Back to Journals » Journal of Inflammation Research » Volume 18
From Brain to Lung: Emerging Insights into Mesenchymal Stem Cell-Derived Extracellular Vesicle-Associated Cargos in Ischemia-Reperfusion Injury
Authors Zhang Y, Shi X, Li S, Yan S, Mei L, Zou Y, Yan C
Received 27 February 2025
Accepted for publication 7 July 2025
Published 25 August 2025 Volume 2025:18 Pages 11645—11666
DOI https://doi.org/10.2147/JIR.S525208
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Dharmappa Krishnappa
Yiqing Zhang,1 Xiaodong Shi,2 Shuang Li,3 Shi Yan,4 Lyu Mei,4 Yuchen Zou,4 Chunhua Yan5
1Department of Integrated Traditional Chinese and Western Medicine (Geriatrics Department), The People’s Hospital Medical Group of Xiangzhou, Zhuhai, Guangdong, People’s Republic of China; 2Shenzhen Cadre and Talent Health Institute, Shenzhen Talent Institute, Shenzhen, Guangdong, People’s Republic of China; 3Department of Respiratory Medicine, The Third People’s Hospital of Longgang District, Shenzhen, Guangdong, People’s Republic of China; 4Pingshan District Central Hospital of Shenzhen, Shenzhen, Guangdong, People’s Republic of China; 5Department of General Medicine, Geriatric Medicine; Pingshan District Central Hospital of Shenzhen, Shenzhen, Guangdong, People’s Republic of China
Correspondence: Chunhua Yan, Department of General Medicine, Geriatric Medicine; Pingshan District Central Hospital of Shenzhen, Pingshan District, Shenzhen, Guangdong, People’s Republic of China, Email [email protected]
Abstract: Ischemic stroke represent major global health challenges due to their high incidence and debilitating consequences. Similarly, lung ischemia-reperfusion injury has gained increasing attention in the medical research community. The emerging concept of the lung-brain axis has shed new light on the complex interplay between these two organ systems during pathological injury, demonstrating that their interaction can significantly exacerbate ischemia- reperfusion damage. Although these conditions exhibit distinct pathophysiological characteristics, they share remarkable similarities in their genetic responses to tissue injury. Recent advancements in regenerative medicine have highlighted the therapeutic potential of mesenchymal stem cells (MSCs) in addressing both ischemic stroke and lung ischemia-reperfusion injury. Particularly, extracellular vesicles (EVs) derived from MSCs have emerged as a promising therapeutic avenue. These MSC-derived EVs exert their protective effects through multiple mechanisms, including immunomodulation, anti-inflammatory actions, anti-apoptotic effects, oxidative stress regulation, and promotion of angiogenesis. This comprehensive review aims to systematically examine the therapeutic roles of MSC-derived EVs in both cerebral and pulmonary ischemia-reperfusion injuries, while emphasizing their shared pathological mechanisms. By elucidating these common pathways, we hope to provide novel perspectives and research directions for both clinical and basic investigations into lung-brain ischemia-reperfusion injury.
Keywords: lung, brain, ischemia-reperfusion injury, mesenchymal stem cell, extracellular vesicles
Introduction
Ischemia-reperfusion injury involves a series of intricate and interconnected pathophysiological events that play a key role in various human diseases, including major health concerns like stroke, myocardial infarction, organ transplantation, acute kidney injury and lung injury.1–5 The cessation of blood flow leads to ischemia, causing rapid damage to metabolically active tissues. In the absence of organ cooling, irreversible injury can occur within minutes, with the extent of damage varying depending on the specific organ involved.6 Paradoxically, restoring blood flow initiates a cascade of events that can exacerbate cellular injury. Upon reperfusion, the biochemical and molecular alterations create conditions conducive to free radical-mediated damage. During reperfusion, when oxygen is introduced into ischemic and hypoxic tissues, it reacts with substances such as NADPH oxidase to produce a large amount of free radicals, which further exacerbates the damage to the ischemic and hypoxic tissues.7–9 The vascular endothelium plays a crucial role in inflammation by regulating the adhesion and infiltration of harmful leukocytes.10 Cerebral and lung ischemia-reperfusion injuries are major contributors to debilitating diseases and mortality in medicine, remaining among the most common causes of these outcomes.4 Cerebral ischemia-reperfusion injury typically results from conditions like stroke. Lung ischemia-reperfusion injury often occurs during procedures such as lung transplantation or certain types of surgery.11 The damage results from inflammatory responses, oxidative stress, and endothelial dysfunction, leading to pulmonary edema, impaired gas exchange, and potential organ failure. Although lung and brain ischemia-reperfusion injuries have significant differences in etiology, they share a high degree of homogeneity in pathology. Recent research has been exploring new methods to reduce the damage caused to organs by ischemia-reperfusion injury. Among these, extracellular vesicles (EVs) have become a hot topic in research due to their abilities to regulate intercellular communication and promote tissue repair.12,13
EVs, are vesicles enclosed by a lipid bilayer membrane, released by all cell types under both normal and pathological conditions.14 EVs are generally classified into three types based on their size and biogenesis: exosomes (less than 150 nm in diameter), microvesicles (MVs) (ranging from 100 to 1000 nm in diameter), and apoptotic bodies (ranging from 0.8 to 5.0 µm in diameter).15,16 These EVs sourced from almost all biological fluids, such as blood, urine, saliva, cerebrospinal fluid, and lavage fluid.17–19 EVs are increasingly recognized as an important mode of intercellular communication, facilitating the transfer of proteins, lipids, bioactive metabolites, RNA and DNA between cells.20 Cargo RNAs of extracellular vesicles consists of various biotypes, which represent a specific subset of the RNA content from the originating cell. In a few cases, DNA has been identified.21 But overall, EVs are primarily enriched with small RNAs, and other species such as mRNAs, miRNAs, non-coding RNAs, and circular RNAs.22 These nucleic acids are shuttled between cells and can regulate key aspects of the onset and progression of ischemia-reperfusion injury.23 Due to their altered expression following ischemia, these nucleic acids have emerged as potential biomarkers and novel therapeutic targets.24–26 This review provides a comprehensive overview of recent literature on the role of EVs-associated cargos, in the progression between cerebral and lung ischemia-reperfusion injuries. It examines the cellular and subcellular origins of EVs-associated cargos, their cellular targets, and their biological effects, with a focus on their potential as biomarkers and therapeutic targets.
Brain and Lung are Eminent Players for Ischemia-Reperfusion Injury with a Tight Interaction
Ischemia-reperfusion injury is a systemic condition characterized by a complex interplay of interrelated pathophysiological processes.27 These include acute heart failure, cerebral dysfunction, lung injury, gastrointestinal impairment, systemic inflammatory response syndrome, and the progression to multiple organ dysfunction syndrome.27,28 Due to the unique characteristics of the brain-lung axis, the brain and lungs are prominent and interrelated players in various diseases, including ischemia-reperfusion injury.29–31 For instance, Dong et al identified a U-shaped association between low-density lipoprotein cholesterol levels and the risk of futile reperfusion in acute ischemic stroke following endovascular thrombectomy. Notably, stroke-associated pneumonia was recognized as a significant mediating mechanism in this study.32 Research indicates that following cerebral ischemia-reperfusion injury, inflammatory cells and factors from the affected area can penetrate the blood-brain barrier and enter systemic circulation, leading to secondary lung injury.33 Moreover, stroke may impact the stability of the immune system, suppressing immune responses and consequently resulting in the development of immune-related pneumonia.34 Correspondingly, diseases associated with pulmonary ischemia-reperfusion can also adversely affect the brain.35 For instance, pulmonary embolism may cause respiratory failure, leading to systemic hypoxia and subsequently causing cerebral hypoxic damage.36 Additionally, after pulmonary ischemia-reperfusion injury, a multitude of inflammatory cells (such as macrophages) are activated, and a significant release of inflammatory factors (like TNF-α, proteases, etc.) occurs.37,38 Once these inflammatory cells and factors enter the bloodstream, they may disrupt the blood-brain barrier and damage the endothelial cells of cerebral blood vessels, ultimately causing harm to brain tissue.37 In the context of brain abnormalities caused by hypoxia, the relationship between respiratory diseases, particularly chronic respiratory conditions, and brain dysfunction has been extensively studied.39–41 Lung ischemia-reperfusion and its impact in nervous system is still a popular research field especially in the context that lung transplantation has been established worldwide as the last treatment for end-stage respiratory failure.5 Furthermore, the nervous system is highly sensitive to oxygen supply, and despite the introduction of therapeutic hypothermia, meaningful neurological recovery remains poor in over 80% of the 500,000 cardiac arrest cases occurring annually in the United States.42–44 Therefore, the brain-lung axis highlights the widely studied interaction between the brain and lungs in pathophysiology, making it essential to consider brain and lung injury separately from other systems when researching ischemia-reperfusion injury. This is not only because they are crucial and sensitive participants in ischemia-reperfusion injury, but also due to their complex interaction, which will be discussed in detail.
Pulmonary complications are common after severe brain injury, it includes pneumonia, pleural effusion, acute lung injury, acute respiratory distress syndrome, and neurogenic pulmonary edema.45 First, brain injury can lead to lung dysfunction through mobility impairment, such as aspiration-associated pneumonia, hypostatic pneumonia, and neuromuscular respiratory failure.46,47 In detail, in the acute phase following brain injury, neuroinflammatory responses trigger many inflammatory cell infiltrations and the release of inflammatory factors. These inflammatory cells and factors not only exacerbate neural damage in the brain but also compromise the integrity of the blood–brain barrier. Once these inflammatory cells and factors enter the bloodstream, they can induce systemic inflammatory responses and may further aggravate lung injury.48,49 Following ischemic injury of brain, patients are prone to developing pulmonary infections due to factors such as dysphagia and stroke-related immune suppression.50,51 Studies have shown that approximately 8.5% to 14.3% of stroke patients will develop secondary pneumonia. In the intensive care unit, this proportion rises to as high as 28%.52 Although the recovery of neurological function following ischemia-reperfusion may improve stroke-associated pneumonia, the brain injury that occurs after reperfusion could also potentially worsen pulmonary infections.11,53 Secondly, in the context of neurological regulation, brain injury can induce acute pulmonary edema by increasing intracranial pressure, which elevates sympathetic activity, resulting in heightened vascular resistance and changes in blood vessel tone.30 In summary, following brain injury, particularly ischemic brain injury, a variety of mechanisms come into play to profoundly affect the lungs. These mechanisms interact and work in concert, ultimately not only significantly increasing the risk of pulmonary injury and infection but also exerting highly adverse effects on patient prognosis.
Conversely, pulmonary dysfunction can also worsen cerebral hypoxia, creating a vicious cycle that further exacerbates intracranial pressure through the role of inflammation and reactive oxygen species.54 The situation might worsens if lung ischemia-reperfusion participates in this cycle, not only by aggravating hypoxia after ischemia-reperfusion through non-specific alveolar damage and pulmonary edema,28 but also by directly releasing numerous harmful factors, such as endotoxemia, ROS, cell death-related molecules, and inflammatory factors into circulation, thereby enhancing brain dysfunction.55–57
MSCs and MSC-EVs
Mesenchymal stem cells (MSCs) are a type of multipotent, self-renewing, adult stem cells with multidirectional differentiation potential and immune-regulating functions.58 MSCs are known for their unique immunomodulatory properties, which allow them to evade immune recognition and suppress immune responses.59 Due to their regenerative potential, MSCs have emerged as a promising therapeutic strategy in regenerative medicine.60 MSCs with different origins have been reported to be used in the treatment of ischemic stroke and lung ischemia injury.59,61–66 However, challenges such as donor heterogeneity, immunogenicity, potential carcinogenic risks, effective delivery, and ethical challenges of MSCs must be addressed. As a result, the further advancement of stem cell therapy faces significant obstacles. Recent evidence suggests that the beneficial effects of stem cell therapy are likely driven by the paracrine mechanisms of MSCs, particularly through the secretion of EVs.67,68 MSC-EVs participate in intercellular communication and modulate the functions of cells and tissues by carrying and releasing a variety of bioactive substances, including proteins, DNA, and RNA.69 Compared with MSCs, MSC-EVs offer higher safety, lower immunogenicity, and easier accessibility. Leveraging these advantages, MSC-EVs have been widely applied in the treatment of various diseases, such as cerebral ischemic injury, pulmonary injury, renal injury, tumor progression, osteoarthritis, fracture healing, and degenerative bone diseases, demonstrating great potential for clinical applications.70
MSC-EVs are typically classified into two groups based on their biogenesis: EVs formed by inward budding of the endolysosomal membrane during the maturation of multivesicular endosomes and EVs formed by direct shedding from the plasma membrane.71,72 MSC-EVs not only carry MSC-specific markers, including CD29, CD73, CD90, CD44, and CD105, but also display typical EVs markers such as CD63, CD9, and CD81, which facilitate their migration to injured and inflamed tissues.73 For example, Zhao et al reported that inhalation of MSC-EVs can attenuate acute lung injury in mice through anti-inflammatory and anti-oxidant activity primarily by modulating Nrf2.74 Besides, the membrane of MSC-EVs is rich in cholesterol, sphingomyelin, ceramide, and lipid raft proteins, which facilitate membrane fusion with target cells and allow MSC-EVs to travel through the body, overcoming biological barriers.75 After being taken up by target cells, MSC-EVs can release their cargo of bioactive substances, such as proteins and cytokines, thereby modulating immune responses and promoting tissue regeneration and repair.76 Around 40 functional proteins have been identified as responsible for the protective effects of MSC-EVs.73 In a mouse model of acute myocardial infarction, MSC-EVs overexpressing AKT exhibited elevated levels of platelet-derived growth factor D (PDGF-D), enhancing the angiogenic effect of MSC-EVs and thereby improving cardiac function.77 Huang et al demonstrated that MSC-EVs alleviated cerebral ischemia/reperfusion injury by preventing neural cell death through the inhibition of caspase-9 and caspase-3.77,78 By delivering miRNAs and other biological active molecules, MSC-EVs can influence immunomodulation, tissue regeneration, apoptosis inhibition, and tumor progression to promote their survival and regeneration.79 Thus, MSC-EVs could serve as a promising alternative to MSC therapy.
MSC-EVs Therapy in Brain Ischemia-Reperfusion Injury
Inhibition of Inflammatory Responses
The onset of neuroinflammation is both beneficial for the clearance of necrotic tissue and can lead to the destruction of the nervous system. MSCs and MSC-EVs can act on different components in the CNS (Central nervous system) to inhibit inflammatory responses and protecting the CNS from damage. Microglia are the resident immune cells of the CNS, and play essential roles in neuroinflammation.80 MSCs can suppress the activation of pro-inflammatory microglia and reduce the release of inflammatory factors, thereby inhibiting neuroinflammation.81–84 Cui et al administered the isolated umbilical cord mesenchymal stem cell-derived extracellular vesicles (UC-MSCs-EVs) into the ventricles of a rat model. The result indicated that MSC-EVs can suppress the activation of microglia and promote the recovery of neurological function.85 By administering Bone Marrow Mesenchymal Stem Cell-derived EVs (BMSC-EVs) into the MCAO (Middle Cerebral Artery Occlusion) model, Liu et al discovered that BMSC-EVs can suppress the NLRP3 inflammasome in neurons, thereby inhibiting inflammatory responses.86 Furthermore, their research indicates that MSC-EVs can induce the transition of pro-inflammatory microglia (M1) to anti-inflammatory phenotypes (M2), thereby suppressing neuro-inflammation.86 Specific research findings by Dong et al indicated that miR-23a-3p, derived from human umbilical cord mesenchymal stem cells exosomes (hUCMSCs-EVs), inhibit the activation of microglia and M1 polarization.87 In addition to the aforementioned information, the intracellular signaling pathway mechanisms by which MSC-EVs regulate the activity of microglia are also gradually being unveiled. Zhang et al discovered that the administration of hUMSC-EVs into a mouse model of ischemic injury reduced the volume of the infarct and suppressed the activation of microglia.88 Furthermore, hUMSC-EVs derived miR-146a-5p can inhibit the NF-κB signaling pathway, reducing the production of inflammatory factors in glial cells.88 MSC-EVs inhibit the microglial NRF2/NF-κB/NLRP3 signaling pathway to reduce the expression of inflammatory factors (IL-6 and TNF-α) and the activity of inflammatory cells, thereby alleviating inflammatory responses and oxidative stress.89 MSC-EVs not only regulate the activation status and functions of microglia and neurons but also modulate astrocytes. Astrocytes, another crucial type of immune cell in neuro-inflammation, are also regulated by MSC-EVs. Xian et al found that MSC-EVs can alleviate the inflammatory response of activated astrocytes, and their mechanism of action may involve the modulation of the Nrf2-NF-κB signaling pathway.90 By using BMSCs-miR-138-5p to treat MCAO, Deng et al found that BMSCs-miR-138-5p could be integrated into astrocytes and reduce the expression of their inflammatory factors.91 Further research indicates that MSC-EVs-miR-138-5p promote astrocytes proliferation and inhibit inflammatory responses by regulating LCN2.91 Therefore, MSC-EVs inhibit inflammatory responses by protecting neuronal cells, regulating the polarization of microglia, suppressing the release of inflammatory factors, modulating the activity of astrocytes, and regulating intracellular signaling pathways in neurons, thereby promoting the repair of damage.
Inhibition of Apoptosis
Apoptosis, a form of programmed cell death, is induced by damage and inflammatory responses that occur subsequent to ischemia-reperfusion. Such cell death, a consequence of injury and inflammation, results in the loss of cells within the CNS, notably neurons. Consequently, the inhibition of cellular apoptosis can reduce the detrimental effects of post-ischemic reperfusion injury.92 Over recent years, a multitude of studies have documented the suppressive effects of MSC-EVs on apoptosis, as well as their protective impact against ischemic damage. Ye et al demonstrated that MSC-EVs reduce apoptosis and enhance the migratory ability of neural cells.93 Similarly, by intranasal administration of MSC-EVs, Ikeda et al found that MSC-EVs can reduce cellular apoptosis and promote neuronal survival.94 MSC-EVs can also inhibit apoptosis in neural cells by regulating intracellular signaling pathways. In this context, caspase family proteins play a crucial role in regulating cellular apoptosis.95 Liu et al found that MSC-EVs can inhibit apoptosis in neural cells by suppressing caspase family proteins.96 Xiao et al found that miR-134 derived from BMSC-EVs can inhibit the expression of caspase-8 and suppress oligodendrocyte apoptosis.97 Kang et al found that BMSCs-miR-23b-3p could inhibit the expression of NEK7 on microglia and suppress microglial pyroptosis.98 Further experiments demonstrated that MSC-EVs-miR-23b-3p can inhibit pyroptosis in microglia by suppressing NEK7.98 Moreover, MSC-EVs reduce pyroptosis in neural cells by targeting Absent in Melanoma 2(AIM2) through miR-193b-5p.99 Similarly, MSC-EVs-miR-22-3p can inhibit the KDM6B/BMP2/BMF axis to reduce neuronal apoptosis and infarction volume.100 Besides, MSC-EVs-miR-145 can significantly inhibit the expression of FOXO1 therefor inhibiting apoptosis in OGD/R-treated BV2 cells.101 MSC-EVs-miR-26a-5p also inhibit microglia apoptosis by suppressing the expression of CDK6, thereby alleviating cerebral ischemia-reperfusion injury.102 In summary, these studies have unveiled the crucial roles of MSC-EVs and their secreted miRNAs in modulating neuroinflammation and promoting neuronal survival. The findings demonstrate that MSC-EVs can effectively regulate microglia and neurons, and significantly alleviate ischemia-reperfusion injury. These discoveries offer novel insights and potential therapeutic strategies for the treatment of cellular damage in neurological diseases (Figure 1 and Table 1).
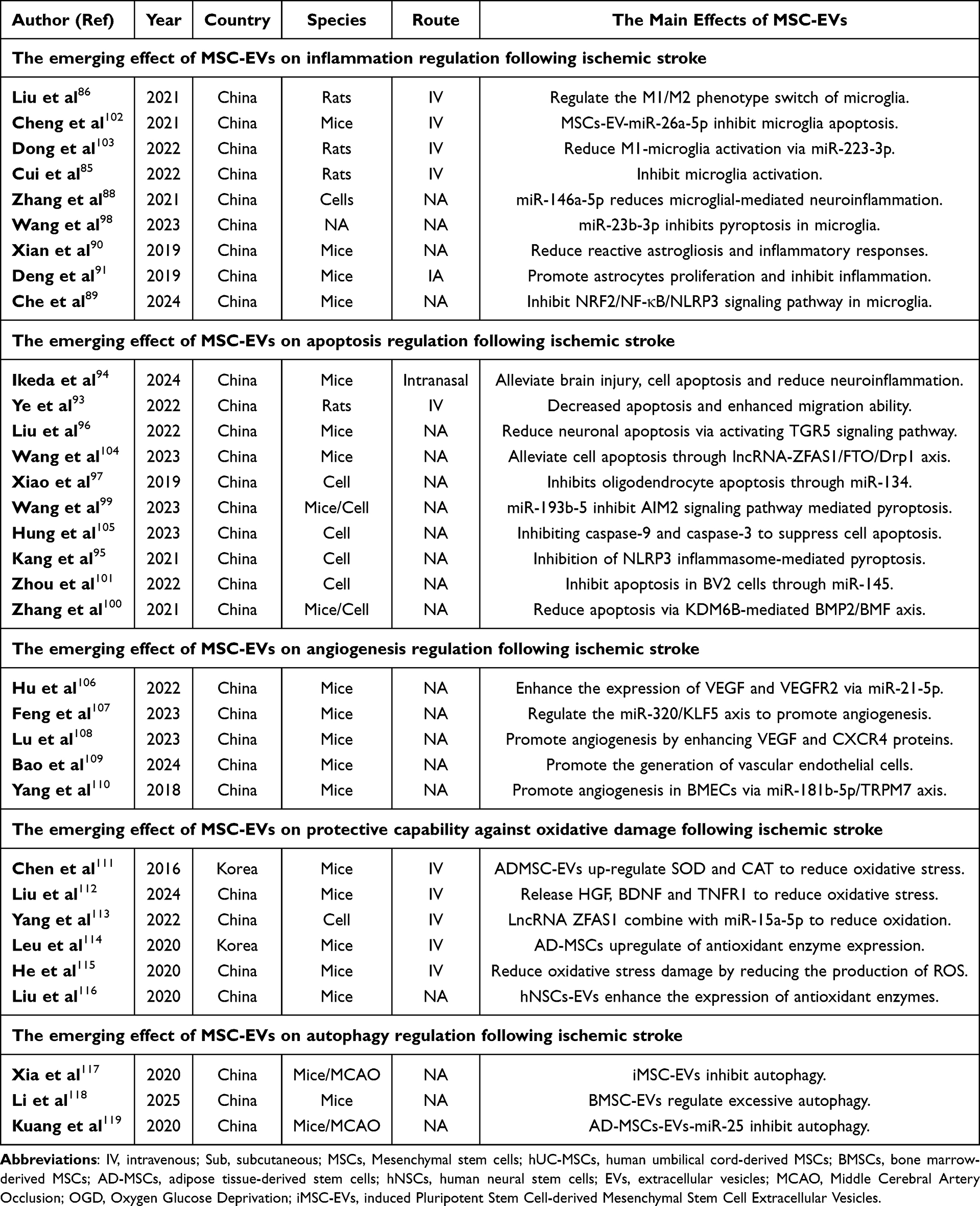 |
Table 1 MSC-EVs Therapy in Brain Ischemia-Reperfusion Injury |
 |
Figure 1 The mechanisms of action of extracellular vesicles (EVs) derived from different types of mesenchymal stem cells (MSCs) in cerebral ischemia-reperfusion injury. MSCs from various sources, including hUC-MSCs, BMSCs, OM-MSCs, (hiPS-MSCs, AD-MSCs, and hNSCs, secrete EVs. These EVs contain various factors, such as miR-193b-5p, miR-22-3p, miR-145, miR-26a-5p, lncRNA ZFAS1, miR-23a-3p, miR-146a-5p, circDLGAP4, miR-138-5p, miR-134, miR-23b-3p, miR-21-5p, VEGF, CXCR4, miR-181b-5p, miR-25, and others. These factors can exert the following effects: The factors can influence apoptosis in neural cells by acting on pathways involving AIM2, FOXO1, KDM6B/BMP2/BMF, CDK6, Caspase8, and NEK7. The factors can affect angiogenesis by acting on VEGF, VEGFR2, ANG I, the PTEN/Akt axis, TRPM7 axis, STAT3, CXCR4, and the miR-320/KLF5 pathway. They can influence autophagy by acting on miR-25, Nrf2, STAT3, and Cav-1. miR-15a-5p can influence oxidative stress. E. They can affect inflammation by acting on pathways involving IL-6 and TNF. Abbreviations: MSCs, Mesenchymal stem cells; hUC-MSCs, human umbilical cord-derived MSCs; BMSCs, bone marrow-derived MSCs; OM-MSCs, oral mucosa-derived MSCs; AD-MSCs, adipose tissue-derived stem cells; hNSCs, human neural stem cells; EVs, extracellular vesicles; hiPS-MSC, Human Induced Pluripotent Stem Cell-derived Mesenchymal Stromal Cells. |
Promotion of Angiogenesis
In brain ischemia-reperfusion injury, angiogenesis enables the formation of new blood vessels and regulates neovascularization, thereby reducing the necrosis of ischemic and hypoxic tissues and promoting the recovery of neurological functions. Hu et al found that in MCAO, BMSC-EVs could reduce infarct volume, increase microvascular density, and upregulate the expression of miR-21-5p.106 In vitro study, the demonstrated that BMSCs-EVs-miR-21-5p upregulate microvascular density and enhance the expression levels of VEGF, VEGFR2, Ang-1, and Tie-2, thereby promoting angiogenesis.106 Similarly, human-induced pluripotent stem cell-derived MSC-EVs (hiPS-MSC-EVs) enhance angiogenesis in the ischemic region by expressing VEGF and CXCR4 proteins.108 Various signaling pathways and axis also play important roles in angiogenesis. Such as Feng et al discovered that hUC-MSCs-Exos-circDLGAP4 can elevate the expression levels of circDLGAP4 and, moreover, hUC-MSCs-Exos-circDLGAP4 can suppress the miR-320/KLF5 signaling pathway.107 Therefore, the hUC-MSCs-EVs promote angiogenesis after cerebral ischemia-reperfusion by regulating the miR-320/KLF5 axis.107 Bao et al found that BMSC-EVs can improve functional recovery of the brain after ischemia and promote vascular regeneration.109 Further experiments have demonstrated that BMSC-EVs promote angiogenesis after cerebral ischemic injury by releasing miR-486 and modulating the PTEN/Akt signaling pathway.109 Besides, Yang et al discovered that exosomes derived from adipose-derived stem cells (ADSCs-EVs) promote the generation of brain microvascular endothelial cells (BMECs) in rats after OGD through the miR-181b-5p/TRPM7 axis.110 The aforementioned studies have revealed that various types of MSC-EVs promote angiogenesis following cerebral ischemia-reperfusion injury by carrying multiple miRNAs to modulate intracellular signaling pathways.
Protective Capability Against Oxidative Damage
Oxidative stress is a crucial factor contributing to cellular and tissue damage following ischemic stroke. After ischemic stroke, cells and tissues are disrupted, leading to a significant increase and accumulation of ROS and other oxidative substances.120 The accumulation of these oxidative substances can cause damage to cellular organelles, cell death, and tissue destruction, thereby leading to adverse outcomes.121 Therefore, regulating the oxidative stress response and reducing the harmful effects of oxidative stress can improve the prognosis of ischemic stroke.121 In recent years, MSC- EVs have been shown to modulate oxidative stress following ischemic stroke, attenuate the detrimental effects of oxidative stress, and protect cells and tissues.122 NcRNAs derived from MSC-EVs also exhibit antioxidant and anti-inflammatory effects. Yang et al have shown that lncRNA ZFAS1 from MSC-EVs can bind to miRNA-15a-5p in damaged cells and regulate its downstream signaling pathways, thereby effectively alleviating cell and tissue damage caused by oxidative stress.113
ADMSC-EVs have emerged as a promising therapeutic direction for ischemic stroke due to their antioxidant properties.111 Chen et al injected ADMSC-EVs into MCAO model and assessed the therapeutic prognosis. The study found that oxidative stress markers, such as malondialdehyde (MDA) and superoxide dismutase (SOD), were significantly reduced in the MCAO model treated with ADMSC-EVs.111 Therefore, Chen et al demonstrated that ADMSC-EVs have antioxidant effects and can reduce oxidative damage.111 Additionally, Leu et al administered ADMSCs intravenously into the MCAO model. Their results indicated that both ADMSCs and ADMSC-EVs could reduce oxidative stress markers in brain tissue following ischemic stroke.114
hUC-MSCs also have the ability to regulate oxidative responses, and hUC-MSC-EVs are the main components through which they exert this regulatory effect.112 Liu et al injected hUC-MSCs into MCAO model to investigate their effects on oxidative stress.112 The results showed that hUC-MSC-EVs reduced MDA levels and increased SOD levels, thereby enhancing antioxidant capacity.112 Additionally, tumor necrosis factor receptor 1 (TNFR1) further upregulated the expression of quinone oxidoreductase 1 (NQO1) and heme oxygenase-1 (HO-1) to achieve antioxidant effects.112 The mechanisms underlying the effects of hUC-MSC-EVs on ischemic stroke remain to be further explored.
Olfactory Mucosa Mesenchymal Stem Cells (OM-MSCs) and their OM-MSCs-EVs also exhibit antioxidant properties. He et al injected OM-MSCs and OM-MSCs-EVs into the MCAO model and assessed their effects.115 The results demonstrated that oxidative stress markers (such as SOD and MDA) were reduced in the MCAO model, along with decreased cell death and tissue damage.115 The experiments by He et al confirmed the protective effects of OM-MSCs and OM-MSCs-EVs on brain cells and tissues following ischemic stroke, laying the foundation for further exploration of the underlying mechanisms.115
Human neural stem cells (hNSCs) and hNSCs-EVs exhibit neuroprotective properties and play a protective role following ischemic stroke. Liu et al treated an OGD model with hNSCs-EVs and investigated their effects.116 The experiments demonstrated that hNSCs-EVs can activate the Nrf2 signaling pathway, thereby promoting the expression of antioxidant genes.116 Following OGD treatment, levels of oxidative stress markers such as SOD and MDA were significantly reduced in cells treated with hNSCs-EVs.116 Through these mechanisms, hNSCs-EVs mitigate oxidative stress following ischemic stroke, thereby protecting neural tissue and improving the prognosis of ischemic stroke.
Regulation on Autophagy
After ischemic stroke, ischemia and hypoxia cause damage to cells and tissues. Damaged cells use autophagy to remove necrotic and damaged organelles and clear toxic substances within the cells.123 This protects cells from destruction by harmful substances, safeguards neurons, and improves the prognosis of ischemic stroke.123 Autophagy is a self-regulatory mechanism within cells that degrades damaged organelles and clears harmful substances via lysosomes.124 Following ischemic stroke, hypoxia leads to the damage of cellular organelles and the accumulation of harmful substances within cells.124,125 Moderate autophagy is protective for cells. However, excessive autophagy can lead to cell death and exacerbate damage to the nervous system.123 Kuang et al demonstrated that ADMSC-EVs administration in MCAO models alleviated ischemic injury by reducing autophagy markers (LC3-II/LC3-I ratio and p62 levels) through miR-25-mediated regulation of ATG7 expression.119 Similarly, Liu et al reported that hNSC-EVs exerted neuroprotective effects post-ischemic stroke through dual mechanisms: Nrf2 pathway activation and autophagy modulation (upregulation of LC3 and p62 expression).116 These findings collectively indicate that EV-mediated autophagy regulation represents a crucial therapeutic mechanism against ischemic injury, with different EV sources employing distinct molecular pathways to enhance cellular clearance and neuronal protection.
Xia et al administered MSC-EVs in MCAO models to explore their effects on autophagy and neurological outcomes, revealing that iMSC-EVs activated the STAT3 signaling pathway.117 This activation promoted angiogenesis, indicating iMSC-EVs exert neuroprotective effects and improve ischemic stroke prognosis through autophagy inhibition-mediated angiogenesis enhancement. Beyond vascular regeneration and repair, the BBB is a vital component of the nervous system and plays a crucial role in the prognosis of ischemic stroke.126 After stroke, the blood-brain barrier can maintain the stability of the brain’s internal environment, suppressing inflammatory responses, and facilitating tissue repair. Following ischemic stroke, lysosomes in damaged cells degrade tight-junction proteins in endothelial cells, such as ZO-1 and Occludin.118 This leads to the disruption of the BBB, thereby worsening the prognosis of ischemic stroke.126,127 Li et al injected BMSCs-EVs into OGD and MCAO models and observed the expression of tight-junction proteins and Caveolin-1 (Cav-1).118 The results indicated that BMSCs-EVs could promote the expression of tight-junction proteins and reduce the impact of cellular autophagy on these proteins, thereby preserving the integrity of the BBB.118 Additionally, BMSCs-EVs were able to directly target Cav-1 and inhibit autophagy mediated by Cav-1.118 Figure 1 and the accompanying Table 1 provide comprehensive data on MSC-EVs’ therapeutic effects in ischemic stroke.
MSC-EVs Therapy in Lung Ischemia-Reperfusion Injury
A growing body of research highlights the pivotal role of MSC-EVs in influencing the outcomes of lung ischemia-reperfusion injury. To comprehensively assess this area, a systematic search was conducted to identify relevant studies examining the therapeutic effects of MSC-EVs, including exosomes, on lung ischemia-reperfusion injury. Key databases such as PubMed, the Cochrane Library (accessed up to November 2024), Web of Science, and EMBASE (covering studies from 1990 to November 2024) were utilized. Search terms included combinations of “lung”, “extracellular vesicles”, “exosomes”, “MSCs”, and “ischemia-reperfusion injury”, employing Boolean operators to optimize results. The review adhered to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. To ensure thoroughness, reference lists of the retrieved articles were manually screened for additional eligible studies. This iterative process was repeated until no new publications were identified. Ultimately, eight studies met the inclusion criteria, originating from the United States, Italy, and China, and published between 2017 and 2024.128–136 These studies identified various cargos carried by MSC-derived EVs in different functional states, which exhibited distinct effects on the prognosis of lung ischemia-reperfusion injury. Specific ncRNAs identified as cargos included miR-21-5p, miR-381, miR-335, lncRNA‑ZFAS1, miR-202-5p. These cargos demonstrated either beneficial or detrimental influences on injury outcomes, as detailed in Figure 2 and Table 2.
 |
Table 2 Preclinical Studies Evaluating the Impact of MSC-Derived Extracellular Vesicles on Lung Ischemia-Reperfusion Injury |
 |
Figure 2 In lung ischemia-reperfusion injury, MSC-EVs exert protective effects on lung tissue. BM-MSC-EVs suppress inflammatory responses by releasing miR-38, miR-335, TNF-α, IL-1β, and miR-21-5p. AD-MSCs-EVs inhibit inflammatory responses by releasing miR-206 to suppress the NF-κB pathway. hUC-MSCs-EVs reduce inflammation by inhibiting IL-17, TNF-α, CXCL1, HMGB1, and TNF-α. BM-MSCs-EVs inhibit pyroptosis by releasing miR-202-5p to regulate CMPK2. BM-MSC-EVs suppress apoptosis by releasing miR-335, miR-21-5p, and LncRNA-ZFAS1. AD-MSCs-EVs inhibit apoptosis by releasing miR-206 to suppress CXCL1. Abbreviations: MSCs, Mesenchymal stem cells; EVs, extracellular vesicles; BMSCs, bone marrow-derived MSCs; AD-MSCs, adipose tissue-derived stem cells; hNSCs, human neural stem cells. |
EVs Derived from BM-Derived Mesenchymal Stem Cells
BM-MSCs are multipotent stromal cells with immense potential in regenerative medicine.137 These cells, derived from bone marrow, possess multilineage differentiation potential and can differentiate into various cell types, including osteoblasts, chondrocytes, adipocytes, and myocytes.138 Moreover, BM-MSCs exhibit robust secretory functions, capable of secreting a variety of cytokines, such as vascular endothelial growth factor (VEGF) and insulin-like growth factor (IGF).138 Therefore, BM-MSCs hold great potential for applications in tissue repair and regeneration.138 Renowned for their strong immunomodulatory and anti-inflammatory properties, BM-MSCs exhibit low immunogenicity, enabling their use in both autologous and allogeneic treatments. They produce various bioactive molecules, such as cytokines and growth factors, which regulate immune responses, enhance angiogenesis, and support tissue healing.139 BM-MSCs can modulate immune responses to suppress inflammation, while also promoting the repair of endothelial cells, improving tissue blood supply, and thereby reducing the degree of fibrosis.140,141 Their therapeutic effects are mediated through mechanisms like direct differentiation, paracrine signaling, and immune modulation. Despite their advantages, challenges like donor-dependent variability, limited proliferation capacity, and risks of unintended differentiation pose obstacles to their clinical application.142 To overcome these limitations, EVs derived from BM-MSCs are being developed as a cell-free therapeutic alternative, offering comparable benefits without the complications associated with cell-based therapies. BM-MSCs continue to be a foundational element of regenerative medicine, with ongoing advancements aimed at refining their clinical efficacy and broadening their therapeutic applications.
Increasing evidence suggests that EVs derived from BM-MSCs can suppress inflammatory responses, inhibit apoptosis and pyroptosis, and promote the differentiation of regulatory T cells.128–136 In terms of anti-inflammatory effects, EVs derived from BM-MSCs not only suppress inflammation in lung microvascular endothelial cells in vitro but also effectively reduce inflammatory responses in lung tissue in vivo. Li et al and Gao et al indicated that EVs derived from BM-MSCs carrying miR-21-5p and lncRNA‑ZFAS1 can inhibit the inflammation in the pulmonary endothelial cells, respectively.130,131 Additionally, EVs derived from BM-MSCs carrying miR-21-5p can inhibit M1 polarization of alveolar macrophages and reduce the secretion of inflammatory cytokines such as HMGB1, IL-8, IL-1β, IL-6, IL-17, and TNF-α.131 When BM-MSCs are pre-challenged by hypoxia/reoxygenation, the expression level of miR-21-5p in the exosomes they secrete is significantly increased, thereby enhancing the anti-inflammation effects of these MSC-derived exosomes.131 Conversely, pre-treatment of BM-MSCs with miR-21-5p antagomir results in an opposing effect, reducing the therapeutic benefits of the exosomes.131 From a tissue perspective, BMSC-derived EVs can help alleviate lung inflammation after ischemia-reperfusion injury by upregulating miR-335 expression.136 This mechanism activates the NF-κB signaling pathway. Additionally, both lncRNA-ZFAS1 and miR-21-5p are involved in the inflammatory suppression at the lung tissue level, further contributing to the therapeutic effects of BMSC-derived EVs.130,131
Pyroptosis is a form of programmed cell death that is triggered by intracellular pathogen infections and is associated with an inflammatory response. The excessive activation of pyroptosis promotes the release of inflammatory factors and intracellular oxidative substances, thereby exacerbating the inflammatory response.143 Therefore, it is one of the key factors contributing to lung ischemia-reperfusion injury. In their study, Sun et al demonstrated that BM-MSC-derived EVs containing miR-202-5p can inhibit pyroptosis and mitigate lung injury caused by ischemia-reperfusion.135 They identified that miR-202-5p targets CMPK2, a critical regulator of pyroptosis.135 Mechanistically, the interaction between miR-202-5p and CMPK2 was validated using dual-luciferase reporter assays and RNA immunoprecipitation. The results confirm that BMSC-derived EVs containing miR-202-5p effectively suppress pyroptosis and prevent the progression of lung ischemia-reperfusion injury by targeting CMPK2135 (Figure 2 and Table 2).
Apoptosis is a form of programmed cell death that plays a crucial role in maintaining cellular homeostasis and eliminating damaged or dysfunctional cells. Pyroptosis and apoptosis are both forms of programmed cell death. The main difference between them is that apoptosis is typically an orderly and relatively mild process, in which cells maintain their basic structure during death and do not trigger a strong inflammatory response.144 In contrast, pyroptosis is an inflammatory form of cell death, characterized by cell rupture and the release of inflammatory factors, thereby inducing an inflammatory response.145 In ischemia-reperfusion injury, both apoptosis and pyroptosis can be activated, but their impacts on tissues differ. Therefore, intervening in the distinct mechanisms of apoptosis and pyroptosis can help alleviate ischemia-reperfusion injury.146 In studies examining cell death, the TUNEL assay is commonly used to detect apoptosis, and the apoptosis index is calculated to quantify the level of cell death.135 Additionally, the protein levels of apoptosis-related proteins, such as caspase-3, cleaved-caspase-3, caspase-9, and cleaved-caspase-9, are often assessed to further evaluate the extent of apoptosis.136 Zhang et al demonstrated that EVs BM-MSCs regulate the NF-κB signaling pathway and significantly reduce the level of apoptosis in lung tissues after ischemia-reperfusion injury in rats. This protective effect is mediated by miR-335, which plays a critical role in reducing lung damage and apoptosis. These findings highlight the therapeutic potential of BM-MSC-derived EVs in mitigating apoptosis and promoting tissue repair in ischemia-reperfusion injury.135 Li et al131 further showed that MSC-derived EVs alleviated ischemia-reperfusion injury-induced apoptosis in primary murine pulmonary endothelial cells by suppressing both intrinsic and extrinsic apoptotic pathways. This effect was mediated through miR-21-5p, which targets PTEN and PDCD4. Additionally, artificially overexpressing PTEN or PDCD4 in vitro significantly reduced the anti-apoptotic effect of MSC-EVs, highlighting the crucial role of these targets in the protective action of MSC-EVs against apoptosis.131 The regulation of cell apoptosis by MSC-EVs extends beyond these findings. Liu et al132 demonstrated that exosomes derived from MSCs downregulate TLR4 and NF-κB signaling pathways, thereby reducing ischemia-reperfusion injury-induced lung damage and apoptosis. Additionally, Gao et al130 revealed that BMSC-EVs, enriched with lncRNA-ZFAS1, alleviate apoptosis in lung microvascular endothelial cells following pulmonary ischemia-reperfusion injury. This effect is mediated through the UPF1-dependent mRNA decay of FOXD1, highlighting a novel mechanism by which MSC-derived exosomes protect against lung injury and cell death.
EVs Derived from Adipose-Derived Mesenchymal Stem Cells
AD-MSCs have emerged as a highly promising option for treating lung diseases, thanks to their robust regenerative abilities and potent anti-inflammatory and immunomodulatory properties.147,148 Sourced from adipose tissue, AD-MSCs are not only easy to obtain but also exhibit a higher proliferative capacity compared to BM-MSCs, making them a practical and efficient choice for therapeutic applications.149 Studies have demonstrated that AD-MSCs may surpass BM-MSCs in effectiveness for cell-based therapies targeting ischemia-reperfusion injury. A comparative study assessing the outcomes of intravenously administered allogenic AD-MSCs and BM-MSCs in IRI models highlighted the superior therapeutic potential of AD-MSCs.150
AD-MSCs have shown efficacy across a range of lung conditions, including acute lung injury, acute respiratory distress syndrome, chronic obstructive pulmonary disease, pulmonary fibrosis, and bronchopulmonary dysplasia.151,152 These cells mitigate lung damage by reducing inflammation, preventing fibrosis, and enhancing alveolar repair, demonstrating a particularly critical role in treating IRI-related lung injury.153 Despite their promise, challenges remain, such as inconsistencies in treatment protocols, safety concerns, and the need for clear evidence of clinical efficacy.154 Innovative approaches using AD-MSC-EVs offer a compelling alternative to direct cell therapy. These EVs deliver bioactive molecules, such as miRNAs, cytokines, and growth factors, through paracrine mechanisms, driving tissue repair, alleviating oxidative stress, and modulating immune responses.155–158 As ongoing research and clinical trials refine these therapies, AD-MSCs and their derivatives are poised to become central to the future of regenerative medicine, particularly for lung ischemia-reperfusion injury and related disorders.
EVs derived from AD-MSCs act as biomimetic nanocarriers for therapeutic immunomodulation, effectively attenuating epithelial cell activation and neutrophil infiltration in the lungs following ischemia-reperfusion injury. Cai et al128 proposed that the immunomodulatory potential of AD-MSC-derived EVs could be further enhanced by incorporating specific cargos, offering a more efficient approach to mitigating post-transplant lung ischemia-reperfusion injury. The authors emphasize the therapeutic potential of targeting miR-206 alongside the immunomodulatory properties of EVs to reduce pulmonary inflammation, edema, and injury after IR injury.128 Elevated miR-206 expression was identified as an early marker and potential biomarker for lung IR injury, with confirmation from human bronchoalveolar lavage samples of post-lung transplant patients and experimental murine models.128 Enriching EVs with miR-206 inhibitors provided significant protection against lung inflammation and injury in two murine models of ischemia-reperfusion injury. The protective effects of miR-206 inhibition were primarily driven by the modulation of alveolar type II epithelial cell activation and the reduction of CXCL1 production.128
EVs Derived from Human Umbilical Cord-Derived Mesenchymal Stem Cells
hUC-MSCs are multipotent stem cells extracted from the umbilical cord tissue, particularly from Wharton’s jelly.159,160 These cells have garnered significant interest in regenerative medicine due to their remarkable properties, including high proliferation capacity, multipotency, and low immunogenicity.161 hUC-MSCs are capable of differentiating into various cell types, such as osteoblasts, adipocytes, and chondrocytes, making them highly suitable for tissue repair and regeneration.161–163 Additionally, they possess powerful immunomodulatory abilities that can help treat autoimmune diseases, reduce inflammation, and prevent transplant rejection.164,165 Their low expression of major histocompatibility complex antigens also minimizes the risk of immune rejection.161 hUC-MSCs are easily obtained from discarded umbilical cord tissue, providing an ethically viable and abundant source of stem cells. EVs derived from hUC-MSCs are increasingly recognized for their therapeutic potential in regenerative medicine. One of the key advantages of these EVs is their ability to deliver the regenerative benefits of the parent stem cells without requiring direct cell transplantation. As a cell-free therapy, hUC-MSC-EVs offer significant benefits, including reduced immune rejection risks and simpler administration.
Studies have shown that EVs derived from hUC-MSCs can effectively mitigate lung inflammation and injury caused by ischemia-reperfusion and enhance the reconditioning of donor lungs during ex vivo lung perfusion.134 As indicated by Stone et al,134 treatment with EVs derived from hUC-MSCs significantly mitigated lung dysfunction and injury in mice, as evidenced by reduced edema, neutrophil infiltration, and myeloperoxidase levels compared to the ischemia-reperfusion group alone. Additionally, EV treatment led to a notable reduction in pro-inflammatory cytokines, including IL-17, TNF-α, CXCL1, and HMGB1, while enhancing the expression of keratinocyte growth factor, prostaglandin E2, and IL-10 in the bronchoalveolar lavage fluid after ischemia-reperfusion.134 Moreover, EVs effectively downregulated IL-17 production by invariant natural killer T cells and HMGB1 and TNF-α production by macrophages following ischemia-reperfusion.134
EV-Associated Cargos Involved in Both Conditions Demonstrate Significant Overlaps in Their Modes of Action
Despite the diverse pathophysiological processes occurring in different tissues and organs associated with these two disorders, a common feature is the role of EVs as carriers for regulating ischemia-reperfusion injury progress via various cargos mechanisms. Interestingly, certain EV-associated cargos seem to participate in both conditions by modulating respective signaling pathways. These overlapping cargos demonstrate different protective effects on disease outcomes. Giving significant attention to uncovering potential shared cargos may offer new insights into tissue remodeling processes and help identify therapeutic targets for lung and brain ischemia-reperfusion injury. Based on the aforementioned intervention studies involving EV-associated cargos, a total of 2 lncRNAs (miR-21-5p and lncRNA‑ZFAS1) have also been identified, with strong evidence suggesting their involvement in both pathophysiological conditions.166
BM-MSC-Derived ZFAS1 Plays a Role in Both Pathological Conditions
ZFAS1, a long noncoding RNA, highlights the intricate and multifaceted roles of lncRNAs in disease biology, including cancer and other pathological conditions.167 Initially characterized as downregulated in human breast cancer,168 it suggested a tumor-suppressive role in specific contexts. However, subsequent research has revealed a more complex expression pattern for ZFAS1. For instance, it has been found overexpressed in various other cell types, including BM-MSCs, where it has distinct functional implications.113,130,169 In ischemia-reperfusion injury, ZFAS1 might contribute to cellular protection, potentially by modulating stress response pathways or regulating mitochondrial function. In the context of ischemic stroke, Yang et al113 demonstrated that BMSC-secreted EVs carrying ZFAS1 played a protective role in neural injury. Specifically, EV-ZFAS1 promoted cell proliferation while simultaneously mitigating oxidative stress, apoptosis, and inflammation in BV-2 cells subjected to oxygen-glucose deprivation/reoxygenation conditions.113 However, these protective effects were abolished when miR-15a-5p was overexpressed, indicating a regulatory interaction between ZFAS1 and miR-15a-5p. Furthermore, in vivo experiments revealed that BMSC-derived EV-ZFAS1 effectively reduced oxidative stress, cerebral infarct size, and inflammatory responses in a mouse model of MCAO.113 Moreover, ZFAS1 also alleviates inflammatory responses by inhibiting the p65/IκBα inflammatory pathway. These findings underscore the therapeutic potential of BMSC-released EV-ZFAS1 in mitigating ischemic stroke-induced damage by modulating oxidative stress and inflammation. Beyond this, a study further highlighted ZFAS1 as a pivotal mediator in the mechanisms by which BM-MSCs promote neural functional recovery after stroke. Wang et al104 demonstrated that BM-MSC-derived EVs containing KLF4 alleviated infarct size, neuronal damage, and apoptosis in a mouse model of MCAO. This neuroprotective effect was achieved via the lncRNA-ZFAS1/FTO/Drp1 axis, underscoring the critical role of ZFAS1 in the therapeutic actions of BM-MSCs against stroke-induced injury. Similarly, in the context of lung ischemia-reperfusion injury, Gao et al130 demonstrated that ZFAS1 delivered by BMSC-EVs promotes the decay of FOXD1 mRNA, leading to the subsequent inactivation of Galectin-3 through direct interaction with UPF1. This mechanism effectively attenuates pulmonary ischemia-reperfusion injury. These findings collectively indicate that BMSC-derived EVs carrying ZFAS1 can facilitate improved prognosis in ischemia-reperfusion injury-related diseases, as evidenced by their protective effects in both cerebral and pulmonary ischemia-reperfusion injuries. Under these two pathological conditions, ZFAS1 exerts a protective role in both cases. However, the mechanisms underlying this protective effect are not the same. In cerebral ischemia-reperfusion injury, ZFAS1 mainly functions by inhibiting cell apoptosis, reducing infarct size, and suppressing inflammatory responses, thereby exerting its protective effect. In pulmonary ischemia-reperfusion injury, ZFAS1 primarily protects cells in the damaged areas and reduces cell apoptosis and injury by regulating the FOXD1/Galectin-3 axis. Future research needs to further explore the specific mechanisms of action of ZFAS1 under different pathological conditions in order to develop targeted therapeutic strategies.
BM-MSC-Derived miR-21-5p Plays a Role in Both Pathological Conditions
miR-21-5p is a well-established microRNA that regulates numerous biological processes, including cell growth, apoptosis, and inflammation. It is a member of the miR-21 family, which has been closely linked to the progression of various diseases, particularly ischemia-reperfusion injuries.170 In the context of ischemia-reperfusion injury, miR-21-5p plays a vital role in modulating the cellular response to ischemic stress.171 Its expression is often increased during ischemia-reperfusion injury, where it provides protective benefits by preventing cell death, mitigating oxidative damage, and regulating inflammatory responses.131 miR-21-5p exerts these effects by targeting key molecules such as PTEN and PDCD4, and activating cell survival pathways like PI3K/Akt.131,172,173 It also supports tissue recovery and angiogenesis, aiding in the repair process following ischemia-reperfusion injury. However, excessive levels of miR-21-5p can have negative consequences, such as promoting fibrosis and abnormal tissue remodeling, especially in organs like the heart.174,175 Thus, while miR-21-5p holds promise as a therapeutic target for ischemia-reperfusion injury, its complex role necessitates careful evaluation to balance its beneficial and potentially harmful effects in clinical treatments.
After ischemic stroke, the high expression of miR-21-5p in AD-MSCs and BM-MSCs provides significant protection against ischemia-reperfusion injuries in the brain and lungs. For example, based on a previous study, Hu et al106 showed that systemic administration of BM-MSC-derived EVs could reduce cerebral ischemia injury and promote angiogenesis in mice. This treatment resulted in the upregulation of proangiogenic proteins, including VEGF, VEGFR2, Ang-1, and Tie-2, as well as increased expression of miR-21-5p in the ischemic boundary regions. To further investigate the role of miR-21-5p, the researchers transfected BM-MSCs with a miR-21-5p inhibitor. The results confirmed that BM-MSC-derived EVs enhanced angiogenesis in HUVECs in vitro by transferring miR-21-5p. In addition, Liu et al demonstrated that miR-21-5p in EVs derived from AD-MSCs alleviates the inflammatory response following cerebral ischemia by modulating the PIK3R1/PI3K/AKT signaling pathway.173 Meanwhile, Li et al176 investigated the impact of AD-MSC administration on blood-brain barrier integrity, apoptosis, inflammation, and the activity of the miR-21-3p/MAT2B axis in ischemic stroke. Additionally, it provided neuroprotection after ischemic stroke by promoting the polarization of M2 microglia. miR-21-5p derived from AD-MSCs improved BBB condition, inhibited apoptosis, and suppressed inflammation.176 In the context of lung ischemia-reperfusion injury, as shown by Li et al,131 murine BM-MSC were subjected to hypoxia or treated with a miR-21-5p antagomir to either enhance or suppress miR-21-5p levels in MSC-derive EVs. The results demonstrated that MSC-EVs significantly alleviated lung ischemia-reperfusion injury in mice, and this protective effect was closely linked to miR-21-5p. Further evidence has confirmed that MSC-Exo-miR-21-5p can target PTEN and PDCD4, thereby inhibiting cell apoptosis and alleviating ischemia-reperfusion injury. In primary cultures of murine pulmonary endothelial cells, both MSC-Exo and miR-21-5p agomir effectively reduced apoptosis caused by oxidative stress. Additionally, they partially mitigated the pro-inflammatory “M1” polarization of alveolar macrophages triggered by hypoxia-reoxygenation procedure, highlighting their anti-inflammatory and cytoprotective properties.131
Conclusion
In ischemic stroke, MSC-EVs can inhibit inflammatory responses, oxidative stress, and cellular autophagy and pyroptosis by releasing extracellular vesicles, while also promoting angiogenesis in damaged tissues. Similarly, in pulmonary ischemia-reperfusion injury, MSC-EVs can suppress inflammatory responses, autophagy, and pyroptosis. Thus, MSC-EVs have demonstrated significant protective effects in both cerebral and pulmonary ischemia-reperfusion injuries. Through comparison, analysis, and summary, this review identifies two non-coding RNAs derived from MSC-EVs (miR-21-5p and lncRNA-ZFAS1), indicating their involvement in both pathological conditions and their similar mechanisms of action. This finding offers new insights and directions for research on ischemia-reperfusion injury and the lung-brain axis, providing a foundation for further exploration of their specific mechanisms.
Abbreviations
MSCs-EVs, mesenchymal stem cell-derived extracellular vesicles; EVs, extracellular vesicles; MVs, microvesicles; ROS, reactive oxygen species; MSCs, mesenchymal stem cells; PDGF-D, platelet-derived growth factor D; CNS, Central nervous system; UC-MSCs-EVs, umbilical cord mesenchymal stem cell-derived extracellular vesicles; BMSC-EVs, Bone Marrow Mesenchymal Stem Cell-derived Exosomes; MCAO, Middle Cerebral Artery Occlusion; hUCMSCs-EVs, human umbilical cord mesenchymal stem cells exosomes; OGD, Oxygen and Glucose Deprivation; AIM2, Absent in Melanoma 2; hiPS-MSC-EVs, Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cell Extracellular Vesicles; ADSCs-EVs, adipose-derived stem cells; BMECs, brain microvascular endothelial cells; MDA, malondialdehyde; SOD, superoxide dismutase; TNFR1, tumor necrosis factor receptor 1; NQO1, quinone oxidoreductase 1; HO-1, oxygenase-1; OM-MSCs, Olfactory Mucosa Mesenchymal Stem Cells; hNSCs human neural stem cells; Cav-1, Caveolin-1; PRISMA, Reporting Items for Systematic Reviews and Meta-Analyses.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Funding
There is no funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pefanis A, Ierino FL, Murphy JM, Cowan PJ. Regulated necrosis in kidney ischemia-reperfusion injury. Kidney Int. 2019;96(2):291–301. PubMed PMID: 31005270. doi:10.1016/j.kint.2019.02.009
2. Ibáñez B, Heusch G, Ovize M, Van de Werf F. Evolving therapies for myocardial ischemia/reperfusion injury. J Am Coll Cardiol. 2015;65(14):1454–1471. PubMed PMID: 25857912. doi:10.1016/j.jacc.2015.02.032
3. Hermann DM, Xin W, Bähr M, Giebel B, Doeppner TR. Emerging roles of extracellular vesicle-associated non-coding RNAs in hypoxia: insights from cancer, myocardial infarction and ischemic stroke. Theranostics. 2022;12(13):5776–5802. PubMed PMID: 35966580; PubMed Central PMCID: PMCPMC9373807. doi:10.7150/thno.73931
4. Xin W, Qin Y, Lei P, Zhang J, Yang X, Wang Z. From cerebral ischemia towards myocardial, renal, and hepatic ischemia: exosomal miRNAs as a general concept of intercellular communication in ischemia-reperfusion injury. Mol Ther Nucleic Acids. 2022;29:900–922. PubMed PMID: 36159596; PubMed Central PMCID: PMCPMC9464648. doi:10.1016/j.omtn.2022.08.032
5. Chen-Yoshikawa TF. Ischemia-reperfusion injury in lung transplantation. Cells. 2021;10(6). PubMed PMID: 34071255; PubMed Central PMCID: PMCPMC8228304. doi:10.3390/cells10061333
6. Anaya-Prado R, Toledo-Pereyra LH, Lentsch AB, Ward PA. Ischemia/reperfusion injury. J Surg Res. 2002;105(2):248–258. PubMed PMID: 12125704. doi:10.1006/jsre.2002.6385
7. Werns SW, Lucchesi BR. Free radicals and ischemic tissue injury. Trends Pharmacol Sci. 1990;11(4):161–166. PubMed PMID: 2185587. doi:10.1016/0165-6147(90)90068-j
8. Bugger H, Pfeil K. Mitochondrial ROS in myocardial ischemia reperfusion and remodeling. Biochim Biophys Acta Mol Basis Dis. 2020;1866(7):165768. PubMed PMID: 32173461. doi:10.1016/j.bbadis.2020.165768
9. Zhang M, Liu Q, Meng H, et al. Ischemia-reperfusion injury: molecular mechanisms and therapeutic targets. Signal Transduct Target Ther. 2024;9(1):12. PubMed PMID: 38185705; PubMed Central PMCID: PMCPMC10772178. doi:10.1038/s41392-023-01688-x
10. Kalogeris T, Baines CP, Krenz M, Korthuis RJ. Cell biology of ischemia/reperfusion injury. Int Rev Cell Mol Biol. 2012;298:229–317. PubMed PMID: 22878108; PubMed Central PMCID: PMCPMC3904795. doi:10.1016/b978-0-12-394309-5.00006-7
11. Zhang Q, Jia M, Wang Y, Wang Q, Wu J. Cell death mechanisms in cerebral ischemia-reperfusion injury. Neurochem Res. 2022;47(12):3525–3542. PubMed PMID: 35976487. doi:10.1007/s11064-022-03697-8
12. Bister N, Pistono C, Huremagic B, Jolkkonen J, Giugno R, Malm T. Hypoxia and extracellular vesicles: a review on methods, vesicular cargo and functions. J Extracell Vesicles. 2020;10(1):e12002. PubMed PMID: 33304471; PubMed Central PMCID: PMCPMC7710128. doi:10.1002/jev2.12002
13. Choudhry H, Harris AL. Advances in hypoxia-inducible factor biology. Cell Metab. 2018;27(2):281–298. PubMed PMID: 29129785. doi:10.1016/j.cmet.2017.10.005
14. Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023;33(8):667–681. PubMed PMID: 36737375; PubMed Central PMCID: PMCPMC10363204. doi:10.1016/j.tcb.2023.01.002
15. Sidhom K, Obi PO, Saleem A. A review of exosomal isolation methods: is size exclusion chromatography the best option? Int J Mol Sci. 2020;21(18). PubMed PMID: 32899828; PubMed Central PMCID: PMCPMC7556044. doi:10.3390/ijms21186466
16. Hoang DM, Pham PT, Bach TQ, et al. Stem cell-based therapy for human diseases. Signal Transduct Target Ther. 2022;7(1):272. PubMed PMID: 35933430; PubMed Central PMCID: PMCPMC9357075. doi:10.1038/s41392-022-01134-4
17. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9(1):27. PubMed PMID: 38311623; PubMed Central PMCID: PMCPMC10838959. doi:10.1038/s41392-024-01735-1
18. Pan Y, Liu Y, Wei W, Yang X, Wang Z, Xin W. Extracellular vesicles as delivery shippers for noncoding RNA-based modulation of angiogenesis: insights from ischemic stroke and cancer. Small. 2023;19(17):e2205739. PubMed PMID: 36592424. doi:10.1002/smll.202205739
19. Xu H, Wang B, Li A, Wen J, Su H, Qin D. Mesenchymal stem cells-based cell-free therapy targeting neuroinflammation. Aging Dis. 2024;15(3):965–976. PubMed PMID: 38722791; PubMed Central PMCID: PMCPMC11081161. doi:10.14336/ad.2023.0904
20. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. PubMed PMID: 29339798. doi:10.1038/nrm.2017.125
21. Waldenström A, Gennebäck N, Hellman U, Ronquist G. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS One. 2012;7(4):e34653. PubMed PMID: 22506041; PubMed Central PMCID: PMCPMC3323564. doi:10.1371/journal.pone.0034653
22. Abels ER, Breakefield XO. Introduction to extracellular vesicles: biogenesis, RNA cargo selection, content, release, and uptake. Cell Mol neurobiol. 2016;36(3):301–312. PubMed PMID: 27053351; PubMed Central PMCID: PMCPMC5546313. doi:10.1007/s10571-016-0366-z
23. Li C, Xu X. Biological functions and clinical applications of exosomal non-coding RNAs in hepatocellular carcinoma. Cell Mol Life Sci. 2019;76(21):4203–4219. PubMed PMID: 31300868; PubMed Central PMCID: PMCPMC11105530. doi:10.1007/s00018-019-03215-0
24. Han C, Yang J, Sun J, Qin G. Extracellular vesicles in cardiovascular disease: biological functions and therapeutic implications. Pharmacol Ther. 2022;233:108025. PubMed PMID: 34687770; PubMed Central PMCID: PMCPMC9018895. doi:10.1016/j.pharmthera.2021.108025
25. Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell. 2016;30(6):836–848. PubMed PMID: 27960084; PubMed Central PMCID: PMCPMC5157696. doi:10.1016/j.ccell.2016.10.009
26. Hu S, Hu Y, Yan W. Extracellular vesicle-mediated interorgan communication in metabolic diseases. Trends Endocrinol Metab. 2023;34(9):571–582. PubMed PMID: 37394346. doi:10.1016/j.tem.2023.06.002
27. Wu MY, Yiang GT, Liao WT, et al. Current mechanistic concepts in ischemia and reperfusion injury. Cell Physiol Biochem. 2018;46(4):1650–1667. PubMed PMID: 29694958. doi:10.1159/000489241
28. de Perrot M, Liu M, Waddell TK, Keshavjee S. Ischemia-reperfusion-induced lung injury. Am J Respir Crit Care Med. 2003;167(4):490–511. PubMed PMID: 12588712. doi:10.1164/rccm.200207-670SO
29. Bajinka O, Simbilyabo L, Tan Y, Jabang J, Saleem SA. Lung-brain axis. Crit Rev Microbiol. 2022;48(3):257–269. PubMed PMID: 34348558. doi:10.1080/1040841X.2021.1960483
30. Kim JT, Song K, Han SW, et al. Modeling of the brain-lung axis using organoids in traumatic brain injury: an updated review. Cell Biosci. 2024;14(1):83. PubMed PMID: 38909262; PubMed Central PMCID: PMCPMC11193205. doi:10.1186/s13578-024-01252-2
31. Mai N, Miller-Rhodes K, Knowlden S, Halterman MW. The post-cardiac arrest syndrome: a case for lung-brain coupling and opportunities for neuroprotection. J Cereb Blood Flow Metab. 2019;39(6):939–958. PubMed PMID: 30866740; PubMed Central PMCID: PMCPMC6547189. doi:10.1177/0271678X19835552
32. Dong M, Li M, Guo Y, Xu S, An K. U-shaped association between low-density lipoprotein cholesterol levels and risk of futile reperfusion mediated by stroke-associated pneumonia in acute ischemic stroke after endovascular thrombectomy. Clin Neurol Neurosurg. 2024;243:108399. PubMed PMID: 38901376. doi:10.1016/j.clineuro.2024.108399
33. Stevens RD, Puybasset L. The brain-lung-brain axis. Intensive Care Med. 2011;37(7):1054–1056. PubMed PMID: 21544691. doi:10.1007/s00134-011-2233-1
34. Hosang L, Canals RC, van der Flier FJ, et al. The lung microbiome regulates brain autoimmunity. Nature. 2022;603(7899):138–144. PubMed PMID: 35197636. doi:10.1038/s41586-022-04427-4
35. Xu D, Zhao M, Liu G, et al. The vagus nerve-dependent lung-brain axis mediates brain demyelination following acute lung injury. Brain Behav Immun Health. 2025;44:100966. PubMed PMID: 40028232; PubMed Central PMCID: PMCPMC11871466. doi:10.1016/j.bbih.2025.100966
36. Witzenrath M, Kuebler WM. The lung-brain axis in ventilator-induced brain injury: enter IL-6. Am J Respir Cell Mol Biol. 2021;65(4):339–340. PubMed PMID: 34153209; PubMed Central PMCID: PMCPMC8525199. doi:10.1165/rcmb.2021-0233ED
37. Villalba N, Ma Y, Gahan SA, et al. Lung infection by Pseudomonas aeruginosa induces neuroinflammation and blood-brain barrier dysfunction in mice. J Neuroinflammation. 2023;20(1):127. PubMed PMID: 37245027; PubMed Central PMCID: PMCPMC10223932. doi:10.1186/s12974-023-02817-7
38. He M, Liu Y, Guan Z, Li C, Zhang Z. Neuroimaging insights into lung disease-related brain changes: from structure to function. Front Aging Neurosci. 2025;17:1550319. PubMed PMID: 40051465; PubMed Central PMCID: PMCPMC11882867. doi:10.3389/fnagi.2025.1550319
39. Sharma S, Culebras A. Sleep apnoea and stroke. Stroke Vasc Neurol. 2016;1(4):185–191. PubMed PMID: 28959482; PubMed Central PMCID: PMCPMC5435217. doi:10.1136/svn-2016-000038
40. Morgan AD, Sharma C, Rothnie KJ, Potts J, Smeeth L, Quint JK. Chronic obstructive pulmonary disease and the risk of stroke. Ann Am Thorac Soc. 2017;14(5):754–765. PubMed PMID: 28459623; PubMed Central PMCID: PMCPMC5427743. doi:10.1513/AnnalsATS.201611-932SR
41. Gong P, Jia HY, Li R, et al. Downregulation of Nogo-B ameliorates cerebral ischemia/reperfusion injury in mice through regulating microglia polarization via TLR4/NF-kappaB pathway. Neurochem Int. 2023;167:105553. PubMed PMID: 37230196. doi:10.1016/j.neuint.2023.105553
42. Bernard SA, Gray TW, Buist MD, et al. Treatment of comatose survivors of out-of-hospital cardiac arrest with induced hypothermia. N Engl J Med. 2002;346(8):557–563. PubMed PMID: 11856794. doi:10.1056/NEJMoa003289
43. Hoesch RE, Koenig MA, Geocadin RG. Coma after global ischemic brain injury: pathophysiology and emerging therapies. Crit Care Clin. 2008;24(1):25–44,vii–viii. PubMed PMID: 18241777. doi:10.1016/j.ccc.2007.11.003
44. Yang K, Zeng L, Ge A, et al. A systematic review of the research progress of non-coding RNA in neuroinflammation and immune regulation in cerebral infarction/ischemia-reperfusion injury. Front Immunol. 2022;13:930171. PubMed PMID: 36275741; PubMed Central PMCID: PMCPMC9585453. doi:10.3389/fimmu.2022.930171
45. Lee K, Rincon F. Pulmonary complications in patients with severe brain injury. Crit Care Res Pract. 2012;2012:207247. PubMed PMID: 23133746; PubMed Central PMCID: PMCPMC3485871. doi:10.1155/2012/207247
46. Rees J. Guillain-Barré syndrome. Clinical manifestations and directions for treatment. Drugs. 1995;49(6):912–920. PubMed PMID: 7641605. doi:10.2165/00003495-199549060-00005
47. Labeit B, Michou E, Hamdy S, et al. The assessment of dysphagia after stroke: state of the art and future directions. Lancet Neurol. 2023;22(9):858–870. PubMed PMID: 37596008. doi:10.1016/S1474-4422(23)00153-9
48. Jurcau A, Simion A. Neuroinflammation in cerebral ischemia and ischemia/reperfusion injuries: from pathophysiology to therapeutic strategies. Int J Mol Sci. 2021;23(1). PubMed PMID: 35008440; PubMed Central PMCID: PMCPMC8744548. doi:10.3390/ijms23010014
49. Chen W, Li D. Reactive oxygen species (ROS)-responsive nanomedicine for solving ischemia-reperfusion injury. Front Chem. 2020;8:732. PubMed PMID: 32974285; PubMed Central PMCID: PMCPMC7472733. doi:10.3389/fchem.2020.00732
50. Westendorp WF, Dames C, Nederkoorn PJ, Meisel A. Immunodepression, infections, and functional outcome in ischemic stroke. Stroke. 2022;53(5):1438–1448. PubMed PMID: 35341322. doi:10.1161/STROKEAHA.122.038867
51. Labeit B, Michou E, Trapl-Grundschober M, et al. Dysphagia after stroke: research advances in treatment interventions. Lancet Neurol. 2024;23(4):418–428. PubMed PMID: 38508837. doi:10.1016/S1474-4422(24)00053-X
52. Westendorp WF, Nederkoorn PJ, Vermeij JD, Dijkgraaf MG, van de Beek D. Post-stroke infection: a systematic review and meta-analysis. BMC Neurol. 2011;11:110. PubMed PMID: 21933425; PubMed Central PMCID: PMCPMC3185266. doi:10.1186/1471-2377-11-110
53. Guo X, Liu R, Jia M, Wang Q, Wu J. Ischemia reperfusion injury induced blood brain barrier dysfunction and the involved molecular mechanism. Neurochem Res. 2023;48(8):2320–2334. PubMed PMID: 37017889. doi:10.1007/s11064-023-03923-x
54. Chacón-Aponte AA, Durán-Vargas ÉA, Arévalo-Carrillo JA, et al. Brain-lung interaction: a vicious cycle in traumatic brain injury. Acute Crit Care. 2022;37(1):35–44. PubMed PMID: 35172526; PubMed Central PMCID: PMCPMC8918716. doi:10.4266/acc.2021.01193
55. Mai N, Prifti L, Rininger A, Bazarian H, Halterman MW. Endotoxemia induces lung-brain coupling and multi-organ injury following cerebral ischemia-reperfusion. Exp Neurol. 2017;297:82–91. PubMed PMID: 28757259; PubMed Central PMCID: PMCPMC5612886. doi:10.1016/j.expneurol.2017.07.016
56. Pak O, Sydykov A, Kosanovic D, et al. Lung ischaemia-reperfusion injury: the role of reactive oxygen species. Adv Exp Med Biol. 2017;967:195–225. PubMed PMID: 29047088. doi:10.1007/978-3-319-63245-2_12
57. Capuzzimati M, Hough O, Liu M. Cell death and ischemia-reperfusion injury in lung transplantation. J Heart Lung Transplant. 2022;41(8):1003–1013. PubMed PMID: 35710485. doi:10.1016/j.healun.2022.05.013
58. Ding DC, Chang YH, Shyu WC, Lin SZ. Human umbilical cord mesenchymal stem cells: a new era for stem cell therapy. Cell Transplant. 2015;24(3):339–347. PubMed PMID: 25622293. doi:10.3727/096368915x686841
59. Doeppner TR, Hermann DM. Mesenchymal stem cells in the treatment of ischemic stroke: progress and possibilities. Stem Cells and Cloning: Advances and Applications. 2010;3:157–163. PubMed PMID: 24198521; PubMed Central PMCID: PMCPMC3781740. doi:10.2147/sccaa.S7820
60. Zou J, Yang W, Cui W, et al. Therapeutic potential and mechanisms of mesenchymal stem cell-derived exosomes as bioactive materials in tendon-bone healing. J Nanobiotechnology. 2023;21(1):14. PubMed PMID: 36642728; PubMed Central PMCID: PMCPMC9841717. doi:10.1186/s12951-023-01778-6
61. Maria Ferri AL, Bersano A, Lisini D, Boncoraglio G, Frigerio S, Parati E. Mesenchymal stem cells for ischemic stroke: progress and possibilities. Curr. Med. Chem. 2016;23(16):1598–1608. PubMed PMID: 26898654. doi:10.2174/0929867323666160222113702
62. Yang Z, Zhu L, Li F, Wang J, Wan H, Pan Y. Bone marrow stromal cells as a therapeutic treatment for ischemic stroke. Neuroscience Bulletin. 2014;30(3):524–534. PubMed PMID: 24817388; PubMed Central PMCID: PMCPMC5562610. doi:10.1007/s12264-013-1431-y
63. Wakabayashi K, Nagai A, Sheikh AM, et al. Transplantation of human mesenchymal stem cells promotes functional improvement and increased expression of neurotrophic factors in a rat focal cerebral ischemia model. J Neurosci Res. 2010;88(5):1017–1025. PubMed PMID: 19885863. doi:10.1002/jnr.22279
64. Zheng W, Honmou O, Miyata K, et al. Therapeutic benefits of human mesenchymal stem cells derived from bone marrow after global cerebral ischemia. Brain Res. 2010;1310:8–16. PubMed PMID: 19913518. doi:10.1016/j.brainres.2009.11.012
65. Liu YY, Chiang CH, Hung SC, et al. Hypoxia-preconditioned mesenchymal stem cells ameliorate ischemia/reperfusion-induced lung injury. PLoS One. 2017;12(11):e0187637. PubMed PMID: 29117205; PubMed Central PMCID: PMCPMC5678873. doi:10.1371/journal.pone.0187637
66. Nakajima D, Watanabe Y, Ohsumi A, et al. Mesenchymal stromal cell therapy during ex vivo lung perfusion ameliorates ischemia-reperfusion injury in lung transplantation. J Heart Lung Transpl. 2019;38(11):1214–1223. PubMed PMID: 31474491. doi:10.1016/j.healun.2019.07.006
67. Sun H, Pratt RE, Hodgkinson CP, Dzau VJ. Sequential paracrine mechanisms are necessary for the therapeutic benefits of stem cell therapy. Am J Physiol Cell Physiol. 2020;319(6):C1141–c50. PubMed PMID: 33026832. doi:10.1152/ajpcell.00516.2019
68. Lui PPY. Mesenchymal stem cell-derived extracellular vesicles for the promotion of tendon repair - an update of literature. Stem Cell Rev Rep. 2021;17(2):379–389. PubMed PMID: 32785869. doi:10.1007/s12015-020-10023-8
69. Lai P, Weng J, Guo L, Chen X, Du X. Novel insights into MSC-EVs therapy for immune diseases. Biomark Res. 2019;7:6. PubMed PMID: 30923617; PubMed Central PMCID: PMCPMC6423844. doi:10.1186/s40364-019-0156-0
70. Mizenko RR, Feaver M, Bozkurt BT, et al. A critical systematic review of extracellular vesicle clinical trials. J Extracell Vesicles. 2024;13(10):e12510. PubMed PMID: 39330928; PubMed Central PMCID: PMCPMC11428870. doi:10.1002/jev2.12510
71. Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol. 1983;97(2):329–339. PubMed PMID: 6309857; PubMed Central PMCID: PMCPMC2112509. doi:10.1083/jcb.97.2.329
72. Nicodemou A, Bernátová S, Čeháková M, Danišovič Ľ. Emerging roles of mesenchymal stem/stromal-cell-derived extracellular vesicles in cancer therapy. Pharmaceutics. 2023;15(5). PubMed PMID: 37242693; PubMed Central PMCID: PMCPMC10221697. doi:10.3390/pharmaceutics15051453
73. Qiu G, Zheng G, Ge M, et al. Functional proteins of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res Ther. 2019;10(1):359. PubMed PMID: 31779700; PubMed Central PMCID: PMCPMC6883709. doi:10.1186/s13287-019-1484-6
74. Zhao R, Wang L, Wang T, Xian P, Wang H, Long Q. Inhalation of MSC-EVs is a noninvasive strategy for ameliorating acute lung injury. J Control Release. 2022;345:214–230. PubMed PMID: 35307508. doi:10.1016/j.jconrel.2022.03.025
75. Harrell CR, Jovicic N, Djonov V, Arsenijevic N, Volarevic V. Mesenchymal stem cell-derived exosomes and other extracellular vesicles as new remedies in the therapy of inflammatory diseases. Cells. 2019;8(12). PubMed PMID: 31835680; PubMed Central PMCID: PMCPMC6952783. doi:10.3390/cells8121605
76. Tan N, Xin W, Huang M, Mao Y. Mesenchymal stem cell therapy for ischemic stroke: novel insight into the crosstalk with immune cells. Front Neurol. 2022;13:1048113. PubMed PMID: 36425795; PubMed Central PMCID: PMCPMC9679024. doi:10.3389/fneur.2022.1048113
77. Ma J, Zhao Y, Sun L, et al. Exosomes derived from Akt-modified human umbilical cord mesenchymal stem cells improve cardiac regeneration and promote angiogenesis via activating platelet-derived growth factor D. Stem Cells Transl Med. 2017;6(1):51–59. PubMed PMID: 28170176; PubMed Central PMCID: PMCPMC5442756. doi:10.5966/sctm.2016-0038
78. Zhou C, Zhou F, He Y, Liu Y, Cao Y. Exosomes in cerebral ischemia-reperfusion injury: current perspectives and future challenges. Brain Sci. 2022;12(12). PubMed PMID: 36552117; PubMed Central PMCID: PMCPMC9776031. doi:10.3390/brainsci12121657
79. Qiu G, Zheng G, Ge M, et al. Mesenchymal stem cell-derived extracellular vesicles affect disease outcomes via transfer of microRNAs. Stem Cell Res Ther. 2018;9(1):320. PubMed PMID: 30463593; PubMed Central PMCID: PMCPMC6249826. doi:10.1186/s13287-018-1069-9
80. Woodburn SC, Bollinger JL, Wohleb ES. The semantics of microglia activation: neuroinflammation, homeostasis, and stress. J Neuroinflammation. 2021;18(1):258. doi:10.1186/s12974-021-02309-6
81. Ishizaka S, Horie N, Satoh K, Fukuda Y, Nishida N, Nagata I. Intra-arterial cell transplantation provides timing-dependent cell distribution and functional recovery after stroke. Stroke. 2013;44(3):720–726. PubMed PMID: 23362081. doi:10.1161/strokeaha.112.677328
82. Yamaguchi S, Horie N, Satoh K, et al. Age of donor of human mesenchymal stem cells affects structural and functional recovery after cell therapy following ischaemic stroke. J Cereb Blood Flow Metab. 2018;38(7):1199–1212. PubMed PMID: 28914133; PubMed Central PMCID: PMCPMC6434451. doi:10.1177/0271678x17731964
83. Wang LQ, Lin ZZ, Zhang HX, et al. Timing and dose regimens of marrow mesenchymal stem cell transplantation affect the outcomes and neuroinflammatory response after ischemic stroke. CNS Neurosci Ther. 2014;20(4):317–326. PubMed PMID: 24393245; PubMed Central PMCID: PMCPMC6493061. doi:10.1111/cns.12216
84. Nakajima M, Nito C, Sowa K, et al. Mesenchymal stem cells overexpressing interleukin-10 promote neuroprotection in experimental acute ischemic stroke. Mol Ther Methods Clin Dev. 2017;6:102–111. PubMed PMID: 28725658; PubMed Central PMCID: PMCPMC5502709. doi:10.1016/j.omtm.2017.06.005
85. Cui L, Luo W, Jiang W, et al. Human umbilical cord mesenchymal stem cell-derived exosomes promote neurological function recovery in rat after traumatic brain injury by inhibiting the activation of microglia and astrocyte. Regener Ther. 2022;21:282–287. PubMed PMID: 36092501; PubMed Central PMCID: PMCPMC9440059. doi:10.1016/j.reth.2022.07.005
86. Liu X, Zhang M, Liu H, et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cerebral ischemia-reperfusion injury-induced neuroinflammation and pyroptosis by modulating microglia M1/M2 phenotypes. Exp Neurol. 2021;341:113700. PubMed PMID: 33741350. doi:10.1016/j.expneurol.2021.113700
87. Dong C, Chen M, Cai B, Zhang C, Xiao G, Luo W. Mesenchymal stem cell-derived exosomes improved cerebral infarction via transferring miR-23a-3p to activate microglia. Neuromol Med. 2022;24(3):290–298. PubMed PMID: 35001328. doi:10.1007/s12017-021-08686-8
88. Zhang Z, Zou X, Zhang R, et al. Human umbilical cord mesenchymal stem cell-derived exosomal miR-146a-5p reduces microglial-mediated neuroinflammation via suppression of the IRAK1/TRAF6 signaling pathway after ischemic stroke. Aging. 2021;13(2):3060–3079. PubMed PMID: 33479185; PubMed Central PMCID: PMCPMC7880318. doi:10.18632/aging.202466
89. Che J, Wang H, Dong J, et al. Human umbilical cord mesenchymal stem cell-derived exosomes attenuate neuroinflammation and oxidative stress through the NRF2/NF-κB/NLRP3 pathway. CNS Neurosci Ther. 2024;30(3):e14454. PubMed PMID: 37697971; PubMed Central PMCID: PMCPMC10916441. doi:10.1111/cns.14454
90. Xian P, Hei Y, Wang R, et al. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice. Theranostics. 2019;9(20):5956–5975. PubMed PMID: 31534531; PubMed Central PMCID: PMCPMC6735367. doi:10.7150/thno.33872
91. Deng Y, Chen D, Gao F, et al. Exosomes derived from microRNA-138-5p-overexpressing bone marrow-derived mesenchymal stem cells confer neuroprotection to astrocytes following ischemic stroke via inhibition of LCN2. J Biol Eng. 2019;13:71. PubMed PMID: 31485266; PubMed Central PMCID: PMCPMC6714399. doi:10.1186/s13036-019-0193-0
92. Fricker M, Tolkovsky AM, Borutaite V, Coleman M, Brown GC. Neuronal cell death. Physiol Rev. 2018;98(2):813–880. PubMed PMID: 29488822; PubMed Central PMCID: PMCPMC5966715. doi:10.1152/physrev.00011.2017
93. Ye YC, Chang ZH, Wang P, et al. Infarct-preconditioning exosomes of umbilical cord mesenchymal stem cells promoted vascular remodeling and neurological recovery after stroke in rats. Stem Cell Res Ther. 2022;13(1):378. PubMed PMID: 35902882; PubMed Central PMCID: PMCPMC9330700. doi:10.1186/s13287-022-03083-9
94. Ikeda T, Kawabori M, Zheng Y, et al. Intranasal administration of mesenchymal stem cell-derived exosome alleviates hypoxic-ischemic brain injury. Pharmaceutics. 2024;16(4). PubMed PMID: 38675108; PubMed Central PMCID: PMCPMC11053690. doi:10.3390/pharmaceutics16040446
95. Kang X, Jiang L, Chen X, et al. Exosomes derived from hypoxic bone marrow mesenchymal stem cells rescue OGD-induced injury in neural cells by suppressing NLRP3 inflammasome-mediated pyroptosis. Exp Cell Res. 2021;405(1):112635. PubMed PMID: 34051241. doi:10.1016/j.yexcr.2021.112635
96. Liu Z, Li X, Ye Z, Lin H. Neuroprotective effect of exosomes derived from bone marrow mesenchymal stem cells via activating TGR5 and suppressing apoptosis. Biochem Biophys Res Commun. 2022;593:13–19. PubMed PMID: 35051777. doi:10.1016/j.bbrc.2022.01.039
97. Xiao Y, Geng F, Wang G, Li X, Zhu J, Zhu W. Bone marrow-derived mesenchymal stem cells-derived exosomes prevent oligodendrocyte apoptosis through exosomal miR-134 by targeting caspase-8. J Cell Biochem. 2019;120(2):2109–2118. PubMed PMID: 30191592. doi:10.1002/jcb.27519
98. Wang J, Sun H, Guo R, et al. Exosomal miR-23b-3p from bone mesenchymal stem cells alleviates experimental autoimmune encephalomyelitis by inhibiting microglial pyroptosis. Exp Neurol. 2023;363:114374. PubMed PMID: 36907352. doi:10.1016/j.expneurol.2023.114374
99. Wang Y, Chen H, Fan X, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-193b-5p reduces pyroptosis after ischemic stroke by targeting AIM2. J Stroke Cerebrovasc Dis. 2023;32(8):107235. PubMed PMID: 37393689. doi:10.1016/j.jstrokecerebrovasdis.2023.107235
100. Zhang Y, Liu J, Su M, Wang X, Xie C. Exosomal microRNA-22-3p alleviates cerebral ischemic injury by modulating KDM6B/BMP2/BMF axis. Stem Cell Res Ther. 2021;12(1):111. PubMed PMID: 33546766; PubMed Central PMCID: PMCPMC7863295. doi:10.1186/s13287-020-02091-x
101. Zhou H, Zhou J, Teng H, Yang H, Qiu J, Li X. MiR-145 enriched exosomes derived from bone marrow-derived mesenchymal stem cells protects against cerebral ischemia-reperfusion injury through downregulation of FOXO1. Biochem Biophys Res Commun. 2022;632:92–99. PubMed PMID: 36206599. doi:10.1016/j.bbrc.2022.09.089
102. Cheng C, Chen X, Wang Y, et al. MSCs‑derived exosomes attenuate ischemia-reperfusion brain injury and inhibit microglia apoptosis might via exosomal miR-26a-5p mediated suppression of CDK6. Mol Med. 2021;27(1):67. PubMed PMID: 34215174; PubMed Central PMCID: PMCPMC8254277. doi:10.1186/s10020-021-00324-0
103. Jinglei H, Zengxiu Z, Dongxia X, Huiping L. The efficacy of budesonide in combination with azithromycin in the treatment of pediatric mycoplasma pneumonia and its impact on inflammatory marker levels. Chinese J Clin Ration Drug Use. 2022;15(07):129–131.
104. Wang QS, Xiao RJ, Peng J, Yu ZT, Fu JQ, Xia Y. Bone marrow mesenchymal stem cell-derived exosomal KLF4 alleviated ischemic stroke through inhibiting N6-methyladenosine modification level of Drp1 by targeting lncRNA-ZFAS1. Mol Neurobiol. 2023;60(7):3945–3962. PubMed PMID: 37002530. doi:10.1007/s12035-023-03301-2
105. Huang X, Ding J, Li Y, et al. Exosomes derived from PEDF modified adipose-derived mesenchymal stem cells ameliorate cerebral ischemia-reperfusion injury by regulation of autophagy and apoptosis. Exp Cell Res. 2018;371(1):269–277. PubMed PMID: 30142325. doi:10.1016/j.yexcr.2018.08.021
106. Hu H, Hu X, Li L, et al. Exosomes derived from bone marrow mesenchymal stem cells promote angiogenesis in ischemic stroke mice via upregulation of MiR-21-5p. Biomolecules. 2022;12(7). PubMed PMID: 35883438; PubMed Central PMCID: PMCPMC9313463. doi:10.3390/biom12070883
107. Feng J, He W, Xia J, et al. Human umbilical cord mesenchymal stem cells-derived exosomal circDLGAP4 promotes angiogenesis after cerebral ischemia-reperfusion injury by regulating miR-320/KLF5 axis. FASEB j. 2023;37(3):e22733. PubMed PMID: 36723877. doi:10.1096/fj.202201488R
108. Lu G, Su X, Wang L, et al. Neuroprotective effects of human-induced pluripotent stem cell-derived mesenchymal stem cell extracellular vesicles in ischemic stroke models. Biomedicines. 2023;11(9). PubMed PMID: 37760991; PubMed Central PMCID: PMCPMC10525838. doi:10.3390/biomedicines11092550
109. Bao H, Mao S, Hu X, et al. Exosomal miR-486 derived from bone marrow mesenchymal stem cells promotes angiogenesis following cerebral ischemic injury by regulating the PTEN/Akt pathway. Sci Rep. 2024;14(1):18086. PubMed PMID: 39103424; PubMed Central PMCID: PMCPMC11300871. doi:10.1038/s41598-024-69172-2
110. Yang Y, Cai Y, Zhang Y, Liu J, Xu Z. Exosomes secreted by adipose-derived stem cells contribute to angiogenesis of brain microvascular endothelial cells following oxygen-glucose deprivation in vitro through MicroRNA-181b/TRPM7 axis. Journal of Molecular Neuroscience: MN. 2018;65(1):74–83. PubMed PMID: 29705934. doi:10.1007/s12031-018-1071-9
111. Chen KH, Chen CH, Wallace CG, et al. Intravenous administration of xenogenic adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes markedly reduced brain infarct volume and preserved neurological function in rat after acute ischemic stroke. Oncotarget. 2016;7(46):74537–74556. PubMed PMID: 27793019; PubMed Central PMCID: PMCPMC5342685. doi:10.18632/oncotarget.12902
112. Liu G, Wang D, Jia J, et al. Neuroprotection of human umbilical cord-derived mesenchymal stem cells (hUC-MSCs) in alleviating ischemic stroke-induced brain injury by regulating inflammation and oxidative stress. Neurochem Res. 2024;49(10):2871–2887. PubMed PMID: 39026086. doi:10.1007/s11064-024-04212-x
113. Yang H, Chen J. Bone marrow mesenchymal stem cell-derived exosomes carrying long noncoding RNA ZFAS1 alleviate oxidative stress and inflammation in ischemic stroke by inhibiting microRNA-15a-5p. Metab Brain Dis. 2022;37(7):2545–2557. PubMed PMID: 35907132. doi:10.1007/s11011-022-00997-4
114. Leu S, Lin YC, Yuen CM, et al. Adipose-derived mesenchymal stem cells markedly attenuate brain infarct size and improve neurological function in rats. J Transl Med. 2010;8:63. PubMed PMID: 20584315; PubMed Central PMCID: PMCPMC2913939. doi:10.1186/1479-5876-8-63
115. He J, Liu J, Huang Y, et al. Olfactory mucosa mesenchymal stem cells alleviate cerebral ischemia/reperfusion injury via golgi apparatus secretory pathway Ca(2+) -ATPase Isoform1. Front Cell Dev Biol. 2020;8:586541. PubMed PMID: 33195239; PubMed Central PMCID: PMCPMC7661436. doi:10.3389/fcell.2020.586541
116. Liu Q, Tan Y, Qu T, et al. Therapeutic mechanism of human neural stem cell-derived extracellular vesicles against hypoxia-reperfusion injury in vitro. Life Sci. 2020;254:117772. PubMed PMID: 32437794. doi:10.1016/j.lfs.2020.117772
117. Xia Y, Ling X, Hu G, et al. Small extracellular vesicles secreted by human iPSC-derived MSC enhance angiogenesis through inhibiting STAT3-dependent autophagy in ischemic stroke. Stem Cell Res Ther. 2020;11(1):313. PubMed PMID: 32698909; PubMed Central PMCID: PMCPMC7374834. doi:10.1186/s13287-020-01834-0
118. Li Y, Quan X, Hu J, et al. BMSCs-derived small extracellular vesicles antagonize cerebral endothelial Caveolin-1 driven autophagic degradation of tight-junction proteins to protect blood-brain barrier post-stroke. Int J Biol Sci. 2025;21(2):842–859. PubMed PMID: 39781452; PubMed Central PMCID: PMCPMC11705626. doi:10.7150/ijbs.101937
119. Kuang Y, Zheng X, Zhang L, et al. Adipose-derived mesenchymal stem cells reduce autophagy in stroke mice by extracellular vesicle transfer of miR-25. J Extracell Vesicles. 2020;10(1):e12024. PubMed PMID: 33304476; PubMed Central PMCID: PMCPMC7710129. doi:10.1002/jev2.12024
120. Zhao Y, Zhang X, Chen X, Wei Y. Neuronal injuries in cerebral infarction and ischemic stroke: from mechanisms to treatment (Review). Int J Mol Med. 2022;49(2). PubMed PMID: 34878154; PubMed Central PMCID: PMCPMC8711586. doi:10.3892/ijmm.2021.5070
121. Orellana-Urzúa S, Rojas I, Líbano L, Rodrigo R. Pathophysiology of ischemic stroke: role of oxidative stress. Curr Pharm Des. 2020;26(34):4246–4260. PubMed PMID: 32640953. doi:10.2174/1381612826666200708133912
122. Tao T, Liu M, Chen M, et al. Natural medicine in neuroprotection for ischemic stroke: challenges and prospective. Pharmacol Ther. 2020;216:107695. PubMed PMID: 32998014. doi:10.1016/j.pharmthera.2020.107695
123. Pei X, Li Y, Zhu L, Zhou Z. Astrocyte-derived exosomes suppress autophagy and ameliorate neuronal damage in experimental ischemic stroke. Exp Cell Res. 2019;382(2):111474. PubMed PMID: 31229506. doi:10.1016/j.yexcr.2019.06.019
124. Hong SB, Yang H, Manaenko A, Lu J, Mei Q, Hu Q. Potential of exosomes for the treatment of stroke. Cell Transplant. 2019;28(6):662–670. PubMed PMID: 30520322; PubMed Central PMCID: PMCPMC6686440. doi:10.1177/0963689718816990
125. Waseem A, Saudamini HR, Janowski M, Raza SS. Mesenchymal stem cell-derived exosomes: shaping the next era of stroke treatment. Neuroprotection. 2023;1(2):99–116. PubMed PMID: 38283953; PubMed Central PMCID: PMCPMC10811806. doi:10.1002/nep3.30
126. Candelario-Jalil E, Dijkhuizen RM, Magnus T. Neuroinflammation, stroke, blood-brain barrier dysfunction, and imaging modalities. Stroke. 2022;53(5):1473–1486. PubMed PMID: 35387495; PubMed Central PMCID: PMCPMC9038693. doi:10.1161/strokeaha.122.036946
127. Jiang X, Andjelkovic AV, Zhu L, et al. Blood-brain barrier dysfunction and recovery after ischemic stroke. Prog Neurobiol. 2018;163-164:144–171. PubMed PMID: 28987927; PubMed Central PMCID: PMCPMC5886838. doi:10.1016/j.pneurobio.2017.10.001
128. Cai J, Gehrau R, Tu Z, et al. MicroRNA-206 antagomiR‒enriched extracellular vesicles attenuate lung ischemia‒reperfusion injury through CXCL1 regulation in alveolar epithelial cells. J Heart Lung Transpl. 2020;39(12):1476–1490. PubMed PMID: 33067103; PubMed Central PMCID: PMCPMC7704771. doi:10.1016/j.healun.2020.09.012
129. Gao C, Chen L, Xie XY, He XF, Shen J, Zheng L. Bone marrow mesenchymal stem cells-derived exosomal miR-381 alleviates lung ischemia-reperfusion injury by activating Treg differentiation through inhibiting YTHDF1 expression. Cell. Signalling. 2024;124:111440. PubMed PMID: 39357613. doi:10.1016/j.cellsig.2024.111440
130. Gao C, Xu YJ, Meng ZX, Gu S, Zhang L, Zheng L. BMSC-derived exosomes carrying lncRNA-ZFAS1 alleviate pulmonary ischemia/reperfusion injury by UPF1-mediated mRNA decay of FOXD1. Molecular neurobiology. 2023;60(5):2379–2396. PubMed PMID: 36652050. doi:10.1007/s12035-022-03129-2
131. Li JW, Wei L, Han Z, Chen Z. Mesenchymal stromal cells-derived exosomes alleviate ischemia/reperfusion injury in mouse lung by transporting anti-apoptotic miR-21-5p. Eur J Pharmacol. 2019;852:68–76. PubMed PMID: 30682335. doi:10.1016/j.ejphar.2019.01.022
132. Liu J, Chen T, Lei P, Tang X, Huang P. Exosomes released by bone marrow mesenchymal stem cells attenuate lung injury induced by intestinal ischemia reperfusion via the TLR4/NF-κB pathway. Int J Med Sci. 2019;16(9):1238–1244. PubMed PMID: 31588189; PubMed Central PMCID: PMCPMC6775266. doi:10.7150/ijms.35369
133. Lonati C, Bassani GA, Brambilla D, et al. Mesenchymal stem cell-derived extracellular vesicles improve the molecular phenotype of isolated rat lungs during ischemia/reperfusion injury. J Heart Lung Transpl. 2019;38(12):1306–1316. PubMed PMID: 31530458. doi:10.1016/j.healun.2019.08.016
134. Stone ML, Zhao Y, Robert Smith J, et al. Mesenchymal stromal cell-derived extracellular vesicles attenuate lung ischemia-reperfusion injury and enhance reconditioning of donor lungs after circulatory death. Respir Res. 2017;18(1):212. PubMed PMID: 29268735; PubMed Central PMCID: PMCPMC5740880. doi:10.1186/s12931-017-0704-9
135. Sun ZL, You T, Zhang BH, Liu Y, Liu J. Bone marrow mesenchymal stem cell-derived exosomes miR-202-5p inhibited pyroptosis to alleviate lung ischemic-reperfusion injury by targeting CMPK2. Kaohsiung J Med Sci. 2023;39(7):688–698. PubMed PMID: 37092308. doi:10.1002/kjm2.12688
136. Zhang B, Meng C, Kang JY, Zhou HC. Exosomes derived from bone marrow mesenchymal stem cells regulate NF-κB pathway and reduce lung ischemia-reperfusion injury in rats by miR-335]. Sheng li xue bao. Acta physiologica Sinica. 2024;76(2):247–256. PubMed PMID: 38658374.
137. Schäfer R, Spohn G, Baer PC. Mesenchymal stem/stromal cells in regenerative medicine: can preconditioning strategies improve therapeutic efficacy? Transfus Med Hemother. 2016;43(4):256–267. PubMed PMID: 27721701; PubMed Central PMCID: PMCPMC5040900. doi:10.1159/000447458
138. Vasiliadis AV, Galanis N. Human bone marrow-derived mesenchymal stem cells from different bone sources: a panorama. Stem Cell Investig. 2020;7:15. PubMed PMID: 32964008; PubMed Central PMCID: PMCPMC7492710. doi:10.21037/sci-2020-013
139. Jovic D, Yu Y, Wang D, et al. A brief overview of global trends in MSC-based cell therapy. Stem Cell Rev Rep. 2022;18(5):1525–1545. PubMed PMID: 35344199; PubMed Central PMCID: PMCPMC8958818. doi:10.1007/s12015-022-10369-1
140. Aslam M, Baveja R, Liang OD, et al. Bone marrow stromal cells attenuate lung injury in a murine model of neonatal chronic lung disease. Am J Respir Crit Care Med. 2009;180(11):1122–1130. PubMed PMID: 19713447; PubMed Central PMCID: PMCPMC2784417. doi:10.1164/rccm.200902-0242OC
141. Wilson JG, Liu KD, Zhuo H, et al. Mesenchymal stem (stromal) cells for treatment of ARDS: a Phase 1 clinical trial. Lancet Respir Med. 2015;3(1):24–32. PubMed PMID: 25529339; PubMed Central PMCID: PMCPMC4297579. doi:10.1016/s2213-2600(14)70291-7
142. Herberts CA, Kwa MS, Hermsen HP. Risk factors in the development of stem cell therapy. J Transl Med. 2011;9:29. PubMed PMID: 21418664; PubMed Central PMCID: PMCPMC3070641. doi:10.1186/1479-5876-9-29
143. Sun J, Li Y. Pyroptosis and respiratory diseases: a review of current knowledge. Front Immunol. 2022;13:920464. PubMed PMID: 36248872; PubMed Central PMCID: PMCPMC9561627. doi:10.3389/fimmu.2022.920464
144. Ketelut-Carneiro N, Fitzgerald KA. Apoptosis, pyroptosis, and necroptosis-oh my! The many ways a cell can die. J Mol Biol. 2022;434(4):167378. PubMed PMID: 34838807. doi:10.1016/j.jmb.2021.167378
145. Yuan J, Ofengeim D. A guide to cell death pathways. Nat Rev Mol Cell Biol. 2024;25(5):379–395. PubMed PMID: 38110635. doi:10.1038/s41580-023-00689-6
146. Newton K, Strasser A, Kayagaki N, Dixit VM. Cell death. Cell. 2024;187(2):235–256. PubMed PMID: 38242081. doi:10.1016/j.cell.2023.11.044
147. Zhao X, Wu J, Yuan R, et al. Adipose-derived mesenchymal stem cell therapy for reverse bleomycin-induced experimental pulmonary fibrosis. Sci Rep. 2023;13(1):13183. PubMed PMID: 37580529; PubMed Central PMCID: PMCPMC10425426. doi:10.1038/s41598-023-40531-9
148. Zuk PA, Zhu M, Ashjian P, et al. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13(12):4279–4295. PubMed PMID: 12475952; PubMed Central PMCID: PMCPMC138633. doi:10.1091/mbc.e02-02-0105
149. Strioga M, Viswanathan S, Darinskas A, Slaby O, Michalek J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012;21(14):2724–2752. PubMed PMID: 22468918. doi:10.1089/scd.2011.0722
150. Gutiérrez-Fernández M, Rodríguez-Frutos B, Ramos-Cejudo J, et al. Effects of intravenous administration of allogenic bone marrow- and adipose tissue-derived mesenchymal stem cells on functional recovery and brain repair markers in experimental ischemic stroke. Stem Cell Res Ther. 2013;4(1):11. PubMed PMID: 23356495; PubMed Central PMCID: PMCPMC3706777. doi:10.1186/scrt159
151. Zhidu S, Ying T, Rui J, Chao Z. Translational potential of mesenchymal stem cells in regenerative therapies for human diseases: challenges and opportunities. Stem Cell Res Ther. 2024;15(1):266. PubMed PMID: 39183341; PubMed Central PMCID: PMCPMC11346273. doi:10.1186/s13287-024-03885-z
152. Cheng SL, Lin CH, Yao CL. Mesenchymal stem cell administration in patients with chronic obstructive pulmonary disease: state of the science. Stem Cells Int. 2017;2017:8916570. PubMed PMID: 28303154; PubMed Central PMCID: PMCPMC5337878. doi:10.1155/2017/8916570
153. Miceli V, Bulati M, Gallo A, et al. Role of mesenchymal stem/stromal cells in modulating ischemia/reperfusion injury: current state of the art and future perspectives. Biomedicines. 2023;11(3). PubMed PMID: 36979668; PubMed Central PMCID: PMCPMC10045387. doi:10.3390/biomedicines11030689
154. Velikova T, Dekova T, Miteva DG. Controversies regarding transplantation of mesenchymal stem cells. World J Transplant. 2024;14(2):90554. PubMed PMID: 38947963; PubMed Central PMCID: PMCPMC11212595. doi:10.5500/wjt.v14.i2.90554
155. Manzoor T, Saleem A, Farooq N, et al. Extracellular vesicles derived from mesenchymal stem cells - a novel therapeutic tool in infectious diseases. Inflamm Regen. 2023;43(1):17. PubMed PMID: 36849892; PubMed Central PMCID: PMCPMC9970864. doi:10.1186/s41232-023-00266-6
156. Alcaraz MJ, Compañ A, Guillén MI. Extracellular vesicles from mesenchymal stem cells as novel treatments for musculoskeletal diseases. Cells. 2019;9(1). PubMed PMID: 31906087; PubMed Central PMCID: PMCPMC7017209. doi:10.3390/cells9010098
157. Vonk LA, van Dooremalen SFJ, Liv N, et al. Mesenchymal stromal/stem cell-derived extracellular vesicles promote human cartilage regeneration in vitro. Theranostics. 2018;8(4):906–920. PubMed PMID: 29463990; PubMed Central PMCID: PMCPMC5817101. doi:10.7150/thno.20746
158. Zhang X, Liu T, Ran C, et al. Immunoregulatory paracrine effect of mesenchymal stem cells and mechanism in the treatment of osteoarthritis. Front Cell Dev Biol. 2024;12:1411507. PubMed PMID: 39129785; PubMed Central PMCID: PMCPMC11310049. doi:10.3389/fcell.2024.1411507
159. Kim DW, Staples M, Shinozuka K, Pantcheva P, Kang SD, Borlongan CV. Wharton’s jelly-derived mesenchymal stem cells: phenotypic characterization and optimizing their therapeutic potential for clinical applications. Int J Mol Sci. 2013;14(6):11692–11712. PubMed PMID: 23727936; PubMed Central PMCID: PMCPMC3709752. doi:10.3390/ijms140611692
160. Todtenhaupt P, Franken LA, Groene SG, et al. A robust and standardized method to isolate and expand mesenchymal stromal cells from human umbilical cord. Cytotherapy. 2023;25(10):1057–1068. PubMed PMID: 37516948. doi:10.1016/j.jcyt.2023.07.004
161. Ullah I, Subbarao RB, Rho GJ. Human mesenchymal stem cells - current trends and future prospective. Biosci Rep. 2015;35(2). PubMed PMID: 25797907; PubMed Central PMCID: PMCPMC4413017. doi:10.1042/bsr20150025
162. Fong CY, Richards M, Manasi N, Biswas A, Bongso A. Comparative growth behaviour and characterization of stem cells from human Wharton’s jelly. Reprod Biomed Online. 2007;15(6):708–718. PubMed PMID: 18062871. doi:10.1016/s1472-6483(10)60539-1
163. Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3(3):301–313. PubMed PMID: 18786417. doi:10.1016/j.stem.2008.07.003
164. Fan CG, Zhang QJ, Zhou JR. Therapeutic potentials of mesenchymal stem cells derived from human umbilical cord. Stem Cell Rev Rep. 2011;7(1):195–207. PubMed PMID: 20676943. doi:10.1007/s12015-010-9168-8
165. Zeng Y, Wu Z, Chen G, et al. Peripheral injection of hUC-MSCs in the treatment of acute liver failure: a pre-clinical cohort study in rhesus monkeys. Stem Cells Int. 2024;2024:4654912. PubMed PMID: 39045027; PubMed Central PMCID: PMCPMC11265939. doi:10.1155/2024/4654912
166. Yang Y, Duan Y, Yue J, et al. Exosomes: an innovative therapeutic target for cerebral ischemia-reperfusion injury. Front Pharmacol. 2025;16:1552500. PubMed PMID: 40206077; PubMed Central PMCID: PMCPMC11979243. doi:10.3389/fphar.2025.1552500
167. Wu H, Qin W, Lu S, et al. Long noncoding RNA ZFAS1 promoting small nucleolar RNA-mediated 2’-O-methylation via NOP58 recruitment in colorectal cancer. Mol Cancer. 2020;19(1):95. PubMed PMID: 32443980; PubMed Central PMCID: PMCPMC7243338. doi:10.1186/s12943-020-01201-w
168. Hansji H, Leung EY, Baguley BC, et al. ZFAS1: a long noncoding RNA associated with ribosomes in breast cancer cells. Biology Direct. 2016;11(1):62. PubMed PMID: 27871336; PubMed Central PMCID: PMCPMC5117590. doi:10.1186/s13062-016-0165-y
169. Dong D, Mu Z, Zhao C, Sun M. ZFAS1: a novel tumor-related long non-coding RNA. Can Cell Inter. 2018;18:125. PubMed PMID: 30186041; PubMed Central PMCID: PMCPMC6122210. doi:10.1186/s12935-018-0623-y
170. Wu F, Huang W, Tan Q, et al. ZFP36L2 regulates myocardial ischemia/reperfusion injury and attenuates mitochondrial fusion and fission by LncRNA PVT1. Cell Death Dis. 2021;12(6):614. PubMed PMID: 34131106; PubMed Central PMCID: PMCPMC8206151. doi:10.1038/s41419-021-03876-5
171. Wang FD, Ding Y, Zhou JH, et al. Gamma-aminobutyric acid enhances miR-21-5p loading into adipose-derived stem cell extracellular vesicles to alleviate myocardial ischemia-reperfusion injury via TXNIP regulation. World J Stem Cells. 2024;16(10):873–895. PubMed PMID: 39493825; PubMed Central PMCID: PMCPMC11525649. doi:10.4252/wjsc.v16.i10.873
172. Yu Z, Wen Y, Jiang N, et al. TNF-α stimulation enhances the neuroprotective effects of gingival MSCs derived exosomes in retinal ischemia-reperfusion injury via the MEG3/miR-21a-5p axis. Biomaterials. 2022;284:121484. PubMed PMID: 35378413. doi:10.1016/j.biomaterials.2022.121484
173. Liu C, Yin T, Zhang M, et al. Function of miR-21-5p derived from ADSCs-exos on the neuroinflammation after cerebral ischemia. J Stroke Cerebrovasc Dis. 2024;33(8):107779. PubMed PMID: 38768666. doi:10.1016/j.jstrokecerebrovasdis.2024.107779
174. Parvan R, Hosseinpour M, Moradi Y, Devaux Y, Cataliotti A, Da Silva GJJ. Diagnostic performance of microRNAs in the detection of heart failure with reduced or preserved ejection fraction: a systematic review and meta-analysis. Eur J Heart Fail. 2022;24(12):2212–2225. PubMed PMID: 36161443; PubMed Central PMCID: PMCPMC10092442. doi:10.1002/ejhf.2700
175. Li SS, Wu JJ, Xing XX, et al. Focal ischemic stroke modifies microglia-derived exosomal miRNAs: potential role of mir-212-5p in neuronal protection and functional recovery. Biol Res. 2023;56(1):52. PubMed PMID: 37789455; PubMed Central PMCID: PMCPMC10548705. doi:10.1186/s40659-023-00458-x
176. Li C, Fei K, Tian F, Gao C, Yang S. Adipose-derived mesenchymal stem cells attenuate ischemic brain injuries in rats by modulating miR-21-3p/MAT2B signaling transduction. Croatian Med J. 2019;60(5):439–448. PubMed PMID: 31686458; PubMed Central PMCID: PMCPMC6852138. doi:10.3325/cmj.2019.60.439
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Proteomic Characterization of Extracellular Vesicles from Human Neural Precursor Cells: A Promising Advanced Therapy for Neurodegenerative Diseases

Stricker PEF, de Oliveira NB, Mogharbel BF, Irioda AC, da Rosa NN, Lührs L, Saçaki CS, Munhoz da Rocha I, Alves LR, Poubel SB, Cardoso da Silva J, Carvalho PC, Fischer JSG, de Carvalho KAT
International Journal of Nanomedicine 2025, 20:6675-6699
Published Date: 25 May 2025
Mesenchymal Stem Cell-Derived Extracellular Vesicles: Emerging Therapies for Neurodegenerative Diseases
Chen P, Wang F, Ling B, Zhu Y, Lin H, Huang J, Wang X
International Journal of Nanomedicine 2025, 20:8547-8565
Published Date: 2 July 2025