Back to Journals » Journal of Pain Research » Volume 19
Gender-Specific Behavior Reductions in Spontaneous Locomotive and Rearing in a Rat Model of Paclitaxel-Induced Peripheral Neuropathy: A Pilot Study
Authors Kim SJ ![]() , Cao K
, Cao K ![]() , Dougherty PM, McDonough KE, Javed S
, Dougherty PM, McDonough KE, Javed S
Received 13 September 2025
Accepted for publication 10 March 2026
Published 16 May 2026 Volume 2026:19 565512
DOI https://doi.org/10.2147/JPR.S565512
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Serena Jiyeon Kim,1,* Kevin Cao,2,* Patrick M Dougherty,1 Kathleen E McDonough,1 Saba Javed1
1Department of Pain Medicine, Division of Anesthesiology, Critical Care & Pain Medicine, The University of Texas MD Anderson Cancer Center, Houston, TX, 77030, USA; 2Department of Medicine, Baylor College of Medicine, Houston, TX, 77030, USA
*These authors contributed equally to this paper
Correspondence: Serena Jiyeon Kim, Department of Pain Medicine, Division of Anesthesiology, Critical Care & Pain Medicine, The University of Texas MD Anderson Cancer Center, Houston, TX, 77030, USA, Tel +1 832-581-1432, Email [email protected]
Introduction: Chemotherapy-induced peripheral neuropathy (CIPN) from agents like paclitaxel causes sensory loss and functional impairment, with limited treatment options. To better model patient-relevant outcomes, we used open field video tracking in rodents to assess gait, anxiety, and exploratory behavior. This approach provides a more comprehensive assessment of CIPN-related deficits beyond pain responses.
Methods and Materials: Adult male and female Sprague Dawley rats were given paclitaxel (Taxol; 24 mg/kg cumulative dose) or a vehicle over 8 days. Their distance traveled, time spent in the center and outer zones of the OF test, and rearing behaviors were recorded using a PhenoTyper 4500 OF system before and after treatment. Behavioral outcomes were analyzed using 2-way repeated measures analysis of variance (ANOVA) with significance set at p < 0.05.
Results: Paclitaxel treatment significantly reduced locomotor activity and rearing behavior in rats, reflecting functional impairments relevant to human CIPN. A three-way ANOVA revealed sex-specific effects, prompting separate analyses by sex. Taxol-treated males showed marked decreases in both distance traveled and rearing, while females exhibited reductions in locomotion but not rearing. No consistent changes were observed in anxiety-like behavior, highlighting the selective impact of paclitaxel on motor and exploratory functions.
Discussion/Conclusion: This pilot study demonstrated that paclitaxel-treated rats exhibited significant reductions in spontaneous behaviors such as distance traveled and rearing activity. These ethologically relevant behavioral measures reflect the clinical symptoms of CIPN in humans and highlight the utility of OF activity as an appropriate method for tracking ongoing pain and treatment efficacy in preclinical models of PIPN.
Keywords: chemotherapy-induced peripheral neuropathy, paclitaxel, open field test, locomotor activity, rearing behavior, spontaneous pain behavior, behavioral tracking
Introduction
Chemotherapy-induced peripheral neuropathy (CIPN) is a dose-limiting toxicity of various chemotherapy agents, characterized by distal symmetric sensory loss, neuropathic pain, and functional impairment in a predominantly stocking-and-glove distribution.1,2 Prevalence of CIPN among patients receiving neurotoxic chemotherapy ranges from 19% to over 85%. Furthermore, 68% of cases occur within the first month of starting chemotherapy, 60% occur after 3 months, and 30% occur at 6 months or longer.3–5 About 30% of patients with CIPN need dose reduction, treatment delay, or treatment discontinuation because of intolerable neuropathy or functional impairment during neurotoxic chemotherapy.6,7 Duloxetine is the only agent recommended by the American Society of Clinical Oncology that has shown limited efficacy in treating painful CIPN. With no approved agent for prevention, dose modification remains the primary strategy to reduce neurotoxicity.8–10 However, this approach increases overall morbidity by inadequately controlling neuropathic symptoms and by risking inferior cancer outcomes.5,11
Taxanes, particularly paclitaxel, are strongly associated with chemotherapy-induced peripheral neuropathy (CIPN), affecting up to 60–70% of treated patients.5,12,13 Axonal degeneration, loss of intraepidermal nerve fibers, mitochondrial dysfunction, oxidative stress, and neuroinflammation, all contribute to the development of paclitaxel-induced peripheral neuropathy (PIPN).14–17 PIPN in humans results in clinically significant sensory and motor impairments, including reduced vibration and proprioception, diminished light touch sensation, balance disturbances, slower gait speed, difficulty with fine motor tasks, and an increased risk of falls.
Current rodent models of PIPN exhibit clinical and molecular features of human PIPN: mechanical allodynia, sensory loss, and intraepidermal nerve fiber degeneration.16,18–20 These models primarily rely on behavioral assays such as von Frey filament testing for mechanical hypersensitivity, the acetone test for cold allodynia, and histological analyses of intraepidermal nerve fiber density and dorsal root ganglia (DRG) pathology.5,21–23
However, a critical gap remains in assays capturing functional impairments observed in patients, specifically, gait disturbances, activity avoidance, and other neurobehavioral changes, limiting translational relevance. To address this translational gap, in the present pilot study, we used open field (OF) video tracking to capture these phenotypic changes in rodents, allowing us to quantitatively analyze locomotor activity (distance travelled), anxiety-related behavioral changes (time spent in center vs. outer zone of the OF test), and exploratory/motivational behavior (rearing). These behaviors are analogous to human functional deficits: reduced locomotion reflects slower gait and mobility limitations, avoidance of open areas reflects cautious movement or fall risk, and decreased rearing mirrors reduced engagement in daily activities. This approach expands the behavioral phenotype beyond reflexive pain responses, providing a preclinical model of PIPN in evaluation of potential therapeutic interventions targeting these relevant functional deficits. Our data supports the use of OF video tracking as a relatively simple, objective method to assess CIPN-related functional deficits.
Materials and Methods
Ethical Approval and Animal Welfare
All experimental animal protocols were approved by the Institutional Animal Care and Use Committee at The University of Texas M.D. Anderson Cancer Center, and were used in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. The procedures employed in this pilot study were designed to minimize discomfort to the animals and use the minimum number of animals needed for statistical analysis.
Paclitaxel Administration
Adult male and female Sprague Dawley rats that were 8–10 weeks of age were used in our behavioral experiments. Pharmaceutical-grade paclitaxel (Taxol; Teva Pharmaceuticals Industries, Tel Aviv, Israel) was diluted with sterile saline from the original stock concentration of 6 mg/mL (in 1:1 Cremophor EL: ethanol) and administered at a dosage of 6 mg/kg intraperitoneally every other day for a total of 4 injections, resulting in a final cumulative dose of 24 mg/kg. Control animals received an equivalent volume of a vehicle consisting only of equal amounts of Cremophor EL and ethanol diluted with saline at the same concentration as that vehicle in the paclitaxel solution.
Behavioral Test
Animal behavior was recorded using a PhenoTyper 4500 OF activity box (Noldus Information Technology, Leesburg, VA), which uses EthoVision video tracking software for the capture and analysis of spontaneous behaviors (Figure 1). The Noldus PhenoTyper is a home-cage monitoring system that can reliably track prolonged, naturalistic rodent behavior with minimal observer interference.24
 |
Figure 1 Noldus PhenoTyper [KM1] PT4500 open-field activity box. |
Behavioral studies were blinded. Specifically, the investigator doing the open field tests did not know which animals were vehicle treated, and which were paclitaxel treated at the time of testing. However, it is important to note that one of the benefits of using this open field behavioral testing is that blinding is not completely necessary, as all behavior is recorded and interpreted by the computer software, and thus is not dependent on subjective judgements by the investigator in the same way as other behavioral tests such as vonFrey.
Day 8 was chosen as an endpoint, because at this day, peak behavioral responses to paclitaxel are known to be present based on previous studies. We chose not to go past this date both for ethical and humane treatment reasons, as well as to prevent the animals from becoming overly acclimated to the testing chambers, thus impacting results dependent on exploratory behaviors.
Each animal was placed in a separate chamber measuring 18.5 inches long x 18.5 inches wide x 26 inches high with a camera inserted into the top, directly above the center of the chamber to capture animal movements. The behaviors recorded in our pilot study included distance traveled, time spent in the center zone and the outer zones of the OF, and rearing. Behavioral recordings were completed for 30 minutes in a quiet room from 9:00 AM to 11:00 AM with the overhead room lights on. A baseline recording of animal activity in the OF box was taken on Day 0 (before paclitaxel administration), and post-treatment recording was taken on Day 8 (24 hours after the final paclitaxel injection) (Figure 2).
 |
Figure 2 Raw Open Field Tracking Data. |
Results
General Observations and Pilot Study Timeline
Sex Differences and Interaction Effects
The literature on the translation of behavioral outcomes from the rat OF test to human studies of CIPN is growing. Researchers have shown increasing validity of rats’ locomotor activity, as measured using the OF test, as a surrogate for human physical activity and mobility, exploratory behavior and motivation, and anxiety-like behavior, with new models and methodologies continuing to enhance its translational relevance.
Furthermore, sexually dimorphic responses in sensory neuron excitability and behavioral outcomes in rats, especially in locomotor activity and anxiety-like behaviors on OF following intraperitoneal paclitaxel administration, suggest that males and females exhibit differential neurobiological and behavioral adaptations to CIPN.25–27 To determine the effect of paclitaxel on locomotor activity and of sex on OF test results in our pilot study animals, we performed 3-way ANOVA to assess the effect of time, treatment, sex, and their interaction on individual OF test parameters. This test revealed a significant main effect of sex on distance traveled (F(1,20) = 14; P = 0.0011), rearing (F(1,20) = 10; P = 0.0045), and time spent in the outer zone (F(1,20) = 8.8; P = 0.0077). Subsequently, we found a significant interaction of sex and treatment on distance travelled (F(1,20) = 7.2, P = 0.014) and rearing ((F(1,20) = 11, P = 0.0031) (Figure 3). Thus, we split up the male and female rats for subsequent analyses.
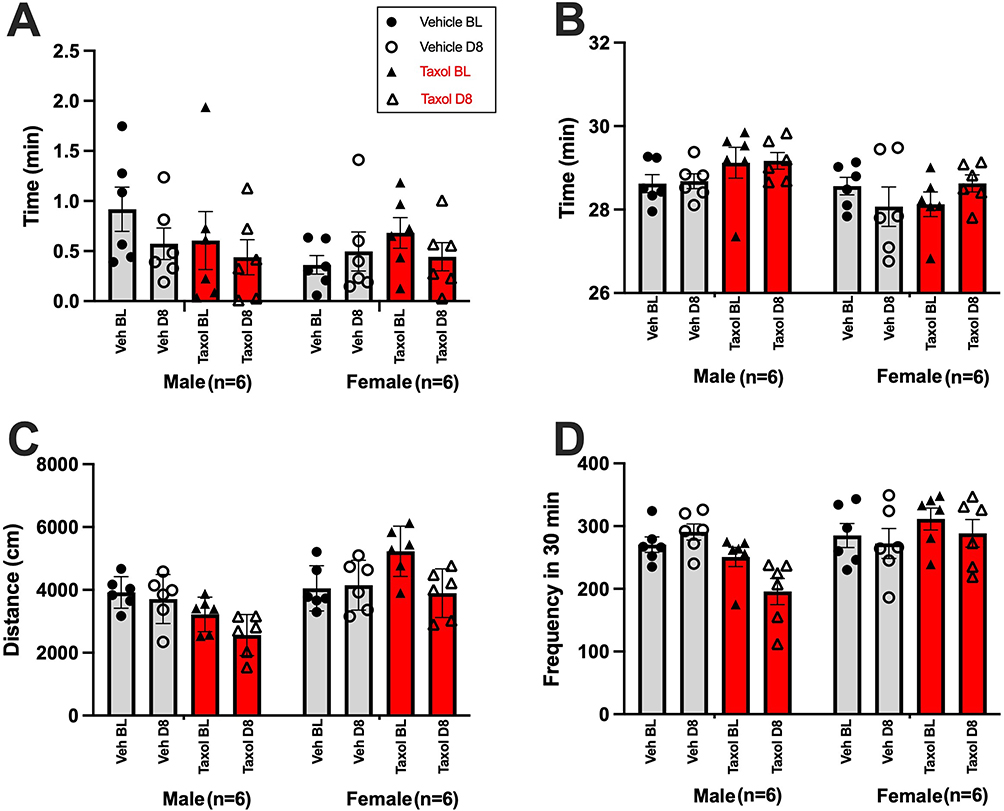 |
Figure 3 Mean ± SEM for (A) Center Zone: time spent in the center zone, (B)Outer Zone: time spent in the outer zone, (C) Distance: total distance traveled, and (D) Rearing: rearing frequency, combining both male and female groups. Behavioral data were analyzed using three-way ANOVAs with treatment (Veh vs. Taxol), time (BL vs. D8), and sex (male vs. female) as independent variables. |
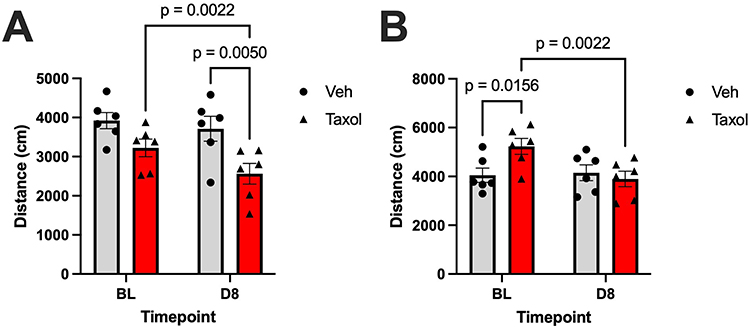 |
Figure 4 Mean distance traveled (± SEM) at baseline (BL) and day 8 (D8) following treatment with either vehicle (Veh) or paclitaxel (Taxol). (A) Distance Male: Distance travelled by male rats. Two-way anova reveals a significant main effect of time (F (1, 10) = 14, P = 0.0035), and treatment (F (1, 10) = 7.2, P = 0.023). (B) Distance Female: Distance travelled by female rats. Two-way anova reveals a significant main effect of time (F (1, 10) = 7.1, P = 0.024), and interaction of time x treatment (F (1, 10) = 9.7, P = 0.011). Post hoc pairwise comparisons (Fisher’s LSD test) are shown in the bar graphs. Sample size per group: n = 6. |
Effects of Paclitaxel on Locomotor Activity in the OF Test
Intraperitoneal administration of paclitaxel in rat models reliably induces chemotherapy-induced peripheral neuropathy (CIPN), which is characterized by the development of mechanical allodynia, cold hypersensitivity, and persistent sensory deficits.28,29 Previous studies demonstrated rats’ locomotor activity as measured in the OF test to be a valid surrogate for human functional impairments and decreased physical activity, reflecting both pain-related behavioral suppression and possible sensorimotor deficits.30
In male rats, 2-way ANOVA revealed significant main effects of time (F(1,10) = 14; P = 0.0035) and treatment (F(1,10) = 7.2; P = 0.023) on distance traveled. Post hoc analysis demonstrated that continuous injection of paclitaxel but not the vehicle significantly reduced locomotor activity as measured according to distance traveled before treatment (BL) compared with day 8 (t = 4.1; p = 0.0022). Moreover, paclitaxel-treated rats traveled markedly less than vehicle-treated rats did following 4 paclitaxel injections (t = 3.2; p = 0.0050), further confirming the negative effect of paclitaxel on locomotor activity.
We observed a similar effect in female rats, with 2-way ANOVA revealing a significant main effect of time (F(1,10) = 7.1; P = 0.024) and interaction of time and treatment (F(1,10) = 9.7; P = 0.011) on distance traveled (F(1,10) = 9.66; P = 0.0111). Indeed, continuous paclitaxel injection significantly reduced the distance traveled by female rats at day 8 when compared with baseline (t = 4.1; p = 0.0022), whereas vehicle injection did not cause any significant differences in the distance traveled between baseline and day 8.
Changes in Rearing Behavior Following Treatment with Paclitaxel
We have observed decreased rearing behavior in rat models following intraperitoneal administration of paclitaxel to induce CIPN. This reduction in rearing is interpreted as a marker of both functional impairment and pain-related behavioral suppression in the OF test. Specifically, paclitaxel-treated rats exhibit a significant decline in vertical exploratory activity (rearing), which is temporally associated with the onset and persistence of mechanical and cold hypersensitivity as well as ongoing pain-like behaviors.
In male rats, 2-way ANOVA revealed a significant main effect of treatment (F(1,10) = 11.25; P = 0.0073) and interaction of time and treatment (F(1,10) = 6.883; P = 0.025) on rearing frequency. Post hoc analysis demonstrated a significant decrease in rearing frequency in paclitaxel-treated rats from baseline to day 8 (t = 2.7; p = 0.022). Furthermore, paclitaxel-treated rats had markedly decreased rearing frequency compared with that for vehicle-treated rats following 4 paclitaxel injections (t = 4.3; p = 0.00040), further supporting paclitaxel’s effect of behavioral suppression.
In female rats, 2-way ANOVA revealed no significant main effects of either time or treatment on rearing frequency (all P > 0.5). As such, the interaction of time and treatment and the differences within and between the paclitaxel- and vehicle-treated groups was negligible.
The data reveal a clear sex-specific effect of paclitaxel on rearing behavior, suggesting that male rats are more susceptible to paclitaxel-induced behavioral suppression than female rats are. In male rats, the significant main effect of treatment and interaction of time and treatment demonstrate that treatment with paclitaxel not only reduced rearing frequency but also but did so progressively over time. The post-hoc findings further confirmed this, demonstrating both a within-group reduction in rearing frequency from baseline to day 8 and a between-group difference when compared with vehicle-treated controls. In contrast, we did not observe a significant effect of paclitaxel treatment on rearing frequency in paclitaxel-treated female rats.
These sex-specific trajectories suggest differential sensitivity of rats to paclitaxel’s behavioral effects and highlight the importance of sex as a biological variable in assessing neuropathy-related outcomes.
Time Spent in Inner and Outer Zones During OF Testing
Time spent in the inner versus outer zones during OF testing in rodent models of paclitaxel-induced CIPN is translationally significant because increased time in the outer zone and decreased time in the inner zone are robust indicators of anxiety-like and pain-suppressed behaviors, which are clinically relevant comorbidities in patients with CIPN.
Rodents naturally prefer the periphery (outer zone) of an OF owing to their innate aversion to open, brightly lit spaces; increased time in the outer zone and decreased time in the inner zone reflect heightened anxiety.31
In both male and female rats, 2-way ANOVA revealed a lack of statistically significant main effects of treatment or time, and interaction between time and treatment on time spent in the center zone of the OF test (all P > 0.05). These findings demonstrated that neither treatment with paclitaxel nor the passage of time influenced anxiety-related behavior, as measured according to center zone exploration.
We observed the same finding regarding time spent in the outer zone for male rats. Two-way ANOVA revealed a lack of statistically significant main effects of treatment or time, and interaction between time and treatment on time spent in the center zone of the OF test (all P > 0.05). In female rats, despite no significant main effects of time or treatment on zone activity, we detected a significant effect of interaction of time and treatment (F (1, 10) = 5.4, P = 0.042), indicating that the effect of treatment on behavior varied depending on the time point. Despite this interaction, post hoc analyses did not reveal any significant differences in specific group comparisons (P > 0.05), suggesting that the interaction effect reflects a subtle or trend-level pattern in zone activity not captured by individual pairwise contrasts.
Discussion
In this pilot exploratory study, we sought to characterize the toxic effects of PIPN using ethologically relevant measures and tracking neurobehavioral changes in a rat model following an 8 days of exposure to paclitaxel. Our findings demonstrate a significant decrease in distance traveled in both male and female rats and reduced rearing activity in male rats treated with paclitaxel (Figures 2–5). These spontaneous pain-related behavioral changes offer valuable insight into the persistent effects of paclitaxel-induced damage that extend beyond traditionally tested evoked pain responses. Therefore, these metrics may offer a more clinically relevant model for evaluation of CIPN, where patients report disruptions in voluntary movement and daily activity rather than only mechanical hypersensitivity or stimulus-evoked pain.
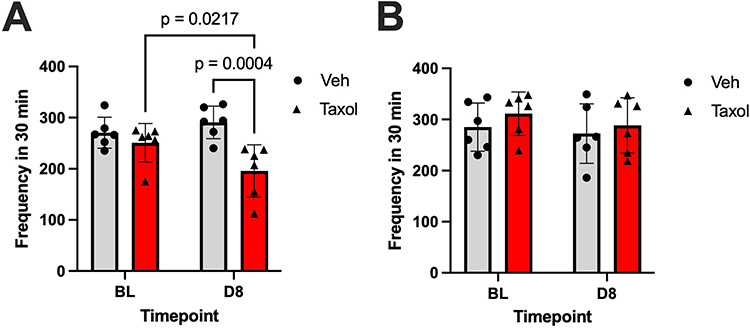 |
Figure 5 Mean rearing frequency (± SEM) at baseline (BL) and day 8 (D8) following treatment with either vehicle (Veh) or paclitaxel (Taxol). (A) Rearing – Frequency, Male. Two-way anova reveals a significant main effect of treatment (F (1, 10) = 11.25, P = 0.0073), and interaction of time x treatment (F (1, 10) = 6.883, P = 0.025). (B) Rearing – Frequency, Female. Post hoc pairwise comparisons (Fisher’s LSD test) are shown in the bar graphs. Sample size per group: n = 6. |
A key consideration in interpreting these findings is whether spontaneous pain-related behavioral alterations reflect motor impairment or sensory pain. Authors have reported a lack of significant motor dysfunction in rodent models, particularly following treatment with low-dose paclitaxel treatment as measured by using grip strength or rotarod performance tests.4,30,32 Moreover, the presence of damaged mitochondria in sensory neurons in the dorsal root ganglia but not in the motor neurons in the ventral root further suggests sensory impairment as the etiology of decreased voluntary movement in rats with PIPN.32 In clinical settings, patients with CIPN tend to exhibit sensory impairment (eg., pain, tingling, numbness) more prominently than motor dysfunction, although at higher cumulative doses of paclitaxel some motor deficits have been observed.33–35 Hence, the reduction in spontaneous activities such as distance traveled and rearing in an OF test most likely mirrors the persistent pain rather than motor deficits. Therefore, these behavioral assessments may be useful in modeling the chronic, ongoing discomfort experienced by patients with CIPN.
The reduction in distance traveled and rearing behavior by our study animals may be explained by various cellular and molecular mechanisms of PIPN that were previously elucidated. Paclitaxel’s mechanism of action involves binding to and stabilizing microtubules which in turn disrupts normal assembly and disassembly of microtubules and subsequently axonal transport.36 Consequently, axonal degeneration occurs, particularly in long distal sensory axons.36 Other studies implicated mitochondrial dysfunction and reactive oxygen species as having roles in the development and maintenance of paclitaxel-induced pain by demonstrating a temporal correlation between mitochondrial dysfunction and pain behavior.21,37 Another widely acknowledged mechanism of PIPN is the increased neuroinflammation and glial activation in the dorsal root ganglia and spinal dorsal horn, which cause hypersensitivity and pain.38–41 Notably, blocking inflammatory mediators can attenuate CIPN symptoms and compensatory behaviors, further supporting these mechanisms.42
Studies have well established that time spent in the center zone and outer zone in the OF test is indicative of anxiety-like behavior. In other words, a rat that spends more time exploring the center zone displays less anxiety-like behavior than one that remains close to the edge of the OF (outer-zone).43,44 Despite not reaching statistical significance, paclitaxel-treated female rats showed a reduced amount of time spent in the center zone and a slightly increased amount in the periphery (Figure 6). Authors previously reported delayed onset of neurobehavioral side effects in mice which may explain the lack of significance in our current findings.45 Previous studies support the findings in the present pilot study, suggesting that there may be a correlation between paclitaxel administration and emotion regulation. For example, in a recent study by Pathak and Singh (2025), female rats exhibited heightened fear response and exaggerated anxious behaviors in OF tests after treatment with paclitaxel as demonstrated by their preference for safer behaviors and avoidance of risk-taking activity.28 In alignment with our findings, Toma et al (2017) and Liu et al (2022) also found that mice spent more time in outer zones than in the inner zones following paclitaxel administration.45,46 The link between psychopathological side effects of chemotherapeutic agents, including anxiety and depression, is well established. The exact pathophysiology underlying anxiety in CIPN cases is still under investigation and future studies with longer follow-up times are needed to more accurately evaluate the effects of paclitaxel on neurobehavior in rodent models.
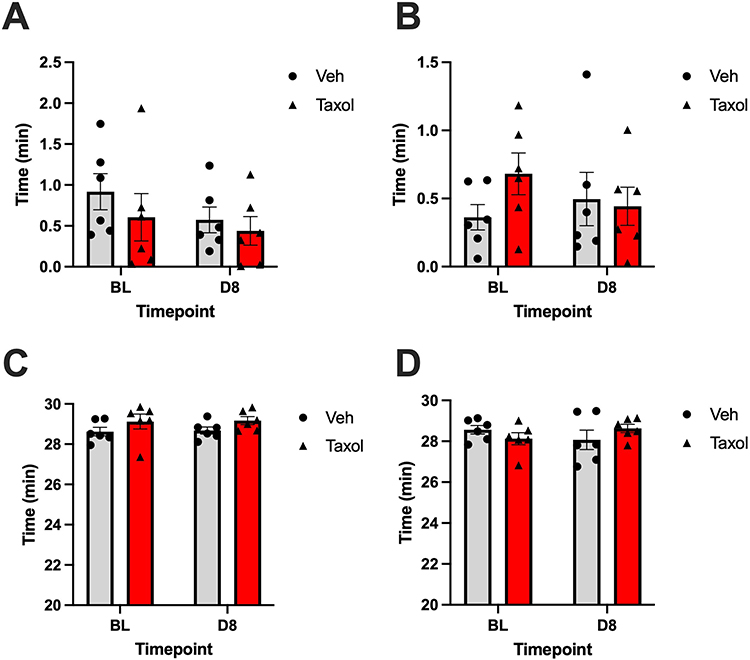 |
Figure 6 Mean zone activity (± SEM) at baseline (BL) and day 8 (D8) following treatment with either vehicle (Veh) or paclitaxel (Taxol). (A) Time Spent in Center Zone – Cumulative (min), Male. (B) Time Spent in Center Zone – Cumulative (min), Female. (C) Time Spent in Outer Zone – Cumulative (min), Male. (D) Time Spent in Outer Zone – Cumulative (min), Female. No significant post hoc pairwise comparisons (Fisher’s LSD test) were found. Sample size per group: n = 6. |
This pilot study had several limitations. Because of the small sample size, the findings should be interpreted conservatively, and their generalizability may be limited. When drawing conclusions using ANOVA results, the small sample size (n=6) poses challenges due to the inability to assess normality and homogeneity of variance assumptions. In future studies, it is necessary to increase the sample size due to the considerable variation in our current pilot study. Although OF activity provides valuable insight into spontaneous, ethologically relevant behaviors, we did not include conventional nociceptive assays such as the von Frey test in this pilot study. The absence of an evoked mechanical sensitivity measure limits our ability to directly confirm the development of paclitaxel-induced mechanical allodynia. Future studies will incorporate von Frey testing in parallel to further validate spontaneous behavioral changes against established CIPN phenotyping methods. Moreover, we observed significant behavioral changes, but we did not study the underlying mechanisms of these changes. Furthermore, given that we tracked the behavioral changes at only one time point, our understanding of longer term and cumulative paclitaxel exposure is limited.
In conclusion, we identified preliminary differences in gait parameters such as distance traveled and rearing behaviors according to sex in paclitaxel-treated rats. These findings indicate the importance of establishing objective, clinically relevant measures in preclinical models that accurately reflect the sustained pain and sensory deficits experienced by patients with CIPN. The findings support the potential feasibility of using an objective, accessible methodology with which to assess the progression or improvement of ongoing symptoms due to CIPN. Given the limitations, larger, adequately powered studies are needed to confirm these trends and explore whether such measures may ultimately contribute to more translationally meaningful assessments of CIPN. Moreover, we expect that behavioral changes would be improved by treatment of CIPN, and we plan to test this in later studies.
Data Sharing Statement
The data that support the findings of this study are available on request from the corresponding author.
Ethical Approval
All experimental animal protocols were approved by the Institutional Animal Care and Use Committee at The University of Texas M.D. Anderson Cancer Center and were used in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals.
Acknowledgments
We thank Donald R. Norwood, scientific editor, Research Medical Library, for editing this article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agreed to be accountable for all aspects of the work.
Funding
This study did not receive any funding.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Han Y, Smith MT. Pathobiology of cancer chemotherapy-induced peripheral neuropathy (CIPN). Front Pharmacol. 2013;4:4. doi:10.3389/fphar.2013.00156
2. Zajączkowska R, Kocot-Kępska M, Leppert W, Wrzosek A, Mika J, Wordliczek J. Mechanisms of chemotherapy-induced peripheral neuropathy. Int J Mol Sci. 2019;20(6):1451. doi:10.3390/ijms20061451
3. D’Souza RS, Saini C, Hussain N, Javed S, Prokop L, Her YF. Global estimates of prevalence of chronic painful neuropathy among patients with chemotherapy-induced peripheral neuropathy: systematic review and meta-analysis of data from 28 countries, 2000–24. Reg Anesth Pain Med. 2025;106229. doi:10.1136/rapm-2024-106229
4. Seretny M, Currie GL, Sena ES, et al. Incidence, prevalence, and predictors of chemotherapy-induced peripheral neuropathy: a systematic review and meta-analysis. Pain. 2014;155(12):2461–11. doi:10.1016/j.pain.2014.09.020
5. Staff NP, Grisold A, Grisold W, Windebank AJ. Chemotherapy-induced peripheral neuropathy: a current review. Ann Neurol. 2017;81(6):772–781. doi:10.1002/ana.24951
6. hertz DL, Dockter TJ, Satele DV, Loprinzi CL, Le-Rademacher J. Neuropathy severity at the time of oxaliplatin treatment alteration in patients with colon cancer (Alliance A151912). Support Care Cancer. 2021;29(12):7855–7863. doi:10.1007/s00520-021-06371-x
7. Nyrop KA, Deal AM, Reeder-Hayes KE, et al. Patient-reported and clinician-reported chemotherapy-induced peripheral neuropathy in patients with early breast cancer: current clinical practice. Cancer. 2019;125(17):2945–2954. doi:10.1002/cncr.32175
8. Desforges AD, Hebert CM, Spence AL, et al. Treatment and diagnosis of chemotherapy-induced peripheral neuropathy: an update. Biomed Pharmacother. 2022;147:112671. doi:10.1016/j.biopha.2022.112671
9. Fradkin M, Batash R, Elmaleh S, et al. Management of peripheral neuropathy induced by chemotherapy. Curr Med Chem. 2019;26(25):4698–4708. doi:10.2174/0929867326666190107163756
10. Loprinzi CL, Lacchetti C, Bleeker J, et al. Prevention and management of chemotherapy-induced peripheral neuropathy in survivors of adult cancers: asco guideline update. J Clin Oncol. 2020;38(28):3325–3348. doi:10.1200/JCO.20.01399
11. Miltenburg NC, Boogerd W. Chemotherapy-induced neuropathy: a comprehensive survey. Cancer Treat Rev. 2014;40(7):872–882. doi:10.1016/j.ctrv.2014.04.004
12. Da Costa R, Passos GF, Quintão NLM, et al. Taxane-induced neurotoxicity: pathophysiology and therapeutic perspectives. Br J Pharmacol. 2020;177(14):3127–3146. doi:10.1111/bph.15086
13. Park SB, Kwok JB, Asher R, et al. Clinical and genetic predictors of paclitaxel neurotoxicity based on patient- versus clinician-reported incidence and severity of neurotoxicity in the ICON7 trial. Ann Oncol. 2017;28(11):2733–2740. doi:10.1093/annonc/mdx491
14. Cirrincione AM, Pellegrini AD, Dominy JR, et al. Paclitaxel-induced peripheral neuropathy is caused by epidermal ROS and mitochondrial damage through conserved MMP-13 activation. Sci Rep. 2020;10(1):3970. doi:10.1038/s41598-020-60990-8
15. Gornstein EL, Schwarz TL. Neurotoxic mechanisms of paclitaxel are local to the distal axon and independent of transport defects. Exp Neurol. 2017;288:153–166. doi:10.1016/j.expneurol.2016.11.015
16. Hanna R, Graur A, Sinclair P, et al. Proteomic analysis of dorsal root ganglia in a mouse model of paclitaxel-induced neuropathic pain. Zhu X ed.. PLoS One. 2024;19(9):e0306498. doi:10.1371/journal.pone.0306498
17. Eun SGJ. Towards a mechanistic understanding of axon transport and endocytic changes underlying paclitaxel-induced peripheral neuropathy. Exp Neurol. 2023;359:114258. doi:10.1016/j.expneurol.2022.114258
18. Liu C, Berryman E, Zakur D, et al. A novel endpoint for the assessment of chemotherapy-induced peripheral neuropathy in rodents: biomechanical properties of peripheral nerve. J Appl Toxicol. 2018;38(2):193–200. doi:10.1002/jat.3513
19. Pennypacker SD, Fonseca MM, Morgan JW, et al. Methods and protocols for chemotherapy-induced peripheral neuropathy (CIPN) mouse models using paclitaxel. Methods in Cell Bio. 2022;168:277–298. doi:10.1016/bs.mcb.2021.12.019
20. Wozniak KM, Vornov JJ, Wu Y, et al. Peripheral neuropathy induced by microtubule-targeted chemotherapies: insights into acute injury and long-term recovery. Cancer Res. 2018;78(3):817–829. doi:10.1158/0008-5472.CAN-17-1467
21. Duggett NA, Griffiths LA, McKenna OE, et al. Oxidative stress in the development, maintenance and resolution of paclitaxel-induced painful neuropathy. Neuroscience. 2016;333:13–26. doi:10.1016/j.neuroscience.2016.06.050
22. Huehnchen P, Boehmerle W, Endres M. Assessment of paclitaxel induced sensory polyneuropathy with “catwalk” automated gait analysis in mice. PLoS One. 2013;8(10):e76772. doi:10.1371/journal.pone.0076772
23. Materazzi S, Fusi C, Benemei S, et al. TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pflüg Arch - Eur J Physiol. 2012;463(4):561–569. doi:10.1007/s00424-011-1071-x
24. Grieco F, Bernstein BJ, Biemans B, et al. Measuring behavior in the home cage: study design, applications, challenges, and perspectives. Front Behav Neurosci. 2021;15:735387. doi:10.3389/fnbeh.2021.735387
25. Ferrari LF, Araldi D, Green PG, Levine JD. Marked sexual dimorphism in neuroendocrine mechanisms for the exacerbation of paclitaxel-induced painful peripheral neuropathy by stress. Pain. 2020;161(4):865–874. doi:10.1097/j.pain.0000000000001798
26. Li H, Ward SJ. Paclitaxel-associated mechanical sensitivity and neuroinflammation are sex-, time-, and site-specific and prevented through cannabigerol administration in c57bl/6 mice. Int J Mol Sci. 2024;25(8):4277. doi:10.3390/ijms25084277
27. Villalba-Riquelme E, De La Torre-Martínez R, Fernández-Carvajal A, Ferrer-Montiel A. Paclitaxel in vitro reversibly sensitizes the excitability of IB4(−) and IB4(+) sensory neurons from male and female rats. Br J Pharmacol. 2022;179(14):3693–3710. doi:10.1111/bph.15809
28. Pathak D, Singh KP. Neurobehavioral manifestations in female rats after intermittent exposure to an anticancer agent, paclitaxel. Behav Pharmacol. 2025;36:276–289. doi:10.1097/FBP.0000000000000833
29. Sun W, Hao Y, Li R, et al. Comparative Transcriptome of Dorsal Root Ganglia Reveals Distinct Etiologies of Paclitaxel- and Oxaliplatin-induced Peripheral Neuropathy in Rats. Neuroscience. 2023;516:1–14. doi:10.1016/j.neuroscience.2023.02.007
30. Griffiths LA, Duggett NA, Pitcher AL, Flatters SJL. Evoked and ongoing pain-like behaviours in a rat model of paclitaxel-induced peripheral neuropathy. Pain Res Manag. 2018;2018:1–11. doi:10.1155/2018/8217613
31. Kraeuter AK, Guest PC, Sarnyai Z. the open field test for measuring locomotor activity and anxiety-like behavior. In: Guest PC, editor. Pre-Clinical Models. Vol 1916. Methods in Molecular Biology. Springer New York; 2019:99–103. doi:10.1007/978-1-4939-8994-2_9
32. Xiao WH, Zheng H, Zheng FY, Nuydens R, Meert TF, Bennett GJ. Mitochondrial abnormality in sensory, but not motor, axons in paclitaxel-evoked painful peripheral neuropathy in the rat. Neuroscience. 2011;199:461–469. doi:10.1016/j.neuroscience.2011.10.010
33. Augusto C, Pietro M, Cinzia M, et al. Peripheral neuropathy due to paclitaxel: study of the temporal relationships between the therapeutic schedule and the clinical quantitative score (QST) and comparison with neurophysiological findings. J Neurooncol. 2008;86(1):89–99. doi:10.1007/s11060-007-9438-8
34. Chen X, Gan Y, Au NPB, Ma CHE. Current understanding of the molecular mechanisms of chemotherapy-induced peripheral neuropathy. Front Mol Neurosci. 2024;17:1345811. doi:10.3389/fnmol.2024.1345811
35. Velasco R, Argyriou AA, Marco C, et al. Serum neurofilament levels correlate with electrodiagnostic evidence of axonal loss in paclitaxel-induced peripheral neurotoxicity. J Neurol. 2023;270(1):531–537. doi:10.1007/s00415-022-11377-4
36. Klein I, Lehmann H. Pathomechanisms of paclitaxel-induced peripheral neuropathy. Toxics. 2021;9(10):229. doi:10.3390/toxics9100229
37. Flatters SJL, Dougherty PM, Colvin LA. Clinical and preclinical perspectives on Chemotherapy-Induced Peripheral Neuropathy (CIPN): a narrative review. Br J Anaesth. 2017;119(4):737–749. doi:10.1093/bja/aex229
38. Li Y, Zhang H, Zhang H, Kosturakis AK, Jawad AB, Dougherty PM. Toll-like receptor 4 signaling contributes to paclitaxel-induced peripheral neuropathy. J Pain. 2014;15(7):712–725. doi:10.1016/j.jpain.2014.04.001
39. Zhang H, Boyette-Davis JA, Kosturakis AK, et al. Induction of Monocyte Chemoattractant Protein-1 (MCP-1) and Its Receptor CCR2 in Primary Sensory Neurons Contributes to Paclitaxel-Induced Peripheral Neuropathy. J Pain. 2013;14(10):1031–1044. doi:10.1016/j.jpain.2013.03.012
40. Zhang H, Li Y, De Carvalho-Barbosa M, et al. Dorsal root ganglion infiltration by macrophages contributes to paclitaxel chemotherapy-induced peripheral neuropathy. J Pain. 2016;17(7):775–786. doi:10.1016/j.jpain.2016.02.011
41. Zhang H, Yoon SY, Zhang H, Dougherty PM. Evidence that spinal astrocytes but not microglia contribute to the pathogenesis of paclitaxel-induced painful neuropathy. J Pain. 2012;13(3):293–303. doi:10.1016/j.jpain.2011.12.002
42. Al-Mazidi S, Alotaibi M, Nedjadi T, Chaudhary A, Alzoghaibi M, Djouhri L. Blocking of cytokines signalling attenuates evoked and spontaneous neuropathic pain behaviours in the paclitaxel rat model of chemotherapy-induced neuropathy. Eur J Pain. 2018;22(4):810–821. doi:10.1002/ejp.1169
43. Çevik ÖS, Yıldırım DD, Uzun C, Horata E. Contribution of distinctive outcome measures to the assessment of anxiety in the open field: a meta-analysis of factors mediating open-field test variability in rodent models of anxiety. Behav Brain Res. 2025;490:115612. doi:10.1016/j.bbr.2025.115612
44. Ennaceur A, Michalikova S, Chazot PL. Models of anxiety: responses of rats to novelty in an open space and an enclosed space. Behav Brain Res. 2006;171(1):26–49. doi:10.1016/j.bbr.2006.03.016
45. Toma W, Kyte SL, Bagdas D, et al. Effects of paclitaxel on the development of neuropathy and affective behaviors in the mouse. Neuropharmacology. 2017;117:305–315. doi:10.1016/j.neuropharm.2017.02.020
46. Liu J, Li D, Huang J, et al. Glutamatergic neurons in the amygdala are involved in paclitaxel-induced pain and anxiety. Front Psychiatry. 2022;13:869544. doi:10.3389/fpsyt.2022.869544
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.