Back to Journals » International Journal of Women's Health » Volume 17
Exploring the Potential Causal Relationship of Galectin Levels and Gynecologic Cancers: A Two-Sample Mendelian Randomization Study
Authors Dong H, Nie S, Ni N, Guo L, Liu Q
Received 5 January 2025
Accepted for publication 3 November 2025
Published 2 December 2025 Volume 2025:17 Pages 5017—5034
DOI https://doi.org/10.2147/IJWH.S515826
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Vinay Kumar
Hongguo Dong,* Sipei Nie,* Na Ni, Ling Guo, Qiaoling Liu
Department of Gynecology and Obstetrics, Affiliated Jiangning Hospital of Nanjing Medical University, Nanjing, Jiangsu, 211100, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qiaoling Liu, Department of Gynecology and Obstetrics, Affiliated Jiangning Hospital of Nanjing Medical University, Nanjing, Jiangsu, People’s Republic of China, Email [email protected]
Background: Previous studies have reported the correlation between single galectin levels and gynecologic cancers. The present study used the Mendelian randomization (MR) technique to evaluate the causal relationship between galectin levels and gynecologic cancers.
Methods: Data on galectin levels (3394 individuals of the Gal-3 dataset, 3301 individuals of Gal-1, Gal-2, Gal-4, Gal-7, Gal-8, Gal-9, and Gal-10 datasets) and gynecologic cancers were obtained from the IEU Open genome-wide association study (GWAS) project, Ovarian Cancer Association Consortium (OCAC) and FinnGen consortium. Two-sample MR was performed to determine the causal relationship between galectin levels and gynecologic cancers. Meanwhile, a bi-directional MR analysis was also conducted to examine the direction of the causal relations. The results were expressed as odds ratios (ORs) and 95% confidence intervals (CIs).
Results: Gal-3 (OR=1.001, 95% CI: 1.000– 1.001) and Gal-8 (OR=1.001, 95% CI: 1.000– 1.002) may indicate small effects on cervical cancer. Gal-1 was suggested as risk factors for ovarian cancer (OR=1.108, 95% CI: 1.033– 1.188), high grade serous ovarian cancer (OR=1.106, 95% CI: 1.019– 1.202), and invasive mucinous ovarian cancer (OR=1.279, 95% CI: 1.041– 1.573). Gal-9 showed modest effect in ovarian cancer (OR=1.070, 95% CI: 1.000– 1.144), but significantly associated with an elevated risk of mucinous ovarian cancer (OR=1.242, 95% CI: 1.015– 1.520). Gal-10 was suggested as a protective factor against ovarian cancer (OR=0.929, 95% CI: 0.867– 0.994) and clear cell ovarian cancer (OR=0.803, 95% CI: 0.653– 0.987). Gal-2 (OR=0.779, 95% CI: 0.654– 0.929) and Gal-4 (OR=0.786, 95% CI: 0.639– 0.966) were found as protective factors for malignant neoplasm of corpus uteri. Gal-2 (OR=0.921, 95% CI: 0.851– 0.997) and Gal-10 (OR=0.892, 95% CI: 0.815– 0.977) were suggested as protective factors against endometrial cancer (endometrioid histology). Similarly, higher Gal-4 was also associated with decreased risk of endometrial cancer (OR=0.902, 95% CI: 0.840– 0.969) and endometrial cancer (endometrioid histology) (OR=0.846, 95% CI: 0.778– 0.920).
Conclusion: The study explored potential causal relationships between galectin levels and gynecologic cancers. While some associations were modest, the consistent directionality across sensitivity analyses and the biological plausibility of galectins in tumor immunology suggest that these proteins warrant further investigation as potential biomarkers and therapeutic targets. Future research should validate these findings in clinical cohorts and explore underlying mechanisms.
Keywords: galectin, gynecologic cancers, risk factor, protective factor, Mendelian randomization
Introduction
Gynecologic cancers, including ovarian, endometrial, and cervical cancers, represent a significant global health burden, accounting for substantial morbidity and mortality among women worldwide.1 Despite advances in screening, diagnosis, and treatment, the prognosis for patients with advanced-stage gynecologic cancers remains poor. Identification of novel biomarkers and a deeper understanding of the underlying molecular mechanisms driving these malignancies are therefore critical to improving early detection, prognosis, and therapeutic strategies. One such promising avenue of research lies in exploring galectins, a family of β-galactoside-binding proteins, which have been implicated in various pathological processes, including tumorigenesis, immune evasion, and metastasis.2,3
Galectins were the first family of glycan binding proteins for which immune regulatory activity. In vertebrates, more than 16 galectins have been characterized (galectin-1–galectin-16).4 It has been reported that galectins play varying roles in distinct tissue types and have been implicated in different diseases, including cancer.5 Galectins are involved in numerous cellular functions, such as cell adhesion, migration, proliferation, and apoptosis, and their dysregulation has been linked to cancer progression and poor clinical outcomes.2 Among these 16 members in galectins family, Gal-1, Gal-3 and Gal-9 have been awarded a key role in tumorigenesis and progression in cancers. Gal1 was the first identified member of the galectin family, which has been ubiquitously involved in promoting tumour progression, immune escape, and metastatic dissemination. Given the limitations and toxic side effects associated with current treatment options, alternative strategies targeting Gal-1 have been explored for their therapeutic potential.6 Galectin-3 is located within the cytoplasm and the nucleus but can be transported onto the cell surface, extra-cellular space, and the circulation via nonclassical secretory pathways. Gal-3 is involved with several cellular activities, such as apoptosis, cell migration, proliferation, angiogenesis and cancer cell stemness.7 Interestingly, Gal-9 exhibits multifaceted roles that are intricately linked to the location of its binding partners at the cellular level. Within the cell, Gal-9 regulates cell adhesion molecules and enhances adhesion, thereby reducing the capacity for tumor invasion and metastasis. Conversely, extracellular Gal-9 serves a completely different role, particularly within the tumor microenvironment.8 Despite the accumulating studies pointing to galectin’s role in cancer biology, establishing a definitive causal relationship between galectin expression and the onset or progression of gynecologic cancers remains elusive. Previous studies reported Gal-1 level was significantly higher in patients with ovarian cancer than in health and closely associated with the invasion and metastasis of ovarian cancer.9,10 In addition, higher Gal-1 and Gal-9 levels were also observed in ovarian tumor samples and were associated with disease progression and worse survival.10,11 Besides, Gal-3, −7,-8 and −9 were also reported to be involved in tumorigenesis and progression of ovarian cancer.12 In vitro experiments also showed that the high expression of Gal-1 and Gal-3 could promote the growth of cervical cancer cells.13,14 Gal-2 and Gal-4 show opposite biological effects in different cancers, which may be related to the expression of different galectin-binding glycans.15–17 However, due to the fact that there are some conflicting evidence of these galectins existing in gynecologic cancers, further researches should be carried out to investigate the detailed function.
Mendelian randomization (MR), which leverages genetic variants as instrumental variables to infer causal relationships between exposures and outcomes, has emerged as a powerful tool in epidemiology.18 By utilizing genetic variants as instrumental variables, MR circumvents some confounding and reverse causation dilemmas that often plague traditional observational studies.19 Furthermore, as the genetic code is not influenced by environmental factors or preclinical diseases, it is also less susceptible to bias caused by reverse causation. MR has been widely used in the etiology of gynecological cancers.20,21 Recent MR studies have shown that Gal-3 is a risk factor for heart failure, Parkinson’s disease, and gestational hypertension.22–24 The relationship between galectin levels and gynecology cancers remains unclear. The study aimed to investigate whether galectin levels are causally associated with the risk of gynecologic cancers, including cervical, ovarian, endometrial cancers, and malignant neoplasm of corpus uteri. In the present study, we used data on galectin levels (Gal-1, Gal-2, Gal-3, Gal-4, Gal-7, Gal-8, Gal-9, and Gal-10) from the IEU Open genome-wide association study (GWAS) project and gynecologic cancers were obtained from GWAS project, Ovarian Cancer Association Consortium (OCAC) and FinnGen consortium to explore the potential causal relationship of galectin levels and gynecologic cancers by two-sample MR analysis. Meanwhile, a bi-directional MR analysis was also conducted to examine the direction of the causal relations. The findings would provide valuable insights into the molecular mechanisms underpinning gynecological cancer pathogenesis and highlight potential biomarkers or therapeutic targets for intervention.
Methods
Study Design
A univariable two-sample MR method was used to investigate the relationship between galectin levels and gynecologic cancers. The three assumptions of this study are presented in Figure 1. The single nucleotide polymorphisms (SNPs) that were selected as instrumental variables (IVs) were supposed to fulfill the three assumptions: (1) SNPs showed an association with the galectin levels, (2) SNPs were not linked to any confounder variables between exposure and outcomes, and (3) SNPs only affect the outcome via the exposure.
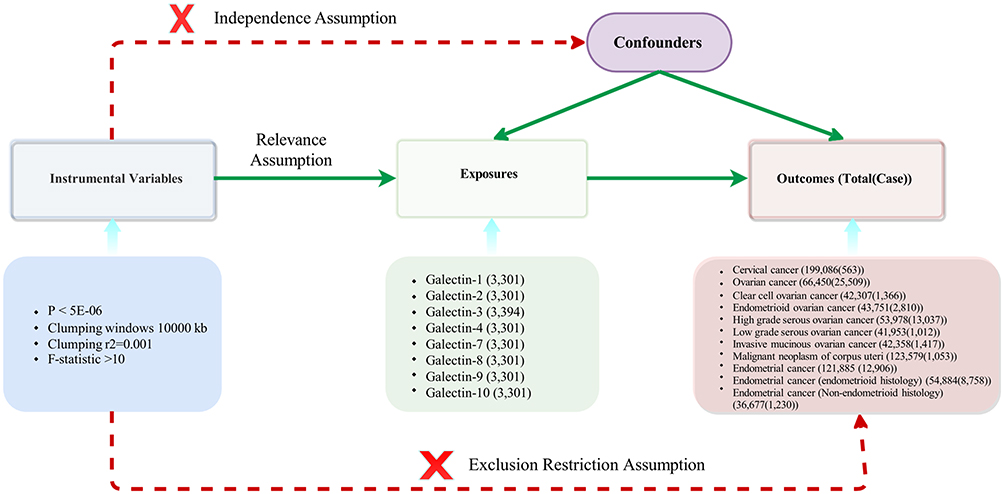 |
Figure 1 The directed acyclic graph of exposures and outcomes. |
Data Sources
For serum galectin, summary statistics data of SNPs were extracted from a large-scale genome-wide association study (GWAS). The Gal-3 dataset includes 3394 individuals of European ancestry.24 And 3301 European ancestry individuals were included in the Gal-1, Gal-2, Gal-4, Gal-7, Gal-8, Gal-9, and Gal-10 datasets.25 Data concerning cervical cancer (563 cases of 199086 individuals), endometrial cancer (12,906 cases of 21,885 individuals), endometrial cancer (endometrioid histology) (8758 cases of 54884 individuals), and endometrial cancer (Non-endometrioid histology) (1230 cases of 36,677 individuals) were sourced from the MRC Integrative Epidemiology Unit (IEU) Open GWAS database.26 Data on ovarian cancer (25509 cases of 66450 individuals), clear cell ovarian cancer (1366 cases of 42307 individuals), endometrioid ovarian cancer (2810 cases of 43,751 individuals), high grade serous ovarian cancer (13037 cases of 53978 individuals), low grade serous ovarian cancer (1012 cases of 41953 individuals), and invasive mucinous ovarian cancer (1417 cases of 42358 individuals) were obtained from the Ovarian Cancer Association Consortium (OCAC).27 Publicly available summary statistics from the FinnGen consortium (www.finbb.fi) provided data on the malignant neoplasm of corpus uteri (1053 cases of 123,579 individuals). Detailed information is expressed in Table S1. The original GWASs have obtained adequate patient consent and ethical approval. Due to the use of de-identified data, ethical approval was not required for our study.
Selection of SNPs
First, SNPs were selected of those associated with galectin at a genome-wide significance threshold (P <5×10−6) from the corresponding datasets to ensure a close relationship existed between the IVs and galectin levels. Then, to eliminate linkage disequilibrium (LD), the clumping process was implemented with R2 < 0.001 and kb = 10000. Furthermore, to ensure the effect of the SNPs on the galectin correspond to the same allele as the effect of the outcome. As the same letters on the forward and reverse strands, palindromic SNPs were deleted to prevent unexpected biases. Finally, the estimated variance in galectin explained by each SNP and the corresponding F statistics were calculated to evaluate the strength of the IVs. To ensure the use of robust genetic instruments, the F statistics should exceed a threshold of 10.28 The detailed screening process of SNPs was shown in Table S2.
Statistical Analysis IVs
A two-sample MR method was performed to explore the causal direction of the relationship between galectin levels and gynecologic cancers. In the primary analysis, we computed a Wald ratio for each genetic variant and summarized these estimates through the inverse-variance weighted (IVW) method. The multiplicative random-effects IVW method offers a succinct estimation while considering potential heterogeneity among SNP-derived Wald ratio estimates.29 Random-effects IVW models are utilized in the presence of heterogeneity; otherwise, a fixed-effect model is used. The heterogeneity was detected using Cochran’s Q statistics and the two-sample MR package between instruments. A Q larger than the number of instruments minus one provides evidence for heterogeneity and invalid instruments, or Q statistics significant at a P-value <0.05 can imply the presence of heterogeneity. Alternative methods were conducted, such as sensitivity analysis, including weighted median, weighted mode, and MR-pleiotropy residual sum and outlier (MR-PRESSO). Leave-one-out analysis was used to detect heterogeneity across the SNPs, while the MR-Egger intercept test was used to detect potential horizontal pleiotropy. A P-value of >0.05 suggested no evidence of heterogeneity or pleiotropy. MR Steiger directionality tests were conducted to examine whether exposure was directionally causal for the outcome. The results were expressed as odds ratios (ORs) and 95% confidence intervals (CIs). Statistical analyses were performed using R version 4.2.0 (Institute for Statistics and Mathematics, Vienna, Austria) with the R package “TwoSampleMR”.
Results
Causal Associations Between Galectin Levels and Cervical Cancer
By exploring the potential causal relationship between galectin levels and cervical cancer by analysis, Gal-3 and Gal-8 showed modest effect in cervical cancer. As shown in Figure 2, Gal-3 (OR=1.001, 95% CI: 1.000–1.001) (Power=0.4%) and Gal-8 (OR=1.001, 95% CI: 1.000–1.002) (Power=1.3%) might be associated with an increased risk of cervical cancer. The results of MR-Steiger test are both “true” (p <0.05). Apart from these two galectin levels showing association with cervical cancer, the remaining galectin species are not causally associated with cervical cancer. Figures S1 and S2 were the scatter plots of the MR analysis.
 |
Figure 2 The forest plot of MR analysis between galectin levels and cervical cancer. A total of 8 galectin types were included, two of which were significantly associated with cervical cancer (Gal-3 and Gal-8). |
Causal Associations Between Galectin Levels and Ovarian Cancer
In the MR analysis of the association between galectin levels and ovarian cancer, Gal-1 was suggested as a risk factor for ovarian cancer (OR=1.108, 95% CI: 1.033–1.188, power: 70.7%), high grade serous ovarian cancer (OR=1.106, 95% CI: 1.019–1.202, power: 43.9%), and invasive mucinous ovarian cancer (OR=1.279, 95% CI: 1.041–1.573, power: 45.4%). In ovarian cancer, Gal-9 showed a possible weak effect (OR=1.070, 95% CI: 1.000–1.144, power: 29.9%). This association was nominally significant (P<0.05) but did not meet multiple testing thresholds. The results of MR-PRESSO test and the radial MR also suggested that Gal-9 was associated with an elevated risk of ovarian cancer. The results of weighted median, weighted mode test are consistent with this finding. Besides, Gal-9 was associated with an elevated risk of invasive mucinous ovarian cancer (OR=1.242, 95% CI: 1.015–1.520, power: 30.6%). Conversely, Gal-10 was suggested as a protective factor against ovarian cancer (OR=0.929, 95% CI: 0.867–0.994, power: 20%) and clear cell ovarian cancer (OR=0.803, 95% CI: 0.653–0.987, power: 58.3%). The MR forest plot was depicted in Figure 3, while the scatter plots were illustrated in Figures S3–S9. The results of MR-Steiger test are “true” in above analysis.
 |
Figure 3 The forest plot of MR analysis between galectin levels and ovarian cancer. A total of 8 galectin types were included, three of which were significantly associated with ovarian cancer (Gal-1, Gal-9, and Gal-10). |
Causal Associations Between Galectin Levels and Malignant Neoplasm of Corpus Uteri
In the MR analysis examining the causal associations between galectin levels and malignant neoplasm of corpus uteri, Gal-2 (OR=0.779, 95% CI: 0.654–0.929, power: 26.2%) and Gal-4 (OR=0.786, 95% CI: 0.639–0.966, power: 13.3%) were suggested as protective factors for malignant neoplasm of corpus uteri. Apart from the two galectins showing potential causal relationships with malignant neoplasm of corpus uteri. The results of MR-Steiger test are “true” in above analysis. The remaining galectins were found to have no significant causal relationship with malignant neoplasm of corpus uteri. The MR forest plot was presented in Figure 4, while the scatter plots were illustrated in Figures S10 and S11.
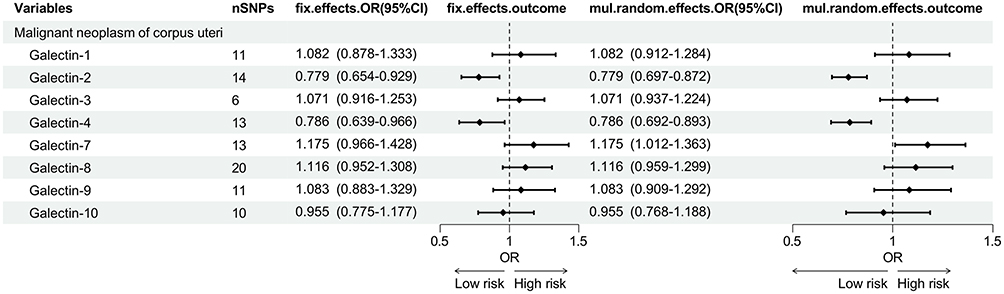 |
Figure 4 The forest plot of MR analysis between galectin levels and malignant neoplasm of corpus uteri. A total of 8 galectin types were included, two of which were significantly associated with malignant neoplasm of corpus uteri (Gal-2 and Gal-4). |
Causal Associations Between Galectin Levels and Endometrial Cancer
In the MR analysis examining the causal associations between galectin levels and endometrial cancer, as depicted in Figure 5 and S12–15, Gal-2 (OR=0.921, 95% CI: 0.851–0.997, power: 50.7%) and Gal-10 (OR=0.892, 95% CI: 0.815–0.977, power: 60.4%) were suggested as protective factors against endometrial cancer (endometrioid histology). Similarly, higher Gal-4 was also associated with decreased risk of endometrial cancer (OR=0.902, 95% CI: 0.840–0.969, power: 52.1%) and endometrial cancer (endometrioid histology) (OR=0.846, 95% CI: 0.778–0.920, power: 76.4%). The results of MR-Steiger test are “true” in above analysis.
 |
Figure 5 The forest plot of MR analysis between galectin levels and endometrial cancer. A total of 8 galectin types were included, three of which were significantly associated with ovarian cancer (Gal-2, Gal-4, and Gal-10). |
Sensitivity Analysis
The causal relationship with gynecologic cancers exhibits heterogeneity only in “Gal-2 with cervical cancer or low grade serous ovarian cancer”. In the remaining analyses, no significant heterogeneity was observed with a P-value >0.05 in Cochran Q tests. The MR-Egger intercept test indicates that the causal relationship between Gal-3 and endometrioid ovarian cancer exhibits horizontal pleiotropy. Therefore, the causal relationship between Gal-3 and endometrioid ovarian cancer is excluded. More results about sensitivity analysis are illustrated in Table 1. The leave-one-out analysis results suggested that no single SNP dominates the causal relationship (Figures S16–S30). MR-Steiger filtering detected no invalid genetic instruments for these analyses. Weighted mode, weighted median methods, MR-PRESSO, and radial tests yield similar causal estimates for magnitude and direction (Tables 2 and 3).
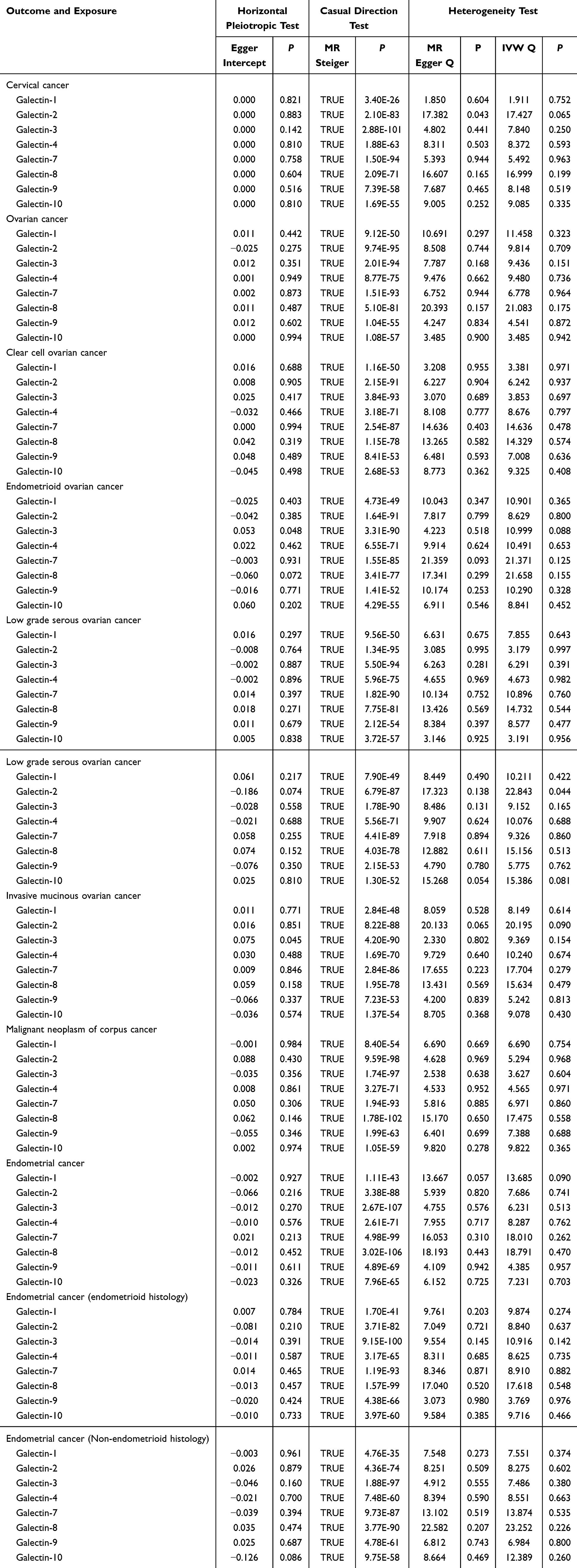 |
Table 1 The Sensitivity Analysis of the Causal Relationship Between Galectin Levels and Gynecologic Cancers |
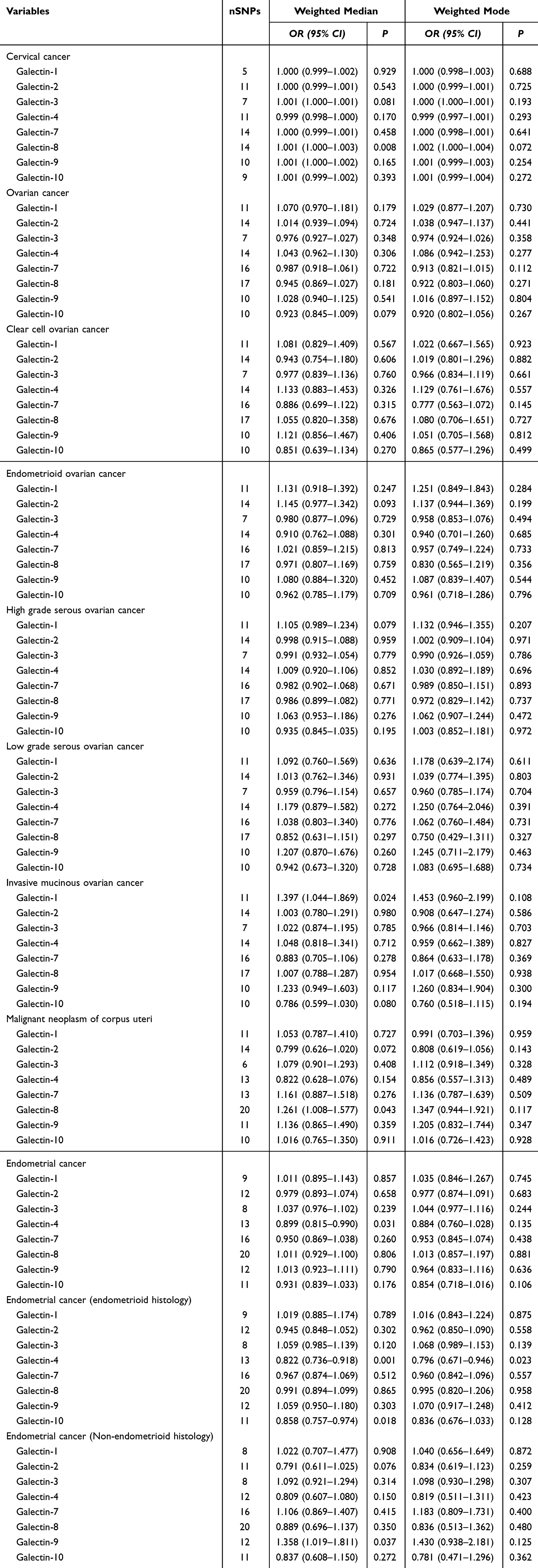 |
Table 2 Weighted Median and Weighted Mode Tests in MR |
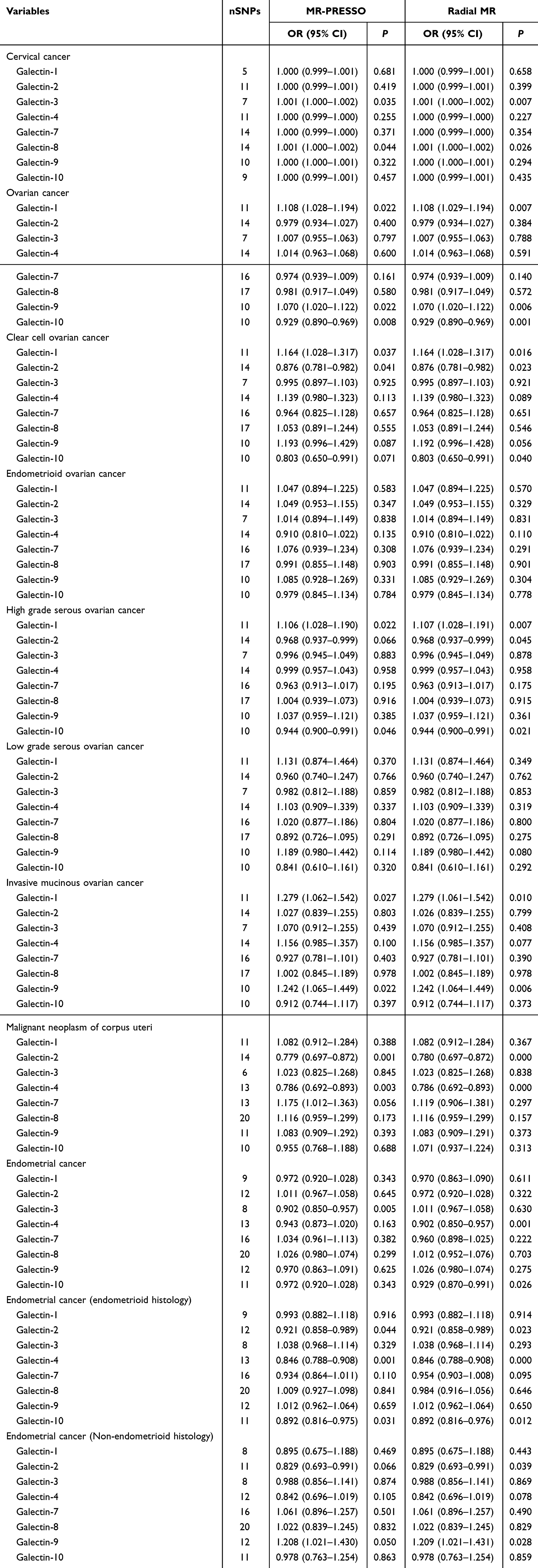 |
Table 3 The MR-PRESSO and Radial Test in MR |
Discussion
Our study explored the potential causal relationships between various galectin levels and the incidence of gynecology cancers, including ovarian, cervical, endometrial cancer, and malignant neoplasm of corpus uteri. We found that several galectins-Gal-1, −2, −3, −4, −8, −9, and −10 showed significant associations with the incidence of these cancers, suggesting a potential role for galectins in the pathogenesis of gynecologic malignancies. The findings provide novel insights into the molecular mechanisms underlying gynecologic cancer development and emphasize the importance of galectins as potential biomarkers or therapeutic targets.
Elevated Gal-1 levels were associated with higher incidences of ovarian cancer, particularly high grade serous ovarian cancer and invasive mucinous ovarian cancer. Gal-1 has been implicated in various aspects of tumor biology, including cell adhesion, migration, and immune escape.30 Gal-1 enhances metastasis and epithelial–mesenchymal transition (EMT) of ovarian cancer cells via promoting the activation of the mitogen-activated protein kinase (MAPK) Jun N-terminal kinase (JNK)/p38 signaling pathway, while EMT is a critical process for metastasis and tumor progression.31 Moreover, Gal-1 has been shown to suppress anti-tumor immune responses, particularly by impairing T-cell function and fostering a tumor-promoting microenvironment.32 The findings suggest that Gal-1 may be a better surrogate biomarker in ovarian cancer.9
In contrast, elevated Gal-2 was associated with a lower incidence of malignant neoplasm of corpus uteri and endometrial cancer, especially of endometrioid histology. Gal-2’s role in cancer remains unclear, but some studies suggest it may act as a tumor suppressor in certain contexts.15 Tumor inoculated with Gal-2 transfected triple-negative breast cancer 4T1 cells in BALB/c mice exhibited significantly accelerated growth compared to control vector-transfected cells, administration of an anti-Gal-2 antibody reduced tumor proliferation.33 Gal-2 binding to the Thomsen-Friedenreich antigen on mucin 1 (MUC1) enhanced cancer cell adhesion to the endothelium, promoting the secretion of metastasis-related cytokines.34,35 Gal-2 may regulate the immune response in the tumor microenvironment, and its increased expression could potentially restore immune surveillance, leading to a reduced incidence of cancers.
Our findings indicated Gal-3 and Gal-8 level may be associated with an increased risk of cervical cancer. However, their effect sizes (OR=1.001) suggest very small effects. The lower power of theses results might be limited by the small sample size of cancer cases. The results of weighted median, weighted mode test are consistent with the results of IVW method, which verifying the robustness of these associations. Thus, a larger sample size is needed to prove the causal relationship. Gal-3 is a multifunctional protein that plays a central role in inflammation, immune response, and tumor progression. It is related to promoting angiogenesis, cell proliferation, and survival, particularly through interactions with cell surface receptors like the epidermal growth factor receptor (EGFR).36 A case–control study in China also found allele C of rs4652 and allele T of rs11125 in the Gal-3 gene may be risk factors for cervical cancer.37 The upregulation in cervical cancer may facilitate tumor growth and progression through the modulation of the tumor microenvironment, and overexpression of those proteins was indicative of poor prognosis in patients with cervical cancer.7,38,39
Additionally, Gal-3 has been characterized as a promoter of EMT, a process linked to enhanced invasiveness in carcinoma cells.7 Gal-8 binds to glycoproteins on the surface of cancer cells, modulating cell–cell interactions and facilitating tumor cell migration and invasion.40,41 In cervical cancer, elevated Gal-8 expression has been linked to poor prognosis, possibly due to its role in promoting cell survival and resistance to apoptotic signals.42 Moreover, Gal-8 may interact with integrins and other adhesion molecules to promote the EMT process, further contributing to the metastatic potential of cervical cancer cells.43
Elevated Gal-4 levels were associated with a lower incidence of malignant neoplasm of corpus uteri, as well as endometrial cancer and its endometrioid histology. Though limited studies in gynecology cancers about Gal-4, the findings still indicate its anti-tumor properties.16 The expression of LGALS4 coded the Gal-4, was upregulated in 66.1% of the patients having ovarian carcinomas with a positive correlation to CA19.9, which suggests a potential role of Gal-4 in ovarian carcinomas.11,44 Gal-4 in cancer pathophysiology indicates the involvement of Gal-4 in the Wnt/β-catenin signaling pathway, leading to cell cycle progression, proliferation, and metastasis of cancer cells.16 In endometrial cancer, Gal-4 may inhibit tumor progression by promoting apoptosis and reducing angiogenesis.
Interestingly, no significant relationship was found between Gal-7 and gynecologic cancers. Unlike other members of the Galectin family, Gal-7 appears to have a more restricted role. Menkhorst et al reported Gal-7 production increased in endometrial cancer with increasing cancer grade.45 In cervical cancer, Gal-7 induced the transformation of HeLa cells to more invasive cells via elevating matrix metalloproteinase (MMP)-9 expression, which may be related to the p38 MAPK signaling pathway.32 Liu et al reported Gal-7 promotes cisplatin sensitivity, and it would be a potential target for potentiating efficacy in cervical cancer chemotherapy. This lack of association with gynecological malignancies may indicate a need for more focused research on the diverse functions of Gal-7.46
Elevated Gal-9 levels were associated with a higher incidence in especially invasive mucinous ovarian cancer and also showed a possible effect in ovarian cancer. Despite the issue of statistical significance boundaries, the association direction between galectin-9 and ovarian cancer is consistent with previous mechanism studies, and may provide potential target clues for future research. Gal-9 staining predominantly occurs in the cytoplasm of ovarian cancer cells.32 High Gal-9 expression is significantly linked to lower tumor stages, earlier International Federation of Gynecology and Obstetrics stages, and younger patient age.47 Gal-9 plays a role in various biological processes, including immune regulation and tumor progression. In ovarian cancer, Gal-9 may promote tumor growth and metastasis by modulating immune cell infiltration and the tumor microenvironment.32 Additionally, Gal-9 interacts with multiple immune checkpoints and has been shown to suppress T-cell responses, facilitating immune evasion.48
Finally, elevated Gal-10 levels were associated with a decreased incidence of ovarian cancer, including clear cell ovarian cancer and endometrial cancer with endometrioid histology. Most Gal-10-related studies were focused on its relationship with eosinophilic diseases.49,50 While less is known about Gal-10, it has been reported to play a role in inhibiting cell proliferation and inducing apoptosis, suggesting a potential tumor-suppressive function similar to that of Gal-4.51 The associations we observed may reflect its role in cellular processes that mitigate tumor progression.
The differential effects of galectins in different types of gynecologic cancers, as observed in this study, may reflect the unique molecular characteristics of each cancer type. For example, ovarian cancer is often associated with immune suppression and metastasis, which could explain the role of Gal-1 and Gal-9 in promoting cancer progression. In contrast, endometrial cancer may involve different regulatory mechanisms, with Gal-2 and Gal-4 potentially playing protective roles by enhancing immune surveillance and inhibiting tumor cell invasion.
Our study is the first to investigate the potential causal relationship between galectin levels and gynecology cancers using a two-sample MR approach, utilizing a large volume of GWAS data. The concordance across pleiotropy-robust methods strengthens causal inference though unmeasured confounding cannot be ruled out. Further, MR-Egger intercept and noted consistency across methods, which supported the primary IVW result, reducing concerns about directional pleiotropy.
However, there are limitations. The galectin data were derived from individuals of European ancestry, which may restrict the generalizability of the findings to other populations. Additionally, the lack of individual-level data prevents a thorough exploration of the non-linear causal relationship between galectin levels and gynecologic cancers. The further exploration is needed to articulate translational implications beyond genetic associations. Such as measuring plasma and tissue galectin levels in clinical prospective cohorts to assess prognostic values. For the significant galectins which exhibit predictive values in gynecologic cancers, the molecular functions and regulating mechanisms deserve further exploration by cell and animal assays. Screening for specific inhibitors/agonists and evaluate their effects for clinical application.
Conclusion
Our study highlights the potential causal relationships between galectin levels and gynecology cancers. Elevated levels of Gal-1, −3, −8, and −9 might be associated with an increased risk of ovarian cancer, particularly high grade serous and invasive mucinous subtypes, as well as cervical cancer. Conversely, elevated Gal-2 and Gal-4 levels appeared to reduce the risk of malignant neoplasm of corpus uteri and endometrial cancer, especially endometrioid histology. Gal-7 showed no significant association with gynecology cancers, while elevated Gal-10 was linked to a reduced incidence of ovarian and endometrial cancers. Although our findings were consistent across sensitivity analyses (MR-Egger, weighted median etc) and accounted for pleiotropy, the limitation of data and potential residual confounding mean that these associations should be interpreted as suggestive rather than definitive evidence of causality. Further experimental and clinical validation is needed to confirm these relationships and explore the underlying biological mechanisms. If validated, galectins could serve as promising biomarkers or therapeutic targets in gynecologic oncology.
Data Sharing Statement
All data generated or analyzed during this study are available from the (https://gwas.mrcieu.ac.uk/). Ovarian Cancer Association Consortium (OCAC), and Publicly available summary statistics from the FinnGen consortium (www.finbb.fi).
Ethical Approval
All data conducted in this study were legally obtained from public dateset, and were analyzed without interfering with public behavior. All data used in this study are anonymous. This study is exempt from approval based on item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects.
Acknowledgments
We would like to thank the genome-wide association study (GWAS) for the offering related data regarding the paper.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
Nanjing Health Science and Technology Development Special Foundation (YKK23226) supported our study.
Disclosure
The authors declared that they have no competing interests.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Johannes L, Jacob R, Leffler H. Galectins at a glance. J Cell Sci. 2018;131(9). doi:10.1242/jcs.208884
3. Girotti MR, Salatino M, Dalotto-Moreno T, et al. Sweetening the hallmarks of cancer: galectins as multifunctional mediators of tumor progression. J Exp Med. 2020;217(2). doi:10.1084/jem.20182041
4. Liu F, Stowell SR. The role of galectins in immunity and infection. Nat Rev Immunol. 2023;23(8):479–494. doi:10.1038/s41577-022-00829-7
5. Kamarudin AA, Abu N. Revisiting galectin-1, −3, −4, and −9 as biotargets for colorectal cancer. Biochem Cell Biol. 2025;103:1–10. doi:10.1139/bcb-2024-0218
6. Griffiths A, Udomjarumanee P, Georgescu A-S, et al. The immunomodulatory role of Galectin-1 in the tumour microenvironment and strategies for therapeutic applications. Cancers. 2025;17(11):1888. doi:10.3390/cancers17111888
7. Nangia-Makker P, Hogan V, Raz A. Galectin-3 and cancer stemness. Glycobiology. 2018;28(4):172–181. doi:10.1093/glycob/cwy001
8. So CY, Li Y, Chow KT. New insights on Galectin-9 expression in cancer prognosis: an updated systemic review and meta-analysis. PLoS One. 2025;20(3):e0320441. doi:10.1371/journal.pone.0320441
9. Masoodi M, Shah ZA, Beigh AH, et al. Galectin-1 as a predictive biomarker in ovarian cancer. J Ovarian Res. 2021;14(1):123. doi:10.1186/s13048-021-00874-1
10. Chen L, Yao Y, Sun L, et al. Galectin-1 promotes tumor progression via NF-kappaB signaling pathway in epithelial ovarian cancer. J Cancer. 2017;8(18):3733–3741. doi:10.7150/jca.20814
11. Mohamed RM, Emam A, Abdelfattah MM, et al. Assessment of galectins −1, −3, −4, −8, and −9 expression in ovarian carcinoma patients with clinical implications. World J Surg Oncol. 2022;20(1):276. doi:10.1186/s12957-022-02738-4
12. Shimada C, Xu R, Al-Alem L, et al. Galectins and ovarian cancer. Cancers. 2020;12(6):1421. doi:10.3390/cancers12061421
13. Chetry M, Song Y, Pan C, et al. Effects of Galectin-1 on biological behavior in cervical cancer. J Cancer. 2020;11(6):1584–1595. doi:10.7150/jca.38538
14. Chauhan R, Malhotra L, Gupta A, et al. Bergenin inhibits growth of human cervical cancer cells by decreasing Galectin-3 and MMP-9 expression. Sci Rep. 2024;14(1):15287. doi:10.1038/s41598-024-64781-3
15. Negedu MN, Duckworth CA, Yu L. Galectin-2 in health and diseases. Int J Mol Sci. 2022;24(1):341. doi:10.3390/ijms24010341
16. Kaur S, Singh J, Kaur M. Multifaceted role of galectin-4 in cancer: a systematic review. Eur J Clin Invest. 2023;53(7):e13987. doi:10.1111/eci.13987
17. Xu D, Guo L, Zhang S, et al. LGALS2 suppresses the progression of papillary thyroid carcinoma by regulating the PI3K/AKT pathway. Gland Surg. 2022;11(9):1518–1528. doi:10.21037/gs-22-452
18. Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014;23(R1):R89–98. doi:10.1093/hmg/ddu328
19. Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362:k601. doi:10.1136/bmj.k601
20. Guo J, Wu Q-J, Liu F-H, et al. Review of Mendelian randomization studies on endometrial cancer. Front Endocrinol. 2022;13:783150. doi:10.3389/fendo.2022.783150
21. Chang X, Liu S, Han L. Mendelian randomization analysis to elucidate the causal relationship between small molecule metabolites and ovarian cancer risk. Front Oncol. 2023;13:1291033. doi:10.3389/fonc.2023.1291033
22. Henry A, Gordillo-Marañón M, Finan C, et al. Therapeutic targets for heart failure identified using proteomics and Mendelian randomization. Circulation. 2022;145(16):1205–1217. doi:10.1161/CIRCULATIONAHA.121.056663
23. Wang Y, Shi X, Yin Y, et al. Association between neuroinflammation and Parkinson’s disease: a comprehensive Mendelian randomization study. Mol Neurobiol. 2024;61(12):10216–10226. doi:10.1007/s12035-024-04197-2
24. Schuermans A, Truong B, Ardissino M, et al. Genetic associations of circulating cardiovascular proteins with gestational hypertension and preeclampsia. JAMA Cardiol. 2024;9(3):209–220. doi:10.1001/jamacardio.2023.4994
25. Sun BB, Maranville JC, Peters JE, et al. Genomic atlas of the human plasma proteome. Nature. 2018;558(7708):73–79. doi:10.1038/s41586-018-0175-2
26. O’Mara TA, Glubb DM, Amant F, et al. Identification of nine new susceptibility loci for endometrial cancer. Nat Commun. 2018;9(1):3166. doi:10.1038/s41467-018-05427-7
27. Phelan CM, Kuchenbaecker KB, Tyrer JP, et al. Identification of 12 new susceptibility loci for different histotypes of epithelial ovarian cancer. Nat Genet. 2017;49(5):680–691. doi:10.1038/ng.3826
28. Burgess S, Thompson SG. Avoiding bias from weak instruments in Mendelian randomization studies. Int J Epidemiol. 2011;40(3):755–764. doi:10.1093/ije/dyr036
29. Bowden J, Del Greco M F, Minelli C, et al. A framework for the investigation of pleiotropy in two-sample summary data Mendelian randomization. Stat Med. 2017;36(11):1783–1802. doi:10.1002/sim.7221
30. Yu X, Qian J, Ding L, Yin S, Zhou L, Zheng S. Galectin-1: a traditionally immunosuppressive protein displays context-dependent capacities. Int J Mol Sci. 2023;24(7):6501.
31. Zhu J, Zheng Y, Zhang H, et al. Galectin-1 induces metastasis and epithelial-mesenchymal transition (EMT) in human ovarian cancer cells via activation of the MAPK JNK/p38 signalling pathway. Am J Transl Res. 2019;11(6):3862–3878.
32. Chetry M, Saroj T, Hu X, et al. The role of galectins in tumor progression, treatment and prognosis of gynecological cancers. J Cancer. 2018;9(24):4742–4755. doi:10.7150/jca.23628
33. Ji P, Gong Y, Jin M-L, et al. In vivo multidimensional CRISPR screens identify Lgals2 as an immunotherapy target in triple-negative breast cancer. Sci Adv. 2022;8(26):eabl8247. doi:10.1126/sciadv.abl8247
34. Chen C, Duckworth CA, Fu B, et al. Circulating galectins −2, −4 and −8 in cancer patients make important contributions to the increased circulation of several cytokines and chemokines that promote angiogenesis and metastasis. Br J Cancer. 2014;110(3):741–752. doi:10.1038/bjc.2013.793
35. Sindrewicz P, Lian L, Yu L. Interaction of the oncofetal thomsen-friedenreich antigen with galectins in cancer progression and metastasis. Front Oncol. 2016;6:79. doi:10.3389/fonc.2016.00079
36. Wu K, Kuo C-M, Huang E-Y, et al. Extracellular galectin-3 facilitates colon cancer cell migration and is related to the epidermal growth factor receptor. Am J Transl Res. 2018;10(8):2402–2412.
37. Fang S, Feng Y, Li M. Correlations of Galectin-3 gene polymorphisms with risk and prognosis of cervical cancer in chinese populations: a case-control study. Oncol Res Treat. 2017;40(9):533–539. doi:10.1159/000475502
38. Kumar R, Mandal S, Arora P, et al. The expression of p16 and galectin-3 in cervical intraepithelial neoplasia (CIN) and squamous cell carcinoma (SCC) uterine cervix. J Obstet Gynaecol. 2021;41(5):785–790. doi:10.1080/01443615.2020.1803235
39. Ma J, Zhang X, He G, et al. The relationship between cervical precancerous lesion galectin-3 and p27 protein expression and clinical prognosis. Oncol Lett. 2018;15(2):1533–1536. doi:10.3892/ol.2017.7503
40. Troncoso MF, Elola MT, Blidner AG, et al. The universe of galectin-binding partners and their functions in health and disease. J Biol Chem. 2023;299(12):105400. doi:10.1016/j.jbc.2023.105400
41. Wang Y, Sun Y, Li H, et al. Galectin-8 alters immune microenvironment and promotes tumor progression. Am J Cancer Res. 2023;13(6):2517–2529.
42. Beyer S, Wehrmann M, Meister S, et al. Galectin-8 and −9 as prognostic factors for cervical cancer. Arch Gynecol Obstet. 2022;306(4):1211–1220. doi:10.1007/s00404-022-06449-9
43. Oyanadel C, Holmes C, Pardo E, et al. Galectin-8 induces partial epithelial-mesenchymal transition with invasive tumorigenic capabilities involving a FAK/EGFR/proteasome pathway in Madin-Darby canine kidney cells. Mol Biol Cell. 2018;29(5):557–574. doi:10.1091/mbc.E16-05-0301
44. Cao Z, Guo X. The role of galectin-4 in physiology and diseases. Protein Cell. 2016;7(5):314–324. doi:10.1007/s13238-016-0262-9
45. Menkhorst E, Griffiths M, Van Sinderen M, et al. Galectin-7 is elevated in endometrioid (type I) endometrial cancer and promotes cell migration. Oncol Lett. 2018;16(4):4721–4728. doi:10.3892/ol.2018.9193
46. Liu C, Zhou L, Chen J, et al. Galectin-7 promotes cisplatin efficacy by facilitating apoptosis and G3BP1 degradation in cervical cancer. Biochem Pharmacol. 2023;217:115834. doi:10.1016/j.bcp.2023.115834
47. Schulz H, Kuhn C, Hofmann S, et al. Overall survival of ovarian cancer patients is determined by expression of Galectins-8 and −9. Int J Mol Sci. 2018;19(1):323. doi:10.3390/ijms19010323
48. Imai Y, Hasegawa K, Matsushita H, et al. Expression of multiple immune checkpoint molecules on T cells in malignant ascites from epithelial ovarian carcinoma. Oncol Lett. 2018;15(5):6457–6468. doi:10.3892/ol.2018.8101
49. Tomizawa H, Yamada Y, Arima M, et al. Galectin-10 as a potential biomarker for eosinophilic diseases. Biomolecules. 2022;12(10):1385. doi:10.3390/biom12101385
50. Yoshimura H, Takeda Y, Shirai Y, et al. Galectin-10 in serum extracellular vesicles reflects asthma pathophysiology. J Allergy Clin Immunol. 2024;153(5):1268–1281. doi:10.1016/j.jaci.2023.12.030
51. Liu D, Zhu H, Li C. Galectins and galectin-mediated autophagy regulation: new insights into targeted cancer therapy. Biomark Res. 2023;11(1):22. doi:10.1186/s40364-023-00466-9
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.