Back to Journals » International Journal of Nanomedicine » Volume 20
Extracellular Vesicles in Sarcoma: Implications for Tumor Progression and Therapy
Authors Wang Y ![]() , Gao S
, Gao S ![]() , Zhang G, Xiao W, Xu B
, Zhang G, Xiao W, Xu B ![]() , Li W
, Li W ![]()
Received 31 July 2025
Accepted for publication 17 October 2025
Published 14 November 2025 Volume 2025:20 Pages 13759—13779
DOI https://doi.org/10.2147/IJN.S557436
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kamakhya Misra
Yue Wang,1,2,* Shaozhi Gao,3,* Geng Zhang,3 Wei Xiao,3 Baoyiyu Xu,3 Wengang Li3
1Central Laboratory, Affiliated Hospital of Putian University, School of Basic Medicine, Putian University, Putian, Fujian Province, 351100, People’s Republic of China; 2Key Laboratory of Translational Tumor Medicine in Fujian Province, Putian University, Putian, Fujian Province, 351100, People’s Republic of China; 3Cancer Research Center, Xiang’an Hospital of Xiamen University, School of Medicine, Xiamen University, Xiamen, Fujian Province, 361005, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wengang Li, Email [email protected]
Abstract: Sarcoma, a heterogeneous group of malignancies arising from mesenchymal tissues, poses significant clinical challenges due to its aggressive behavior, early metastasis, and resistance to conventional therapies. Recent advancements highlight the pivotal role of extracellular vesicles (EVs)—nanoscale particles secreted by tumor and stromal cells—in modulating sarcoma progression through intercellular communication within the tumor microenvironment. This review comprehensively examines the multifaceted functions of EVs in sarcoma pathogenesis, focusing on their subtype-specific mechanisms (eg, osteosarcoma, liposarcoma) in promoting epithelial-mesenchymal transition (EMT), angiogenesis, and immune evasion. Additionally, we discuss the emerging potential of EVs as non-invasive biomarkers for early diagnosis and prognosis, as well as their engineered applications in targeted drug delivery and immunotherapy. By integrating cutting-edge research, this article underscores the translational value of EVs in overcoming current therapeutic limitations and provides a roadmap for future investigations into EV-based precision strategies for sarcoma management.
Keywords: extracellular vesicles, sarcoma, tumor microenvironment, biomarkers, therapy
Introduction
Sarcoma, a malignant tumor originating from mesenchymal tissues (eg, bone, cartilage, fat, muscle, and blood vessels), accounts for approximately 1% of adult malignancies and 15% of pediatric cancers.1 Based on histological origins, sarcomas are classified into subtypes such as osteosarcoma, liposarcoma, rhabdomyosarcoma, Kaposi’s sarcoma, and Ewing sarcoma, exhibiting complex biological behaviors and significant heterogeneity.2 Despite surgical resection combined with radiotherapy and chemotherapy remaining the primary treatment modalities, the high invasiveness, early metastatic propensity, and chemotherapy resistance of sarcomas have led to stagnant 5-year survival rates, particularly in patients with advanced or metastatic disease.3 Furthermore, the toxicity of conventional therapies to normal tissues and postoperative recurrence underscore the urgent need for novel precision therapies.
In recent years, the role of the tumor microenvironment (TME) in sarcoma progression has emerged as a research focus. As key mediators of intercellular communication within the TME, extracellular vesicles (EVs)—carrying bioactive molecules such as proteins, nucleic acids, and lipids—have been identified as critical regulators of tumor growth, metastasis, angiogenesis, and immune evasion.4 Studies demonstrate that EVs secreted by sarcoma cells can remodel stromal cell functions by transferring oncogenic miRNAs (eg, miR-21, miR-135b), thereby promoting tumor invasion.5,6 Simultaneously, EV-specific surface markers (eg, CD63, CD9) offer potential non-invasive biomarkers for sarcoma diagnosis and prognosis.7,8 Additionally, engineered EVs loaded with chemotherapeutic agents or targeting molecules exhibit promise as “natural nanocarriers” to overcome tumor barriers and enhance therapeutic precision.9 However, the precise mechanisms of EVs in sarcomas, subtype-specific functions, and clinical translation remain largely unresolved.
The evolving landscape of EV research in sarcoma therapy has witnessed several key milestones over the past decades (Figure 1). Extracellular vesicles were first described as early as 1967, referred to as “platelet dust” and linked to physiological processes such as bone mineralization and coagulation.10 Follow-up research in the 1990s revealed their significant regulatory roles within the immune system.11 Research specific to sarcomas began to emerge in the late 2000s, with initial studies focusing on the basic characterization of sarcoma-derived EVs and the identification of exosome-associated proteins in secretory factors from murine dendritic cell sarcoma (DCS).12 By the 2010s, investigations expanded to uncover the functional roles of EVs in sarcoma metastasis, drug resistance, and immune evasion. For example, seminal studies around 2011 demonstrated that EVs from osteosarcoma cells could promote pulmonary metastasis via miRNA transfer.13 During the latter half of the 2010s, EV-based biomarkers—such as miR-25-3p, PD-L1, and N-cadherin—were identified as promising diagnostic and prognostic tools in sarcoma.14,15 Most recently, from 2020 onward, the field has increasingly shifted toward therapeutic applications, including the development of engineered EVs as drug delivery systems and EV-based vaccines, paving the way for future clinical trials.16,17
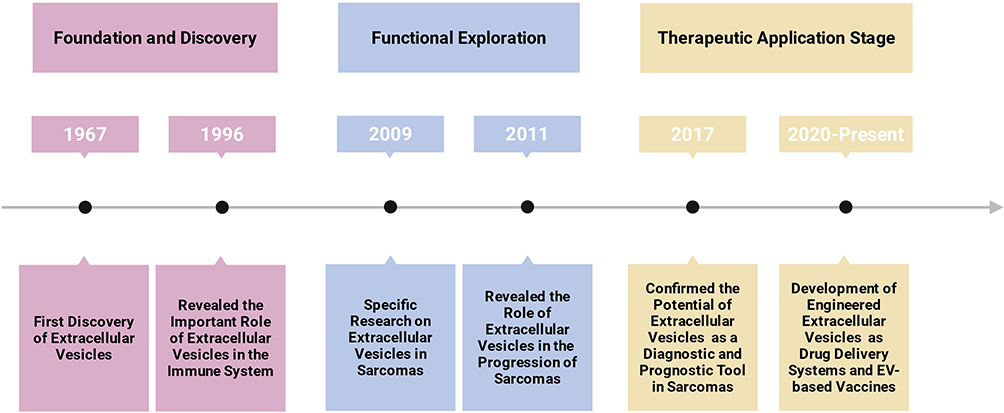 |
Figure 1 Timeline of key milestones in extracellular vesicle research for sarcoma. The schematic illustrates the evolution from the initial discovery of EVs to their current application in sarcoma therapy, highlighting pivotal discoveries in biological mechanisms, diagnostic biomarker development, and the advent of engineered EV therapeutics. Created with BioRender.com. Abbreviation: EV, Extracellular vesicle. |
This review systematically summarizes the multifaceted roles of EVs in sarcoma pathogenesis, with a focus on molecular mechanisms in subtypes such as osteosarcoma and liposarcoma (eg, EMT regulation, angiogenesis activation). It also critically evaluates recent advances in EV-based diagnostic strategies and engineered therapeutic applications. By integrating cutting-edge research, this article aims to provide a comprehensive perspective on the complex EV network in sarcomas and establish a theoretical foundation for developing EV-centric precision diagnostic and therapeutic approaches.
Extracellular Vesicles
Extracellular vesicles (EVs) are heterogeneous membrane-bound particles secreted by diverse cell types, serving as critical mediators of intercellular communication by transporting functional cargos—including nucleic acids, proteins, and lipids—to recipient cells, thereby modulating physiological and pathological processes.18,19
Based on their biogenesis and cellular origin, EVs are classified into three major subtypes: exosomes (40–100 nm in diameter), which originate from the fusion of multivesicular bodies (MVBs) with the plasma membrane and are marked by tetraspanins, Alix, and TSG101; microvesicles (100–1000 nm), formed by outward budding of the plasma membrane and regulated by calcium-dependent enzymes and ARF6 signaling; and apoptotic bodies (50–5000 nm), released during late apoptosis and characterized by DNA-histone complexes.20–24 Exosome biogenesis involves the endosomal sorting complex required for transport (ESCRT), which regulates ubiquitinated protein sorting and intraluminal vesicle (ILV) formation, though alternative pathways, such as plasma membrane budding in T cells, have also been proposed.25–30 Microvesicle generation, in contrast, relies on calcium-triggered scramblase activation and ARF6-mediated molecular loading.31–33 This structural and mechanistic diversity underpins the functional versatility of EVs in physiological and pathological contexts.
EVs exert their biological roles through the selective transfer of molecular cargo. They encapsulate nucleic acids (eg, miRNA, mRNA, mtDNA), proteins (eg, kinases, integrins, Rab GTPases), and lipids, which are shielded from degradation and delivered to target cells.34–39 Notably, EVs carry oncogenic molecules such as PD-L1 and tumor-promoting receptors, enabling immune evasion and angiogenesis in cancer.40,41 Their protein cargo also includes metabolic enzymes (eg, GAPDH), cytoskeletal components (eg, actin), and chaperones, which regulate cellular homeostasis and survival.35–37 Crucially, the functional impact of EVs depends on their origin—such as tumor-derived “oncosomes” or platelet-derived EVs—and the physiological state of parent cells, positioning them as dynamic biomarkers and therapeutic targets in diseases like cancer.21,22
To unravel these functional roles, robust isolation methods are essential. Current techniques prioritize balancing purity, yield, and scalability. Differential ultracentrifugation (dUC), the traditional gold standard, separates EVs based on size and density but risks aggregation or membrane damage.42–44 Refinements like density gradient ultracentrifugation (DGUC) improve specificity by leveraging buoyant density differences, while size-exclusion chromatography (SEC) offers a gentler, non-destructive alternative.45–47
Tangential flow filtration (TFF) has gained attention as a scalable and efficient method that uses recirculating flow across a membrane to separate EVs based on size while minimizing fouling and preserving vesicle integrity. TFF is particularly advantageous for processing large volumes, such as cell culture supernatants or plasma, and offers high yield and reproducibility, making it suitable for both research and clinical-scale applications.48,49 However, it may require additional purification steps to achieve high purity due to potential co-isolation of similarly sized particles.49
Immunoaffinity-based methods, such as antibody-coated magnetic beads, achieve superior purity by targeting surface markers (eg, CD63)—a strategy directly informed by EVs classification studies—but are limited by the lack of universal markers and low recovery rate.50–52 Emerging technologies, like Lucia Casadei’s microfluidic device combining size-based filtration and immunoaffinity capture, show promise for clinical applications but require optimization for complex biofluids.53 Together, these evolving methodologies highlight the interplay between EVs biology and technological innovation, driving advancements in both basic research and translational medicine.
The Role of Extracellular Vesicles in Tumor
Extracellular vesicles (EVs) exhibit spatiotemporal heterogeneity driven by pathological and microenvironmental cues, with cancer patients showing a 2-fold elevation in circulating EVs (~4000 trillion) compared to healthy individuals (~2000 trillion).54 However, not all cancer types show consistently elevated levels of circulating EVs, a discrepancy attributed to substantial inter-cancer heterogeneity in EVs biogenesis and secretion mechanisms, as well as technical variations in EVs isolation and characterization methods.55–57 For instance, studies employing different isolation techniques (ultracentrifugation vs precipitation-based kits) have reported conflicting results regarding EVs abundance in certain malignancies like prostate and ovarian cancer.58,59 This methodological dependency underscores the need for standardized protocols in EVs research.
Tumor extracellular matrix (ECM) stiffness amplifies EV secretion via mechanosensitive pathways: Akt/Rab8 activation in breast cancer and YAP/TAZ-THBS1 signaling in other malignancies coordinate matrix metalloproteinase-dependent EV release.60,61 EV biodistribution favors liver, spleen, and lungs, governed by membrane composition (eg, CD44 in gastric/pancreatic cancers) that mediates metastasis and immune evasion.62–64 Hypoxia and therapy-induced stress further enhance EV secretion, exemplified by irradiated esophageal cancer EVs conferring radioresistance via cyclin B1/CDK1-mediated cell cycle arrest and PI3K/AKT/FOXO3A survival signaling.65,66
EVs orchestrate tumor-stroma crosstalk to initiate and sustain oncogenesis. Prostate and ovarian cancer-derived EVs convert fibroblasts into cancer-associated fibroblasts (CAFs), with aggressive variants (eg, SKOV3 EVs) exhibiting superior CAF-activating capacity.67–69 Activated CAFs reciprocally secrete EVs that drive metabolic remodeling and proliferation, establishing a feedforward tumorigenic loop.69 Breast cancer EVs deliver Dicer, an miRNA-processing enzyme, and CRISPR-mediated Dicer silencing in EVs attenuates tumor growth, directly linking EV cargo to oncogenic transformation.70 Leukemia-derived EVs activate NF-κB in macrophages, upregulating IL-8 to enhance gastric cancer invasiveness, while pharmacological NF-κB inhibition reverses these effects.71 Chronic myelogenous leukemia (CML) EVs stimulate bone marrow stromal cells to secrete IL-8, activating CXCR1/CXCR2 receptors on leukemia cells to perpetuate disease progression.72–74
Metastatic is critically regulated by EVs-mediated premetastatic niche formation. Melanoma-derived EVs prime sentinel lymph nodes through extracellular matrix remodeling and secretion of angiogenic factors such as VEGF, thereby establishing a permissive microenvironment for tumor cell colonization.75,76 In breast cancer, CD81-enriched EVs secreted by cancer-associated fibroblasts activate Wnt-planar cell polarity (PCP) signaling in malignant cells, enhancing metastatic spread.77 Horizontal transfer of metastatic potential is exemplified by Cre-LoxP tracking studies: highly aggressive tumor cells deliver Cre mRNA via EVs to less malignant counterparts, imparting migratory and invasive capabilities detectable through intravital imaging.78 Furthermore, colorectal cancer EVs drive persistent stromal reprogramming of mesenchymal stem cells (MSCs), characterized by sustained proliferation, enhanced migratory activity, and heightened invasive capacity—effects that endure even following EV withdrawal.79
Emerging evidence highlights EVs’ dual roles as tumor promoters and potential therapeutic vehicles. While PD-L1+ breast cancer EVs propagate immune evasion,80 senescent stromal EVs upregulate ABCB4 to drive chemoresistance.81,82 Conversely, EVs may deliver immunostimulatory cargo or be engineered (eg, CD44-targeted membranes, CRISPR-edited miRNAs) for precision therapy.83 Their functional duality, shaped by donor cell status and microenvironmental stressors, underscores the need for single-EV analyses to resolve context-specific roles.
The Functions and Mechanisms of Extracellular Vesicles in Sarcoma
Sarcoma, a highly heterogeneous malignancy originating from mesenchymal tissues, exhibits remarkable diversity in both biological behaviors and molecular profiles. This heterogeneity is reflected not only in complex driver genetic alterations—such as chromosomal translocations, fusion protein expression, and epigenetic dysregulation—but also in starkly contrasting clinical phenotypes, ranging from locally indolent growth to early systemic metastasis, and from chemosensitivity to therapy-resistant recurrence, with prognosis spanning extremes. For instance, certain sarcoma subtypes are defined by specific molecular events (eg, EWSR1-FLI1 fusion)84–86 that dictate malignant transformation pathways, while others rely on polygenic cooperative mutations (eg, TP53/RB1 mutations in osteosarcoma)87–90 or viral oncogenic factors (eg, HHV-8 in Kaposi sarcoma)91–93 to drive progression. Such diversity further extends to the dynamic interplay within the tumor microenvironment, exemplified by the functional divergence of extracellular vesicles (EVs): some sarcomas utilize EVs to deliver oncogenic non-coding RNAs to remodel distant organ microenvironments,87–90 whereas others exploit EV surface immune checkpoint molecules to evade immune surveillance, thereby promoting sarcoma progression and metastasis (Figure 2).91–94 In rhabdomyosarcoma, particularly the pleomorphic subtype (PRMS), EVs may contribute to early metastatic spread and poor prognosis through complex molecular events.95,96 The subtype-specific molecular hallmarks and clinical behaviors of sarcomas are summarized in Table 1, along with a preliminary outline of their distinct EV-mediated mechanisms. The following sections (4.1–4.5) will focus on the subtype-specific mechanisms of EVs in sarcomas.
 |
Table 1 Classification, Key Features and EVs-Mediated Mechanisms of Major Sarcoma Subtype |
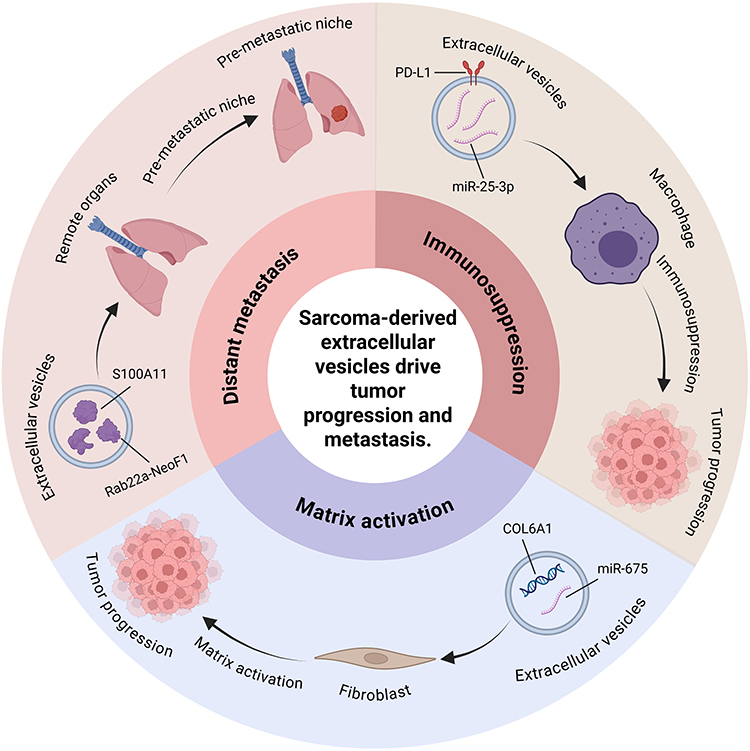 |
Figure 2 Roles of sarcoma-derived extracellular vesicles in tumor microenvironment, cancer progression and metastasis. Sarcoma-derived extracellular vesicles carry specific molecules, such as miR-675, S100A11, PD-L1, and miR-25-3p, which enable immune evasion, activate fibroblasts to remodel the tumor microenvironment, and collectively promote immunosuppression, matrix activation, and distant metastasis. Created with BioRender.com. |
Extracellular Vesicles in Liposarcoma
Liposarcoma (LPS), the most prevalent histological subtype of soft tissue sarcomas, comprises four distinct entities: well-differentiated (WDLPS), dedifferentiated (DDLPS), myxoid/round cell (MRC), and pleomorphic LPS. Among these, DDLPS exhibits particularly aggressive clinical behavior characterized by frequent local recurrence and distant metastasis.94 Emerging research highlights the pivotal role of extracellular vesicles (EVs) in mediating the interplay between liposarcoma cells and their microenvironment, with recent studies focusing on two key mechanisms: oncogenic miRNA-mediated crosstalk and systemic delivery of tumor-promoting cargo.
A hallmark of LPS pathogenesis is the secretion of EVs enriched with miR-25-3p and miR-92a-3p, which orchestrate tumor progression through dual microenvironmental modulation. These exosomes are selectively packaged by LPS cells and internalized by tumor-associated macrophages (TAMs), triggering TLR7/8-dependent activation that drives IL-6 secretion.97,109 The resulting pro-inflammatory cascade not only enhances cancer cell proliferation and invasion but also establishes a feedforward loop sustaining malignant transformation of adipose tissue.97 Concurrently, miR-25-3p exerts additional oncogenic effects by suppressing preadipocyte differentiation via NF-κB pathway inhibition and promoting angiogenesis through vascular permeability alterations, thereby remodeling the stromal landscape to favor tumor growth.110,111 Notably, the stability and tumor-specific enrichment of these exosomal miRNAs position them as promising liquid biopsy biomarkers for detecting subclinical recurrence before radiological progression.97
The microenvironmental reprogramming capacity of LPS-derived EVs is further exemplified in DDLPS, where exosomes deliver oncogenic MDM2 DNA and other molecular cargo to adjacent stromal cells.98 Upon internalization by preadipocytes, these vesicles induce malignant transformation through coordinated p53 pathway suppression and MMP2 upregulation, effectively enhancing recipient cell proliferation and migratory potential.98 This systemic delivery mechanism underscores the dual role of DDLPS-EVs as both local signaling mediators and systemic metastasis facilitators.
Collectively, these findings establish that liposarcoma-derived extracellular vesicles (EVs) function as pleiotropic regulators of tumor progression, exerting their effects via miRNA-mediated immunomodulation, stromal remodeling, and systemic dissemination of oncogenic cargo. The integration of these pathways underscores EV-driven intercellular communication as a pivotal therapeutic vulnerability in liposarcoma, with heightened relevance in the clinically aggressive DDLPS subtype.
Extracellular Vesicles in Ewing Sarcoma
Ewing sarcoma (EWS), the second most common malignant bone tumor in children and adolescents, is molecularly defined by the EWSR1-FLI1 fusion oncogene resulting from the t(11;22)(q24;q12) translocation in 95% of cases.84–86 Beyond its cell-autonomous oncogenic effects, emerging evidence highlights extracellular vesicles (EVs) as critical mediators of tumor progression through intercellular communication and microenvironmental reprogramming.
EWS-derived EVs carry unique molecular cargo, including the EWS-FLI1 transcript itself, which transfers oncogenic signals specifically between EWS cells to amplify tumorigenicity.84 Functional studies reveal these EVs are enriched in neurotransmitter signaling components and G-protein-coupled receptor pathway mRNAs, suggesting their role in sustaining a pro-tumorigenic state.99 Notably, CD99—a surface marker pivotal in EWS differentiation—modulates EV composition in a bidirectional manner.100,112 CD99-positive EVs deliver oncogenic miRNAs to drive tumor growth, while CD99-negative EVs exhibit elevated miR-34a and miR-199a-3p, which suppress NF-κB/AP-1 signaling to inhibit metastasis but paradoxically enhance neural differentiation and chemoresistance.100,112,113 This duality underscores EVs as context-dependent regulators of tumor plasticity.
The immunomodulatory role of EWS-EVs further complicates disease progression. Recent studies demonstrate that EV-packaged retrotransposon RNAs (eg, HSAT2/3) activate cGAS-TBK1 innate immune signaling, inducing proinflammatory cytokine release while simultaneously suppressing dendritic cell maturation markers (CD80/CD86) and T-cell effector functions.101,114 This creates an immunosuppressive niche conducive to metastasis. Similarly, EV-mediated transfer of the lncRNA FOXP4-AS1 correlates with adverse clinical outcomes by reshaping the tumor immune microenvironment.115 These findings align with observations that EWS-EVs reprogram stromal fibroblasts into cancer-associated fibroblasts (CAFs) and expand myeloid-derived suppressor cells, collectively establishing a pro-metastatic ecosystem.116
Metastatic processes are further amplified by EV-driven heterogeneity. For instance, IGF2BP3—an RNA-binding protein selectively packaged into EWS-EVs—enhances migratory capacity by modulating PI3K/Akt signaling through miRNA redistribution.117 Clinically, elevated IGF2BP3 levels predict poor prognosis, emphasizing its role in EV-mediated metastatic spread.117 Together, these mechanisms illustrate how EWS-EVs orchestrate a complex crosstalk network, balancing oncogenic signaling, immune evasion, and stromal activation to fuel disease progression.
Extracellular Vesicles in Osteosarcoma
Osteosarcoma (OS), a malignant bone tumor arising from primitive mesenchymal cells, exhibits aggressive progression marked by early pulmonary metastasis, which historically accounted for over 90% of mortality prior to chemotherapy advancements.87–90 Extracellular vesicles (EVs) have emerged as central mediators of OS pathogenesis, dynamically coordinating tumor-stroma crosstalk to drive malignancy. OS-derived EVs transport functionally active miRNAs such as miR-21-5p and miR-148-5p, which suppress apoptosis-associated genes, while miR-675 enhances fibroblast invasiveness via CALN1 downregulation, collectively amplifying tumorigenic signaling.6,102,118 These EV cargoes exhibit metastatic selectivity, as evidenced by elevated miR-25-3p and miR-148a in circulating EVs from OS patients, whose levels correlate with tumor growth kinetics.14,119,120 Complementing miRNA-driven mechanisms, EVs transfer oncoproteins like urokinase plasminogen activator (uPA/uPAR), whose expression parallels pulmonary metastatic propensity.121 Beyond direct tumor modulation, OS-EVs remodel the tumor microenvironment (TME) through dual stromal reprogramming: COL6A1-enriched exosomes induce fibroblast-to-CAF transformation via IL-6/IL-7 secretion, activating TGF-β/COL6A1 cascades to enhance invasiveness,122 while H3K27ac-activated COL6A1 simultaneously suppresses STAT-mediated immunity and primes TGF-β-dependent CAFs to establish immune-evasive metastatic niches.122
The immunosuppressive effects of EVs further synergize with metastatic progression. Chemotherapy itself paradoxically fuels immune evasion, as doxorubicin-treated OS cells release EVs that activate IL-1β signaling to upregulate PD-L1, creating a checkpoint-enriched immunosuppressive milieu.103 Concurrently, EVs orchestrate pre-metastatic niche formation through myeloid cell hijacking: S100A11-containing EVs recruit granulocytic MDSCs to lungs,5 while Rab22a-NeoF1 fusion protein-loaded exosomes promote macrophage-dependent niche preparation via HSP90-dependent sorting, activating RhoA/STAT3 pathways to license metastasis.123 This immune subversion is compounded by EV-mediated macrophage polarization, where Tim-3 or CD47 interactions skew macrophages toward pro-tumorigenic M2 phenotypes, effectively silencing antitumor M1 responses.124,125 Counteracting these mechanisms, therapeutic EVs from mesenchymal stem cells deliver miR-22 to suppress Twist1/CADM1 signaling, demonstrating potential to reverse tumor invasiveness.126
Within bone ecosystems, EVs exert context-dependent regulation of osteolytic and osteogenic processes. OS-derived exosomes armed with miR-501-3p exacerbate bone destruction by enhancing RANKL/NFATc1-driven osteoclastogenesis through PTEN suppression,127 while MNNG/HOS cell-derived EVs further impair bone integrity via anti-osteoblastogenic and pro-angiogenic effects.128 In contrast, osteoblast-derived exosomes counteract tumor growth by modulating the URG4/WIF1-Wnt axis to promote OS cell differentiation, offering a paradigm for differentiation therapy.129 This duality extends to chemoresistance mechanisms, where EVs act as molecular couriers for adaptive survival signals. Doxorubicin-resistant OS cells disseminate miR-143-3p via exosomes to confer chemoresistance in sensitive cells, with elevated exosomal levels predicting poor therapeutic outcomes.104 Similarly, BMSC-EVs transport lncRNA NORAD to enhance OS aggressiveness through miR-30c-5p sponging,130 while miR-144-3p-enriched EVs inhibit ferroptosis via ZEB1 targeting, creating a pro-survival niche.131 Strategies targeting EV biogenesis show therapeutic promise, as IRF5-mediated suppression of EV secretion disrupts pre-metastatic niche formation and correlates with improved patient survival.132
Extracellular Vesicles in Rhabdomyosarcoma
Rhabdomyosarcoma (RMS), the most common pediatric soft tissue sarcoma, presents as two biologically distinct subtypes: embryonal (ERMS) and alveolar (ARMS), which differ markedly in clinical presentation and prognosis. Notably, the PAX3-FOXO1 fusion oncoprotein serves as a critical prognostic determinant, with its presence strongly correlating with unfavorable outcomes. While multimodal therapy achieves good responses in localized disease, metastatic RMS remains therapeutic recalcitrant a clinical challenge that underscores the urgent need to understand its molecular drivers.95,96
Emerging evidence implicates extracellular vesicles (EVs) as pivotal mediators of RMS progression through bidirectional tumor-stroma communication. Proteomic profiling of EVs from five RMS cell lines revealed 80 unique protein components, including previously unrecognized molecules like BMP1, CDKN2A, and ITGA7 that functionally converge on pro-metastatic processes such as tumor cell proliferation and invasion.133 This molecular repertoire is complemented by specific microRNA cargo: both ERMS and ARMS-derived EVs consistently carry miR-1246 and miR-1268, which directly induce migratory and invasive phenotypes in stromal fibroblasts and endothelial cells.105
Transmembrane proteins play critical roles in EV-mediated pathological processes in rhabdomyosarcoma (RMS). CD147, which is highly expressed in RMS cells and their secreted exosomes, promotes tumor cell invasion through autocrine signaling. Additionally, CD147 facilitates stromal fibroblast activation via exosome-mediated paracrine communication, as evidenced by studies showing that CD147 knockdown reduces RMS cell invasion by over 50% in vitro.106 Recent studies further identified CD147 as another transmembrane glycoprotein overexpressed in metastatic RMS. Exosomes carrying CD147 enhance tumor invasiveness by inducing fibroblast proliferation, migration, and invasion through intercellular signaling.106 The coordinated actions of CD147 through EV-mediated pathways underscore the importance of transmembrane proteins in regulating stromal interactions during RMS progression.
Extracellular Vesicles in Kaposi’s Sarcoma
Kaposi sarcoma (KS), an angioproliferative malignancy driven by human herpesvirus 8 (HHV-8/KSHV) infection, is characterized by spindle cell proliferation and aberrant vascular formation.91,92 While classically presenting as cutaneous lesions, it exhibits visceral involvement in immunocompromised hosts such as HIV/AIDS patients, underscoring its dependence on immune dysregulation for progression.93 Emerging insights into extracellular vesicle (EV)-mediated intercellular communication reveal their multifaceted roles in KSHV pathogenesis through viral persistence and immune modulation.
Central to this process is the KSHV-induced hyperproduction of EVs, mechanistically linked to viral upregulation of Rab27b—a key regulator of exosome biogenesis.134 This EV hypersecretion not only facilitates viral miRNA transfer but also delivers pro-survival factors to sustain infected cell viability and promote persistent infection.107 Paradoxically, while KSHV+ EVs lack direct infectivity, they orchestrate profound microenvironmental remodeling. Notably, excessive mitochondrial DNA (mtDNA) expulsion via these EVs depletes intracellular mtDNA reservoirs, impairing mitochondrial antiviral signaling (MAVS) pathways and compromising host defense mechanisms.134 Concurrently, proteomic analyses reveal EV-mediated trafficking of viral lytic proteins that enhance endothelial cell motility while suppressing adhesion—a functional reprogramming that drives angiogenic dysregulation characteristic of KS lesions.135
The immunomodulatory payload of KSHV+ EVs further exemplifies viral hijacking of host machinery. BCBL-1-derived EVs carry IFI16, an inflammasome sensor paradoxically repurposed to cleave IL-1β, thereby subverting IL-1β-mediated inflammatory responses and establishing immune-tolerant niches.108 This immune evasion is synergized by KSHV-encoded viral miRNAs that reprogram recipient cell transcriptomes.134 Intriguingly, B cell-derived EVs from infected hosts exhibit selective enrichment of glycolytic enzymes, reflecting KSHV’s metabolic rewiring of host cells toward aerobic glycolysis—a hallmark of viral oncogenesis.136
Collectively, extracellular vesicles (EVs) deliver diverse molecular cargoes across sarcoma subtypes, modulating recipient cell phenotypes and enhancing tumor aggressiveness through multifaceted mechanisms (Figure 3). A comparative analysis further reveals that although EVs carry distinct cargoes in different sarcoma subtypes, they exhibit significant convergence in the pathogenic mechanisms (as summarized in Figure 3 and Table 1):
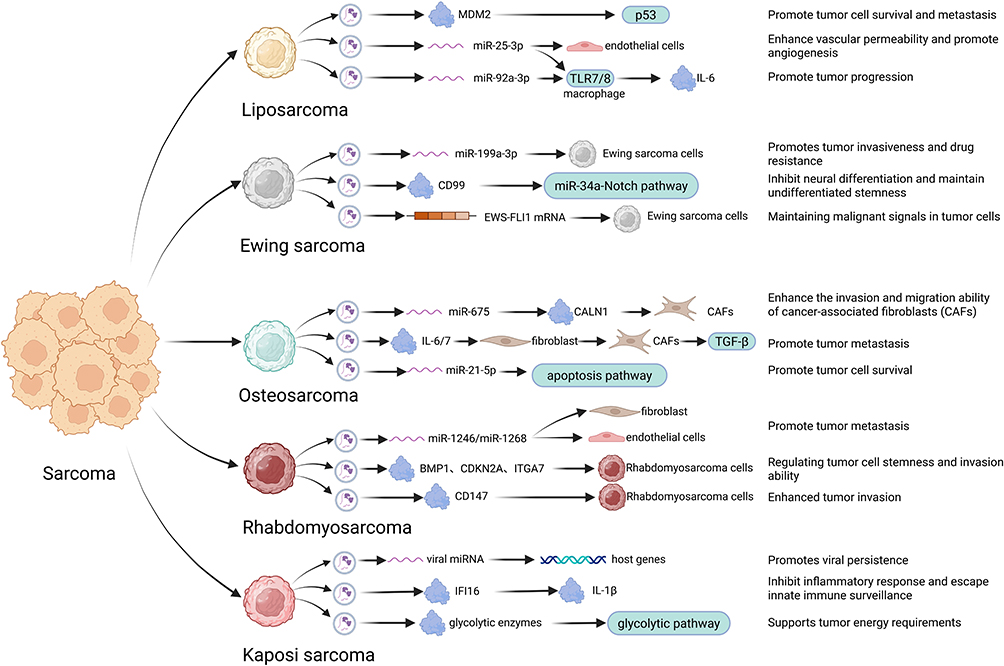 |
Figure 3 The functions and mechanisms of extracellular vesicles in Sarcoma. Extracellular vesicles have several functions and mechanisms cross various types of sarcomas. Liposarcoma-derived EVs: Promote tumor cell survival and metastasis, Enhance vascular permeability and promote angiogenesis; Ewing sarcoma-derived EVs: Promotes tumor invasiveness and drugresistance, Inhibit neural differentiation and maintain undifferentiated stemness, Maintaining malignant signals in tumor cells; Osteosarcoma-derived EVs: Enhance the invasion and migration ability of cancer-associated fibroblasts (CAFs), Promote tumor metastasis, Promote tumor cell survival; Rhabdomyosarcoma-derived EVs: Promote tumor metastasis, Regulating tumor cell stemness and invasion ability, Enhanced tumor invasion; Kaposi’s sarcoma-derived EVs: Promotes viral persistence, Inhibit inflammatory response and escape innate immune surveillance, Supports tumor energy requirements. Portrayed by BioRender.com. |
TME Reprogramming and Stromal Activation
A fundamental function of sarcoma-derived EVs is the education and activation of stromal cells to create a pro-tumorigenic niche. This is exemplified by osteosarcoma (OS)-EVs inducing fibroblast-to-CAF transformation via COL6A1/IL-6/IL-7 signals,122 liposarcoma (LPS)-EVs activating tumor-associated macrophages (TAMs) via miR-25-3p/TLR7/IL-6 signaling,97 and Ewing sarcoma (EWS)-EVs reprogramming fibroblasts and expanding myeloid-derived suppressor cells.116 Despite the distinct molecular triggers, the outcome is consistent: the creation of a supportive, reactive stroma that enhances tumor growth and invasion.
Metastatic Cascade Activation
EVs facilitate metastasis through pre-metastatic niche formation and enhancement of invasive potential. Osteosarcoma EVs carrying Rab22a-NeoF1 or S100A11 promote lung colonization by recruiting and educating myeloid cells,5,123 paralleled by liposarcoma EVs that disseminate oncogenic MDM2 DNA to induce proliferative and migratory phenotypes in stromal cells.98 This mechanism is conserved in Ewing sarcoma, where EVs transfer IGF2BP3 to enhance cell motility via PI3K/Akt signaling.117
Immune Evasion
A common feature across sarcomas is EV-mediated immunosuppression. Osteosarcoma EVs upregulate PD-L1 following chemotherapy,103 while Kaposi sarcoma EVs exploit viral miRNAs and deliver IFI16 to cleave IL-1β, thereby subverting inflammatory responses.108,134 Ewing sarcoma EVs further impair dendritic cell maturation and suppress T-cell effector functions, illustrating a recurrent theme of EVs as vehicles for innate and adaptive immune suppression.101,114
Transfer of Oncogenic Drivers
EVs facilitate the horizontal transfer of oncogenic molecules that define or drive specific sarcomas. This is most striking in fusion-driven sarcomas: EWS-EVs can carry the EWSR1-FLI1 fusion transcript itself, amplifying oncogenic signals,84,86 while OS-EVs transfer the Rab22a-NeoF1 fusion protein.123 In LPS, EVs deliver oncogenic miRNAs such as miR-25-3p.97 This intercellular transmission amplifies oncogenic signaling and enhances tumor heterogeneity and adaptability.
In summary, EVs from diverse sarcoma subtypes drive malignant progression through a convergent set of mechanisms, centered on reprogramming of the tumor microenvironment, initiation of the metastatic cascade, induction of immunosuppression, and intercellular transmission of oncogenic signals. This mechanistic unity not only offers an integrated perspective on EV function in sarcomas, but also suggests that targeting key steps in EV-mediated communication—such as EV biogenesis, cargo loading, or receptor uptake—may represent a potential broad-spectrum therapeutic strategy applicable across multiple sarcoma subtypes.
Dual Roles of Extracellular Vesicles in Sarcoma: Tumor Promotion and Suppression
While the preceding sections have detailed the extensive pro-tumorigenic roles of sarcoma-derived EVs, a growing body of evidence underscores their functional duality, wherein EVs from specific sources can also inhibit sarcoma progression. This tumor-suppressive capacity primarily manifests through two mechanisms: the intrinsic anti-tumor properties of EVs derived from normal or engineered stromal cells, and the context-dependent cargo that can counteract oncogenic signaling.
A prominent mechanism involves EVs originating from mesenchymal stem cells (MSCs), such as those derived from bone (BMSCs) or adipose tissue (AD-MSCs). These EVs can deliver a repertoire of tumor-suppressive nucleic acids to sarcoma cells. In osteosarcoma (OS), BMSCs-EVs have been demonstrated to carry multiple inhibitory cargos. For instance, they transfer miR-206 into OS cells, which directly targets neurensin-2 (NRSN2), leading to the suppression of the ERK1/2-Bcl-xL pathway and consequent inhibition of OS cell malignant progression in vitro and in vivo.137 Similarly, AD-MSCs engineered to secrete miR-101-enriched EVs effectively deliver this miRNA to OS cells. The delivered miR-101 directly targets BCL6, inhibiting cell invasion and migration in vitro and significantly reducing pulmonary metastasis in vivo, highlighting its therapeutic potential.138 Furthermore, circ-0000190, predominantly packaged into EVs from normal cells, can be transferred to OS cells. There, it acts as a competitive endogenous RNA (ceRNA) for miR-767-5p, thereby alleviating the suppression of TET1 and ultimately inhibiting malignant behavior.139 Beyond OS, in Ewing sarcoma (EWS), BMSCs-EVs carrying the long non-coding RNA LINC00847 have been shown to inhibit the proliferation, migration, and invasion of EWS cells.140 These findings, complemented by reports of MSC-EVs delivering miR-22 to suppress the Twist1/CADM1 axis in OS,126 collectively demonstrate a broad tumor-suppressive role for MSC-derived EVs across sarcoma subtypes.
Secondly, the functional output of EVs can be shifted toward tumor suppression by altering the molecular makeup of the parent sarcoma cell itself. A key example is observed in Ewing sarcoma. Proteomic analysis reveals that silencing the surface marker CD99 in EWS cells significantly alters the protein profile of their secreted EVs. EVs derived from CD99-low EWS cells carry a protein signature associated with inhibited migration and pro-immunostimulatory effects, thereby exerting an anti-tumor function upon transfer to recipient cells.100,113 This aligns with earlier findings that CD99-negative EWS cells secrete EVs enriched with miR-34a and miR-199a-3p, microRNAs that suppress the NF-κB/AP-1 pathway to inhibit metastatic behavior.112,113
Additionally, EVs from healthy stromal cells can inherently counteract sarcoma growth through distinct pathways. For example, osteoblast-derived EVs modulate the URG4/WIF1-Wnt signaling axis, promoting osteogenic differentiation of OS cells and suppressing their malignant phenotype, representing a natural defensive mechanism within the bone microenvironment.129
In summary, the role of EVs in sarcoma is not monolithic. Beyond being potent tumor promoters, EVs can function as vehicles for tumor-suppressive signals, either derived from normal/engineered tissues or reprogrammed from tumor cells themselves. Recognizing this context-dependent duality is critical for developing balanced EV-based diagnostic and therapeutic strategies, as the ultimate function of an EV is contingent upon its cellular origin, specific cargo, and the recipient cell environment.
Extracellular Vesicles in Sarcoma Diagnosis and Therapy
Extracellular vesicles (EVs) have attracted considerable interest for their potential as both tools and targets in sarcoma management, spanning diagnostic and therapeutic applications. Their contributions range from non-invasive biomarker discovery to serving a dual role as innate therapeutic agents/antitumor mediators and as intervention targets. Additionally, EVs are being engineered into sophisticated delivery systems for targeted therapy. The following section outlines key advances in these areas.
Extracellular Vesicles for Sarcoma Diagnosis and Prognosis
Extracellular vesicles (EVs), particularly exosomes, have emerged as pivotal mediators in sarcoma biology and clinical management. These nanoscale vesicles secreted by tumor and stromal cells carry tumor-specific biomolecules (proteins, miRNAs, DNA) that reflect dynamic changes in the tumor microenvironment (TME), offering unprecedented opportunities for minimally invasive diagnosis and prognostic stratification.141,142 The diagnostic power of EVs stems from their ability to encapsulate disease-specific molecular signatures. Proteomic analyses reveal that serum EV protein levels (eg, CD63, ENO-1) are significantly elevated in sarcoma patients compared to benign tumor cases and healthy controls, with distinct profiles enabling discrimination between osteosarcoma subtypes and metastatic states.8,15 Notably, sarcoma-derived EVs transport oncogenic drivers such as the Rab22a-NeoF1 fusion protein and its partner PYK2, which orchestrate pre-metastatic niche formation in osteosarcoma.124 Similarly, Ewing sarcoma-derived EVs exhibit unique surface markers like ENO-1, whose serum levels correlate with tumor burden and treatment response.143
Beyond protein biomarkers, EV-encapsulated nucleic acids provide exceptional specificity for molecular subtyping. Cross-sarcoma miRNA profiling reveals distinct expression patterns, with miR-335-5p upregulation in alveolar rhabdomyosarcoma (ARMS) EVs marking advanced disease stages,144 and a 62-miRNA signature in Ewing sarcoma EVs detecting 80% of cases while identifying novel EWS-FLI1 translocation variants.100,145 DNA-based diagnostics further expand this potential, as evidenced by HSATI/HSATII repeats in osteosarcoma EVs achieving diagnostic AUC≥0.90 for early detection.146 These molecular findings are being translated into clinical practice through technological breakthroughs in EV analysis.
Innovations like the “Click Beads” platform enable oncogenic mutation detection with 95% tissue biopsy concordance.147 Microfluidic innovations such as ZnO-nanorod chips allow simultaneous EV quantification and metastasis-associated vimentin detection in osteosarcoma,148 while partition-less digital immunoassays (μTIP-dELISA) achieve 97% diagnostic accuracy for Ewing sarcoma through multiplexed EV biomarker detection.149 Label-free techniques combining SERS and MALDI-TOF MS further enhance diagnostic precision by resolving cellular origin-specific EV signatures.150
The dynamic nature of EV biomarkers further enhances their clinical utility in therapeutic monitoring. Chemotherapy induces measurable changes in EV cargo composition: metastatic osteosarcoma patients show altered exosomal KIT and SPRY4 protein levels post-treatment, while downregulation of specific EV proteins predicts immediate drug responsiveness.151 This temporal resolution capability, combined with minimally invasive sampling, positions EVs as real-time sentinels of treatment efficacy. Importantly, the integration of spatial information (through TME-derived molecular profiles) and temporal data (via longitudinal biomarker tracking) provides a multidimensional perspective on tumor behavior. By bridging molecular heterogeneity with clinical observability, EV-based diagnostics are redefining sarcoma management—from enabling early detection through subtype-specific signatures to guiding adaptive treatment strategies via dynamic biomarker surveillance.
Leveraging the Innate Properties of Extracellular Vesicles for Sarcoma Therapy
Beyond diagnostic applications, the inherent biological properties of extracellular vesicles (EVs) themselves offer multiple strategies for sarcoma therapy. Firstly, thanks to their excellent biocompatibility and low immunogenicity, even unmodified, natural EVs are being explored as efficient delivery systems for chemotherapeutic drugs. For instance, doxorubicin-loaded mesenchymal stem cell-derived EVs demonstrated enhanced uptake by tumor cells and reduced cardiotoxicity compared to the free drug in in vitro osteosarcoma models, achieving superior anti-tumor efficacy and safety.152 Additionally, a recent study established an optimal 3D culture system for antler reserve mesenchymal cells to produce EVs (3D-RM-EVs).153 3D-RM-EVs loaded with ifosfamide and etoposide significantly enhanced tumor suppression in an in vivo osteosarcoma model compared to chemotherapy drugs alone, while also reducing weight loss, highlighting the potential of natural EVs as drug delivery vehicles.153
Secondly, EVs derived from immune cells, such as dendritic cells, show great potential in activating anti-tumor immunity. Dendritic cell-derived exosomes (Dex) are enriched on their surface with MHC-peptide complexes, co-stimulatory molecules, and adhesion molecules, enabling them to efficiently present tumor antigens to T cells, thereby stimulating a specific cellular immune response.154,155 In an experimental fibrosarcoma model, the injection of tumor antigen-loaded Dex has been shown to inhibit tumor growth and establish long-term immune memory.156 This strategy lays the foundation for developing EV-based sarcoma vaccines.
Furthermore, given the critical role of tumor-derived EVs (TEVs) in sarcoma progression, metastasis, and immunosuppression, targeting the EVs themselves has emerged as a novel therapeutic approach. Research indicates that sarcoma EVs mediate organotropic metastasis by expressing specific integrins (eg, integrin αvβ5), providing a molecular basis for interfering with their targeted delivery.35 More profoundly, Baglio et al revealed that osteosarcoma EV-educated mesenchymal stem cells (MSCs) were induced to produce interleukin-6 (IL-6), promoting tumor growth and lung metastasis. Blocking the IL-6 receptor with an antibody (tocilizumab) significantly reversed this tumor-promoting effect.157 Another study confirmed that the antibody drug bevacizumab could inhibit osteosarcoma angiogenesis and tumor growth in vitro and in vivo by targeting and suppressing the oncogenic lncRNA MIAT within EVs, thereby blocking its downstream miR-613/GPR158 signaling axis.158 Thus, neutralizing TEVs or inhibiting their cargo represents an innovative indirect therapeutic modality to disrupt tumor communication.
In summary, the innate properties of EVs themselves offer multiple promising strategies for sarcoma therapy. These include leveraging their features as efficient delivery systems for chemotherapeutic drugs to enhance efficacy and reduce toxicity; utilizing EVs derived from immune cells, such as dendritic cell-derived exosomes, to activate anti-tumor immunity, laying the groundwork for EV-based vaccines; and targeting tumor-derived EVs themselves or their harmful cargo to disrupt their roles in tumor progression, metastasis, and immunosuppression. Collectively, these approaches highlight the significant potential of multi-faceted EV-based therapeutic modalities in combating sarcoma.
Engineered Extracellular Vesicles for Sarcoma Therapy
Extracellular vesicles (EVs), particularly exosomes and engineered microvesicles (EMs), have emerged as versatile nanoplatforms for sarcoma therapy, leveraging their intrinsic biocompatibility, cell-targeting specificity, and capacity to overcome biological barriers. Building upon the foundational roles of native EVs, engineering strategies further augment their therapeutic potential by precision-loading of cargo and enhancing targetability. These engineered EVs are increasingly designed to deliver therapeutic payloads—ranging from chemotherapeutics to nucleic acids—with enhanced precision to tumor microenvironments (TMEs). For instance, McNamara et al harnessed tumor-derived exosomes to encapsulate chemotherapeutic agents via miRNA scaffolding, achieving superior tumor targeting and drug accumulation compared to conventional liposomal systems, thereby suppressing tumor growth and angiogenesis in preclinical models.159 Similarly, Wei et al demonstrated that mesenchymal stem cell (MSC)-derived exosomes delivered doxorubicin to osteosarcoma cells via SDF1-CXCR4 axis-mediated homing, significantly enhancing antitumor efficacy while mitigating cardiotoxicity.160 These studies underscore the potential of EV-based drug delivery to improve therapeutic windows in sarcoma.
A critical advancement lies in the engineering of EVs for multimodal therapeutic effects. Engineered exosomes can simultaneously reverse chemoresistance and induce tumor-specific cell death pathways. For example, Du et al developed NGR peptide-modified cancer-associated fibroblast (CAF)-derived exosomes to deliver autophagy-promoting peptides, which circumvented osteosarcoma chemoresistance by activating autophagy-dependent ferroptosis.161 Complementary to this, bone-targeted exosome nanoparticles functionalized with SDSSD peptides were shown to trigger ferroptosis in osteosarcoma through Keap1/Nrf2/GPX4 pathway activation, characterized by ROS accumulation, Fe2+ aggregation, and lipid peroxidation.162 This dual-action strategy—targeting both drug resistance and cell death mechanisms—highlights the sophistication of next-generation EV therapeutics.
To further enhance tumor specificity, surface modification strategies have been widely adopted. Sasmita et al engineered folate-conjugated osteoblast-derived EVs to co-deliver HDAC1 siRNA and zoledronic acid, achieving precise targeting of osteosarcoma cells while overcoming chemoresistance.163 Similarly, Xin et al developed c(RGDyK)-modified exosomes for αvβ integrin-targeted delivery of lncRNA MEG3, which suppressed osteosarcoma proliferation and metastasis via miR-185-5p regulation.17 These modifications not only improve EV accumulation in tumors but also minimize off-target effects, a critical consideration for clinical translation.
Antiangiogenic therapy represents another promising application. Wang et al revealed that bevacizumab inhibits osteosarcoma angiogenesis by blocking serum EV-transferred MIAT and enhancing miR-613-mediated GPR158 downregulation.158 In parallel, Yu et al engineered hepatocyte-derived small EVs (sEVs) loaded with VEGFR2-targeting siRNA, which outperformed the antiangiogenic drug apatinib in suppressing pulmonary metastatic osteosarcoma.164 Conversely, Palinski et al identified that sarcoma-derived EVs promote neovascularization by activating endothelial P2X4 receptors, suggesting that P2X4 blockade could synergize with EV-based antiangiogenic therapies.165
Recent studies have significantly advanced the frontiers of extracellular vesicle (EV)-based therapeutics for sarcoma, with notable progress spanning novel decoy approaches, production and engineering enhancements, and theranostic integrations. A prominent decoy strategy for osteosarcoma involves the use of saponin-treated, cargo-eliminated small EVs (CE-sEVs). Although oncogenic cargo is removed, these CE-sEVs retain their inherent lung fibroblast targeting capability.166 They function by competitively inhibiting the uptake of tumor-promoting osteosarcoma-derived EVs, thereby preventing pulmonary pre-metastatic niche formation and improving survival in preclinical models.166
In parallel, advancements in EV production and targeting have further enhanced their therapeutic potential. In drug loading, high-pressure homogenization has been used to encapsulate doxorubicin into MSC-derived small EVs (DXR-MSC-EVs), improving drug uptake efficiency and tumor toxicity while maintaining a favorable safety profile toward healthy cells.167 Three-dimensional culture of natural killer (NK) cells can increase EV yield, while modification with iRGD peptides enhances tumor-specific targeting. Additionally, a sequentially loaded EV system containing doxorubicin and the antitumor agent ABT-263 has been developed to induce senescence and clearance of osteosarcoma cells. This strategy effectively suppresses tumor growth and metastasis while reducing systemic toxicity.168 Another study developed a microvesicles platform modified with Ephrin alpha 2 (EphA2)-targeted peptides (YSAYPDSVPMMS, YSA) and loaded with methotrexate. This system achieved precise drug delivery in an osteosarcoma model, significantly enhancing antitumor efficacy while minimizing side effects.169
Beyond these approaches, theranostic platforms are evolving to combine diagnosis and treatment in a single system. One innovative example is a platform integrating 124I-labeled bone marrow MSC EVs with CRISPR/Cas9 ribonucleoproteins targeting KCNJ2. This 124I@EVs-Cas9 system permits in vivo tracking while simultaneously inhibiting osteosarcoma proliferation and metastasis via KCNJ2 suppression and HIF-1α ubiquitination-mediated degradation.170
Despite these preclinical advances, challenges remain in scaling EV therapeutics for clinical use. Key limitations include optimizing isolation protocols, enhancing payload stability, and evading immune clearance.141 For instance, Chen et al demonstrated that exosomes loaded with rifampin (EXO-RIF) induce mitochondrial fission-mediated apoptosis in osteosarcoma, yet achieving consistent drug encapsulation efficiency remains technically demanding.171 Future research must prioritize mechanistic studies to elucidate EV-mediated signaling across sarcoma subtypes and refine engineering strategies for improved targeting and delivery.172
In summary, engineered EVs represent a transformative approach for sarcoma therapy, integrating drug delivery, resistance reversal, and microenvironment modulation (Figure 4). While early-phase clinical trials are pending, the convergence of synthetic biology, nanotechnology, and tumor biology holds promise for translating these innovations into precision therapies for sarcoma patients.
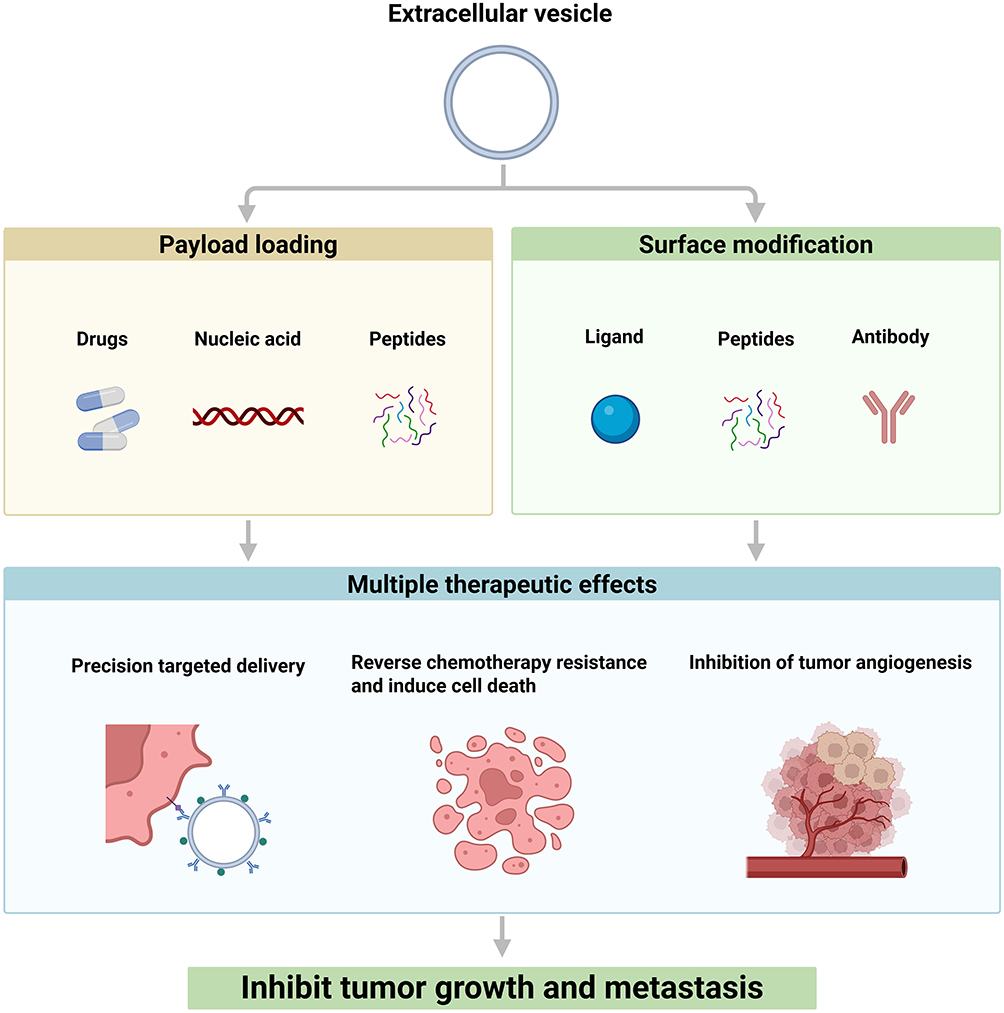 |
Figure 4 Engineered extracellular vesicles for sarcoma therapy. Engineered extracellular vesicles are developed through payload loading (eg, drugs, nucleic acids) and surface modification (eg, ligands, antibodies) to achieve targeted delivery, reverse chemoresistance, inhibit tumor angiogenesis, and ultimately suppress tumor growth and metastasis in sarcoma. Created with BioRender.com. |
Discussion
Extracellular vesicles (EVs) have emerged as pivotal mediators in sarcoma biology, orchestrating tumor progression, metastasis, and therapy resistance through dynamic intercellular communication. This review highlights the multifaceted roles of EVs across sarcoma subtypes, underscoring their capacity to transfer oncogenic cargo (eg, miRNAs, fusion proteins, and immune checkpoint molecules) to remodel the tumor microenvironment (TME), activate stromal cells, and suppress antitumor immunity. Subtype-specific mechanisms, such as osteosarcoma-derived EVs promoting pre-metastatic niche formation via Rab22a-NeoF1 fusion proteins or liposarcoma EVs driving immune evasion through miR-25-3p/TLR7 signaling, exemplify the heterogeneity of EV-mediated pathways in sarcoma pathogenesis.
The translational potential of EVs is twofold. First, their unique molecular signatures—enriched in tumor-specific proteins, nucleic acids, and surface markers—position EVs as promising non-invasive biomarkers for early diagnosis, prognosis, and real-time therapeutic monitoring. Innovations in EV isolation and analysis, including microfluidic platforms and multiplexed immunoassays, are accelerating their clinical adoption. Second, engineered EVs demonstrate remarkable versatility as targeted drug delivery systems, capable of overcoming chemoresistance, reversing immunosuppression, and suppressing metastasis through tailored cargo (eg, CRISPR-edited miRNAs, antiangiogenic siRNAs). However, challenges such as scalable production, payload stability, and immune clearance must be addressed to advance these therapies into clinical trials.
Future research should prioritize elucidating the context-dependent roles of EVs across sarcoma subtypes, leveraging single-EV profiling and spatial transcriptomics to resolve their functional heterogeneity. Additionally, interdisciplinary efforts integrating synthetic biology, nanotechnology, and tumor immunology will be critical to optimizing EV-based strategies for precision oncology. By bridging mechanistic insights with technological innovation, EV-centric approaches hold transformative potential to redefine sarcoma management, offering hope for overcoming the therapeutic limitations that have long plagued this aggressive disease.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Natural Science Foundation of China (82272935), the Natural Science Foundation of Fujian Province (2025J08239), the Young and Middle-aged Teachers Educational Scientific Research Program of Fujian Province (JAT241104) and the Startup Fund for Advanced Talents of Putian University (2024148).
Disclosure
The authors declare no competing interests.
References
1. von Mehren M, Kane JM, Bui MM, et al. NCCN guidelines insights: soft tissue sarcoma, version 1.2021. J National Compr Cancer Network. 2020;18(12):1604–1612. doi:10.6004/jnccn.2020.0058
2. Sbaraglia M, Bellan E, Dei Tos AP. The 2020 WHO classification of soft tissue tumours: news and perspectives. Pathologica. 2021;113(2):70–84. doi:10.32074/1591-951x-213
3. Kantidakis G, Litière S, Neven A, et al. New benchmarks to design clinical trials with advanced or metastatic liposarcoma or synovial sarcoma patients: an EORTC- Soft Tissue and Bone Sarcoma Group (STBSG) meta-analysis based on a literature review for soft-tissue sarcomas. Eur J Cancer. 2022;174:261–276. doi:10.1016/j.ejca.2022.07.010
4. Cocks A, Martinez-Rodriguez V, Del Vecchio F, et al. Diverse roles of EV-RNA in cancer progression. Semi Cancer Biol. 2021;75:127–135. doi:10.1016/j.semcancer.2020.11.022
5. Deng CZ, Xu YY, Chen HM, et al. Extracellular-vesicle-packaged S100A11 from osteosarcoma cells mediates lung premetastatic niche formation by recruiting gMDSCs. Cell Rep. 2024;43(2):113751. doi:10.1016/j.celrep.2024.113751
6. Raimondi L, De Luca A, Gallo A, et al. Osteosarcoma cell-derived exosomes affect tumor microenvironment by specific packaging of microRNAs. Carcinogenesis. 2020;41(5):666–677. doi:10.1093/carcin/bgz130
7. Zhai C, Xu J, Yang Y, et al. Heterogeneous analysis of extracellular vesicles for osteosarcoma diagnosis. Anal Chem. 2024;96(23):9486–9492. doi:10.1021/acs.analchem.4c00941
8. Yokoo S, Fujiwara T, Yoshida A, et al. Liquid biopsy targeting monocarboxylate transporter 1 on the surface membrane of tumor-derived extracellular vesicles from synovial sarcoma. Cancers. 2021;13(8):1823. doi:10.3390/cancers13081823
9. Zhang X, Zhang H, Gu J, et al. Engineered extracellular vesicles for cancer therapy. Adv Mater. 2021;33(14):e2005709. doi:10.1002/adma.202005709
10. Wolf P. The nature and significance of platelet products in human plasma. Br J Haematol. 1967;13(3):269–288. doi:10.1111/j.1365-2141.1967.tb08741.x
11. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi:10.1084/jem.183.3.1161
12. Chen Y, Gu B, Wu S, et al. Using enrichment index for quality control of secretory protein sample and identification of secretory proteins. J Mass Spectrom. 2009;44(3):397–403. doi:10.1002/jms.1517
13. Osaki M, Takeshita F, Sugimoto Y, et al. MicroRNA-143 regulates human osteosarcoma metastasis by regulating matrix metalloprotease-13 expression. Mol Ther. 2011;19(6):1123–1130. doi:10.1038/mt.2011.53
14. Fujiwara T, Uotani K, Yoshida A, et al. Clinical significance of circulating miR-25-3p as a novel diagnostic and prognostic biomarker in osteosarcoma. Oncotarget. 2017;8(20):33375–33392. doi:10.18632/oncotarget.16498
15. Wang J, Zhang H, Sun X, et al. Exosomal PD-L1 and N-cadherin predict pulmonary metastasis progression for osteosarcoma patients. J Nanobiotechnol. 2020;18(1):151. doi:10.1186/s12951-020-00710-6
16. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16(7):748–759. doi:10.1038/s41565-021-00931-2
17. Huang X, Wu W, Jing D, et al. Engineered exosome as targeted lncRNA MEG3 delivery vehicles for osteosarcoma therapy. J Control Release. 2022;343:107–117. doi:10.1016/j.jconrel.2022.01.026
18. Fusco C, De Rosa G, Spatocco I, et al. Extracellular vesicles as human therapeutics: a scoping review of the literature. J Extracell Vesicles. 2024;13(5):e12433. doi:10.1002/jev2.12433
19. Zaborowski MP, Balaj L, Breakefield XO, Lai CP. Extracellular vesicles: composition, biological relevance, and methods of study. Bioscience. 2015;65(8):783–797. doi:10.1093/biosci/biv084
20. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
21. Yáñez-Mó M, Siljander PRM, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4(1):27066. doi:10.3402/jev.v4.27066
22. Smyth IM, Bertram JF. Seminars in cell and developmental biology. Semin Cell Dev Biol. 2019;91:84–85. doi:10.1016/j.semcdb.2018.11.003
23. Riancho J, Sánchez-Juan P. Circulating extracellular vesicles in human disease. N Engl J Med. 2018;379(22):2180. doi:10.1056/NEJMc1813170
24. György B, Szabó TG, Pásztói M, et al. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol Life Sci. 2011;68(16):2667–2688. doi:10.1007/s00018-011-0689-3
25. Biller SJ, Schubotz F, Roggensack SE, Thompson AW, Summons RE, Chisholm SW. Bacterial vesicles in marine ecosystems. Science. 2014;343(6167):183–186. doi:10.1126/science.1243457
26. Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013;126(Pt 24):5553–5565. doi:10.1242/jcs.128868
27. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–1247. doi:10.1126/science.1153124
28. Wubbolts R, Leckie RS, Veenhuizen PTM, et al. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J Biol Chem. 2003;278(13):10963–10972. doi:10.1074/jbc.M207550200
29. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30(1):255–289. doi:10.1146/annurev-cellbio-101512-122326
30. Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–593. doi:10.1038/nri2567
31. Muralidharan-Chari V, Clancy J, Plou C, et al. ARF6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr Biol. 2009;19(22):1875–1885. doi:10.1016/j.cub.2009.09.059
32. D’Souza-Schorey C, Chavrier P. ARF proteins: roles in membrane traffic and beyond. Nat Rev Mol Cell Biol. 2006;7(5):347–358. doi:10.1038/nrm1910
33. Booth AM, Fang Y, Fallon JK, Yang J-M, Hildreth JEK, Gould SJ. Exosomes and HIV Gag bud from endosome-like domains of the T cell plasma membrane. J Cell Biol. 2006;172(6):923–935. doi:10.1083/jcb.200508014
34. Li S, Xu J, Qian J, Gao X. Engineering extracellular vesicles for cancer therapy: recent advances and challenges in clinical translation. Biomater Sci. 2020;8(24):6978–6991. doi:10.1039/d0bm01385d
35. Hoshino A, Costa-Silva B, Shen T-L, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527(7578):329–335. doi:10.1038/nature15756
36. Raiborg C, Wenzel EM, Pedersen NM, et al. Repeated ER-endosome contacts promote endosome translocation and neurite outgrowth. Nature. 2015;520(7546):234–238. doi:10.1038/nature14359
37. Sinha S, Hoshino D, Hong NH, et al. Cortactin promotes exosome secretion by controlling branched actin dynamics. J Cell Biol. 2016;214(2):197–213. doi:10.1083/jcb.201601025
38. Öztürk S, Elçin AE, Koca A, Elçin YM. Therapeutic applications of stem cells and extracellular vesicles in emergency care: futuristic perspectives. Stem Cell Rev Rep. 2021;17(2):390–410. doi:10.1007/s12015-020-10029-2
39. Bebelman MP, Smit MJ, Pegtel DM, Baglio SR. Biogenesis and function of extracellular vesicles in cancer. Pharmacol Ther. 2018;188:1–11. doi:10.1016/j.pharmthera.2018.02.013
40. Theodoraki M-N, Yerneni SS, Hoffmann TK, Gooding WE, Whiteside TL. Clinical significance of PD-L1+ exosomes in plasma of head and neck cancer patients. Clin Cancer Res. 2018;24(4):896–905. doi:10.1158/1078-0432.CCR-17-2664
41. Yang Y, Li C-W, Chan L-C, et al. Exosomal PD-L1 harbors active defense function to suppress T cell killing of breast cancer cells and promote tumor growth. Cell Res. 2018;28(8):862–864. doi:10.1038/s41422-018-0060-4
42. Morris-Love J, Atwood WJ. Complexities of JC polyomavirus receptor-dependent and -independent mechanisms of infection. Viruses. 2022;14(6):1130. doi:10.3390/v14061130
43. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–9420. doi:10.1016/S0021-9258(18)48095-7
44. An M, Wu J, Zhu J, Lubman DM. Comparison of an optimized ultracentrifugation method versus size-exclusion chromatography for isolation of exosomes from human serum. J Proteome Res. 2018;17(10):3599–3605. doi:10.1021/acs.jproteome.8b00479
45. Guan S, Yu H, Yan G, Gao M, Sun W, Zhang X. Characterization of urinary exosomes purified with size exclusion chromatography and ultracentrifugation. J Proteome Res. 2020;19(6):2217–2225. doi:10.1021/acs.jproteome.9b00693
46. Linares R, Tan S, Gounou C, Arraud N, Brisson AR. High-speed centrifugation induces aggregation of extracellular vesicles. J Extracell Vesicles. 2015;4(1):29509. doi:10.3402/jev.v4.29509
47. Stam J, Bartel S, Bischoff R, Wolters JC. Isolation of extracellular vesicles with combined enrichment methods. J Chromatogr B Analyt Technol Biomed Life Sci. 2021;1169:122604. doi:10.1016/j.jchromb.2021.122604
48. Busatto S, Vilanilam G, Ticer T, et al. Tangential flow filtration for highly efficient concentration of extracellular vesicles from large volumes of fluid. Cells. 2018;7(12):273. doi:10.3390/cells7120273
49. Visan KS, Lobb RJ, Ham S, et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J Extracell Vesicles. 2022;11(9):e12266. doi:10.1002/jev2.12266
50. Kowal J, Arras G, Colombo M, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A. 2016;113(8):E968–E977. doi:10.1073/pnas.1521230113
51. Multia E, Tear CJY, Palviainen M, Siljander P, Riekkola M-L. Fast isolation of highly specific population of platelet-derived extracellular vesicles from blood plasma by affinity monolithic column, immobilized with anti-human CD61 antibody. Anal Chim Acta. 2019;1091:160–168. doi:10.1016/j.aca.2019.09.022
52. Zarovni N, Corrado A, Guazzi P, et al. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches. Methods. 2015;87:46–58. doi:10.1016/j.ymeth.2015.05.028
53. Casadei L, Choudhury A, Sarchet P, et al. Cross-flow microfiltration for isolation, selective capture and release of liposarcoma extracellular vesicles. J Extracell Vesicles. 2021;10(4):e12062. doi:10.1002/jev2.12062
54. Kalluri R. The biology and function of exosomes in cancer. J Clin Invest. 2016;126(4):1208–1215. doi:10.1172/JCI81135
55. Lee YJ, Chae S, Choi D. Monitoring of single extracellular vesicle heterogeneity in cancer progression and therapy. Front Oncol. 2023;13:1256585. doi:10.3389/fonc.2023.1256585
56. Gai C, Pomatto MAC, Grange C, Deregibus MC, Camussi G. Extracellular vesicles in onco-nephrology. Exp Mol Med. 2019;51(3):1–8. doi:10.1038/s12276-019-0213-7
57. Gardiner C, Di Vizio D, Sahoo S, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles. 2016;5(1):32945. doi:10.3402/jev.v5.32945
58. Pang B, Zhu Y, Ni J, et al. Extracellular vesicles: the next generation of biomarkers for liquid biopsy-based prostate cancer diagnosis. Theranostics. 2020;10(5):2309–2326. doi:10.7150/thno.39486
59. Zheng X, Wang X, Li X. Extracellular vesicle-based liquid biopsy holds great promise for the management of ovarian cancer. Biochim Biophys Acta Rev Cancer. 2020;1874(1):188395. doi:10.1016/j.bbcan.2020.188395
60. Wu B, Liu D-A, Guan L, et al. Stiff matrix induces exosome secretion to promote tumour growth. Nat Cell Biol. 2023;25(3):415–424. doi:10.1038/s41556-023-01092-1
61. Patwardhan S, Mahadik P, Shetty O, Sen S. ECM stiffness-tuned exosomes drive breast cancer motility through thrombospondin-1. Biomaterials. 2021;279:121185. doi:10.1016/j.biomaterials.2021.121185
62. Wang M, Yu W, Cao X, et al. Exosomal CD44 transmits lymph node metastatic capacity between gastric cancer cells via YAP-CPT1A-mediated FAO reprogramming. Front Oncol. 2022;12:860175. doi:10.3389/fonc.2022.860175
63. Xie Z, Gao Y, Ho C, et al. Exosome-delivered CD44v6/C1QBP complex drives pancreatic cancer liver metastasis by promoting fibrotic liver microenvironment. Gut. 2022;71(3):568–579. doi:10.1136/gutjnl-2020-323014
64. Choi H, Choi Y, Yim HY, Mirzaaghasi A, Yoo J-K, Choi C. Biodistribution of exosomes and engineering strategies for targeted delivery of therapeutic exosomes. Tissue Eng Regen Med. 2021;18(4):499–511. doi:10.1007/s13770-021-00361-0
65. Park JE, Tan HS, Datta A, et al. Hypoxic tumor cell modulates its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and exosomes. Mol Cell Proteomics. 2010;9(6):1085–1099. doi:10.1074/mcp.M900381-MCP200
66. Du X, Zhang X, Dong J, et al. Irradiation-induced exosomal HMGB1 to confer radioresistance via the PI3K/AKT/FOXO3A signaling pathway in ESCC. J Transl Med. 2022;20(1):507. doi:10.1186/s12967-022-03720-0
67. Paggetti J, Haderk F, Seiffert M, et al. Exosomes released by chronic lymphocytic leukemia cells induce the transition of stromal cells into cancer-associated fibroblasts. Blood. 2015;126(9):1106–1117. doi:10.1182/blood-2014-12-618025
68. Webber JP, Spary LK, Sanders AJ, et al. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene. 2015;34(3):290–302. doi:10.1038/onc.2013.560
69. Giusti I, Di Francesco M, D’Ascenzo S, et al. Ovarian cancer-derived extracellular vesicles affect normal human fibroblast behavior. Cancer Biol Ther. 2018;19(8):722–734. doi:10.1080/15384047.2018.1451286
70. Melo SA, Sugimoto H, O’Connell JT, et al. Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell. 2014;26(5):707–721. doi:10.1016/j.ccell.2014.09.005
71. Wu L, Zhang X, Zhang B, et al. Exosomes derived from gastric cancer cells activate NF-κB pathway in macrophages to promote cancer progression. Tumour Biol. 2016;37(9):12169–12180. doi:10.1007/s13277-016-5071-5
72. Corrado C, Raimondo S, Saieva L, Flugy AM, De Leo G, Alessandro R. Exosome-mediated crosstalk between chronic myelogenous leukemia cells and human bone marrow stromal cells triggers an interleukin 8-dependent survival of leukemia cells. Cancer Lett. 2014;348(1–2):71–76. doi:10.1016/j.canlet.2014.03.009
73. Zhang X, Yuan X, Shi H, Wu L, Qian H, Xu W. Exosomes in cancer: small particle, big player. J Hematol Oncol. 2015;8(1):83. doi:10.1186/s13045-015-0181-x
74. Waugh DJJ, Wilson C. The interleukin-8 pathway in cancer. Clin Cancer Res. 2008;14(21):6735–6741. doi:10.1158/1078-0432.CCR-07-4843
75. Hood JL, Pan H, Lanza GM, Wickline SA. Paracrine induction of endothelium by tumor exosomes. Lab Invest. 2009;89(11):1317–1328. doi:10.1038/labinvest.2009.94
76. Hood JL, San RS, Wickline SA. Exosomes released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 2011;71(11):3792–3801. doi:10.1158/0008-5472.CAN-10-4455
77. Luga V, Zhang L, Viloria-Petit AM, et al. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell. 2012;151(7):1542–1556. doi:10.1016/j.cell.2012.11.024
78. Zomer A, Maynard C, Verweij FJ, et al. In Vivo imaging reveals extracellular vesicle-mediated phenocopying of metastatic behavior. Cell. 2015;161(5):1046–1057. doi:10.1016/j.cell.2015.04.042
79. Lugini L, Valtieri M, Federici C, et al. Exosomes from human colorectal cancer induce a tumor-like behavior in colonic mesenchymal stromal cells. Oncotarget. 2016;7(31):50086–50098. doi:10.18632/oncotarget.10574
80. Qiu Y, Yang Y, Yang R, et al. Activated T cell-derived exosomal PD-1 attenuates PD-L1-induced immune dysfunction in triple-negative breast cancer. Oncogene. 2021;40(31):4992–5001. doi:10.1038/s41388-021-01896-1
81. Song YH, Warncke C, Choi SJ, et al. Breast cancer-derived extracellular vesicles stimulate myofibroblast differentiation and pro-angiogenic behavior of adipose stem cells. Matrix Biol. 2017;60-61:190–205. doi:10.1016/j.matbio.2016.11.008
82. Han L, Long Q, Li S, et al. Senescent stromal cells promote cancer resistance through SIRT1 loss-potentiated overproduction of small extracellular vesicles. Cancer Res. 2020;80(16):3383–3398. doi:10.1158/0008-5472.CAN-20-0506
83. Batrakova EV, Kim MS. Using exosomes, naturally-equipped nanocarriers, for drug delivery. J Control Release. 2015;219:396–405. doi:10.1016/j.jconrel.2015.07.030
84. Choi EYK, Gardner JM, Lucas DR, McHugh JB, Patel RM. Ewing sarcoma. Sem Diagnostic Pathol. 2014;31(1):39–47. doi:10.1053/j.semdp.2014.01.002
85. Kunisada T, Nakata E, Fujiwara T, et al. Soft-tissue sarcoma in adolescents and young adults. Int J Clin Oncol. 2023;28(1):1–11. doi:10.1007/s10147-022-02119-7
86. Tsugita M, Yamada N, Noguchi S, et al. Ewing sarcoma cells secrete EWS/Fli-1 fusion mRNA via microvesicles. PLoS One. 2013;8(10):e77416. doi:10.1371/journal.pone.0077416
87. Tang J, He J, Feng C, Tu C. Exosomal MiRNAs in osteosarcoma: biogenesis and biological functions. Front Pharmacol. 2022;13:902049. doi:10.3389/fphar.2022.902049
88. Liu S, Liu C, Wang Y, et al. The role of programmed cell death in osteosarcoma: from pathogenesis to therapy. Cancer Med. 2024;13(10):e7303. doi:10.1002/cam4.7303
89. Wu F, Huang Y, Huang X, et al. 99mTc-MIBI scintigraphy for the preoperative assessment of histological response to neoadjuvant chemotherapy in patients with osteosarcoma: a systematic review and a bivariate meta-analysis. Front Oncol. 2020;10:762. doi:10.3389/fonc.2020.00762
90. Fang X, Zhu L, Wu Q, Zhong X, Chen Y, Zhu W. The influence of MDR1 expression regulated by miR-138 through TRPS1 signaling pathway on multidrug resistance of osteosarcoma and formation of bacterial infection biofilm. Cell Mol Biol. 2023;69(5):51–57. doi:10.14715/cmb/2023.69.5.9
91. Chang Y, Cesarman E, Pessin MS, et al. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science. 1994;266(5192):1865–1869. doi:10.1126/science.7997879
92. Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N Engl J Med. 1995;332(18):1186–1191. doi:10.1056/NEJM199505043321802
93. Simonart T, Van Vooren JP, Meuris S. Kaposi’s sarcoma. N Engl J Med. 2000;343(8):582. author reply 583-582; author reply 584.
94. Bill KLJ, Casadei L, Prudner BC, Iwenofu H, Strohecker AM, Pollock RE. Liposarcoma: molecular targets and therapeutic implications. Cell Mol Life Sci. 2016;73(19):3711–3718. doi:10.1007/s00018-016-2266-2
95. Malempati S, Hawkins DS. Rhabdomyosarcoma: review of the Children’s Oncology Group (COG) Soft-Tissue Sarcoma Committee experience and rationale for current COG studies. Pediatr Blood Cancer. 2012;59(1):5–10. doi:10.1002/pbc.24118
96. Dantonello TM, Int-Veen C, Schuck A, et al. Survival following disease recurrence of primary localized alveolar rhabdomyosarcoma. Pediatr Blood Cancer. 2013;60(8):1267–1273. doi:10.1002/pbc.24488
97. Casadei L, Calore F, Creighton CJ, et al. Exosome-derived miR-25-3p and miR-92a-3p stimulate liposarcoma progression. Cancer Res. 2017;77(14):3846–3856. doi:10.1158/0008-5472.CAN-16-2984
98. Casadei L, Calore F, Braggio DA, et al. MDM2 derived from dedifferentiated liposarcoma extracellular vesicles induces MMP2 production from preadipocytes. Cancer Res. 2019;79(19):4911–4922. doi:10.1158/0008-5472.CAN-19-0203
99. Miller IV, Raposo G, Welsch U, et al. First identification of Ewing’s sarcoma-derived extracellular vesicles and exploration of their biological and potential diagnostic implications. Biol Cell. 2013;105(7):289–303. doi:10.1111/boc.201200086
100. De Feo A, Manfredi M, Mancarella C, et al. CD99 modulates the proteomic landscape of Ewing sarcoma cells and related extracellular vesicles. Int J Mol Sci. 2024;25(3):1588. doi:10.3390/ijms25031588
101. Ruzanov P, Evdokimova V, Pachva MC, et al. Oncogenic ETS fusions promote DNA damage and proinflammatory responses via pericentromeric RNAs in extracellular vesicles. J Clin Invest. 2024;134(9). doi:10.1172/JCI169470
102. Gong L, Bao Q, Hu C, et al. Exosomal miR-675 from metastatic osteosarcoma promotes cell migration and invasion by targeting CALN1. Biochem Biophys Res Commun. 2018;500(2):170–176. doi:10.1016/j.bbrc.2018.04.016
103. Yati S, Silathapanasakul A, Thakaeng C, et al. Extracellular vesicle-mediated IL-1 signaling in response to doxorubicin activates PD-L1 expression in osteosarcoma models. Cells. 2022;11(6):1042. doi:10.3390/cells11061042
104. Cai T, Zhang C, Zhan T. Transfer of exosomal microRNAs confers doxorubicin resistance in osteosarcoma cells. Mol Med Rep. 2023;27(4). doi:10.3892/mmr.2023.12973
105. Ghayad SE, Rammal G, Ghamloush F, et al. Exosomes derived from embryonal and alveolar rhabdomyosarcoma carry differential miRNA cargo and promote invasion of recipient fibroblasts. Sci Rep. 2016:637088. doi:10.1038/srep37088.
106. Fahs A, Hussein N, Zalzali H, et al. CD147 promotes tumorigenesis via exosome-mediated signaling in rhabdomyosarcoma. Cells. 2022;11(15):2267. doi:10.3390/cells11152267
107. Jeon H, Kang SK, Lee MJ, et al. Rab27b regulates extracellular vesicle production in cells infected with Kaposi’s sarcoma-associated herpesvirus to promote cell survival and persistent infection. J Microbiol. 2021;59(5):522–529. doi:10.1007/s12275-021-1108-6
108. Singh VV, Kerur N, Bottero V, et al. Kaposi’s sarcoma-associated herpesvirus latency in endothelial and B cells activates gamma interferon-inducible protein 16-mediated inflammasomes. J Virol. 2013;87(8):4417–4431. doi:10.1128/jvi.03282-12
109. Kok VC, Yu -C-C. Cancer-derived exosomes: their role in cancer biology and biomarker development. Int J Nanomed. 2020;15:8019–8036. doi:10.2147/IJN.S272378
110. Zeng Z, Li Y, Pan Y, et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat Commun. 2018;9(1):5395. doi:10.1038/s41467-018-07810-w
111. Syed SN, Brüne B. Exosomal and non-exosomal MicroRNAs: new kids on the block for cancer therapy. Int J Mol Sci. 2022;23(9):4493. doi:10.3390/ijms23094493
112. Ventura S, Aryee DNT, Felicetti F, et al. CD99 regulates neural differentiation of Ewing sarcoma cells through miR-34a-Notch-mediated control of NF-κB signaling. Oncogene. 2016;35(30):3944–3954. doi:10.1038/onc.2015.463
113. De Feo A, Sciandra M, Ferracin M, et al. Exosomes from CD99-deprived Ewing sarcoma cells reverse tumor malignancy by inhibiting cell migration and promoting neural differentiation. Cell Death Dis. 2019;10(7):471. doi:10.1038/s41419-019-1675-1
114. Gassmann H, Schneider K, Evdokimova V, et al. Ewing sarcoma-derived extracellular vesicles impair dendritic cell maturation and function. Cells. 2021;10(8):2081. doi:10.3390/cells10082081
115. Xiong J, Wu L, Huang L, et al. LncRNA FOXP4-AS1 promotes progression of Ewing sarcoma and is associated with immune infiltrates. Front Oncol. 2021:11718876. doi:10.3389/fonc.2021.718876.
116. Pachva MC, Lai H, Jia A, Rouleau M, Sorensen PH. Extracellular vesicles in reprogramming of the Ewing sarcoma tumor microenvironment. Front Cell Develop Biol. 2021;9726205. doi:10.3389/fcell.2021.726205
117. Mancarella C, Giusti V, Caldoni G, et al. Extracellular vesicle-associated IGF2BP3 tunes Ewing sarcoma cell migration and affects PI3K/Akt pathway in neighboring cells. Cancer Genet Ther. 2023;30(9):1285–1295. doi:10.1038/s41417-023-00637-8
118. Jerez S, Araya H, Hevia D, et al. Extracellular vesicles from osteosarcoma cell lines contain miRNAs associated with cell adhesion and apoptosis. Gene. 2019;710:246–257. doi:10.1016/j.gene.2019.06.005
119. Cai H, Zhao H, Tang J, Wu H. Serum miR-195 is a diagnostic and prognostic marker for osteosarcoma. J Surg Res. 2015;194(2):505–510. doi:10.1016/j.jss.2014.11.025
120. Ma W, Zhang X, Chai J, Chen P, Ren P, Gong M. Circulating miR-148a is a significant diagnostic and prognostic biomarker for patients with osteosarcoma. Tumour Biol. 2014;35(12):12467–12472. doi:10.1007/s13277-014-2565-x
121. Endo-Munoz L, Cai N, Cumming A, et al. Progression of osteosarcoma from a non-metastatic to a metastatic phenotype is causally associated with activation of an autocrine and paracrine uPA axis. PLoS One. 2015;10(8):e0133592. doi:10.1371/journal.pone.0133592
122. Zhang Y, Liu Z, Yang X, et al. H3K27 acetylation activated-COL6A1 promotes osteosarcoma lung metastasis by repressing STAT1 and activating pulmonary cancer-associated fibroblasts. Theranostics. 2021;11(3):1473–1492. doi:10.7150/thno.51245
123. Zhong L, Liao D, Li J, et al. Rab22a-NeoF1 fusion protein promotes osteosarcoma lung metastasis through its secretion into exosomes. Signal Transduct Target Ther. 2021;6(1):59. doi:10.1038/s41392-020-00414-1
124. Cheng Z, Wang L, Wu C, Huang L, Ruan Y, Xue W. Tumor-derived exosomes induced M2 macrophage polarization and promoted the metastasis of osteosarcoma cells through Tim-3. Arch Med Res. 2021;52(2):200–210. doi:10.1016/j.arcmed.2020.10.018
125. Griffin KH, Mizenko RR, Arun V, Carney RP, Leach JK. Extracellular vesicles from highly metastatic osteosarcoma cells induce pro-tumorigenic macrophage phenotypes. Adv Biol. 2024;8(6):e2300577. doi:10.1002/adbi.202300577
126. Ruan Q, Wang C, Wu Y, Zhu Q. Exosome microRNA-22 inhibiting proliferation, migration and invasion through regulating Twist1/CADM1 axis in osteosarcoma. Sci Rep. 2024;14(1):761. doi:10.1038/s41598-023-50612-4
127. Lin L, Wang H, Guo W, He E, Huang K, Zhao Q. Osteosarcoma-derived exosomal miR-501-3p promotes osteoclastogenesis and aggravates bone loss. Cell Signal. 2021;82:109935. doi:10.1016/j.cellsig.2021.109935
128. Ucci A, Cappariello A, Ponzetti M, et al. Anti-osteoblastogenic, pro-inflammatory and pro-angiogenic effect of extracellular vesicles isolated from the human osteosarcoma cell line MNNG/HOS. Bone. 2021;153:116130. doi:10.1016/j.bone.2021.116130
129. Leng Y, Li J, Long Z, et al. Osteoblast-derived exosomes promote osteogenic differentiation of osteosarcoma cells via URG4/Wnt signaling pathway. Bone. 2024;178:116933. doi:10.1016/j.bone.2023.116933
130. He H, Ding M, Li T, et al. Bone mesenchymal stem cell-derived extracellular vesicles containing NORAD promote osteosarcoma by miR-30c-5p. Lab Invest. 2022;102(8):826–837. doi:10.1038/s41374-021-00691-6
131. Jiang M, Jike Y, Liu K, et al. Exosome-mediated miR-144-3p promotes ferroptosis to inhibit osteosarcoma proliferation, migration, and invasion through regulating ZEB1. Mol Cancer. 2023;22(1):113. doi:10.1186/s12943-023-01804-z
132. Roberts BK, Li DI, Somerville C, et al. IRF5 suppresses metastasis through the regulation of tumor-derived extracellular vesicles and pre-metastatic niche formation. Sci Rep. 2024;14(1):15557. doi:10.1038/s41598-024-66168-w
133. Rammal G, Fahs A, Kobeissy F, et al. Proteomic profiling of rhabdomyosarcoma-derived exosomes yield insights into their functional role in paracrine signaling. J Proteome Res. 2019;18(10):3567–3579. doi:10.1021/acs.jproteome.9b00157
134. Jeon H, Yoo SM, Choi HS, et al. Extracellular vesicles from KSHV-infected endothelial cells activate the complement system. Oncotarget. 2017;8(59):99841–99860. doi:10.18632/oncotarget.21668
135. Chugh PE, Sin SH, Ozgur S, et al. Systemically circulating viral and tumor-derived MicroRNAs in KSHV-associated malignancies. PLoS Pathogens. 2013;9(7):e1003484. doi:10.1371/journal.ppat.1003484
136. Meckes DG, Gunawardena HP, Dekroon RM, et al. Modulation of B-cell exosome proteins by gamma herpesvirus infection. Proc Natl Acad Sci U S A. 2013;110(31):E2925–E2933. doi:10.1073/pnas.1303906110
137. Keremu A, Aila P, Tusun A, Abulikemu M, Zou X. Extracellular vesicles from bone mesenchymal stem cells transport microRNA-206 into osteosarcoma cells and target NRSN2 to block the ERK1/2-Bcl-xL signaling pathway. Eur J Histochem. 2022;66(3). doi:10.4081/ejh.2022.3394
138. Zhang K, Dong C, Chen M, et al. Extracellular vesicle-mediated delivery of miR-101 inhibits lung metastasis in osteosarcoma. Theranostics. 2020;10(1):411–425. doi:10.7150/thno.33482
139. Li S, Pei Y, Wang W, Liu F, Zheng K, Zhang X. Extracellular nanovesicles-transmitted circular RNA has_circ_0000190 suppresses osteosarcoma progression. J Cell Mol Med. 2020;24(3):2202–2214. doi:10.1111/jcmm.14877
140. Huang L, Xiong J, Fu J, et al. Bone marrow mesenchymal stem cell-derived exosomal LINC00847 inhibits the proliferation, migration, and invasion of Ewing sarcoma. J Clin Trans Res. 2022;8(6):563–576.
141. Ye H, Hu X, Wen Y, et al. Exosomes in the tumor microenvironment of sarcoma: from biological functions to clinical applications. J Nanobiotechnol. 2022;20(1):403. doi:10.1186/s12951-022-01609-0
142. Chicón-Bosch M, Tirado OM. Exosomes in bone sarcomas: key players in metastasis. Cells. 2020;9(1):241. doi:10.3390/cells9010241
143. Uotani K, Fujiwara T, Ueda K, et al. Identification of ENO-1 positive extracellular vesicles as a circulating biomarker for monitoring of Ewing sarcoma. Cancer Sci. 2024;115(11):3660–3671. doi:10.1111/cas.16343
144. Di Paolo V, Paolini A, Galardi A, et al. Plasma-derived extracellular vesicles miR-335-5p as potential diagnostic biomarkers for fusion-positive rhabdomyosarcoma. J Exp Clin Cancer Res. 2024;43(1):282. doi:10.1186/s13046-024-03197-3
145. Crow J, Samuel G, Farrow E, et al. MicroRNA content of Ewing sarcoma derived extracellular vesicles leads to biomarker potential and identification of a previously undocumented EWS-FLI1 translocation. Biomark Insights. 2022;17:11772719221132693. doi:10.1177/11772719221132693
146. Cambier LD, Stachelek K, Triska M, et al. Extracellular vesicle-associated repetitive element DNAs as candidate osteosarcoma biomarkers. Sci Rep. 2021;11(1):94. doi:10.1038/s41598-020-77398-z
147. Sun N, Tran B, Peng ZS, et al. Coupling lipid labeling and click chemistry enables isolation of extracellular vesicles for noninvasive detection of oncogenic gene alterations. Adv Sci. 2022;9(14):2105853. doi:10.1002/advs.202105853
148. Xu YQ, Bao QY, Yu SX, et al. A novel microfluidic chip for fast, sensitive quantification of plasma extracellular vesicles as biomarkers in patients with osteosarcoma. Front Oncol. 2021:11709255. doi:10.3389/fonc.2021.709255.
149. Wen YJ, Li YT, Cheng SB, et al. Partition-less digital immunoassay using configurable topographic nanoarrays for extracellular vesicle diagnosis of Ewing sarcoma. Acs Nano. 2025;19(12):11973–11986. doi:10.1021/acsnano.4c16904
150. Han ZZ, Yi J, Yang Y, et al. SERS and MALDI-TOF MS based plasma exosome profiling for rapid detection of osteosarcoma. Analyst. 2021;146(21):6496–6505. doi:10.1039/d1an01163d
151. Atay S, Wilkey DW, Milhem M, Merchant M, Godwin AK. Insights into the proteome of gastrointestinal stromal tumors-derived exosomes reveals new potential diagnostic biomarkers. Mol Cell Proteomics. 2018;17(3):495–515. doi:10.1074/mcp.RA117.000267
152. Wei H, Chen J, Wang S, et al. A nanodrug consisting of doxorubicin and exosome derived from mesenchymal stem cells for osteosarcoma treatment in vitro. Int J Nanomed. 2019;14:8603–8610. doi:10.2147/ijn.S218988
153. Hu P, Ying J, Wang Y, et al. Extracellular vesicles derived from 3D cultured antler stem cells serve as a new drug vehicle in osteosarcoma treatment. Cell Transplant. 2023;32:9636897231219830. doi:10.1177/09636897231219830
154. Pitt JM, André F, Amigorena S, et al. Dendritic cell-derived exosomes for cancer therapy. J Clin Invest. 2016;126(4):1224–1232. doi:10.1172/jci81137
155. Luo S, Chen J, Xu F, Chen H, Li Y, Li W. Dendritic cell-derived exosomes in cancer immunotherapy. Pharmaceutics. 2023;15(8):2070. doi:10.3390/pharmaceutics15082070
156. Morisaki S, Onishi H, Morisaki T, et al. Dendritic cell-derived exosomes induce monocyte antigen-presentation and immune amplification in neoantigen vaccine therapy. Front Immunol. 2025;16:1565696. doi:10.3389/fimmu.2025.1565696
157. Baglio SR, Lagerweij T, Pérez-Lanzón M, et al. Blocking tumor-educated MSC paracrine activity halts osteosarcoma progression. Clin Cancer Res. 2017;23(14):3721–3733. doi:10.1158/1078-0432.Ccr-16-2726
158. Wang BD, Yu XJ, Hou JC, et al. Bevacizumab attenuates osteosarcoma angiogenesis by suppressing MIAT encapsulated by serum-derived extracellular vesicles and facilitating miR-613-mediated GPR158 inhibition. Cell Death Dis. 2022;13(3):272. doi:10.1038/s41419-022-04620-3
159. McNamara RP, Eason AB, Zhou YJ, et al. Exosome-encased nucleic acid scaffold chemotherapeutic agents for superior anti-tumor and anti-angiogenesis activity. Acs Bio Med Chem Au. 2022;2(2):140–149. doi:10.1021/acsbiomedchemau.1c00030
160. Wei H, Chen F, Chen J, et al. Mesenchymal stem cell derived exosomes as nanodrug carrier of doxorubicin for targeted osteosarcoma therapy via SDF1-CXCR4 axis. Int J Nanomed. 2022;17:3483–3495. doi:10.2147/ijn.S372851
161. Du JX, Meng XW, Yang MH, et al. NGR-modified CAF-derived exos targeting tumor vasculature to induce ferroptosis and overcome chemoresistance in osteosarcoma. Adv Sci. 2025;12(12). doi:10.1002/advs.202410918
162. Chen W, Li Z, Yu N, et al. Bone-targeting exosome nanoparticles activate Keap1 / Nrf2 / GPX4 signaling pathway to induce ferroptosis in osteosarcoma cells. J Nanobiotechnol. 2023;21(1):355. doi:10.1186/s12951-023-02129-1
163. Samal S, Panda GP, Shyamal S, Das K, Dash M. Surface engineered osteoblast-extracellular vesicles serve as an efficient carrier for drug and small RNA to actively target osteosarcoma. ACS Biomater Sci Eng. 2024;10(12):7466–7481. doi:10.1021/acsbiomaterials.4c00952
164. Yu LF, Fan GT, Wang QY, et al. In vivo self-assembly and delivery of VEGFR2 siRNA-encapsulated small extracellular vesicles for lung metastatic osteosarcoma therapy. Cell Death Dis. 2023;14(9):626. doi:10.1038/s41419-023-06159-3
165. Palinski W, Monti M, Camerlingo R, et al. Lysosome purinergic receptor P2X4 regulates neoangiogenesis induced by microvesicles from sarcoma patients. Cell Death Dis. 2021;12(9):797. doi:10.1038/s41419-021-04069-w
166. Lin S, Shu L, Guo Y, et al. Cargo-eliminated osteosarcoma-derived small extracellular vesicles mediating competitive cellular uptake for inhibiting pulmonary metastasis of osteosarcoma. J Nanobiotechnol. 2024;22(1):360. doi:10.1186/s12951-024-02636-9
167. Romano E, Perut F, Avnet S, et al. Mesenchymal stem cells-derived small extracellular vesicles and their validation as a promising treatment for chondrosarcoma in a 3D model in vitro. Biotechnol Bioeng. 2025;122(3):667–676. doi:10.1002/bit.28909
168. Yue X, Cui J, Ren S, et al. NK-cell-derived extracellular vesicles engineered to carry senolytics eliminate chemotherapy-induced senescent osteosarcoma cells. J Extracell Vesicles. 2025;14(7):e70123. doi:10.1002/jev2.70123
169. Wang Z, He Z, Wan J, Chen A, Cheng P, Zhu W. EphA2-specific microvesicles derived from tumor cells facilitate the targeted delivery of chemotherapeutic drugs for osteosarcoma therapy. J Nanobiotechnol. 2024;22(1):89. doi:10.1186/s12951-024-02372-0
170. Pan Y, Yang X, Zeng Z, et al. (124)I-labelled BMSC-derived extracellular vesicles deliver CRISPR/Cas9 ribonucleoproteins with a GFP-reporter system to inhibit osteosarcoma proliferation and metastasis. J Extracell Vesicles. 2025;14(7):e70130. doi:10.1002/jev2.70130
171. Chen WK, Lin WP, Yu NC, et al. Activation of dynamin-related protein 1 and induction of mitochondrial apoptosis by exosome-rifampicin nanoparticles exerts anti-osteosarcoma effect. Int J Nanomed. 2022;17:5431–5446. doi:10.2147/ijn.S379917
172. Min L, Shen J, Tu CQ, Hornicek F, Duan ZF. The roles and implications of exosomes in sarcoma. Cancer Metastasis Rev. 2016;35(3):377–390. doi:10.1007/s10555-016-9630-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.