Back to Journals » International Journal of Nanomedicine » Volume 21
Extracellular Vesicles in Gestational Diabetes Mellitus: Pathogenesis, Diagnosis, and Therapy
Authors Zhang X, Zhang Z, Jin L, Huang Y
Received 9 April 2026
Accepted for publication 2 July 2026
Published 16 July 2026 Volume 2026:21 615762
DOI https://doi.org/10.2147/IJN.S615762
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Farooq A. Shiekh
Xiaojuan Zhang,1 Ziwen Zhang,2 Limin Jin,3 Yimin Huang4
1Department of Pharmaceutics, College of Medicine, Jiaxing University, Jiaxing, People’s Republic of China; 2Department of Obstetrics, Jiaxing Maternity and Child Health Care Hospital, Affiliated Women and Children Hospital, Jiaxing University, Jiaxing, People’s Republic of China; 3Department of Clinical Laboratory, Jiaxing Maternity and Child Health Care Hospital, Affiliated Women and Children Hospital, Jiaxing University, Jiaxing, People’s Republic of China; 4Department of Central Laboratory, Jiaxing Maternity and Child Health Care Hospital, Affiliated Women and Children Hospital, Jiaxing University, Jiaxing, People’s Republic of China
Correspondence: Limin Jin, Department of Clinical Laboratory, Jiaxing Maternity and Child Health Care Hospital, Affiliated Women and Children Hospital, Jiaxing University, Jiaxing, People’s Republic of China, Email [email protected] Yimin Huang, Department of Central Laboratory, Jiaxing Maternity and Child Health Care Hospital, Affiliated Women and Children Hospital, Jiaxing University, Jiaxing, People’s Republic of China, Email [email protected]
Abstract: Gestational diabetes mellitus (GDM) is the most common metabolic complication of pregnancy, with a continuously rising global prevalence. However, current diagnostic and therapeutic strategies are limited by delayed detection and insufficient targeting. Extracellular vesicles (EVs), as nanoscale messengers that mediate intercellular communication, have recently been implicated in GDM pathogenesis and exhibit dual potential as liquid biopsy biomarkers and drug delivery vehicles. This review systematically integrates research findings on EVs in GDM, constructs an EV-mediated multi-organ crosstalk network framework, and comparatively evaluates the diagnostic and therapeutic potential of EVs derived from the placenta, adipose tissue, blood, breast milk, and urine. And further discussed current challenges and future directions for clinical translation of EV-based strategies. It provides an integrative perspective on the pathogenesis, precision diagnosis, and treatment of GDM and offers critical guidance for advancing the clinical translation of EV-based strategies.
Keywords: extracellular vesicles, gestational diabetes mellitus, multi-organ crosstalk, biomarkers, therapeutic strategies
Introduction
Gestational diabetes mellitus (GDM) is defined as glucose intolerance that is first recognized during pregnancy. It is one of the most common metabolic complications of pregnancy.1 In recent years, the global prevalence of GDM has been rising consistently, affecting 5% to 17% of pregnancies, and this upward trend is expected to continue.2,3 GDM affects approximately 21 million live births annually. Prevalence estimates range from 5% to 25%, depending on diagnostic criteria, ethnic background, and socioeconomic factors.4–6 GDM not only increases the risk of maternal preeclampsia (PE), cesarean delivery, and future type 2 diabetes,7–9 but also significantly increases the incidence of fetal macrosomia, neonatal hypoglycemia, and long-term metabolic disorders.6,10
The diagnosis of GDM has traditionally relied on the oral glucose tolerance test (OGTT), which is typically performed between 24 and 28 weeks of gestation.11 However, this approach has several limitations. First, the diagnosis occurs late in pregnancy, and exposure to hyperglycemia in the first trimester may already have adverse effects on fetal development. Evidence suggests that early hyperglycemia is associated with severe neonatal complications.3,12 Second, diagnostic criteria are not fully standardized, leading to diagnostic heterogeneity. For example, the Diabetes in Pregnancy Study Group India (DIPSI) method widely used in India has lower sensitivity compared to the World Health Organization (WHO) criteria, which may result in many missed cases.13 Third, the OGTT is cumbersome, requiring overnight fasting and multiple blood draws, which reduces patient compliance. Although alternative markers such as glycated hemoglobin and fasting plasma glucose have been explored for first trimester screening, their limited sensitivity and risk of false negatives preclude their use as replacement for the OGTT as a standard diagnostic tool.3 Emerging technologies such as continuous glucose monitoring, vibrational spectroscopy, and machine learning models show potential for early detection, but their high cost and insufficient clinical validation limit current clinical application.14–16 Therefore, developing early, non-invasive, and highly sensitive diagnostic tools remains a critical goal in GDM.
Current management of GDM is based primarily on lifestyle interventions, including medical nutrition therapy and regular physical activity. Approximately 70 to 85% of women achieve glycemic control through non-pharmacological interventions.3 For women who do not meet glycemic targets, insulin is widely recognized as the first-line pharmacological treatment, with strong evidence supporting its safety during pregnancy.17 However, insulin therapy has limitations, such as the burden of injections and suboptimal patient acceptance, leading to suboptimal adherence in some patients.18 Furthermore, the safety of oral antidiabetic agents such as metformin and glyburide during pregnancy remains controversial. A network meta-analysis showed that metformin effectively controls blood glucose and reduces the risk of neonatal hypoglycemia and preeclampsia, but it may increase the risk of small-for-gestational-age infants.19 Glyburide is associated with a higher incidence of neonatal hypoglycemia.18 Thus, the development of novel, safe, and convenient therapeutic strategies represents a critical priority for GDM management.
Extracellular vesicles (EVs) are nanoscale membrane-bound vesicles released by cells, ranging from 30 to 1000 nm in diameter. They carry bioactive molecules such as proteins, lipids, mRNA, and microRNA (miRNA), and mediate intercellular communication.20,21 According to the MISEV 2023 guidelines from the International Society for Extracellular Vesicles, these vesicles are collectively referred to as small extracellular vesicles (sEVs) unless their endosomal origin is explicitly demonstrated.22 EVs derived from the maternal-fetal interface are involved in embryo implantation, placental development, and the establishment of maternal-fetal immune tolerance. Studies have shown that EVs from the fetal-placental unit deliver fetal antigens to maternal secondary lymphoid tissues, where they are recognized by maternal T cells, representing a key mechanism for maintaining maternal-fetal immune tolerance.23 Placenta-derived EVs enter the maternal circulation as early as six weeks of gestation, and their concentration increases with gestational age. Their molecular cargo reflects the state of the placental microenvironment.24 Recently, EVs have emerged as a focus in liquid biopsy research due to their stability in body fluids, disease-specific cargo, and ability to reflect the pathological state of cells.25 In GDM, the molecular signatures of EVs are closely associated with insulin resistance, inflammatory responses, and placental dysfunction, highlighting their potential as early diagnostic biomarkers.26 Furthermore, EVs from various sources offer promising therapeutic options for GDM, owing to their low immunogenicity, potential for engineering, and flexible routes of administration.25
This review systematically summarizes the role of EVs in the pathogenesis of GDM, recent advances in diagnostic biomarkers, and the therapeutic potential of EVs from different sources, and further discusses the path toward clinical translation. To systematically capture the research progress in this field, we searched the PubMed and Web of Science databases for literature published up to 2026. The search combined the terms “extracellular vesicles” or “exosomes” with “gestational diabetes mellitus” or “GDM”. Inclusion criteria were original research articles and high-quality reviews addressing EVs in GDM pathogenesis, diagnostic biomarkers, or therapeutic strategies. We prioritized studies that provided mechanistic insights or quantitative diagnostic performance, and studies that focused only on other pregnancy complications without addressing GDM were excluded.
In recent years, several reviews have summarized EV research in GDM. However, most have focused on placental mechanisms,27 other specific complications,28 or diagnostic applications.26 Compared with previously published reviews, our study offers the following contributions. (1) Systematic integration of a pathological framework of EV-mediated multi-organ crosstalk involving β-cells, placenta, liver, and adipose tissue. (2) Comprehensive coverage of both diagnostic and therapeutic applications, with a detailed discussion of the therapeutic strategies of EVs from various sources and their clinical translation prospects. (3) Emphasis on the key challenges in the clinical translation of EVs, including quality control, safety, and large-scale manufacturing. Through this synthesis, this review aims to provide a new perspective on the precise diagnosis and treatment of GDM.
Biological Basis of EVs
Biogenesis and Characteristics of EVs
EVs are a general term for membrane-bound vesicles released by cells. They can be classified into different subtypes based on their biogenesis pathway, size, and release mechanism.29 In the MISEV 2023 guidelines, the International Society for Extracellular Vesicles recommends that vesicles smaller than 200 nm in diameter be referred to as sEVs unless their endosomal origin is clearly demonstrated, vesicles that originate from endosomal multivesicular bodies are traditionally termed exosomes.22
The size of EVs typically ranges from 30 to 1000 nm. Among these, sEVs are predominantly 30 to 150 nm in diameter and exhibit a characteristic cup-shaped morphology under transmission electron microscopy. Nanoparticle tracking analysis can be used to measure their size distribution and concentration.22,30 EVs carry a variety of bioactive molecules, including proteins, lipids, mRNA, miRNA, circular RNA (circRNA), and DNA.31 Their molecular cargo reflects the physiological or pathological state of the originating cell. EVs derived from different cell types have characteristic protein and nucleic acid compositions.32
EV biogenesis relies on two main pathways: the endosomal sorting complexes required for transport (ESCRT)-dependent pathway and the ESCRT-independent pathway.33 In the ESCRT-dependent pathway, the plasma membrane invaginates to form early endosomes. The endosomal membrane then buds inward to form intraluminal vesicles (ILVs), and the endosome matures into a multivesicular body (MVB). When MVBs fuse with the plasma membrane, ILVs are released into the extracellular space and become EVs.34 The ESCRT machinery consists of four subcomplexes, including ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III. These subcomplexes cooperatively recognize and sort cargo proteins and mediate endosomal membrane budding and vesicle formation.35 The ESCRT-independent pathway involves lipid metabolism, particularly the synthesis of ceramides. Ceramides can induce membrane domains formation and curvature, promoting ILV generation.36 Tetraspanins (such as CD9, CD63, and CD81) also contribute to ESCRT-independent vesicle formation by interacting to form microdomains and participating in cargo sorting and vesicle biogenesis.37
Physiological Roles of EVs During Pregnancy
EVs play key roles in the physiological processes of pregnancy. They mediate maternal-fetal crosstalk, placental development, and the maintenance of immune tolerance (Figure 1).38,39 The maternal-fetal interface represents the most complex site of cellular communication during pregnancy. EVs serve as important information carriers, transmitting signals between the mother and the fetus.40
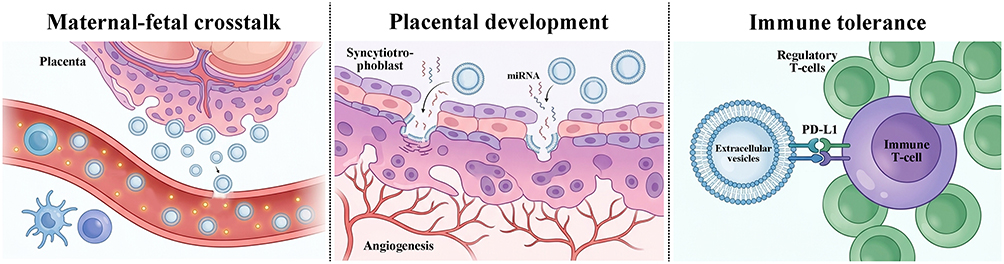 |
Figure 1 Role of extracellular vesicles (EVs) in maternal-fetal crosstalk, placental development, and immune tolerance during pregnancy. Placenta-derived enter the maternal circulation and their concentration increases with advancing gestational age. By delivering fetal antigens to maternal secondary lymphoid tissues, these EVs initiate maternal–fetal crosstalk and promote immune tolerance. Syncytiotrophoblast-derived EVs carry specific miRNAs and proteins that regulate trophoblast cell fusion and syncytialization, thereby maintaining the placental barrier and facilitating placental angiogenesis. In immune tolerance, EVs express surface molecules including PD-L1 ligands. Engagement of PD-L1 with PD-1 on maternal T cells inhibits T cell activation and induces the differentiation of regulatory T cells. Together with EV-encapsulated miRNAs that modulate maternal immune cell functions, these mechanisms collectively establish a protective barrier against fetal immune rejection. |
In maternal-fetal communication, placenta-derived exosomes enter the maternal circulation as early as the sixth week of gestation, and their concentration increases with gestational age.41 The molecular cargo of these exosomes reflects the status of the placental microenvironment and can be internalized by maternal immune cells, thereby modulating maternal adaptation to pregnancy.42,43 Studies have shown that exosomes from the fetoplacental unit deliver fetal antigens to maternal secondary lymphoid tissues, where they are recognized by maternal T cells. This mechanism is essential for establishing maternal immune tolerance to fetal antigens.44
In placental development, EVs regulate trophoblast proliferation, differentiation, and invasion. Syncytiotrophoblast-derived exosomes carry specific miRNAs and proteins that regulate trophoblast fusion and syncytialization, thereby maintaining the placental barrier function.45 Placenta-derived EVs also promote angiogenesis by carrying pro-angiogenic molecules. In normal pregnancy, placenta-derived EVs stimulate endothelial cell migration and tube formation, contributing to the formation and remodeling of placental vasculature. In pregnancy complications such as preeclampsia, the angiogenic balance of placenta-derived EVs is disrupted, resulting in vascular dysfunction.46
In the maintenance of immune tolerance, placenta-derived EVs play a central role. Syncytiotrophoblast-derived EVs carry a unique immunosuppressive protein signature, including NKG2D receptor ligands, apoptosis-inducing molecules, and programmed death ligand 1 (PD-L1).47 These molecules can inhibit the cytotoxicity of maternal natural killer cells, induce apoptosis of activated immune cells, and promote the differentiation of regulatory T cells, thereby protecting the fetus from maternal immune system attack.47 Studies have demonstrated that placenta-derived bEVs express PD-L1 on their surface. Upon interaction with maternal T cells, these PD-L1 molecules bind to programmed death receptor 1 (PD-1) expressed on T cells, leading to the inhibition of T cell activation and the promotion of regulatory T cell differentiation. This PD-L1/PD-1 signaling axis is critical for establishing and maintaining maternal-fetal immune tolerance.48,49 In addition, C19MC family miRNAs carried by placenta-derived EVs regulate maternal immune cell function, further reinforcing immune tolerance.50 Together, these mechanisms establish an immunological barrier that protects the fetus from immune rejection.
EVs Mediate the Pathogenesis of GDM
In normal pregnancy, EVs are involved in maternal-fetal crosstalk, placental development, and immune tolerance maintenance. However, in GDM, both the molecular cargo and function of EVs undergo characteristic alterations, mediating pathological processes across multiple organs (Figure 2).
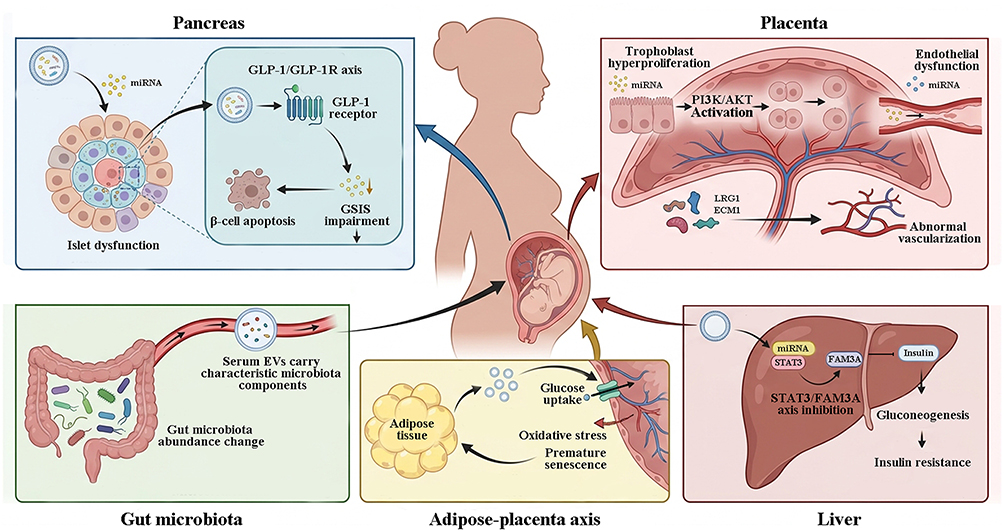 |
Figure 2 Extracellular vesicle-mediated multi-organ pathological network in gestational diabetes mellitus. (1) GDM‑derived EVs carry specific miRNAs that activate the GLP‑1/GLP‑1R axis, leading to premature functional maturation of pancreatic β‑cells and concurrently promoting β‑cell apoptosis. Further results in impaired glucose‑stimulated insulin secretion (GSIS) and failure of islet compensation. (2) Upregulation of EV‑associated miR‑135a‑5p activates the PI3K/AKT pathway, promoting trophoblast hyperproliferation, whereas downregulation of miR‑140‑3p and miR‑574‑3p impairs endothelial cell function. In addition, EVs carry LRG1 and ECM1 proteins that induce aberrant placental angiogenesis and disrupt placental barrier integrity. (3) Circulating EVs contain miRNAs that target the STAT3/FAM3A axis, suppressing insulin signaling, enhancing hepatic gluconeogenesis, and exacerbating hepatic insulin resistance. (4) Adipose tissue‑derived EVs transport miRNAs that promote placental glucose uptake, while NOX4 induces placental oxidative stress and premature senescence, establishing a positive feedback loop of metabolic dysregulation between adipose tissue and the placenta. (5) serum EVs from GDM patients carry characteristic microbial components, with increased abundance of Actinobacteria and Firmicutes and decreased abundance of Proteobacteria and Bacteroidetes. Gut microbiota‑derived EVs may participate in GDM pathogenesis via the gut–placenta axis. Collectively, EV‑mediated aberrant inter‑organ communication across these multiple sites forms the complex pathological network underlying GDM. The flat-ended black arrow represent inhibition. |
EVs Mediate β-Cell Dysfunction and Islet Developmental Abnormalities
The compensatory function of pancreatic β-cells is a key mechanism for maintaining normal glucose homeostasis during pregnancy. In normal pregnancy, β-cells counteract physiological insulin resistance through enhanced proliferation and insulin secretion. Failure of this compensation leads to the development of GDM.3 β-cell dysfunction, characterized by impaired glucose-stimulated insulin secretion, increased apoptosis, and aberrant islet development, is commonly observed in patients with GDM.2
Functional studies have demonstrated that EVs directly participate in the regulation of glucose homeostasis during pregnancy. EVs derived from healthy pregnant women promote glucose-stimulated insulin secretion from islets and induce peripheral insulin resistance. In contrast, EVs from women with GDM fail to promote insulin secretion and instead exacerbate insulin resistance.51 Continuous infusion of GDM-derived EVs into pregnant mice via a mini-osmotic pump for four days successfully induced glucose intolerance, whereas EVs from healthy pregnant women stimulated glucose-stimulated insulin secretion from islets.52 EVs derived from GDM patients can directly impair β-cell function. Studies have shown that EVs isolated from umbilical cord blood of GDM patients induce glucose intolerance and hyperinsulinemia in healthy pregnant mice. The mechanism involves activation of the glucagon-like peptide-1 (GLP-1)/GLP-1 receptor axis within the islets, stimulating premature functional maturation of β-cells through the Akt-mTORC1-pS6 pathway, thereby disrupting normal islet developmental timing.52 This finding provides a mechanistic explanation for the increased risk of diabetes in offspring of mothers with GDM. However, as this study is based on animal models, whether the same process occurs in humans remains to be verified.
Furthermore, specific miRNAs play critical roles in EV-mediated β-cell injury. In vitro and animal models have shown that placental exosomes from GDM patients are enriched in miR-320b, which increases β-cell apoptosis and reduces glucose-stimulated insulin secretion by targeting downstream genes.53 In early pregnancy, the expression of maternal serum-derived exosomal miR-149-3p and miR-455-3p is closely associated with pancreatic β-cell function, and these miRNAs are involved in regulating insulin secretion and signaling pathways.54 Additionally, miR-7-19488 is significantly upregulated in GDM-derived EVs. This miRNA targets the mRNA of phosphoinositide-3-kinase regulatory subunit 2 (PIK3R2), thereby interfering with signaling pathways that regulate islet development and causing aberrant islet development in the offspring.55
EVs Mediate Placental Dysfunction
The placenta is a central organ for maintaining pregnancy, responsible for maternal-fetal substance exchange, hormone synthesis, and immune regulation. Normal placental development depends on the proliferation, migration, invasion, and angiogenesis of trophoblast cells.39 In GDM, the placenta exhibits pathological changes, including excessive villous vascularization, increased syncytial knot formation, and elevated release of inflammatory factors. These changes are closely associated with insulin resistance and maternal metabolic dysregulation.56,57
EVs from multiple sources can mediate placental dysfunction. Placenta-derived EVs participate in trophoblast proliferation, differentiation, and angiogenesis, maintaining placental barrier function. Dysregulation of these processes underlies placental dysfunction in GDM. miR-135a-5p expression is significantly increased in placenta-derived EVs from GDM patients. In vitro functional experiments demonstrate that this miRNA activates the phosphoinositide-3-kinase/protein kinase B (PI3K/AKT) signaling pathway by targeting sirtuin 1 (SIRT1), thereby promoting excessive proliferation and migration of trophoblast cells and disrupting normal placental development.58 These findings suggest that miR-135a-5p may be contribute to placental dysfunction, but the causal relationship remains to be established through in vivo gene intervention experiments. Conversely, miR-140-3p and miR-574-3p are downregulated in placenta-derived EVs from GDM patients. These two miRNAs inhibit the proliferation, migration, and tube formation of human umbilical vein endothelial cells by targeting vascular endothelial growth factor.59 Chemerin induces placental inflammation by recruiting macrophages and promoting the release of interleukin-18 and interleukin-1β, thereby downregulating miR-140-3p and miR-574-3p expression and contributing to placental dysfunction in GDM.59
CircRNAs carried by exosomes also participate in placental dysfunction. circ-PNPT1 is highly expressed in placental tissues from GDM patients and in trophoblast cells exposed to high glucose. It inhibits trophoblast cell proliferation, migration, and invasion while promoting apoptosis by sponging miR-889-3p and upregulating PAK1 expression. This circRNA can be packaged into EVs and transferred to surrounding untreated cells, mediating the spread of pathological signals.60
Adipose tissue-derived EVs are also involved in placental function regulation. In pregnancies complicated by obesity, adipocytes-derived exosomes carry NADPH oxidase 4 (NOX4). When delivered to trophoblast cells, they induce DNA oxidative damage, cellular senescence, and impaired proliferation and invasion, ultimately leading to premature placental senescence and adverse pregnancy outcomes.61 miR-515-5p expression is upregulated in adipose tissue-derived EVs from GDM patients. This miRNA promotes glucose uptake in placental trophoblast cells and is associated with excessive fetal growth.62,63 Proteomic analysis revealed that 122 proteins are upregulated in adipose tissue-derived exosomes from GDM patients. These proteins are primarily enriched in pathways associated with mitochondrial dysfunction and oxidative phosphorylation, and they upregulate the expression of aldolase B, phosphoglycerate kinase 2, and glucokinase in placental cells, thereby affecting placental glucose metabolism.64
Umbilical cord blood-derived EVs also contribute to abnormal placental angiogenesis in GDM. Leucine-rich α-2-glycoprotein 1 (LRG1) and extracellular matrix protein 1 (ECM1) are significantly upregulated in umbilical cord blood exosomes from GDM patients. These two proteins promote endothelial cell proliferation and migration, thereby inducing abnormal placental vascularization and contributing to macrosomia in GDM.65,66
EVs Mediate Hepatic Insulin Resistance
The liver is a central organ for maintaining glucose homeostasis, regulating blood glucose levels through glycogen synthesis, gluconeogenesis, and glucose output.67 The hepatic insulin signaling pathway is impaired in patients with GDM, leading to increased gluconeogenesis and elevated hepatic glucose output, which exacerbate maternal hyperglycemia.68
Circulating EVs can directly induce hepatic insulin resistance. In vitro experiments revealed that when hepatocytes were co-cultured with EVs isolated from the plasma of GDM patients, they exhibited increased gluconeogenesis, reduced energy metabolism, and upregulated oxidative stress.69 Mechanistic studies identified miR-1299 as the predominant differentially expressed miRNA in circulating EVs from GDM patients. This miRNA directly targets the 3′ untranslated region of signal transducer and activator of transcription 3 (STAT3) and inhibits the insulin signaling pathway through regulation of the STAT3/ FAM3A (family with sequence similarity 3 member A) axis.69 Functional experiments confirmed that miR-1299 increases insulin resistance by modulating mitochondrial function and oxidative stress in hepatocytes. In animal experiments, mice injected with EVs from GDM patients developed glucose intolerance, insulin resistance, and downregulation of hepatic STAT3 and FAM3A expression, further validating the EV-mediated mechanism of hepatic insulin resistance.69
EVs Mediate Adipose-Placenta-Maternal Metabolism Axis
Metabolic regulation during pregnancy involves coordinated crosstalk among multiple organs, including adipose tissue, placenta, and liver. EVs serve as important mediators of inter-organ communication and play a critical role in GDM pathology.70
Adipose tissue-derived EVs participate in the regulation of placental glucose metabolism. The number of adipose tissue-derived exosomes is significantly increased in GDM patients, and this increase positively correlates with fetal birth weight.71 Proteomic analysis revealed altered expression of proteins involved in sirtuin signaling, oxidative phosphorylation, and mechanistic target of rapamycin (mTOR) signaling pathways in adipose tissue-derived exosomes from GDM patients. Functional experiments confirmed that, compared with exosomes from healthy pregnant women, exosomes from GDM patients increased the expression of genes related to glycolysis and gluconeogenesis in placental cells, suggesting that adipose tissue-derived exosomes contribute to GDM pathology by regulating placental metabolism.71
Placenta-derived EVs also participate in maternal metabolic regulation. The number of syncytiotrophoblast-derived EVs released from the placenta is significantly increased in GDM patients, and these EVs carry active dipeptidyl peptidase IV (DPPIV) on their surface.72 DPPIV is a key enzyme that degrades GLP-1,73 which regulates glucose-dependent insulin secretion. DPPIV activity is increased eightfold in placenta-derived EVs from GDM patients compared with that in EVs from healthy pregnant women, and these EVs can degrade GLP-1 in vitro, suggesting that placenta-derived EVs may influence maternal insulin secretion through degradation of GLP-1, thereby contributing to insulin resistance in GDM.72 Together, these findings establish an EV-mediated adipose-placenta-maternal metabolism regulatory network.
Bacterial EVs Participate in GDM Pathology
Gut microbiota dysbiosis is closely associated with the development and progression of GDM, and bacterial extracellular vesicles (bEVs) serve as important mediators of microbiota-host communication.74 bEVs are nanoscale vesicles secreted by both Gram-negative and Gram-positive bacteria. They carry active molecules such as lipopolysaccharides, flagellin, nucleic acids, and metabolites, and can cross the intestinal barrier to enter the circulation, affecting distal organ function.75 A recent clinical cohort study has revealed that maternal factors during pregnancy shape the taxonomic profiles of bEVs. The study found that antibiotic use during pregnancy was associated with reduced species diversity of bEVs. Furthermore, in women with overweight/obesity, GDM, or gestational weight gain exceeding 15 kg, bEVs in both amniotic fluid and feces were markedly altered, and numerous bEVs were depleted in these individuals.76 This work provides clinical evidence directly linking maternal factors to shifts in the bEV landscape, suggesting that bEVs represent a critical link between maternal metabolic dysregulation in pregnancy and host responses.
Key evidence indicates that bEVs can cross biological barriers and directly communicate with the fetus. bEVs can be detected in the amniotic fluid of healthy pregnant women, and their molecular characteristics resemble those of bEVs found in the maternal gut microbiota.77 In mouse experiments, bEVs derived from the maternal gut microbiota were found to reach the amniotic cavity, and these bEVs may play a critical role in priming the prenatal immune system.77
Furthermore, characteristic microbiota components are present in circulating EVs from GDM patients. 16S rRNA sequencing analysis of serum EVs from GDM patients and normal pregnant women revealed increased relative abundances of Actinobacteria and Firmicutes, decreased abundances of Proteobacteria and Bacteroidetes, and an elevated Firmicutes/Bacteroidetes ratio in the GDM group.78 Functional prediction analysis showed that the microbial functions in serum EVs from both groups were enriched in amino acid metabolism, carbohydrate metabolism, energy metabolism, and cofactor and vitamin metabolism pathways. This finding provides the first evidence that circulating EVs from GDM patients carry microbiota-derived components, suggesting that bEVs may participate in GDM pathology.78
In vitro experiments have further demonstrated that pathogenic bEVs can directly impair placental function. Outer membrane vesicles derived from Porphyromonas gingivalis are internalized by human trophoblast cells, where they inhibit trophoblast migration and invasion, and induce placental metabolic reprogramming.79,80 In animal experiments, treatment with P. gingivalis outer membrane vesicles resulted in reduced placental and fetal weights and decreased expression of the glucose transporter GLUT1 in the placenta.80 Additionally, probiotic-derived bEVs play protective roles in metabolic regulation. For instance, Akkermansia muciniphila-derived bEVs enhance intestinal tight junction function, reduce gut permeability, and improve glucose tolerance and insulin resistance in high-fat diet-induced diabetic mice.81,82 A recent study has further confirmed that A. muciniphila-derived bEVs significantly protect streptozotocin (STZ)-induced diabetic mice by regulating glucose homeostasis, attenuating oxidative stress, and inducing immune tolerance, thereby effectively reducing blood glucose levels and ameliorating insulin resistance.83 In a mouse model of PE, A. muciniphila-derived bEVs were transported from the gastrointestinal tract to the placenta and attenuated preeclamptic symptoms by promoting trophoblast invasion and spiral artery remodeling through activation of the EGFR-PI3K-AKT signaling pathway.84 However, no study to date has directly established a causal relationship between alterations in bEVs and the development of GDM. The specific mechanisms by which bEVs contribute to GDM, the key effector molecules involved, and their therapeutic potential remain to be further elucidated.
To integrate these mechanisms, we summarize the key molecules cargoes and pathological effects of EVs across multiple organs (Table 1). However, it should be noted that current evidence is largely based on correlational or preclinical exploratory studies. The causal relationship between EV cargo and disease progression remains to be determined. Most studies rely on in vitro cell experiments or animal models. These models cannot fully recapitulate the complex metabolic environment of human GDM. For example, while EVs from GDM patients can induce glucose intolerance in pregnant mice,52 significant differences in placental structure and metabolic regulation between mice and humans limit the clinical translation of such findings. Research on placental dysfunction has been largely descriptive, cataloguing differential expression of EV-associated miRNAs or proteins.59,64–66 The mechanism by which these molecules lead to functional changes remains unclear. Moreover, investigations into bEVs have merely reported associations between microbial components in serum EVs and GDM,78 without functional experiments that demonstrate direct effects of bEVs on the placenta or pancreas. Although multiple EV molecules involved in GDM pathology have been identified (Table 1), it remains unclear whether EVs from different organs have synergistic or cascading effects, or how they dynamically participate in the onset and progression of GDM. In summary, large-scale prospective cohort studies, combined with gain- and loss-of-function approaches and mechanistic validation in human participants, are needed to establish causal links between specific EV cargo and GDM progression.
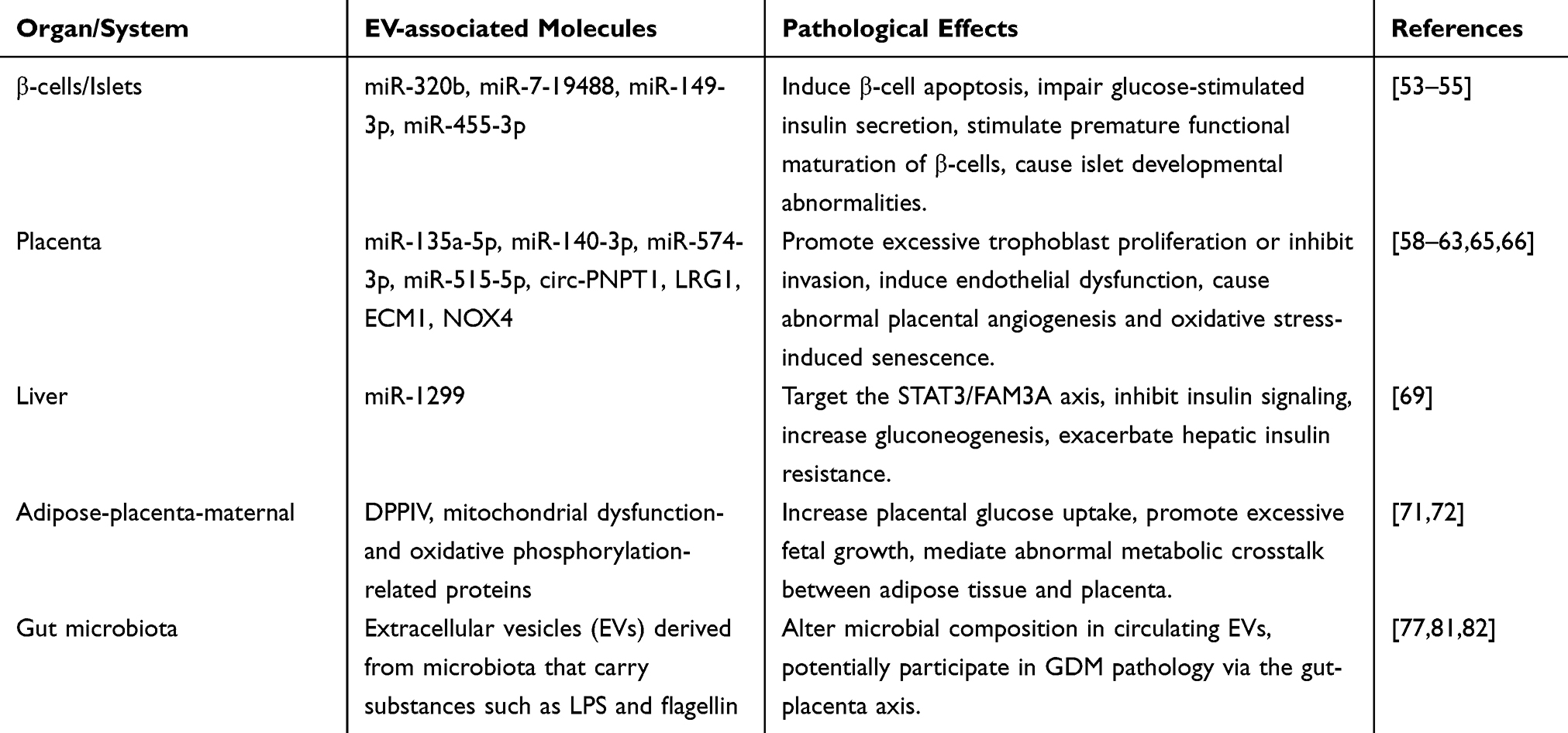 |
Table 1 Summary of Extracellular Vesicles -Mediated Pathological Mechanisms in GDM Across Multiple Organs |
Emerging Roles of EVs in GDM Diagnosis and Therapy
Advantages of EVs as Liquid Biopsy Biomarkers
Liquid biopsy is a non-invasive technique for disease diagnosis and monitoring through the detection of circulating biomarkers in body fluids.85 EVs exhibit unique advantages as liquid biopsy biomarkers. First, EVs are widely and stably present in body fluids such as blood, urine, saliva, and breast milk.86 Their lipid bilayer membrane structure protects internal nucleic acids and proteins from degradation, making them suitable for long-term storage and transport.87 Second, the molecular cargo carried by EVs reflects the physiological and pathological state of their originating cells and exhibits tissue specificity. Placenta-derived EVs can be detected in the maternal circulation as early as six weeks of gestation, and their concentration increases with gestational age, providing a time window for early diagnosis of pregnancy-related disorders.26 Third, changes in exosomal molecular profiles often precede the appearance of clinical symptoms, enabling early warning of disease.88 Additionally, detecting specific EV subpopulations from defined cellular origins allows precise localization of disease sites, overcoming the limited tissue specificity of traditional serum biomarkers.89 EVs hold considerable promise as liquid biopsy biomarkers in GDM, and multiple studies have identified characteristic changes in exosomal molecular cargo in patients with GDM.90,91 The concentration of placenta-derived EVs in the maternal circulation is significantly higher in patients with GDM than in normal pregnancies, with increases of approximately 2.2-fold, 1.5-fold, and 1.8-fold at 11–14 weeks, 22–24 weeks, and 32–36 weeks of gestation, respectively. These EVs induce endothelial cells to release pro-inflammatory cytokines, and changes in their concentration are closely associated with disease progression.70 However, most of these studies are based on small cohorts, and EV isolation and quantification methods have not yet been standardized, which limits the comparability of results across studies. Therefore, although the finding of an increased EV concentration holds clinical potential, it requires validation through large-scale multicenter studies.
Non-Coding RNA Based Molecular Markers
miRNAs are a class of non-coding RNAs approximately 22 nucleotides in length that participate in various physiological and pathological processes by regulating gene expression.92 miRNAs carried by EVs exhibit high stability and high abundance, making them ideal diagnostic biomarkers. Multiple studies have revealed characteristic alterations in miRNA expression profiles in circulating EVs from GDM patients. In plasma EVs from GDM patients, miR-423-5p expression is upregulated, while miR-122-5p, miR-148a-3p, miR-192-5p, and miR-99a-5p are downregulated. The combination of these five miRNAs achieved an area under the curve (AUC) of 0.82 for the early prediction of GDM. Further studies have shown that these miRNAs are involved in the regulation of the insulin and AMPK signaling pathways, which are closely associated with metabolic dysregulation in GDM.93
The miRNA expression profiles in circulating EVs from GDM patients exhibit dynamic changes across gestation. In the first trimester, 92% of the detected miRNAs in plasma exosomes from GDM patients were significantly elevated compared to those in normal pregnancies, while no significant differences were observed between the two groups in the second trimester, suggesting that the first trimester is the optimal time window for exosomal miRNA detection.90 Ten miRNAs (including miR-122-5p, miR-132-3p, miR-136-5p, miR-210-3p, and miR-29a-3p) in the first trimester serum EVs were significantly increased in GDM patients, and bioinformatics analysis indicated that these miRNAs are involved in trophoblast proliferation and differentiation as well as insulin secretion and glucose transport regulation.94
miRNAs in EVs from various body fluids have been found to have diagnostic value. In urine-derived placental EVs from GDM patients, the expression of miR-516-5p, miR-517-3p, miR-518-5p, miR-222-3p, and miR-16-5p was downregulated in the third trimester. These miRNAs are involved in the regulation of multiple metabolic pathways, suggesting that urinary EVs may serve as a non-invasive sample source for GDM diagnosis.95 The abundance of miR-148a, miR-30b, let-7a, and let-7d was reduced in breast milk exosomes from GDM patients, with miR-148a showing a negative correlation with infant body weight, body fat percentage, and fat mass, suggesting that breast milk EV miRNAs may serve as biomarkers for evaluating the impact of GDM on offspring.96 The expression of maternal serum-derived exosomal miR-149-3p and miR-455-3p in early pregnancy is closely associated with pancreatic β-cell function, and these miRNAs are involved in insulin secretion and signaling, lipolysis, and adipokine signaling pathways.54 Furthermore, placenta-derived exosomal miR-3665 and miR-6727-5p regulate genes involved in pregnancy immune tolerance and pathways associated with placental dysfunction.54 These findings suggest that early-pregnancy serum exosomal miRNA profiles can reflect pancreatic β-cell functional status and hold potential for the early prediction of GDM.
It is worth noting that the vast majority of the above studies lack external validation, and the reported AUC values only reflect diagnostic performance within a single cohort. Without independent cohort validation, the true predictive value of these biomarkers remains uncertain. Furthermore, differences in EV isolation methods, sequencing platforms, and data analysis workflows across studies have led to poor consistency in the lists of candidate miRNAs. There is an urgent need for multicenter, prospective validation studies using standardized protocols.
CircRNAs refer to non-coding RNAs with covalently closed loop structures that exhibit high stability and tissue specificity and are abundant in EVs.97 Jiang et al revealed that the expression of hsa_circRNA_0039480 and hsa_circRNA_0026497 is significantly upregulated in plasma exosomes from GDM patients and is positively correlated with oral glucose tolerance test levels The AUC of hsa_circRNA_0039480 for diagnosing GDM was 0.704 in the first trimester and reached 0.898 in the second trimester. The combination of these two circRNAs achieved an AUC of 0.754 for diagnosis in the first trimester, demonstrating promising early diagnostic potential.98
CircRNA expression profiling in umbilical cord blood EVs revealed that 229 circRNAs were significantly upregulated and 278 circRNAs were downregulated in GDM patients. These differentially expressed circRNAs are involved in metabolic processes, growth, and developmental regulation pathways, and most circRNAs possess miRNA-binding sites, suggesting their potential involvement in GDM pathology through miRNA sponging.99 circ-PNPT1 is highly expressed in placental tissues from GDM patients and in high-glucose-induced trophoblast cells. It can be packaged into EVs and transferred to surrounding cells. This circRNA inhibits trophoblast cell proliferation, migration, and invasion while promoting apoptosis by sponging miR-889-3p and upregulating PAK1 expression.60 The aberrant expression of circ-PNPT1 not only participates in GDM pathogenesis but also provides a theoretical basis for its use as a diagnostic biomarker. However, studies on these circRNA biomarkers also lack external validation and involve small sample sizes. Moreover, circRNA detection methods have not yet been standardized, which may affect the reproducibility of the results.
Long non-coding RNAs (lncRNAs) are non-coding RNAs longer than 200 nucleotides that are abundant in EVs and participate in various gene expression regulatory processes.100 The expression profiles of mRNAs and lncRNAs in umbilical cord blood exosomes from GDM patients are significantly altered, and these differentially expressed lncRNAs may participate in GDM development and fetal growth by regulating related gene expression.101 Although research on lncRNAs as diagnostic biomarkers for GDM is still in its early stages, their tissue specificity and disease-associated changes suggest potential diagnostic value.
Protein Based Clinical Markers
Proteomic analysis provides a rich resource for EV-based diagnostic biomarkers in GDM. Proteomic analyses of total plasma EVs and placenta-derived EVs from GDM patients have revealed that differentially expressed proteins are primarily involved in complement cascades, immune regulation, inflammation, and coagulation pathways. At 12–16 weeks of gestation, 36 differentially expressed proteins were identified in total plasma EVs, and 34 were identified in placenta-derived sEVs.102 Hub proteins identified from these analyses hold significant value for GDM prediction and diagnosis.
Proteomic analysis of serum EV from GDM patients revealed significant changes in 78 proteins, primarily involving complement and coagulation cascades, platelet activation, prothrombotic factors, and cholesterol metabolism. Complement C3, C5, C4B, C4BPB, and C4BPA were downregulated, while C7, C9, and F12 were upregulated. These changes are consistent with the chronic low-grade inflammation and vascular dysfunction characteristic of GDM.103 Proteomic analysis of syncytiotrophoblast-derived EVs showed that 56 proteins were differentially expressed in medium/large EVs, with positive regulation of the cytoskeleton being the most significantly enriched biological process, suggesting that EV protein profiles may serve as potential biomarkers for GDM diagnosis.91
Nevertheless, there is significant inconsistency in the proteomic findings across different studies. For example, Kandzija et al91 used an ex vivo placental perfusion model and found no differentially expressed proteins in small syncytiotrophoblast-derived EVs but identified differences in medium/large sEVs. In contrast, other studies using plasma EVs reported a large number of differentially expressed proteins.103 These discrepancies may arise from differences in sample sources, isolation methods, and mass spectrometry analysis strategies. These contradictory findings indicate that reliable and reproducible protein biomarkers are currently lacking, and there is an urgent need for multicenter standardized studies.
Diagnostic Value of EVs from Different Sources
EVs derived from different tissues carry tissue-specific molecular cargoes, providing multidimensional information for GDM diagnosis (Table 2). Placenta-derived EVs serve as the most direct readout of placental function. Their presence in the maternal circulation as early as six weeks of gestation provides an opportunity for early assessment.26 Quantitative analysis has revealed a progressive increase in placenta-derived EV concentration across gestation, with GDM patients showing consistently higher levels than healthy controls in each trimester.70 Proteomic profiling of these EVs during early and mid-gestation has identified differentially expressed proteins enriched in complement cascades, immune regulation, inflammation, and coagulation pathways.102 Additionally, placenta-derived EVs carry C19MC family miRNAs, including miR-516-5p, miR-517-3p, and miR-518-5p, which exhibit altered expression patterns in GDM and are known to be involved in maternal-fetal immune tolerance and placental development.95
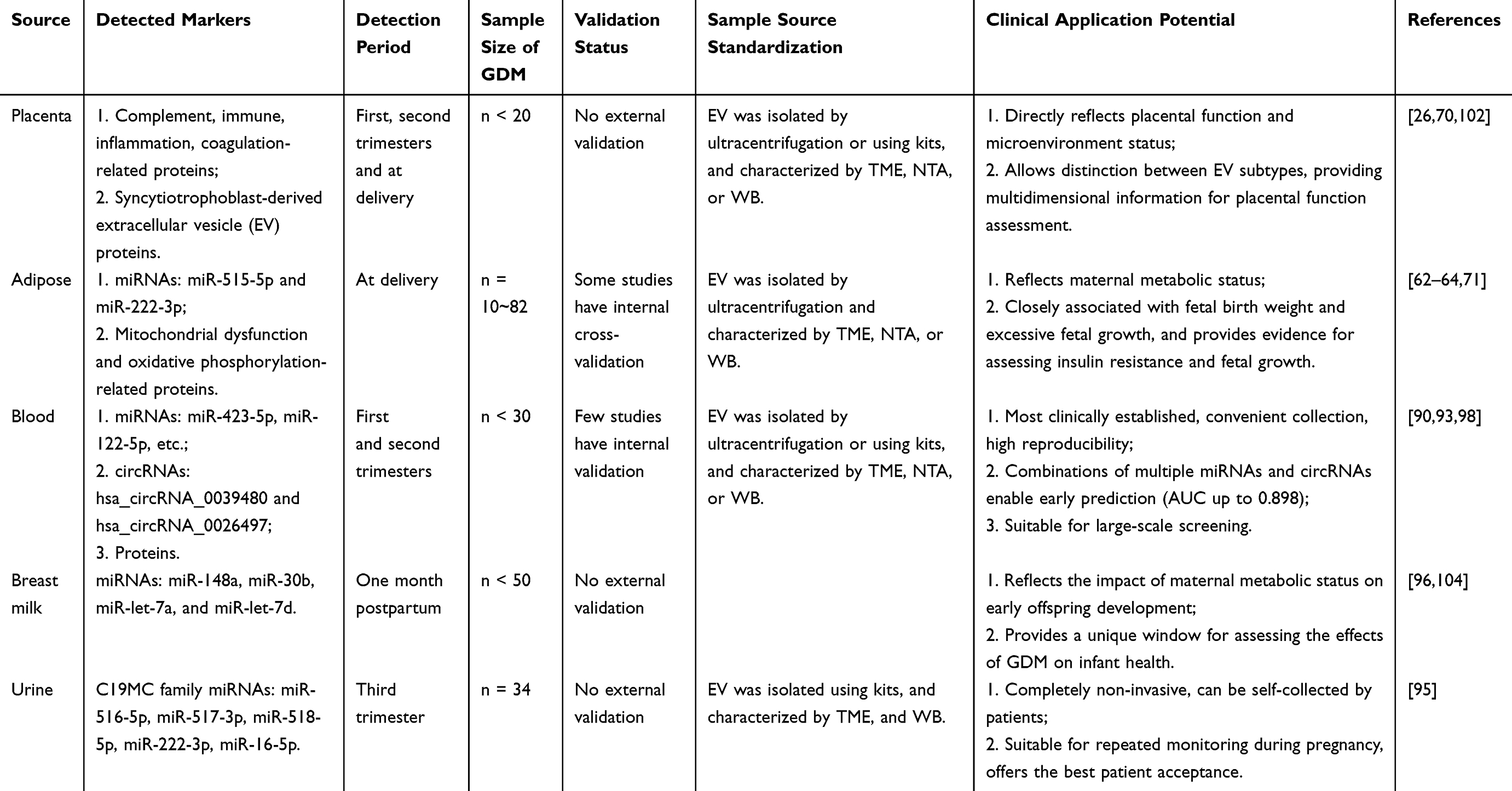 |
Table 2 Diagnostic Value of Extracellular Vesicles from Different Sources in Gestational Diabetes Mellitus |
Adipose tissue-derived EVs capture information about maternal metabolic status. Their abundance is elevated 1.7-fold in GDM pregnancies compared to normal controls, and this increase correlates positively with birth weight Z-scores.71 Proteomic analysis has revealed that the upregulated proteins in these exosomes are predominantly associated with mitochondrial dysfunction and oxidative phosphorylation pathways.64 Specific miRNA cargoes within adipose tissue-derived EVs, such as miR-515-5p, promote placental glucose uptake and correlate with excessive fetal growth,62,63 while miR-222-3p has been linked to estrogen-mediated insulin resistance.105
Blood-derived exosomes represent the most extensively characterized diagnostic samples in clinical practice. Multiple molecular panels combining miRNAs, circRNAs, and proteins have demonstrated diagnostic value.90,93,98 While blood collection is convenient and allows repeated sampling, the heterogeneous nature of circulating EVs requires careful subpopulation analysis using tissue-specific markers to enhance diagnostic precision.26 Breast milk-derived EVs offer a unique perspective on the impact of maternal metabolic status on offspring development. Reduced abundance of specific miRNAs, including miR-148a, miR-30b, let-7a, and let-7d, has been observed in milk EVs from GDM mothers, with miR-148a levels correlating with infant body composition.96,104 Additionally, urine-derived EVs represent a completely non-invasive diagnostic approach. They contain placenta-derived EV populations, as evidenced by the detection of C19MC family miRNAs such as miR-516-5p and miR-517-3p in urine samples from pregnant women.95 Urine collection is simple and patient-friendly, making it suitable for repeated monitoring throughout pregnancy, although standardized collection and preservation protocols remain to be established.
A comparison of diagnostic value across these sources reveals distinct strengths, placenta-derived EVs offer the highest specificity for placental function assessment, blood-derived EVs are the most clinically established, urine-derived EVs provide optimal patient compliance, and breast milk-derived EVs uniquely inform offspring health outcomes. Integrating molecular signatures from multiple EV sources may enable a multidimensional diagnostic approach for GDM.
Although the studies described above demonstrate the great potential of EVs as diagnostic biomarkers for GDM, multiple obstacles must be overcome for their clinical translation. First, most studies have small sample sizes and lack independent external validation cohorts, which may lead to overestimation of the reported AUC values. Second, EV isolation and characterization methods are not uniform, techniques such as ultracentrifugation, size exclusion chromatography, and polymer precipitation differ significantly in EV yield, purity, and integrity, directly affecting the reliability of downstream analyses. Third, differences in detection platforms and data analysis workflows across studies make cross-validation of candidate biomarkers difficult. Finally, reference ranges and diagnostic thresholds based on EV biomarkers have not yet been established, preventing direct application in clinical decision-making. Therefore, future studies should adopt multicenter, prospective designs, follow standardized operating procedures recommended by the MISEV 2023 guidelines, and perform rigorous independent validation.
Advantages of Therapeutic EVs
As novel drug delivery systems, therapeutic EVs offer several unique advantages. First, EVs have low immunogenicity, and their lipid bilayer membrane structure prevents rapid clearance by the immune system, thereby prolonging circulation time in vivo.106 Unlike live cell-based formulations, EVs carry no risk of replication and therefore exhibit a superior safety profile.22 EVs can also be derived from autologous cells, thereby avoiding the immune rejection often associated with allogeneic transplantation.107 EVs are easy to store and transport, and lyophilized EVs can be stored at room temperature for extended periods, facilitating clinical translation.22 Moreover, EVs can be engineered for targeted delivery through surface modification with targeting ligands or by loading therapeutic nucleic acids or drugs.108 Finally, EVs offer flexible administration routes. They can be delivered intravenously for systemic distribution or administered orally, intranasally, or locally for targeted delivery. Notably, plant-derived EVs and bEVs exhibit good oral bioavailability.109
Therapeutic Strategies Using EVs from Different Sources
Stem Cell-Derived Exosomes
Stem cell-derived exosomes retain the therapeutic activity of stem cells while avoiding the risks associated with cell transplantation, making them important candidate strategies for GDM therapy. Human umbilical cord mesenchymal stromal cell-derived exosomes (hucMSC-Exos) have shown promising therapeutic effects in GDM. Intravenous injection of hucMSC-Exos in GDM mice significantly improved glucose metabolism abnormalities and insulin resistance. The mechanism involves circ_0046060 carried by these exosomes upregulating G6PC2 expression by sponging miR-338-3p, thereby regulating glucose metabolism.110 In vitro experiments showed that hucMSC-Exos carrying miR-455-3p target calcium/calmodulin-dependent protein kinase 2 inhibitor 1 (CAMK2N1), protecting high glucose-induced trophoblast cell injury, promoting cell proliferation and migration, and inhibiting apoptosis.111
Applications of stem cell-derived exosomes in other pregnancy complications provide supporting evidence for GDM therapy. Human umbilical cord mesenchymal stem cell-derived exosomes alleviate antiphospholipid antibody-induced trophoblast injury and placental dysfunction by delivering miR-146a-5p to inhibit the TRAF6/ nuclear factor κB (NF-κB) axis.112 Amniotic fluid-derived mesenchymal stem cell EVs containing miR-146a-5p and miR-548e-5p reduce lipopolysaccharide-induced inflammatory responses in trophoblast cells by regulating NF-κB, AKT, and mitogen-activated protein kinase signaling pathways.113 These studies indicate that stem cell-derived exosomes exert protective effects in pregnancy-related disorders through mechanisms including anti-inflammation, anti-apoptosis, and the promotion of cellular functions. However, all of the above therapeutic evidence is derived from GDM animal models or in vitro cell experiments, and no clinical studies on stem cell-derived exosomes for GDM therapy have been reported to date.
Plant-Derived EVs
Plant-derived EVs have emerged as a novel therapeutic strategy in recent years, offering advantages such as high oral bioavailability, a favorable safety profile, and potential for large-scale production.109 It must be clearly emphasized that no direct studies on plant-derived EVs for GDM therapy have been published to date, all evidence for plant-derived EVs presented in this section is derived from type 2 diabetes. Although GDM shares some pathophysiological similarities with type 2 diabetes, including insulin resistance and β-cell dysfunction,114,115 the pregnancy-specific placental factors and hormonal environment cannot be overlooked. Therefore, the following evidence serves only as mechanistic insights and future hypotheses, and does not represent proven efficacy for GDM.
In type 2 diabetes animal models, a biomimetic hydrogen nanogenerator coated with ginger-derived EVs significantly improved insulin resistance and hepatic steatosis in mice, inhibited pancreatic β-cell dedifferentiation, and increased the proportion of pancreatic β-cells. Mechanistic studies revealed that this nanoplatform increased the abundance of Lactobacillus and levels of tryptophan metabolites, activating the aryl hydrocarbon receptor/interleukin-22 pathway and improving intestinal barrier function.116 Garlic-derived exosome-like nanoparticles can “train” intestinal A. muciniphila to release functional outer membrane vesicles, reversing high-fat diet-induced type 2 diabetes. The mechanism involves increased levels of Amuc-1100 and P9 proteins in outer membrane vesicles, promoting glucagon-like peptide-1 secretion, while also increasing phosphatidylcholine levels and inhibiting cGas/STING-mediated inflammatory responses.117
In GDM, Lycium barbarum polysaccharide has been shown to improve symptoms in GDM mice by modulating miRNA expression profiles in plasma EVs.118 Although this study did not directly use plant-derived EVs, it provides mechanistic evidence that plant active ingredients may exert effects through exosomal pathways. Overall, the application of plant-derived EVs in GDM therapy is still at the hypothesis stage. Future studies must first directly evaluate the efficacy and safety of plant-derived EVs in animal models of GDM, and clarify whether they exert their effects through the same gut-placenta axis mechanism. In addition, standardized preparation, quality control, and pharmacokinetic studies of plant-derived EVs are urgently needed.
Conclusion
Nowadays, substantial progress has been made in elucidating the role of EVs in GDM. They have been shown to participate in β-cell dysfunction, placental dysfunction, hepatic insulin resistance, and multi-organ crosstalk involving adipose tissue, the placenta, and the liver. These findings have deepened the understanding of GDM pathophysiology.119,120 Currently, multiple studies have identified differentially expressed miRNAs, circRNAs, and protein cargoes in circulating EVs from GDM patients, underscoring their potential for early diagnosis.26 In addition, stem cell-derived exosomes have shown efficacy in improving glucose metabolism and insulin resistance in animal models, while plant-derived EVs have garnered significant attention owing to their high oral bioavailability and favorable safety profile.106 However, despite these advances in basic research, the clinical translation of EVs faces numerous challenges.
The clinical translation of EVs must overcome challenges in quality control and standardization. Methods for EV isolation, purification, and characterization remain unstandardized. Different laboratories use various techniques such as ultracentrifugation, size exclusion chromatography, polymer precipitation, or immunoaffinity capture. These methods differ significantly in EV yield, purity, size distribution, and contaminant residues.22,121 The MISEV 2023 guidelines published by the International Society for Extracellular Vesicles provide foundational standards for EV research. These guidelines emphasize the importance of selecting appropriate isolation methods based on the research purpose and performing thorough characterization of EV size, concentration, and marker expression.122 However, the implementation of these guidelines varies across studies, limiting the comparability and reproducibility of research results. Furthermore, EV source, extraction method, and storage conditions all affect batch consistency. The establishment of standardized operating procedures is a prerequisite for clinical translation.22
Safety assessment poses a critical barrier to the clinical translation of EVs. This is particularly important for pregnancy-related applications, given more stringent safety requirements. GDM treatment involves both the mother and the fetus, and any therapeutic intervention must consider potential effects on both individuals. Although stem cell-derived exosomes avoid the risks associated with live cell transplantation, their long-term safety still necessitates systematic evaluation.119 Plant-derived EVs have garnered interest because of their natural origin and good biocompatibility. Existing studies have shown no overt toxicity of plant EVs in animal models, but systematic toxicological evaluation, including acute toxicity, subchronic toxicity, and genotoxicity studies, is still needed.123 Safety profiles may vary among EVs from different sources, warranting further evaluation.
In addition, current EV production relies primarily on cell culture, which yields low quantities at high cost, thereby limiting the ability to meet clinical demands.124 Plant-derived EVs hold promise for large-scale production because of their natural abundance, but optimizing of extraction processes and improving purification efficiency remain urgent challenges.109,123 Good manufacturing practice (GMP)-compliant production processes for EVs have not yet been fully established. This includes standardization of upstream cell or plant material culture, scale-up of downstream purification processes, establishment of quality control systems, and development of final formulation methods.108,124 Moreover, the stability and storage conditions of EV formulations influence their clinical applicability. Lyophilization can improve EV storage stability, but the effects of the lyophilization process on EV structure and function require systematic investigation.125
Future Perspectives
Based on current research progress and technical bottlenecks, the following strategies, prioritized according to their clinical translational feasibility, are expected to drive future development.
(1) Advance the development of oral formulations for plant-derived EVs. Plant-derived EVs represent the most promising strategy for GDM therapy owing to their natural origin, high safety profile, suitability for oral administration, and potential for large-scale extraction, rendering them highly feasible for clinical translation. Research should focus on the stability of plant-derived EVs in the gastrointestinal tract, elucidating the mechanisms underlying they resistance to degradation by gastric acid and digestive enzymes. Meanwhile, oral delivery systems such as enteric coatings or hydrogels should be developed for plant-derived EVs to improve their intestinal targeting and bioavailability.
(2) Optimize the targeting efficiency of engineered exosomes. Although engineered stem cell-derived exosomes entail higher costs, their strong targeting capability and clear efficacy make them uniquely valuable for precision therapy. Placenta-specific targeting ligands, such as antibodies or peptides against surface markers on syncytiotrophoblasts, should be further screened and optimized, and efficient surface modification methods should also be established to enable targeted delivery. However, given the complex engineering steps and high production costs, clinical translation of this strategy may proceed more slowly.
(3) Explore the application of bEVs as postbiotic formulations. bEVs represent an emerging research direction. As postbiotic formulations, they offer advantages including suitability for oral administration and the ability to target the gut microbiota. Although no direct studies on probiotic EVs for GDM therapy have been conducted, the discovery of microbiota components in serum EVs from GDM patients provides a theoretical foundation for this approach. Methods for isolating and purifying bEVs should first be established, and the molecular composition and functional characteristics of bEVs from different probiotic strains should be characterized. Oral delivery strategies suitable for bEVs should be developed to protect their active components from degradation in the digestive tract. Given that this field remains in the proof‑of‑concept stage and lacks direct evidence for GDM, its clinical translation represents a long‑term exploratory direction.
Although clinical translation still faces obstacles related to quality control, safety assessment, and large-scale production, the establishment of standardized production systems and comprehensive safety evaluation is expected to enable EVs to become new tools for the precision diagnosis and treatment of GDM.
Abbreviations
GDM, gestational diabetes mellitus; PE, preeclampsia; OGTT, oral glucose tolerance test; DIPSI, Diabetes in Pregnancy Study Group India; WHO, World Health Organization; EVs, extracellular vesicles; miRNA, microRNA; sEVs, small extracellular vesicles; circRNA, circular RNA; ESCRT, endosomal sorting complexes required for transport; ILVs, intraluminal vesicles; MVB, multivesicular body; mTOR, mechanistic target of rapamycin; PD-L1, programmed death ligand 1; PD-1, programmed death receptor 1; GLP-1, glucagon-like peptide-1; PIK3R2, phosphoinositide-3-kinase regulatory subunit 2; PI3K/AKT, phosphoinositide-3-kinase/protein kinase B; SIRT1, sirtuin 1 NOX4, NADPH oxidase 4; LRG1, leucine-rich α-2-glycoprotein 1; ECM1, extracellular matrix protein 1; STAT3, signal transducer and activator of transcription 3; FAM3A, family with sequence similarity 3 member A; DPPIV, dipeptidyl peptidase IV; bEVs, bacterial extracellular vesicles; AUC, area under the curve; lncRNAs, long non-coding RNAs; hucMSC-Exos, human umbilical cord mesenchymal stromal cell-derived exosomes; CAMK2N1, calmodulin-dependent protein kinase 2 inhibitor 1; NF-κB, nuclear factor κB; plant-derived EVs, Plant-derived EVs; GMP, good manufacturing practice.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Zhejiang Provincial Natural Science Foundation of China under Grant No. LQN25H160047. Medical and Health Science Program of Zhejiang Province under Grant No. 2024KY1694.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. American Diabetes Association Professional Practice Committee for D. 2. Diagnosis and classification of diabetes: standards of care in diabetes-2026. Diabetes Care. 2026;49(Supplement_1):S27–19. doi:10.2337/dc26-S002
2. Sweeting A, Hannah W, Backman H, et al. Epidemiology and management of gestational diabetes. Lancet. 2024;404(10448):175–192. doi:10.1016/S0140-6736(24)00825-0
3. Punnose J, Gobl CS, Glastras SJ, Tura A. Editorial: recent advances in gestational diabetes: diagnosis, treatment and prevention. Front Endocrinol. 2026;17:1773541. doi:10.3389/fendo.2026.1773541
4. Yu J, Chen Z, Chen Y, et al. Neglected sleep quality problems in women with gestational diabetes mellitus: a multi-center cross-sectional study. BMC Pregnancy Childbirth. 2024;24(1):794. doi:10.1186/s12884-024-06940-y
5. Senbanjo OC, Akinlusi FM, Rabiu KA. Prevalence and determinants of gestational diabetes mellitus at a tertiary health facility in Lagos, Nigeria. Niger Postgrad Med J. 2026;33(1):103–110. doi:10.4103/npmj.npmj_117_25
6. Hannah W, Deepa M, Ram U, et al. Prevalence and risk factors of early gestational diabetes mellitus (EGDM) in Indians: the STRiDE study. Diabet Res Clin Pract. 2026;231:113023. doi:10.1016/j.diabres.2025.113023
7. Juan J, Sun Y, Wei Y, et al. Progression to type 2 diabetes mellitus after gestational diabetes mellitus diagnosed by IADPSG criteria: systematic review and meta-analysis. Front Endocrinol. 2022;13:1012244. doi:10.3389/fendo.2022.1012244
8. Retnakaran R, Ye C, Hanley AJ, Connelly PW, Sermer M, Zinman B. Subtypes of gestational diabetes and future risk of pre-diabetes or diabetes. EClinicalMedicine. 2021;40:101087. doi:10.1016/j.eclinm.2021.101087
9. Erkamp JS, Geurtsen ML, Duijts L, et al. Associations of maternal early-pregnancy glucose concentrations with placental hemodynamics, blood pressure, and gestational hypertensive disorders. Am J Hypertens. 2020;33(7):660–669. doi:10.1093/ajh/hpaa070
10. Simmons D, Immanuel J, Hague WM, et al. Treatment of gestational diabetes mellitus diagnosed early in pregnancy. N Engl J Med. 2023;388(23):2132–2144. doi:10.1056/NEJMoa2214956
11. Carlson AL, Beck RW, Li Z, et al. Continuous glucose monitoring-measured glucose levels during oral glucose tolerance testing in pregnancy. Diabetes Technol Ther. 2025;27(6):439–448. doi:10.1089/dia.2024.0563
12. Punnose J, Sukhija K, Rijhwani RM. Intermediate hyperglycemia in early pregnancy: a South Asian perspective. World J Diab. 2023;14(5):573–584. doi:10.4239/wjd.v14.i5.573
13. Saxena P, Verma P, Goswami B. Comparison of diagnostic accuracy of non-fasting DIPSI and HbA1c with fasting WHO criteria for diagnosis of gestational diabetes mellitus. J Obstet Gynaecol India. 2017;67(5):337–342. doi:10.1007/s13224-017-0962-y
14. Bergenstal RM. Continuous glucose monitoring: transforming diabetes management step by step. Lancet. 2018;391(10128):1334–1336. doi:10.1016/S0140-6736(18)30290-3
15. Zale A, Mathioudakis N. Machine learning models for inpatient glucose prediction. Curr Diab Rep. 2022;22(8):353–364. doi:10.1007/s11892-022-01477-w
16. Caixeta DC, Lima C, Xu Y, et al. Monitoring glucose levels in urine using FTIR spectroscopy combined with univariate and multivariate statistical methods. Spectrochim Acta A Mol Biomol Spectrosc. 2023;290:122259. doi:10.1016/j.saa.2022.122259
17. Toledano Y, Hadar E, Hod M. Pharmacotherapy for hyperglycemia in pregnancy - The new insulins. Diabet Res Clin Pract. 2018;145:59–66. doi:10.1016/j.diabres.2018.04.035
18. Costa DN, Malek R, Davis SN. A safety review of approved oral agents for treating type 2 diabetes and associated diseases in pregnant patients. Expert Opin Drug Saf. 2026;25(2):259–276. doi:10.1080/14740338.2025.2545929
19. Yu DQ, Xu GX, Teng XY, et al. Glycemic control and neonatal outcomes in women with gestational diabetes mellitus treated using glyburide, metformin, or insulin: a pairwise and network meta-analysis. BMC Endocr Disord. 2021;21(1):199. doi:10.1186/s12902-021-00865-9
20. Xie Y, Dang W, Zhang S, et al. The role of exosomal noncoding RNAs in cancer. Mol Cancer. 2019;18(1):37. doi:10.1186/s12943-019-0984-4
21. Liu Q, Li D, Pan X, Liang Y. Targeted therapy using engineered extracellular vesicles: principles and strategies for membrane modification. J Nanobiotechnol. 2023;21(1):334. doi:10.1186/s12951-023-02081-0
22. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
23. Iqbal Z, Rehman K, Mahmood A, et al. Exosome for mRNA delivery: strategies and therapeutic applications. J Nanobiotechnol. 2024;22(1):395. doi:10.1186/s12951-024-02634-x
24. Zhang M, Xing J, Zhao S, Chen H, Yin X, Zhu X. Engineered extracellular vesicles in female reproductive disorders. Biomed Pharmacother. 2023;166:115284. doi:10.1016/j.biopha.2023.115284
25. Su X, Wang H, Li Q, Chen Z. Extracellular vesicles: a review of their therapeutic potentials, sources, biodistribution, and administration routes. Int J Nanomed. 2025;20:3175–3199. doi:10.2147/IJN.S502591
26. Ahmed M, Temilola DO, Matjila M, Chetty M. Small extracellular vesicles in gestational diabetes mellitus: current landscape and emerging diagnostic horizons. J Extracell Biol. 2026;5(3):e70120. doi:10.1002/jex2.70120
27. James-Allan LB, Devaskar SU. Extracellular vesicles and their role in gestational diabetes mellitus. Placenta. 2021;113:15–22. doi:10.1016/j.placenta.2021.02.012
28. Farrelly R, Kennedy MG, Spencer R, Forbes K. Extracellular vesicles as markers and mediators of pregnancy complications: gestational diabetes, pre-eclampsia, preterm birth and fetal growth restriction. J Physiol. 2023;601(22):4973–4988. doi:10.1113/JP282849
29. Lotvall J, Hill AF, Hochberg F, et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International Society for Extracellular Vesicles. J Extracell Vesicles. 2014;3:26913. doi:10.3402/jev.v3.26913
30. Thery C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
31. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
32. Mathieu M, Martin-Jaular L, Lavieu G, Thery C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17. doi:10.1038/s41556-018-0250-9
33. Hessvik NP, Llorente A. Current knowledge on exosome biogenesis and release. Cell Mol Life Sci. 2018;75(2):193–208. doi:10.1007/s00018-017-2595-9
34. Hurley JH. ESCRTs are everywhere. EMBO J. 2015;34(19):2398–2407. doi:10.15252/embj.201592484
35. Henne WM, Buchkovich NJ, Emr SD. The ESCRT pathway. Dev Cell. 2011;21(1):77–91. doi:10.1016/j.devcel.2011.05.015
36. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–1247. doi:10.1126/science.1153124
37. Mathieu M, Nevo N, Jouve M, et al. Specificities of exosome versus small ectosome secretion revealed by live intracellular tracking of CD63 and CD9. Nat Commun. 2021;12(1):4389. doi:10.1038/s41467-021-24384-2
38. Mitchell MD, Peiris HN, Kobayashi M, et al. Placental exosomes in normal and complicated pregnancy. Am J Obstet Gynecol. 2015;213(4 Suppl):S173–181. doi:10.1016/j.ajog.2015.07.001
39. Nishi K, Modi D. Placental exosomes in pregnancy and preeclampsia. Am J Reprod Immunol. 2024;91(5):e13857. doi:10.1111/aji.13857
40. Czernek L, Düchler M. Exosomes as messengers between mother and fetus in pregnancy. Int J Mol Sci. 2020;21:4264. doi:10.3390/ijms21124264
41. Bai K, Li X, Zhong J, et al. Placenta-derived exosomes as a modulator in maternal immune tolerance during pregnancy. Front Immunol. 2021;12:671093. doi:10.3389/fimmu.2021.671093
42. Tong M, Abrahams VM, Chamley LW. Immunological effects of placental extracellular vesicles. Immunol Cell Biol. 2018;96:714–722. doi:10.1111/imcb.12049
43. Schuster J, Cheng SB, Padbury J, Sharma S. Placental extracellular vesicles and pre-eclampsia. Am J Reprod Immunol. 2021;85(2):e13297. doi:10.1111/aji.13297
44. Powell JS, Larregina AT, Shufesky WJ, et al. Fetoplacental extracellular vesicles deliver conceptus-derived antigens to maternal secondary lymphoid tissues for immune recognition. JCI Insight. 2025;10(10). doi:10.1172/jci.insight.186335.
45. Ohara T, Horie A, Nakamura M, et al. Galectin-3 in endometrial small extracellular vesicles promotes cytotrophoblast cell fusion during the implantation phase. Cell Commun Signal. 2025;24(1):41. doi:10.1186/s12964-025-02586-1
46. Gebara N, Correia Y, Wang K, Bussolati B. Angiogenic properties of placenta-derived extracellular vesicles in normal pregnancy and in preeclampsia. Int J Mol Sci. 2021;22(10):5402. doi:10.3390/ijms22105402
47. Mincheva-Nilsson L. Immunosuppressive protein signatures carried by syncytiotrophoblast-derived exosomes and their role in human pregnancy. Front Immunol. 2021;12:717884. doi:10.3389/fimmu.2021.717884
48. Sabapatha A, Gercel-Taylor C, Taylor DD. Specific isolation of placenta-derived exosomes from the circulation of pregnant women and their immunoregulatory consequences. Am J Reprod Immunol. 2006;56(5–6):345–355. doi:10.1111/j.1600-0897.2006.00435.x
49. Kshirsagar SK, Alam SM, Jasti S, et al. Immunomodulatory molecules are released from the first trimester and term placenta via exosomes. Placenta. 2012;33(12):982–990. doi:10.1016/j.placenta.2012.10.005
50. Chaiwangyen W, Murrieta-Coxca JM, Favaro RR, et al. MiR-519d-3p in trophoblastic cells: effects, targets and transfer to allogeneic immune cells via extracellular vesicles. Int J Mol Sci. 2020;21(10):3458. doi:10.3390/ijms21103458
51. James-Allan LB, Rosario FJ, Barner K, et al. Regulation of glucose homeostasis by small extracellular vesicles in normal pregnancy and in gestational diabetes. FASEB J. 2020;34(4):5724–5739. doi:10.1096/fj.201902522RR
52. James-Allan LB, Rosario FJ, Madi L, et al. A novel technique using chronic infusion of small extracellular vesicles from gestational diabetes mellitus causes glucose intolerance in pregnant mice. Clin Sci. 2022;136(21):1535–1549. doi:10.1042/CS20220484
53. Wang Y, Yuan Y, Shen S, Ge Z, Zhu D, Bi Y. Placenta-derived exosomes exacerbate beta cell dysfunction in gestational diabetes mellitus through delivery of miR-320b. Front Endocrinol. 2023;14:1282075. doi:10.3389/fendo.2023.1282075
54. Razo-Azamar M, Nambo-Venegas R, Quevedo IR, et al. Early-pregnancy serum maternal and placenta-derived exosomes miRNAs vary based on pancreatic beta-cell function in GDM. J Clin Endocrinol Metab. 2024;109(6):1526–1539. doi:10.1210/clinem/dgad751
55. Xu C, Zou L, Wang L, et al. Gestational diabetes mellitus-derived miR-7-19488 targets PIK3R2 mRNA to stimulate the abnormal development and maturation of offspring-islets. Life Sci. 2025;363:123369. doi:10.1016/j.lfs.2025.123369
56. Xu P, Dong S, Wu L, et al. Maternal and placental DNA methylation changes associated with the pathogenesis of gestational diabetes mellitus. Nutrients. 2022;15(1):70. doi:10.3390/nu15010070
57. Luo Y, Qin Y, Kong L, et al. Clinical and pathological characteristics of gestational diabetes mellitus with different insulin resistance. J Diabetes Complications. 2024;38(8):108796. doi:10.1016/j.jdiacomp.2024.108796
58. Zhang Q, Ye X, Xu X, Yan J. Placenta-derived exosomal miR-135a-5p promotes gestational diabetes mellitus pathogenesis by activating PI3K/AKT signalling pathway via SIRT1. J Cell Mol Med. 2023;27(23):3729–3743. doi:10.1111/jcmm.17941
59. Zhang L, Wu Q, Zhu S, et al. Chemerin-induced down-regulation of placenta-derived exosomal miR-140-3p and miR-574-3p promotes umbilical vein endothelial cells proliferation, migration, and tube formation in gestational diabetes mellitus. Cells. 2022;11(21):3457. doi:10.3390/cells11213457
60. Zhang L, Zeng M, Tang F, Chen J, Cao D, Tang ZN. Circ-PNPT1 contributes to gestational diabetes mellitus (GDM) by regulating the function of trophoblast cells through miR-889-3p/PAK1 axis. Diabetol Metab Syndr. 2021;13(1):58. doi:10.1186/s13098-021-00678-9
61. Tao Y, Chen W, Xu H, et al. Adipocyte-derived exosomal NOX4-mediated oxidative damage induces premature placental senescence in obese pregnancy. Int J Nanomed. 2023;18:4705–4726. doi:10.2147/IJN.S419081
62. Jayabalan N, Nair S, Lai A, et al. Extracellular vesicle-associated miR-515-5p from adipose tissue regulates placental metabolism and fetal growth in gestational diabetes mellitus. Cardiovasc Diabetol. 2025;24(1):205. doi:10.1186/s12933-025-02739-z
63. Jayabalan N, Scholz-Romero K, Guanzon D, Lai A, Salomon C. 1972-P: adipose tissue–derived exosomal microRNA regulates placental glucose uptake in gestational diabetes mellitus pregnancies. Diabetes. 2020;69(Supplement 1):1972–P. doi:10.2337/db20-1972-P
64. Jayabalan N, Ormazabal V, Lai A, Lappas M, Salomon C. Proteomic profile of adipose tissue-derived exosomes and their potential role on placental glucose metabolism in gestational diabetes mellitus. Diabetes. 2018;67(Sup1):1. doi:10.2337/db18-165-LB
65. Yao J, Duan T, Wang K. 1388-P: umbilical cord blood-derived exosomal LRG1 and ECM1 induce abnormal placental angiogenesis in women with gestational diabetes mellitus. Diabetes. 2019;68(Sup1):1. doi:10.2337/db19-1388-P
66. Yao J, Chang X, He Q, Li H, Duan T, Wang K. Exosome enriched leucine-rich alpha-2-glycoprotein-1 and extracellular matrix protein 1 proteins induce abnormal placental angiogenesis in pregnant mice. Placenta. 2023;143:45–53. doi:10.1016/j.placenta.2023.09.008
67. Petersen MC, Shulman GI. Mechanisms of insulin action and insulin resistance. Physiol Rev. 2018;98(4):2133–2223. doi:10.1152/physrev.00063.2017
68. Catalano PM. Trying to understand gestational diabetes. Diabet Med. 2014;31(3):273–281. doi:10.1111/dme.12381
69. Chen X, Tao X, Wang M, et al. Circulating extracellular vesicle-derived miR-1299 disrupts hepatic glucose homeostasis by targeting the STAT3/FAM3A axis in gestational diabetes mellitus. J Nanobiotechnol. 2024;22(1):509. doi:10.1186/s12951-024-02766-0
70. Salomon C, Scholz-Romero K, Sarker S, et al. Gestational diabetes mellitus is associated with changes in the concentration and bioactivity of placenta-derived exosomes in maternal circulation across gestation. Diabetes. 2016;65(3):598–609. doi:10.2337/db15-0966
71. Jayabalan N, Lai A, Ormazabal V, et al. Adipose tissue exosomal proteomic profile reveals a role on placenta glucose metabolism in gestational diabetes mellitus. J Clin Endocrinol Metab. 2019;104(5):1735–1752. doi:10.1210/jc.2018-01599
72. Kandzija N, Zhang W, Motta-Mejia C, et al. Placental extracellular vesicles express active dipeptidyl peptidase IV; levels are increased in gestational diabetes mellitus. J Extracell Vesicles. 2019;8(1):1617000. doi:10.1080/20013078.2019.1617000
73. Zhang M, He S, Han C, et al. High-throughput screening of DPPIV inhibitors antagonizing GLP-1 degradation using an enzymatic activated fluorescent probe. Anal Chem. 2025;97(1):262–270. doi:10.1021/acs.analchem.4c04093
74. Huang J, Yu Y, Feng Z, et al. Cross - kingdom dialogue of microbial messengers: multi - target regulatory mechanisms and therapeutic strategies of gut microbiota - derived extracellular vesicles in metabolic diseases. Int J Nanomed. 2025;20:12573–12591. doi:10.2147/IJN.S548624
75. Zhou Y, Sun Y, Yin P, Zuo S, Li H, Cao K. Bacterial extracellular vesicles: emerging mediators of gut-liver axis crosstalk in hepatic diseases. Front Cell Infect Microbiol. 2025;15:1620829. doi:10.3389/fcimb.2025.1620829
76. Turunen J, Kyro K, Virta J, et al. Maternal factors associate with microbiota-derived extracellular vesicle profiles in pregnancy: a clinical cohort study. BMC Med. 2026;24(1):352. doi:10.1186/s12916-026-04960-3
77. Kaisanlahti A, Turunen J, Byts N, et al. Maternal microbiota communicates with the fetus through microbiota-derived extracellular vesicles. Microbiome. 2023;11(1):249. doi:10.1186/s40168-023-01694-9
78. Chang CJ, Bai YC, Jiang H, et al. Microbiome analysis of serum extracellular vesicles in gestational diabetes patients. Acta Diabetol. 2025;62(3):329–341. doi:10.1007/s00592-024-02358-2
79. Lara B, Sassot M, Calo G, et al. Extracellular vesicles of Porphyromonas gingivalis disrupt trophoblast cell interaction with vascular and immune cells in an in vitro model of early placentation. Life. 2023;13(10):14 doi:10.3390/life13101971.
80. Lara B, Loureiro I, Gliosca L, et al. Porphyromonas gingivalis outer membrane vesicles shape trophoblast cell metabolism impairing functions associated to adverse pregnancy outcome. J Cell Physiol. 2023;238(11):2679–2691. doi:10.1002/jcp.31138
81. Chelakkot C, Choi Y, Kim DK, et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med. 2018;50(2):e450. doi:10.1038/emm.2017.282
82. He K, An F, Zhang H, et al. Akkermansia muciniphila: a potential target for the prevention of diabetes. Foods. 2024;14(1):23. doi:10.3390/foods14010023
83. Chang Y, Zhang C, Li Z, et al. Extracellular vesicles from Akkermansia muciniphila protect streptozotocin-induced diabetic mice by regulating glucose homeostasis, oxidative stress, and immune tolerance. Front Immunol. 2026;17:1739048. doi:10.3389/fimmu.2026.1739048
84. Chen Y, Ou Z, Pang M, et al. Extracellular vesicles derived from Akkermansia muciniphila promote placentation and mitigate preeclampsia in a mouse model. J Extracell Vesicles. 2023;12(5):e12328. doi:10.1002/jev2.12328
85. Heitzer E, Haque IS, Roberts CES, Speicher MR. Current and future perspectives of liquid biopsies in genomics-driven oncology. Nat Rev Genet. 2019;20(2):71–88. doi:10.1038/s41576-018-0071-5
86. Wang X, Guo J, Yu P, et al. The roles of extracellular vesicles in the development, microenvironment, anticancer drug resistance, and therapy of head and neck squamous cell carcinoma. J Exp Clin Cancer Res. 2021;40(1):35. doi:10.1186/s13046-021-01840-x
87. Skotland T, Sagini K, Sandvig K, Llorente A. An emerging focus on lipids in extracellular vesicles. Adv Drug Deliv Rev. 2020;159:308–321. doi:10.1016/j.addr.2020.03.002
88. Nguyen CM, Sallam M, Islam MS, et al. Placental exosomes as biomarkers for maternal diseases: current advances in isolation, characterization, and detection. ACS Sens. 2023;8(7):2493–2513. doi:10.1021/acssensors.3c00689
89. Liu X, Qiu H. Placenta-derived exosomes and gestational diabetes mellitus. Diab Metabolic Syndrome Obes. 2022;15(000):14. doi:10.2147/DMSO.S363226
90. Guanzon D, Lai A, Scholz-Romero K, Zuñiga F, Salomon C. Circulating exosomal miRNA signature in pregnancies with gestational diabetes mellitus across gestation. Diabetes. 2018;67(Supplement 1):166–LB. doi:10.2337/db18-166-LB
91. Kandzija N, Payne S, Cooke WR, Seedat F, Fischer R, Vatish M. Protein profiling of placental extracellular vesicles in gestational diabetes mellitus. Int J Mol Sci. 2024;25(4):e0267564. doi:10.3390/ijms25041947
92. Bartel DP. Metazoan MicroRNAs. Cell. 2018;173(1):20–51. doi:10.1016/j.cell.2018.03.006
93. Ye Z, Wang S, Huang X, et al. Plasma exosomal miRNAs associated with metabolism as early predictor of gestational diabetes mellitus. Diabetes. 2022;71(11):2272–2283. doi:10.2337/db21-0909
94. Gillet V, Ouellet A, Stepanov Y, et al. miRNA profiles in extracellular vesicles from serum early in pregnancies complicated by gestational diabetes mellitus. J Clin Endocrinol Metab. 2019;104(11):5157–5169. doi:10.1210/jc.2018-02693
95. Herrera-Van Oostdam AS, Toro-Ortiz JC, Lopez JA, et al. Placental exosomes isolated from urine of patients with gestational diabetes exhibit a differential profile expression of microRNAs across gestation. Int J Mol Med. 2020;46(2):546–560. doi:10.3892/ijmm.2020.4626
96. Shah KB, Fields DA, Pezant NP, et al. Gestational diabetes mellitus is associated with altered abundance of exosomal microRNAs in human milk. Clin Ther. 2022;44(2):172–185.e171. doi:10.1016/j.clinthera.2022.01.005
97. Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–691. doi:10.1038/s41576-019-0158-7
98. Jiang B, Zhang J, Sun X, et al. Circulating exosomal hsa_circRNA_0039480 is highly expressed in gestational diabetes mellitus and may be served as a biomarker for early diagnosis of GDM. J Transl Med. 2022;20(1):5. doi:10.1186/s12967-021-03195-5
99. Cao M, Zhang L, Lin Y, et al. Circular RNA expression profiles in umbilical cord blood exosomes from normal and gestational diabetes mellitus patients. Biosci Rep. 2020;40(11). doi:10.1042/BSR20201946.
100. Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016;17(1):47–62. doi:10.1038/nrg.2015.10
101. Cao M, Zhang L, Lin Y, et al. Differential mRNA and long noncoding RNA expression profiles in umbilical cord blood exosomes from gestational diabetes mellitus patients. DNA Cell Biol. 2020;39(11):2005–2016. doi:10.1089/dna.2020.5783
102. Lin J, Zhao D, Liang Y, et al. Proteomic analysis of plasma total exosomes and placenta-derived exosomes in patients with gestational diabetes mellitus in the first and second trimesters. BMC Pregnancy Childbirth. 2024;24(1):713. doi:10.1186/s12884-024-06919-9
103. Bernea EG, Suica VI, Uyy E, et al. Exosome proteomics reveals the deregulation of coagulation, complement and lipid metabolism proteins in gestational diabetes mellitus. Molecules. 2022;27(17):5502. doi:10.3390/molecules27175502
104. Zheng Z, Mo J, Lin F, et al. Milk exosomes from gestational diabetes mellitus (GDM) and healthy parturient exhibit differential miRNAs profiles and distinct regulatory bioactivity on hepatocyte proliferation. Mol Nutr Food Res. 2023;67(16):e2300005. doi:10.1002/mnfr.202300005
105. Zhonghua S, Chun Z, Xirong G, et al. Differential expression of microRNAs in omental adipose tissue from gestational diabetes mellitus subjects reveals miR-222 as a regulator of ERα expression in estrogen-induced insulin resistance. Endocrinology. 2014;155(5):1982–1990. doi:10.1210/en.2013-2046
106. Zheng S, Georgiou HM, Kokkinos MI, Mirabito Colafella KM, Brennecke SP, Kalionis B. Therapeutic potential of extracellular vesicles for treating human pregnancy disorders. Extracell Vesicles Circ Nucl Acids. 2025;6(2):287–309. doi:10.20517/evcna.2025.07
107. S ELA, Mager I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12(5):347–357. doi:10.1038/nrd3978
108. Lener T, Gimona M, Aigner L, et al. Applying extracellular vesicles based therapeutics in clinical trials: an ISEV position paper. J Extracell Vesicles. 2015;4:30087. doi:10.3402/jev.v4.30087
109. Xiao X, Guo Y, Msomi NZ, Islam MS, Chu M. Exosome-like nanoparticles extracted from plant cells for diabetes therapy. Int J Mol Sci. 2025;26(18):9155. doi:10.3390/ijms26189155
110. Cao M, Bu C, Zhang J, et al. Exosomal circular RNA hsa_circ_0046060 of umbilical cord mesenchymal stromal cell ameliorates glucose metabolism and insulin resistance in gestational diabetes mellitus via the miR-338-3p/G6PC2 axis. Int J Endocrinol. 2022;2022:9218113. doi:10.1155/2022/9218113
111. Zhang Y, Du M, Zhu B, Ma R, Zhang J. Human umbilical cord mesenchymal stem cell-derived exosomal miR-455-3p targets CAMK2N1 to improve trophoblast cell injury in gestational diabetes mellitus. Diabetol Metab Syndr. 2025;17(1):138. doi:10.1186/s13098-025-01692-x
112. Lv Q, Wang Y, Tian W, et al. Exosomal miR-146a-5p derived from human umbilical cord mesenchymal stem cells can alleviate antiphospholipid antibody-induced trophoblast injury and placental dysfunction by regulating the TRAF6/NF-κB axis. J Nanobiotechnol. 2023;21:419. doi:10.1186/s12951-023-02179-5
113. Yang C, Lim W, Park J, Park S, You S, Song G. Anti-inflammatory effects of mesenchymal stem cell-derived exosomal microRNA-146a-5p and microRNA-548e-5p on human trophoblast cells. Mol Hum Reprod. 2019;25(11):755–771. doi:10.1093/molehr/gaz054
114. Yao D, GangYi Y, QiNan W. Autophagic dysfunction of beta cell dysfunction in type 2 diabetes, a double-edged sword. Genes Dis. 2021;8(4):438–447. doi:10.1016/j.gendis.2020.03.003
115. Ellerbrock J, Spaanderman B, Drongelen JV, et al. Role of beta cell function and insulin resistance in the development of gestational diabetes mellitus. Nutrients. 2022;14(12):2444. doi:10.3390/nu14122444
116. Wang X, Tian R, Liang C, et al. Biomimetic nanoplatform with microbiome modulation and antioxidant functions ameliorating insulin resistance and pancreatic beta-cell dysfunction for T2DM management. Biomaterials. 2025;313:122804. doi:10.1016/j.biomaterials.2024.122804
117. Sundaram K, Teng Y, Mu J, et al. Outer membrane vesicles released from garlic exosome-like nanoparticles (GaELNs) train gut bacteria that reverses type 2 diabetes via the gut-brain axis. Small. 2024;20(20):e2308680. doi:10.1002/smll.202308680
118. Xiao Y, Chen W, Chen R, et al. Exosomal microRNA expression profiling analysis of the effects of Lycium Barbarum polysaccharide on gestational diabetes mellitus mice. Evid Based Complement Alternat Med. 2020;2020:2953502. doi:10.1155/2020/2953502
119. Nair S, Ormazabal V, Carrion F, Handberg A, McIntyre HD, Salomon C. Extracellular vesicle-mediated targeting strategies for long-term health benefits in gestational diabetes. Clin Sci. 2023;137(16):1311–1332. doi:10.1042/CS20220150
120. Nair S, Razo-Azamar M, Jayabalan N, et al. Advances in extracellular vesicles as mediators of cell-to-cell communication in pregnancy. Cytokine Growth Factor Rev. 2024;76:86–98. doi:10.1016/j.cytogfr.2023.12.004
121. Stawarska A, Bamburowicz-Klimkowska M, Runden-Pran E, et al. Extracellular vesicles as next-generation diagnostics and advanced therapy medicinal products. Int J Mol Sci. 2024;25(12):29. doi:10.3390/ijms25126533
122. Pfaffl MW. How laboratory guidelines promote the validity of circulating extracellular vesicle-associated nucleic acid biomarker signatures in liquid biopsy. Int J Mol Sci. 2025;26(24):12115. doi:10.3390/ijms262412115
123. Li C, Zhao L, Wang X, Wu J, Chen H, Lu Q. Biogenesis, preparation, characterization, therapeutic mechanisms and safety evaluation of plant-derived exosome-like nanovesicles in the treatment of ulcerative colitis. J Nanobiotechnol. 2026 doi:10.1186/s12951-026-04209-4.
124. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nature Nanotechnol. 2021;16(7):748–759. doi:10.1038/s41565-021-00931-2
125. Yuan F, Li YM, Wang Z. Preserving extracellular vesicles for biomedical applications: consideration of storage stability before and after isolation. Drug Delivery. 2021;28(1):1501–1509. doi:10.1080/10717544.2021.1951896
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Salivary Extracellular Vesicles in Detection of Head and Neck Cancers: A Systematic Review
Owecki W, Wojtowicz K, Nijakowski K
International Journal of Nanomedicine 2025, 20:6757-6775
Published Date: 26 May 2025