Back to Journals » Cancer Management and Research » Volume 11
Function of microRNA-145 and mechanisms underlying its role in malignant tumor diagnosis and treatment
Authors Ye D ![]() , Shen ZS
, Shen ZS ![]() , Zhou SH
, Zhou SH ![]()
Received 22 October 2018
Accepted for publication 28 December 2018
Published 22 January 2019 Volume 2019:11 Pages 969—979
DOI https://doi.org/10.2147/CMAR.S191696
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lu-Zhe Sun
Dong Ye,1,2 Zhisen Shen,2 Shuihong Zhou1
1Department of Otorhinolaryngology – Head and Neck Surgery, The First Affiliated Hospital, College of Medicine, Zhejiang University, Hangzhou, Zhejiang, China; 2Department of Otorhinolaryngology –Head and Neck Surgery, Lihuili Hospital, Ningbo University, Ningbo, Zhejiang, China
Abstract: miRNAs are single-stranded small RNAs that do not encode proteins. They can combine complementarily with the 3’-UTRs of target gene mRNA molecules to promote targeted mRNA degradation or inhibit mRNA translation, thereby regulating gene expression at the post-transcriptional level. MiRNAs participate in regulation of cell cycling, growth, apoptosis, differentiation, and stress responses. MiRNA-145 (miR-145) is a tumor suppressor that targets various tumor-specific genes and proteins, thereby influencing related signaling pathways. MiR-145 not only regulates tumor growth, invasion, and metastasis, but is also important for tumor angiogenesis and tumor stem cell proliferation. Here, we review the roles and mechanisms of miR-145 in the diagnosis and treatment of malignant tumors. Published data confirm that miR-145 expression in various tumors is significantly lower than that in normal tissues and that overexpression of miR-145 inhibits the growth of different tumor cells, significantly reduces the ability of tumors to spread, and improves sensitivity to chemotherapeutic drugs. We conclude that miR-145 is a potential marker for use in the early diagnosis and prognostic evaluation of patients with cancer, has a role as a tumor suppressor, and is a promising cancer treatment target candidate.
Keywords: miRNA-145, tumor, proliferation, metastasis, mechanism
Introduction
The mechanisms underlying the occurrence and development of malignant tumors are of great research interest, particularly those that influence the pathogenesis, diagnosis, and treatment of tumors. Protein coding genes are the primary effectors and regulators of tumor occurrence and development; however, the emergence of epigenetics has revealed numerous regulatory mechanisms and effects on protein function that occur after gene transcription. Epigenetic control pathways involving non-coding RNA molecules are an area of increasing research interest, with specific relevance to miRNAs. MiRNAs are widespread in eukaryotic organisms, where they are closely associated with the expression and regulation of tumor-associated genes and influence the occurrence and development of tumors.1
MiRNAs are small, single-stranded RNA molecules that do not encode proteins.2 Non-coding single-stranded RNA precursors, which are hairpin structures of approximately 70–90 nucleotides, are processed by the Dicer enzyme to produce miRNAs of about 22 residues. These miRNAs can be complementary with the 3’-UTRs of target mRNA molecules and they may promote degradation of target mRNAs or inhibit their translation, thereby mediating post-transcriptional gene regulation. MiRNAs can influence cell cycling, growth, apoptosis, differentiation, and stress responses, and are important as gene regulators, particularly in cancer.3,4 Approximately 5,300 miRNA target genes have been reported to date, and 52.5% of known miRNAs are located in genome regions related to cancer or fragile sites;5,6 a fragile site is a site-specific gap or fracture region that occurs in a metaphase chromosome when DNA synthesis is partially suppressed, which is susceptible to rearrangements in response to external factors, and predisposes cells to cancer. Fragile sites are usually stable in somatic cells; however, they are frequently absent or rearranged in tumor cells. By regulating target gene signaling pathways influencing tumor growth, invasion, metastasis, and angiogenesis, miRNAs can function as oncogenes or tumor suppressor genes.
miRNA-145 (miR-145) is a tumor suppressor expressed in various tumors, including prostate, bladder, colon, ovarian, and esophageal cancers, at lower levels than those in normal tissues.7–11 As a tumor suppressor gene, miR-145 inhibits tumor cell proliferation, invasion, and metastasis,12,13 increases tumor cell sensitivity to chemotherapeutic drugs,14 and regulates tumor occurrence and development. Detailed studies suggest that miR-145 is potentially an ideal marker for diagnostic and prognostic evaluation of tumors and may represent a new cancer treatment target. This article reviews the reported roles of miR-145 in the diagnosis and treatment of malignant tumors and the mechanisms underlying these.
Structure and biological characteristics of miR-145
MiR-145 maps to chromosome 5q32-33 and is 4.08 kb in length. The sequence is highly conserved, relative to other non-coding small molecule RNAs. Depending on the direction of transcription, the miR-145 locus can generate two transcripts, miR-145-3p (the passenger strand) and miR-145-5p (the guide strand). The miR-145-3p transcript is processed to produce miRNAs of ~22 nucleotides (miRBase database number: MIMAT0004601), whereas miR-145-5p generates 23 nucleotide fragments (miRBase database number: MIMAT0000437). MiR-145 maps in a cluster close to miR-143, and the two transcripts have similar roles and are thought to be co-transcribed.15 The miR-145 chromosome locus is close to a very important tumor fragile site at 5q31, and implicated in tumor occurrence and development, as it is frequently deleted during the chromosome rearrangements that accompany malignant transformation, leading to its low expression levels in tumor tissues.6
MiR-145 was first detected in mouse heart,16 and subsequently, in human colorectal cancer and other tissues.17 Mesodermal tissues, such as the uterus, ovary, testis, prostate, and heart, are relatively rich in miR-145;18 however, the transcript is expressed at low levels in various tumor tissues, including colon, breast, prostate, lung, liver, bladder, and ovarian cancers, along with pituitary adenoma and B-cell lymphoma, among others.13 There is evidence that miR-145 protects against tumorigenesis and can regulate tumor growth, invasion, metastasis, and angiogenesis via complete or incomplete complementation with the 3′ non-coding region of target mRNA molecules, resulting in degradation of the target mRNAs and effects on their translation levels.
MiR-145 and tumor cell growth
Both the sequence and tissue-specific transcription of miR-145 are highly conserved, and this miRNA regulates oncogene expression at the transcriptional and post-transcriptional levels, thereby reducing tumor growth by blocking the cell cycle and promoting tumor cell apoptosis.
MiR-145 exerts its effects on tumor cells through regulation of downstream target genes, including: octamer-binding transcription factor 4 (OCT4), SRY-box 9 (SOX9), C-MYC, YES, EGFR, insulin-like growth factor, mucin 1 (MUC1), adenovirus EIB19000 interaction protein 3, signal transducer and activator of transcription 1 (STAT1), tumor necrosis factor super family member 10 (TNFSF10), and others.19–24 MiR-145 also regulates the differentiation of human embryonic stem cells and inhibits proliferation of smooth muscle cells by targeting OCT4, SOX2, and Krüppel-like factor 4 (KLF4).25
Cho et al showed that miR-145 negatively regulates the expression of the folate receptor and NUDT1 at the mRNA and protein levels, respectively,26 and cell proliferation is inhibited in miR-145-transfected lung adenocarcinoma cells. Zhong et al27 studied the expression of three different miRNAs in patients with non-small-cell lung cancer (NSCLC) and conducted quantitative analysis of three kinds of miRNA molecules; miR-145 was identified as mainly involved in regulating tumor cell behavior through the RAS/ERK and PI3K/AKT signaling pathways. Further, miR-145 and miR-143 attenuate tumor cell growth by inhibiting the ERK5/c-MYC and p68/p72/β-catenin signaling pathways in Apc(Min) mice with tumors of the gut.28 Moreover, Xu et al showed that miR-145 inhibits tumor cell growth and angiogenesis in patients with colon cancer by targeting mammalian target of rapamycin (mTOR)/p70S6K1 signaling,29 while Wang et al reported that overexpression of sentrin-specific protease 1 (SENP1), which promotes the proliferation, infiltration, and metastasis of tumor cells, is an important feature of bladder cancer30 and, in a nude mouse model of bladder cancer, miR-145 inhibited tumor formation by interfering with SENP1.
MiR-145 in tumor invasion and metastasis
Local invasion and distant metastasis are important characteristics of malignant tumors and the main cause of death from cancer, and miR-145 can regulate these processes.31
Sachdeva et al showed that miR-145 exerts its function in a cell-specific manner.32 Although miR-145 inhibits cell growth in MCF-7 and HCT-116 cells, it has no significant effect on cell growth in metastatic breast cancer cell lines. However, miR-145 significantly suppresses cell invasion in these cells. Using luciferase reporter assays, they showed that miR-145 targets the tumor metastasis gene, MUC1, and inhibits tumor metastasis by silencing its expression. Gotte et al reported that miR-145 regulates the movement of breast cancer cells by targeting expression of the cell adhesion molecule, JAM-A, and the actin-crosslinking protein, fascin, which influence cell cytoskeletal rearrangement and nuclear rotation.33
MiR-145 also negatively regulates N-cadherin levels by binding directly to its 3’-UTR and overexpression of miR-145 silences N-cadherin expression, inhibiting invasion and metastasis of a lung adenocarcinoma cell line.34
MiR-145 and tumor angiogenesis
Tumor angiogenesis has a key role in tumor growth and metastasis, along with patient prognosis. Tumor angiogenesis and anti-tumor angiogenesis are studied with the aim of cancer prevention and treatment, and many investigations have shown that miR-145 is important in this process.35
The miR-145 gene locus is close to that of miR-143 and, importantly, their transcription is coordinated.15 Both transcripts directly participate in the regulation of vascular smooth muscle cell differentiation by maintaining the cell in a mature and shrinkable state.36 Cordes et al reported that miR-145 and miR-143 control the expression of KLF4, myocardin, and Elk-1, thereby promoting the differentiation of vascular smooth muscle cells and inhibiting their proliferation.15
MiR-145 and tumor stem cells
MiR-145 not only directly modulates the biological characteristics of tumor cells, but can also control tumor stem cells, a subset of tumor cells with stem cell characteristics, that are the source of tumor growth, metastasis, and recurrence. Targeting of tumor stem cells has become a new approach for cancer treatment. Xu et al reported that miR-145 regulates the expression of cell reprogramming genes, including OCT4, SOX2, and KLF4, which maintain stem cell function and have characteristics of oncogenes.25 MiR-145 also inhibits human embryonic stem cell regeneration, promotes their differentiation, and is important for tumorigenesis and tumor development. Simultaneously, OCT4 inhibits miR-145 expression, forming an expression-regulation negative feedback loop. Research into this phenomenon will provide new information regarding the role of miR-145 in tumor stem cells, and it is of great significance for the treatment of tumors. A study of bone marrow cells showed that stable knockdown of miR-145 following its overexpression in CD34+ cells can lead to myelodysplastic (5q−) syndrome.37
Relationship between miR-145 and tumors
MiR-145 has been studied extensively in the context of tumor cell growth inhibition, and has become increasingly important in tumor diagnosis, prognostic assessment, and targeted therapy.38 MiR-145 may function as a tumor suppressor gene that is expressed in various tumor tissues, including ovarian, cervical, breast, and colorectal cancers, at significantly lower levels than those in normal tissue.39–42 Consequently, miR-145 is expected to be useful as an early tumor diagnostic and prognostic marker. In addition, overexpression of miR-145 inhibits the proliferation and metastasis of tumor cells, and it functions as a tumor suppressor gene and improves the sensitivity to chemotherapeutic drugs; hence, it is expected to serve as a novel target for cancer treatment.
Progress in the diagnosis of malignant tumors by analysis of miR-145
Studies on associations of miR-145 with specific tumors are ongoing. As a diagnostic tool with high accuracy and efficiency, miR-145 has potential for application in tumor diagnosis; however, this approach remains at the research stage.43
MiR-145 may be an ideal marker for early diagnosis. Boufraqech et al used reverse-transcription PCR (RT-PCR) to quantitatively detect miR-145 expression in 75 samples from cases with thyroid cancer, demonstrating that miR-145 expression levels were significantly higher in benign tissue than in malignancies.44 Blood miR-145 levels are markedly increased in patients with thyroid cancer and show a specific gradient of venous concentration, suggesting that miR-145 may be useful as an accessory biomarker for thyroid carcinoma diagnosis. Moreover, Peng et al demonstrated that miR-145 and miR-378* are potential early diagnostic markers of colorectal cancer.45
The clinical signs of malignant pleural mesothelioma (MPM) are difficult to distinguish from those of reactive mesothelial proliferation. Andersen et al used quantitative RT-PCR to analyze 742 miRNA molecules in tumor tissues and corresponding non-neoplastic pleural mesothelial tissues, and found that levels of miR-145, miR-126, miR-143, and miR-652 were significantly reduced in tumor tissues and that these four miRNAs can be used as markers of MPM.46
Gits et al showed that miR-145 levels in liposarcomas were significantly lower than those in normal adipose tissue, with marked differences between different tumor subtypes, suggesting that miR-145 levels can be used for objective auxiliary diagnosis of liposarcoma.47
Ethnic variation in the usefulness of miR-145 as a marker for diagnosing tumors has been described, with the marker exhibiting higher sensitivity and specificity in Caucasian than East Asian patients in a meta-analysis, which demonstrates that miR-145 has high accuracy in distinguishing between patients with and without lymph node metastases of various cancers.43
MiR-145 levels are closely associated with prognosis of patients with cancer. Campayo et al determined miR-145 and miR-367 levels in tumor tissues from 70 patients with NSCLC, and found that the average time to recurrence for patients with low miR-145 levels was 18.4 months, whereas that for patients with high miR-145 levels was 28.2 months.48 In contrast, the recurrence period for patients with high levels of miR-367 was shorter than that of patients with low levels. Hence, miR-145 and miR-367 levels can be used as predictors of postoperative recurrence in patients with NSCLC.
Wu et al showed that miR-145 targets matrix metalloproteinase-11 (MMP-11) to inhibit the proliferation and metastasis of renal cell carcinoma (RCC), suggesting that miR-145 could be used as an early predictor of RCC metastasis.49
Kim et al found that low levels of miR-145 are significantly correlated with recurrence and survival rates in patients with ovarian cancer (HR=8.62, P=0.039), and low miR-145 expression levels were detected in ovarian cancer patients with poor prognosis.50
Levels of miR-145 in cervical cancer samples are significantly lower than those in adjacent non-cancer tissues, and low expression of miR-145 is closely associated with advanced International Federation of Gynecology and Obstetrics stage, lymph node metastasis, vascular infiltration, and shorter survival; hence, MiR-145 is a potential prognostic marker for cervical cancer.51 Low miR-145 expression is also associated with the diameter and location of rectal cancer and esophageal carcinoma stage.52,53
Xia et al54 reported that differences in expression levels of miR-130b, miR-145, and miR-223 in patients with adult T cell lymphoma (ATL) are associated with differences in patient survival rates. Multivariate analysis indicated that low miR-145 levels are associated with deterioration of patients with ATL and closely related to poor prognosis in these individuals. Tang et al also showed that low miR-145 levels are closely related to invasion and metastasis in osteosarcoma and can be used as an independent prognostic marker.55
Naito et al reported high levels of miR-145 expression in patients with advanced stage scirrhous-type gastric cancer (P=0.0156), and significant correlation of high miR-145 expression with adverse prognosis (P=0.0438).56
Overall, these studies demonstrate that levels of miR-145 expression in patients with cancer are strongly correlated with prognosis. In the majority of tumor types, low miR-145 levels are associated with worse prognosis.
Progress in miR-145 research aimed at treatment of malignant tumors
Various studies have reported targeting miR-145 and related genes and proteins for drug delivery, and the consequent inhibition of tumor cell proliferation and metastasis, pointing to new possibilities for tumor treatment.
Chondrosarcoma is the most common type of adult bone tumor. The effects of traditional chemotherapy and radiotherapy on this type of tumor are limited, hence the main treatment option is surgery. Mak et al reported that ETS variant 5 (ETV5) activates MMP-2 expression in chondrosarcoma, which enhances bone matrix resorption.57 Levels of SOX9, ETV5, and MMP-2 in cells were significantly reduced by stable transfection of miR-145 into chondrosarcoma cells by lentivirus, demonstrating the potential of miR-145 for use in the clinical treatment of chondrosarcoma.
He et al showed that overexpression of prostate-specific transcript 1 (PCGEM1) promotes proliferation and metastasis of prostate cancer cells, along with resistance of tumor cells to chemotherapeutic agents. MiR-145 and PCGEM1 exhibit double-negative feedback regulation, where overexpression of miR-145 inhibits that of PCGEM1, indicating a potential new approach for treating prostate cancer by targeting miR-145 or PCGEM1.58
MiR-145 regulates expression of key factors (HIF2α, VEGF, MMP-9, and CCND1) in RCC progression, and hence, enhanced miR-145 expression inhibits RCC progression.59
Li et al reported that miR-145 expression is significantly reduced in rectal cancer tissues and can effectively inhibit the proliferation and metastasis of rectal cancer cells.60 This finding provides a theoretical basis for treating rectal cancer by targeting this tumor suppressor. Ibrahim et al evaluated substitution therapy with miR-145 and miR-133a mediated by polyethylene imide in a colorectal cancer model, which had the therapeutic effect of promoting tumor cell apoptosis and inhibiting proliferation.61
Huang et al reported that miR-145 reduces the expression of various oncogenes and tumor biomarkers in pancreatic cancer cell lines, including SET nuclear proto-oncogene, replication protein A1, mini chromosome maintenance complex component 2, ATP-binding cassette, subfamily C1, non-erythrocyte-type β-membrane contractile protein (SPTBN1), and serine palmitoyltransferase long chain base subunit 1.62 As these proteins function in tumor growth inhibition, miR-145 may be an ideal target for tumor therapy.
Restoration of miR-145 expression can effectively promote apoptosis of breast cancer cells expressing wild-type TP53 or estrogen receptor, suggesting that miR-145 therapy may be effective in patients with breast tumors expressing wild-type TP53- or estrogen receptor.63
MiR-145 levels are lower in NSCLC than in normal lung cells, and AP27i145 replication is negatively correlated with miR-145 expression. Oncolytic herpes simplex virus (HSV)-1 selectively inhibits cell proliferation and colony formation in samples from patients with NSCLC. The combination of AP27i145 and radiotherapy is more likely to kill cancer cells than either treatment alone; hence, oncolytic HSV-1, which is regulated by miR-145, is an effective target for treatment of NSCLC.64
When miR-145 and related genes are targeted for treatment, they can reduce or even counteract chemotherapeutic resistance to some drugs, making them ideal targets for cancer drug therapy. Zhong et al27showed that miR-145 has an important role in reducing the apoptosis tolerance of tumor cells and that its expression enhances sensitivity to chemotherapeutic drugs, such as gefitinib, in patients with NSCLC.
Cioce et al20 reported that cell invasiveness and resistance to the chemotherapeutic drug, pemetrexed, decreased in an MPM cell line overexpressing miR-145. Elevated OCT4 levels are considered one of the most important indices of cancer cell chemotherapy resistance; however, increased miR-145 levels can diminish expression of OCT4 and its target gene ZEB1, thus offsetting the rise in OCT4 levels caused by pemetrexed treatment.
Pancreatic cancer is a major cause of cancer deaths worldwide. Despite recent progress in research and treatment, difficulty in early disease detection and lack of effective treatments mean that the prognosis for patients with this type of cancer remains poor. Gemcitabine is a first-line drug for pancreatic cancer; however, chemotherapy resistance is a serious problem. Lin et al showed that p70S6K1 has an important role in drug resistance.14 Acting as a tumor suppressor, miR-145 directly inhibits p70S6K1 expression in pancreatic cancer, constituting an effective treatment regimen. Khan et al found that miR-145 inhibits the proliferation and metastasis of pancreatic cancer cells and improves sensitivity to gemcitabine. MiR-145 may target the MUC13 3’-UTR, thereby inhibiting intracellular MUC13 and downstream ERBB2 expression.65
Zhu et al reported that miR-145 is imported into ovarian cancer SKOV3/PTX cells, decreasing intracellular levels of cyclin-dependent kinase 6 and SP1;66 levels of ATP binding cassette subfamily B1 and retinoblastoma 1 also decrease in response to miR-145. These changes could induce aggregation of antitumor drugs in cells, and a G1 cell cycle blockade, consequently improving the sensitivity of tumor cells to taxol in vitro and in vivo. In addition, reactivation of miR-145 expression by demethylation mediated by the DNA methylase inhibitor, 5-Aza-2′-deoxycytidine, may also increase susceptibility of tumor cells to taxol.
MiR-145 regulatory factors
Environmental factors influencing miR-145 expression
Various factors affect the expression of miR-145 and regulate the proliferation and metastasis of tumor cells and their sensitivity to chemotherapeutic drugs. MiR-145 expression levels are affected by carcinogens in a rat model, with expression in lung tissues decreasing significantly after 28 days of exposure to environmental cigarette smoke. MiR-145 is also regulated in response to stress, as well as cell apoptosis, proliferation, and angiogenesis.67 Hepatitis B virus (HBV) infection promotes cell proliferation, reduces miR-145 expression, and participates in regulating cell transformation, leading to HBV-associated hepatocellular carcinoma (HCC).68
Factors regulating transcription and processing of miR-145
The tumor protein, P53, and forkhead box O1/3 (FOXO1/3) promote transcription of precursor miR-145 (pre-miR-145), whereas Ras-responsive element-binding protein 1 (RREB1) and CCAAT/enhancer-binding protein-β inhibit its transcription. During miR-145 maturation, P53 and breast cancer 1 proteins have catalytic roles, whereas the human RNA methyltransferase, BCDIN3, is inhibitory .69 P53 primarily functions in Drosha–p68 complex formation to promote pri-miRNA shearing into pre-miRNAs; however, mutated P53 lacks this function, leading to inhibition of miR-145 expression.
Adjustment of colon cancer-associated transcript-2 (CCAT2) levels regulate miR-145 expression in the HCT-116 and HT-29 rectal cancer cell lines, confirming that CCAT2 is a negative regulator of miR-145. CCAT2 is enriched in the nuclei of HCT-116 cells and closely associated with pre-miR-145 expression. CCAT2 selectively inhibits miR-145 maturation in vitro by diminishing pre-miR-145 export to the cytoplasm by the Dicer enzyme, preventing pre-miR-145 shearing .70
In lung cancer cells, EGFR downregulates miR-145 expression via the ERK1/2 pathway.71 MiR-145 expression regulation can be achieved through hypermethylation of the miR-145 promoter region in esophageal squamous cell carcinoma (ESCC).72
Mechanisms underlying miR-145 function
Tumor development is a complex process involving multiple factors and stages, and the accumulation of multiple gene mutations. Consequently, the causes and mechanisms underlying the infinite proliferation of tumor cells are highly complex and include inactivation of tumor suppressor genes, overexpression of oncogenes, uncontrolled cell cycling, and tolerance to apoptosis. MiR-145 inhibits tumor growth by reducing oncogene expression, blocking of the cell cycle, and promoting tumor cell apoptosis. One important reason for the rapid proliferation of tumor cells is the activation of proto-oncogenes; miR-145 targets and inhibits multiple oncogenes or proteins, including c-MYC, FLI1, YES, and IRS-1, thereby exerting a tumor growth inhibitory effect.
MiRNAs have multiple downstream targets, and different miRNAs may act on the same target. Lee et al reported that high miR-145 expression diminishes levels of tumor metastasis-related gene expression, including ADAM22, SPOCK3, PLAUR, SLC7A5, and FASCN1.73
MiR-145 targeted oncogenes
Sachdeva et al showed that P53 inhibits c-MYC expression via miR-145 and proposed the theory that miR-145 blocks the cell cycle.74 According to this theory, P53 regulation of P21 is an important pathway that blocks the cell cycle; P21 expression is inhibited in tumor cells due to high expression of c-MYC, thus blocking the P53/P21 signaling pathway. When miR-145 expression is restored, c-MYC levels decrease and the inhibition of P21 is relieved, thereby restoring P53/P21 signaling.
Kent et al reported that miR-143 and miR-145 are involved in positive feedback loops in the RAS cancer gene signaling pathway.75 MiR-143 inhibits expression of the RAS subtype, KRAS, while miR-145 represses RREB1 expression. In contrast, RREB1 inhibits the miR-143/145 promoter and reduces miR-143/145 expression, and low miR-143/145 expression levels are important for KRAS-meditated cell transformation.
MiR-145 has a tumor-inhibiting effect; however, the mechanisms underlying its involvement in the proliferation, metastasis, and invasion of tumor cells are not entirely clear. Patients with bladder cancer express low levels of miR-145, which inhibits tumor cell proliferation by targeting FSCN1.8 In prostate cancer, miR-145 inhibits the proliferation, invasion, and metastasis of tumor cells via the same target.76
MiR-145 targeted apoptosis-related factors
Methylation of the miR-145 promoter in RCC results in low levels of this miRNA in this type of cancer. Expression of high miR-145 levels in RCC cells leads to inhibition of tumor cell proliferation and markedly increases cisplatin-induced apoptosis, possibly by inhibiting the expression of ADAM17, c-MYC, or other downstream genes.77,78
MiR-145 expression levels directly affect the tolerance of tumor cells to apoptosis-stimulating factors. Ostenfeld et al reported that overexpression of miR-145 in urothelial carcinoma cells strongly induced apoptosis, and that it simultaneously activated caspase-dependent and non-caspase-dependent apoptotic pathways. It has been speculated that the low levels of miR-145 expression in urothelial carcinoma may be one reason for tumor-induced apoptosis escape.79
MiR-145 also inhibits the proliferation of tumor cells in NSCLC by targeting c-MYC.80
Wang et al showed that miR-145 levels are significantly lower in MCF-7 breast cancer cells compared with normal mammary epithelial (MCF10A) cells.81 RTKN expression is decreased in MCF-7 cells, indicating that miR-145 inhibits the proliferation of breast cancer cells by targeting RTKN.
DNA fragmentation is an important feature of apoptosis, which primarily occurs as a result of DFF40 degradation of chromosomal DNA. Under normal circumstances, DFF40 and DFF45 are bound to one another in the cell in an inactive state. When apoptosis occurs, a series of enzymes promote a cascade, comprising the activation of caspase-3, cleavage of DFF45 by activated caspase-3, and release of DFF40, leading to DNA fragmentation. Zhang et al reported that miR-145 directly inhibits DFF45 expression, thereby causing DNA fragmentation and inducing apoptosis in colon cancer.82
Other tumor-related factors targeted by miR-145
MiRNAs can also contribute to disorders involving the onset and development of cancers. MiR-145 expression is significantly decreased in HCC cell lines and negatively correlated with high levels of insulin receptor substrate 1 (IRS1), a key factor in the tumorigenic insulin-like growth factor pathway. IRS1 is a direct target of miR-145, and increased miR-145 expression inhibits tumor cell proliferation by lowering IRS1 levels and inhibiting the downstream AKT/FOXO1 signaling pathway. The lower miR-145 expression in HCC has been suggested as a potential molecular mechanism underlying carcinogenesis.83
MiR-145 inhibits the invasion of U87 glioma cells by inhibiting the RhoA/ROCK1 pathway, where ROCK1, a newly discovered miR-145 target, promotes invasion of glioma cells.84
Tumors in which miR-145 is overexpressed
MiR-145 is also overexpressed in a few tumor types, including esophageal adenocarcinoma (EAC) and glioma, where it enhances cell proliferation, invasion, and metastasis. In one study, no significant difference was observed in cell proliferation or 5-fluorouracil resistance between an miR-145 EAC cell line treatment and control groups, and miR-145 enhanced the resistance of SK-GT-4 cells to cisplatin. There were clear differences in cell invasion, adhesion, and anti-apoptosis effects in the OE33, FLO-1, and SK-GT-4 cell lines, and invasion in the miR-145 treatment group was significantly greater than that in the control group. MiR-145 overexpression in ESCC prevents tumor cell proliferation and invasion, whereas overexpression of miR-145 in EAC enhances invasion and anti-apoptosis effects.85
MiR-145 expression is increased, and that of srGAP1 decreased, in invasive glioblastoma (IM3) cells, whereas downstream G-proteins remain active, thus promoting an invasive phenotype.86
Summary
The known common tumor-related factors that miR-145 can target are as follows: PI3K/AKT,27 ROCK1,87 caspase,79 c-MYC,74RTKN,81RREB1,75ER-alpha,63 DFF45,82MUC1,32JAM-A,33fascin,33ELK1,15OCT4,25 SOX2,25KLF4,25YES,21STAT1,21IRS-1,22FLI1,88 and TNFSF10.11 MiR-145 target molecules differ in various types of tumor and tissue. Target molecules and effects of their targeting by miR-145 in common tumors are presented in Table 1.
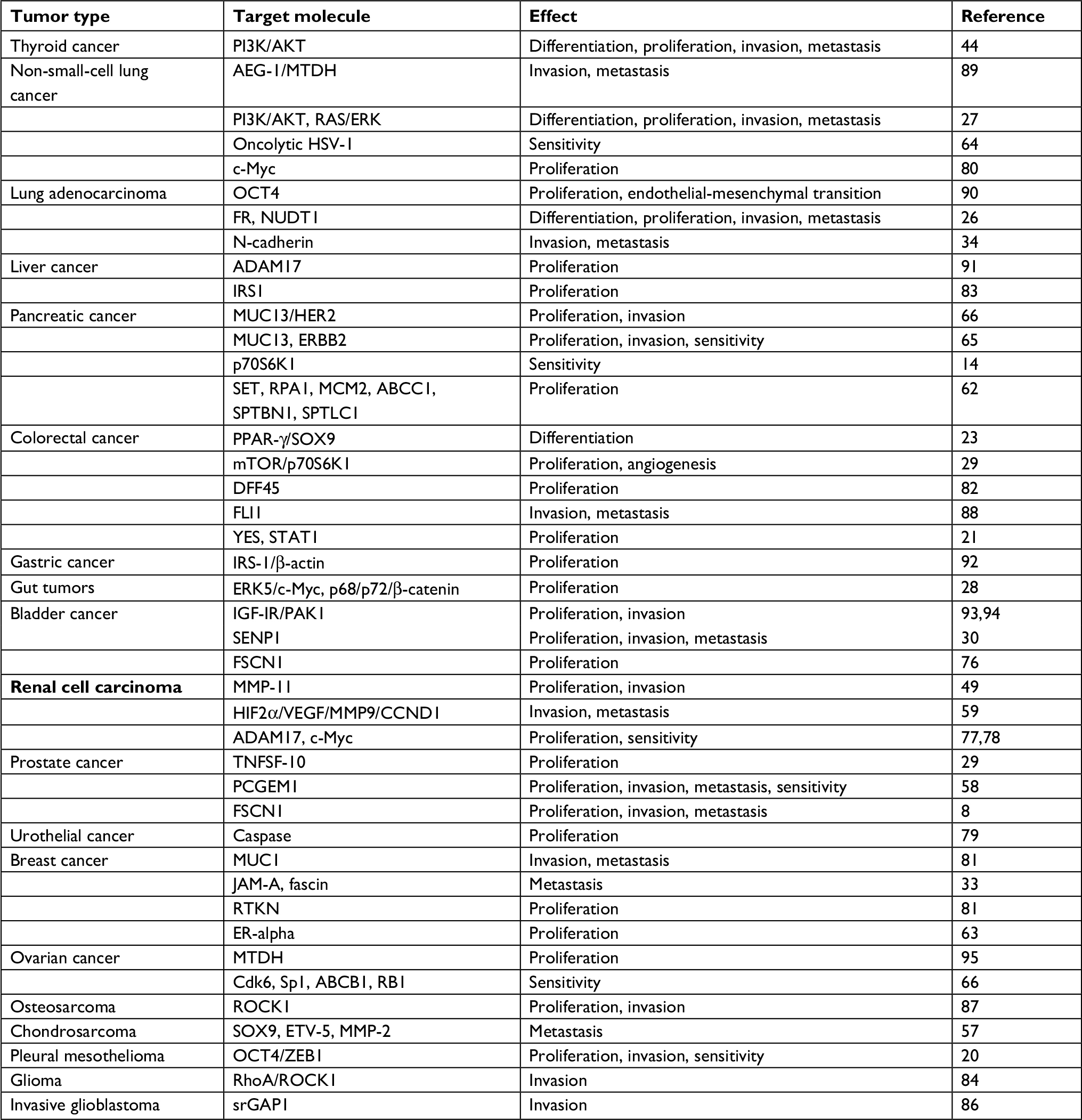 | Table 1 MiR-145 target molecules and consequences of their targeting in common tumors |
MiR-145 not only regulates tumor growth, invasion, and metastasis, but also plays an important role in tumor angiogenesis and tumor stem cells; hence, with further research, miR-145 has potential for application as a treatment tool in clinical oncology.
Conclusion
In summary, miR-145 has important functions in the proliferation, apoptosis, invasion, and metastasis of tumor cells. This is evidenced by findings showing that miR-145 is downregulated in numerous tumors compared with normal tissues, and that overexpression of miR-145 inhibits the growth of multiple tumor cells and functions as a tumor suppressor gene. Consequently, miR-145 is expected to be a marker for the early diagnosis and prognostic assessment of tumors. Furthermore, transfection with miR-145 can inhibit the proliferation and metastasis of tumor cells and improve sensitivity to chemotherapeutic drugs, potentially providing new tumor treatment approaches. The majority of studies of miR-145 in malignant tumors have been limited to investigation of its expression spectrum and its specific targets vary among tumors and have differing effects. The detailed mechanisms of miR-145 target gene activity, and the interactions among target genes in the context of tumorigenesis and development, have yet to be fully elucidated, and further study is required to better understand how miR-145 participates in tumor occurrence and development. Nevertheless, continuous improvements in understanding and in-depth research into miR-145 and its potential as a diagnostic marker bring hope for novel interventions to overcome chemotherapy resistance and develop targeted biological therapies for the diagnosis and treatment of cancers.
Acknowledgments
This work was supported by grants from the Scientific Innovation Team Project of Ningbo (Grant No. 2012B82019), the Ningbo Social Developmental Key Research Project (Grant No. 2012C5015), the Natural Science Foundation of Ningbo (Grant Nos. 2013A610217; 2017A610236; 2018A610361), the Leading and Top Talents Training Projects of Ningbo (NBLJ201801032), Ningbo Health Branding Subject Fund (PPXK2018-02), and the Medical and Health Training Project of Zhejiang Province (Grant Nos. 2015RCB025; 2018RC063).
Disclosure
The authors report no conflicts of interest in this work.
References
Babashah S, Soleimani M. The oncogenic and tumour suppressive roles of microRNAs in cancer and apoptosis. Eur J Cancer. 2011;47(8):1127–1137. | ||
Small EM, Frost RJ, Olson EN. MicroRNAs add a new dimension to cardiovascular disease. Circulation. 2010;121(8):1022–1032. | ||
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. | ||
Meltzer PS. Cancer genomics: small RNAs with big impacts. Nature. 2005;435(7043):745–746. | ||
Bentwich I. Identifying human microRNAs. Curr Top Microbiol Immunol. 2008;320:257–269. | ||
Calin GA, Sevignani C, Dumitru CD, et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Nat Acad Sci USA. 2004;101(9):2999–3004. | ||
Arndt GM, Dossey L, Cullen LM, et al. Characterization of global microRNA expression reveals oncogenic potential of miR-145 in metastatic colorectal cancer. BMC Cancer. 2009;9(1):374. | ||
Chiyomaru T, Enokida H, Tatarano S, et al. MiR-145 and miR-133a function as tumour suppressors and directly regulate FSCN1 expression in bladder cancer. Br J Cancer. 2010;102(5):883–891. | ||
Iorio MV, Visone R, Di Leva G, et al. MicroRNA signatures in human ovarian cancer. Cancer Res. 2007;67(18):8699–8707. | ||
Kano M, Seki N, Kikkawa N, et al. MiR-145, miR-133a and miR-133b: tumor-suppressive miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J Cancer. 2010;127(12):2804–2814. | ||
Zaman MS, Chen Y, Deng G, et al. The functional significance of microRNA-145 in prostate cancer. Br J Cancer. 2010;103(2):256–264. | ||
Lu Y, Chopp M, Zheng X, Katakowski M, Buller B, Jiang F. MiR-145 reduces ADAM17 expression and inhibits in vitro migration and invasion of glioma cells. Oncol Rep. 2013;29(1):67–72. | ||
Sachdeva M, Mo YY. miR-145-mediated suppression of cell growth, invasion and metastasis. Am J Trans Res. 2010;2:170–180. | ||
Lin Y, Ge X, Wen Y, et al. MiRNA-145 increases therapeutic sensibility to gemcitabine treatment of pancreatic adenocarcinoma cells. Oncotarget. 2016;7(43):70857–70868. | ||
Cordes KR, Sheehy NT, White MP, et al. MiR-145 and miR-143 regulate smooth muscle cell fate and plasticity. Nature. 2009;11:705–710. | ||
Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T. Identification of novel genes coding for small expressed RNAs. Science. 2001;294(5543):853–858. | ||
Michael MZ, O’ Connor SM, van Holst Pellekaan NG, Young GP, James RJ. Reduced accumulation of specific microRNAs in colorectal neoplasia. Mol Cancer Res. 2003;1(12):882–891. | ||
Landgraf P, Rusu M, Sheridan R, et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell. 2007;129(7):1401–1414. | ||
Chen X, Gong J, Zeng H, et al. MicroRNA145 targets BNIP3 and suppresses prostate cancer progression. Cancer Res. 2010;70(7):2728–2738. | ||
Cioce M, Ganci F, Canu V, et al. Protumorigenic effects of miR-145 loss in malignant pleural mesothelioma. Oncogene. 2014;33(46):5319–5331. | ||
Gregersen LH, Jacobsen AB, Frankel LB, Wen J, Krogh A, Lund AH. MicroRNA-145 targets yes and STAT1 in colon cancer cells. PLoS One. 2010;5(1):e8836. | ||
La Rocca G, Badin M, Shi B, et al. Mechanism of growth inhibition by MicroRNA 145: the role of the IGF-I receptor signaling pathway. J Cell Physiol. 2009;220(2):485–491. | ||
Panza A, Votino C, Gentile A, et al. Peroxisome proliferator-activated receptor γ-mediated induction of microRNA-145 opposes tumor phenotype in colorectal cancer. Biochim Biophys Acta. 2014;1843(6):1225–1236. | ||
Shi B, Sepp-Lorenzino L, Prisco M, Linsley P, Deangelis T, Baserga R. Micro RNA 145 targets the insulin receptor substrate-1 and inhibits the growth of colon cancer cells. J Biol Chem. 2007;282(45):32582–32590. | ||
Xu N, Papagiannakopoulos T, Pan G, Thomson JA, Kosik KS. MicroRNA-145 regulates Oct4, Sox2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell. 2009;137(4):647–658. | ||
Cho WC, Chow AS, Au JS. MiR-145 inhibits cell proliferation of human lung adenocarcinoma by targeting EGFR and NUDT1. RNA Biol. 2011;8(1):125–131. | ||
Zhong M, Ma X, Sun C, Chen L. MicroRNAs reduce tumor growth and contribute to enhance cytotoxicity induced by gefitinib in non-small cell lung cancer. Chem Biol Interact. 2010;184(3):431–438. | ||
Takaoka Y, Shimizu Y, Hasegawa H, et al. Forced expression of miR-143 represses ERK5/c-Myc and p68/p72 signaling in concert with miR-145 in gut tumors of Apc(Min) mice. PLoS One. 2012;7(8):e42137. | ||
Xu Q, Liu LZ, Qian X, et al. MiR-145 directly targets p70S6K1 in cancer cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res. 2012;40(2):761–774. | ||
Wang C, Tao W, Ni S, et al. Tumor-suppressive microRNA-145 induces growth arrest by targeting SENP1 in human prostate cancer cells. Cancer Sci. 2015;106(4):375–382. | ||
Nicoloso MS, Spizzo R, Shimizu M, Rossi S, Calin GA. MicroRNAs-the micro steering wheel of tumour metastases. Nat Rev Cancer. 2009;9(4):293–302. | ||
Sachdeva M, Mo YY. MicroRNA-145 suppresses cell invasion and metastasis by directly targeting mucin 1. Cancer Res. 2010;70(1):378–387. | ||
Götte M, Mohr C, Koo CY, et al. miR-145-dependent targeting of junctional adhesion molecule A and modulation of fascin expression are associated with reduced breast cancer cell motility and invasiveness. Oncogene. 2010;29(50):6569–6580. | ||
Mo D, Yang D, Xiao X, Sun R, Huang L, Xu J. MiRNA-145 suppresses lung adenocarcinoma cell invasion and migration by targeting N-cadherin. Biotechnol Lett. 2017;39(5):701–710. | ||
Daubman S. MicroRNAs in angiogenesis and vascular smooth muscle cell function. Circ Res. 2010;106(3):423–425. | ||
Boettger T, Beetz N, Kostin S, et al. Acquisition of the contractile phenotype by murine arterial smooth muscle cells depends on the Mir143/145 gene cluster. J Clin Invest. 2009;119(9):2634–2647. | ||
Starczynowski DT, Kuchenbauer F, Argiropoulos B, et al. Identification of miR-145 and miR-146a as mediators of the 5q- syndrome phenotype. Nat Med. 2010;16(1):49–58. | ||
Bartels CL, Tsongalis GJ. MicroRNAs: novel biomarkers for human cancer. Clin Chem. 2009;55(4):623–631. | ||
Dip N, Reis ST, Srougi M, Dall’Oglio MF, Leite KR. Expression profile of microRNA-145 in urothelial bladder cancer. Int Braz J Urol. 2013;39(1):95–102. | ||
du L, Pertsemlidis A. microRNAs and lung cancer: tumors and 22-mers. Cancer Metastasis Rev. 2010;29(1):109–122. | ||
Garzon R, Calin GA, Croce CM. MicroRNAs in cancer. Annu Rev Med. 2009;60(1):167–179. | ||
Huang L, Lin JX, Yu YH, Zhang MY, Wang HY, Zheng M. Downregulation of six microRNAs is associated with advanced stage, lymph node metastasis and poor prognosis in small cell carcinoma of the cervix. PLoS One. 2012;7(3):e33762. | ||
Hou Y, Wang X, Chen Y, Mu S. MicroRNA-145 as ideal biomarker for the diagnosis of various carcinomas. Tumor Biol. 2015;36(4):2641–2649. | ||
Boufraqech M, Zhang L, Jain M, et al. miR-145 suppresses thyroid cancer growth and metastasis and targets Akt3. Endocr Relat Cancer. 2014;21(4):517–531. | ||
Peng J, Xie Z, Cheng L, et al. Paired design study by real-time PCR: miR-378* and miR-145 are potent early diagnostic biomarkers of human colorectal cancer. BMC Cancer. 2015;15(1):158. | ||
Andersen M, Grauslund M, Ravn J, Sørensen JB, Andersen CB, Santoni-Rugiu E. Diagnostic potential of miR-126, miR-143, miR-145, and miR-652 in malignant pleural mesothelioma. J Mol Diagn. 2014;16(4):418–430. | ||
Gits CM, van Kuijk PF, Jonkers MB, et al. MicroRNA expression profiles distinguish liposarcoma subtypes and implicate miR-145 and miR-451 as tumor suppressors. Int J Cancer. 2014;135(2):348–361. | ||
Campayo M, Navarro A, Viñolas N, et al. Low miR-145 and high miR-367 are associated with unfavourable prognosis in resected nonsmall cell lung cancer. Eur Respir J. 2013;41(5):1172–1178. | ||
Wu D, Li M, Wang L, et al. microRNA-145 inhibits cell proliferation, migration and invasion by targeting matrix metallopeptidase-11 in renal cell carcinoma. Mol Med Rep. 2014;10(1):393–398. | ||
Kim TH, Song JY, Park H, et al. miR-145, targeting high-mobility group A2, is a powerful predictor of patient outcome in ovarian carcinoma. Cancer Lett. 2015;356(2 Pt B):937–945. | ||
Azizmohammadi S, Safari A, Azizmohammadi S, et al. Molecular identification of miR-145 and miR-9 expression level as prognostic biomarkers for early-stage cervical cancer detection. QJM. 2017;110(1):11–15. | ||
Slaby O, Svoboda M, Fabian P, et al. Altered expression of miR-21, miR-31, miR-143 and miR-145 is related to clinicopathologic features of colorectal cancer. Oncology. 2007;72(5–6):397–402. | ||
Wu BL, Xu LY, du ZP, et al. MiRNA profile in esophageal squamous cell carcinoma: downregulation of miR-143 and miR-145. World J Gastroenterol. 2011;17(1):79–88. | ||
Xia H, Yamada S, Aoyama M, et al. Prognostic impact of microRNA-145 down-regulation in adult T-cell leukemia/lymphoma. Hum Pathol. 2014;45(6):1192–1198. | ||
Tang M, Lin L, Cai H, Tang J, Zhou Z. MicroRNA-145 downregulation associates with advanced tumor progression and poor prognosis in patients suffering osteosarcoma. Onco Targets Ther. 2013;6:833–838. | ||
Naito Y, Yasuno K, Tagawa H, et al. MicroRNA-145 is a potential prognostic factor of scirrhous type gastric cancer. Oncol Rep. 2014;32(4):1720–1726. | ||
Mak IW, Singh S, Turcotte R, Ghert M. The epigenetic regulation of SOX9 by miR-145 in human chondrosarcoma. J Cell Biochem. 2015;116(1):37–44. | ||
He JH, Zhang JZ, Han ZP, Wang L, Lv YB, Li YG. Reciprocal regulation of PCGEM1 and miR-145 promote proliferation of LNCaP prostate cancer cells. J Exp Clin Cancer Res. 2014;33:72. | ||
Chen Y, Sun Y, Rao Q, Xu H, Li L, Chang C. Androgen receptor (AR) suppresses miRNA-145 to promote renal cell carcinoma (RCC) progression independent of VHL status. Oncotarget. 2015;6(31):31203–31215. | ||
Li C, Xu N, Li YQ, Wang Y, Zhu ZT. Inhibition of SW620 human colon cancer cells by upregulating miRNA-145. World J Gastroenterol. 2016;22(9):2771–2778. | ||
Ibrahim AF, Weirauch U, Thomas M, Grünweller A, Hartmann RK, Aigner A. MicroRNA replacement therapy for miR-145 and miR-33a is efficacious in a model of colon carcinoma. Cancer Res. 2011;71(15):5214–5224. | ||
Huang TC, Renuse S, Pinto S, et al. Identification of miR-145 targets through an integrated omics analysis. Mol Biosyst. 2015;11(1):197–207. | ||
Spizzo R, Nicoloso MS, Lupini L, et al. miR-145 participates with TP53 in a death-promoting regulatory loop and targets estrogen receptor-alpha in human breast cancer cells. Cell Death Differ. 2010;17(2):246–254. | ||
Li JM, Kao KC, Li LF, et al. MicroRNA-145 regulates oncolytic herpes simplex virus-1 for selective killing of human non-small cell lung cancer cells. Virol J. 2013;10:241. | ||
Khan S, Ebeling MC, Zaman MS, et al. MicroRNA-145 targets MUC13 and suppresses growth and invasion of pancreatic cancer. Oncotarget. 2014;5(17):7599–7609. | ||
Zhu X, Li Y, Xie C, et al. miR-145 sensitizes ovarian cancer cells to paclitaxel by targeting Sp1 and CDK6. Int J Cancer. 2014;135(6):1286–1296. | ||
Izzotti A, Calin GA, Arrigo P, Steele VE, Croce CM, De Flora S. Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke. FASEB J. 2009;23(3):806–812. | ||
Gao F, Sun X, Wang L, Tang S, Yan C. Downregulation of microRNA-145 caused by hepatitis B virus X protein promotes expression of Cul5 and contributes to pathogenesis of hepatitis B virus-associated hepatocellular carcinoma. Cell Physiol Biochem. 2015;37(4):1547–1559. | ||
Cui SY, Wang R, Chen LB. MicroRNA-145: a potent tumour suppressor that regulates multiple cellular pathways. J Cell Mol Med. 2014;18(10):1913–1926. | ||
Yu Y, Nangia-Makker P, Farhana L, Majumdar APN. A novel mechanism of lncRNA and miRNA interaction: CCAT2 regulates miR-145 expression by suppressing its maturation process in colon cancer cells. Mol Cancer. 2017;16(1):155. | ||
Guo YH, Gao FH, Shi J, Yuan HH, Jiang B. EGFR-ERK signaling pathway down-regulates miRNA-145 in lung cancer cells. Zhonghua Zhong Liu Za Zhi. 2013;35(3):187–192. | ||
Harada K, Baba Y, Ishimoto T, et al. Suppressor microRNA-145 is epigenetically regulated by promoter hypermethylation in esophageal squamous cell carcinoma. Anticancer Res. 2015;35(9):4617–4624. | ||
Lee SJ, Kim SJ, Seo HH, et al. Over-expression of miR-145 enhances the effectiveness of HSVtk gene therapy for malignant glioma. Cancer Lett. 2012;320(1):72–80. | ||
Sachdeva M, Zhu S, Wu F, et al. P53 represses c-Myc through induction of the tumor suppressor miR-145. Proc Natl Acad Sci USA. 2009;106(9):3207–3212. | ||
Kent OA, Chivukula RR, Mullendore M, et al. Repression of the miR-143/145 cluster by oncogenic Ras initiates a tumor-promoting feed-forward pathway. Genes Dev. 2010;24(24):2754–2759. | ||
Fuse M, Nohata N, Kojima S, et al. Restoration of miR-145 expression suppresses cell proliferation, migration and invasion in prostate cancer by targeting FSCN1. Int J Oncol. 2011;38(4):1093–1101. | ||
Doberstein K, Steinmeyer N, Hartmetz AK, et al. MicroRNA-145 targets the metalloprotease ADAM17 and is suppressed in renal cell carcinoma patients. Neoplasia. 2013;15(2):218–IN31. | ||
Gan B, Lim C, Chu G, et al. FoxOs enforce a progression checkpoint to constrain mTORC1-activated renal tumorigenesis. Cancer Cell. 2010;18(5):472–484. | ||
Ostenfeld MS, Bramsen JB, Lamy P, et al. miR-145 induces caspase-dependent and -independent cell death in urothelial cancer cell lines with targeting of an expression signature present in TA bladder tumors. Oncogene. 2010;29(7):1073–1084. | ||
Chen Z, Zeng H, Guo Y, et al. miRNA-145 inhibits non-small cell lung cancer cell proliferation by targeting c-myc. J Exp Clin Cancer Res. 2010;29(1):151. | ||
Wang S, Bian C, Yang Z, et al. miR-145 inhibits breast cancer cell growth through RTKN. Int J Oncol. 2009;34(5):1461–1466. | ||
Zhang J, Guo H, Qian G, et al. MiR-145, a new regulator of the DNA fragmentation factor-45 (DFF45)-mediated apoptotic network. Mol Cancer. 2010;9(1):211. | ||
Wang Y, Hu C, Cheng J, et al. MicroRNA-145 suppresses hepatocellular carcinoma by targeting IRS1 and its downstream Akt signaling. Biochem Biophys Res Commun. 2014;446(4):1255–1260. | ||
Wan X, Cheng Q, Peng R, et al. ROCK1, a novel target of miR-145, promotes glioma cell invasion. Mol Med Rep. 2014;9(5):1877–1882. | ||
Derouet MF, Liu G, Darling GE. MiR-145 expression accelerates esophageal adenocarcinoma progression by enhancing cell invasion and anoikis resistance. PLoS One. 2014;9(12):e115589. | ||
Koo S, Martin G, Toussaint LG. MicroRNA-145 promotes the phenotype of human glioblastoma cells selected for invasion. Anticancer Res. 2015;35(6):3209–3215. | ||
Lei P, Xie J, Wang L, Yang X, Dai Z, Hu Y. microRNA-145 inhibits osteosarcoma cell proliferation and invasion by targeting ROCK1. Mol Med Rep. 2014;10(1):155–160. | ||
Zhang J, Guo H, Zhang H, et al. Putative tumor suppressor miR-145 inhibits colon cancer cell growth by targeting oncogene Friend leukemia virus integration 1 gene. Cancer. 2011;117(1):86–95. | ||
Wang M, Wang J, Deng J, Li X, Long W, Chang Y. MiR-145 acts as a metastasis suppressor by targeting Metadherin in lung cancer. Med Oncol. 2015;32(1):344. | ||
Hu J, Qiu M, Jiang F, et al. MiR-145 regulates cancer stem-like properties and epithelial-to-mesenchymal transition in lung adenocarcinoma-initiating cells. Tumor Biol. 2014;35(9):8953–8961. | ||
Liu Y, Wu C, Wang Y, et al. MicroRNA-145 inhibits cell proliferation by directly targeting ADAM17 in hepatocellular carcinoma. Oncol Rep. 2014;32(5):1923–1930. | ||
Takagi T, Iio A, Nakagawa Y, Naoe T, Tanigawa N, Akao Y. Decreased expression of microRNA-143 and -145 in human gastric cancers. Oncology. 2009;77(1):12–21. | ||
Kou B, Gao Y, Du C, et al. miR-145 inhibits invasion of bladder cancer cells by targeting PAK1. Urol Oncol. 2014;32(6):846–854. | ||
Zhu Z, Xu T, Wang L, et al. MicroRNA-145 directly targets the insulin-like growth factor receptor I in human bladder cancer cells. FEBS Lett. 2014;588(17):3180–3185. | ||
Dong R, Liu X, Zhang Q, et al. miR-145 inhibits tumor growth and metastasis by targeting Metadherin in high-grade serous ovarian carcinoma. Oncotarget. 2014;5(21):10816–10829. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.