Back to Journals » Infection and Drug Resistance » Volume 19
Exploring Novel Gene Mutations Associated with Fluoroquinolone Resistance in Mycobacterium tuberculosis: Gene Sequencing Results from 275 Clinical Isolates in Chongqing, China
Authors Zheng X, Zhou T, Zeng R, Lu Y, Yang S, Ouyang J, Xu J, Yu J, Deng J, Chen Y
Received 22 December 2025
Accepted for publication 30 March 2026
Published 9 April 2026 Volume 2026:19 590990
DOI https://doi.org/10.2147/IDR.S590990
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Sandip Patil
Xiaoman Zheng,1,* Tingting Zhou,1,* Rong Zeng,2 Yanqiu Lu,1 Sen Yang,1 Jing Ouyang,1 Jintian Xu,2 Jifang Yu,2 Jiaoyu Deng,2 Yaokai Chen1
1Clinical Research Center, Chongqing Public Health Medical Center, Chongqing, People’s Republic of China; 2Key Laboratory of Special Pathogens and Biosafety, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yaokai Chen, Email [email protected] Jiaoyu Deng, Email [email protected]
Purpose: This study aimed to investigate whether mutations in the rv2005c, PPE59, and recC genes are associated with fluoroquinolone (FQ) resistance in Mycobacterium tuberculosis (Mtb), particularly in strains lacking mutations in the known resistance genes gyrA and gyrB.
Methods: A total of 275 clinical isolates, consisting of 217 pre-extensively drug-resistant strains and 58 fluoroquinolone-sensitive strains, were included in this study. Gene sequencing was performed on all isolates, and functional assessments through both overexpression and knockout experiments of the gene were conducted to evaluate its effect on fluoroquinolone susceptibility.
Results: Results: Among 275 isolates, no mutations were detected in PPE59 or recC. In the 217 pre-XDR isolates, gyrA mutations were found in 214 (98.6%), predominantly at codons 94, 90, and 91. Only two isolates harbored synonymous mutations in rv2005c (P235P), both with concurrent gyrA D94G mutations. Overexpression or knockout of rv2005c in Mtb did not alter FQ susceptibility. Notably, two FQ-resistant isolates lacked mutations in any of the five genes sequenced, suggesting the presence of unknown resistance mechanisms.
Conclusion: Our results showed that mutations of the rv2005c, PPE59 and recC genes may not contribute to FQ resistance in clinical isolates of Mtb in Chongqing, China. Furthermore, our study results also showcased the complexity of mechanisms of FQ resistance in clinical Mtb isolates.
Keywords: Mycobacterium tuberculosis, fluoroquinolone resistance, drug susceptibility, gene mutations, mechanism
Introduction
Mycobacterium tuberculosis (Mtb) is the single infectious agent responsible for tuberculosis (TB), which is a chronic infectious disease, and is the leading cause of death in humans from a single source of infection.1 According to estimates made by the World Health Organization (WHO), the scale of the latently Mtb-infected population worldwide is approximately two billion, with an incident rate of 127/100,000 globally. New TB cases identified globally numbered 9.87 million in 2020.1 Meanwhile, the estimated number of new TB cases in China in 2020 was 842,000, with an incident rate of 59/100,000.1 The emergence and evolution of drug-resistant tuberculosis (DR-TB) in contemporary times often complicates TB treatment and control.2 The burden of DR-TB remains substantial worldwide. According to the World Health Organization Global Tuberculosis Report 2024, the estimated number of multidrug-resistant or rifampicin-resistant tuberculosis (MDR/RR-TB) cases globally was 400,000 in 2023, with a treatment success rate of only 68%.3 In China, a recent national drug-resistant tuberculosis survey among adolescents and young adults reported that among new pulmonary tuberculosis cases, the proportion of multi-drug resistant (MDR)-TB was 3.18% (95% CI: 2.37–4.15) for adolescents and 3.76% (95% CI: 3.03–4.60) for young adults; among previously treated patients, these proportions increased to 11.25% (95% CI: 5.28–20.28) and 11.05% (95% CI: 6.88–16.55), respectively4. Of particular concern is the rising resistance to fluoroquinolones, which are core components of MDR-TB treatment regimens. A large-scale global genomic analysis of 7,890 M. tuberculosis isolates from 77 countries revealed substantial non-susceptibility to second-line agents, particularly fluoroquinolones such as ciprofloxacin and ofloxacin.5 These findings underscore the urgent need to continuously monitor fluoroquinolone (FQ) resistance and investigate its underlying mechanisms across different geographical settings.
In 1996, the WHO included the FQ class of drugs in the anti-TB treatment regimen for the first time, and since then FQ drugs have gradually become the core drugs for MDR-TB treatment.6 In 2016, the WHO listed FQ drugs as the first-choice drug group for MDR-TB treatment.3 With extensive global utilization of FQs, Mtb resistance to FQ drugs has inevitably been steadily increasing. The principal mechanism of FQ resistance in Mtb is mutations in the quinolone resistance-determining region (QRDR) of the gyrA and gyrB genes. As a consequence of these mutations, FQ drugs are unable to act on the DNA helicase of Mtb.7 Mutations in the QRDR of the gyrA gene account for 60% to 90% of the FQ resistance phenotype of clinically isolated pre-extensive drug resistant (Pre-XDR) Mtb strains.8 This implies that approximately 10% to 40% of phenotypically resistant isolates lack mutations in the QRDR of gyrA or gyrB, suggesting the involvement of alternative mechanisms such as efflux pumps or other novel genetic determinants. Despite some clinical isolates being resistant to FQs, they do not exhibit mutations in the gyrA and gyrB genes, indicating that other cryptic mechanisms of resistance exist, and remain to be unidentified.
In a recent study, Chaiyachat et al, used whole genome sequencing analysis in an attempt to identify novel gene mutations associated with FQ resistance, and observed that the rv2005c, PPE59, and recC genes statistically correlate with FQ drug resistance.9 Notably, the rv2005c gene, which encodes a universal stress protein (USP), has previously been implicated in resistance to aminoglycosides, raising the possibility that it may play a broader role in drug resistance, including against FQs.10 However, the mutational types and frequencies of drug resistance genes to FQs in Mtb vary significantly under the influence of different environmental factors.11,12 Whether the rv2005c, PPE59, and recC genes contribute FQ drug resistance in clinical isolates of Mtb in countries with a high burden of TB, such as China, remains unknown. In the present study, we sequenced the gene mutations of the QRDR and the rv2005c, PPE59, and recC gene loci in 217 Pre-XDR strains of Mtb and 58 fluoroquinolone-sensitive strains of Mtb, and we also investigated the association between the identified gene mutations (excluding the gyrA and gyrB genes) and phenotypic FQ resistance by performing gene over-expression and gene knock out in Mtb. The specific research question addressed in this study was whether mutations in the rv2005c, PPE59, or recC genes are associated with phenotypic fluoroquinolone resistance in clinical Mtb isolates from Chongqing, China.
Materials and Methods
Clinical Mtb Isolates
All clinical isolates of Mtb used in this study were collected at Chongqing Public Health Medical Center between June 2020 and October 2022. A total of 217 pre-extensively drug-resistant (Pre-XDR) and 58 FQ-sensitive isolates were selected for further study. Mycobacterial drug susceptibility testing (DST) was performed using the proportion method on Lowenstein-Jensen (LJ) solid medium in accordance with the WHO 2021 guidelines for the diagnosis and treatment of drug-resistant tuberculosis. MDR-TB was defined as strains resistant to at least rifampicin (RIF) and isoniazid (INH). Pre-XDR-TB was defined as MDR-TB strains additionally resistant to any one of the FQ drugs (including moxifloxacin [MOX], levofloxacin [LVX], and ofloxacin [OFX]) but susceptible to bedaquiline and linezolid. FQ-sensitive isolates were defined as strains susceptible to all tested FQ drugs (MOX, LVX, OFX) with no resistance to first-line anti-TB drugs.
All the 275 clinical Mtb isolates included in this study were obtained from distinct individual patients, and no multiple samples were collected from the same patient, ensuring the independence of all experimental samples.
Bacterial Strains, Plasmids, and Culture Conditions
Mtb H37Rv or H37Ra was cultured in Middlebrook 7H9 broth (Difco, Becton, Dickinson and company, Sparks, MD, United States) supplemented with 10% oleic acid-albumin-dextrose-catalase (OADC, Difco), 0.5% glycerol, and 0.05% Tween 80 (Sigma) at 37°C, or incubated on 7H10 agar medium (Difco) supplemented with 10% OADC and 0.5% glycerol at 37°C. Clinical isolates were cultured on Lowenstein–Jensen (LJ) medium (Zhuhai YinKe Medical Engineering Co., China), prepared according to the standard formulation containing defined amounts of mineral salts, asparagine, glycerol, homogenized egg, and malachite green. Mycobacterium smegmatis mc2 155 was grown in Middlebrook 7H9 broth supplemented with 0.5% glycerol, and 0.05% Tween 80 (Sigma) at 37°C, with shaking (100 rpm) or on 7H10 agar medium (Difco) supplemented with 0.5% glycerol at 37°C. Escherichia coli strains DH5α and HB101 were cultured in LB medium at 37°C, with shaking at 220 rpm overnight or on LB agar medium at 37°C for 24h. Plasmid pMV261 and the E.coli BL21 strain, which is specifically designed for the expression of plasmid pMV261, were generously provided by the TB research group at the Wuhan Institute of Virology, Chinese Academy of Sciences.
Mycobacterial Genomic DNA
DNA isolation was conducted using the HiPure Mycobacterial DNA Kit (Magen, Guangzhou, China). One milliliter of bacterial liquid culture of each strain was extracted and transferred to a 1.5 mL Eppendorf tube. Bacterial cells were collected by centrifugation (10000 xg, 10 min), washed twice with sterile water, and re-suspended in 200µL of buffer ATL containing 10µg/mL lysozyme and 10µg/mL proteinase K. Subsequently, bacterial cells were heated (56°C for 30 min, 95°C for 30 min), and then centrifuged at 10000 xg for 3 min. The supernatant was transferred to a new 1.5 mL Eppendorf tube, and 200µL buffer GXP solution and 200µL anhydrous ethanol were added and mixed evenly. After centrifugation (10,000 xg, 1 min), genomic DNA was collected on a silica membrane, washed 3 times with the buffer GW2 solution, then dissolved in 40µL of sterile water, and subsequently stored at −20°C until use.
Gene Amplification and Sequencing Analysis
All primers used in the study are detailed in Table 1. The gyrA QRDR region was amplified by using the primer pairs gyrA-F and gyrA-R (Table 1). The gyrB QRDR region was amplified by PCR using gyrB-F and gyrB-R (Table 1). The rv2005c region was amplified by PCR using the primer pairs rv2005c-F1 and rv2005c-R1. The PPE59 region was amplified by PCR using the primer pairs PPE59-F and PPE59-R. The recC region was amplified by PCR using the primer pairs recC-F and recC-R. The 30-μL amplification reaction mixture containing 2μL genomic DNA, 15μL 2× AceTaq Master Mix, 2.4μL 10μM PCR primer mixture and 10.6μL distilled water. The PCR parameters included one denaturation cycle (95°C, 5 min) followed by 35 cycles of denaturation (95°C, 15 sec), annealing (56°C for gyrA, 54°C for gyrB, 65°C for rv2005c, 67°C for PPE59, and 67°C for recC, 45 sec), primer extension (72°C, 45 sec), and a final extension at 72°C for 10 min.
 |
Table 1 Primers Used in This Study |
Successful PCR products were sent to the Tsingke Biotechnology Company (Chongqing, China) for sequencing. Mutations in the gyrA QRDR region, the gyrB QRDR region, rv2005c, PPE59, and recC were analyzed using SnapGene software through comparison with the corresponding gene sequence of H37Rv, and also with clinically susceptible strains isolated from Chongqing Public Health Medical Center.
Statistical Analysis
Associations between gene mutations and fluoroquinolone resistance were assessed by calculating odds ratios (OR) with 95% confidence intervals (CI) using 2×2 contingency tables. Due to the low frequency of mutations in some genes, Fisher’s exact test was performed to obtain exact CIs. An OR with a 95% CI that did not cross 1.0 was considered indicative of a potential association. All statistical analyses were conducted using SPSS version 26.0 (IBM, Armonk, NY, USA). The two phenotypically resistant isolates lacking mutations in all sequenced genes were excluded from the association analysis, as they lacked candidate genetic determinants and are the subject of a separate ongoing investigation.
Construction of the rv2005c Gene Knockout Strains in H37Ra and H37Rv
Because the rv2005c gene on the genome of the knockout strain (Δrv2005c) was replaced by a hygromycin box of approximately 3.7kb, the PCR fragment of the Δrv2005c strain (5.2kb) was larger than that of the wild strain (2.3kb). Additionally, hygromycin resistance acquired by the genome was also one of the screening markers. A colony that is able to grow on a 7H10 plate containing hygromycin after four weeks of culture is considered a positive monoclonal for the rv2005c gene knockout. The positive monoclonals from the plate were then inoculated into 7H9 medium with an identical hygromycin concentration.
Construction of H37Ra pMV261::rv2005c and H37Rv pMV261::rv2005c
Genomic DNA of H37Rv was extracted, and rv2005c was amplified with primers (Table 1, rv2005c-F2, rv2005c-R2) containing restriction sites EcoRI and HindIII. The fragment was ligated to the vector plasmid pMV261, cloned onto E.coli cells, and identified by double enzyme digestion. After the application of rv2005c overexpression was successfully identified by double enzyme digestion, an appropriate amount of H37Ra pMV261::rv2005c, H37Rv pMV261::rv2005c and fresh bacterial liquid of the logarithmic growth stage were extracted, and the recombinant plasmid was sent for sequencing analysis and verification.
Drug Susceptibility Tests
Mycobacterial cells were grown in 7H9 liquid culture (with 10% OADC) to an optical density at 600 nm (OD600) of 0.5–1.0. The culture was then serially diluted 10-fold in fresh 7H9 medium with 10% OADC. The 10−3 or 10−4 dilution, corresponding to approximately 105 CFU/mL, was used as the inoculum. Aliquots of 10 μL of the diluted suspension were plated onto 7H10 agar plates containing various concentrations of the following antimicrobial drugs, ie., streptomycin (Sm) (0, 0.125, 0.25, 0.5, 1, 2, 4, and 8 μg/mL), RIF (0, 0.01, 0.02, 0.04, 0.08, 0.16, 0.32, and 0.64 μg/mL), MOX (0, 0.025, 0.05, 0.1, 0.2, 0.4, 0.8, 1.6, 3.2, and 6.4 μg/mL), ofloxacin (OFX) (0, 0.025, 0.05, 0.1, 0.2, 0.4, 0.8, and 1.6 μg/mL), and LVX (0, 0.025, 0.05, 0.1, 0.2, 0.4, 0.8 1.6, 3.2, and 6.4 μg/mL). All antibiotics were purchased from Sigma-Aldrich and were solubilized according to the manufacturer’s recommendations. Cultures were incubated at 37°C for 21 days. The MIC was defined as the lowest required concentration of antibiotics to inhibit the growth of 99% of bacterial CFUs.
Antibiotic Killing Assays
Mycobacterial cells were grown to mid-log phase (OD600 0.5 to 1.0), and diluted to the cell density of OD600=0.1 with fresh 7H9 medium. The antibiotics were added to 10 mL diluted bacterial solution, and incubated at 37°C. Then, aliquots of the samples were taken and plated on 7H10 medium without any drug after serial dilutions at time intervals of 0, 3, 7, 14, and 21 days. Each set of experiments contained three sample replicates, and was repeated three times. Graphs for antibiotic kill curves were prepared using Graphpad Prism 9.0, and mean values with standard deviations (SD) were plotted against time.
Results
Sample Characteristics
Characteristics of Mtb clinical strains used in the study are shown in Table 2. The majority (73.82%) of the strains were isolated from sputum samples. Meanwhile, 50.91% of the strains were isolated from patients in the 41–65 year age group, demonstrating that TB is prevalent in the middle-aged population. The proportion of strains isolated from male patients (66.91%) was approximately double that from female patients (33.09%).
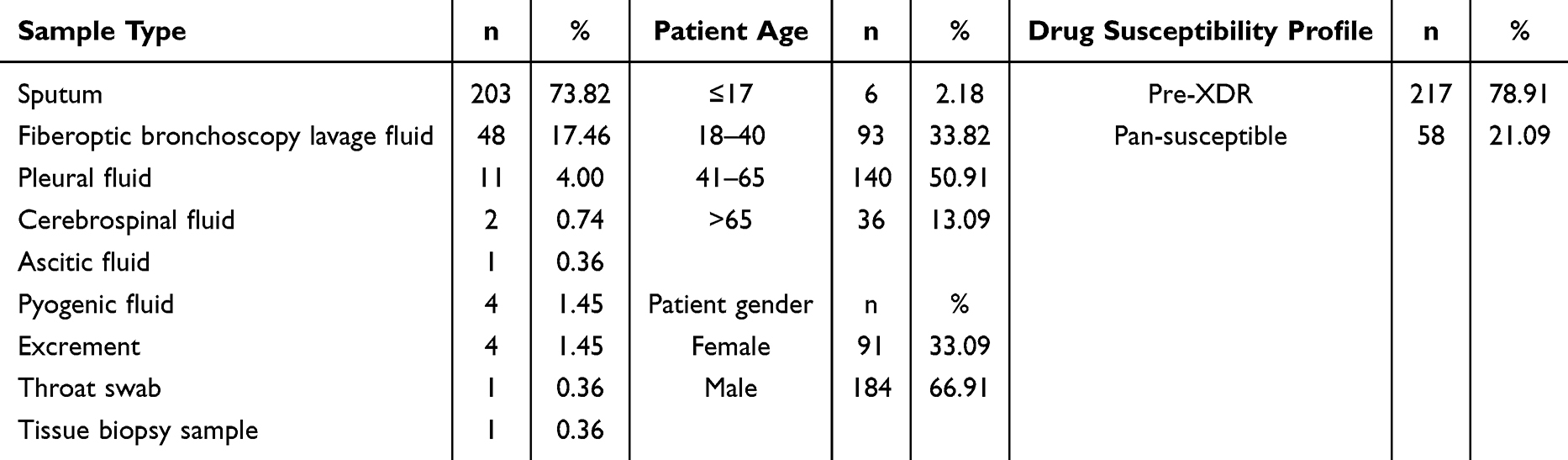 |
Table 2 Stratification of the 275 Isolates Used in This Study |
Successful Construction of the rv2005c Gene Knockout Strains in H37Ra and H37Rv
The Δrv2005c strain DNA was extracted via the CTAB method, and the genomic DNA sequence was validated by PCR using the rv2005c-LF and rv2005c-RR primers (Table 1). Our results demonstrated that the rv2005c gene knockout strains were successfully constructed in both H37Ra and H37Rv (Figure 1).
 |
Figure 1 PCR verification of the knock-out of the rv2005c gene. (A) Schematic map of the rv2005c gene knockout strategy. (B) Agarose gel electrophoresis of PCR products for rv2005c gene knockout verification. Lane 1, DNA molecular weight marker; Lane 2, wild-type (WT) M. tuberculosis strain (2.3 kb PCR product); Lane 3, H37Ra rv2005c gene knockout strains (H37RaΔrv2005c). Lane 4 and Lane 5, H37Rv rv2005c gene knockout strains (H37RvΔrv2005c). bp, base pair. |
Mutations in gyrA, gyrB, rv2005c, PPE59, and recC
The SNPs in the gyrA, gyrB, rv2005c, PPE59, and recC genes of the clinically isolated sensitive strains were first sorted out, and results indicated that a double mutation (G61C and G2003A) of gyrA was present (Table 3). Thus, this double mutation was classified as a background mutation and was not included in the subsequent SNP analysis of Pre-XDR strains. Then, SNPs in the five genes of 217 Pre-XDR clinical isolates were collated. As shown in Table 4, mutations in the gyrA QRDR had the highest frequency (97.69%, 212/217). A single mutation was identified in the gyrA QRDR of 174 isolates (174/217, 80.18%), a double mutation was identified in the gyrA QRDR of 13 isolates (13/217, 5.99%), and a triple mutation was identified in the gyrA QRDR of one isolate (1/217, 0.42%). Twenty two isolates had mutations in both the gyrA and the gyrB genes (22/217, 10.14%). Two isolates harbored mutations in both gyrA and rv2005c (2/217, 0.92%). Additionally, mutations in three strains occurred in the non-QRDR of gyrA and gyrB (3/217, 1.38%). No mutations in the PPE59 and recC genes were observed. Two of the FQ-resistant isolates were observed to have no mutations in any of the gyrA QRDR, gyrB QRDR, rv2005c, PPE59, or recC genes.
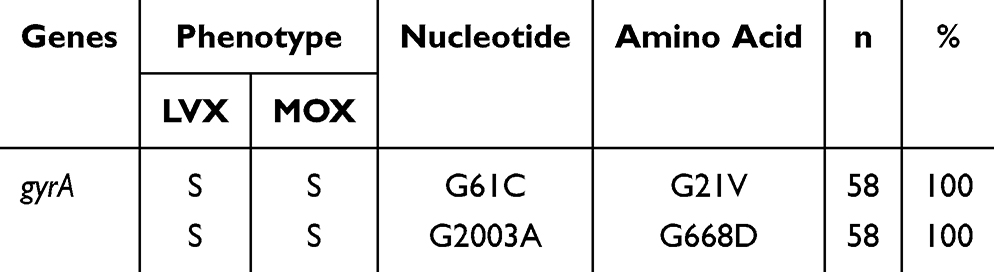 |
Table 3 Statistics on Target Gene SNPs and Drug Resistance of 58 Clinically Isolated Sensitive Strains |
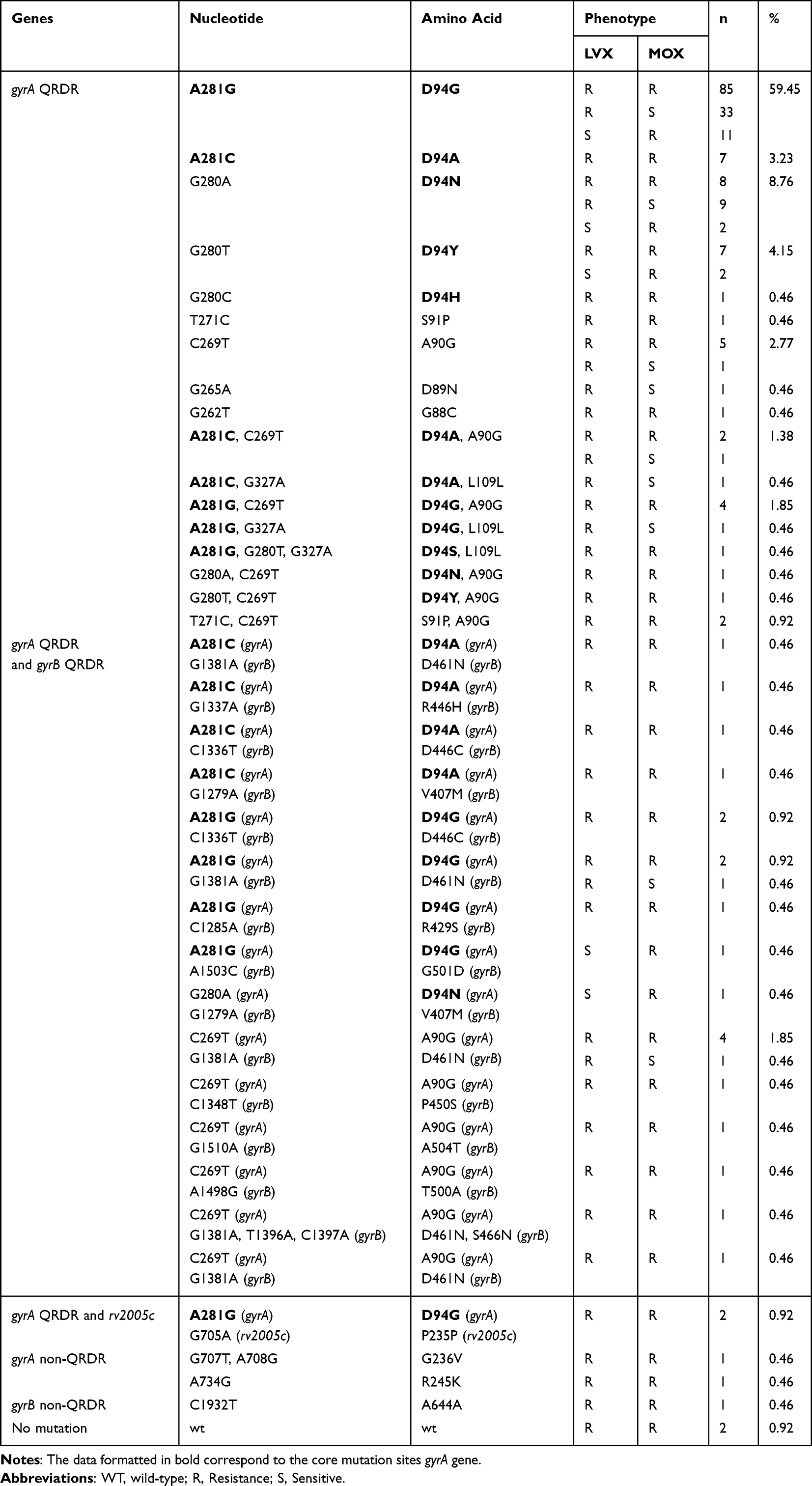 |
Table 4 Clinical Isolation of Pre-XDR Strain Target Gene SNP and Drug Resistance Statistics |
Association Between Gene Mutations and FQ Resistance
To quantify the strength of association between individual gene mutations and fluoroquinolone resistance, odds ratios (OR) with 95% confidence intervals (CI) were calculated (Table 5). Mutations in the gyrA QRDR were strongly associated with FQ resistance (OR = 45.3; 95% CI: 12.7–161.2). In contrast, mutations in gyrB QRDR showed no significant association (OR = 1.8; 95% CI: 0.6–5.4), as the confidence interval crossed 1.0. For rv2005c, the OR was 1.0 (95% CI: 0.1–9.8), indicating no association with FQ resistance. No mutations were detected in PPE59 or recC in any of the FQ-resistant isolates, precluding association analysis for these genes. The two isolates without any identified mutations were excluded from this analysis as they lacked candidate genetic determinants.
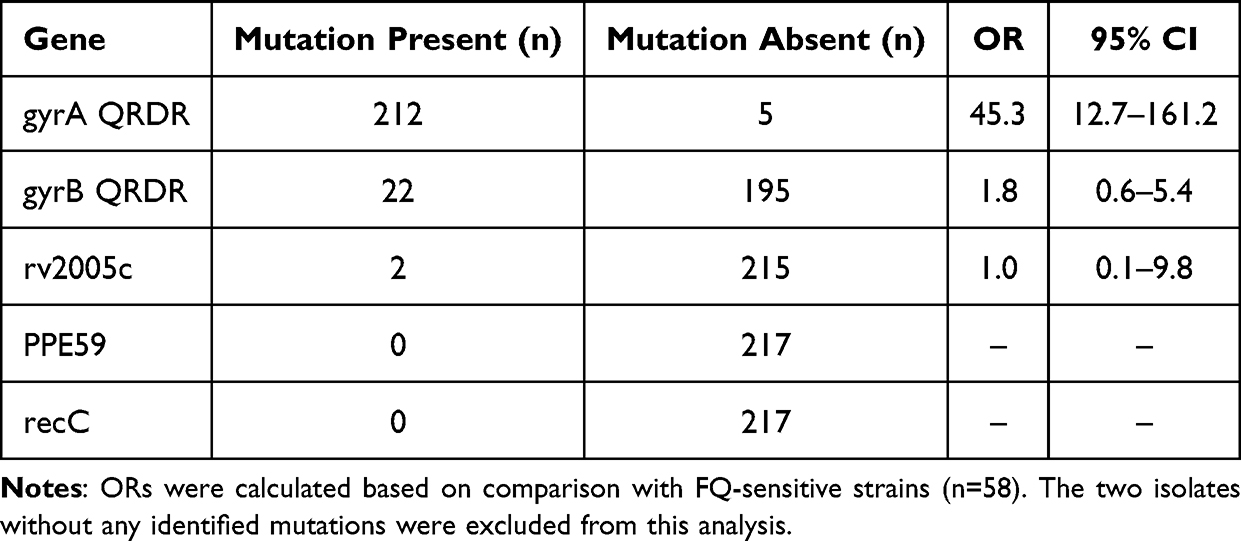 |
Table 5 Association Between Gene Mutations and Fluoroquinolone Resistance in 217 Pre-XDR Clinical Isolates |
The Effect of the rv2005c Gene on Susceptibility of Mtb to FQs
The effect of the rv2005c gene on susceptibility of Mtb to different FQs (MOX, LVX, and OFX) was tested, with utilization of RIF and Sm as positive controls. As shown in Table 6, no differences were observed in susceptibility to all drugs among all strains tested.
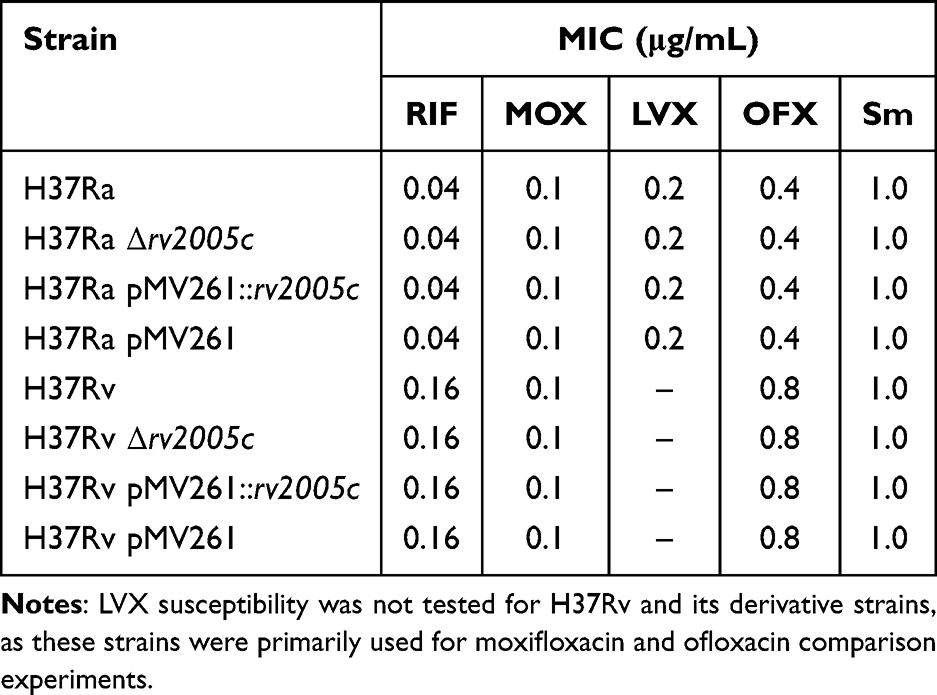 |
Table 6 MICs of Four Drugs Against Different Mycobacterial Strains |
Effect of the rv2005c Gene on the Bactericidal Effect of FQs
To further probe the impact of rv2005c on the anti-tuberculosis efficacy of FQs, killing curves of FQs were compared among different Mtb strains by using RIF and Sm as control drugs. As shown in Figure 2, deletion or over-expression of rv2005c was observed to not influence the bactericidal effect of any of the drugs tested.
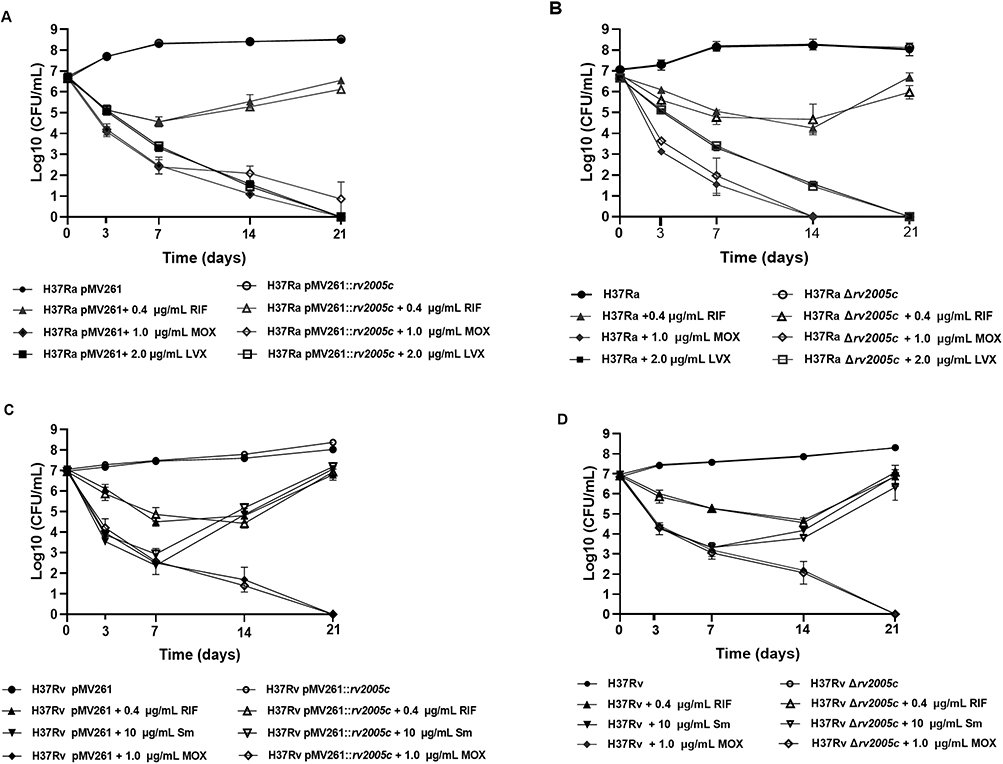 |
Figure 2 Effect of rv2005c on the bactericidal effect of FQs. (A) Bactericidal curves of H37Ra pMV261 strains and rv2005c-overexpressing H37Ra strains treated with RFP, MOX and LVX. (B) Bactericidal curves of H37Ra strains and rv2005c-knockout H37Ra strains treated with RFP, MOX and LVX. (C) Bactericidal curves of H37Rv pMV261 strains and rv2005c-overexpressing H37Rv strains treated with RFP, Sm and MOX. (D) Bactericidal curves of H37Rv strains and rv2005c-knockout H37Rv strains treated with RFP, Sm and Mox. |
Discussion
China has a significant burden of Mtb-related disease. According to the WHO Global Tuberculosis Report for 2023, the estimated number of new TB cases in China was 748,000, which accounts for 7.1% of the total estimated new cases worldwide (approximately 10.06 million), and placing China as the third-largest contributor to new TB cases globally. The report indicated that 65% of these cases are in males aged 15 and older, 30% are in females aged 15 and older, and 5% are in children under 15 years old.13 An analysis of the age-specific reported incidence of tuberculosis from China’s Tuberculosis Information Management System, covering the period from 2008 to 2018, revealed that the reported rate of tuberculosis in males (84.4%) was more than double that in females (38.6%).14 This data is consistent with clinical information regarding Pre-XDR strains collected in the present study. In our study, we analyzed 217 clinical isolates of Pre-XDR TB collected in Chongqing, China, from 2020 to 2022, along with the basic demographic characteristics of the affected population. Our results indicated the presence of a significant number of Pre-XDR TB patients among middle-aged individuals, with 2.81% of Pre-XDR TB patients being teenagers, and the youngest of whom was only seven years old. The fraction of male patients was approximately twice that of female patients, indicating a greater risk of disease among males. Furthermore, the drug resistance rates of MOX and LVX in Pre-XDR clinical strains were observed to be elevated, with the resistance rate for MOX being higher than that of LVX. These observations suggest that future TB prevention and control efforts should prioritize the screening and treatment of FQ drug resistance in middle-aged TB patients. Additionally, vigilance is necessary with respect to the risk of Pre-XDR TB transmission among children, and enhanced screening and treatment of Pre-XDR TB in male TB patients should also be emphasized.
FQs act primarily on DNA helicase in the Mtb organism, which is a type II topoisomerase composed of 4 subunits. The gyrA and gyrB genes encode two A subunits and two B subunits, respectively.15 The gyrA gene of Mtb has a total length of 2516 bp and encodes 838 amino acids. Most scholars believe that the QRDR of gyrA ranges from 74 to 113 codons, and mutations in this region are the most important mechanism that confers resistance to FQ drugs in Mtb. Amino acid mutations in this region at positions 88, 90, 91, and 94 are the most common mutation sites for gene mutations related to FQ drug resistance.16–18 The gyrB gene, located upstream of the gyrA gene, is 2027 bp in length and encodes 675 amino acids. The existing literature indicates that the QRDR of gyrB spans codons 461—499 and 500—538.8,16 The SNP sites and drug resistance of 217 Pre-XDR clinical isolates indicate that gyrA mutation frequency was highest in our samples from Chongqing, and this was primarily associated with FQ drug resistance. It is known that the SNP sites in gyrA are concentrated, and that the SNP types at a particular site are rich and diverse. With identical mutations at a particular site, different strains have different degrees of drug resistance to MOX and LVX. In the collected clinical strains, 22 strains were observed to have mutations in the gyrB QRDR, and these strains also had concurrent mutations in the gyrA QRDR. No strains with a solitary mutation in the gyrB QRDR were identified in this study. From the results of the present study, it can be seen that current Mtb FQ resistance in Chongqing is primarily caused by the gyrA QRDR mutation. When determining whether an Mtb isolate from a particular TB patient is resistant to FQ, it remains a priority to test whether the Mtb gyrA QRDR mutation is present in Mtb organisms isolated from that patient.
In one study using whole genome sequencing analysis to search for new mutated Mtb genes associated with FQ drug resistance, three new genes, ie., rv2005c, PPE59 and recC were identified.19 The total length of the rv2005c gene is 887bp, and the encoded rv2005c protein is a member of the Mtb Universal stress proteins (USPs) family.10 In the clinical isolates collected in the present study, only two strains were observed to have the synonymous mutation of rv2005c (P235P), and the gyrA D94G mutation was also observed in these two strains. The results of subsequent drug sensitivity studies and bactericidal curve tests indicated that there were no significant changes in the MIC and bactericidal curves of Mtb to MOX and LVX, including when the rv2005c gene was knocked out or overexpressed. Based on these observations, it can be inferred that the rv2005c gene does not play a role in Mtb resistance to MOX and LVX. Additionally, the absence of SNP sites in the PPE59 and recC genes of clinical isolates were observed in our study. The results observed in our study do not concur with observations reported previously in the literature, indicating that further investigations are warranted to demonstrate whether the rv2005c,PPE59, and recC genes are, indeed, associated with the Mtb FQ resistance mechanism.
In our collected Pre-XDR strains, two strains did not have nucleotide site mutations in the gyrA QRDR, gyrB QRDR, rv2005c, PPE59, and recC genes; however, these two strains were observed to be resistant to MOX and LVX, suggesting that a novel FQ resistance mechanism is likely to exist. In the future, our research team hopes to explore the mechanisms of FQ resistance in the two strains via whole genome sequencing, proteomics, and other investigational methods.
Nonetheless, these results should be interpreted with caution, and a number of limitations to this study should be borne in mind. Firstly, it is known that there are significant differences in TB drug resistance among different regions within China.20 The samples used in this study were all taken from patients with TB in Chongqing, China; thus, our study samples were limited by the regional TB resistance characteristics inherent to TB cases in Chongqing, China. Future studies should expand sample sources to include TB cases from different regions of China, and from different countries. Secondly, although we isolated some strains that did not undergo mutation but still showed resistance, this outcome warrants further study and validation as our sample size was relatively small. Therefore, future follow-up studies warrant larger sample sizes in order to more accurately explore novel fluoroquinolone resistance mechanisms.
Our findings align with studies from other high-burden TB settings while revealing geographic heterogeneity. Within China, surveillance in Shanghai identified acquired resistance as the dominant mechanism, with pre-diagnostic FQ exposure strongly associated with resistance (aOR 4.31 for single prescription),21 while a study in Heyuan reported FQ resistance rates of 3.86% among drug-resistant TB patients.22 In Brazil, 92.7% of FQ-resistant MDR isolates harbored gyrA mutations,23 closely mirroring our finding of 98.6% gyrA mutation prevalence. However, the absence of rv2005c, PPE59, and recC mutations in our Chongqing cohort contrasts with a Thai report identifying these as novel resistance-associated genes,19 suggesting such alternative mechanisms may be geographically restricted.6 Clinically, the high prevalence of gyrA mutations validates the continued use of rapid molecular assays targeting the gyrA QRDR for FQ resistance detection.24 However, the two FQ-resistant isolates (0.92%) without mutations in any tested genes underscore a critical diagnostic gap: exclusive reliance on genotypic methods may miss a small but clinically significant proportion of resistant strains.25 Therefore, phenotypic drug susceptibility testing should remain essential, particularly in patients with unexplained treatment failure despite negative molecular results.
Conclusion
In conclusion, our findings demonstrate that mutations in the gyrA QRDR are the primary mechanism underlying fluoroquinolone resistance in clinical Mtb isolates from Chongqing, China. Functional analysis confirmed that the rv2005c gene does not contribute to FQ resistance in Chongqing clinical isolates. Notably, two FQ-resistant strains (0.92%, 2/217) harbored no mutations in any of the tested genes (gyrA, gyrB, rv2005c, PPE59, recC), suggesting the existence of alternative, as-yet-unidentified resistance mechanisms. Further investigations, including whole-genome sequencing and proteomic approaches, are warranted to elucidate these novel resistance pathways.
Data Sharing Statement
The gene sequence data supporting the findings of this study have been deposited in the NCBI database (https://www.ncbi.nlm.nih.gov): gyrA (Gene ID: 887105), gyrB (Gene ID: 887081), rv2005c (Gene ID: 888831), PPE59 (Gene ID: 887630), and recC (Gene ID: 888008).
Ethics Statement
This study utilized archived strains isolated from clinical samples; no human subjects or identifiable human data were included. The Institutional Ethics Committee of Chongqing Public Health Medical Center approved this study (Permit Number: 2024-088-01-KY), and an exemption from written informed consent was obtained. We confirm that all methods were conducted in compliance with the relevant ethical guidelines and regulations.
Acknowledgments
We would like to thank all the laboratory staff of the study for their cooperation in giving us access to the records to extract the data.
Author Contributions
All authors made a significant contribution to the work reported, whether in the conception, study design, execution, acquisition of data, analysis, and interpretation, or all these areas, took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Chongqing Medical Scientific Research Project (Joint project of Chongqing Health Commission and Science and Technology Bureau, No. 2025QNXM052; 2024MSXM120), the Medical Research Project of Chongqing Science & Technology Bureau, China (No. CSTB2023TIAD-KPX0063-3), the Open Project of the Chongqing Key Laboratory of Highly Pathogenic Microbes (No. 2025ZDSYSZD001), and the Prevention and Control of Emerging and Major Infectious Diseases–National Science and Technology Major Project (No. 2025ZD01904904; 2025ZD01905202).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. World Health Organization (WHO). Global Tuberculosis Report 2021. Geneva: World Health Organization; 2021.
2. Lange C, Dheda K, Chesov D, Mandalakas AM, Udwadia Z, Horsburgh CR. Management of drug-resistant tuberculosis. Lancet. 2019;394(10202):953–12. doi:10.1016/S0140-6736(19)31882-3
3. WHO Guidelines Approved by the Guidelines Review Committee. WHO Treatment Guidelines for Drug-Resistant Tuberculosis, 2016 Update. Geneva:World Health Organization; 2016.
4. Wang S, Ou X, Zhou Y, et al. Drug-resistant pulmonary tuberculosis among adolescents and young adults in China. Biomed Environ Sci. 2026;39(2):131–145. doi:10.3967/bes2025.159
5. Alhusseini LB, Hassan TH, Jaafar FN, Kouhsari E, Sholeh M. Global whole-genome–based genomic insights into Mycobacterium tuberculosis: clonal dominance, sequence-type structure, and antimicrobial resistance-virulence landscapes. Infect Genet Evol. 2026;105894. doi:10.1016/j.meegid.2026.105894
6. World Health Organization (WHO). The Role of Fluoroquinolones in the Treatment of Tuberculosis: Position Paper. Geneva: World Health Organization; 1997.
7. Zhang H, Yang J, Zhang Z, et al. Patterns and trends of primary drug-resistant tuberculosis in Chongqing, China, from 2012 to 2020. Medicine (Baltimore). 2023;102(10):e33230. doi:10.1097/MD.0000000000033230
8. Avalos E, Catanzaro D, Catanzaro A, et al. Frequency and geographic distribution of gyrA and gyrB mutations associated with fluoroquinolone resistance in clinical Mycobacterium tuberculosis isolates: a systematic review. PLoS One. 2015;10(3):e0120470. doi:10.1371/journal.pone.0120470
9. Chaiyachat P, Chaiprasert A, Nonghanphithak D, et al. Whole-genome analysis of drug-resistant Mycobacterium tuberculosis reveals novel mutations associated with fluoroquinolone resistance.International. J Antimicrobial Agents. 2021;2:14. doi:10.1016/j.ijantimicag.2021.106385
10. Sharma D, Lata M, Faheem M, et al. Role of M. tuberculosis protein rv2005c in the aminoglycosides resistance. Microb Pathog. 2019;132:150–155. doi:10.1016/j.micpath.2019.05.001
11. Castro RAD, Ross A, Kamwela L, et al. The genetic background modulates the evolution of fluoroquinolone-resis-tance in Mycobacterium tuberculosis. Mol Biol Evol. 2020;37:
12. Hoshide M, Qian L, Rodrigues C, et al. Geographical differences associated with single-nucleotide polymorphisms (SNPs) in nine gene targets among resistant clinical isolates of Mycobacterium tuberculosis. J Clin Microbiol. 2014;52(5):
13. World Health Organization. Global tuberculosis report 2023. Geneva: World Health Organization, 2023. Available from: https://www.who.int/.
14. Li T, Li J, Du X, et al. Age-specific pulmonary tuberculosis notification rates - China, 2008-2018. China CDC Wkly. 2022;4(38):841–846. doi:10.46234/ccdcw2022.176
15. Takiff HE, Salazar L, Guerrero C, et al. Cloning and nucleotide sequence of Mycobacterium tuberculosis gyrA and gyrB genes and detection of quinolone resistance mutations. Antimicrob Agent Chemother. 1994;38(4):773–780. doi:10.1128/AAC.38.4.773
16. Malik S, Willby M, Sikes D, et al. New insights into fluoroquinolone resistance in Mycobacterium tuberculosis: functional genetic analysis of gyrA and gyrB mutations. PLoS One. 2012;7(6):e39754. doi:10.1371/journal.pone.0039754
17. Kabir S, Tahir Z, Mukhtar N, et al. Fluoroquinolone resistance and mutational profile of gyrA in pulmonary MDR tuberculosis patients. BMC Pulm Med. 2020;20(1):138. doi:10.1186/s12890-020-1172-4
18. Momen G, Aainouss A, Lamaammal A, et al. Molecular characterization of mutations associated with resistance to second line drugs in Mycobacterium tuberculosis patients from Casablanca, Morocco. Rev Inst Med Trop Sao Paulo. 2021:63e19. 10.1590/S1678-9946202163019.
19. Chaiyachat P, Chaiprasert A, Nonghanphithak D, et al. Whole-genome analysis of drug-resistant Mycobacterium tuberculosis reveals novel mutations associated with fluoroquinolone resistance. Int J Antimicrob Agents. 2021;58(3):106385. doi:10.1016/j.ijantimicag.2021.106385
20. Hairong H, Nan D, Tingting Y, et al. Cross-sectional whole-genome sequencing and epidemiological study of multidrug-resistant Mycobacterium tuberculosis in China. Clin Infect Dis. 2018;69(3):3. doi:10.1093/cid/ciy883
21. Zhang Y, Li D, Liu J, et al. Association between pre-diagnostic fluoroquinolone exposure and possible acquired fluoroquinolone resistance in Mycobacterium tuberculosis in Shanghai: an EHR-based case-control study using whole-genome sequencing. J Infect. 2025;91(5):106648. doi:10.1016/j.jinf.2025.106648
22. Qiu D, Ma J, Liu Z, et al. Prevalence and molecular characterization of drug-resistant Mycobacterium tuberculosis in Heyuan City in China. Front Cell Infect Microbiol. 2025;15:1586938. doi:10.3389/fcimb.2025.1586938
23. Oliveira RS, Brandao AP, Ferreira FMDA, et al. Fluoroquinolone and second-line injectable resistance among rifampicin- and isoniazid-resistant Mycobacterium tuberculosis clinical isolates: a molecular study from a high-burden setting. Microorganisms. 2025;13(11):2470. doi:10.3390/microorganisms13112470
24. Zhang R, Bao X, Bao F, et al. Evaluation of the Xpert MTB/XDR test for detection of isoniazid, fluoroquinolones, and second-line injectable drugs resistance to Mycobacterium tuberculosis-Anhui Province, China. PLoS One. 2025;20(9):e0331264. doi:10.1371/journal.pone.0331264
25. Armstrong DT, Baird ECE, Pretty L, et al. Diagnostics and new treatment regimens for TB: can the Xpert MTB/XDR assay fill the gap for fluoroquinolone testing? J Clin Microbiol. 2025;63(10):e0064325. doi:10.1128/jcm.00643-25
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.