Back to Journals » OncoTargets and Therapy » Volume 9
Expression of PER, CRY, and TIM genes for the pathological features of colorectal cancer patients
Authors Wang Y, Cheng Y, Yu G, Jia B, Hu Z, Zhang L
Received 23 September 2015
Accepted for publication 22 January 2016
Published 5 April 2016 Volume 2016:9 Pages 1997—2005
DOI https://doi.org/10.2147/OTT.S96925
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Daniele Santini
Yong Wang,1 Yunsheng Cheng,1 Gang Yu,1 Benli Jia,1 Zhihang Hu,1 Lijiu Zhang2
1Department of General Surgery, 2Department of Gastroenterology, The Second Affiliated Hospital of Anhui Medical University, Hefei, People’s Republic of China
Abstract: As typical clock gene machinery, period (PER1, PER2, and PER3), cryptochrome (CRY1 and CRY2), and timeless (TIM), could control proliferation, cellular metabolism, and many key functions, such as recognition and repair of DNA damage, dysfunction of the circadian clock could result in tumorigenesis of colorectal cancer (CRC). In this study, the expression levels of PER1, PER2, and PER3, as well as CRY1, CRY2, and TIM in the tumor tissue and apparently healthy mucosa from CRC patients were examined and compared via quantitative real-time polymerase chain reaction. Compared with the healthy mucosa from CRC patients, expression levels of PER1, PER2, PER3, and CRY2 in their tumor tissue are much lower, while TIM level was much enhanced. There was no significant difference in the CRY1 expression level. High levels of TIM mRNA were much prevalent in the tumor mucosa with proximal lymph nodes. CRC patients with lower expression of PER1 and PER3 in the tumor tissue showed significantly poorer survival rates. The abnormal expression levels of PER and TIM genes in CRC tissue could be related to the genesis process of the tumor, influencing host–tumor interactions.
Keywords: colorectal cancer, cancer chronotherapy, period genes, cryptochrome genes, timeless gene
Introduction
The time-dependent pattern of variation exists in most biological processes and functions of living organisms.1 The circadian rhythm is defined as the rhythmic patterns of oscillation with a period of ~24 hours. The circadian timing system, containing central and peripheral oscillators,2 is responsible for the generation of the rhythmic variation. The central pacemaker and master oscillator, located at the suprachiasmatic nuclei of the brain, are entrained to the environmental light–dark cycle via photon, which is conveyed by the retinohypothalamic tract and synchronizes slave oscillators in peripheral tissues.3,4
Up to now, several biological clock genes have been confirmed, such as ARNTL1, clock (circadian locomotor output cycles kaput), period 1, 2 and 3 (PER1–3), cryptochrome (CRY1 and CRY2), timeless (TIM), timeless-interacting protein (TIPIN), and CSNK1Ε (encoding casein kinase 1-epsilon, CK1ε). The molecular components, which generate circadian rhythms of the circadian/biological clock, constitute a unique collaboration mechanism of genes and proteins via transcriptional, translational, and posttranslational procedures.
PER1–3, CRY1 and CRY2 can be transcriptionally activated by the CLOCK and ARNTL1, which are the basic helix–loop–helix/Period, Aryl-hydrocarbon-receptor, Single minded (PAS) transcription factors, heterodimerizing and binding to the elements of E-box enhancer in the gene promoters. In contrast, CRY and PER proteins form the repression complex, translocating back into the nucleus and interacting with CLOCK and ARNTL1, resulting in the loss of their activity.5,6 As a core circadian clock gene in Drosophila melanogaster, TIM is also retained in mammals. However, the influence of mammalian circadian system on clock function is not yet clear. By interacting with the components of the DNA replication system, TIM could regulate DNA replication processes, which are essential for ataxia telangiectasia and Rad3-related-checkpoint kinase 1 (ATR-Chk1)-mediated as well as ataxia telangiectasia mutated–checkpoint kinase 2 (ATMChk2)-mediated signaling and S-phase arrest.7 As more and more incidence of various cancers has been reported by epidemiological studies, the temporal organization variation in body, defined as the circadian disruption, is nowadays considered to be an important risk factor for cancer.8,9
Approximately 5%–15% genome-wide mRNA expressions, including key cell cycle regulators, tumor suppressors genes, and oncogenes, are circadian rhythmic, which are driven by biological clock genes, whose expression regulates the timing of DNA repair, apoptosis, and cell proliferation. The cycle progression of the cells is tightly regulated by the circadian system, including the control of cell proliferation and apoptosis, as well as clocking the transcription and posttranslational modification for key proteins. The deregulated cell proliferation may occur in case of the circadian clock disruption, implicating in many types of cancers.10,11 Overexpression of PER1 in cancer cell lines of humans leads to the reduction of colony formation and clonogenic expansion and altered expression of transcriptional target genes, such as MYC and p21. For the PER2, a mutation has been investigated to accelerate the generation of intestinal polyp in APCMin/+ mice, and an increase in hyperplasia and neoplasia under γ-radiation has been shown in PER2-null mice.12,13 Based on the analysis of two different mouse models of breast cancer, PER3 was putatively considered to be a tumor suppressor gene. Moreover, for patients with estrogen receptor-positive tumors treated with tamoxifen, and especially for those without chemotherapy, breast cancer recurrence has been observed to be related to the deletion and/or reduction in the expression of PER3 gene.14
In humans, deregulation or polymorphism of the PER, CRY, and TIM genes is associated with a number of neoplastic and hemolymphoprolipherative diseases.15,16 Recently, there is a great deal of interest in the alteration of clock gene and clock-controlled gene expression in colorectal cancer (CRC) patients.17,18 CRC accounts for ~10% among all kinds of cancers, becoming the third most common cancer and the fourth most common for death all over the world.19,20 In this study, the expression levels of PER and CRY genes in human CRC tissues and matched normal mucosa were evaluated to explore the relationship between their expressions in tissue with cancer and the clinical and pathological features of CRC patients.
Patients and methods
Patients
Primary tumor samples and matched normal tissues were obtained from 19 recently diagnosed CRC patients (13 men and six women) in our hospital who had had curative surgery. Clinical data, tumor characteristics, location, and staging of these patients are listed in Table 1. All tissue specimens were collected between 9 am and 5 pm in a same day (9 am to 11 am five samples, 11 am to 1 pm four samples, 1 pm to 3 pm six samples, and 3 pm to 5 pm four samples) and immediately put into liquid nitrogen and stored at −80°C. This study was approved by the Ethics Committee in the Second Affiliated Hospital of Anhui Medical University. All patients offered their informed written consent.
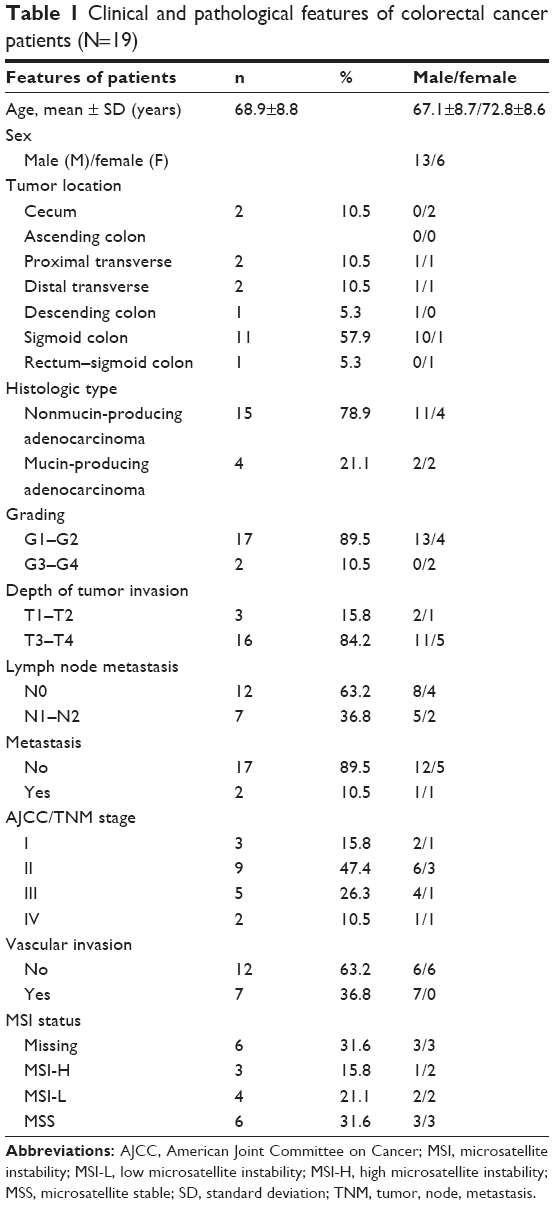 | Table 1 Clinical and pathological features of colorectal cancer patients (N=19) |
Extraction of RNA from fresh frozen tissue and synthesis of the first-strand cDNA
The total RNA from ~150 mg to 200 mg fresh frozen tissues was isolated by phenol extraction. The amount of extracted RNA was weighted by Nano Drop Spectrophotometer, while Agilent 2100 Bioanalyzer was applied for monitoring RNA integrity after subsequent digestion by DNaseI. Then, 1.0 μg of total RNA was reverse transcribed by using the High-Capacity cDNA Archive Kit.
Quantitative real-time reverse transcriptase-polymerase chain reaction assay
The differential expressions of the genes in tumor tissue matched to normal mucosa of CRC samples were assessed by quantitative real-time reverse transcriptase-polymerase chain reaction (qRT-PCR) assay. Human QuantiTec Primers Assay was applied for the determination of PER1, PER2, PER3, CRY1, CRY2, and TIM. All qRT-PCRs with a 25 μL final volume and three replicates were performed on a QuantiFast SYBR Green PCR kit and run in an ABI PRISM 7700 Sequence Detection System under conditions at 50°C for 2 minutes and 95°C for 10 minutes, as well as 40 cycles at 95°C for 15 seconds and 60°C for 1 minute. The threshold cycle (Ct) values were acquired. Expression levels of the target genes were normalized by glutaraldehyde-3-phosphate dehydrogenase housekeeping control gene,21 and the relative amount of mRNA in each target gene comparing with glutaraldehyde-3-phosphate dehydrogenase was calculated by average 2−ΔΔCt method.22 The results are shown in Table 2.
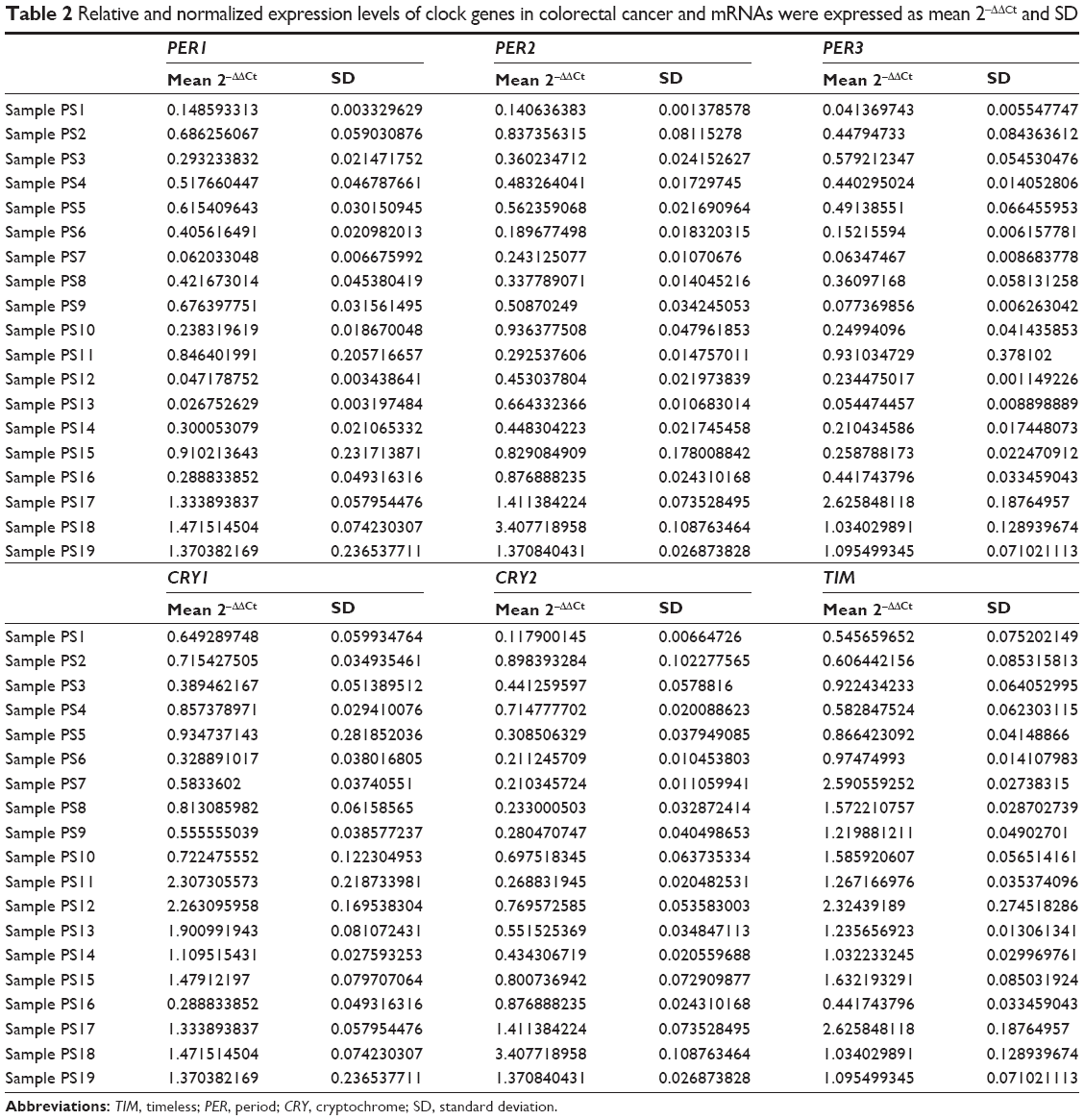 | Table 2 Relative and normalized expression levels of clock genes in colorectal cancer and mRNAs were expressed as mean 2−ΔΔCt and SD |
Microsatellite instability
The Bethesda panel of microsatellite (BAT25, BAT26, D5S346, D17S250, and D2S123) was applied for the microsatellite instability (MSI) analysis, which was evaluated by means of multiplex PCR and polyacrylamide gel electrophoresis analysis. Then, the tumors were accordingly classified into the following categories: microsatellite stable, low microsatellite instability (MSI-L), and high microsatellite instability (MSI-H).23
Statistical analysis
The SPSS Statistical Package was used for the statistical analyses. Comparing with gene expression levels of the normal mucosa, those from the adjacent CRC tissues were calculated by the formula 2−ΔΔCt, with the reported values of median, 25th percentile (Q1), and 75th percentile (Q3). The normal distribution of the continuous variables was verified by the Shapiro–Wilk test and the one-sample Kolmogorov–Smirnov test. By addressing with a nonparametric Wilcoxon signed-rank test, the statistical significance of the up- or downregulation for non-normal distribution of the 2−ΔΔCt transformation was assessed. The differences among the groups for normally distributed variables were compared by Student’s t-test or by analysis of variance, while those for non-normally distributed variables were compared by the Mann–Whitney rank sum test or the Kruskal–Wallis rank sum test. The correlation between mRNA expression levels and clinical and pathological features was evaluated by Spearman’s test, and the survival rates were calculated by the Kaplan–Meier method via censored data analysis. A P-value <0.05 was considered as statistically significant.
Results and discussion
The expression levels of the core clock genes (PER1, PER2, PER3, CRY1, CRY2, and TIM) from 19 paired normal and colon cancer tissues were evaluated, in order to identify those differentially expressed in primary CRC. Figure 1 presents the relative expression levels of clock genes in CRC tissue samples according to the data in Table 2. Comparing with normal tissue after normalizing to 1, the following five genes in the tumor samples were down-regulated: PER1 (median =0.42, Q1–Q3=0.24–0.85, P=0.002), PER2 (median =0.51, Q1–Q3=0.34–0.88, P=0.011), PER3 (median =0.36, Q1–Q3=0.15–0.58, P=0.003), and CRY2 (median =0.55, Q1–Q3=0.27–0.90, P=0.012). The CRY1 gene did not show different expression (median =0.93, Q1–Q3=0.65–1.48, P=0.600). It is obviously shown in Figure 1 that TIM was upregulated (median =1.22, Q1–Q3=0.92–1.63, P=0.044).
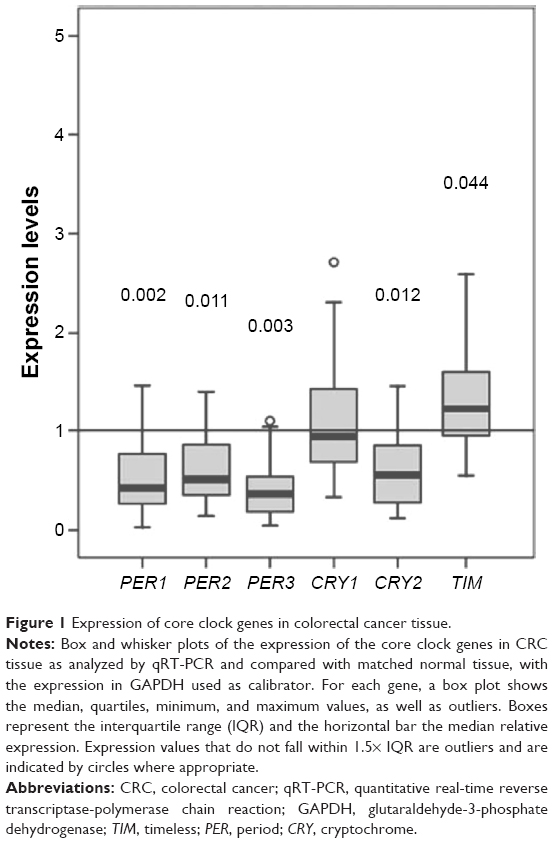 | Figure 1 Expression of core clock genes in colorectal cancer tissue. |
The association between gene expression levels (PER, CRY, and TIM) and clinical and pathological features, such as patient age and sex, tumor location and stage, and MSI status, is listed in Table 3. A significant association was observed for the CRY1 and TIM genes. In particular, lower expression level of CRY1 in the tumor mucosa was found in the 68–75-year-old subjects (P=0.026) and female patients (P=0.005), whereas higher expression level of CRY1 in the tumor mucosa was found in cancers located in the distal colorectal segments (P=0.015), which was confirmed by Spearman’s correlation (r=0.521, P=0.02, slope =0.519) (Figure 2). A significant association could be observed between high TIM mRNA expression level in tumor mucosa and stages III–IV (P=0.005), involving lymph nodes (P=0.005), in particular, of proximal lymph nodes (P=0.013), and DNA mismatch repair proficiency and MSI (P=0.015) (Table 3).
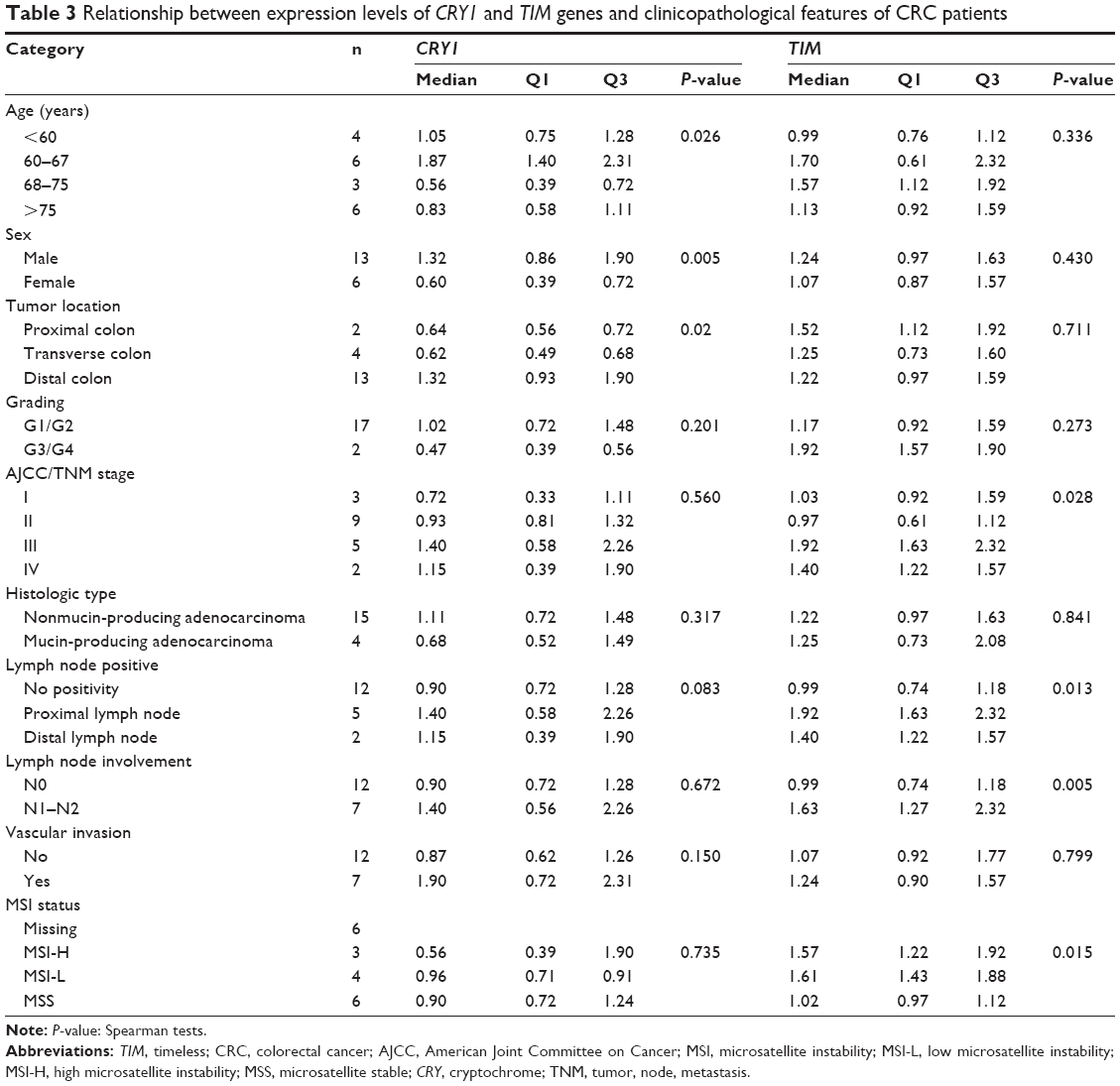 | Table 3 Relationship between expression levels of CRY1 and TIM genes and clinicopathological features of CRC patients |
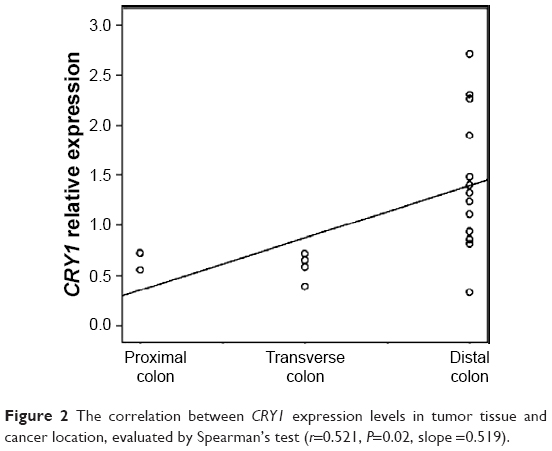 | Figure 2 The correlation between CRY1 expression levels in tumor tissue and cancer location, evaluated by Spearman’s test (r=0.521, P=0.02, slope =0.519). |
CRC patients with lower expression of PER1 (P=0.010) and PER3 (P=0.010) in the tumor tissue showed significantly poorer survival rates, according to the Kaplan–Meier method for the analysis of censored data. No statistically significant decrease in survival could be evidenced in patients with low expression level of PER2 (P=0.143), CRY1 (P=0.143), CRY2 (P=0.236), and TIM (P=0.491) in the tumor tissue (Figure 3).
 | Figure 3 Cumulative survival of patients according to the expression levels of PER1, PER2, PER3, CRY1, CRY2, and TIM in tumor tissue of CRC patients. |
The core clock genes drive and activate downstream clock-controlled genes and the control of tissue/organ function.24,25 In addition, altered expression levels of PER and CRY genes have been evidenced in intestinal biopsies of diseased intestinal segments of patients affected by ulcerative colitis and Crohn’s disease, and inflammation is considered to favor the development of neoplastic disease. Considering the reported involvement of the circadian clock in several cancers, qRT-PCR was applied to examine the expressions of PER1, PER2, PER3, CRY1, CRY2, and TIM in CRC and matched normal colorectal tissues. The expression level of some clock genes in tumor tissue was found to be significantly decreased, as in the case of PER1, PER2, PER3, and CRY2, whereas that of TIM (P=0.044) was higher, and poorer survival rate was associated with lower expression levels of PER1 and PER3 in the tumor tissue in a statistically significant way.
Our results are in agreement with previous reports describing alterations in the expression of clock genes in CRC. In particular, the decreased expression of PER genes seems to be most relevant for the process of oncogenesis and tumor progression. PER1 and PER2 are related to the pathways for ATM-Chk1/Chk2 DNA damage response, β-catenin modulate, and proliferation of colon and noncolon cancer cells. In contrast, clock function may be altered by intestinal tumorigenesis, leading to increase in β-catenin-destabilizing PER2 protein. PER1 could directly interact with ATM in vitro as a response to radiation. In mice, PER2 mutation would result in change in the temporal gene expression for regulation of the cell cycle and tumor suppression (C-MYC, CYCLIN A, CYCLIN D1, GADD45A, and MDM-2), DNA-damage response deregulation, accelerating intestinal polyp formation in APCMin/+ mice, and increase in neoplastic growth.26,27 We found no statistically significant difference in the CRY1 expression level between CRC and matched normal tissue. CK1ε phosphorylates PER and CRY proteins and β-catenin, thereby facilitating their ubiquitination and proteasomal degradation. In vitro and in vivo studies strongly confirm that CKIε is a key factor in the early stages of tumorigenesis, predisposing to colon cancer. As tumor cells have more dependence on the kinase activity of CK1ε than normal ones, a specific kinase inhibitor to CK1ε could induce tumor cell-selective cytotoxicity.28,29
A correlation between low expression of PER1 gene and liver metastasis and the relationship between high expression of PER2 gene and significantly better outcomes were reported in a previous study.30 In our study, CRY1 mRNA expression levels in tissues of CRC were significantly related to patient age (with the lowest levels detected in the age range of 68–75 years), sex (with the lowest levels detected in female patients), and cancer location (with the highest levels detected in tumors located in the distal colon).
The clock gene machinery, which controls the system functions of hepatic, intestinal, renal detoxification, and xenobiotic detoxification, exhibits circadian variation of activity, determining time-dependent toxicity of xenobiotics and drugs. Time-of-day-dependent variation in drug toxicity, metabolism, and effectiveness provides the basis for the specific 24-hour period timing of drug administration during the chronomodulated cancer chemotherapy of advanced-stage CRC.
For example, diurnally active patients deliver oxaliplatin in the afternoon and fluorouracil and leucovorin at night.31 For the sex dependency, female CRC patients have been reported to have shorter survival and greater toxicities, according to European Organization for Research and Treatment of Cancer (EORTC) Chronotherapy Group trial.32 The male and female mammals (including humans) have different xenobiotic detoxification and metabolic pathways. The change of sexual dimorphism in hepatic drug metabolism could be observed in double mutant CRY1−/− CRY2−/− male mice. Once the CRY genes are inactivated, the male and female mice have similar expression levels of sex-specific liver products, such as several cytochrome P450 enzymes.33 By transferring this evidence to humans, it could be suggested that the decreased expression level of the CRY1 gene in female CRC patients might lead to the different median survival and the increase in the toxicity after the administration of chronic-modulated chemotherapy.
Comparing with the normal mucosa, the TIM mRNA levels observed in the CRC tissue were significantly related to American Joint Committee on Cancer (AJCC)/TNM stage (highest levels in TNM stages III–IV), lymph node involvement (highest levels in the case of positive lymph nodes, especially proximal lymph node involvement), and MSI (highest levels in MSI-H and MSI-L). Most of the cytotoxic anticancer drugs would damage DNA and activate DNA checkpoints for approving the attempted DNA repair, which is important to the survival of the cells. However, the cytotoxicity of anticancer drugs may be reduced. For ATM-dependent Chk2-mediated signaling of doxorubicin-induced DNA double-strand breaks, TIM plays an essential role. Moreover, the arresting of doxorubicin-induced G(2)/M cell cycle would be significantly attenuated by downregulation of TIM siRNA, sensitizing cancer cells to doxorubicin-induced cytotoxicity. Hence, the variation in drug sensitivity could be predicted by TIM mutation in human cancers. In order to enhance the cytotoxic effectiveness of chemotherapeutic drugs for activating DNA response pathways in cancer cells, the TIM inhibition becomes a potential novel target for anticancer drugs.34,35 In our study, TIM expression was also significantly associated with MSI. Approximately 15% of CRCs are diagnosed by defects in mismatch repair system of DNA, leading to MSI and generating many substitution, insertion, deletion, and mutations. As mainly targeting to the microsatellite sequences, these mutations could lead to reading frame variation, further resulting in truncation or alterations in protein. The mRNA expressions with such frameshift mutations could be decreased with the presence of premature stop codons, leading to some mutant mRNA degradation via the nonsense-mediated decay pathway.7,36 As better outcome with irinotecan-containing regimens was shown in MSI-H tumors than with 5-fluorouracil-containing treatments, the MSI status of CRC patients could affect the response to adjuvant chemotherapy.37 In MSI-H and MSI-L patients, the increase in the TIM expressions may be associated with the tumorigenesis process in CRC and reduction in the response to adjuvant chemotherapy.
Conclusion
In conclusion, there are differences in the expression levels of PER, CRY, and TIM genes in CRC tissues compared with matched normal ones, and the altered expression might influence the process of carcinogenesis and various aspects of host–tumor characteristics and interactions.
Disclosure
The authors report no conflicts of interest in this work.
References
Lemmer B. Discoveries of rhythms in human biological functions: a historical review. Chronobiol Int. 2009;26:1019–1068. | ||
Duguay D, Cermakian N. The crosstalk between physiology and circadian clock proteins. Chronobiol Int. 2009;26:1479–1513. | ||
Hastings MH, Reddy AB, Maywood ES. A clockwork web: circadian timing in brain and periphery, in health and disease. Nat Rev Neurosci. 2003;4:649–661. | ||
Jud C, Chappuis S, Revell VL, et al. Age-dependent alterations in human PER2 levels after early morning blue light exposure. Chronobiol Int. 2009;26:1462–1469. | ||
Agostino PV, Harrington ME, Ralph MR, Golombek DA. Casein kinase-1-epsilon (CK1epsilon) and circadian photic responses in hamsters. Chronobiol Int. 2009;26:126–133. | ||
Nagoshi E, Saini C, Bauer C, Laroche T, Naef F, Schibler U. Circadian gene expression in individual fibroblasts: cell-autonomous and self-sustained oscillators pass time to daughter cells. Cell. 2004;119:693–705. | ||
Smith KD, Fu MA, Brown EJ. Tim-Tipin dysfunction creates an indispensible reliance on the ATR-Chk1 pathway for continued DNA synthesis. J Cell Biol. 2009;187:15–23. | ||
Erren TC, Groβ JV, Meyer-Rochow VB. Light, clocks, mood, and cancer: consolidation and novel tests of latitude and instability hypotheses. Chronobiol Int. 2011;28:471–473. | ||
Kloog I, Haim A, Stevens RG, Portnov BA. Global co-distribution of light at night (LAN) and cancers of prostate, colon, and lung in men. Chronobiol Int. 2009;26:108–125. | ||
Chu G, Yoshida K, Narahara S, et al. Alterations of circadian clockworks during differentiation and apoptosis of rat ovarian cells. Chronobiol Int. 2011;28:477–487. | ||
Hayashida S, Kuramoto Y, Koyanagi S, et al. Peroxisome proliferator-activated receptor-α mediates high-fat, diet-enhanced daily oscillation of plasminogen activator inhibitor-1 activity in mice. Chronobiol Int. 2010;27:1735–1753. | ||
Im JS, Jung BH, Kim SE, Lee KH, Lee JK. Per3, a circadian gene, is required for Chk2 activation in human cells. FEBS Lett. 2010;584:4731–4734. | ||
Yang X, Wood PA, Ansell C, Hrushesky WJ. Circadian time-dependent tumor suppressor function of period genes. Integr Cancer Ther. 2009;8:309–316. | ||
Climent J, Perez-Losada J, Quigley DA, et al. Deletion of the PER3 gene on chromosome 1p36 in recurrent ER-positive breast cancer. J Clin Oncol. 2010;28:3770–3778. | ||
Alhopuro P, Björklund M, Sammalkorpi H, et al. Mutations in the circadian gene CLOCK in colorectal cancer. Mol Cancer Res. 2010;8:952–960. | ||
Yang X, Wood PA, Ansell CM, et al. Beta-catenin induces beta-TrCP-mediated PER2 degradation altering circadian clock gene expression in intestinal mucosa of ApcMin/+ mice. J Biochem. 2009;145:289–297. | ||
Krugluger W, Brandstaetter A, Kállay E, et al. Regulation of genes of the circadian clock in human colon cancer: reduced period-1 and dihydropyrimidine dehydrogenase transcription correlates in high-grade tumors. Cancer Res. 2007;67:7917–7922. | ||
Mostafaie N, Kállay E, Sauerzapf E, et al. Correlated downregulation of estrogen receptor beta and the circadian clock gene Per1 in human colorectal cancer. Mol Carcinog. 2009;48:642–647. | ||
Haggar FA, Boushey RP. Colorectal cancer epidemiology: incidence, mortality, survival, and risk factors. Clinlical Colon Rectal Surgery. 2009;22:191–197. | ||
Bijan M-D, Azadeh S. An overview of colorectal cancer survival rates and prognosis in Asia. World J Gastrointest Oncol. 2012;4:71–75. | ||
Barber RD, Harmer DW, Coleman RA, Clark BJ. GAPDH as a housekeeping gene: analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genomics. 2005;21:389–395. | ||
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T). Methods. 2001;25:402–408. | ||
Boland CR, Thibodeau SN, Hamilton SRA. National Cancer Institute workshop on microsatellite instability for cancer detection and familial predisposition: development of international criteria for the determination of microsatellite instability in colorectal cancer. Cancer Res. 1998;58:5248–5257. | ||
Hrushesky WJ, Grutsch J, Wood PA, et al. Circadian clock manipulation for cancer prevention and control and the relief of cancer symptoms. Integr Cancer Ther. 2009;8:387–397. | ||
Yamamura Y, Yano I, Kudo T, Shibata S. Time-dependent inhibitory effect of lipopolysaccharide injection on Per1 and Per2 gene expression in the mouse heart and liver. Chronobiol Int. 2010;27:213–232. | ||
Antoch MP, Kondratov RV, Takahashi JS. Circadian clock genes as modulators of sensitivity to genotoxic stress. Cell Cycle. 2005;4:901–907. | ||
Yang X, Wood PA, Ansell CM, et al. The circadian clock gene Per1 suppresses cancer cell proliferation and tumor growth at specific times of day. Chronobiol Int. 2009;26:1323–1339. | ||
Modak C, Chai J. Potential of casein kinase I in digestive cancer screening. World J Gastrointest Oncol. 2009;15:26–33. | ||
Yang WS, Stockwell BR. Inhibition of casein kinase 1-epsilon induces cancer-cell-selective, PERIOD2-dependent growth arrest. Genome Biol. 2008;9:R92. | ||
Oshima T, Takenoshita S, Akaike M, et al. Expression of circadian genes correlates with liver metastasis and outcomes in colorectal cancer. Oncol Rep. 2011;25:1439–1446. | ||
Levi F, Schibler U. Circadian rhythms: mechanisms and therapeutic implications. Annu Rev Pharmacol Toxicol. 2007;47:593–628. | ||
Giacchetti S, Bjarnason G, Garufi C, et al; European Organisation for Research and Treatment of Cancer Chronotherapy Group. Phase III trial comparing 4-day chronomodulated therapy versus 2-day conventional delivery of fluorouracil, leucovorin, and oxaliplatin as first-line chemotherapy of metastatic colorectal cancer: The European Organisation for Research and Treatment of Cancer Chronotherapy Group. J Clin Oncol. 2006;24:3562–3569. | ||
Bur IM, Cohen-Solal AM, Carmignac D, et al. The circadian clock components CRY1 and CRY2 are necessary to sustain sex dimorphism in mouse liver metabolism. J Biol Chem. 2009;284:9066–9073. | ||
Yang X, Wood PA, Hrushesky WJ. Mammalian TIMELESS is required for ATM-dependent CHK2 activation and G2/M checkpoint control. J Biol Chem. 2010;29:3030–3034. | ||
Kemp MG, Akan Z, Yilmaz S, et al. Tipin-replication protein A interaction mediates Chk1 phosphorylation by ATR in response to genotoxic stress. J Biol Chem. 2010;285:16562–16571. | ||
Unsal-Kaçmaz K, Chastain PD, Qu PP, et al. The human Tim/Tipin complex coordinates an Intra-S checkpoint response to UV that slows replication fork displacement. Mol Cell Biol. 2007;27:3131–3142. | ||
Bertagnolli MM, Niedzwiecki D, Compton CC, et al. Microsatellite instability predicts improved response to adjuvant therapy with irinotecan, fluorouracil, and leucovorin in stage III colon cancer: cancer and Leukemia Group B Protocol 89803. J Clin Oncol. 2009;27:1814–1821. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.