Back to Journals » Journal of Inflammation Research » Volume 19
Fuzheng Jiedu Formula Ameliorates Acute Lung Injury by Modulating Gut Microbiota to Enhance Short-Chain Fatty Acid
Authors Chen J, Pan S, Huo W, Wang W, Tan Z, Wu Y, Kong Y, Yin C, Gan K, Zhao M, Gao M, Xia Q, Li J, Lu Y, Yang R, Liu Y
Received 28 July 2025
Accepted for publication 25 December 2025
Published 10 February 2026 Volume 2026:19 556752
DOI https://doi.org/10.2147/JIR.S556752
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Jiankun Chen,1,2,* Simin Pan,2,* Wanhua Huo,2 Weiye Wang,2 Zhaoqi Tan,2 Yuan Wu,1 Yunqi Kong,2 Chubo Yin,2 Kao Gan,2,3 Meng Zhao,4 Ming Gao,2,5 Qinghua Xia,6 Jiqiang Li,1 Yue Lu,7 Rongyuan Yang,8,9 Yuntao Liu3,8,9
1Department of The Third General, The Second Affiliated Hospital Guangzhou University of Chinese Medicine (Guangdong Provincial Hospital of Chinese Medicine), Guangzhou, People’s Republic of China; 2The Second Clinical College of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 3Department of Emergency, The Second Affiliated Hospital Guangzhou University of Chinese Medicine (Guangdong Provincial Hospital of Chinese Medicine), Guangzhou, People’s Republic of China; 4School of Pharmaceutical Sciences, Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 5Department of Emergency, Nanhai District Hospital of Chinese Medicine (Guangdong Provincial Hospital of Integrated Chinese and Western Medicine), Foshan, People’s Republic of China; 6Department of Respiratory, Qingyuan Hospital of Chinese Medicine, Qingyuan, People’s Republic of China; 7Research Team of Bio-Molecular and System Biology of Chinese Medicine, Guangdong Academy of Traditional Chinese Medicine, Guangzhou, Guangdong Province, People’s Republic of China; 8State Key Laboratory of Traditional Chinese Medicine Syndrome, The Second Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 9Guangdong Provincial Key Laboratory of Research on Emergency in TCM, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Rongyuan Yang, Email [email protected] Yuntao Liu, Email [email protected]
Aim of Study: The objective of this research is to clarify the mechanism by which FZJDF mitigates lipopolysaccharide (LPS)- ALI through the enhancement of short-chain fatty acids (SCFAs) production by Clostridium butyricum (C. butyricum), and the modulation of the gut microbiota-gut-lung axis.
Materials and Methods: The mice were treated with FZJDF for 7 days and then treated with LPS. The therapeutic efficacy of FZJDF against LPS-induced ALI was evaluated through lung-to-weight ratio, Inflammatory factor, pathological changes. Additionally, the intestinal barrier function was evaluated by analyzing tight junction protein expression levels. 16S rRNA gene sequencing was employed to monitor alterations in the intestinal microbiome and pulmonary microbiota, while gas chromatography-mass spectrometry (GC-MS) and ultra-performance liquid chromatography (UPLC) were utilized to quantify the concentrations of SCFAs. Ultimately, the necessity of C. butyricum for FZJDF’s therapeutic influence was confirmed through antibiotic-mediated gut microbiota depletion.
Results: FZJDF significantly decreased lung-to-weight ratio and reducing inflammatory cell infiltration of neutrophils. Furthermore, it significantly elevated the expression levels of tight junction proteins. It is plausible that FZJDF may improve intestinal and lung microecological imbalance and stimulate the synthesis of SCFAs. Notably, we determine C. butyricum as the crucial bacterium for the role of FZJDF in gut barrier repair and suppression of lung inflammation in ALI mice. The use of antibiotics led to the repair of the intestinal barrier and a failure in SCFAs production, whereas C. butyricum colonization restores the therapeutic effect of FZJDF in ALI mice, further confirming that FZJDF attenuates ALI.
Conclusion: Our results imply that FZJDF could exert its palliative effect in ALI by regulating the intestinal microbiota to increase the production of SCFAs, which in turn inhibits neutrophil-mediated inflammatory responses. These findings support microbiota-targeted traditional medicine as a translational strategy for acute lung injury.
Keywords: fuzhengjiedu formula, LPS-induced ALI, short-chain fatty acids, gut-lung axis
Introduction
Acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) are recognized as a category of pulmonary disorders characterized by extensive inflammatory leukocyte infiltration, hemorrhagic exudation, and pulmonary edema. Both primary and secondary triggers, including inhalation trauma, pneumonia, sepsis, injury, pancreatitis, and transfusion-related events, can precipitate ALI/ARDS.1,2 Studies indicate that the fatality rate associated with ALI/ARDS is around 40%. Inflammation is a pivotal factor in the etiology of ALI/ARDS.3 A substantial aggregation of immune cells at injury sites initiates a sequence of inflammatory signaling pathways and the release of multiple pro-inflammatory cytokines, which in turn compromises the alveolar epithelial integrity and increases the permeability of the alveolar-capillary membrane. Consequently, this leads to the accumulation of protein-rich edema fluid in the pulmonary interstitium, resulting in pulmonary edema and tissue damage.4,5 Current therapeutic approaches for ALI/ARDS primarily involve mechanical ventilation and fluid management, supplemented with glucocorticoids, pulmonary vasodilators administered via inhalation, and extracorporeal membrane oxygenation.6,7 Despite these interventions, the prognosis for most ALI/ARDS patients remains poor, underscoring the need for novel therapeutic strategies and medications.
Recent investigations have documented that a disruption in the intestinal microbiota correlates with heightened fatality rates due to respiratory infections. The gut-lung axis underscores the complex interplay between the intestinal and pulmonary systems.8 The gut acts as a bacterial reservoir within the organism. Under normal physiological conditions, the intestinal barrier, enteric microflora, and microbial byproducts constitute an intestinal ecosystem, preserving a state of equilibrium. In cases of pulmonary and intestinal injury, particular pathogens can penetrate the epithelial barrier, gain access to the bloodstream, and be recognized by immune cells from both the gastrointestinal and respiratory tracts, thus triggering a cascade of localized and systemic inflammatory responses.9 The epithelial barrier is crucial for maintaining the structural integrity and immune balance within both the gastrointestinal and respiratory tracts.
Short-chain fatty acids (SCFAs), primarily resulting from gut microbial fermentation of dietary fiber, include acetic, propionic, and butyric acids that bolster the intestinal epithelial barrier and modulate immune responses by influencing the functionality and maturation of both local and systemic immune cells.10 Previous research has indicated that mice with ALI exhibit a marked reduction in SCFA concentrations relative to healthy counterparts. Furthermore, contemporary research has revealed that adding SCFAs to drinking water reduces lung inflammation, oxidative stress, and metabolic issues in older mice, while also lessening the severity and inflammatory mechanisms of ALI.11
Consequently, the intestinal microbiota and its derivatives, especially SCFAs, merit further investigation within the framework of the gut-lung axis. C. butyricum, an obligate anaerobe and butyrate-yielding rod bacterium, is consistently found throughout the gastrointestinal tract in both humans and animals. It is capable of breaking down indigestible dietary fiber to yield SCFAs and has been utilized in medical and healthcare settings for many decades.12 Research suggests that C. butyricum not only aids in the repair of damaged intestinal mucosa13 and preserves gut equilibrium but also ameliorates colitis by increasing levels of anti-inflammatory factors in mouse models.14 Hence, we have chosen C. butyricum as a candidate probiotic for the current research.
The Fuzheng jiedu formula (FZJDF) serves as an experiential treatment employed as a complementary medicine for COVID-19, specifically tailored for patients presenting with pneumonia and pronounced deficiency patterns.15 FZJDF comprises eight herbs: Aconitum carmichaelii Debeaux (Danfupian), Zingiber officinale Rosc. (Ganjiang), Glycyrrhiza uralensis Fisch. (Gancao), Lonicera japonica Thunb. (Jinyinhua), Gleditsia sinensis Lam. (Zaojiaoci), Ficus simplicissima Lour. (Wuzhimaotao), Pogostemon cablin (Blanco) Benth. (Guanghuoxiang), and Citrus reticulata Blanco (Chenpi). The side roots of Aconitum carmichaelii Debeaux, a popular herbal remedy in China, Japan, and Korea, are formally listed in the pharmacopeias of these nations.16 In China, “Danfupian” and its derivatives possess a spicy-sweet taste, are characterized by a warm, poisonous quality, and are known for their ability to revive Yang energy, counteract collapse, enhance Fire and Yang elements, expel Cold, and alleviate pain.17 Zingiber officinale Rosc., a prevalent dual-purpose plant in both medicinal and culinary domains, has been historically utilized to alleviate gastrointestinal issues such as queasiness and disgorgement since ancient times.18 Glycyrrhiza uralensis Fisch., which is known for its resolve phlegm and relieve cough, relieve spasm and pain, is listed in the Russian Pharmacopoeia and the American compendium of approved medicinal substances, and has subsequently been included in the official drug registries of numerous countries.19 Prior studies have demonstrated the efficacy of FZJDF in COVID-19 therapy and in animal-derived ALI models, along with reduced herbal toxicity and fewer adverse effects.20 However, microbiota-directed interventions for ALI remain experimental and often lack defined microbial or metabolic targets. We therefore hypothesized that FZJDF alleviates ALI by enriching C. butyricum and thereby increasing gut-derived SCFAs.
Methods and Materials
Materials
FZJDF was manufactured by the Guangzhou university of Chinese Medicine Science and Technology Industrial Park Co., Ltd. (Guangzhou, China). Dexamethasone was procured from Tianjin Xinyi JINjin Pharmaceutical Co. Ltd. (Tianjin, China). Lipopolysaccharide (LPS, L2880) was acquired from Sigma-Aldrich (MO, USA). C. butyricum was obtained from BNCC (Beijing, China). Kits for the enzyme-linked immunosorbent assay (ELISA) of IL-6 and IL-1β were supplied by Jiangsu Enzyme Industry Co., Ltd. (Jiangsu, China). The polyclonal antibody for occludin was purchased from Proteintech Group, Inc. (Chicago, IL, USA). Antibodies against ZO-1, myeloperoxidase, and the secondary antibody goat anti-rabbit IgG H&L (HRP) were sourced from Abcam (Cambridge, UK). Ly-6G (1A8) Rat mAb was obtained from CST (Boston, USA). The C. butyricum FISH Probe (CY3 Labeled) was provided by Guangzhou Exon Biotechnology Co., Ltd (Guangzhou, China).
Preparation and Chemical Profiles of FZJDF
FZJDF (batch number: S20231123) was manufactured by the Guangzhou university of Chinese Medicine Science and Technology Industrial Park Co., Ltd. The preparation followed the method previously described by Lu.20,21
Animals and LPS-Induced ALI
100 specific pathogen-free male Balb/c mice, weighing 20–22 g, were sourced from Beijing Vital River Laboratory Animal Technology Co., Ltd. in Beijing, China. They were housed in specific pathogen-free conditions with a humidity of 55% ± 2%, a temperature of 22 ± 2°C, and a 12-hour light/dark cycle at Guangdong Provincial Hospital of Chinese Medicine. The study complied with international Ethical Guidelines and the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. All animal experiments were implemented following the guidelines approved by the Ethics Committee of Guangzhou University of Chinese Medicine (No. ZYD-2024-162).
The study was executed in three distinct stages. In the initial phase, 42 mice were categorized into six groups: a control, an LPS (0.5 mg/kg), a dexamethasone (DXM, 1.5 mg/kg), and three FZJDF treatment groups (FZJDF_L, 3.3 g/kg; FZJDF_M, 6.6 g/kg; FZJDF_H, 13.2 g/kg) (Figure 1A). Mice acclimated to the experimental setup and diet for 7 days before oral FZJDF administration at specified doses. Control and LPS groups received isotonic saline. After a 7-day dosing period, LPS and treatment groups were intratracheally instilled with LPS, while controls received saline. Mice were euthanized 24 hours post-LPS instillation for tissue collection.
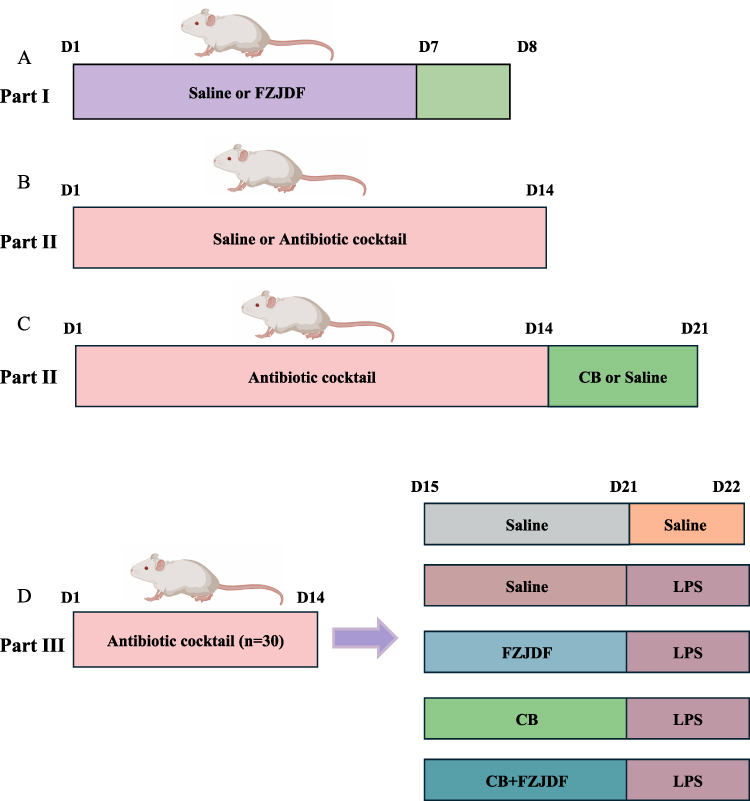 |
Figure 1 Schematic representation of animal experimental design. (A) Part I animal experiments: FZJDF protection against LPS-induced ALI. Mice were gavaged with FZJDF or vehicle for 7 days (D1–D7). On D7, LPS (or saline control) was administered intratracheally; tissues and blood were collected 24 h later (D8). (B) Part II animal experiments: ABX-mediated PGF induction. Animals received a broad-spectrum antibiotic cocktail (ABX) or regular water (NC, n = 6) throughout D1–D14. (C) Part II animal experiments: After ABX treatment (D1–D14), mice were gavaged daily with C. butyricum (CB, n = 8) or vehicle (PGF, n = 8) for 7 days; all samples were collected on day 21 (D21). (D) Part III animal experiments: Mice were rendered PGF with ABX (D1–D14), then received C. butyricum, FZJDF, or vehicle (D15–D21). LPS was instilled on D21; lungs and blood were collected 24 h later (D22). |
In the subsequent phase, as illustrated in the diagrammatic representation of the experimental design (Figure 1B and C), 6 pseudo germ-free (PGF) mice were developed for the antibiotic cocktail (ABX) group by administering a sterilized mixture of antibiotics (0.1mL/10g) including vancomycin (50 mg/kg), neomycin sulfate (100 mg/kg), metronidazole (100 mg/kg), and ampicillin (100 mg/kg) A control group of six mice was given distilled water. After 14 days, mice from both ABX and control groups were euthanized, and tissues were collected. The analysis confirmed the PGF model’s success. The remaining 16 ABX mice were divided into a natural recovery group (given sterile saline orally) and a C. butyricum group (given C. butyricum orally). Each received 0.2 mL of saline or C. butyricum daily for a week. After the 7-day treatment, mice were euthanized, tissues were gathered, and the results confirmed effective C. butyricum colonization.
In the final phase of the study (Figure 1D), 30 mice were systematically allocated to five groups: the pseudo germ-free group (PGF), the LPS group, the LPS+FZJDF group (LPS+FZJDF), the LPS+CB group (C. butyricum + LPS), and the LPS+CB+FZJDF group (C. butyricum + LPS + FZJDF, 13.2 g/kg). The LPS+FZJDF and LPS+CB+FZJDF groups were given 13.2 g/kg FZJDF orally daily for 7 days. Except for the PGF group, all mice underwent intratracheal LPS injections. Twenty-four hours post-LPS, all mice were euthanized, and samples were collected.
C. Butyricum Culture and Treatment
C. butyricum was cultivated under anaerobic conditions in Reinforced Clostridium Medium (RCM) at a temperature of 37°C for an extended period until it reached the exponential growth phase, achieving a bacterial concentration of 0.5 at an optical density of 600 nm (A600). Subsequently, the microbial cells were collected via centrifugation at 8000 g for a duration of 3 minutes and reconstituted in sterile saline solution to achieve the desired experimental concentration of 5×10^8 colony-forming units per milliliter (CFU/mL).22
Histopathological Evaluation (HE)
Lung tissues and colonic samples were procured, rinsed with PBS, and then fixed in a 4% paraformaldehyde solution. Following this, they were dehydrated using a series of ethanol concentrations, impregnated with paraffin, sectioned, and subsequently stained with HE. Pathological changes were examined under a light microscope (OLYMPUS, Tokyo, Japan).
Immunofluorescence
The lung and colon tissue sections, embedded in paraffin, underwent deparaffinized and were rehydrated using a graded series of alcohol solutions. Following antigen retrieval with citrate buffer, the sections were immunostained with appropriate primary and secondary antibodies. Finally, the tissues were counterstained with DAPI and examined using a fluorescence microscope (OLYMPUS, Tokyo, Japan).
Immunohistochemistry
Sections of lung and colon tissues, embedded in paraffin, were cleared of paraffin using xylene and underwent a rehydration process with a sequence of alcohol solutions of escalating concentrations. Following this, the sections were subjected to treatment with citrate buffer to retrieve antigens. Autochthonous peroxidase activity was neutralized with a 10% solution of bovine serum albumin for a duration of 30 minutes, preceding an extended period of incubation with the primary antibody overnight. On the subsequent day, the further incubated with a secondary antibody for a period of 1 hour. Visualization was achieved with DAB reagent, and tissue morphology was assessed using a light microscope (OLYMPUS, Tokyo, Japan).
16S rDNA Gene Sequencing
Upon completion of the experimental treatments, lung tissue and fresh fecal specimens were harvested from the mice and preserved at a temperature of −80°C for 16S rDNA amplicon sequencing. The targeted amplification of a distinct segment of the bacterial 16S rDNA gene was performed utilizing primers that incorporated adapter and barcode sequences, succeeded by a two-stage PCR process. The PCR amplicons were consolidated and subjected to sequencing on the Illumina PE250 sequencing platform utilizing the NovaSeq 6000 instrument (San Diego, CA, USA).
SCFAs Quantification
For the analysis of colonic content, samples were vortexed with distilled water, and the supernatant was extracted and diluted for Gas chromatography-mass spectrometry (GC-MS) analysis using an Agilent 7820 system with a DB-FFAP column (30 m × 0.25 mm × 0.25 μm). The oven temperature was programmed from 70°C to 100°C at 6°C/min, and a quadrupole mass spectrometer was used for SIM detection (m/z 30–550). Data was acquired with MassHunter software and processed with Quant-My-Way (Agilent Technologies).
For serum analysis, samples were mixed with extraction medium, centrifuged, and derivatized with EDC and 3NPH at 40°C for 30 minutes. The extracts were analyzed by Ultra-performance liquid chromatography-Orbitrap mass spectrometer system (UPLC, Vanquish; MS, QE) on a Waters BEH C18 column (50×2.1 mm, 1.8 μm) with a gradient elution program. High-resolution mass spectrometry (HRMS) was performed on a Q Exactive mass spectrometer with heated ESI source (Thermo Fisher Scientific), using Xcalibur software for data acquisition and TraceFinder for processing (Thermo Scientific).
Fluorescence in Situ Hybridization (FISH)
Paraffin-embedded colon tissue sections were prepared for FISH to assess bacterial infiltration. A CY3-tagged probe specific to C. butyricum was used alongside a FITC kit from Guangzhou Exon Biotechnology for detection.
Statistical Analysis
Results are expressed as mean ± SEM. Analysis was performed with GraphPad Prism 9.0 software. Data underwent normality testing, with nonparametric analyses employed for non-normal distributions. Group differences were evaluated using Kruskal–Wallis or one-way ANOVA and pairwise comparisons between groups were conducted using Tukey’s test or Dunnett’s T3 test. A threshold of P < 0.05 was set for statistical significance.
Result
FZJDF Exerts Protective Effect Against LPS-Induced Acute Lung Injury in vivo
In the control group, mouse lung tissues exhibited a pale pink appearance, with no evident hemorrhagic spots or ecchymosis, and a soft texture. Conversely, LPS stimulation caused the lung tissue to become dark red, with surface hyperemia, visible hemorrhagic spots, ecchymosis, and a slightly firmer texture. These alterations were ameliorated following treatment with FZJDF and DXM (Figure 2A). Additionally, pulmonary edema was quantified by measuring the ratio of lung-to-body weight ratio, which was elevated in LPS-treated mice and reduced in FZJDF-treated animals (Figure 2B). Serum levels of key inflammatory cytokines in ALI pathogenesis were measured by ELISA. LPS-induced ALI markedly increased IL-1β and IL-6 levels, which were reduced by FZJDF and DXM, with the highest FZJDF dose showing the greatest reduction (Figure 2C).
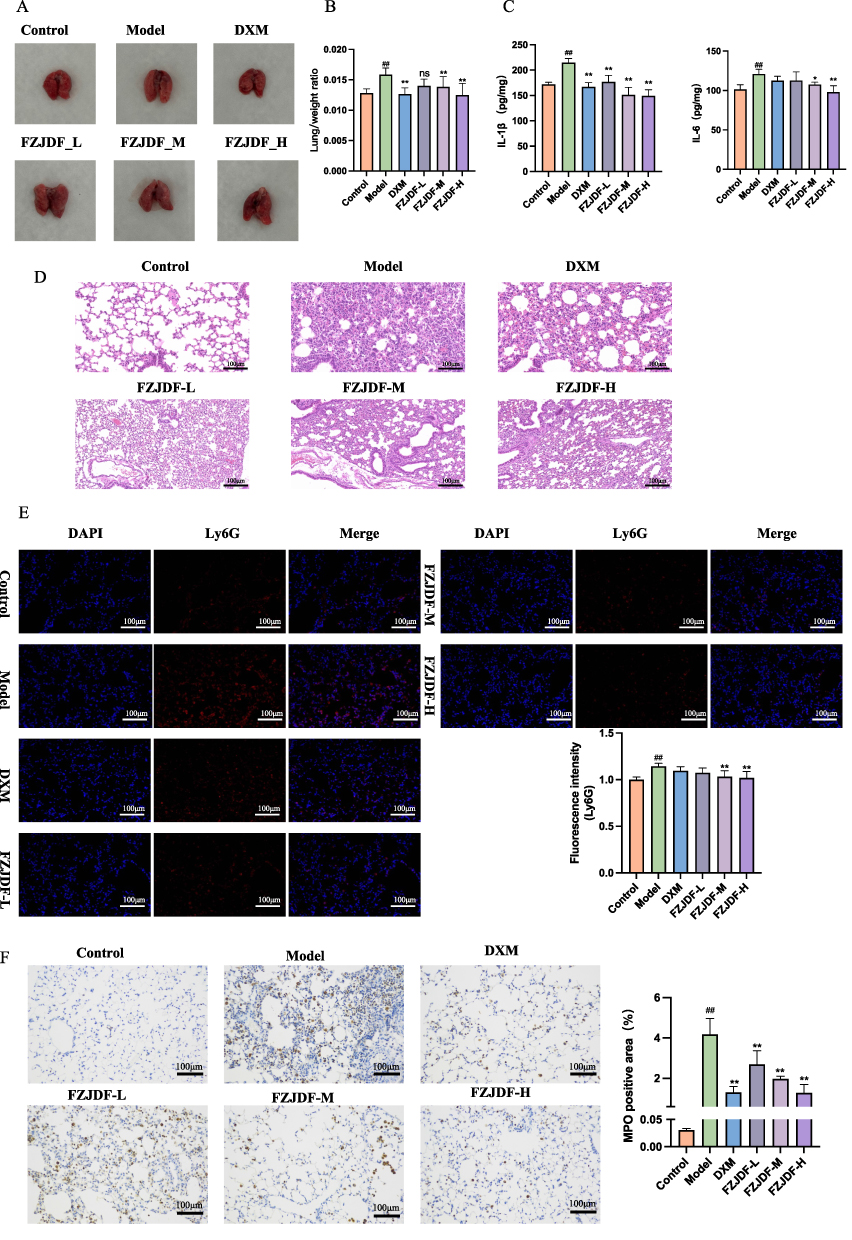 |
Figure 2 FZJDF treatment ameliorated LPS-induced ALI (A) Appearance of lung tissues in each group. (B) Lung/weight ratio in each group (n=6). (C) Serum level of IL-1β and IL-6 in each group (n=5). (D) H&E staining of the lungs (×200). (E) Ly6G were observed by immunofluorescence (×200, n=5, red: Ly6G-positive, blue: DAPI). (F) MPO were observed by Immunohistochemistry (×200, n=5). The data are mean ± SD. FZJDF_L, low dose of FZJDF (3.3g/kg); FZJDF_M, medium dose of FZJDF (6.6g/kg); FZJDF_H, high dose of FZJDF (13.2g/kg); ##P<0.01 vs the control group; *P<0.05, **P<0.01 vs the model group; ns, not significant. |
Histopathological analysis of FZJDF-treated LPS-induced ALI mice showed that the model group exhibited interstitial edema, alveolar wall thickening, and substantial inflammatory cell infiltration. FZJDF and DXM treatment alleviated these changes, with high-dose FZJDF being most effective (Figure 2D). Furthermore, immunofluorescence and immunohistochemistry were performed using anti-Ly6G and anti-MPO antibodies to assess neutrophil infiltration in the lungs (Figure 2E and F). Staining results revealed that LPS treatment significantly increased neutrophil recruitment, whereas FZJDF treatment notably reduced neutrophil infiltration. These findings suggest that high-dose FZJDF effectively maintains alveolar-vascular barrier integrity and reduces excessive pulmonary inflammation in ALI mice.
FZJDF Enhances the Impaired Intestinal Barrier Function in LPS-Induced ALI Mice
Inflammation systemic to the body, stemming from pulmonary damage, impaired the intestinal barrier’s continuity, thus enabling the migration of microorganisms and their byproducts. HE staining revealed that the administration of FZJDF maintained the structural integrity of the colonic epithelium in mice (Figure 3A). Concurrently, the abundance of intestinal tight junction proteins was assessed using immunofluorescence and immunohistochemistry (Figure 3B and C). Analysis revealed that LPS exposure significantly decreased ZO-1 and Occludin expression compared to the control group. Notably, FZJDF treatment significantly upregulated the expression of these proteins compared to the model group. Collectively, these findings indicate that FZJDF ameliorates intestinal barrier dysfunction, which is closely associated with the translocation of gut microbiota.
 |
Figure 3 FZJDF can improve the impaired intestinal barrier function in LPS-induced ALI mice (A) H&E staining of colonic tissue (red arrows indicated the infiltration of inflammatory cells ×200). (B) ZO-1 was observed by immunofluorescence (×200, n=5, red: ZO-1-positive, blue: DAPI). (C) Occludin were observed by Immunohistochemistry (×200, n=5). The data are mean ± SD. FZJDF_L, low dose of FZJDF (3.3g/kg); FZJDF_M, medium dose of FZJDF (6.6g/kg); FZJDF_H, high dose of FZJDF (13.2g/kg); ##P<0.01 vs the control group. *P<0.05, **P<0.01 vs the model group. |
Effect of FZJDF on the Intestinal Microflora of LPS-Induced ALI Mice
The lung-gut axis hypothesis suggests that intestinal microbiota changes can markedly affect pulmonary conditions, suggesting a regulatory role of the gut microbiota in lung disease progression. Accordingly, this study examined the effects of FZJDF on the gut microbiome in LPS-induced ALI mice. Results showed that FZJDF supplementation significantly increased the diversity and abundance of fecal microbes reduced in the model group (Figure 4A). Principal coordinate analysis (PCoA) based on Bray-Curtis distances indicated that LPS disrupted the gut microbiome, an effect mitigated by FZJDF (Figure 4B).
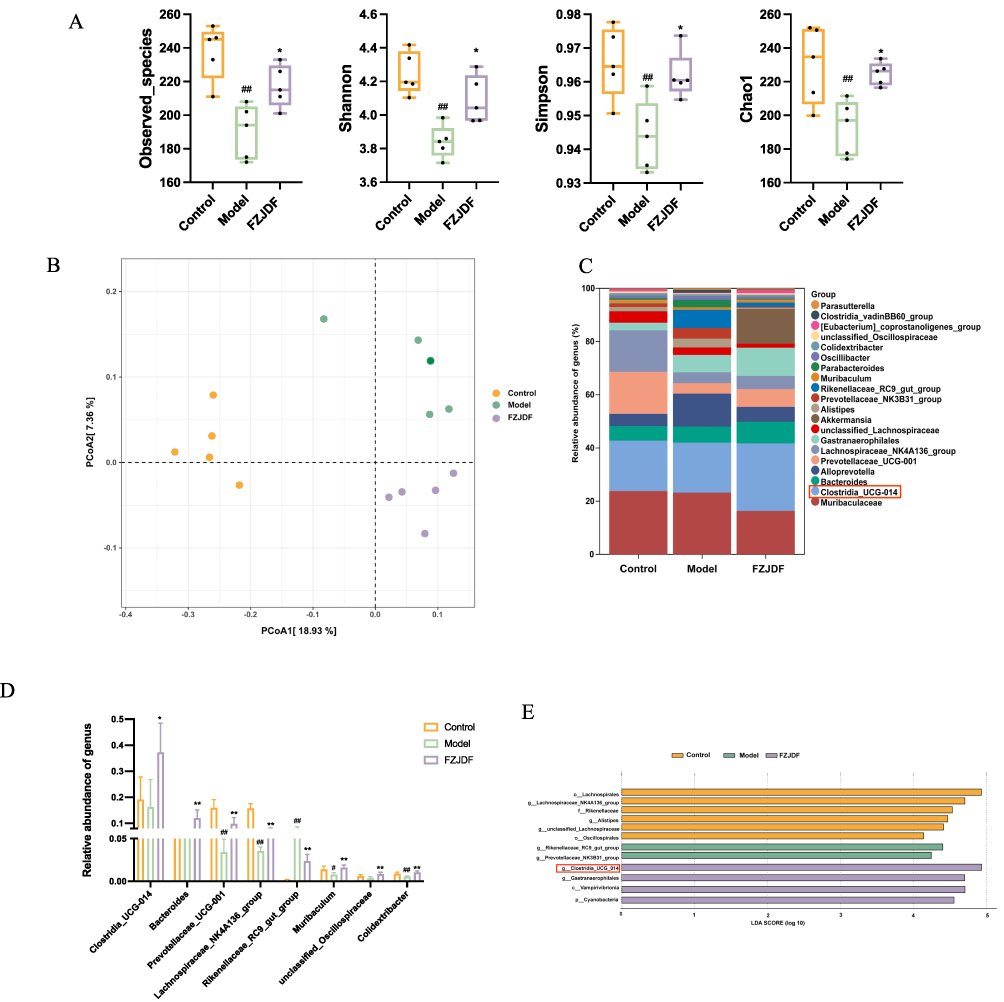 |
Figure 4 Effect of FZJDF on the intestinal microflora of LPS-induced ALI mice. (A) Alpha diversity analysis of gut bacterial richness from different group. (B) PCoA analysis of intestinal flora in different groups. (C) Community structure analysis of each group at the genus level. (D). Relative abundance of the genus levels. (E) LEfSe analysis of the control, model, and FZJDF groups. The data are mean ± SD (n=5). FZJDF_L, low dose of FZJDF (3.3g/kg); FZJDF_M, medium dose of FZJDF (6.6g/kg); FZJDF_H, high dose of FZJDF (13.2g/kg); Clostridia_UCG_014 indicated by the red rectangle box, #P<0.05, ##P<0.01 vs the control group. *P<0.05, **P<0.01 vs the model group. |
To further explore the role of microbiota in FZJDF’s effects on ALI, a taxonomic profiling of bacteria at the genus level was conducted. The model group had a higher prevalence of Rikenellaceae_RC9_gut_group and lower levels of Prevotellaceae_UCG_001, Lachnospiraceae_NK4A136_group, Colidextribacter, unclassified_Oscillospiraceae, and Clostridium_UCG_014, which were more abundant in the FZJDF group, with Clostridium_UCG_014 showing a significant increase (Figure 4C and D). LEfSe analysis confirmed the substantial difference in Clostridium _UCG_014 relative abundance between the FZJDF and model groups (Figure 4E).
Effect of FZJDF on the Lung Microflora of LPS-Induced ALI Mice
To explore the correlation between lung microbiota alterations and LPS-induced lung injury, a 16S rDNA analysis was conducted to assess microbial diversity in control, model, and FZJDF-treated groups. No significant differences were found in species richness or diversity indices such as Shannon, Simpson, and Chao1 among the groups. However, non-metric multidimensional scaling (NMDS) based on weighted UniFrac distances revealed clear distinctions between group microbial compositions (Figure 5A). The Firmicutes/Bacteroidetes ratio, indicative of inflammation, was lower in the model group than in controls (Figures 5B and C). FZJDF treatment significantly increased this ratio, suggesting a potential therapeutic effect.
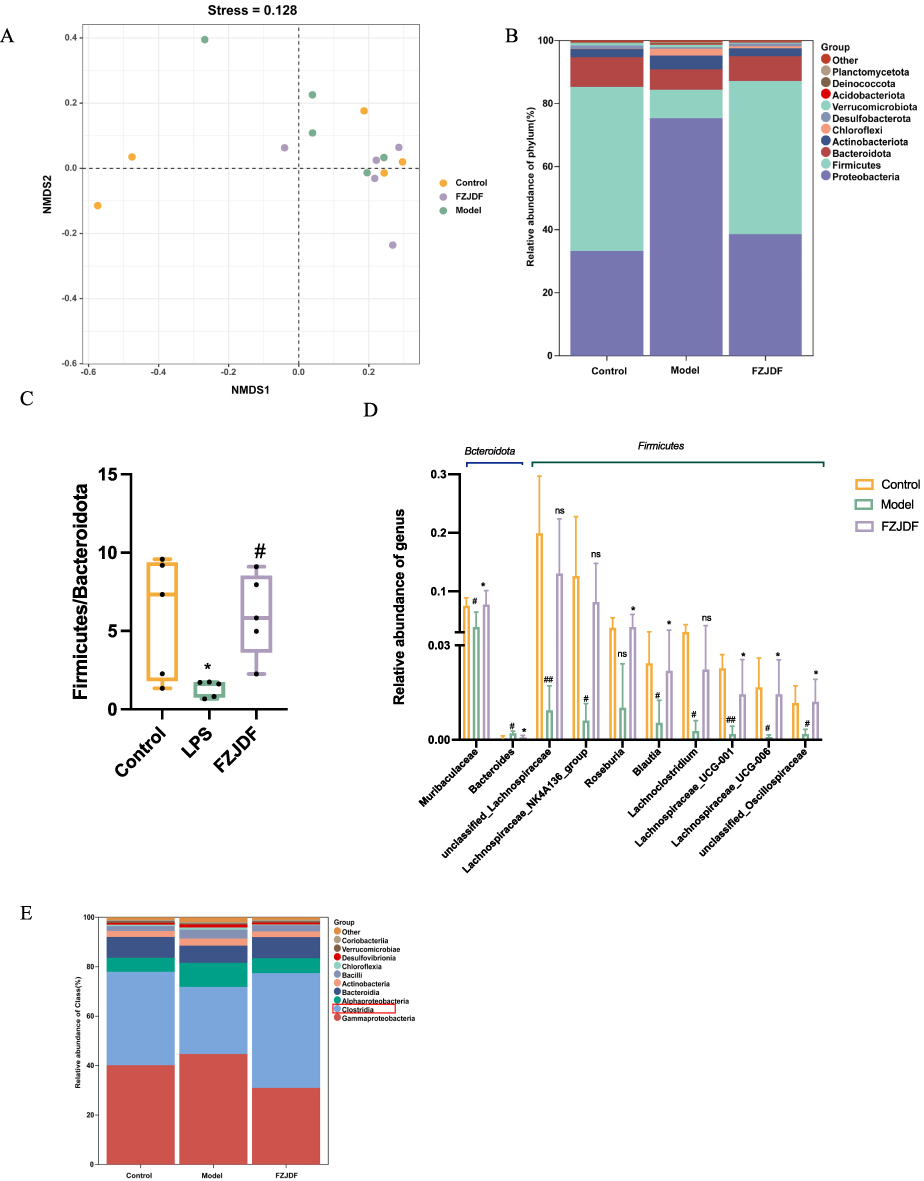 |
Figure 5 Effect of FZJDF on the lung microflora of LPS-induced ALI mice. (A) PCoA analysis of lung microflora. (B) Community structure analysis of each group at the Phylum level. (C) the relative ratio between Firmicutes and Bacteroidetes. (D) Representative species of Firmicutes family and Bacteroidetes family in lung microbial. (E) Community structure analysis of each group at the Class level. The data are mean ± SD (n=5). FZJDF_L, low dose of FZJDF (3.3g/kg); FZJDF_M, medium dose of FZJDF (6.6g/kg); FZJDF_H, high dose of FZJDF (13.2g/kg); Clostridia indicated by the red rectangle box, #P<0.05, ##P<0.01 vs the control group; *P<0.05, vs the model group; ns, not significant. |
At the genus level, the model group exhibited reduced relative abundances of Muribaculaceae, unclassified_Lachnospiraceae, Lachnospiraceae_NK4A136_group, Roseburia, Blautia, Lachnoclostridium, Lachnospiraceae_UCG-001, Lachnospiraceae_UCG-006, unclassified_Oscillospiraceae, and an increase in Bacteroides. Conversely, the FZJDF group had increased relative abundances of Muribaculaceae, Roseburia, Blautia, Lachnospiraceae_UCG-001, Lachnospiraceae_UCG-006, unclassified_Oscillospiraceae, and a decrease in Bacteroides relative to the model group (Figure 5D). Additionally, class-level changes in Clostridium relative abundance were noted (Figure 5E).
FZJDF Enhanced the Production of SCFAs Through Modulation of the Gut Microbiota
C. butyricum, a member of the genus Clostridium, ferments indigestible dietary fibers into SCFAs, which are recognized for their role in preserving intestinal homeostasis. We conducted an analysis of SCFA concentrations in the intestinal contents, serum, and lung tissue of mice (Figure 6). Notably, the SCFA content in the FZJDF group were markedly elevated in comparison to the model group implying that SCFAs originating from C. butyricum might exert a significant influence on the therapeutic effectiveness of FZJDF in alleviating LPS-induced ALI in mice.
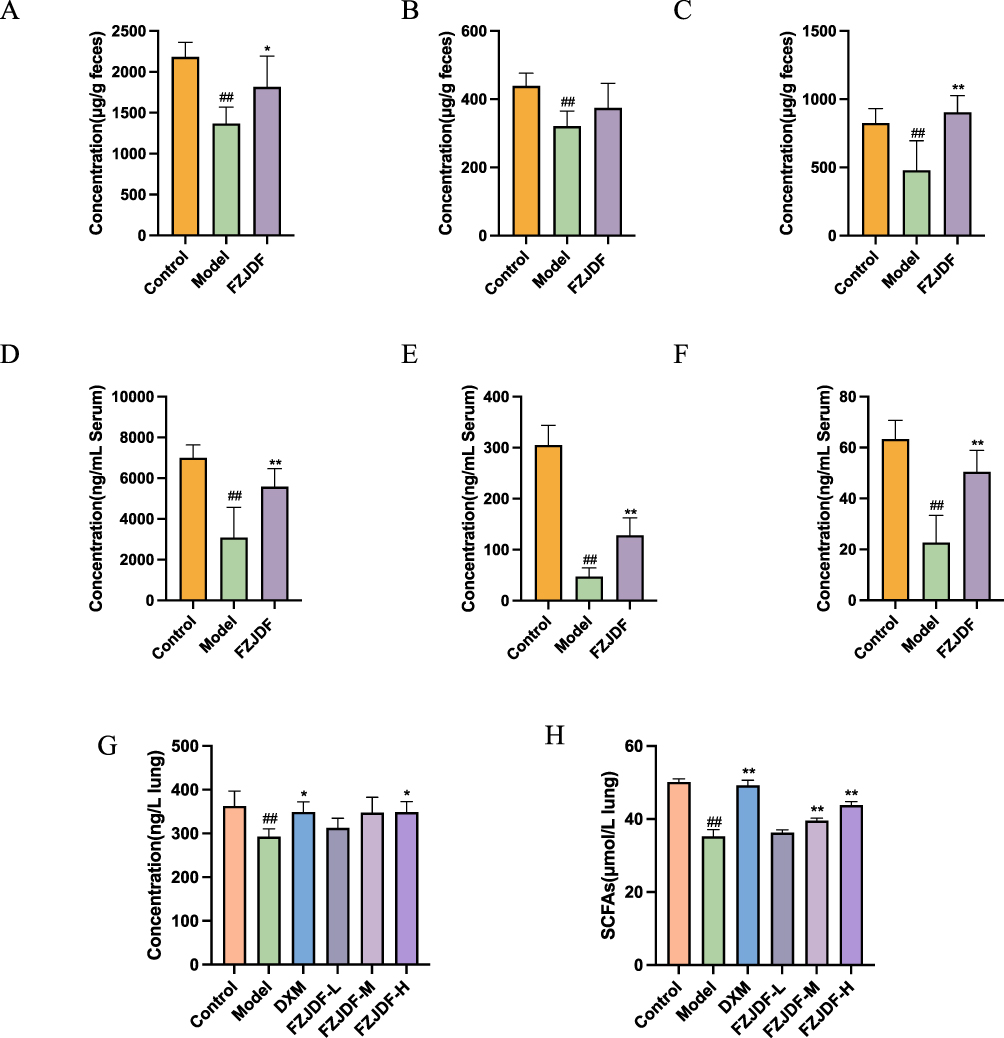 |
Figure 6 FZJDF regulates intestinal microbiota to promote the production of SCFAs. (A) The concentration of acetic acid in intestinal contents of mice (B) The concentration of propionic acid in intestinal contents of mice. (C) The concentration of butyric acid in intestinal contents of mice. (D) The concentration of acetic acid in serum of mice. (E) The concentration of propionic acid in serum of mice. (F) The concentration of butyric acid in serum of mice. (G) The concentration of butyric acid in lung of mice. (H) The concentration of SCFAs in lung of mice. The data are mean ± SD (n=5). FZJDF_L, low dose of FZJDF (3.3g/kg); FZJDF_M, medium dose of FZJDF (6.6g/kg); FZJDF_H, high dose of FZJDF (13.2g/kg); ##P<0.01 vs the control group. *P<0.05, **P<0.01 vs the model group. |
FZJDF Attenuated LPS-Induced ALI Mice by Targeting C. Butyricum
To confirm the role of C. butyricum in FZJDF’s therapeutic effects, ALI was induced in PGF mice after their intestines were colonized with C. butyricum. The impact of FZJDF on ALI was evaluated using the Observed OTUs index and PCoA to measure diversity of the gut microbiota (Figure 7A and B). The results validated the PGF mice model and demonstrated that C. butyricum could efficiently colonize the intestinal tract through oral gavage (Figure 7C).
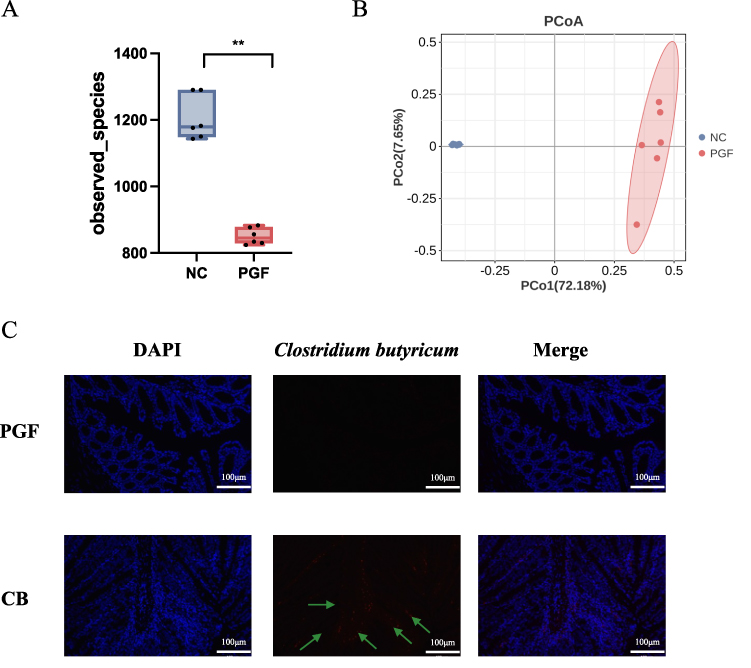 |
Figure 7 Confirmation of the PGF mice model and establishment of C. butyricum in the colonic microbiome. (A) Alpha diversity assessment using the Observed index. (B) PCoA depicting the composition of the intestinal microbiota. (C) Detection of C. butyricum colonization in the murine colon via FISH (×200), (C. butyricum indicated by the green arrows); The data are mean ± SD (n=6). **P<0.01 vs the PGF group. |
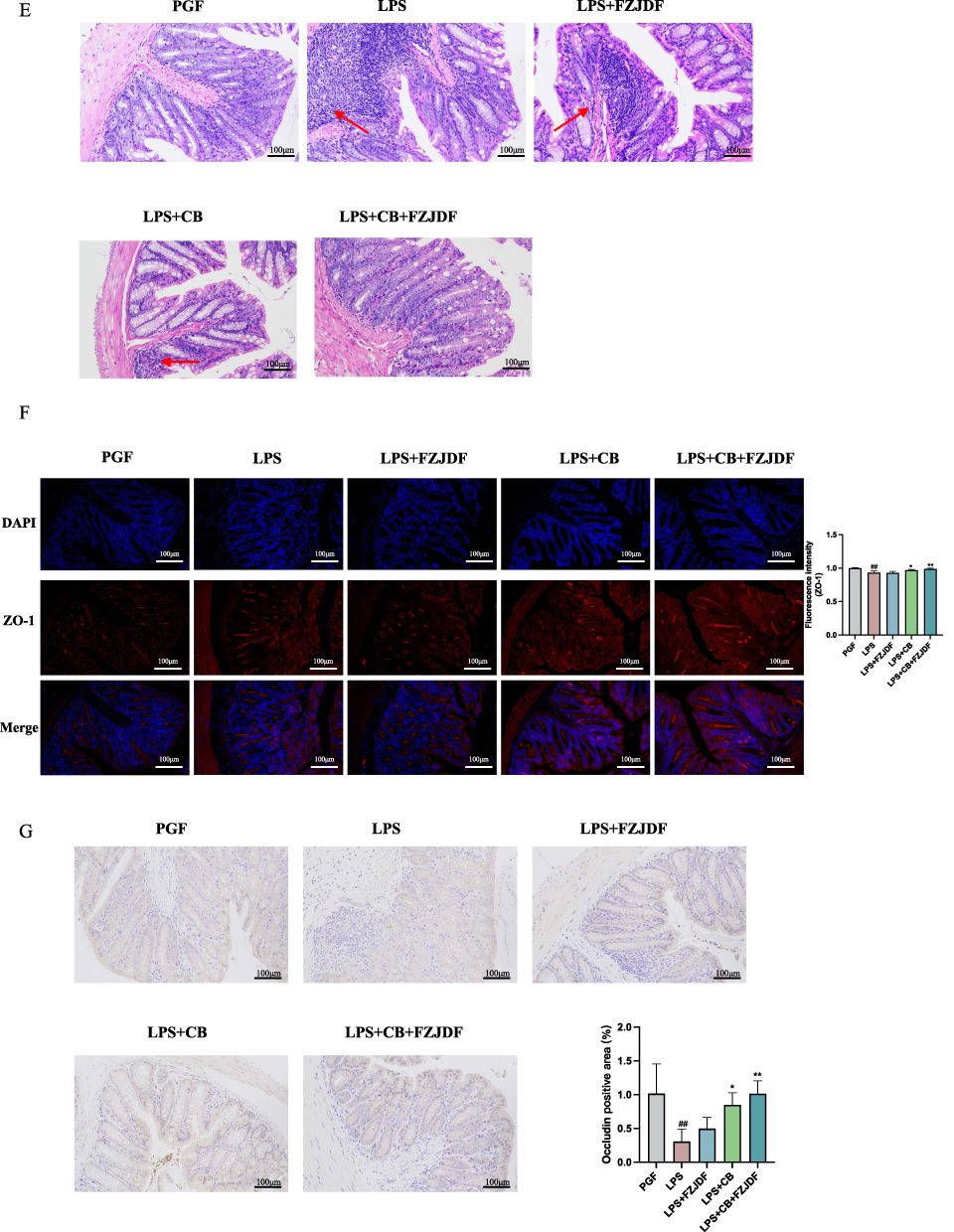
Subsequent examination of the ALI model revealed that the advantageous impacts of FZJDF, including decreases in inflammatory biomarkers and pathological damage, were nearly abolished in the deficiency of intestinal microflora (Figure 8A–D). Moreover, FZJDF treatment influenced the levels of tight junction proteins in mouse intestinal epithelial cells, with no marked differences observed compared to the LPS group (Figure 8E and G). On the contrary, LPS+CB+FZJDF group not only mitigated ALI symptoms but also rejuvenated the intestinal barrier integrity in mice. Concurrently, FZJDF elevated the levels of SCFAs in mice inhabited by C. butyricum (Figure 9). These observations suggest that the therapeutic efficacy of FJZDF on ALI is dependent on the presence of C. butyricum, with FZJDF augmenting the SCFA production by C. butyricum, thereby ameliorating lung inflammation and repairing damaged intestinal mucosa.
Figure 8 continued. Figure 8 FZJDF attenuated LPS-induced ALI mice by targeting C. butyricum. (A) H&E staining of the lungs (×200). (B) Serum level of IL-1β and IL-6 in each group. (C) Ly6G were observed by immunofluorescence (×200, red: Ly6G-positive, blue: DAPI). (D) MPO were observed by Immunohistochemistry (×200). (E) H&E staining of colonic tissue (×200); (The inflammation indicated by the red arrows). (F) ZO-1 was observed by immunofluorescence (×200, red: ZO-1-positive, blue: DAPI). (G) Occludin were observed by Immunohistochemistry (×200). The data are mean ± SD (n=5). LPS+FZJDF, LPS (0.5 mg/kg) + high dose of FZJDF (13.2g/kg); LPS+CB, LPS (0.5 mg/kg) + C. butyricum; LPS+CB+FZJDF, (0.5 mg/kg) + C. butyricum + high dose of FZJDF (13.2g/kg); ##P<0.01 vs the PGF group. *P<0.05, **P<0.01 vs the LPS group.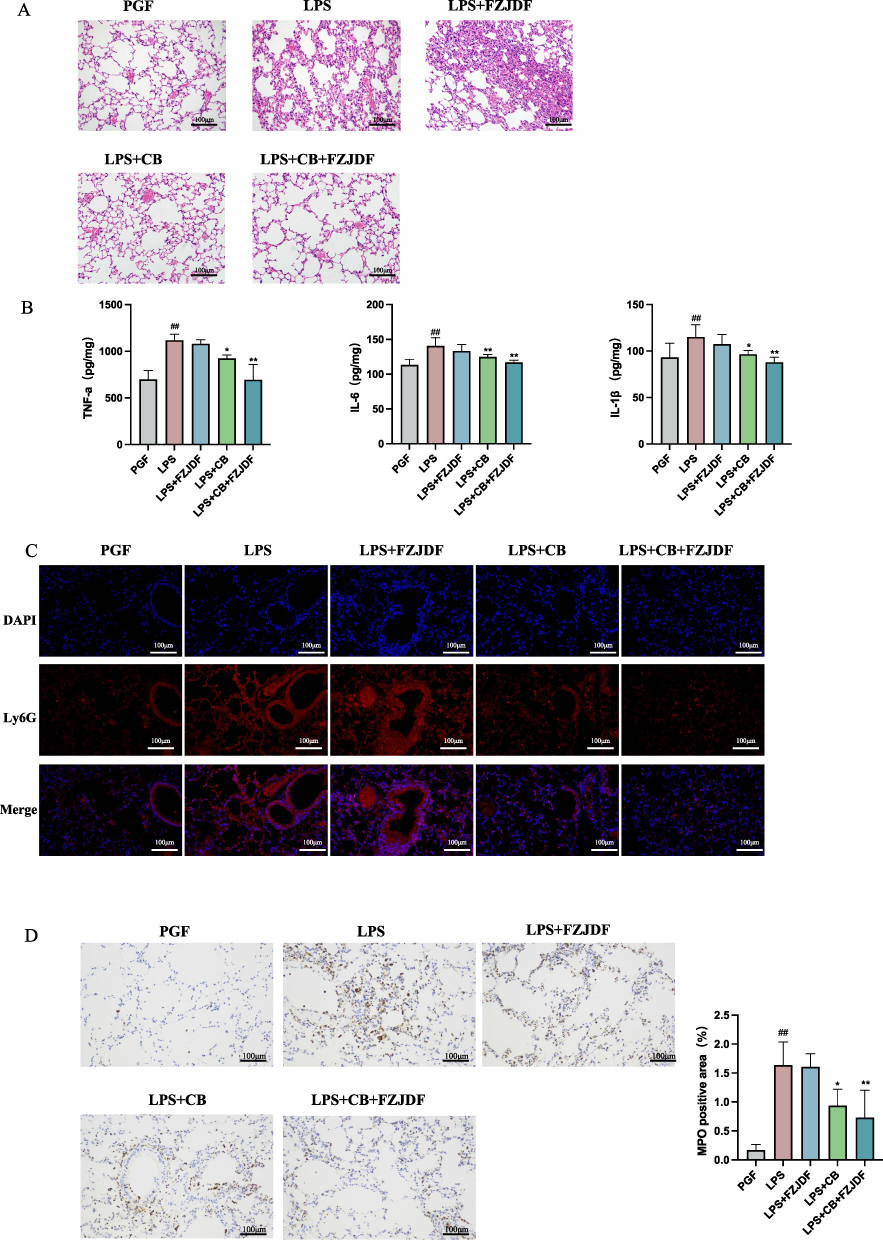
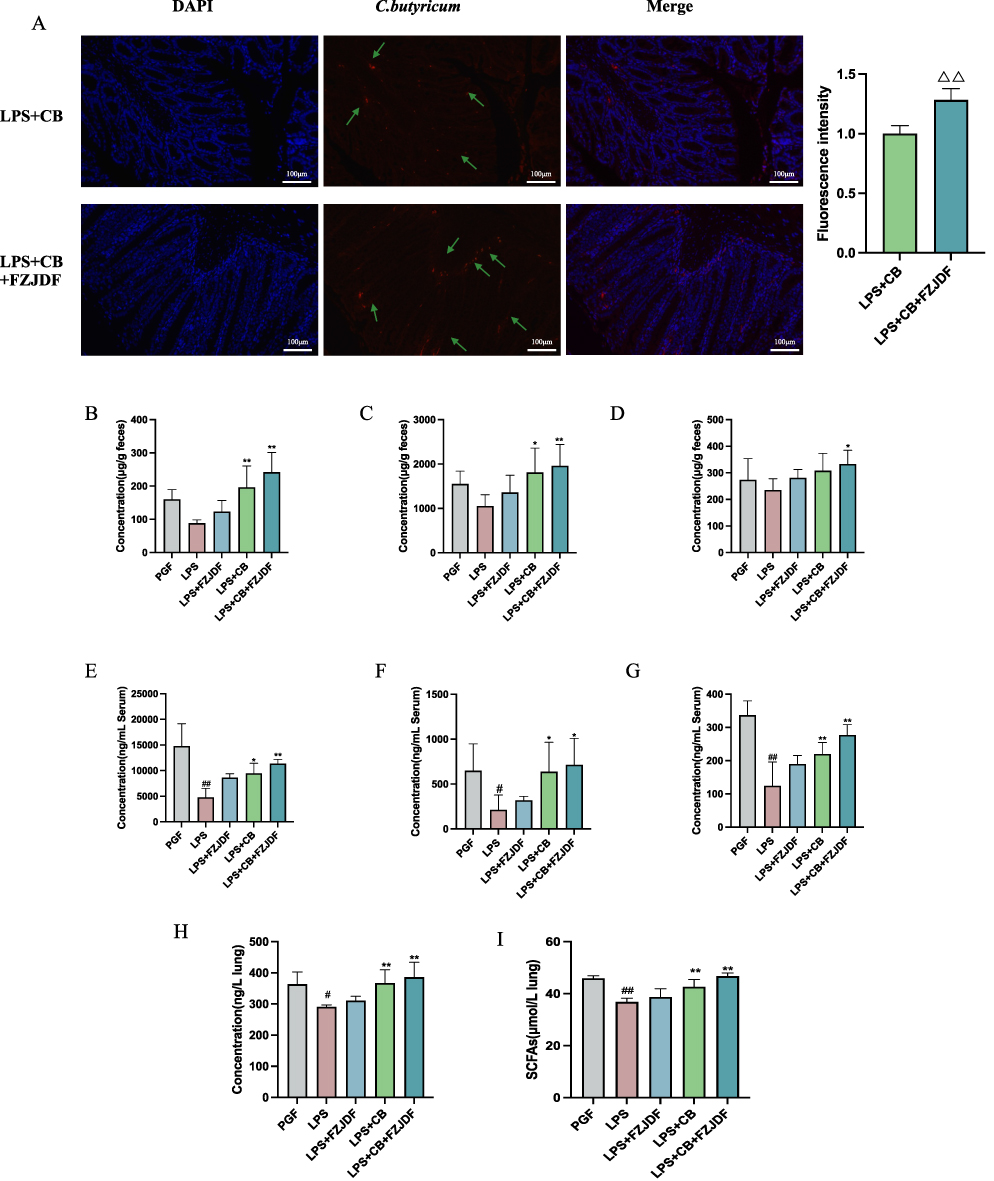 |
Figure 9 FZJDF enhances the proliferation of C. butyricum and the production of SCFAs in PGF mice. (A) FISH was employed to quantify the presence of C. butyricum in the colon of mice; (C. butyricum indicated by the green arrows). (B) The concentration of acetic acid within the intestinal contents of mice. (C) The concentration of propionic acid within the intestinal contents of mice. (D) The concentration of butyric acid within the intestinal contents of mice. (E) The concentration of acetic acid in serum of mice. (F) The concentration of propionic acid in serum of mice. (G) The concentration of butyric acid in serum of mice. (H) The concentration of butyric acid in lung of mice. (I) The concentration of SCFAs in lung of mice. The data are mean ± SD (n=5). LPS+FZJDF, LPS (0.5 mg/kg) + high dose of FZJDF (13.2g/kg); LPS+CB, LPS (0.5 mg/kg) + C. butyricum; LPS+CB+FZJDF, (15mg/kg) + C. butyricum + high dose of FZJDF (13.2g/kg); ΔΔP<0.01 vs the LPS+CB group; #P<0.05, ##P<0.01 vs the PGF group; *P<0.05, **P<0.01 vs the LPS group. |
Discussion
Inflammatory modulators, specifically proinflammatory cytokines, are pivotal in the etiology of ALI and ARDS. Such mediators induce extensive impairment of alveolar epithelial and vascular endothelial cells, escalate vascular permeability, and encourage protein seepage, thus intensifying lung edema.23,24 In our investigation, HE staining revealed that LPS exposure resulted in significant alveolar and interstitial damage in mouse lung tissue. Nonetheless, intervention with FZJDF alleviated alveolar and interstitial edema and curtailed the infiltration of inflammatory cells.
Neutrophils, the initial immune responders at sites of inflammation, phagocytize and eliminate bacteria and fungi, and trigger the mobilization and activation of additional immune cells.25 SCFAs produced by the microbiota influence neutrophil activity and monocyte recruitment in areas of inflammation by regulating the synthesis of inflammatory mediators, making neutrophils an important source of mediators.26 In line with prior research,27 FZJDF administration reduced the concentrations of inflammatory factors, thereby mitigating lung tissue damage. Immunofluorescence assays using anti-MPO and anti-Ly6G antibodies on lung tissues confirmed elevated neutrophil infiltration in ALI mice, with the FZJDF group showing diminished inflammatory cell counts.
Numerous investigations have demonstrated that inflammation throughout the body, triggered by pulmonary damage, can result in heightened intestinal permeability and compromise the intestinal barrier. This heightened permeability allows microbiota and their metabolites to translocate into the lungs, potentially exacerbating lung disease.28 The crosstalk between the gut29 and the lungs30 is strongly correlated with the severity of lung disease, primarily directed from the gut to the lungs, though bidirectional communication is also possible. Intestinal epithelial cells, linked through tight junctions, constitute the initial defense mechanism against microbial invasion by establishing a physical barrier. The intestinal integrity relies on both physical and microbial barriers.31 Previous research has confirmed that the tight junction proteins exhibit decreased expression in LPS-induced mice.32 Our results demonstrate that FZJDF administration increases the levels of these proteins, decreases intestinal permeability, and repairs the intestinal barrier function.
A substantial body of research indicates that the respiratory microbiome is crucial for various pulmonary diseases, encompassing idiopathic pulmonary fibrosis, asthma, and lung cancer.33 Notably, lung flora composition shows a significant increase in Firmicutes following treatment, whereas LPS exposure markedly elevates Proteobacteria abundance.34 Our findings corroborate previous studies, indicating that FZJDF treatment increases the abundance of Firmicutes. The microbial barrier, primarily composed of intestinal microbiota, maintains intestinal mucosal homeostasis and prevents pathogenic bacterial invasion, remaining relatively stable under normal conditions.35 Thus, we hypothesized that FZJDF might regulate the gut microbiota and embarked on pertinent research endeavors. Previous research has shown that inflammatory processes, immune reactions, and additional elements can perturb the intestinal microbiota, diminishing its diversity and augmenting susceptibility to detrimental bacteria.36 In line with these observations, our 16S rDNA sequencing revealed a significant decrease in both the richness and diversity of the gut microbiome in ALI mice. In FZJDF-treated mice, the dominant genera in fecal samples included Clostridia, Gastranaerophilales, Vampirivibrionia, and Cyanobacteria, with a significant increase in Clostridia_UCG_014.
A large body of literature has established that C. butyricum, a bacterium of the Clostridium genus, promotes the production of butyrate among SCFAs.37 As a probiotic inhabiting the gastrointestinal tract, C. butyricum exerts beneficial effects by modulating the intestinal microbiota and curbing inflammation.38,39 For example, C. butyricum strain MIYAIRI 588 increases the quantity of beneficial microbes, enhances intestinal homeostasis, and correlates positively with the effectiveness of immunotherapy in individuals with advanced non-small cell lung cancer.40 Additionally, C. butyricum has been shown to mitigate lung inflammation and modulate immune responses by promoting M2 polarization of pulmonary macrophages, diminishing the secretion of inflammatory factors.41 In summary, C. butyricum exhibits promise as a protective and therapeutic agent in pulmonary pathologies.
Prior research22 has demonstrated that C. butyricum possesses considerable promise in facilitating the regeneration of intestinal epithelial cells, reshaping the intestinal microbiota, and suppressing the synthesis of pro-inflammatory cytokines. Consequently, C. butyricum may be pivotal for the efficacy of FZJDF in alleviating ALI. Our investigation revealed that FZJDF diminished both respiratory and systemic inflammatory responses, as indicated by lower levels of pro-inflammatory cytokines. Furthermore, by supplying energy to colonic cells, FZJDF effectively protected the intestinal mucosa from bacterial translocation into the bloodstream, thus preventing the onset of inflammatory reactions. Given that the primary metabolic products of C. butyricum are SCFAs, we also scrutinized changes in SCFA levels using GC-MS. The results indicated that FZJDF increased SCFA levels. Upon in-depth analysis of the ALI model, we found that the potential benefits of FZJDF, including the reduction of pro-inflammatory markers and attenuation of pathological damage, were significantly attenuated under conditions lacking a gut microbiota. However, in the LPS+CB+FZJDF group, FZJDF not only significantly ameliorated ALI symptoms in C. butyricum-colonized mice, but also promoted the restoration of intestinal barrier function. In addition, FZJDF treatment significantly increased the concentration of SCFAs in C. butyricum colonized mice.
Our findings suggest that FZJDF may contribute to alleviating ALI by modulating the gut microbiota, potentially enhancing the production of SCFAs such as butyrate, which may in turn help suppress neutrophil-mediated inflammatory responses. This finding provides new insights into the potential mechanisms by which FZJDF regulates intestinal flora and ameliorates ALI. This study has certain limitations, particularly regarding the precise mechanisms by which FZJDF exerts its effects. While our findings suggest that FZJDF alleviates ALI by modulating the gut microbiota to SCFA production, it remains unclear whether these effects are primarily mediated through GPR43 signaling. Future studies will expand the sample size, integrate clinical trials to verify the transformation potential, and use receptor-specific inhibition or knockout models to analyze the signal transduction process mediated by GPR43, and clarify the more in-depth mechanism research of FZJDF in the relationship between microbial community changes and immune regulation.
Conclusion
Our research has revealed that FZJDF can alleviate ALI by reshaping the gut microbiota and increasing SCFA levels, which may consequently dampen neutrophil-driven pulmonary inflammation. While these pre-clinical findings identify a promising microbiota–metabolite–immune axis, they require validation in human cohorts. Ongoing work will integrate these basic observations with clinical studies to elucidate the deeper mechanisms of FZJDF and to evaluate its therapeutic potential in patients.
Abbreviations
ALI, Acute lung injury; ARDS, Acute respiratory distress syndrome; ABX, antibiotic cocktail; C. butyricum, Clostridium butyricum; DXM, Dexamethasone; ELISA, Enzyme-linked immunosorbent assay; FISH, Fluorescence in situ hybridization. GC-MS, Gas chromatography-mass spectrometry; HE, Histopathological evaluation; HRMS, High-resolution mass spectrometry; LPS, Lipopolysaccharide; NMDS, Non-metric multidimensional scaling; PGF, Pseudo germ-free; SCFAs, short-chain fatty acids; UPLC, Ultra-performance liquid chromatography.
Data Sharing Statement
Data supporting the findings of this study will be made available upon reasonable request on Yuntao Liu ([email protected]).
Ethics Statement
All animal experiments were implemented following the guidelines approved by the Ethics Committee of Guangzhou University of Chinese Medicine (NO. ZYD-2024-162).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by National Natural Science Foundation of China (U25A20157, 82374392), the National Science and Technology Major Project for the Prevention and Control of New and Major Infectious Diseases (2025ZD01902900, 2025ZD01903002), Science and Technology Planning Project of Guangdong Province (2023B1212060062), Guangdong Provincial Natural Science Foundation (2025A1515011169), Basic and Applied Basic Research of Guangzhou City-University Joint Funding Project (2025A03J4129), Disciplinary Project of Guangzhou University of Chinese Medicine (GZY2025GB0301, GZY2025GB0305, GZY2025GB0410), Open project of Guangdong Provincial Key Laboratory of Research on Emergency in TCM (KF2023JZ06), the Incubation Program for the Science and Technology Development of Chinese Medicine Guangdong Laboratory (HQL2024PZ022), the “Challenge and Response” Postgraduate Innovation Capacity Enhancement Project of the Second Clinical College of Guangzhou University of Chinese Medicine (Document No. 269 of the Office of Guangzhou University of Chinese Medicine, 2024), Special Fund for National Natural Science Research of Guangdong Provincial Hospital of Traditional Chinese Medicine (No. YN2025GZR47).
Disclosure
The authors declare no conflicts of interest in relation to this article.
References
1. Bos LDJ, Ware LB. Acute respiratory distress syndrome: causes, pathophysiology, and phenotypes. Lancet. 2022;400(10358):1145–20. doi:10.1016/S0140-6736(22)01485-4
2. Meyer NJ, Gattinoni L, Calfee CS. Acute respiratory distress syndrome. Lancet. 2021;398(10300):622–637. doi:10.1016/S0140-6736(21)00439-6
3. Wheeler AP, Bernard GR. Acute lung injury and the acute respiratory distress syndrome: a clinical review. Lancet. 2007;369(9572):1553–1564. doi:10.1016/S0140-6736(07)60604-7
4. Spinelli E, Mauri T, Beitler JR, Pesenti A, Brodie D. Respiratory drive in the acute respiratory distress syndrome: pathophysiology, monitoring, and therapeutic interventions. Intensive Care Med. 2020;46(4):606–618. doi:10.1007/s00134-020-05942-6
5. Mokrá D. Acute lung injury - from pathophysiology to treatment. Physiol Res. 2020;69(Suppl 3):S353–s366. doi:10.33549/physiolres.934602
6. Hadaya J, Benharash P. Prone positioning for acute respiratory distress syndrome (ARDS. JAMA. 2020;324(13):1361. doi:10.1001/jama.2020.14901
7. Kaku S, Nguyen CD, Htet NN, et al. Acute respiratory distress syndrome: etiology, pathogenesis, and summary on management. J Intensive Care Med. 2020;35(8):723–737. doi:10.1177/0885066619855021
8. Ma PJ, Wang MM, Wang Y. Gut microbiota: a new insight into lung diseases. Biomed Pharmacother. 2022;155:113810. doi:10.1016/j.biopha.2022.113810
9. Enaud R, Prevel R, Ciarlo E, et al. The gut-lung axis in health and respiratory diseases: a place for inter-organ and inter-kingdom crosstalks. Front Cell Infect Microbiol. 2020;10:9. doi:10.3389/fcimb.2020.00009
10. Tan JK, Macia L, Mackay CR. Dietary fiber and SCFAs in the regulation of mucosal immunity. J Allergy Clin Immunol. 2023;151(2):361–370. doi:10.1016/j.jaci.2022.11.007
11. Hildebrand CB, Lichatz R, Pich A, et al. Short-chain fatty acids improve inflamm-aging and acute lung injury in old mice. Am J Physiol Lung Cell Mol Physiol. 2023;324(4):L480–l492. doi:10.1152/ajplung.00296.2022
12. Chen D, Jin D, Huang S, et al. Clostridium butyricum, a butyrate-producing probiotic, inhibits intestinal tumor development through modulating Wnt signaling and gut microbiota. Cancer Lett. 2020;469:456–467. doi:10.1016/j.canlet.2019.11.019
13. Stoeva MK, Garcia-So J, Justice N, et al. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes. 2021;13(1):1–28. doi:10.1080/19490976.2021.1907272
14. Xie Y, Zhou L, Yao X, Li Y. Protective effects of clostridium butyricum in a murine model of dextran sodium sulfate-induced colitis that involve inhibition of the tlr2 signaling pathway and t helper 17 cells. Am J Med Sci. 2020;360(2):176–191. doi:10.1016/j.amjms.2020.05.021
15. Wang Y, Liu Y, Lv Q, et al. Effect and safety of Chinese herbal medicine granules in patients with severe coronavirus disease 2019 in Wuhan, China: a retrospective, single-center study with propensity score matching. Phytomedicine. 2021;85:153404. doi:10.1016/j.phymed.2020.153404
16. Zhou G, Tang L, Zhou X, Wang T, Kou Z, Wang Z. A review on phytochemistry and pharmacological activities of the processed lateral root of Aconitum carmichaelii Debeaux. J Ethnopharmacol. 2015;160:173–193. doi:10.1016/j.jep.2014.11.043
17. Fu YP, Zou YF, Lei FY, Wangensteen H, Inngjerdingen KT. Inngjerdingen, Aconitum carmichaelii Debeaux: a systematic review on traditional use, and the chemical structures and pharmacological properties of polysaccharides and phenolic compounds in the roots. J Ethnopharmacol. 2022;291:115148. doi:10.1016/j.jep.2022.115148
18. Zhao C, Chen W, Wang D, et al. Ginger (Zingiber officinale Roscoe) preparations for prophylaxis of postoperative nausea and vomiting: a Bayesian network meta-analysis. J Ethnopharmacol. 2023;317:116791. doi:10.1016/j.jep.2023.116791
19. Shikov AN, Narkevich IA, Flisyuk EV, Luzhanin VG, Pozharitskaya ON. Medicinal plants from the 14(th) edition of the Russian Pharmacopoeia, recent updates. J Ethnopharmacol. 2021;268:113685. doi:10.1016/j.jep.2020.113685
20. Lu Y, Wu Y, Huang M, et al. Fuzhengjiedu formula exerts protective effect against LPS-induced acute lung injury via gut-lung axis. Phytomedicine. 2024;123:155190. doi:10.1016/j.phymed.2023.155190
21. Huang YF, Li HY, Guo JX, et al. Simultaneous determination of twenty-nine active compounds in fuzhengjiedu granules by HPLC-QQQ-MS/MS. Heliyon. 2023;9(2):e13675. doi:10.1016/j.heliyon.2023.e13675
22. Zhao M, Xie X, Xu B, et al. Paeonol alleviates ulcerative colitis in mice by increasing short-chain fatty acids derived from Clostridium butyricum. Phytomedicine. 2023;120:155056. doi:10.1016/j.phymed.2023.155056
23. Zhang X, Wei X, Deng Y, et al. Mesenchymal stromal cells alleviate acute respiratory distress syndrome through the cholinergic anti-inflammatory pathway. Signal Transduct Target Ther. 2022;7(1):307. doi:10.1038/s41392-022-01124-6
24. Bhattacharya J, Matthay MA. Regulation and repair of the alveolar-capillary barrier in acute lung injury. Annu Rev Physiol. 2013;75:593–615. doi:10.1146/annurev-physiol-030212-183756
25. Vinolo MA, Rodrigues HG, Hatanaka E, Sato FT, Sampaio SC, Curi R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J Nutr Biochem. 2011;22(9):849–855. doi:10.1016/j.jnutbio.2010.07.009
26. Sina C, Gavrilova O, Förster M, et al. G protein-coupled receptor 43 is essential for neutrophil recruitment during intestinal inflammation. J Immunol. 2009;183(11):7514–7522. doi:10.4049/jimmunol.0900063
27. Tai W, Xu Y, Ding J, et al. Fibrocytes ameliorate acute lung injury by decreasing inflammatory cytokine and chemokine levels and reducing neutrophil accumulation in the lung. Cell Physiol Biochem. 2017;44(4):1526–1536. doi:10.1159/000485647
28. Shen J, Wang S, Huang Y, et al. Lactobacillus reuteri ameliorates lipopolysaccharide-induced acute lung injury by modulating the gut microbiota in mice. Nutrients. 2023;15(19):4256. doi:10.3390/nu15194256
29. Xu Y, Zhu J, Feng B, et al. Immunosuppressive effect of mesenchymal stem cells on lung and gut CD8(+) T cells in lipopolysaccharide-induced acute lung injury in mice. Cell Prolif. 2021;54(5):e13028. doi:10.1111/cpr.13028
30. Sze MA, Tsuruta M, Yang SW, et al. Changes in the bacterial microbiota in gut, blood, and lungs following acute LPS instillation into mice lungs. PLoS One. 2014;9(10):e111228. doi:10.1371/journal.pone.0111228
31. Horowitz A, Chanez-Paredes SD, Haest X, Turner JR. Paracellular permeability and tight junction regulation in gut health and disease. Nat Rev Gastroenterol Hepatol. 2023;20(7):417–432. doi:10.1038/s41575-023-00766-3
32. Zhan L, Pu J, Zheng J, et al. Tetrastigma hemsleyanum Diels et Gilg ameliorates lipopolysaccharide induced sepsis via repairing the intestinal mucosal barrier. Biomed Pharmacother. 2022;148:112741. doi:10.1016/j.biopha.2022.112741
33. Whiteside SA, McGinniss JE, Collman RG. The lung microbiome: progress and promise. J Clin Invest. 2021;131(15). doi:10.1172/JCI150473
34. Bingula R, Filaire M, Radosevic-Robin N, et al. Desired turbulence? gut-lung axis, immunity, and lung cancer. J Oncol. 2017;2017:5035371. doi:10.1155/2017/5035371
35. Ghosh S, Whitley CS, Haribabu B, Jala VR. Regulation of intestinal barrier function by microbial metabolites. Cell Mol Gastroenterol Hepatol. 2021;11(5):1463–1482. doi:10.1016/j.jcmgh.2021.02.007
36. Allaire JM, Crowley SM, Law HT, Chang SY, Ko HJ, Vallance BA. The intestinal epithelium: central coordinator of mucosal immunity. Trends Immunol. 2018;39(9):677–696. doi:10.1016/j.it.2018.04.002
37. Wu J, Zhou B, Pang X, et al. Clostridium butyricum, a butyrate-producing potential probiotic, alleviates experimental colitis through epidermal growth factor receptor activation. Food Funct. 2022;13(13):7046–7061. doi:10.1039/D2FO00478J
38. Zhao X, Yang J, Ju Z, et al. Clostridium butyricum ameliorates salmonella enteritis induced inflammation by enhancing and improving immunity of the intestinal epithelial barrier at the intestinal mucosal level. Front Microbiol. 2020;11:299. doi:10.3389/fmicb.2020.00299
39. Wang Y, Gu Y, Fang K, et al. Lactobacillus acidophilus and Clostridium butyricum ameliorate colitis in murine by strengthening the gut barrier function and decreasing inflammatory factors. Benef Microbes. 2018;9(5):775–787. doi:10.3920/BM2017.0035
40. Socorro TPD, Oka K, Boulard O, et al. The biotherapeutic Clostridium butyricum MIYAIRI 588 strain potentiates enterotropism of Rorγt(+)Treg and PD-1 blockade efficacy. Gut Microbes. 2024;16(1):2315631. doi:10.1080/19490976.2024.2315631
41. Zhu W, Wang J, Zhao N, et al. Oral administration of Clostridium butyricum rescues streptomycin-exacerbated respiratory syncytial virus-induced lung inflammation in mice. Virulence. 2021;12(1):2133–2148. doi:10.1080/21505594.2021.1962137
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.