Back to Journals » Cancer Management and Research » Volume 12
FOXCUT Promotes the Proliferation and Invasion by Activating FOXC1/PI3K/AKT Pathway in Colorectal Cancer
Authors Zhang X, Yi S, Xing G, Wu H, Zhu Y, Guo X, Zhang L
Received 12 May 2020
Accepted for publication 17 June 2020
Published 24 July 2020 Volume 2020:12 Pages 6269—6278
DOI https://doi.org/10.2147/CMAR.S259801
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kenan Onel
Xiaojie Zhang, Shanyong Yi, Guochen Xing, Huili Wu, Ying Zhu, Xiaodan Guo, Lei Zhang
Zhengzhou Central Hospital Affiliated to Zhengzhou University, Zhengzhou 450000, People’s Republic of China
Correspondence: Lei Zhang
Gastrointestinal Surgery, Zhengzhou Central Hospital Affiliated to Zhengzhou University, No. 195, Tongbai Road, Zhengzhou 450000, People’s Republic of China
Email [email protected]
Introduction: Colorectal cancer (CRC) is the third most commonly diagnosed world cancer. Long noncoding RNAs (lncRNAs) serve important regulatory roles in tumorigenesis. However, the contributions of lncRNAs to human CRC remain largely unknown.
Material and Methods: FOXC1 and FOXCUT lncRNA expression levels were detected in a panel of paired specimens obtained from 48 patients’ tissues and cell lines with CRC using RT-qPCR. RNA interference was used to investigate potential correlations between FOXC1 and FOXCUT expression in HT29. Cell proliferation was assessed by MTT assay and EdU incorporation assay. The migration and invasion of CRC cells were detected by transwell assay. Western blot was applied to assess the protein expression and PI3K/AKT signaling pathway.
Results: In this study, a novel long noncoding RNA (FOXCUT) was frequently overexpressed in CRC tissues and cell lines. In addition, the expressions of FOXCUT and FOXC1 were positively correlated. When the expression of FOXCUT was downregulated by small interfering RNA (siRNA), the expression of FOXC1 was also decreased. Moreover, knockdown of FOXCUT significantly inhibited proliferation and invasion of CRC cell lines and resulted in downregulated expression of the matrix metalloproteinase 1 (MMP-1). Mechanistically, FOXCUT promotes the expression of FOXC1 to activate PI3K/AKT signaling pathway for its regulation of cell growth and proliferation.
Conclusion: In summary, our findings indicate that FOXCUT plays an important oncogenic role and may serve as a novel biomarker and therapeutic target in CRC progression.
Keywords: colorectal cancer, FOXCUT, FOXC1, proliferation, invasion
Introduction
Colorectal cancer (CRC), which third in morbidity and second-highest mortality, is one of the most common malignancies.1,2 The incidence of CRC is increasing year by year, which seriously threatens the life and health of the public in China.3 Due to the lack of biomarkers for early diagnosis, prognostic indicators and effective treatment targets, the treatment of CRC has always been a bottleneck. Therefore, it is of great clinical and social significance to find new molecular markers and therapeutic targets to improve the survival rate of patients.
FOXC1 is a member of the transcription factor forkhead box (FOX) protein superfamily.4 FOXC1 regulates tumor-associated genes and is regulated by several mechanisms that control its gene expression and protein activity. Additionally, it affects the cell cycle and proliferation of tumor cells by epithelial-to-mesenchymal transition (EMT), cellular signal transduction, migration of vascular endothelial cell.5 It has been reported that FOXC1 gene is highly expressed in many kinds of tumors,4,6-8 such as pancreatic ductal adenocarcinoma (PDA), basal-like breast cancer (BLBC), esophageal cancer and gastric cancer (ESCC). Importantly, FOXC1 may act as a subtype-specific biomarker for breast cancer and acute myeloid leukemia (AML). It was reported that FOXC1 expression is significantly higher in BLBC than in luminal and HER2 breast cancer.4 High levels of FOXC1 in AML (FAB-M2) exhibit essential relevance with morphological classifications associated with granulocyte differentiation compared with other AML subtypes.9 Furthermore, in most cancer types, high FOXC1 expression may predict clinical features such as malignant clinical manifestations, increased pathological grade, and poor prognosis of patients10,11 and it may be considered as a novel target for therapy. However, the relationship between FOXC1 gene and CRC remains largely unknown.
Long noncoding RNAs (lncRNAs) are now defined as the long RNA transcripts with >200 nucleotides lacking protein encoding potential or with limited peptide coding potential.12–14 Extensive evidence suggests that lncRNAs have vital roles in regulating gene expression at levels of chromatin modification, transcriptional and posttranscriptional regulation. Many lncRNAs have been identified to affect tumorigenesis.13,15-17 A novel lncRNA named FOXC1 promoter upstream transcript (FOXCUT) has been identified to functionally contact its adjacent FOXC1 mRNA and take on the form of ‘lncRNA-mRNA pairs’ in ESCC, oral squamous cell carcinoma (OSCC), BLBC, and nasopharyngeal carcinoma (NPC).18–21 Nevertheless, whether lncRNA plays a role in CRC is not clear yet. In this study, we first explored the expression correlation and clinicopathological significance of FOXCUT and FOXC1 in CRC, so as to provide new clues and research targets for the pathogenesis, prognosis and treatment of CRC.
Materials and Methods
The colon cancer cell lines (Caco-2, HCT116, HT29 and DLD-1) and the human colonic epithelial cells (NCM-460) used for experiments were obtained from Type Culture Collection (CCTCC, China). All cells were regularly authenticated by short tandem repeat (STR) analysis and tested for the absence of Mycoplasma contamination. Human colon cancer tissues and the corresponding adjacent mucosa were obtained from biopsy in a total of 48 consecutive colon cancer cases admitted in our Zhengzhou central hospital, China, from April 2018 to April 2019. CRC diagnosis was histopathologically confirmed, and none of the patients received treatment prior to the biopsy. The utilization of tumor material for research was approved by the ethical committee of Zhengzhou central hospital, which was in accordance with the Declaration of Helsinki. Written informed consent was obtained from the patients. The tissues from each subject were immersed overnight in RNAlater reagent (Thermo Scientific, USA) at 4°C and stored at −80°C.
Cell Culture and Transfection
Colon cancer cell lines and NCM460 were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Invitrogen, Carlsbad, CA, USA), whereas Caco-2, HCT116, HT29 and DLD-1cells were cultivated in RPMI 1640 (21875091 Thermo Fisher Scientific) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco, Life Technologies), 100 U/mL of penicillin, and 100 µg/mL of streptomycin (P4333, Sigma). Cells in this medium were placed in a humidified atmosphere containing 5% CO2 at 37°C.
The sequence of short-hairpin RNA (shRNA) directed against FOXCUT (5ʹ- CACCGGCATTGATGCAAGGATATCC CGAA GGATATCCTTGCATCAATGCC −3ʹ) was ligated into the pLKO.1-Puro vector (TaKaRa Bio, Tokyo, Japan). We packaged lentivirus into human embryonic kidney 293T (HEK293T) cells using Lipofectamine 2000 (Life Technologies, Carlsbad, CA, USA) and the supernatant was collected following the manufacturer’s instructions. Lentiviral particles were used to infect HT29 cells. We established stable cell lines via puromycin selection and then used for subsequent in vitro and in vivo experiments. The PCR product of the FOXC1 ORF was recovered and isolated by gel extraction, then inserted into the pcDNA3.1 Vector (TaKaRa Bio, Tokyo, Japan), and sequenced (Sangon Biotech, Shanghai, China). FOXC1 ORF forward primer containing a BamHI site and the antisense primer containing an EcoRI site to obtain the ORF fragment. For transient transfection assays small interfering RNA (siRNA) duplexes (si-FOXCUT #1 and si-FOXCUT #2), and negative control (NC) RNA duplexes were synthesized (GenePharma, Shanghai, China) by using the primer pairs (Table 1). For transfection of siRNA or pcDNA3.1-FOXC1, HT29 cells were seeded at 2x105 cells/well in 6-well plates (Eppendorf, Hamburg, Germany) 24 h before transfection. When the cells were grown to 80% confluence, cells were transfected a transfection complex consisting of experimental siRNA, pcDNA3.1-FOXC1, complex of siRNA and pcDNA3.1-FOXC1, or control and Lipofectamine® 2000 Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions, and cultured at 37°C and 5% CO2. For insulin-like growth factor 1 (IGF-1) treatment, IGF-1 was reconstituted at 200 ng/mL in sterile phosphate-buffered saline (PBS), cells were incubated with 200 ng/mL.22,23 Then, supernatant was replaced with serum containing growth medium. Cells were lysed and harvested for RNA and protein after 48 h transfection. Recombinant human IGF-1 was purchased from R&D Systems (Minneapolis, USA).
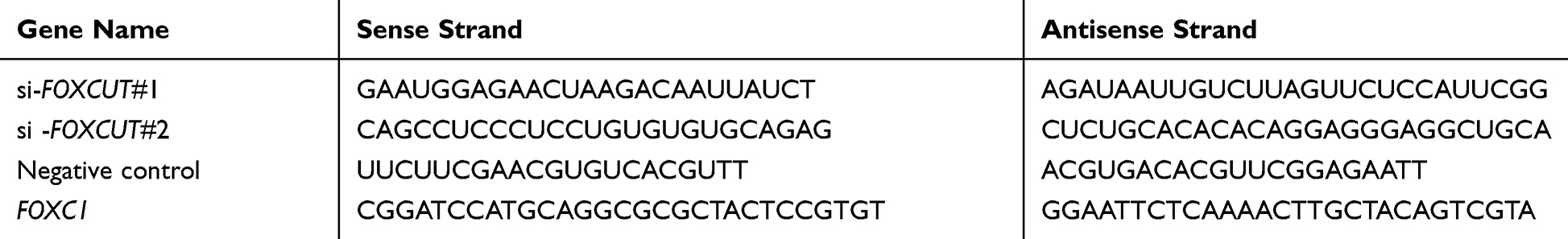 |
Table 1 Sequences for Small Interfering RNA Analysis and Vector Construction |
Quantitative PCR (qPCR)
To quantitatively determine the mRNA expression level of FOXCUT and FOXC1 in colon cancer cells, tissues and colonic epithelial cells, real-time PCR was performed, primers of qPCR were synthesized (Sangon Biotech, Shanghai, China) and primer information are showed in Table 2. Total RNA was extracted from each sample using TRIzol Total RNA reagent (Invitrogen Life Technologies, USA) according to the manufacturer’s protocol. The concentration of RNA samples was measured using a NanoDrop 2000 (Thermo Scientific, USA) and the quality confirmed by agarose gel electrophoresis. Total RNA (1 μg) was reverse transcribed into cDNA with FastKing RT Kit including DNase treatment (Takara, Shiga, Japan). cDNA amplification was performed using the SYBR-Green master mix kit (Tiangen, China) following the manufacturer’s instructions. The reaction mixtures were incubated at 94°C for 5 min, followed by 40 cycles of 94°C for 5 sec,60°C for 34 sec and 72°C for 20 sec. Data were analyzed using the comparative threshold cycle (Ct) method (2−∆∆Ct).24 The ACTB gene was used as an endogenous control. Triplicate wells were performed per sample.
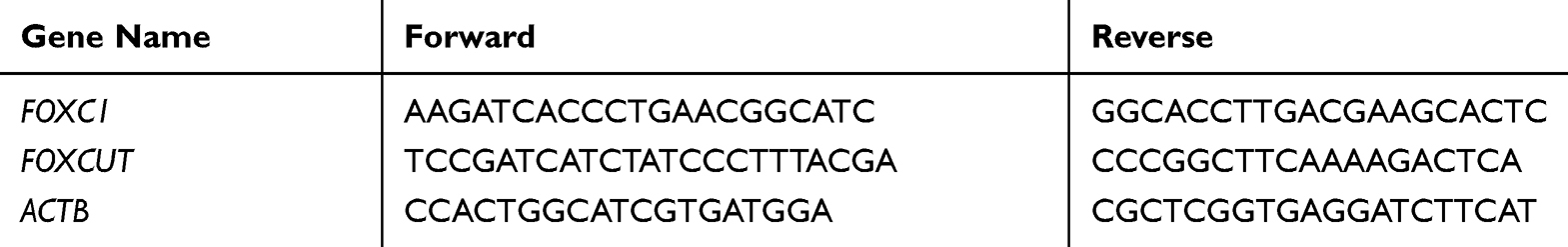 |
Table 2 Primer Sequences for Quantitative Real-Time PCR |
Cell Proliferation Assay
Cell proliferation was analyzed using the MTT assay. Briefly, cells in a 96-well plate were (Eppendorf, Hamburg, Germany) incubated in a humidified 5% CO2 chamber after transfection, followed by addition of 20 µL of MTT dye (2.5 mg/mL) at 0, 24, 48 and 72 h, and subsequently incubated with 150 µL of Dimethyl sulfoxide for 5 min. The absorbance at 570 nm was recorded using an Enzyme Immunoassay Analyzer (Bio-Rad, Hercules, USA).
EdU Incorporation Assay
The cell proliferation was also examined by 5-ethynyl-2′-deoxyuridine (EdU) incorporation assay.25 Briefly, indicated cells were incubated with 50 μM EdU for 2 h at 37°C after 24 h transfection. Cells were then washed with PBS for three times, and fixed with 4% formaldehyde for 30 min at room temperature (RT) according to the manufacturer’s protocol (RiboBio, China). Then, 0.5% Triton X-100 was applied for 10 min at RT to permeabilize cells. Cells were incubated with 1 × Apollo reaction cocktail for 30 min after washing with PBS for three times. DNA was stained with 4′,6-diamidino-2-phenylindole (DAPI) for 30 min and results were visualized with a Zeiss confocal microscope (LSM 700, USA).
Transwell Invasion Assay
Cell invasion was examined using the transwell system chambers with 8 μm pore size polycarbonate membrane (BD Biosciences, San Jose, CA, USA) following the manufacturer’s instructions. In brief, HT-29 cells (5×105 cells/well) were inoculated to an upper chamber and the inserts were coated with Matrigel (BD Biosciences, USA). Six hundred μL of complete medium was present in that bottom chamber. The cells were allowed to invade for 24 h at 37°C with 5% CO2, the medium was removed from the upper chamber. The non-invaded cells were removed from on the upper side of the chamber, whereas the cells on the bottom surface of the surface were fixed with 0.05% crystal violet for 2 h. Finally, the number of invaded cells was counted by using an inverted microscope and the relative number was calculated.
Western Blot Assay
CRC lines were harvested and extracted using RIPA lysis buffer (Thermo Scientific, MA). Antibody and dilutions used in the studies: polyclonal antibodies against PI3K, AKT, p-PI3K, p-AKT (1: 2000, 1: 1000, Cell Signaling, Danvers, MA), monoclonal MMP-1 and FOXC1 (1:2000 Bio-techne, Minnesota, USA), and anti-GAPDH antibody (Sigma-Aldrich, St. Louis, MO) was used at a 1:3000 dilution. Protein bands were quantified using Quantity One 4.5.0 software (Bio-Rad, Hercules, CA, USA). Signals were visualized using ECL Substrates (Thermo Scientific, MA). The relative protein levels in different cell lines were normalized to the concentration of GAPDH. The experiment was done in triplicate.
Animal Studies
All animal experiments were approved by the Animal Care Committee of Zhengzhou University and performed following the guidelines and regulations of the Ethics Committee of Zhengzhou University for the welfare of animals in cancer research. Male BALB/c nude mice (4–5 weeks old) were purchased from Vital River Laboratory Animal Technologies Co. Ltd (Beijing, China). For xenograft experiments, 1×107 cells were subcutaneously injected into the flanks of animals (n = 5 per group). Tumor growth was examined every 4 days for 4 weeks. Tumor volumes were calculated by the equation V(mm3) = a × b × c/2, where a is the length, b is the width, and c is the height. Tumors were harvested for RNA and protein assays.
Statistical Analysis
Data were presented as means ± standard deviation (SD), and GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA, USA) software was used for data statistical analysis. Independent samples t-test (two groups) or one-way ANOVA (no less than three groups) was used to analyse the statistical significance at a significance. * P < 0.05.
Results
FOXCUT and FOXC1 are Overexpressed in CRC Tissues and Cell Lines
FOXCUT and FOXC1 expression levels were detected in a panel of paired specimens obtained from 48 patient’s tissues and cell lines with CRC using RT-qPCR. The results showed that, compared with the adjacent mucosa, the FOXCUT and FOXC1 mRNA level were significantly increased in human colon cancer tissues (Figure 1A and B). Similarly, the expressions of FOXCUT and FOXC1 were significantly higher in four colon cancer cell lines (Caco-2, HCT116, HT29 and DLD-1), compared with those NCM460 (Figure 1C and D). Because FOXCUT and FOXC1 are highly expressed in HT29 cells, these cell lines were exploited for further study. In addition, the relative expression of FOXCUT was positively correlated with that of FOXC1 in the CRC tissue samples. These results suggested that FOXCUT and FOXC1 were highly expressed in CRC in association with cancer progression.
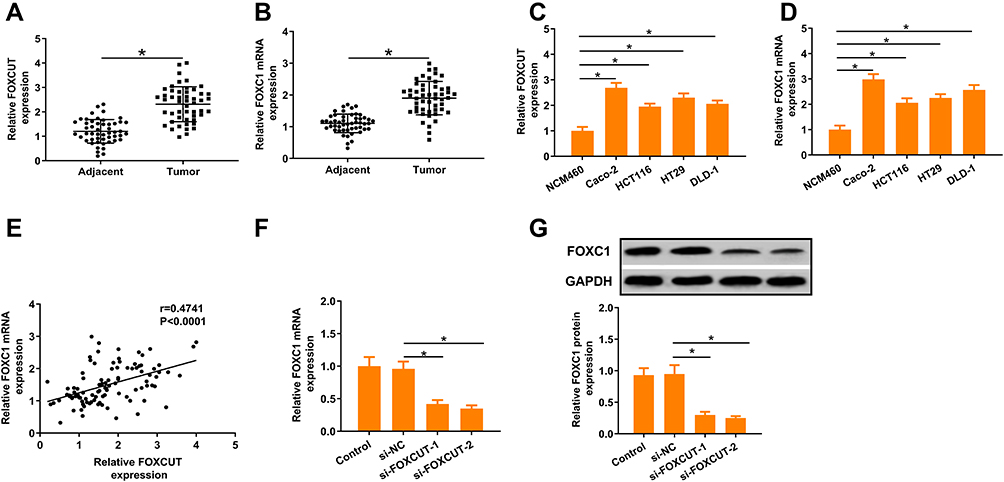 |
Figure 1 FOXCUT and FOXC1 expression is frequently upregulated in CRC cell lines and tissues. (A, B) Expression of FOXCUT and FOXC1 was detected by qPCR in adjacent mucosa and colon cancer tissues (n = 48). (C, D) Abundance of FOXCUT and FOXC1 in CRC cell lines relative to that in the colonic epithelial cell line NCM460. The expression of FOXCUT and FOXC1 was normalized to that in NCM460. The statistical differences between groups were analyzed using independent samples t-test. (E) Linear regression analysis was performed on FOXC1 and lncRNA-FOXCUT expression levels in 48 CRC tissue samples (n = 48). r=0.4741, P<0.0001. (F, G) FOXC1 expressions were significantly attenuated following FOXCUT siRNA transfection both in mRNA and protein levels. Relative expression of the target genes was quantified for each transcript and were normalized with the expression of β-actin by 2− ΔΔCt method. Data show the mean ± SD. * indicates significant differences compared with the control group (P < 0.05). |
To further investigate potential correlations between FOXCUT and FOXC1expression, we used RNA interference to silence FOXCUT expression in HT29, the expression of FOXCUT was detected by RT-qPCR after transfection. The results showed that FOXCUT gene was successfully silenced in HT29 cells after transfection with FOXCUT siRNA (Supplementary Figure S1). Our results also showed that FOXC1 expression was inhibited by si-FOXCUT both in mRNA and protein levels (Figure 1E–G). These results indicate that FOXCUT promoted FOXC1 expression in HT29 cells.
Knockdown of FOXCUT Inhibited the Cell Proliferation and Invasion Ability in CRC
We further investigated the roles of FOXCUT on CRC development. To clarify whether FOXCUT has a functional role in facilitating CRC cell progression, we examined cell proliferative activities and invasive abilities by MTT assays, EdU incorporation assay and transwell invasion assay. The results showed that inhibiting FOXCUT markedly diminished the proliferative activities of HT29 in a time-dependent manner compared to the control group (Figure 2A), the amount of cancer cells was markedly lower in FOXCUT siRNA (Figure 2C). Correspondingly, we found that the invasive potential of HT29 was apparently decreased in FOXCUT siRNA group (Figure 2E). Furthermore, the MMP-1 (matrix metalloproteinase-1) have been identified as important indicators in CRC.26–28 We also found that the expression level of MMP-1 protein was significantly downregulated in FOXCUT siRNA group (Figure 2H). On the contrary, inhibiting FOXCUT markedly diminished the proliferative activities and invasive abilities of HT29 cells when overexpression of FOXC1 largely rescued these defects (Figure 2B, D, F and G; Supplementary Figures S2). Based on the findings above, we suggested that FOXC1 expression is regulated by the FOXCUT, and FOXCUT expression is necessary during the process of proliferation and invasion of HT29 cells.
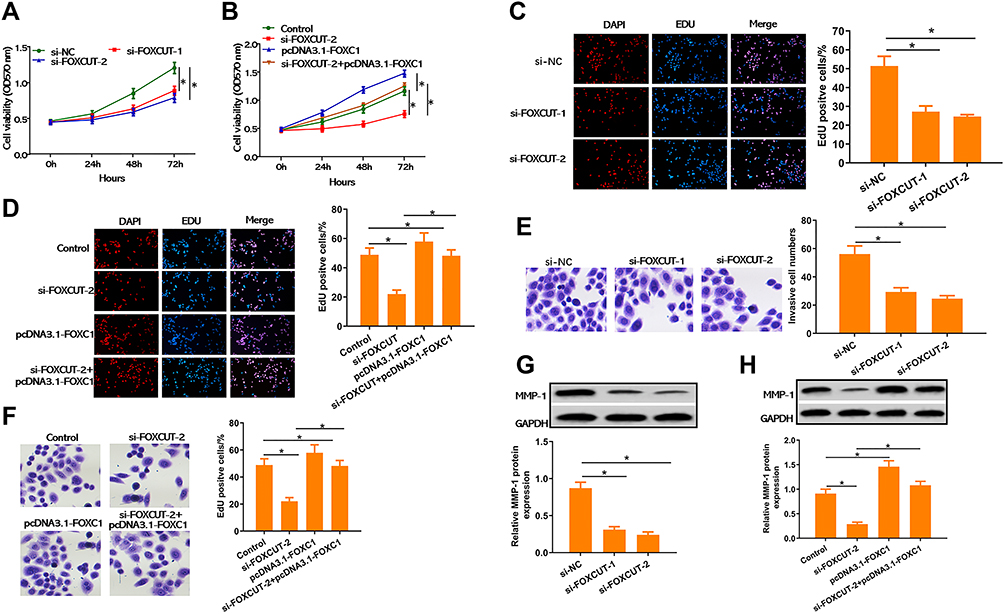 |
Figure 2 Knockdown of FOXCUT inhibited cell proliferation and invasion abilities in HT29. (A, B) Inhibition of FOXCUT reduces growth rate of cervical HT29 cells after treatment with or without si-FOXCUT or pcDNA3.1-FOXC1, determined by MTT assay. (C, D) Representative micrographs of the DAPI/EdU dyeing assay in HT29 cell after treatment with or without si-FOXCUT or pcDNA3.1-FOXC1. (E, F) Representative micrographs (left) and quantification (right) of transwell assay in HT29 cell after treatment with or without si-FOXCUT or pcDNA3.1-FOXC1. (G, H) Western blot and quantification analysis of MMP-1 after treatment with or without si-FOXCUT or pcDNA3.1-FOXC1, normalized to GAPDH. Data show the mean ± SD. * indicates significant differences compared with the control group (P < 0.05). |
Activated of PI3K/AKT Signaling Reversed FOXCUT Silencing Resulted in CRC Development Inhibition
Several studies reported that FOXCUT and FOXC1 through activated PI3K/AKT pathway to regulate proliferation and migration in cancer cells.5,29-31 IGF-1 is known to bind to the IGF-1 receptor (IGF-1R), which is one of the tyrosine kinase receptors that activate the PI3K/AKT signaling pathway upon stimulation. Thus, to further confirm the role of PI3K/AKT signaling during FOXCUT-mediated CRC development in HT29 cells, we measured the activation of key signal proteins, including PI3K and AKT. We found that FOXCUT silencing resulted in blockage of PI3K and AKT protein phosphorylation in HT-29 cells, while the inhibiting effect of FOXCUT silencing significantly diminished of HT29 when overexpression of FOXC1 or treatment with IGF-1 largely rescued these defects (Figure 3A). Consistent with this, MTT proliferation and transwell invasion assays showed that the IGF-1 treatment activated of PI3K/AKT signaling reversed FOXCUT silencing resulted in HT29 cells growth inhibition (Figure 3B and C). Taken together, these results suggest that the PI3K/AKT pathway is involved in FOXCUT-mediated CRC development in HT29 cells.
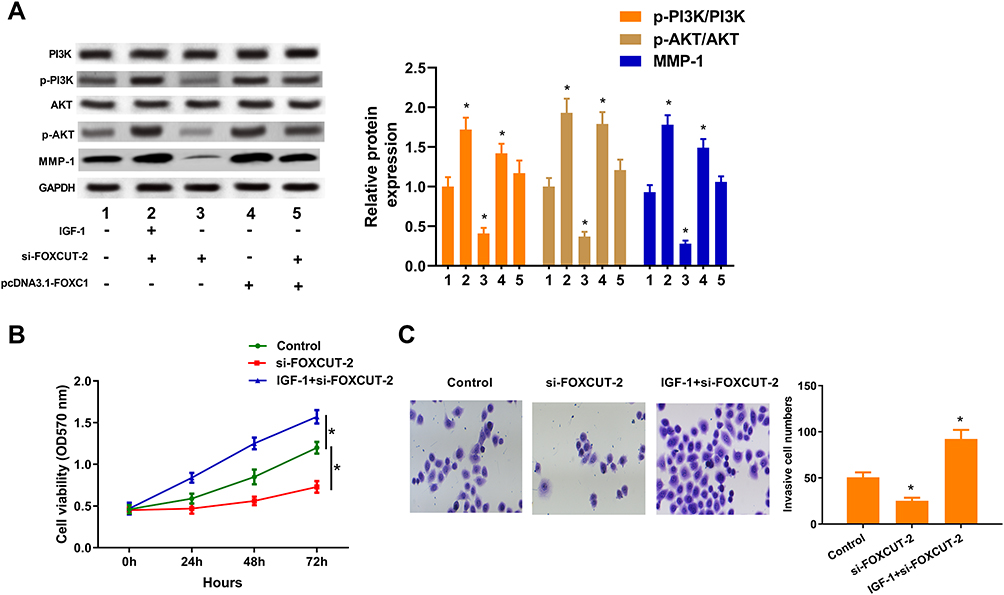 |
Figure 3 Activated of PI3K/AKT signaling reversed FOXCUT silencing resulted in CRC development inhibition. (A) Western blot analysis of PI3K, AKT and MMP-1 expression in HT29 cells after treatment with or without IGF-1 (200 ng/mL), si-FOXCUT or pcDNA3.1-FOXC1 for 48 h. GAPDH was used as a loading control. Right: quantification of p-PI3K/PI3K, p-AKT/AKT and MMP-1 protein levels, normalized to GAPDH. (B) Growth curves of HT29 cells after treatment with or without IGF-1 (200 ng/mL) or si-FOXCUT were determined via MTT assays. (C) Representative micrographs (left) and quantification (right) of transwell assay in HT29 cell after treatment with or without IGF-1 (200 ng/mL) or si-FOXCUT. Data show the mean ± SD. * indicates significant differences compared with the control group (P < 0.05). |
FOXCUT Expression Correlates with CRC Progression in vivo
Towards further defining FOXCUT is in relation to the progression of CRC, we confirmed by in vivo tumor growth assays. Consistent with in vitro results, xenograft tumors grown from FOXCUT silenced HT29 cells had smaller mean volumes and formed more slowly than xenograft tumors grown from control cells (Figure 4A–C). In addition, the protein expression of MMP-1 and FOXC1, the ratio of p-PI3K/PI3K, p-AKT/AKT both significantly decreased in sh-FOXCUT group. Altogether these results indicate that suppression of FOXCUT expression contributed to CRC cell growth inhibition.
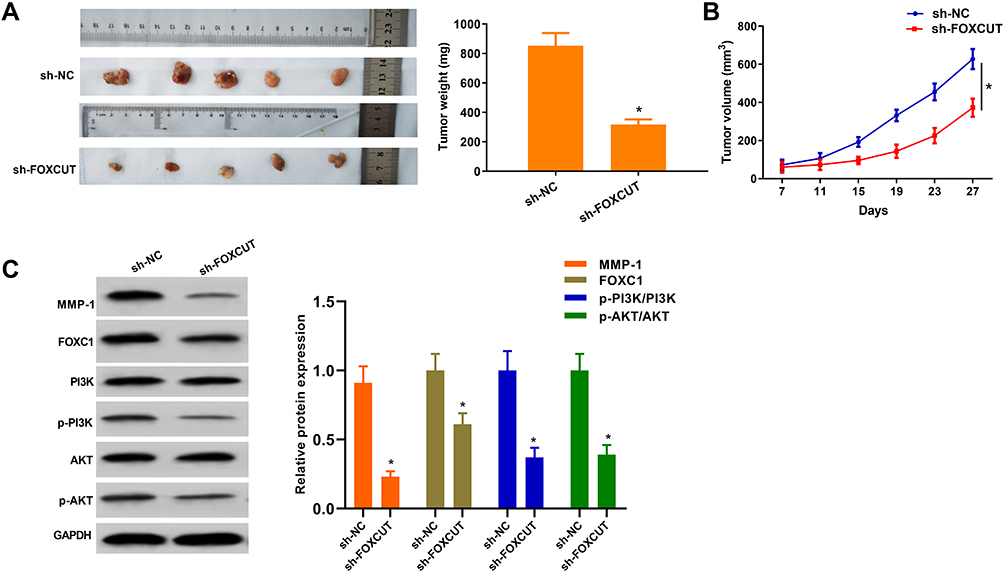 |
Figure 4 FOXCUT expression correlates with CRC progression in vivo. (A) Effects of FOXCUT knockdown on tumor growth after 4 weeks in vivo (n = 5 per group). Upper: negative control cells. Lower: representative images of tumors formed in nude mice subcutaneously injected with FOXCUT-silenced HT29 cells. (B) Growth curves of xenograft tumors after subcutaneous injection of mice with FOXCUT-silenced HT29 or negative control cells. The tumor volumes were measured every 4 days after inoculation. (n = 5). (C) Western blot analysis of MMP-1, FOXC1, PI3K and AKT expression in xenograft tumors after subcutaneous injection of mice with FOXCUT-silenced HT29. GAPDH was used as a loading control. The p-PI3K/PI3K, p-AKT/AKT, MMP-1/GAPDH and FOXC1/GAPDH ratios were calculated. *P < 0.05 compared with the sh-NC. All data are expressed as the mean ± SD. |
Discussion
lncRNAs are considered to be the basis for eukaryotes and essential for cancer progression.12,13,32 Although many lincRNAs have been identified recently, the functional characterization of lincRNAs has just begun. In addition, with the characteristic of non-invasive and humoral stability, lncRNA has become a sensitive biomarker for tumor diagnosis and prognosis. FOXCUT is a novel lncRNA located on upstream of the FOXC1 promoter. Previous studies indicated that the FOXCUT–FOXC1 regulatory network is related to tumorigenesis and cancer progression in breast cancer, oral squamous cell carcinoma and nasopharyngeal carcinoma.18,19,21 Nevertheless, the function of the FOXCUT and FOXC1 in CRC has not been determined. In our study, we explored the expression levels and clinicopathological features of the FOXCUT and FOXC1 in CRC and revealed the effects of FOXCUT in cell proliferation and migration associated with CRC to evaluate its feasibility as a potential therapeutic target.
FOXC1 initially was shown to be a key transcription factor that regulates the development of neural crest embryos.33 Recent studies indicated that FOXC1 was overexpressed in a wide variety of malignant cancers and it can promote tumorigenesis, and tumor progression.4,6-8 FOXC1 expression can be regulated at many levels, including modulation of DNA transcription, post-transcriptional regulation, and posttranslational modifications. lncRNAs exist as novel RNA transcripts, which function as high-level regulators that participated in post-transcriptional regulation.34 FOXCUT is a FOXC1 adjacent lncRNA that belongs to a class of ncRNAs called promoter upstream transcripts (PROMPTs).35 The expression and function of PROMPTs are oftenrelated to the adjacent protein-coding transcripts.21 Several studies reported that FOXCUT may be associated with the regulation of proliferation and invasion in CRC cell lines by increasing FOXC1 gene expression.18,19,21 In this study, we first investigated highly expression of FOXCUT and FOXC1 in clinical CRC tissues and CRC cell lines. In addition, the relative expression of FOXCUT was positively correlated with FOXC1 in the CRC cell lines and tissue samples. Besides, FOXCUT silencing reduced FOXC1 expression and abrogated CRC proliferation and invasion in vitro and in vivo. In contrast, overexpression of FOXC1 reversed FOXCUT silencing resulted in CRC development inhibition. These studies have shown that FOXCUT intensify cancer cell proliferation and invasion by promoting FOXC1 expression.
The PI3K/AKT signaling pathway plays important roles in the proliferation and their epithelial–mesenchymal transition (EMT) of CRC cells.36–39 In melanoma, FOXC1 promoted melanoma by activating MST1R/PI3K/AKT pathway, which is related to poor prognosis.30 It is also reported that FOXC1 promoted proliferation and epithelial–mesenchymal transition through the PI3K-AKT signal pathway in cervical carcinoma.29 However, FOXC1 inhibited migration and invasion in vitro and reduced the pulmonary metastasis in vivo.40 Besides, wang reported that lncRNA AB073614 regulated proliferation and metastasis via the PI3K/AKT signaling pathway in colorectal cancer cells.31 Our further study showed that FOXCUT up-regulated FOXC1 expression to activate PI3K/AKT signaling pathway. We observed that depleting FOXCUT expression decreased PI3K and AKT protein phosphorylation in vitro and in vivo, while the protein level of PI3K and AKT showed no change. In addition, EGF potently increases FOXC1 expression and phosphorylation in BLBC cells through Ras/ERK and phosphatidylinositol PI3K/AKT pathways.41 FOXC1 has also been involved in inflammation-related tumor metastasis. Proinflammatory cytokine interleukin-8 (IL-8) can activate FOXC1 via activation of hypoxia-inducible factor 1α (HIF-1α) by the PI3K/AKT pathway. These results also imply the feedback regulation between FOXC1 and PI3K/AKT pathway cells.42 Considering the complexity of the interaction between FOXC1 and PI3K/AKT, the roles of FOXCUT–FOXC1 in the cell proliferation and invasion deserve further study.
In summary, the present study demonstrated that high expressions of FOXCUT and FOXC1 in CRC. Our results also indicated that FOXCUT positively regulates the expression of FOXC1 and activates PI3K/AKT pathway to regulate cell proliferation and invasion in CRC, which is the first report of FOXCUT promotes CRC progression in vivo. We have identified that FOXCUT and FOXC1 might be acted as tumor promoter that provides potential diagnostic markers and therapeutic targets for CRC.
Abbreviations
BLBC, basal-like breast cancer; CRC, colorectal cancer; EMT, epithelial-to-mesenchymal transition; ESCC, esophageal cancer and gastric cancer; FOX, forkhead box; FOXCUT FOXC1, promoter upstream transcript; HIF-1α, hypoxia-inducible factor 1α; lncRNA, long noncoding RNA; NPC, nasopharyngeal carcinoma; OSCC, oral squamous cell carcinoma; PDA, pancreatic ductal adenocarcinoma; PVDF, polyvinylidene fluoride; siRNA, small interfering RNA; MMP-1, matrix metalloproteinase 1.
Ethical Statement
Our study was approved by the Ethics Review Board of Zhengzhou university. All experiments were performed following the guidelines and regulations of the Ethics Committee of the Zhengzhou University.
Author Contributions
(I) Conception and design: Lei Zhang, Xiaojie Zhang, and Shanyong Yi. (II) Administrative support: Lei Zhang, Xiaojie Zhang; (III) Provision of study materials or patients: All authors; (IV) Collection and assembly of data: Xiaojie Zhang and Shanyong Yi; (V) All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors have no conflicts of interest to declare.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–E86. doi:10.1002/ijc.29210
3. Zhang Y, Chen Z, Li J. The current status of treatment for colorectal cancer in China: a systematic review. Medicine (Baltimore). 2017;96(40):e8242. doi:10.1097/MD.0000000000008242
4. Ray PS, Wang JH, Qu Y, et al. FOXC1 is a potential prognostic biomarker with functional significance in basal-like breast cancer. Cancer Res. 2010;70(10):3870–3876. doi:10.1158/0008-5472.CAN-09-4120
5. Yang Z, Jiang S, Cheng Y, et al. FOXC1 in cancer development and therapy: deciphering its emerging and divergent roles. Ther Adv Med Oncol. 2017;9(12):797–816. doi:10.1177/1758834017742576
6. Wang L, Gu F, Liu C-Y, Wang R-J, Li J, Xu J-Y. Biology J-YXJT. High level of FOXC1 expression is associated with poor prognosis in pancreatic ductal adenocarcinoma. Tumour Biol. 2013;34(2):853–858. doi:10.1007/s13277-012-0617-7
7. Zhu X, Wei L, Bai Y, Wu S, Han S. FoxC1 promotes epithelial-mesenchymal transition through PBX1 dependent transactivation of ZEB2 in esophageal cancer. Am J Cancer Res. 2017;7(8):1642–1653.
8. Xu Y, Shao Q-S, Yao H-B, Jin Y, Ma -Y-Y, Jia,L-H. Overexpression of FOXC1 correlates with poor prognosis in gastric cancer patients. Histopathology. 2014;64(7):963–970. doi:10.1111/his.12347
9. Somerville TD, Wiseman DH, Spencer GJ, et al. Frequent derepression of the mesenchymal transcription factor gene FOXC1 in acute myeloid leukemia. Cancer Cell. 2015;28(3):329–342. doi:10.1016/j.ccell.2015.07.017
10. Han B, Bhowmick N, Qu Y, Chung S, Giuliano AE, Cui X. FOXC1: an emerging marker and therapeutic target for cancer. Oncogene. 2017;36(28):3957–3963. doi:10.1038/onc.2017.48
11. Zhang Y, Lai MD. Pathology MDLJZblxzzCjo. Advances of FOXC1 transcription factor in cancer. Zhonghua Bing Li Xue Za Zhi. 2017;46(5):362–365. doi:10.3760/cma.j.issn.0529-5807.2017.05.022
12. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. doi:10.1038/nrg2521
13. Batista PJ, Chang HY. Long noncoding RNAs: cellular address codes in development and disease. Cell. 2013;152(6):1298–1307. doi:10.1016/j.cell.2013.02.012
14. Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–914. doi:10.1016/j.molcel.2011.08.018
15. Anko ML, Neugebauer KM. Long noncoding RNAs add another layer to pre-mRNA splicing regulation. Mol Cell. 2010;39(6):833–834. doi:10.1016/j.molcel.2010.09.003
16. Yoon JH, Abdelmohsen K, Srikantan S, et al. LincRNA-p21 suppresses target mRNA translation. Mol Cell. 2012;47(4):648–655. doi:10.1016/j.molcel.2012.06.027
17. Gong C, Maquat LE. lncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3′ UTRs via Alu elements. Nature. 2011;470(7333):284–288. doi:10.1038/nature09701
18. Xu YZ, Chen FF, Zhang Y, et al. The long noncoding RNA FOXCUT promotes proliferation and migration by targeting FOXC1 in nasopharyngeal carcinoma. Tumour Biol. 2017;39(6):101042831770605. doi:10.1177/1010428317706054
19. Liu J, Shen L, Yao J, et al. Forkhead box C1 promoter upstream transcript, a novel long non-coding RNA, regulates proliferation and migration in basal-like breast cancer. Mol Med Rep. 2015;11(4):
20. Chen Y, Zhang C, Lu N. A novel long non-coding RNA FOXCUT and mRNA FOXC1 pair promote progression and predict poor prognosis in esophageal squamous cell carcinoma. Int J Clin Exp Pathol. 2014;7(9):
21. Kong X-P, Yao J, Luo W, et al. The expression and functional role of a FOXC1 related mRNA-lncRNA pair in oral squamous cell carcinoma. Mol Cell Biochem. 2014;394(1–2):177–186. doi:10.1007/s11010-014-2093-4
22. Li TT, Zhu D, Mou T, et al. IL-37 induces autophagy in hepatocellular carcinoma cells by inhibiting the PI3K/AKT/mTOR pathway. Mol Immunol. 2017;87:132–140. doi:10.1016/j.molimm.2017.04.010
23. Liu ZQ, Zhao S, Fu WQ. Insulin-like growth factor 1 antagonizes lumbar disc degeneration through enhanced autophagy. Am J Transl Res. 2016;8(10):4346–4353.
24. Whelan JA, Russell NB, Whelan MA. A method for the absolute quantification of cDNA using real-time PCR. J Immunol Methods. 2003;278(1–2):261–269. doi:10.1016/S0022-1759(03)00223-0
25. Fang Z, Zhang L, Liao Q, et al. Regulation of TRIM24 by miR-511 modulates cell proliferation in gastric cancer. J Exp Clin Cancer Res. 2017;36(1):17. doi:10.1186/s13046-017-0489-1
26. Hettiaratchi A, Hawkins NJ, McKenzie G, et al. The collagenase-1 (MMP-1) gene promoter polymorphism - 1607/2G is associated with favourable prognosis in patients with colorectal cancer. Br J Cancer. 2007;96(5):783–792. doi:10.1038/sj.bjc.6603630
27. Zugowski C, Lieder F, Muller A, et al. STAT3 controls matrix metalloproteinase-1 expression in colon carcinoma cells by both direct and AP-1-mediated interaction with the MMP-1 promoter. Biol Chem. 2011;392(5):449–459. doi:10.1515/bc.2011.038
28. Tahara K, Mimori K, Iinuma H, et al. Serum matrix-metalloproteinase-1 is a bona fide prognostic marker for colorectal cancer. Ann Surg Oncol. 2010;17(12):3362–3369. doi:10.1245/s10434-010-1149-2
29. Huang L, Huang Z, Fan Y, He L, Li Q. FOXC1 promotes proliferation and epithelial-mesenchymal transition in cervical carcinoma through the PI3K-AKT signal pathway. Am J Transl Res. 2017;9(3):1297–1306.
30. Wang J, Li L, Liu S, Zhao Y, Du GJO. FOXC1 promotes melanoma by activating MST1R/PI3K/AKT. Oncotarget. 2016;7(51):
31. Wang Y, Kuang H, Xue J, Liao L, Yin F, Zhou X. LncRNA AB073614 regulates proliferation and metastasis of colorectal cancer cells via the PI3K/AKT signaling pathway. Biomed Pharmacother. 2017;93:1230–1237. doi:10.1016/j.biopha.2017.07.024
32. Glover AR, Zhao JT, Ip JC, et al. Long noncoding RNA profiles of adrenocortical cancer can be used to predict recurrence. Endocr Relat Cancer. 2015;22(1):99–109. doi:10.1530/ERC-14-0457
33. Kume T, Deng KY, Winfrey V, Gould DB, Hogan BLMJC, Hogan BLM. The forkhead/winged helix gene Mf1 Is disrupted in the pleiotropic mouse mutation congenital hydrocephalus. Cell. 1998;93(6):
34. Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016;17(1):47–62. doi:10.1038/nrg.2015.10
35. Pascal P, Kristina A, Christensen MS, et al. PROMoter uPstream transcripts share characteristics with mRNAs and are produced upstream of all three major types of mammalian promoters. Nucleic Acids Res. 2011;39(16):
36. Pandurangan AK. Potential targets for prevention of colorectal cancer: a focus on PI3K/Akt/mTOR and Wnt pathways. Asian Pac J Cancer Prev. 2013;14(4):
37. Johnson SM, Gulhati P, Rampy BA, et al. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J Am Coll Surg. 2010;210(767–76):76–78. doi:10.1016/j.jamcollsurg.2009.12.008
38. Bahrami A, Khazaei M, Hasanzadeh M, et al. Therapeutic potential of targeting PI3K/AKT pathway in treatment of colorectal cancer: rational and progress. J Cell Biochem. 2018;119(3):2460–2469. doi:10.1002/jcb.25950
39. Rodgers SJ, Ferguson DT, Mitchell CA, Ooms LM. Regulation of PI3K effector signalling in cancer by the phosphoinositide phosphatases. Biosci Rep. 2017;37(1). doi:10.1042/BSR20160432
40. Du J, Li L, Ou Z, et al. Erratum to: FOXC1, a target of polycomb, inhibits metastasis of breast cancer cells. Breast Cancer Res Treat. 2016;156(3):
41. Jin Y, Han B, Chen J, Wiedemeyer R, Cui X. FOXC1 is a critical mediator of EGFR function in human basal-like breast cancer. Ann Surg Oncol. 2014;21:758–766.
42. Huang W, Chen Z, Zhang L, et al. Interleukin-8 induces expression of FOXC1 to promote transactivation of CXCR1 and CCL2 in hepatocellular carcinoma cell lines and formation of metastases in mice. Gastroenterology. 2015;149(4):1053–67 e14. doi:10.1053/j.gastro.2015.05.058
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.