data from the UK Biobank. Five self-reported sleep behaviors (sleep duration, chronotype, snoring, daytime dozing and insomnia) were used to generate a sleep health score according to the previous study.30 Sleep duration was defined using the following question, “About how many hours sleep do you get in every 24 h? (include naps)”. Then, we categorized the sleep duration as short sleep (<7 h/day), normal (7–8 h/day), and long (≥9 h/day) consistent with previous studies.30 The participants with normal sleep were recoded as code 0 and with short or long sleep were recoded as code 1. Chronotype preference was defined based on the question,
Do you consider yourself to be (1) definitely a ‘morning’ person, (2) more a ‘morning’ than ‘evening’ person, (3) more an ‘evening’ person than a ‘morning’ person, (4) definitely an ‘evening’ person.
We recoded the participants who responded (1) and (2) as code 0 and who responded (3) and (4) as code 1. Insomnia symptom was assessed using the question “Do you have trouble falling asleep at night or do you wake up in the middle of the night?”, participants with answer (1) never/rarely were recoded as code 0 and participants with answer (2) sometimes and (3) usually were recoded as code 1. Snoring was obtained using the question “Does your partner or a close relative or friend complain about your snoring?” with responses of (1) yes and (2) no, participants with the answer (2) were recoded as 0. Daytime dozing was obtained by asking “How likely are you to doze off or fall asleep during the daytime when you don’t mean to? (eg, when working, reading or diving)”, the responders with the answer (0) never/rarely and (1) sometimes were recoded as code 0, and the answer (2) often and (3) all the time were recoded as code 1.
For each sleep behavior, participants with code 0 were defined as low risk factor and received a score of 1. The sleep health score was obtained from the sum scores of the five sleep behaviors (ranges from 0 to 5), with the higher scores representing the healthier overall sleep traits. Overall sleep traits were defined as “healthy overall sleep traits” (sleep health score >3), “intermediate overall sleep traits” (1 health score <3) and “poor overall sleep traits” (sleep health score ≤1) according to the sleep health score. The sleep health score in our study has been validated by the previous studies in the UK Biobank.30
Genome-Wide Association Study for Sleep Health Score in UK Biobank
GWAS analyses were performed in a maximal set of unrelated self-reported White participants in the UK Biobank using the PLINK2.0 software.34 We used KING software (http://people.virginia.edu/?wc9c/KING/) to generate unrelated subjects. Kinship coefficients had been calculated using the genetic data for each pair of participants in the cohort. Duplicate and related individuals were removed. Subjects who had a self-reported sex inconsistent with the genetic gender, who were genotyped but not imputed or who withdrew their consents were also removed. GWAS was performed using a generalize linear regression model under an additive genetic model, using the sleep health score as the dependent variable, using gender, age, first ten principal components of the genome as covariates. Call rates < 0.90, Hardy–Weinberg equilibrium exact test P values < 0.001, or minor allele frequencies (MAFs) < 0.01 were excluded for quality control filters. We applied a nominal genome-wide statistical significance threshold level of p < 5×10−8. Manhattan plots were generated by the “CMplot” R script (https://github.com/YinLiLin/R-CMplot).
Proteome-Wide Association Study
We integrated sleep health score GWAS results with reference human brain proteomes to perform a PWAS of sleep health score using the Functional Summary-based Imputation (FUSION) (https://gusevlab.org/projects/fusion/) pipeline.35 PWAS is used to identify loci that confer sleep health score through their effects on brain protein.28 Human brain proteomes were generated from the dorsolateral prefrontal cortex (dPFC) of 152 participants of European descent recruited by Banner Sun Health Research Institute.36 Proteomic profiling was performed using isobaric tandem mass tag (TMT) peptide labeling and liquid chromatography coupled to mass spectrometry. For proteome quality control, proteins with missing values in more than 50% were excluded, outlier samples were identified and removed through iterative principal component analysis. Regression was used to remove the effects of proteomic sequencing batch, age, sex, postmortem interval and final clinical diagnosis of cognitive status from the confirmation proteomic profiles before estimating the protein weights. After quality control, the proteomic profiles included 8168 proteins, of which 1139 were heritable and their protein weights were included in the PWAS. Individuals from Banner were genotyped using an Affymetrix Precision Medicine Array. After quality control, 152 individuals with proteomic and genetic data were included. More information on proteomic profiling, proteome quality control, genotyping was described in detail previously.37
Transcriptome-Wide Association Study
TWAS is a method to identify gene-trait associations at the level of gene expression by integrating GWAS and gene expression datasets.29 In our study, TWAS of sleep health score was performed by the FUSION tool (https://gusevlab.org/projects/fusion/) through integrating sleep health score GWAS results with precomputed gene expression reference weights of brain (the dorsolateral prefrontal cortex) tissue obtained from TWAS/FUSION website.35 TWAS is capable to identify significant cis genetic correlations between imputed gene expression and sleep health score, aiding identification of additional susceptibility loci. Bayesian sparse linear mixed model (BSLMM) was used to compute the SNP-expression weights in the 1-Mb cis loci of the gene for a given gene.38 The association testing statistics between predicted gene expression and target diseases was calculated as ZTWAS=w’Z/(w’Lw)1/238. Z denotes the scores of sleep health score, w denotes the weights, and L denotes the SNP-correlation (LD) matrix. We set P value <0.05 for statistical significance.
Genetic Correlations with Other Traits and Diseases
We further used LDSC (https://github.com/bulik/ldsc) to estimate the genetic correlation between sleep health score and other traits, by comparing sleep health score GWAS results with GWAS summary statistics of other traits. LDSC is used to estimate heritability and genetic correlation from GWAS summary statistics.39 For the correlation of sleep health score and other specific sleep behaviors, we obtained published GWAS summary statistics for insomnia (N = 386,533 individuals), snoring (N = 359,916), dozing (N = 386,548) and napping (N = 386,577) in the UK biobank.19 Jansen et al19 identified 202 loci implicating 956 genes associated with insomnia, and reported GWASs of other sleep-related traits in the UK biobank. Unhealthy sleep behaviors have coincided with a strong increase in risk factors for psychiatric disorders.4,5,19 Genetic correlation was also estimated between sleep health score and 5 psychiatry disorders: bipolar disorder (BD) (20,352 cases and 31,358 controls),40 major depressive disorder (MDD) (135,458 cases and 344,901 controls),41 attention deficit/hyperactivity disorder (ADHD) (19,099 cases and 34,194 controls),42 schizophrenia (11,260 cases and 24,542 controls),43 autism spectrum disorder (ASD) (18,382 cases and 27,969 controls).44 We also obtained summary statistics for the genetic correlation calculation for the plasma protein (2994 plasma proteins in 3301 individuals of European descent).45 These previous GWAS studies of psychiatry disorders and plasma protein identified multiple loci and provided GWA summary statistics at the PGC website (https://www.med.unc.edu/pgc/results-and-downloads) or (http://www.phpc.cam.ac.uk/ceu/proteins/). As recommended, LD scores from European ancestry individuals from 1000G were calculated for SNPs in the HapMap 3 SNP set and used to calculate genetic correlation.
Results
Genome-Wide Association Analysis
Table 1 shows the baseline characteristics of the study participants according to sleep health score. A total of 336,463 unrelated participants (185,165 females, 56.98 ± 7.93 years) with complete genotype, self-reported sleep behaviors and covariate data from the UK Biobank were available for study. We identified 32 genome-wide significant (P < 5 × 10−8) variants near known candidate genes to be associated with sleep health score in our GWAS, after adjusted for gender, age and first ten principal components of the genome (Figure 1, Supplementary Table 1). For example, the most significant SNP was rs113851554 (P = 3.39 × 10−14) near MEIS1 on chromosome 2, several other significant associations were with variants close to FBXL13 (rs375365692, P = 1.80 × 10−10), MED20 (rs2274578, P = 6.95 × 10−10) and SMAD5 (rs35711851, P = 9.69 × 10−9).
 |
Table 1 Baseline Characteristics of the Study Participants |
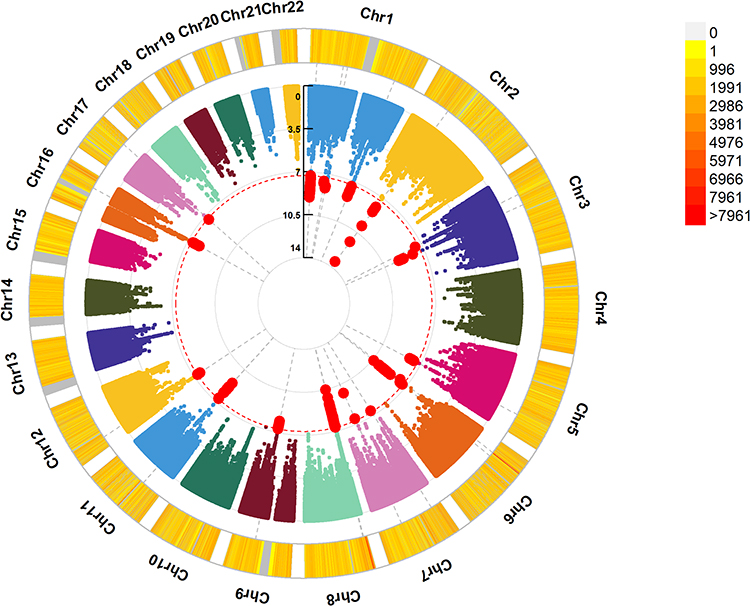 |
Figure 1 Circular Manhattan plot of sleep health score genome-wide association study. The P values are from the sleep health score GWAS results of 336,463 participants from the UK biobank. The line that cross the circle represents the –log10 p value for each genetic variant. The red line on the Manhattan plot represents the genome-wide significance threshold (P < 5×10−8). The colors of the circle represent chromosome density. The plot was generated by the “CMplot” R script (https://github.com/YinLiLin/R-CMplot). The significant genomic loci available in Supplementary Table 1. |
Many of the previous studies conducted GWASs of individual sleep behaviors.15,16,18,19 We compare our results with the three previous individual GWAS studies on sleep traits, including 446,118,22 386,53319 and 237,62723 individuals from the UK Biobank. Supplementary Table 2 presents candidate genes that are associated with sleep health score or associated with individual sleep behaviors including sleep duration, chronotype (morningness), snoring, daytime dozing and insomnia. We observed 2 genes (RPL10P19 and AC022784.1) that determined both the sleep health score and sleep duration, 9 genes (such as FBXL13 and DDI2) that determined both the sleep health score and chronotype (morningness), ROBO2 gene that determined both the sleep health score and snoring, 3 genes (MEIS1, PSMC3 and SMAD5) that determined both the sleep health score and insomnia, but no genes that determined both the sleep health score and daytime dozing. In addition, we observed 18 novel genes that associated with sleep health score, such as MED20, SMAD5. Our results extend the previous genetic studies on individual sleep behaviors by jointly evaluating multiple sleep behaviors.
PWAS of Sleep Health Score Identified 2 Loci at the Protein Level
To identify genes that affect sleep health score through their effects on brain protein abundance, we integrated sleep health score GWAS results with human brain proteomes to perform a PWAS. The PWAS identified 2 genes whose cis-regulated brain protein levels were associated with sleep health score at empirical permutation p-value < 0.05, including HDHD2 (Chr: 18, ZPWAS = 2.633, PPWAS = 0.0146) and GFAP (Chr: 17, ZPWAS = −3.26, PPWAS = 0.0236).
TWAS of Sleep Health Score Identified 2 Loci at the mRNA Level
To further explore genes that were associated with sleep health score at the transcriptional level, we integrated sleep health score GWAS results with precomputed gene expression reference weights of brain (the dorsolateral prefrontal cortex) tissue to perform a TWAS. The TWAS also identified 2 genes whose genetically regulated mRNA expression levels were associated with sleep health score at empirical permutation p-value < 0.05, including ORC4 (Chr: 2, ZTWAS = −2.11, PTWAS = 0.0212) and ZNF732 (Chr: 4, ZTWAS = 2.0659, PTWAS = 0.0349). By comparing the TWAS results on sleep health score with previously published TWAS results on sleep duration22 and insomnia,23 we found that none of the sleep duration and insomnia genes previously identified by TWAS were replicable in our sleep health score TWAS results. We considered 2 genes as novel for which sleep health score SNPs influenced gene expression in brain tissue.
Genetic Correlations with Other Traits
LDSC regression observed several genetic correlation signals between sleep health score and other traits with P values < 0.05 (Figure 2). For specific sleep behaviors, we observed significant negative genetic correlations with insomnia (rg = −0.5679, SE = 0.0331, P = 4.58 × 10−66), snoring (rg = −0.3356, SE = 0.0312, P = 6.60 × 10−27) and dozing (rg = −0.2986, SE = 0.0714, P = 2.91 × 10−5). We also determined genetic correlations with 5 psychiatry disorders and found significant negative genetic correlations with MDD (rg = −0.3664, SE = 0.0427, P = 9.02 × 10−18), ADHD (rg = −0.2506, SE = 0.05, P = 5.33 × 10−7), schizophrenia (rg = −0.1315, SE = 0.0361, P = 0.0003) and ASD (rg = −0.1716, SE = 0.0557, P = 0.0021). Further, we determined genetic correlations with plasma protein and found significant correlations with 9 plasma protein, such as Stabilin-1 (rg = −0.3177, SE = 0.1284, P = 0.0134), Stromelysin-2 (rg = −0.3803, SE = 0.1727, P = 0.0276) and Cytochrome c (rg = −0.227, SE = 0.1112, P = 0.0413). See Table 2 for details.
 |
Table 2 Sleep Health Score Genetic Correlations with Other Traits (P < 0.05) |
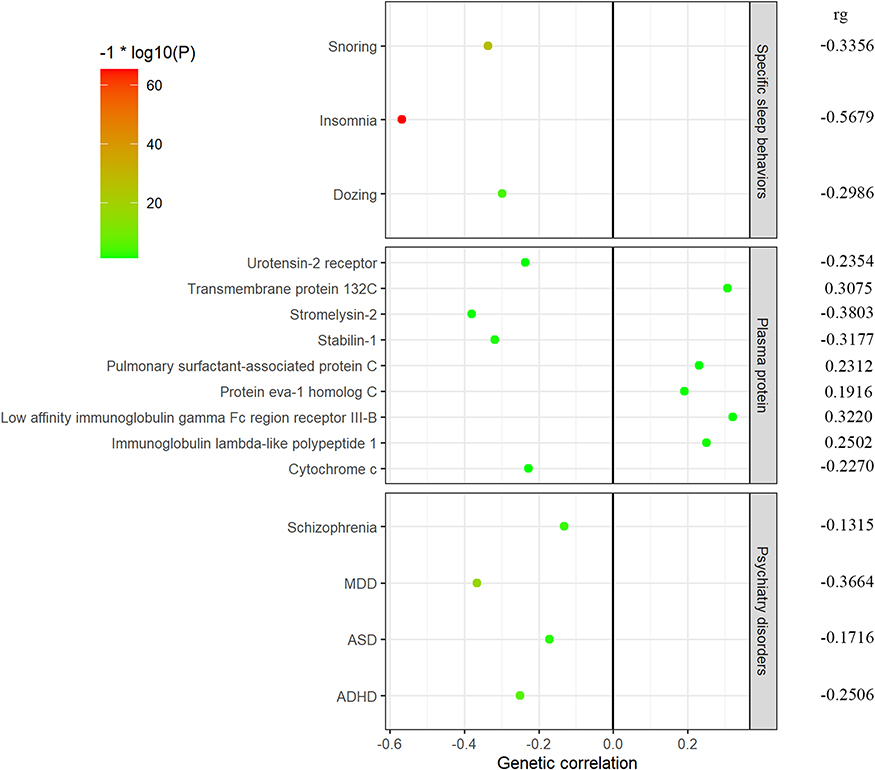 |
Figure 2 Sleep health score genetic correlations with other traits. Genome-wide genetic correlations between sleep health score and published summary statistics for other sleep behaviors, psychiatry disorders and plasma protein. Data presented are genetic correlation (rg) from LDSC analysis. −1*log10(P) represents -log10 (p-value). The P value from high to low is show as the color of the circle from green to red. Full results are described in Table 2. |
Discussion
In this large comprehensive genetic analysis in the UK biobank cohort, we examined the genetic architecture of the sleep health score, which could reflect a more comprehensive sleep phenotype through taking into account the combined impact of five sleep behaviors on health. A total of 336,463 unrelated participants from the UK Biobank were available and we found that 32 genetic loci were associated with the sleep health score in GWAS, such as MEIS1, FBXL13, SMAD5 and MED20. Furthermore, we performed a PWAS to identify genes associated with sleep health score at the protein level and identified two (HDHD2 and GFAP) genes. We also performed a TWAS and identified two (ORC4 and ZNF732) genes that were associated with sleep health score at the mRNA level. Lastly, through LDSC regression study, we demonstrated the shared biology of sleep health score with three sleep behaviors (insomnia, snoring and dozing), four psychiatry disorders (MDD, ADHD, schizophrenia and ASD), and nine plasma protein (such as Stabilin-1, Stromelysin-2 and Cytochrome c).
The strongest association found in GWAS was rs113851554 near MEIS1 gene. MEIS1 belongs to the homeobox and is a member of three-amino-acid loop extension homeodomain transcription factors. The MEIS1 GWAS signal has previously been identified to be associated with insomnia complaints.46 Genetic variants at MEIS1 locus were also found to be associated with restless legs syndrome (RLS), which is a common sleep-related disorder characterized by an uncontrollable urge to move and sometimes painful sensations in the legs.47 RLS could lead to sleep-onset or sleep-maintenance insomnia, and occasionally excessive daytime sleepiness.48 Researchers suggested the hypothesis that MEIS1 may exhibit pleiotropy for insomnia and RLS.46
We found that rs375365692 in the FBXL13 was also one of the most strongly associated variant with sleep health score. FBXL13 encodes a protein in the F-box protein family with leucine-rich repeats and is well known to be associated with the regulation of circadian rhythm.49 FBXL13 is near the PER2, which plays an important role in setting the speed of the molecular clock.50 Mutations of FBXL13 are associated with lengthened circadian periods in mice.51 Other variants identified in GWAS associated with sleep health score were near to genes such as SMAD5, MED20. GWAS recently has shown that SMAD5 is associated with insomnia symptoms.23 SMAD is known to transduce the action of TGF-β superfamily proteins, which play key roles in brain homeostasis, memory formation and neuronal plasticity.52 Mice with reduced expression of SMAD1 and SMAD5 in neural progenitors present with microcephaly and an increased production of early-born cortical neurons.53 MED20 encodes a component of the mediator complex, and is a coactivator involved in the regulated transcription of nearly all RNA polymerase II-dependent genes. Mediator complex is critical for normal neuronal function. Mutation in this gene has been associated with a novel infantile-onset neurodegenerative movement disorder characterized with infantile-onset spasticity and childhood-onset dystonia, progressive basal ganglia degeneration, and brain atrophy.54
In addition, we compared the results of sleep health score GWAS of our study with the three previous individual GWAS studies on sleep traits, including 446,118,22 386,53319 and 237,62723 individuals from the UK Biobank. Some of the genes showing significant associations in previous studies of individual sleep traits were also identified in our study (Supplementary Table 2). For example, MEIS1 and SMAD5 were reported to be associated with insomnia,23 FBXL13 was identified to be related to chronotype, ROBO2 was identified to be related to chronotype and snoring. Although some of the genes previously identified by the GWAS on individual sleep traits were not identified here, such as BAX8, BANK1 of sleep duration,22 RGS16, RNASEL of chronotype,19 DLEU7 of snoring,19 AC095050.1 of dozing,19 HEXIM2/LOC339192, CCDC68/TCF4 of insomnia,23 more novel sleep-related genes were identified by using the overall measure of sleep health. Sleep behaviors are correlated, the human body usually regulates sleep in a holistic way. Sleep behaviors may interact with each other in a compensatory fashion. The available genetic findings are partially limited due to not considering the complex nature of the overall sleep behaviors and focusing on limited sleep traits. Thus, our results extend the previous genetic studies on individual sleep behaviors by jointly evaluating multiple sleep behaviors. However, we dichotomized five sleep factors to create a combination for a sleep health measure for simplicity, which might lead to the loss of sleep information.
PWAS identified two genes (HDHD2 and GFAP) at the brain protein level, which may play an important role in the mechanism of sleep health score. HDHD2 encodes a novel protein found in proteomics analysis of hippocampus from chronic restraint stress (CRS) mouse model. HDHD2 was significantly changed in the hippocampus of animals exposed to CRS.55 Search according to NCBI conserved domain database (CDD) (https://www.ncbi.nlm.nih.gov/cdd), HDHD2 belongs to the HAD-like hydrolase superfamily structurally and is found to be similar to phospholysine phosphohistidine inorganic pyrophosphate phosphatase (LHPP), which is known to associated with depression.56 Sleep abnormalities are one of the critical symptoms of depression.57 However, its function is not completely understood. GFAP gene encodes one of the major intermediate filament proteins of mature astrocytes, which are important during regeneration, synaptic plasticity and reactive gliosis.58 Astrocytes can also modulate the process of sleep homeostasis through the release of adenosine and pro-inflammatory cytokines, and by clearing toxic substances through the glymphatic system.59 Patients with chronic insomnia disorder exhibited increased GFAP serum levels, which could indicate some degree of astrocyte damage.60 Mice with chronic sleep deprivation showed low-grade neuroinflammation, anxiogenic response and recognition memory impairment, accompanied by enhanced expression of GFAP in brain regions.61 Consistent with these evidences, expression level of GFAP was negatively correlated with sleep health score in our PWAS results. We also identified genes to be associated with sleep health score at the mRNA level. ORC4 gene encodes a protein essential for the initiation of the DNA replication in eukaryotic cells. Mutations in ORC4 are linked to Meier-Gorlin syndrome, which is frequently associated with short stature and microcephaly,62 suggesting that this gene may play an important role in normal brain development.
We also compared the TWAS results on sleep health score with previously published TWAS results on sleep duration22 and insomnia.23 Previous TWAS studies identified 38 genes for which sleep duration SNPs influenced gene expression in 11 tissues,22 and 24 genes for which insomnia SNPs influenced gene expression in one or more of the 14 tissues.23 However, none of the sleep duration and insomnia genes previously identified by TWAS were replicable in our sleep health score TWAS results. We identified 2 novel genes for which sleep health score SNPs influenced gene expression in brain tissue. Considering PWAS, we reported the first PWAS research of sleep traits to date.
Negative genetic correlations between sleep health score and a variety of sleep behaviors (insomnia, snoring and dozing) were identified, in which the highest genetic correlation with sleep health score is insomnia. We also found negative genetic correlation evidences between sleep health score and four psychiatry disorders (MDD, ADHD, schizophrenia and ASD), which were consistent with previous studied.63,64 Depression is the most closely genetic related to sleep health score among the four psychiatry disorders. Most depressed patients suffer from sleep abnormalities.57 A cohort study suggested that reduced quantity of sleep increases risk for major depression, which in turn increases risk for decreased sleep.64 There is a strong bidirectional relationship between sleep alterations and depression. Abnormal sleep also accompanies many other psychiatric disorders, such as ADHD. ADHD and sleep problems may interact each other and possible involvement of comorbidity.63
We also observed genetic correlation evidence with nine plasma protein. For example, Stabilin-1, Stromelysin-2 and Cytochrome c showed negative correlations with sleep health score in this study. Cytochrome c oxidase (COX) is necessary for aerobic ATP production by maintaining the proton gradient across the inner mitochondrial membrane. Consistent with our results, higher levels of COX have previously been identified in long-term deprivation relative to sleep.65 Stromelysin-2 is a member of the peptidase M10 family of matrix metalloproteinases (MMPs). MMPs could degrade amyloid precursor protein leading to aggregation of amyloid β.66 Sleep disorders are frequent in Alzheimer’s disease (AD). And previous evidence showed that sleep disturbance might increase the risk of AD by increasing β-amyloid burden.67 Stabilin-1 may function in initiation of inflammation and receptor scavenging as one of the naturally occurring antibodies targeting receptors of the innate immune system, with links to AD.68 Our results might provide new opportunities to the understanding of highly related sleeping behaviors, as well as its relationship with psychiatric disorders and other traits.
To our knowledge, we reported the first genetic investigation of overall sleep health score to date. The overall sleep health score we used integrated several sleep behaviors (sleep duration, chronotype, snoring, daytime dozing and insomnia), and could more powerfully represents overall sleep traits according to the previous study.30 Sleep health score provides a positive frame of reference for sleep, and is also of value in promoting epidemiological findings easier to be interpreted and translated into practice. The overall measure of sleep health in genetic studies may find novel information than directly assessing individual traits. Our results estimated with LDSC suggested significant negative genetic correlations between sleep health score and sleep behaviors (insomnia, snoring and dozing), which could also provide support for the representativeness of the sleep health score. In addition, this study examined mRNA and protein levels associated with sleep health score through PWAS and TWAS.
However, some limitations must be acknowledged. First, our analysis was limited to European ancestries only to avoid population stratification confounding, it should be careful to apply our results to other ethnic groups. Second, analyses used self-reported sleep data from the UK biobank, misclassification of exposures was inevitable Third, we dichotomized five sleep factors to create a combination for a sleep health measure for simplicity, which might lead to the loss of nuance of sleep information and the loss of study power. Forth, UK Biobank is a high quality data resource that allows the use of data on self-reported sleep behavior with a large population size. The comprehensive dataset also allows adjustment for a wide range of confounders including gender, age and the first ten principal components of the genome. However, the study is also subject to limitations. Our baseline sleep behavioral variables were assessed through a self-report method, thus there remains the possibility of residual confounding due to recall and social desirability biases. Fifth, the objective of this study is to explore the genetic architecture of the overall sleep health score, further functional studies are still needed to confirm our finding. Sixth, we were unable to identify any other publicly available cohorts with sufficiently large numbers to allow an adequately powered second group for replication analysis. Confirmation in other independent study samples will be of great interest to replicate our findings.
Conclusion
To conclude, genetic variations in 32 loci are related to sleep health score. Some of the genes showing significant associations with sleep health score were also identified in previous studies of individual sleep behaviors. We identified genes that contribute to sleep health score at the protein level and the mRNA level by PWAS and TWAS. We identified 2 potentially causal genes (HDHD2 and GFAP) that act via modulating their brain protein abundances, in which the GFAP gene plays a role at the mature astrocytes. We further found evidence of genetic correlation between sleep health score and other sleep behaviors, psychiatric traits and plasma protein. Our results advance the understanding of the aetiology and genetic architecture of the sleep health score, refine the understanding of the relationship of sleep health score with sleep behaviors and other traits, and may serve as potential targets for future mechanistic studies of sleep phenotype.
Data Sharing Statement
The UK Biobank data are available through the UK Biobank Access Management System https://www.ukbiobank.ac.uk/.
Ethical Statement
There is no ethical statement.
Acknowledgments
This research is conducted under UK Biobank application 46478.
Funding
This work was supported by the National Natural Scientific Foundation of China (grant number 81922059, 81972980, 81703177).
Disclosure
All authors declare that they have no competing interest.
References
1. Rasch B, Born J. About sleep’s role in memory. Physiol Rev. 2013;93(2):681–766. doi:10.1152/physrev.00032.2012
2. Sateia MJ. International classification of sleep disorders-third edition: highlights and modifications. Chest. 2014;146(5):1387–1394. doi:10.1378/chest.14-0970
3. Besedovsky L, Lange T, Haack M. The sleep-immune crosstalk in health and disease. Physiol Rev. 2019;99(3):1325–1380. doi:10.1152/physrev.00010.2018
4. Carpena MX, Bonilla C, Matijasevich A, et al. Sleep-related traits and attention-deficit/hyperactivity disorder comorbidity: shared genetic risk factors, molecular mechanisms, and causal effects. World J Biol Psychiatry. 2021:1–39. doi:10.1080/15622975.2021.1907719
5. Kivelä L, Papadopoulos MR, Antypa N. Chronotype and psychiatric disorders. Curr Sleep Med Rep. 2018;4(2):94–103. doi:10.1007/s40675-018-0113-8
6. Wintler T, Schoch H, Frank MG, Peixoto L. Sleep, brain development, and autism spectrum disorders: insights from animal models. J Neurosci Res. 2020;98(6):1137–1149. doi:10.1002/jnr.24619
7. Daghlas I, Lane JM, Saxena R, Vetter C. Genetically proxied diurnal preference, sleep timing, and risk of major depressive disorder. JAMA Psychiatry. 2021;78(8):903–910. doi:10.1001/jamapsychiatry.2021.0959
8. Goldstein AN, Walker MP. The role of sleep in emotional brain function. Annu Rev Clin Psychol. 2014;10:679–708. doi:10.1146/annurev-clinpsy-032813-153716
9. Riemann D, Krone LB, Wulff K, Nissen C. Sleep, insomnia, and depression. Neuropsychopharmacology. 2020;45(1):74–89. doi:10.1038/s41386-019-0411-y
10. Troynikov O, Watson CG, Nawaz N. Sleep environments and sleep physiology: a review. J Therm Biol. 2018;78:192–203. doi:10.1016/j.jtherbio.2018.09.012
11. Lopez-Minguez J, Morosoli JJ, Madrid JA, Garaulet M, Ordoñana JR. Heritability of siesta and night-time sleep as continuously assessed by a circadian-related integrated measure. Sci Rep. 2017;7(1):12340. doi:10.1038/s41598-017-12460-x
12. Kocevska D, Barclay NL, Bramer WM, Gehrman PR, Van Someren EJW. Heritability of sleep duration and quality: a systematic review and meta-analysis. Sleep Med Rev. 2021;59:101448. doi:10.1016/j.smrv.2021.101448
13. Leocadio-Miguel MA, Ruiz FS, Ahmed SS, et al. Compared heritability of chronotype instruments in a single population sample. J Biol Rhythms. 2021;36(5):483–490. doi:10.1177/07487304211030420
14. Madrid-Valero JJ, Rubio-Aparicio M, Gregory AM, Sánchez-Meca J, Ordoñana JR. The heritability of insomnia: systematic review and meta-analysis of twin studies. Sleep Med Rev. 2021;58:101437. doi:10.1016/j.smrv.2021.101437
15. Campos AI, García-Marín LM, Byrne EM, Martin NG, Cuéllar-Partida G, Rentería ME. Insights into the aetiology of snoring from observational and genetic investigations in the UK biobank. Nat Commun. 2020;11(1):817. doi:10.1038/s41467-020-14625-1
16. Dashti HS, Daghlas I, Lane JM, et al. Genetic determinants of daytime napping and effects on cardiometabolic health. Nat Commun. 2021;12(1):900. doi:10.1038/s41467-020-20585-3
17. Jones SE, van Hees VT, Mazzotti DR, et al. Genetic studies of accelerometer-based sleep measures yield new insights into human sleep behaviour. Nat Commun. 2019;10(1):1585. doi:10.1038/s41467-019-09576-1
18. Jones SE, Lane JM, Wood AR, et al. Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms. Nat Commun. 2019;10(1):343. doi:10.1038/s41467-018-08259-7
19. Jansen PR, Watanabe K, Stringer S, et al. Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nat Genet. 2019;51(3):394–403. doi:10.1038/s41588-018-0333-3
20. Möller-Levet CS, Archer SN, Bucca G, et al. Effects of insufficient sleep on circadian rhythmicity and expression amplitude of the human blood transcriptome. Proc Natl Acad Sci U S A. 2013;110(12):E1132–E1141. doi:10.1073/pnas.1217154110
21. Noya SB, Colameo D, Brüning F, et al. The forebrain synaptic transcriptome is organized by clocks but its proteome is driven by sleep. Science. 2019;366(6462):6462. doi:10.1126/science.aav2642
22. Dashti HS, Jones SE, Wood AR, et al. Genome-wide association study identifies genetic loci for self-reported habitual sleep duration supported by accelerometer-derived estimates. Nat Commun. 2019;10(1):1100. doi:10.1038/s41467-019-08917-4
23. Lane JM, Jones SE, Dashti HS, et al. Biological and clinical insights from genetics of insomnia symptoms. Nat Genet. 2019;51(3):387–393. doi:10.1038/s41588-019-0361-7
24. Chen Y, Kartsonaki C, Clarke R, et al. Characteristics and correlates of sleep duration, daytime napping, snoring and insomnia symptoms among 0.5 million Chinese men and women. Sleep Med. 2018;44:67–75. doi:10.1016/j.sleep.2017.11.1131
25. Roepke SE, Duffy JF. Differential impact of chronotype on weekday and weekend sleep timing and duration. Nat Sci Sleep. 2010;2010(2):213–220. doi:10.2147/nss.S12572
26. Buysse DJ, Reynolds CF
27. Antunes BM, Campos EZ, Parmezzani SS, Santos RV, Franchini E, Lira FS. Sleep quality and duration are associated with performance in maximal incremental test. Physiol Behav. 2017;177:252–256. doi:10.1016/j.physbeh.2017.05.014
28. Brandes N, Linial N, Linial M. PWAS: proteome-wide association study-linking genes and phenotypes by functional variation in proteins. Genome Biol. 2020;21(1):173. doi:10.1186/s13059-020-02089-x
29. Wainberg M, Sinnott-Armstrong N, Mancuso N, et al. Opportunities and challenges for transcriptome-wide association studies. Nat Genet. 2019;51(4):592–599. doi:10.1038/s41588-019-0385-z
30. Fan M, Sun D, Zhou T, et al. Sleep patterns, genetic susceptibility, and incident cardiovascular disease: a prospective study of 385 292 UK biobank participants. Eur Heart J. 2020;41(11):1182–1189. doi:10.1093/eurheartj/ehz849
31. Li ZH, Zhang PD, Chen Q, et al. Association of sleep and circadian patterns and genetic risk with incident type 2 diabetes: a large prospective population-based cohort study. Eur J Endocrinol. 2021;185(5):765–774. doi:10.1530/eje-21-0314
32. Wang M, Zhou T, Li X, et al. Baseline vitamin D status, sleep patterns, and the risk of incident type 2 diabetes in data from the UK Biobank Study. Diabetes Care. 2020;43(11):2776–2784. doi:10.2337/dc20-1109
33. Bycroft C, Freeman C, Petkova D, et al. The UK biobank resource with deep phenotyping and genomic data. Nature. 2018;562(7726):203–209. doi:10.1038/s41586-018-0579-z
34. Purcell S, Neale B, Todd-Brown K, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81(3):559–575. doi:10.1086/519795
35. Gusev A, Ko A, Shi H, et al. Integrative approaches for large-scale transcriptome-wide association studies. Nat Genet. 2016;48(3):245–252. doi:10.1038/ng.3506
36. Beach TG, Adler CH, Sue LI, et al. Arizona study of aging and neurodegenerative disorders and brain and body donation program. Neuropathology. 2015;35(4):354–389. doi:10.1111/neup.12189
37. Wingo AP, Liu Y, Gerasimov ES, et al. Integrating human brain proteomes with genome-wide association data implicates new proteins in alzheimer’s disease pathogenesis. Nat Genet. 2021;53(2):143–146. doi:10.1038/s41588-020-00773-z
38. Zhou X, Carbonetto P, Stephens M, Visscher PM. Polygenic modeling with bayesian sparse linear mixed models. PLoS Genet. 2013;9(2):e1003264. doi:10.1371/journal.pgen.1003264
39. Bulik-Sullivan B, Finucane HK, Anttila V, et al. An atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47(11):1236–1241. doi:10.1038/ng.3406
40. Stahl EA, Breen G, Forstner AJ, et al. Genome-wide association study identifies 30 loci associated with bipolar disorder. Nat Genet. 2019;51(5):793–803. doi:10.1038/s41588-019-0397-8
41. Wray NR, Ripke S, Mattheisen M, et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat Genet. 2018;50(5):668–681. doi:10.1038/s41588-018-0090-3
42. Demontis D, Walters RK, Martin J, et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat Genet. 2019;51(1):63–75. doi:10.1038/s41588-018-0269-7
43. Pardiñas AF, Holmans P, Pocklington AJ, et al. Common schizophrenia alleles are enriched in mutation-intolerant genes and in regions under strong background selection. Nat Genet. 2018;50(3):381–389. doi:10.1038/s41588-018-0059-2
44. Grove J, Ripke S, Als TD, et al. Identification of common genetic risk variants for autism spectrum disorder. Nat Genet. 2019;51(3):431–444. doi:10.1038/s41588-019-0344-8
45. Sun BB, Maranville JC, Peters JE, et al. Genomic atlas of the human plasma proteome. Nature. 2018;558(7708):73–79. doi:10.1038/s41586-018-0175-2
46. Hammerschlag AR, Stringer S, de Leeuw CA, et al. Genome-wide association analysis of insomnia complaints identifies risk genes and genetic overlap with psychiatric and metabolic traits. Nat Genet. 2017;49(11):1584–1592. doi:10.1038/ng.3888
47. Sarayloo F, Dion PA, Rouleau GA. MEIS1 and restless legs syndrome: a comprehensive review. Front Neurol. 2019;10:935. doi:10.3389/fneur.2019.00935
48. Venkateshiah SB, Ioachimescu OC. Restless legs syndrome. Crit Care Clin. 2015;31(3):459–472. doi:10.1016/j.ccc.2015.03.003
49. Lane JM, Vlasac I, Anderson SG, et al. Genome-wide association analysis identifies novel loci for chronotype in 100,420 individuals from the UK biobank. Nat Commun. 2016;7:10889. doi:10.1038/ncomms10889
50. Kurien P, Hsu PK, Leon J, et al. TIMELESS mutation alters phase responsiveness and causes advanced sleep phase. Proc Natl Acad Sci U S A. 2019;116(24):12045–12053. doi:10.1073/pnas.1819110116
51. Kalmbach DA, Schneider LD, Cheung J, et al. Genetic basis of chronotype in humans: insights from three landmark GWAS. Sleep. 2017;40:2. doi:10.1093/sleep/zsw048
52. Diniz LP, Matias I, Siqueira M, Stipursky J, Gomes FCA. Astrocytes and the TGF-β1 pathway in the healthy and diseased brain: a double-edged sword. Mol Neurobiol. 2019;56(7):4653–4679. doi:10.1007/s12035-018-1396-y
53. Najas S, Pijuan I, Esteve-Codina A, et al. A SMAD1/5-YAP signalling module drives radial glia self-amplification and growth of the developing cerebral cortex. Development. 2020;147(13). doi:10.1242/dev.187005
54. Vodopiutz J, Schmook MT, Konstantopoulou V, et al. MED20 mutation associated with infantile basal ganglia degeneration and brain atrophy. Eur J Pediatr. 2015;174(1):113–118. doi:10.1007/s00431-014-2463-7
55. Choi JE, Lee JJ, Kang W, et al. Proteomic analysis of hippocampus in a mouse model of depression reveals neuroprotective function of Ubiquitin C-terminal Hydrolase L1 (UCH-L1) via stress-induced cysteine oxidative modifications. Mol Cell Proteomics. 2018;17(9):1803–1823. doi:10.1074/mcp.RA118.000835
56. Cui L, Wang F, Yin Z, et al. Effects of the LHPP gene polymorphism on the functional and structural changes of gray matter in major depressive disorder. Quant Imaging Med Surg. 2020;10(1):257–268. doi:10.21037/qims.2019.12.01
57. Wang YQ, Li R, Zhang MQ, Zhang Z, Qu WM, Huang ZL. The neurobiological mechanisms and treatments of REM sleep disturbances in depression. Curr Neuropharmacol. 2015;13(4):543–553. doi:10.2174/1570159x13666150310002540
58. Middeldorp J, Hol EM. GFAP in health and disease. Prog Neurobiol. 2011;93(3):421–443. doi:10.1016/j.pneurobio.2011.01.005
59. Haydon PG. Astrocytes and the modulation of sleep. Curr Opin Neurobiol. 2017;44:28–33. doi:10.1016/j.conb.2017.02.008
60. Zhang P, Li Y-X, Zhang -Z-Z, et al. Astroglial mechanisms underlying chronic insomnia disorder: a Clinical Study. Nat Sci Sleep. 2020;12:693–704. doi:10.2147/NSS.S263528
61. Manchanda S, Singh H, Kaur T, Kaur G. Low-grade neuroinflammation due to chronic sleep deprivation results in anxiety and learning and memory impairments. Mol Cell Biochem. 2018;449(1–2):63–72. doi:10.1007/s11010-018-3343-7
62. de Munnik SA, Hoefsloot EH, Roukema J, et al. Meier-Gorlin syndrome. Orphanet J Rare Dis. 2015;10:114. doi:10.1186/s13023-015-0322-x
63. Hvolby A. Associations of sleep disturbance with ADHD: implications for treatment. Atten Defic Hyperact Disord. 2015;7(1):1–18. doi:10.1007/s12402-014-0151-0
64. Roberts RE, Duong HT. The prospective association between sleep deprivation and depression among adolescents. Sleep. 2014;37(2):239–244. doi:10.5665/sleep.3388
65. Cirelli C, Pfister-Genskow M, McCarthy D, Woodbury R, Tononi G. Proteomic profiling of the rat cerebral cortex in sleep and waking. Arch Ital Biol. 2009;147(3):59–68.
66. Horstmann S, Budig L, Gardner H, et al. Matrix metalloproteinases in peripheral blood and cerebrospinal fluid in patients with Alzheimer’s disease. Int Psychogeriatr. 2010;22(6):966–972. doi:10.1017/S1041610210000827
67. Irwin MR, Vitiello MV. Implications of sleep disturbance and inflammation for Alzheimer’s disease dementia. Lancet Neurol. 2019;18(3):296–306. doi:10.1016/s1474-4422(18)30450-2
68. Giil LM, Vedeler CA, Kristoffersen EK, et al. Antibodies to signaling molecules and receptors in Alzheimer’s disease are associated with psychomotor slowing, depression, and poor visuospatial function. J Alzheimers Dis. 2017;59(3):929–939. doi:10.3233/jad-170245
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.