Back to Journals » Journal of Hepatocellular Carcinoma » Volume 11
GDF15: Immunomodulatory Role in Hepatocellular Carcinoma Pathogenesis and Therapeutic Implications
Received 22 April 2024
Accepted for publication 7 June 2024
Published 19 June 2024 Volume 2024:11 Pages 1171—1183
DOI https://doi.org/10.2147/JHC.S471239
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr David Gerber
Yi-Ning Du,1 Jin-Wei Zhao2
1Department of Medical Sciences, Li Ka-shing School of Medicine, University of Hong Kong, Hong Kong, People’s Republic of China; 2Department of Hepatopancreatobiliary Surgery, Second Hospital of Jilin University, Jilin University, Changchun, Jilin Province, People’s Republic of China
Correspondence: Jin-Wei Zhao, Department of Hepatopancreatobiliary Surgery, Second Hospital of Jilin University, Jilin University, No. 218, Ziqiang Road, Changchun, 130000, Jilin Province, People’s Republic of China, Tel +8615844089385, Email [email protected]
Abstract: Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related deaths globally and the sixth most common cancer worldwide. Evidence shows that growth differentiation factor 15 (GDF15) contributes to hepatocarcinogenesis through various mechanisms. This paper reviews the latest insights into the role of GDF15 in the development of HCC, its role in the immune microenvironment of HCC, and its molecular mechanisms in metabolic dysfunction associated steatohepatitis (MASH) and metabolic associated fatty liver disease (MAFLD)-related HCC. Additionally, as a serum biomarker for HCC, diagnostic and prognostic value of GDF15 for HCC is summarized. The article elaborates on the immunological effects of GDF15, elucidating its effects on hepatic stellate cells (HSCs), liver fibrosis, as well as its role in HCC metastasis and tumor angiogenesis, and its interactions with anticancer drugs. Based on the impact of GDF15 on the immune response in HCC, future research should identify its signaling pathways, affected immune cells, and tumor microenvironment interactions. Clinical studies correlating GDF15 levels with patient outcomes can aid personalized treatment. Additionally, exploring GDF15-targeted therapies with immunotherapies could improve anti-tumor responses and patient outcomes.
Keywords: growth differentiation factor 15, GDF15, hepatocellular carcinoma, HCC, immune suppression, immunotherapy
Introduction
GDF15, also known as macrophage inhibitory factor-1 (MIC-1), prostate-derived factor (PDF), and others, is a member of the transforming growth factor (TGF) superfamily.1–3 It forms a pro-GDF15 homodimer in the endoplasmic reticulum (ER) through disulfide bond formation. After proteolytic secretion in the ER, mature homodimeric GDF15 is released from its propeptide.4
Other members of the GDF family, such as GDF1, GDF3, GDF5, GDF7, and GDF10 are predominantly regulators of cell growth and differentiation in embryonic and adult tissues, and even play a role in skeletal morphogenesis. Silencing of GDF1 expression significantly inhibited the GDF1-SMAD2/3 pathway, demonstrating the role of GDF1 in inhibiting gastric cancer progression.5 GDF3 regulates cell fate in stem cells and early embryos.6 GDF5 would be involved in the pathogenesis of osteoarthritis.7,8 GDF7 can promote the differentiation in vitro of two dorsal sensory interneuron classes.9
GDF15 was first identified from a cDNA library enriched for genes linked to macrophages.1 Paralkar et al discovered it using a probe for human transforming growth factor placental (hTGF-PL), naming it GDF15.10 Its role in cancer has been suggested, showing various functions mainly involved in regulating anti-inflammatory and apoptotic pathways.11
GFRAL is a specific receptor for GDF15, belonging to the GDNF family receptor α (GFR-α) group within the TGF-β superfamily.3 It’s mainly found in hindbrain neurons, exclusively serving as the receptor for GDF15 in the brainstem.12 To instigate intracellular signaling upon GDF15 activation, GFRAL necessitates binding to the co-receptor proto-oncogene tyrosine-protein kinase receptor RET, and signals through the extracellular-signal related kinase (ERK) and AKT/protein kinase B pathways, without activating canonical TGF-β signaling pathways.13,14 The disparity has been attributed to the presence of TGF-β impurities in several commercially available recombinant GDF15 preparations, considering that TGF-β may induce significant biological effects within the millimolar concentration range.15
HCC is the third most prominent contributor to cancer-associated mortality worldwide.16 Data based on prevalence and population attributable scores from different regions suggest that 44% of HCC cases worldwide were attributable to chronic hepatitis B infection, with the majority of cases occurring in Asia.17 Hepatitis C was responsible for 21% of cases.17 The development of HCC is largely determined by a variety of key risk factors, including alcohol, obesity, smoking, persistent infection with hepatitis B and C viruses, and the combined effects of MASH and MAFLD transformation.18,19
GDF15 is associated with obesity, aging, cardiovascular disease and progression of various cancers such as colorectal and ovarian cancers, and GDF15 can inhibit DC to promote immune escape from ovarian cancer.20–23 Recently, GDF15 was found to play an important role in immunosuppression in HCC.24 However, little is known about the interaction between GDF15 and immune cells in the tumor microenvironment.
Some findings showed that GDF15 expression was upregulated in HCC tissues, promoting HCC cell invasion and inhibiting HCC cell apoptosis through the TGF-β/Smad pathway.25–27 In response to the inconsistent findings, subsequent studies have generally concluded that GDF15 promotes HCC, but does not act by acting on the receptor for TGF-β.28 Overall, the inhibitory effect of GDF15 on HCC involves multiple signaling pathways, and the effect of GDF15 on promoting the development of HCC and immune escape proceeds by suppressing immune responses in the tumor microenvironment.
Role of GDF15 in Occurrence and Development of HCC
GDF15 exerts a substantial impact on various activities related to HCC, including proliferation, migration, invasion, angiogenesis, and other processes associated with carcinogenesis. Numerous recent studies have explored the involvement and underlying mechanisms of GDF15 in the carcinogenic process.
Dual Role of GDF15 in HCC Development and the Immune Microenvironment in HSC Fibrosis and MASH/MAFLD-HCC
Like other members of the GDNF receptor family, GFRAL interacts with its co-receptor RET and signals via the ERK and AKT/protein kinase B pathways.28 Xu et al discovered that reducing GDF15 levels in human liver cancer stem-like cells (LCSCs) inhibited the AKT pathway. Additionally, blocking the AKT/GSK-3β/β-catenin pathway reduced GDF15-driven proliferation, colony formation, and invasion in these cells.29 Benkhei et al found through microarray analysis that overexpression of GDF15 could up-regulate epidermal growth factor receptor (EFGR), a key cytokine in Hepatitis C Virus (HCV) induction of HCC, and Dong et al discovered that GDF15 activates VEGFR during the development of HCC.30,31
Yu et al found that ectopic expression of GDF15 in Hep3B cells (human HCC cell line) suppressed proliferation and induced apoptosis, thus demonstrating that GDF15 is a potential tumor suppressor gene.32 The experiment lacked exploration of inhibitory mechanisms. Then Suriben et al complemented GDF15 has been shown to translocate into mitochondria and induce ROS production and cell death in HCC cells.33
GDF15, a key protein induced by the tumor suppressor gene p53, can inhibit tumor growth.34 In 2003, Yang et al discovered that GDF15 is linked to p53 pathway activation in HCT116 human colon cancer cells, a finding also confirmed in p53-wild-type HepG2 and p53-deficient Hep3B cells.35,36 This was evidenced by the upregulation of p21, a cyclin-dependent kinase inhibitor that regulates cell cycle progression and senescence.37
However, p21 also has a pro-tumorigenic role in HCC. Marhenke et, al. found that the absence of p21 impairs hepatocyte proliferation, while p21 promotes HCC proliferation by facilitating the formation of the cell cycle protein D1-CDK4 complex.38 GDF15 is known to inhibit other cancers in its early stages but promote it in later stages.39–41 The dual role of GDF15 in HCC development, potentially influenced by p21’s dual role, is rarely reported.
GDF15 is found in both HSCs and HCC cells. HCC is commonly seen in people with fibrotic livers. HSCs act as fibroblasts that produce extracellular matrix (ECM) proteins and are key players in liver fibrosis.42 Myojin et al clarified a study using a mixed culture of hepatoma cells and HSCs, where they knocked out GDF15 in LX-2 cells (a human HSC cell line). They found that without GDF15, LX-2 cells no longer promoted the growth of hepatoma cells.43 This research demonstrates HSCs promote the proliferation of hepatoma cells through autophagy-dependent production of GDF15. This study shows that HSCs promote hepatoma cell proliferation through autophagy-dependent GDF15 production. Additionally, GDF15 induces collagen synthesis in HSCs, increasing liver fibrosis.44,45
The study showed a significant increase in GDF15 in human HCC, with more GDF15-expressing HSCs in cancerous regions than in non-cancerous ones, indicating the role of GDF15 in promoting liver tumors.43
When the liver is damaged in MASH or MAFLD, HSCs are activated and become myofibroblast-like cells.46,47 These activated HSCs proliferate and secrete extracellular matrix, leading to liver fibrosis and changes in liver structure.46,48 Overactivation of HSCs by GDF15 can worsen liver fibrosis and possibly contribute to HCC development.49 Activated HSCs also promote tumor cell growth, migration, and invasion by secreting factors like HGF and Osteopontin.50,51
In mice without GDF15, severe liver inflammation and fibrosis occur, showing GDF15 helps prevent MASH progression.52 In humans, GDF15 levels correlate with MAFLD severity and indicate fibrosis progression in MASH/MAFLD-HCC.53 Therefore, targeting HSC activation and proliferation may offer new treatment strategies for MASH/MAFLD-HCC. HSCs promote liver fibrosis and possibly tumor development in MASH-HCC, so interventions targeting HSCs could be a new treatment approach.
The Role of GDF15 in HCC Metastasis: Angiogenesis, Metastasis Promotion, and Therapeutic Interactions
Neoangiogenesis supports tumor development, and activating the tyrosine kinase steroid receptor coactivator (Src) is necessary for this process. Treating endothelial cells with recombinant GDF15 increases Src phosphorylation.54 Using the Src inhibitor Saracatinib reduces cell migration, tubular formation, and phosphorylation of mitogen-activated protein kinase (MAPK) Erk1/2, P38MAPK, c-Jun N-terminal kinase, AKT, and nuclear factor kappa-B (NF-κB) in GDF15-treated cells.31,55 Additionally, inhibitors of AKT, P38 MAPK, ERK1/2, or JNK block GDF15-triggered Src signaling.31 These findings suggest GDF15 promotes angiogenesis by activating Src and its downstream signals.
In vivo experiments with nude mice showed that HCC cells damaged by chemotherapy can induce angiogenesis through GDF15.31 GDF15 released by these HCC cells promotes proliferation, migration, and tube formation in endothelial cells, aiding residual tumor growth after chemotherapy.
However, in a diethylnitrosamine-induced HCC mouse model, high GDF15 expression was seen in small HCC foci, but not in advanced tumors. There was no significant difference in tumor area or invasiveness between GDF15 knockout mice and wild-type mice, indicating that suppressing GDF15 did not affect tumor formation, growth, or invasiveness in this model.55
Effects of GDF15 on multiple immune cell subsets on the promotion of neovascularization. GDF15 promotes the differentiation of neutrophil into the N2 phenotype for the production of pro-angiogenic factors.56,57 GDF15 promotes the secretion of multiple pro-angiogenic growth factors (VEGF, FGF and EGF), angiogenic chemokine (C-X-C motif) ligands, and angiogenesis-associated factors (TGF-β and TNF-α) after macrophage polarization towards the M2 phenotype.58
Angiogenesis is essential for providing nutrients and oxygen to growing tumors. Drugs targeting the receptor VEGF or receptor tyrosine kinases can effectively inhibit angiogenesis is one of the main therapeutic strategies for HCC.
GDF15 and Cachexia
Cachexia, marked by severe weight loss and muscle wasting, often occurs in advanced HCC and significantly reduces life expectancy.59 Key contributors to cachexia include inflammation, metabolic dysregulation, and anorexia.60 The combination of anorexia and inflammation disrupts metabolism, leading to cachexia. Elevated levels of circulating GDF15 are linked to reduced food intake in cachexia patients.61
The GDF15/GFRAL pathway is key in regulating energy balance and anorexia. GDF15 reduces appetite, lean body mass (LBM), and fat mass (FM) by binding to the neuroreceptor GFRAL in the brainstem. This binding inhibits food intake, adjusts energy use, and promotes weight loss.62 GDF15 acts on the appetite center in the brainstem and hypothalamus.60 It binds specifically to GFRAL and RET, forming a receptor complex that activates intracellular signaling through phosphorylation.63
The study conducted by Johnen et al provides evidence supporting the role of GDF15 in inducing tumor-related anorexia and weight loss. This effect involves the modulation of various biological factors, including hypothalamic function, ERK1, ERK2, activation of transcription-3, neuropeptide Y, and morphine-like neuropeptides.63
Signaling Pathway of GDF15 in HCC
GDF15 activates the AKT/GSK-3β/β-catenin signaling pathway by inducing AKT phosphorylation, which inhibits GSK-3β. This inhibition prevents β-catenin degradation, causing its accumulation and movement into the nucleus. In the nucleus, β-catenin activates oncogenic targets like cyclin D1, myelocytomatosis (MYC), metalloproteinase-2 (MMP-2), and MMP-7, leading to epithelial-to-mesenchymal transition (EMT), tumor growth, and metastasis.29 The phenomenon of EMT has been identified as a critical determinant in the metastasis and migration of HCC cells.64,65
The mRNA synthesized by the HCV encodes proteins that collaborate with GDF15 to induce the activation of the downstream MAPK pathway.66 The EGFR-ERK signaling pathway significantly impacts HCC progression.67 High EGFR expression in HCC is linked to metastasis and lower survival rates. EGFR activation and MAPK signaling contribute to the expression of pro-inflammatory and angiogenic proteins involved in liver cancer development.
GDF15 significantly increases in HCC cells when exposed to chemotherapy or hypoxia. This upregulation, triggered by DNA damage, activates p38 MAPK, JNK, and ERK1/2 pathways, as shown in Figure 1. In the liver, JNK phosphorylation disrupts mitochondrial function, raises oxidative stress, and produces reactive oxygen species, leading to cell death.68 Consequently, GDF15 has been associated with an unfavorable prognosis in HCC. Urakawa et al found that introducing recombinant human GDF15 increased cell proliferation and activated AkT and ERK1/2 phosphorylation in ESCC cell lines. This suggests GDF15 may be linked to abnormal tumor growth and a poorer prognosis.69
 |
Figure 1 Signaling pathway of the GDF15 expression. GDF15 induces proliferation and pro-angiogenic effects in HCC through the phosphorylation of Src and downstream AKT, MAPK, and NF-κB signaling pathways. GDF15 activates p53 pathway to inhibit HCC.P21 promotes HCC proliferation by facilitating the formation of the cell cycle protein D1-CDK4 complex. Created with BioRender.com. |
Tumor stem cells, known as CSCs, are specific cell groups within tumors that drive invasion, recurrence, metastasis, and resistance to treatments. HSCs play a crucial role in the tumor microenvironment by secreting ECM proteins during liver damage and inflammation, leading to fibrous tissue formation and aiding HCC progression. The interaction between HSCs and CSCs promotes cancer development through various mechanisms.44 Furthermore, GDF15 induces the synthesis of collagen in HSCs, thereby augmenting fibrogenic processes in the liver.45
Immunoregulatory Effect of GDF15 in HCC
The genetic elimination of GDF15 in HCC induces a transition of the immunosuppressive TME to an inflammatory state. GDF15 plays a role in promoting suppressive functionality of immune cells as illustrated in Figure 2.
 |
Figure 2 Effect of GDF15 on immune cells in liver cancer immune microenvironment. GDF15 promotes HCC development in the tumor microenvironment through immune escape, hepatocyte fibrosis, and tumor angiogenesis pathways. GDF15 inhibits M2-type cell action and promotes M2 to M1 conversion, inhibits nTreg-type cell action and promotes nTreg to iTreg reprogramming, inhibits DC maturation and T-cell activation, and inhibits NK cells, which diminishes the pro-apoptotic effects of immune cells and promotes immune escape. GDF15 promotes hepatocyte fibrosis through activation of hepatic stellate cells and will promote the transformation of MASH/MAFLD to HCC. GDF15 promotes the transformation of M1 to M2 and the differentiation of neutrophils into the N1 phenotype, which promotes tumour angiogenesis and exacerbates HCC. Created with BioRender.com. |
Treg Cell
The harmful impact of GDF15 on HCC involves its immunosuppressive role through Treg cells. Wang et al discovered that GDF15 binds to the CD48 receptor on Treg cells, inhibiting the ERK- Activator protein 1 (AP1) pathway downstream of the T cell recepor (TCR).24 This downregulates STIP1 homology and U-Box containing protein 1 (STUB1), preventing FOXP3 polyubiquitination, leading to FOXP3 accumulation and HCC development. GDF15 also reduces phosphorylation levels of lymphocyte-specific protein tyrosine kinase (Lck) and ERK, lowers STUB1 expression, and decreases nuclear levels of cjun and c-fos proteins.24 Unlike TGF-β, GDF15 enhances FOXP3 protein expression without affecting FOXP3 mRNA, enhancing nTreg function and inducing iTreg production to suppress immunosuppressive responses.
GDF15 increases FOXP3 protein expression in both mouse and human CD4+ T cells.70 However, unlike TGF-β, GDF15 does not affect FOXP3 mRNA expression. Instead, GDF15 blocks FOXP3 ubiquitination, enhancing the inhibitory function of natural Treg (nTreg) cells and inducing the production of induced Treg (iTreg) cells to suppress immune responses.24,71
Dendritic Cell (DC)
DCs are crucial for initiating antigen-specific immune responses. GDF15 inhibits DC maturation and function, impairing tumor-specific immune responses. Zhou et al showed that GDF15 prevents surface protrusion formation during DC maturation, reduces the expression of maturation and co-stimulatory factors, and inhibits T cell stimulation and CTL activation by DCs.72 This suppression leads to weaker immune responses against tumors. In ovarian cancer, GDF15 targets CD44 in DCs, decreasing their function and aiding immune escape.23 In HBV-associated HCC, elevated GDF15 near DCs suppresses antigen presentation, contributing to recurrence and poorer prognosis, indicating the role of in immune escape in HCC.73
Macrophage
Hepatic Macrophages play a crucial role in the anti-tumor response during hepatocarcinogenesis. In MAFLD, the loss of CD4+ T lymphocytes in the liver promotes hepatocarcinogenesis, but hepatic macrophages can prevent their interaction.74 Tumor-derived GDF15 facilitates HCC development by inhibiting the TAK1 to NF-κB signaling, reducing TNF and NO synthesis, and thus inhibiting macrophage activity.40,75 This NF-κB/GDF15 axis is crucial in early tumorigenesis for evading immunosurveillance.
However, during HCC progression, macrophages shift from an M2 to an M1 phenotype, with M2 macrophages promoting cancer cell proliferation and invasion by suppressing the adaptive immune system. GDF15 drives macrophages towards an M2 phenotype, promoting HCC and alleviating metabolic syndrome.76,77 Thus, despite the double-edged role of hepatic macrophages in HCC development and progression, GDF15 always acts in the direction of promoting HCC development.
Natural Killer (NK) Cell
NK cells are crucial in viral hepatitis and HCC by killing cells and secreting cytokines like interferon.78 GDF15 helps colon cancer cells evade NK cell surveillance by regulating their activity, in which GDF15 expression is regulated by acetylation.79 In HBV-induced HCC, NK cells have a dual role: they are hepatoprotective and antifibrotic, but their activation can also cause liver damage.80 NK cell-derived IFN-γ promotes HCC development through the EpCAM-EMT axis in HBV.81 However, the role of GDF15 in modulating NK cells in HCC remains unclear.
GDF15 as a Target for HCC Diagnosis, Prognosis, and Treatment
To investigate the changes in GDF15 levels at different stages of HCC development in conjunction with serum and tissue studies of clinical cases, and to investigate whether GDF15 can be used as a biomarker for HCC diagnosis as illustrated in Table 1.
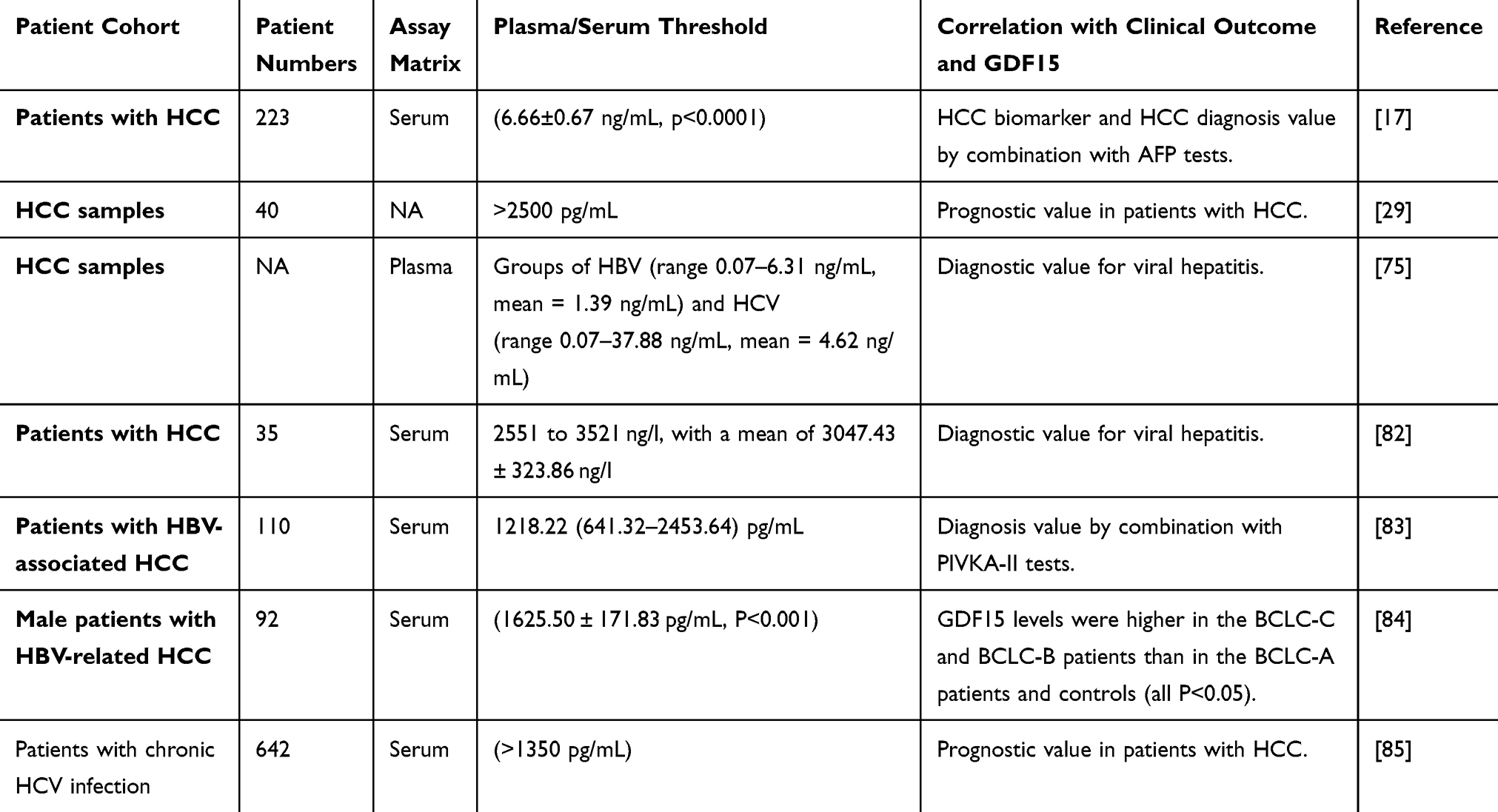 |
Table 1 Expression Level of GDF15 in Peripheral Blood of Patients with HCC |
GDF15 is a Serum Indicator of HCC
Alpha-fetoprotein (AFP) is a common biomarker for HCC, but only 10–20% of early-stage tumors show abnormal AFP levels.86 GDF15 has been shown to be upregulated in HCC, indicating its potential as a diagnostic or prognostic biomarker.82 In healthy individuals, serum GDF15 levels typically range from 0.2 to 1.2 ng/mL, with a median of 0.30 ng/mL.75,87 In HCC patients, GDF15 levels are significantly higher, ranging from 2551 to 3521 ng/L, compared to those without HCC or with negative biopsy results.82
Serum GDF15 has been validated as a biomarker for HBV-HCC. Monitoring GDF15 along with AFP and PIVKA-II enhances HCC diagnosis sensitivity and specificity.83,88 Immunohistochemistry (IHC) testing shows increased GDF15 expression in moderate to severe HCC, correlating with pathological grading, but not with age, gender, or tumor node metastasis (TNM) stage.29,89 The Barcelona Clinic Liver Cancer (BCLC) system, the most reliable staging method for HCC, includes five stages: stage 0 (extremely early), stage A (early), stage B (intermediate), stage C (advanced), and stage D (end-stage).84,90 A study by Chen et al found elevated GDF15 levels in patients with BCLC-B and BCLC-C compared to healthy individuals and those with BCLC-A.91
GDF15 inhibits early prostate cancer development but promotes metastasis in later stages. It regulates GDF15 expression throughout tumor development.92 GDF15 is linked to the progression and poorer survival in HCC.82,88 Elevated GDF15 levels are found in HCV (0.07–37.88 ng/mL) and HBV (0.07–6.31 ng/mL) patients, with much higher levels in some HCC patients (13.83, 35.18, 37.21 ng/mL).75 Therefore, GDF15 can serve as a diagnostic marker for HCC due to its significant up-regulation in infected patients.85
The expression level of GDF15 in blood or tissues affected by HCC are detailed in Table 1.
GDF15 as a Therapeutic Target for Liver Cancer Development
Given the critical role of GDF15 in HCC, inhibiting the function or expression of GDF15 may become an effective therapeutic strategy. By developing drugs targeting GDF15 or its receptor, its signaling pathway in HCC could be blocked, thereby inhibiting the growth and spread of HCC.
The main agents signaling pathways of HCC are detailed in Table 2.
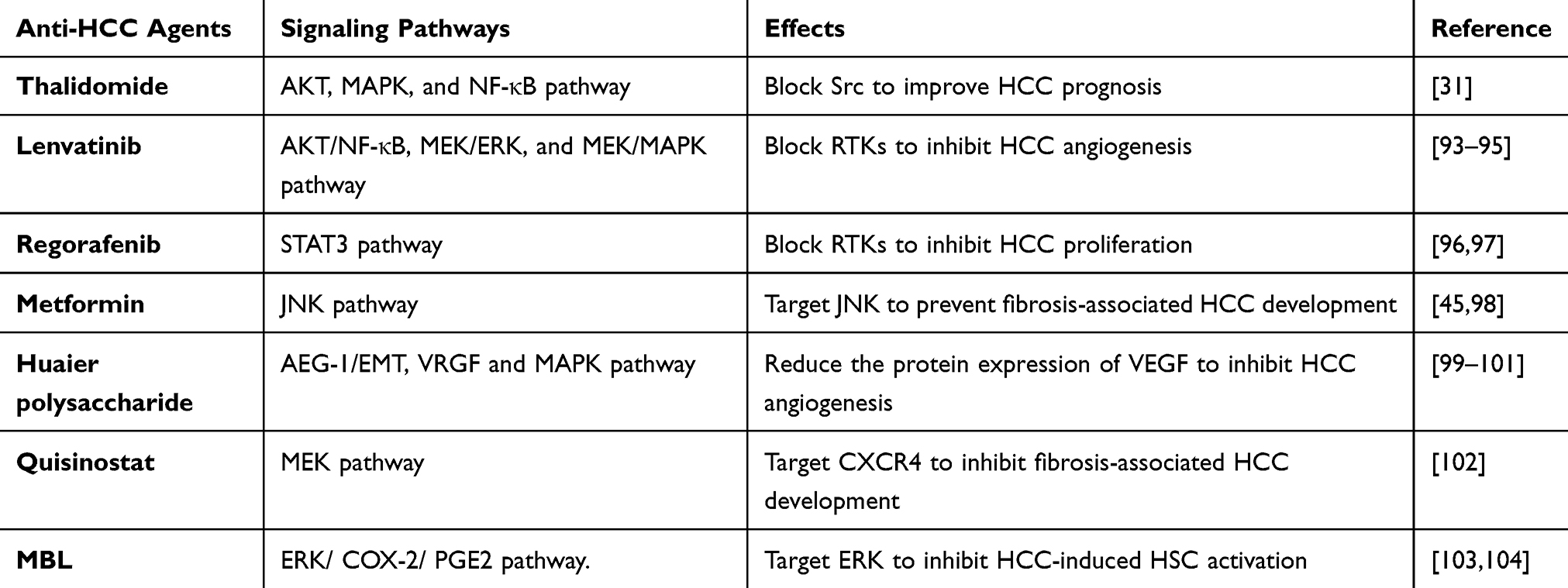 |
Table 2 The Main Agents and Its Mechanisms in HCC |
Lenvatinib
Lenvatinib is a tyrosine kinase inhibitor that blocks receptor dimerization and autophosphorylation, inhibiting pathways like AKT/NF-κB, MEK/ERK, and MEK/MAPK.93,94 GDF15 promotes angiogenesis by binding to vascular endothelial growth factor receptors (VEGFR).95 Lenvatinib inhibits tumor angiogenesis by blocking these receptors. Phase I clinical trials in Japan, Europe, and the US have shown its effectiveness in various solid tumors.105
A Phase I trial of Lenvatinib (12 mg/day) for HCC in 46 patients showed a median time to progression of 7.4 months, median overall survival of 18.7 months, a response rate of 37%, and a disease control rate of 78%.106 These significant results in metastatic HCC led to a Phase III trial comparing Lenvatinib with Sorafenib, the first tyrosine kinase inhibitor (TKI) for metastatic HCC.107,108 Lenvatinib was found to be superior in overall survival, resulting in its approval as a frontline therapy for advanced HCC due to its effectiveness in blocking abnormal signal pathways.94
TKI resistance is a major challenge in advanced HCC management. Most patients on Sorafenib develop resistance within 6 months.109 Although Lenvatinib is as effective as Sorafenib for overall survival, acquired resistance still limits its efficacy. Targeting lipid metabolism pathways is a promising strategy to overcome treatment resistance in HCC.110
Regorafenib
Regorafenib is an oral multikinase inhibitor similar to sorafenib, targeting kinases involved in angiogenesis and tumor growth. It enhances anti-tumor immunity by regulating macrophages and increasing CD8+ T cell activity, switching macrophages from a tumor-promoting M2 phenotype to a tumor-inhibiting M1 phenotype, thereby blocking the role of GDF15 in HCC proliferation.111 Regorafenib also inhibits the STAT3 pathway downstream of GDF15, enhances NK cell activity by upregulating NKG2D ligands, and helps NK cells recognize and kill HCC cells, countering the function of GDF15 in promoting immune escape.96,97
Metformin in Combination with Transarterial Chemoembolization (TACE)
TACE is a key treatment for inoperable HCC. Metformin significantly reduces GDF15 production in HCC cells after treatment by targeting JNK, thereby decreasing fibrosis and GDF15 expression.45,98 This counteracts treatment-induced fibrosis and cirrhosis. Additionally, TACE-induced damage can increase GDF15, promoting tumor angiogenesis, which thalidomide can mitigate by inhibiting AKT, MAPK, and NF-κB signaling via Src inhibition.31 Combining thalidomide with TACE improves HCC prognosis, offering better therapeutic outcomes.
Huaier Polysaccharide (HP)
The poor prognosis and high mortality in HCC result mainly from its tendency to invade and metastasize. Inhibiting these processes effectively can reduce fatality rates. Factors like CSC and EMT contribute to tumor recurrence and metastasis.112 Recent research suggests that LCSCs play a crucial role in HCC relapse and metastasis due to their increased tumorigenicity, metastatic potential, and chemotherapy resistance.113 Notably, HP has been identified as a substance with anti-invasive and metastatic properties, demonstrated in experiments on human HCC cells. HP’s efficacy in impeding HCC progression and metastasis is attributed to its regulatory impact on the astrocyte elevated gene-1 (AEG-1)/EMT and MAPK pathways.99–101
Potential Therapeutic Targets for HCC
Quisinostat, a novel chemotherapy for HCC, shows promising antitumor activity both in vitro and in vivo, especially when combined with sorafenib.114 Sorafenib/MEK inhibitor-loaded nanoparticles targeting C-X-C-Motif receptor 4 (CXCR4) effectively suppress hepatic fibrosis and prevent fibrosis-associated HCC development and liver metastasis.102
Mannan binding lectin (MBL) interacts with HSCs, inhibiting HCC-induced HSC activation by downregulating the ERK/cyclooxygenase-2(COX-2)/ prostaglandin E2 (PGE2) pathway.103,104 Resveratrol inhibits orthotopic HCC tumor growth and reduces the frequency of CD8+CD122+Tregs and M2-like macrophages.115 CD48 is identified as the primary receptor for GDF15 in the immune system, offering a pathway for therapeutic inhibition of GDF15 to achieve successful HCC clearance with minimal side effects.24
Conclusion
This review focuses primarily on the role of GDF15 in the tumor immune microenvironment. GDF15 interacts with its receptor CD48, playing a crucial role in the immune response in cancer and other diseases. Simultaneously, GDF15 directly interacts with immune cells in the tumor microenvironment, primarily through the following mechanisms: disrupting DC activation and maturation; reducing T cell induction and activation by DCs; blocking immune cell extravasation and tumor infiltration; inhibiting T and NK cell cytotoxicity against tumor cells; inducing immune suppression by generating and enhancing regulatory T cell function in tumor tissues. Additionally, in the process of transition from MASH and MAFLD to HCC, GDF15 promotes the occurrence and development of HCC, suggesting that GDF15 may become a new target for anti-tumor immunity. Increasing clinical evidence suggests that serum GDF15 levels can be used for the diagnosis and prognosis of HCC, but substantial clinical data are still needed for confirmation.
Currently, drug research mainly focuses on Lenvatinib and Sorafenib, which act by inhibiting receptor tyrosine kinases (RTKs) to block downstream tumor angiogenesis. However, there is a need for further development of targeted drugs for HCC progression, as GDF15 is involved in the entire process of HCC development, with high GDF15 expression promoting HCC progression and poor prognosis. Interestingly, previous research has highlighted the potential of targeting GDF15 for the treatment of HCC. Therefore, GDF15 provides a new avenue for HCC treatment.
Abbreviations
AEG-1, astrocyte elevated gene-1; AFP, alpha-fetoprotein; AP1, Activator protein 1; BCLC, Barcelona Clinic Liver Cancer; CDK, cyclin-dependent kinases; COX-2, cyclooxygenase-2; CSCs, cancer stem cells; CTL, cytotoxic T-lymphocyte; CXCR4, chemokine C-X-C-motif receptor 4; DC, dendritic cells; EFGR, epidermal growth factor receptor; ECM, extracellular matrix; EMT, epithelial-to-mesenchymal transition; ER, endoplasmic reticulum; ERK, extracellular-signal related kinase; GDF15, growth differentiation factor 15; GFRα-1, GDNF family receptor α-1; GSK, glycogen synthase kinase; HBV-HCC, hepatitis B virus associated hepatocellular carcinoma; HCC, hepatocellular carcinoma; HCV, Hepatitis C Virus; HP, Huaier polysaccharide; HSCs, Hepatic stellate cells; IFN-γ, Interferon-γ; IHC, Immunohistochemistry; iTreg, induced Treg; JNK, c-Jun N-terminal kinase; LBM, lean body mass; Lck, lymphocyte-specific protein tyrosine kinase; LCSCs, liver cancer stem-like cells; MAPK, mitogen-activated protein kinases; MASH, metabolic dysfunction associated steatohepatitis; MASLD, metabolic dysfunction associated fatty liver disease; MBL, mannan binding lectin; MIC-1, macrophage inhibitory factor-1; MMP-2, matrix metalloproteinase-2; MYC, myelocytomatosis; NAG-1, nonsteroidal anti-inflammatory drugs activity gene-1; NF-κB, nuclear factor kappa-B; NK, Natural killer; NKG2D, natural killer cell group 2D; NKG2DL, natural killer cell group 2D ligands; NO, Nitric oxide; NSAID, nonsteroidal anti-inflammatory drug; nTreg, natural Treg; PDF, prostate-derived factor; PGE2, prostaglandin E2; PIVKA-II, protein induced by vitamin K absence or antagonist-II; PTGF-β, placental transforming growth factor-β; RTKs, receptor tyrosine kinases; Src, steroid receptor coactivator; STUB1, STIP1 homology and U-Box containing protein 1; TACE, transarterial chemoembolization; TAK1, TGF-β-activated kinase; TGF, transforming growth factor; TKI, tyrosine kinase inhibitor; TME, tumor microenvironment; TNF, tumor necrosis factor; TNM, tumor node metastasis; VEGFR, vascular endothelial growth factor receptors.
Funding
This work was supported by the Fund for Science and Technology Development Plan Project of Jilin Province, China (20240601022RC).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bootcov MR, Bauskin AR, Valenzuela SM, et al. MIC-1, a novel macrophage inhibitory cytokine, is a divergent member of the TGF-beta superfamily. Proc Natl Acad Sci USA. 1997;94(21):11514–11519. doi:10.1073/pnas.94.21.11514
2. Baek SJ, Kim KS, Nixon JB, Wilson LC, Eling TE. Cyclooxygenase inhibitors regulate the expression of a TGF-beta superfamily member that has proapoptotic and antitumorigenic activities. Mol Pharmacol. 2001;59(4):901–908. doi:10.1124/mol.59.4.901
3. Hsu JY, Crawley S, Chen M, et al. Non-homeostatic body weight regulation through a brainstem-restricted receptor for GDF15. Nature. 2017;550:255–259. doi:10.1038/nature24042
4. Bauskin AR, Zhang HP, Fairlie WD, et al. The propeptide of macrophage inhibitory cytokine (MIC-1), a TGF-beta superfamily member, acts as a quality control determinant for correctly folded MIC-1. EMBO J. 2000;19(10):2212–2220. doi:10.1093/emboj/19.10.2212
5. Yang W, Mok MT, Li MS, et al. Epigenetic silencing of GDF1 disrupts SMAD signaling to reinforce gastric cancer development. Oncogene. 2016;35(16):2133–2144. doi:10.1038/onc.2015.276
6. Levine AJ, Brivanlou AH GDF3, a BMP inhibitor, regulates cell fate in stem cells and early embryos; 2006.
7. Miyamoto Y, Mabuchi A, Shi D, et al. A functional polymorphism in the 5′ UTR of GDF5 is associated with susceptibility to osteoarthritis. Nature Genet. 2007;39(4):529–533. doi:10.1038/2005
8. Capellini TD, Chen H, Cao J, et al. Ancient selection for derived alleles at a GDF5 enhancer influencing human growth and osteoarthritis risk. Nature Genet. 2017;49(8):1202–1210. doi:10.1038/ng.3911
9. Lee KJ, Mendelsohn M, Jessell TM. Neuronal patterning by BMPs: a requirement for GDF7 in the generation of a discrete class of commissural interneurons in the mouse spinal cord. Genes Dev. 1998;12(21):3394–3407. doi:10.1101/gad.12.21.3394
10. Paralkar VM, Vail AL, Grasser WA, et al. Cloning and characterization of a novel member of the transforming growth factor-β/bone morphogenetic protein family. J Biol Chem. 1998;273(22):13760–13767. doi:10.1074/jbc.273.22.13760
11. Emmerson PJ, Duffin KL, Wu X. GDF15 and growth control. Front Physiol. 2018;9:411437. doi:10.3389/fphys.2018.01712
12. Yang L, Chang CC, Sun Z, et al. GFRAL is the receptor for GDF15 and is required for the anti-obesity effects of the ligand. Nat Med. 2017;23(10):1158–1166. doi:10.1038/nm.4394
13. Mullican SE, Lin-Schmidt X, Chin CN, et al. GFRAL is the receptor for GDF15 and the ligand promotes weight loss in mice and nonhuman primates. Nat Med. 2017;23(10):1150–1157. doi:10.1038/nm.4392
14. Takahashi M, Ritz J, Cooper GM. Activation of a novel human transforming gene, ret, by DNA rearrangement. Cell. 1985;42(2):581–588. doi:10.1016/0092-8674(85)90115-1
15. Olsen OE, Skjærvik A, Størdal BF, Sundan A, Holien T. TGF-β contamination of purified recombinant GDF15. PLoS One. 2017;12(11):e0187349. doi:10.1371/journal.pone.0187349
16. Shlomai A, de Jong YP, Rice CM. Virus Associated Malignancies: The Role of Viral Hepatitis in Hepatocellular Carcinoma. Elsevier; 2014:78–88.
17. Baecker A, Liu X, La Vecchia C, Zhang ZF. Worldwide incidence of hepatocellular carcinoma cases attributable to major risk factors. Eur J Cancer Prev. 2018;27(3):205–212. doi:10.1097/cej.0000000000000428
18. Donne R, Lujambio A. The liver cancer immune microenvironment: therapeutic implications for hepatocellular carcinoma. Hepatology. 2023;77(5):1773–1796. doi:10.1002/hep.32740
19. Ogunwobi OO, Harricharran T, Huaman J, et al. Mechanisms of hepatocellular carcinoma progression. World J Gastroenterol. 2019;25(19):2279. doi:10.3748/wjg.v25.i19.2279
20. Liu H, Huang Y, Lyu Y, Dai W, Tong Y, Li Y. GDF15 as a biomarker of ageing. Exp Gerontology. 2021;146:111228. doi:10.1016/j.exger.2021.111228
21. Wang D, Day EA, Townsend LK, Djordjevic D, Jørgensen SB, Steinberg GR. GDF15: emerging biology and therapeutic applications for obesity and cardiometabolic disease. Nat Rev Endocrinol. 2021;17(10):592–607. doi:10.1038/s41574-021-00529-7
22. Li C, Wang J, Kong J, et al. GDF15 promotes EMT and metastasis in colorectal cancer. Oncotarget. 2016;7(1):860. doi:10.18632/oncotarget.6205
23. Gao Y, Xu Y, Zhao S, et al. Growth differentiation factor-15 promotes immune escape of ovarian cancer via targeting CD44 in dendritic cells. Exp Cell Res. 2021;402(1):112522. doi:10.1016/j.yexcr.2021.112522
24. Wang Z, He L, Li W, et al. GDF15 induces immunosuppression via CD48 on regulatory T cells in hepatocellular carcinoma. J Immunoth Can. 2021;9(9). doi:10.1136/jitc-2021-002787
25. M-Z M, Zhang X-H, Li B, Liang T-J, Yang C-M, Liu J-Y. Growth differentiation factor 15 (GDF15) contributes to invasion and anti-anoikis of hepatocellular cancer through TGF-β/Smad-associated signaling. Int J Clin Exp Med. 2018;11(12):12964–12973.
26. Fang L, Li F, Gu C. GDF-15: a multifunctional modulator and potential therapeutic target in cancer. Curr. Pharm. Des. 2019;25(6):654–662. doi:10.2174/1381612825666190402101143
27. Spanopoulou A, Gkretsi V. Growth differentiation factor 15 (GDF15) in cancer cell metastasis: from the cells to the patients. Clin Exp Metastasis. 2020;37(4):451–464. doi:10.1007/s10585-020-10041-3
28. Emmerson PJ, Wang F, Du Y, et al. The metabolic effects of GDF15 are mediated by the orphan receptor GFRAL. Nature Med. 2017;23(10):1215–1219. doi:10.1038/nm.4393
29. Xu Q, H-X X, J-P L, et al. Growth differentiation factor 15 induces growth and metastasis of human liver cancer stem-like cells via AKT/GSK-3β/β-catenin signaling. Oncotarget. 2017;8(10):16972. doi:10.18632/oncotarget.15216
30. Benkheil M, Paeshuyse J, Neyts J, Van Haele M, Roskams T, Liekens S. HCV-induced EGFR-ERK signaling promotes a pro-inflammatory and pro-angiogenic signature contributing to liver cancer pathogenesis. Biochem Pharmacol. 2018;155:305–315. doi:10.1016/j.bcp.2018.07.011
31. Dong G, Zheng QD, Ma M, et al. Angiogenesis enhanced by treatment damage to hepatocellular carcinoma through the release of GDF 15. Cancer Med. 2018;7(3):820–830. doi:10.1002/cam4.1330
32. Yu J, Shen B, Chu ES, et al. Inhibitory role of peroxisome proliferator‐activated receptor gamma in hepatocarcinogenesis in mice and in vitro. Hepatology. 2010;51(6):2008–2019. doi:10.1002/hep.23550
33. Suriben R, Chen M, Higbee J, et al. Antibody-mediated inhibition of GDF15-GFRAL activity reverses cancer cachexia in mice. Nat Med. 2020;26(8):1264–1270. doi:10.1038/s41591-020-0945-x
34. Tsui K-H, Hsu S-Y, Chung L-C, et al. Growth differentiation factor-15: a p53-and demethylation-upregulating gene represses cell proliferation, invasion and tumorigenesis in bladder carcinoma cells. Sci Rep. 2015;5(1):12870. doi:10.1038/srep12870
35. Yang H, Filipovic Z, Brown D, Breit SN, Vassilev LT. Macrophage inhibitory cytokine-1: a novel biomarker for p53 pathway activation. Mol Cancer Ther. 2003;2(10):1023–1029.
36. Shin DY, Kim GY, Kim ND, et al. Induction of apoptosis by pectenotoxin-2 is mediated with the induction of DR4/DR5, Egr-1 and NAG-1, activation of caspases and modulation of the Bcl-2 family in p53-deficient Hep3B hepatocellular carcinoma cells. Oncol Rep. 2008;19(2):517–526.
37. Xiong Y, Hannon GJ, Zhang H, Casso D, Kobayashi R, Beach D. p21 is a universal inhibitor of cyclin kinases. Nature. 1993;366(6456):701–704. doi:10.1038/366701a0
38. Marhenke S, Buitrago-Molina LE, Endig J, et al. p21 promotes sustained liver regeneration and hepatocarcinogenesis in chronic cholestatic liver injury. Gut. 2014;63(9):1501–1512. doi:10.1136/gutjnl-2013-304829
39. Roth P, Junker M, Tritschler I, et al. GDF-15 contributes to proliferation and immune escape of malignant gliomas. Clin Cancer Res. 2010;16(15):3851–3859. doi:10.1158/1078-0432.CCR-10-0705
40. Ratnam NM, Peterson JM, Talbert EE, et al. NF-κB regulates GDF-15 to suppress macrophage surveillance during early tumor development. J Clin Invest. 2017;127(10):3796–3809. doi:10.1172/JCI91561
41. Vaňhara P, Hampl A, Kozubík A, Souček K. Growth/differentiation factor-15: prostate cancer suppressor or promoter? Prost Can Prost Dis. 2012;15(4):320–328. doi:10.1038/pcan.2012.6
42. Carloni V, Luong TV, Rombouts K. Hepatic stellate cells and extracellular matrix in hepatocellular carcinoma: more complicated than ever. Liver Int. 2014;34(6):834–843. doi:10.1111/liv.12465
43. Myojin Y, Hikita H, Sugiyama M, et al. Hepatic stellate cells in hepatocellular carcinoma promote tumor growth via growth differentiation factor 15 production. Gastroenterology. 2021;160(5):1741–1754. e16. doi:10.1053/j.gastro.2020.12.015
44. Quiroz Reyes AG, Lozano Sepulveda SA, Martinez-Acuña N, et al. Cancer stem cell and hepatic stellate cells in hepatocellular carcinoma. Technol Cancer Res Treat. 2023;22:15330338231163677. doi:10.1177/15330338231163677
45. Dong G, Ma M, Lin X, et al. Treatment-damaged hepatocellular carcinoma promotes activities of hepatic stellate cells and fibrosis through GDF15. Exp Cell Res. 2018;370(2):468–477. doi:10.1016/j.yexcr.2018.07.011
46. Bendixen SM, Jakobsgaard PR, Hansen D, et al. Single cell-resolved study of advanced murine MASH reveals a homeostatic pericyte signaling module. J Hepatol. 2024;80(3):467–481. doi:10.1016/j.jhep.2023.11.001
47. Kuchay MS, Choudhary NS, Mishra SK. Pathophysiological mechanisms underlying MAFLD. Diabetes Metab Syndr. 2020;14(6):1875–1887. doi:10.1016/j.dsx.2020.09.026
48. Park S, Hwang S, Sun J, et al. A novel A2a adenosine receptor inhibitor effectively mitigates hepatic fibrosis in a metabolic dysfunction-associated steatohepatitis mouse model. Int J Bio Sci. 2024;20(5):1855. doi:10.7150/ijbs.92371
49. Luo Q, Wang CQ, Yang LY, et al. FOXQ1/NDRG1 axis exacerbates hepatocellular carcinoma initiation via enhancing crosstalk between fibroblasts and tumor cells. Cancer Lett. 2018;417:21–34. doi:10.1016/j.canlet.2017.12.021
50. Trusolino L, Bertotti A, Comoglio PM. MET signalling: principles and functions in development, organ regeneration and cancer. Nat Rev Mol Cell Biol. 2010;11(12):834–848. doi:10.1038/nrm3012
51. Song J, Ge Z, Yang X, et al. Hepatic stellate cells activated by acidic tumor microenvironment promote the metastasis of hepatocellular carcinoma via osteopontin. Cancer Lett. 2015;356(2):713–720. doi:10.1016/j.canlet.2014.10.021
52. Kim KH, Kim SH, Han DH, Jo YS, Lee Y-H, Lee M-S. Growth differentiation factor 15 ameliorates nonalcoholic steatohepatitis and related metabolic disorders in mice. Sci Rep. 2018;8(1):6789. doi:10.1038/s41598-018-25098-0
53. Chung HK, Ryu D, Kim KS, et al. Growth differentiation factor 15 is a myomitokine governing systemic energy homeostasis. J Cell Biol. 2017;216(1):149–165. doi:10.1083/jcb.201607110
54. Gordon EJ, Fukuhara D, Weström S, et al. The endothelial adaptor molecule TSAd is required for VEGF-induced angiogenic sprouting through junctional c-Src activation. Sci Signal. 2016;9(437):ra72. doi:10.1126/scisignal.aad9256
55. Zimmers TA, Jin X, Gutierrez JC, et al. Effect of in vivo loss of GDF-15 on hepatocellular carcinogenesis. J Cancer Res Clin Oncol. 2008;134:753–759. doi:10.1007/s00432-007-0336-4
56. Wei X, Zou S, Xie Z, et al. EDIL3 deficiency ameliorates adverse cardiac remodelling by neutrophil extracellular traps (NET)-mediated macrophage polarization. Cardiov Res. 2022;118(9):2179–2195. doi:10.1093/cvr/cvab269
57. Rochette L, Méloux A, Zeller M, Cottin Y, Vergely C. Functional roles of GDF15 in modulating microenvironment to promote carcinogenesis. Bioch et Bioph Acta. 2020;1866(8):165798. doi:10.1016/j.bbadis.2020.165798
58. Larionova I, Kazakova E, Gerashchenko T, Kzhyshkowska J. New angiogenic regulators produced by TAMs: perspective for targeting tumor angiogenesis. Cancers. 2021;13(13):3253. doi:10.3390/cancers13133253
59. Husaini Y, Tsai VW, Manandhar R, et al. Growth differentiation factor-15 slows the growth of murine prostate cancer by stimulating tumor immunity. PLoS One. 2020;15(6):e0233846. doi:10.1371/journal.pone.0233846
60. Evans WJ, Morley JE, Argilés J, et al. Cachexia: a new definition. Clin Nutr. 2008;27(6):793–799. doi:10.1016/j.clnu.2008.06.013
61. Navarro IBK, Schraner M, Riediger T. Brainstem prolactin-releasing peptide contributes to cancer anorexia-cachexia syndrome in rats. Neuropharmacology. 2020;180:108289. doi:10.1016/j.neuropharm.2020.108289
62. Suzuki H, Mitsunaga S, Ikeda M, et al. Clinical and tumor characteristics of patients with high serum levels of growth differentiation factor 15 in advanced pancreatic cancer. Cancers (Basel). 2021;13(19):4842. doi:10.3390/cancers13194842
63. Ahmed DS, Isnard S, Lin J, Routy B, Routy JP. GDF15/GFRAL pathway as a metabolic signature for cachexia in patients with cancer. J Cancer. 2021;12(4):1125–1132. doi:10.7150/jca.50376
64. Zhang Y, Wang X, Zhang M, Zhang Z, Jiang L, Li L. GDF15 promotes epithelial-to-mesenchymal transition in colorectal [corrected]. Artif Cells Nan Bio. 2018;46(sup2):652–658. doi:10.1080/21691401.2018.1466146
65. Mohan CD, Rangappa S, Nayak SC, Sethi G, Rangappa KS. Paradoxical functions of long noncoding RNAs in modulating STAT3 signaling pathway in hepatocellular carcinoma. Biochim Biophys Acta Rev Cancer. 2021;1876(1):188574. doi:10.1016/j.bbcan.2021.188574
66. Berasain C, Avila MA. The EGFR signalling system in the liver: from hepatoprotection to hepatocarcinogenesis. J Gastroenterol. 2014;49(1):9–23. doi:10.1007/s00535-013-0907-x
67. Komposch K, Sibilia M. EGFR signaling in liver diseases. Int J Mol Sci. 2015;17(1). doi:10.3390/ijms17010030
68. Win S, Than TA, Le BH, García-Ruiz C, Fernandez-Checa JC, Kaplowitz N. Sab (Sh3bp5) dependence of JNK mediated inhibition of mitochondrial respiration in palmitic acid induced hepatocyte lipotoxicity. J Hepatol. 2015;62(6):1367–1374. doi:10.1016/j.jhep.2015.01.032
69. Urakawa N, Utsunomiya S, Nishio M, et al. GDF15 derived from both tumor-associated macrophages and esophageal squamous cell carcinomas contributes to tumor progression via Akt and Erk pathways. Lab Invest. 2015;95(5):491–503. doi:10.1038/labinvest.2015.36
70. Kochetkova I, Thornburg T, Callis G, Pascual DW. Segregated Regulatory CD39+ CD4+ T Cell Function: TGF-β–Producing Foxp3− and IL-10–Producing Foxp3+ cells are interdependent for protection against collagen-induced arthritis. J Immunol. 2011;187(9):4654–4666. doi:10.4049/jimmunol.1100530
71. Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003;299(5609):1057–1061. doi:10.1126/science.1079490
72. Zhou Z, Li W, Song Y, et al. Growth differentiation factor-15 suppresses maturation and function of dendritic cells and inhibits tumor-specific immune response. PLoS One. 2013;8(11):e78618. doi:10.1371/journal.pone.0078618
73. Chen S, Huang C, Liao G, et al. Distinct single-cell immune ecosystems distinguish true and de novo HBV-related hepatocellular carcinoma recurrences. Gut. 2023;72(6):1196–1210. doi:10.1136/gutjnl-2022-328428
74. Ma C, Kesarwala AH, Eggert T, et al. NAFLD causes selective CD4(+) T lymphocyte loss and promotes hepatocarcinogenesis. Nature. 2016;531:7593):253–7. doi:10.1038/nature16969
75. Si Y, Liu X, Cheng M, et al. Growth differentiation factor 15 is induced by hepatitis C virus infection and regulates hepatocellular carcinoma-related genes. PLoS One. 2011;6(5):e19967. doi:10.1371/journal.pone.0019967
76. Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331(6024):1565–1570. doi:10.1126/science.1203486
77. Jung SB, Choi MJ, Ryu D, et al. Reduced oxidative capacity in macrophages results in systemic insulin resistance. Nat Commun. 2018;9(1):1551. doi:10.1038/s41467-018-03998-z
78. Liu P, Chen L, Zhang H. Natural killer cells in liver disease and hepatocellular carcinoma and the NK cell-based immunotherapy. J Immunol Res. 2018;2018:1–8. doi:10.1155/2018/1206737
79. Han B, He J, Chen Q, et al. ELFN1-AS1 promotes GDF15-mediated immune escape of colorectal cancer from NK cells by facilitating GCN5 and SND1 association. Disc Oncol. 2023;14(1):56. doi:10.1007/s12672-023-00675-6
80. Maini MK, Peppa D. NK cells: a double-edged sword in chronic hepatitis B virus infection. Front Immunol. 2013;4:42790. doi:10.3389/fimmu.2013.00057
81. Chen Y, Hao X, Sun R, Wei H, Tian Z. Natural killer cell–derived interferon‐gamma promotes hepatocellular carcinoma through the epithelial cell adhesion molecule–epithelial‐to‐mesenchymal transition axis in hepatitis B virus transgenic mice. Hepatology. 2019;69(4):1735–1750. doi:10.1002/hep.30317
82. Anis ZM, Ahmed AY, Soliman HH, Nagy HM. Serum growth differentiation factor 15 levels as a marker for liver cirrhosis and hepatocellular carcinoma on top of liver cirrhosis. Tanta Med J. 2022;50(4):251–259. doi:10.4103/tmj.tmj_173_20
83. Chen J, Tang D, Xu C, et al. Evaluation of serum GDF15, AFP, and PIVKA-II as diagnostic markers for HBV-associated hepatocellular carcinoma. Lab Med. 2021;52(4):381–389. doi:10.1093/labmed/lmaa089
84. Cillo U, Vitale A, Grigoletto F, et al. Prospective validation of the Barcelona Clinic Liver Cancer staging system. J Hepatol. 2006;44(4):723–731. doi:10.1016/j.jhep.2005.12.015
85. Myojin Y, Hikita H, Tahata Y, et al. Serum growth differentiation factor 15 predicts hepatocellular carcinoma occurrence after hepatitis C virus elimination. Aliment. Pharmacol. Ther. 2022;55(4):422–433. doi:10.1111/apt.16691
86. Villanueva A, Minguez B, Forner A, Reig M, Llovet JM. Hepatocellular carcinoma: novel molecular approaches for diagnosis, prognosis, and therapy. Ann Rev Med. 2010;61:317–328. doi:10.1146/annurev.med.080608.100623
87. Bauskin AR, Brown DA, Kuffner T, et al. Role of macrophage inhibitory cytokine-1 in tumorigenesis and diagnosis of cancer. Cancer Res. 2006;66(10):4983–4986. doi:10.1158/0008-5472.Can-05-4067
88. Liu X, Chi X, Gong Q, et al. Association of serum level of growth differentiation factor 15 with liver cirrhosis and hepatocellular carcinoma. PLoS One. 2015;10(5):1.
89. Wu K, Lin F. Lipid metabolism as a potential target of liver cancer. J Hepatocell Carcinoma. 2024;11:327–346. doi:10.2147/JHC.S450423
90. Vitale A, Morales RR, Zanus G, et al. Barcelona clinic liver cancer staging and transplant survival benefit for patients with hepatocellular carcinoma: a multicentre, cohort study. Lancet Oncol. 2011;12(7):654–662. doi:10.1016/S1470-2045(11)70144-9
91. Chen J, Dai W, Zhu C, Liu H, Li Y, Zhang P. Circulating levels of growth differentiation factor 15 and sex hormones in male patients with HBV-associated hepatocellular carcinoma. Biomed. Pharmacother. 2020;121:109574. doi:10.1016/j.biopha.2019.109574
92. Rybicki BA, Sadasivan SM, Chen Y, et al. Growth and differentiation factor 15 and NF‐κB expression in benign prostatic biopsies and risk of subsequent prostate cancer detection. Cancer Med. 2021;10(9):3013–3025. doi:10.1002/cam4.3850
93. Suyama K, Iwase H. Lenvatinib: a promising molecular targeted agent for multiple cancers. Cancer Control. 2018;25(1):1073274818789361. doi:10.1177/1073274818789361
94. Eso Y, Marusawa H. Novel approaches for molecular targeted therapy against hepatocellular carcinoma. Hepatol Res. 2018;48(8):597–607. doi:10.1111/hepr.13181
95. Hidman J, Larsson A, Thulin M, Karlsson T. Increased plasma GDF15 is associated with altered levels of soluble VEGF receptors 1 and 2 in symptomatic multiple myeloma. Acta Haematolog. 2022;145(3):326–333. doi:10.1159/000519888
96. Duan S, Guo W, Xu Z, et al. Natural killer group 2D receptor and its ligands in cancer immune escape. Mol Cancer. 2019;18:1–14. doi:10.1186/s12943-019-0956-8
97. Tai W-T, Chu P-Y, Shiau C-W, et al. STAT3 mediates regorafenib-induced apoptosis in hepatocellular carcinoma. Clin Cancer Res. 2014;20(22):5768–5776. doi:10.1158/1078-0432.CCR-14-0725
98. Fujita H, Hirose K, Sato M, et al. Metformin attenuates hypoxia‑induced resistance to cisplatin in the HepG2 cell line. Oncol Lett. 2019;17(2):2431–2440. doi:10.3892/ol.2018.9869
99. Zheng J, Li C, Wu X, et al. Huaier polysaccharides suppresses hepatocarcinoma MHCC97-H cell metastasis via inactivation of EMT and AEG-1 pathway. Int J Biol Macromol. 2014;64:106–110. doi:10.1016/j.ijbiomac.2013.11.034
100. Bao H, Liu P, Jiang K, et al. Huaier polysaccharide induces apoptosis in hepatocellular carcinoma cells through p38 MAPK. Oncol Lett. 2016;12(2):1058–1066. doi:10.3892/ol.2016.4686
101. Li C, Wu X, Zhang H, et al. A Huaier polysaccharide restrains hepatocellular carcinoma growth and metastasis by suppression angiogenesis. Int J Biol Macromol. 2015;75:115–120. doi:10.1016/j.ijbiomac.2015.01.016
102. Sung Y-C, Liu Y-C, Chao P-H, et al. Combined delivery of sorafenib and a MEK inhibitor using CXCR4-targeted nanoparticles reduces hepatic fibrosis and prevents tumor development. Theranostics. 2018;8(4):894. doi:10.7150/thno.21168
103. Li J, Li H, Yu Y, et al. Mannan-binding lectin suppresses growth of hepatocellular carcinoma by regulating hepatic stellate cell activation via the ERK/COX-2/PGE2 pathway. Oncoimmunology. 2019;8(2):e1527650. doi:10.1080/2162402X.2018.1527650
104. Wu M, Miao H, Fu R, Zhang J, Zheng W. Hepatic stellate cell: a potential target for hepatocellular carcinoma. Curr Molec Pharmacol. 2020;13(4):261–272. doi:10.2174/1874467213666200224102820
105. Matsuki M, Hoshi T, Yamamoto Y, et al. Lenvatinib inhibits angiogenesis and tumor fibroblast growth factor signaling pathways in human hepatocellular carcinoma models. Cancer Med. 2018;7(6):2641–2653. doi:10.1002/cam4.1517
106. Ikeda K, Kudo M, Kawazoe S, et al. Phase 2 study of lenvatinib in patients with advanced hepatocellular carcinoma. J Gastroenterol. 2017;52:512–519. doi:10.1007/s00535-016-1263-4
107. Cheng A-L, Finn RS, Qin S, et al. Phase III trial of lenvatinib (LEN) vs sorafenib (SOR) in first-line treatment of patients (pts) with unresectable hepatocellular carcinoma (uHCC). Am Soc Clin Oncol. 2017;2017:1.
108. Tang W, Chen Z, Zhang W, et al. The mechanisms of sorafenib resistance in hepatocellular carcinoma: theoretical basis and therapeutic aspects. Signal Transduct Target Thera. 2020;5(1):87. doi:10.1038/s41392-020-0187-x
109. Cheng A-L, Kang Y-K, Chen Z, et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: a phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009;10(1):25–34. doi:10.1016/S1470-2045(08)70285-7
110. Huang X, Wang M, Zhang D, Zhang C, Liu P. Advances in targeted drug resistance associated with dysregulation of lipid metabolism in hepatocellular carcinoma. J Hepatocell Carcinoma. 2024;11:113–129. doi:10.2147/JHC.S447578
111. Abou-Elkacem L, Arns S, Brix G, et al. Regorafenib inhibits growth, angiogenesis, and metastasis in a highly aggressive, orthotopic colon cancer model. Mol Cancer Ther. 2013;12(7):1322–1331. doi:10.1158/1535-7163.MCT-12-1162
112. Mitra A, Mishra L, Li S. EMT, CTCs and CSCs in tumor relapse and drug-resistance. Oncotarget. 2015;6(13):10697. doi:10.18632/oncotarget.4037
113. Pang WC, Poon TP. Cancer stem cell as a potential therapeutic target in hepatocellular carcinoma. Curr Cancer Drug Targets. 2012;12(9):1081–1094. doi:10.2174/156800912803987995
114. He B, Dai L, Zhang X, et al. The HDAC inhibitor quisinostat (JNJ-26481585) supresses hepatocellular carcinoma alone and synergistically in combination with sorafenib by G0/G1 phase arrest and apoptosis induction. Int J Bio Sci. 2018;14(13):1845. doi:10.7150/ijbs.27661
115. Zhang Q, Huang H, Zheng F, et al. Resveratrol exerts antitumor effects by downregulating CD8+ CD122+ Tregs in murine hepatocellular carcinoma. Oncoimmunology. 2020;9(1):1829346. doi:10.1080/2162402X.2020.1829346
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.