Back to Journals » Biologics: Targets and Therapy » Volume 18
Exploring Anticancer Potential of Lactobacillus Strains: Insights into Cytotoxicity and Apoptotic Mechanisms on HCT 115 Cancer Cells
Authors Wang L, Xu Z, Bains A, Ali N, Shang Z, Patil A ![]() , Patil S
, Patil S ![]()
Received 9 May 2024
Accepted for publication 21 September 2024
Published 1 October 2024 Volume 2024:18 Pages 285—295
DOI https://doi.org/10.2147/BTT.S477602
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shein-Chung Chow
Luolin Wang,1,* Zhenglei Xu,1,* Aarti Bains,2 Nemat Ali,3 Zifang Shang,4 Abhinandan Patil,5 Sandip Patil6
1Department of Gastroenterology, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen, Guangdong, People’s Republic of China; 2Deparment of Microbiology, Lovely Professional University, Phagwara, Punjab, India; 3Department of Pharmacology and Toxicology, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia; 4Research Experiment Center, Meizhou People’s Hospital, Meizhou Academy of Medical Sciences, Meizhou, Guangdong, People’s Republic of China; 5Department of Pharmaceutics, D. Y. Patil Education Society (Deemed to be University), Kolhapur, Maharashtra, India; 6Department of Haematology and Oncology, Shenzhen Children’s Hospital, Shenzhen, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Abhinandan Patil, Department of Pharmaceutics, D. Y. Patil Education Society (Deemed to be University), Kadamwadi, Kolhapur, Maharashtra, India, Email [email protected] Sandip Patil, Department of Haematology and Oncology, Shenzhen Children’s Hospital, Shenzhen, Guangdong, People’s Republic of China, Email [email protected]
Introduction: This study aims to systematically assess the anticancer potential of distinct Lactobacillus strains on Human Colorectal Tumor (HCT) 115 cancer cells, with a primary focus on the apoptotic mechanisms involved. Lactobacillus strains were isolated from sheep milk and underwent a meticulous microbial isolation process. Previous research indicates that certain probiotic bacteria, including Lactobacillus species, may exhibit anticancer properties through mechanisms such as apoptosis induction. However, there is limited understanding of how different Lactobacillus strains exert these effects on cancer cells and the underlying molecular pathways involved.
Methods: Cytotoxicity was evaluated through 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assays and exposure durations of Lactobacillus cell-free lyophilized filtrates. Additional apoptotic features were characterized using 4.6-diamidino-2-phenylindole (DAPI) analysis for nuclear fragmentation and Annexin V/PI analysis for apoptosis quantification. Genetic analysis explored the modulation of apoptotic proteins (Bax and Bcl2) in response to Lactobacillus treatment. Whole-genome sequencing (WGS) was performed to understand the genetic makeup of the Lactobacillus strains used in the study.
Results: The study demonstrated a significant reduction in HCT 115 cell viability, particularly with L. plantarum, as evidenced by Sulforhodamine B (SRB) and MTT assays. DAPI analysis revealed nuclear fragmentation, emphasizing an apoptotic cell death mechanism. Annexin V/PI analysis supported this, showing a higher percentage of early and late apoptosis in L. plantarum-treated cells. Genetic analysis uncovered up-regulation of pro-apoptotic protein Bax and down-regulation of anti-apoptotic protein Bcl2 in response to Lactobacillus treatment. WGS study revealed a strain reported to NCBI PRJNA439183.
Discussion: L. plantarum emerged as a potent antiproliferative agent against HCT 115 cancer cells, inducing apoptosis through intricate molecular mechanisms. This study underscores the scientific basis for L. plantarum’s potential role in cancer therapeutics, highlighting its impact on antiproliferation, adhesion, and gene-protein regulation. Further research is warranted to elucidate the specific molecular pathways involved and to evaluate the therapeutic potential of L. plantarum in preclinical and clinical settings.
Keywords: sheep milk, Lactobacillus plantarum, cytotoxicity, apoptosis, human colorectal tumor 115, cancer therapeutics
Introduction
Colorectal cancer (CRC) continues to impose a significant global health burden, necessitating ongoing exploration of innovative strategies for prevention and treatment.1 These are the third most commonly diagnosed cancer and according to recent data, CRC accounts for approximately 10% of all new cancer cases and 9% of all cancer-related deaths globally.2 Presently utilized therapeutic approaches, which encompass surgery, radiotherapy, and chemotherapy, have the potential to induce numerous undesirable side effects.3 These may include but are not limited to clinical conditions. Furthermore, the drugs employed in these treatments face challenges related to their lack of selectivity and the development of chemoresistance.4 These issues pose significant hurdles to the safety and efficacy of cancer therapy, impacting its overall management. Nutrition seems to play a pivotal role in preventing, initiating, and progressing cancer, complementing diverse and effective treatment approaches.5 The digestive system forms an intricate ecosystem hosting a variety of both advantageous and detrimental microorganisms.6 Probiotics, particularly various strains of Lactobacillus, have emerged as potential agents with diverse health benefits, including anticancer properties as elimination of carcinogens, modification of colon physicochemical conditions, metabolic activities in the intestinal microflora, production of anti-mutagenic or anti-tumorigenic compounds, and stimulation of the immune system.7,8 Few lactic acid bacteria (LABs), such as Lactobacillus casei, Lactobacillus rhamnosus, and Lactobacillus acidophilus, control over oncological markers in human CRC and preventive effects on carcinogen-induced colon malignancies in experimental animal models.9 LABs impede the proliferation of cells by interacting with them.10 This study focuses on a comparative assessment of the cytotoxic effects of novel Lactobacillus strains – L. plantarum, L. rhamnosus, and L. acidophilus – specifically targeting the HCT 115 colon cancer cell line. The findings of Gholipour et al suggested that the metabolomics of LABs directly induce toxicity against cancer cells. The primary objective is to unravel and understand the apoptotic mechanisms induced by these probiotic strains, shedding light on their potential as therapeutic interventions against colorectal cancer. The distinct characteristics of L. plantarum, L. rhamnosus, and L. acidophilus warrant investigation, particularly regarding their potential impacts on colon cancer cells. This study aims to provide valuable insights that could have implications for their application as adjuvants in colorectal cancer therapy. The isolation of Lactobacillus strains from sheep milk forms the foundation of our study. This study aims to contribute novel insights into the comparative cytotoxicity of distinct Lactobacillus strains on colon cancer cells. The elucidation of apoptotic mechanisms not only advances our understanding of probiotics’ anticancer potential but also provides a foundation for potential therapeutic applications. The findings hold promise for the evolving field of probiotics in oncology, particularly within the context of colorectal cancer research.
Methods
Sample Collection and Lactobacillus Strains Isolation
A total of 180 sheep milk samples (Indian breed) were collected from local places in the Kolhapur, Sangli, and Admapur areas of Maharashtra, India. For bacterial enumeration, milk samples (1 mL) were kept at −80°C in 15% glycerol before use. Isolation and purification were performed on de Man-Rogosa-Sharpe (MRS) broth and agar medium.11 The samples were inoculated on MRS agar and incubated for a period of 48 hrs under microaerophilic conditions. After incubation, the individual colonies formed were transferred into a sterile MRS broth medium to enrich. The purified cultures were identified using 16sRNA PCR assay and sequencing. The universal primers pair F-5′-CTTGTACACACCGCCCGTCA-3′ and reverse R-5′-CTCAAAACTAAACAAAGTTTC-3′ were used. The PCR protocol was adopted from our laboratory.12
Genome Drafting of Lactobacillus Strains
The identified strains were subjected to DNA extraction. Total DNA was extracted using a PureLink genomic DNA extraction kit (Life Technologies), following the manufacturer’s instructions. The sequencing libraries were prepared using a Nextera XT library kit, and whole-genome sequencing of L. plantarum, L. rhamnosus, and L. acidophilus strain denoted as JRARSH was performed on the Illumina NextSeq 500 sequencing platform (150-bp paired-end reads) with a shotgun strategy.13 The Illumina-generated sequence reads were quality filtered by FastQC v.0.10.1 (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/), and low-quality reads were removed before assembly. The quality-filtered reads were assembled de novo using SPAdes v.3.9,14,15 producing 31 contigs. The gene prediction was performed using the Rapid Annotations using Subsystems Technology (RAST) server v.2.0 (rast.nmpdr.org/), with default parameters.16–18 Predicted genes were annotated using the Prokaryotic Genome Annotation Pipeline (PGAP) v.4.919 and BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Default parameters were used for all software unless otherwise specified.
HCT 115 Cell Culture and Cell-Free Lyophilized Filtrate Preparation
The human colon cancer cell line HCT 115 is developed as monolayers by maintaining it in RPMI (Roswell Park Memorial Institution medium). The procurement of these cells was carried out from NCCS Pune, India. These media are augmented with 100 U/mL streptomycin-penicillin and 11% temperature maintained heated killed fetal bovine serum (FBS). The cells were maintained in a humidified atmospheric condition of 4.6% CO2 at around 36°C (approximately). After 80–85% confluence, cells were treated with the cell-free lyophilized filtrate of the bacterial samples. The L. plantarum, La and L. rhamnosus cultures of Lactobacillus were grown in MRS broth at 37°C overnight with a concentration of 1 X 107~8 cfu count/mL. Later, these cultures were centrifuged at 5100 x g for 5.5 min at 4°C to get pellets. The formed cell pellets were washed with PBS before ultrasonic disruption in a cooling ice bath (4°C) for 15 min at an interval of 28 sec. The formed supernatant is passed through the 0.22 µm pore microfilter (Himedia, USA). Further for cell-free lyophilized filtrates, centrifugation was used to extract the bacterial cells at 5000 rpm for 15 minutes at 4°C when the cultures attained the required OD600. The bacterial pellet was carefully separated from the supernatant, which included the released metabolites and bacterial by-products. A 0.22 μm sterile membrane filter was used to filter the supernatant in order to make sure there were no remaining cellular contamination. Using a freeze dryer, the resultant filtrate—which was devoid of bacterial cells—was lyophilised. After the material was lyophilised, it was reconstituted in sterile phosphate-buffered saline (PBS) to create a concentrated stock solution. This solution was then kept at −80°C until it was needed again. In the tests that followed, cytotoxicity and apoptosis were examined using this cell-free lyophilised filtrate (CFLF). The sterilized filtrate of supernatant was freeze-dried and various concentration was used for cell viability assays.20
Sulforhodamine B (SRB) Assay of L. plantarum, L. rhamnosus, and L. acidophilus
The cellular protein content is measured in SRB assay by determination of cell density. In brief, the cell in the concentration of 1×105−6 cells/mL was incubated for 24 hrs using a 96-well microtitre plate. After incubation, the cells were treated with the cell-free lyophilized filtrate of the L. plantarum, L. rhamnosus and L. acidophilus in concentrations of 10, 20, 30, 40 and 50 (µg/mL). Then, the total medium was incubated at 38°C in 5% CO2 atmospheric conditions for 24 hrs, 48 hrs, and 72 hrs. After the incubation period, the formed cell monolayers were fixed with 9.9% (w/v) trichloroacetic acid. Further, it was stained for 30 min with SRB and for removing the excess dye, 1% (v/v) acetic acid was used. The OD was calculated at 500 nm by dissolving the protein-bound dye in 9.9 mm Tris base solution using a microplate reader for determination of the cell viability. The percentage of cell viability was calculated by comparing with the values of the control well without any LABs samples and using Equation 1. L. acidophilus sample was used as a reference control group.21
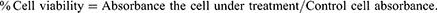
MTT Assay of L. plantarum, L. rhamnosus, and L. acidophilus
The anchorage-dependent viability in the form of the cell growth was determined by the colorimetric assay using [3-(4-5-dimethylthiazol-2yl-)-2, 5diphenyl tetrazolium bromide] MTT reagent. In the MTT assay, the purple formazan colour of the reduced MTT dye is found directly proportional to the viable cell number quantitated by the spectrophotometric study. In brief, 9000–10,500 cells of HCT 115 were seeded (triplicates) in the 96-well flat-bottomed tissue-culture plates for 24 hrs. After 24 hrs, the cells were treated with the cell-free lyophilized filtrate of the bacterial samples of L. plantarum, L. rhamnosus and L. acidophilus in a concentration of 10, 20, 30, 40 and 50 (µg/ mL). The negative control group kept for the assay consisted of a non-inoculated RPMI medium. The cells washed with the PBS solution after 24 hrs post-treatment were inoculated with 5 mg/mL solution of the MTT in PBS. Further, the cells were incubated for 4 hrs at 37°C for the mitochondrial dehydrogenase activity which was later visualized by the purple formazan crystals formation. Finally, in each well 120 µL of DMSO (dimethyl sulfoxide) was added and incubated for 28 min at 37°C. By the spectrophotometric method using a microplate reader, the colour intensity was measured at 570 nm. All the experiments were executed in triplicates (n=3), and the data were represented as percent viability compared to the control.22,23
Nuclear Staining with DAPI of L. plantarum, L. rhamnosus, and L. acidophilus
The cell-specific apoptotic process is characterized by DAPI assay. In HCT 115 cells, the analysis of nuclear fragmentation along with the chromatin condensation was done by DAPI staining using fluorescence microscopy. After treatment of 70–80% confluent HCT 115 cells with cell-free lyophilised filtrate of L. plantarum, L. rhamnosus and L. acidophilus (50 µg/mL in RPMI) for 24 hrs the cells were quickly washed with ice-cold PBS and fixed in ice-chilled acetone: methanol (1:1) mixture for 10 min at 4°C in the dark. The cells after washing with ice-cold PBS were incubated for 20 min with the DNA-specific fluorochrome, DAPI. The ice-cold PBS wash was carried out to remove the excess DAPI. Finally, the cells were observed using a fluorescence microscope at 20–40X resolution. L. acidophilus sample was used as a reference control group.24
Annexin-V and Propidium Iodide (PI) Assay of L. plantarum, L. rhamnosus, and L. acidophilus
HCT 115 cells were cultured in a dish of 60 mm and further treated with the cell-free filtrate of L. plantarum, L. rhamnosus and L. acidophilus (500 µL). After a 24-hr incubation period, the harvested cells were washed with cold PBS. A 1% Annexin binding buffer solution was prepared. 5 µL of the 1 mg/mL PI stock solution was used to prepare a diluted 100 µg/mL working solution of PI. The final solution along with the inoculated cell was centrifuged and the cells were suspended in 1X Annexin-binding buffer after discarding the supernatant solution. The final volume of 100 µL of cells per assay of buffer was prepared by diluting the 1X Annexin-binding buffer to ~1 × 105~6 cells/mL. 1 µL 100 µg/mL PI working solution and 6 µL of Alexa Fluor® 488 Annexin V were added to form 100 µL of cell suspension. Later, the cells were kept at room temperature for about 12 min for the incubation. Finally, the cells were inoculated with the 390~410 µL of 1X Annexin-binding buffer and were placed in an ice bath. The stained cells were evaluated by measuring the fluorescence emission at 520 nm using flow cytometry.20
Statistical Analysis
One way analysis of variance (ANOVA) with Tukey’s multiple comparison tests was used to compare the results of the probiotic and control groups using GraphPad Prism 5.01 software, USA. Standard deviations and significant differences were presented as *p<0.05, **p<0.01.
Results
Lactobacillus Strains and Genomic Study
The 16sRNA sequencing confirmed the three stains as L. plantarum, L. rhamnosus, and L. acidophilus and the phylogenetic tree was prepared (Figure 1A). The final assembly yielded a 3.20-Mb genome with a mean G+C content of 44.5%, an N50 value of 174,093 bp, an L50 of 6, and a total of 3128 genes, of which 2980 are protein-coding genes, 59 are tRNA genes, 5 are rRNA genes (35S rRNAs, 116S rRNA, and 123S rRNA), and 4 are noncoding RNA (ncRNA) genes. The genome also contained 80 pseudogenes. Within the total of 3128 genes, biological functions were defined for 2135 (68.25%). The predicted genes that were identified are associated with cellular metabolism (n =38), protein processing (n =323), energy (n = 276), stress response defence and virulence (n = 182), DNA processing (n = 181), cellular response (n = 163), RNA processing (91), membrane transport (n = 33), the cellular envelope (n = 28), and regulation and cell signalling (n = 18) (Figure 1B). Moreover, the genes were each assigned a putative function. Putative functions included cell adhesions, acid tolerance, bile toxicity, molybdenum cofactor biosynthesis, folate and pteridine biosynthesis, aromatic compound degradation, exopolysaccharide production, riboflavin and bacteriocin production, thiamine and methionine biosynthesis, phosphate metabolism, sulfur metabolism, dormancy and sporulation, and denitrification. This L. plantarum bacterial strain also proves to be an attractive candidate for the metabolic engineering of lignocellulosic biomass to biofuels, owing to its characteristic natural abilities to metabolize the hexose sugars and to tolerate high ethanol and acid concentrations.
 |
Figure 1 (A) Phylogenetic Tree based on 16S rRNA Sequencing Confirms L. plantarum (B1), (L) rhamnosus (C1), and L. acidophilus (D1). (B) Genome Annotation Overview of the Assembled Lactobacillus Genome. Different colors represent various functional categories. Cellular Metabolism (Red), Protein Processing (Purple), Energy (Yellow), Stress Response, Defense, and Virulence (Light Blue), DNA Processing (Dark Blue), Cellular Response (Pink), RNA Processing (Brown), Membrane Transport (Gray), Cellular Envelope (Teal), Regulation and Cell Signaling (Green). |
SRB Assay on HCT 115 Cell Line
The viability study conducted by SRB assay on the HCT 115 cell line shows a decrease in percentage cell viability of approximately 64% in the case of L. plantarum for 50 µg/mL concentration concerning the control group. Similarly, the cell viability results obtained in the case of L. rhamnosus and L. acidophilus show 74% and 88% viability of the HCT 115 cell line for 50 µg/mL with respect to the control group. Time-dependent SRB assay was carried out for a period of 24 hrs, 48 hrs, and 72 hrs. The study reveals that the cell viability decreased with time. L. plantarum shows a decrease in viability approximately from 64% at 24 hrs, 52% at 48 hrs to 48% after 72 hrs as compared to the control group (Figure 2A). Similarly, the cell viability decreased with time in the case of L. rhamnosus with an approximate decrease in viability from 76% at 24 hrs to 72% at 48 hrs and 70% at 72 hrs (Figure 2B). A very negligible decrease in the viability of HCT 115 cell lines are observed in the case of L. acidophilus. Thus, both time-dependent and concentration-dependent decreases in the cancer cell line viability is observed significantly in cases of L. plantarum as compared to L. rhamnosus and L. acidophilus by SRB assay.
 |
Figure 2 (A) Effects of the cell-free lyophilized filtrate of LAB on the viability of HCT 115 cell line for various concentrations by SRB assay. Values are expressed as viability mean ratio ± SD*, n = 3. (B) Effects of the cell-free lyophilized filtrate of LAB on the viability of HCT 115 cell line for various incubation time by SRB assay. Values are expressed as viability mean ratio ± SD*, n = 3. |
MTT Assay
In this assay, the MTT reagent passes through the viable cells and enters into the mitochondria, producing the dark blue/purple insoluble compound called formazan. This is observed only in viable cells, as the yellow tetrazolium salt from the MTT reagent is reduced to coloured purple dye only in the living cells. These viable colored cells get homogenized and soluble because of the use of organic solvent like isopropanol resulting in the formation of the formazan. The dead cells never show these activities thus, this assay gives the count of the living cells in the form of the viability count and determines the number of metabolically active cells. The viability study conducted by MTT assay on HCT 115 cell lines shows a decrease in percentage cell viability, approximately around 40% in the case of the L. plantarum for its highest dose of 50 µg/mL and further, showing the strongest antiproliferative activity as compared to the control group. In the case of L. plantarum, except for the dose of 10 µg/mL the other doses from 20 µg/mL to 50 µg/mL show anti-proliferative activity. Similarly, the cell viability results obtained in the case of L. rhamnosus and L. acidophilus are 74% and 88% on the HCT 115 cell line for its highest dose of 50 µg/mL with respect to the control group. L. acidophilus, which is used as the reference control group, does not show any antiproliferative activity (Figure 3A). Time-dependent MTT assay is carried out for a period of 24 hrs, 48 hrs, and 72 hrs. The study reveals that cell viability decreases with time. L. plantarum shows a decrease in viability count approximately from 41% at 24 hrs, 38% at 48 hrs to 34% after 72 hrs as compared to the control group. Similarly, the cell viabilities decreased with time in the case of L. rhamnosus with an approximate decrease in viability from 72% at 24 hrs, 70% at 48 hrs and 68% at 72 hrs (Figure 3B). While the negligible decrease in the viability in HCT 115 cell lines are observed in the case of the control group. Thus, both time-dependent and concentration-dependent decrease in the cancer cell line viability is observed significantly in the case of L. plantarum as compared to L. rhamnosus and L. acidophilus by MTT assay.
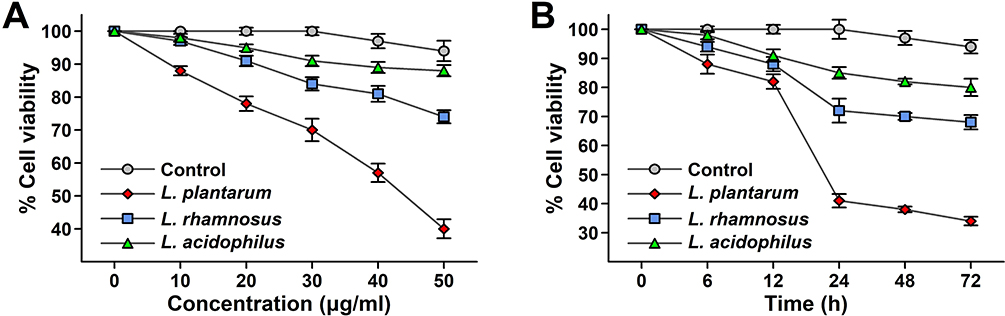 |
Figure 3 (A) Effects of the cell-free lyophilized filtrate of LAB on the viability of HCT 115 cell line for various concentrations by MTT assay. Values are expressed as viability mean ratio ± SD*, n = 3. (B) Effect of the cell-free lyophilized filtrate of LAB on the viability of HCT 115 cell line for various incubation time by MTT assay. Values are expressed as viability mean ratio ± SD*, n = 3. |
DAPI Analysis
The cell-free lyophilized filtrate of all three strains was analyzed for 24 hrs on HCT 115 cell lines. Significant apoptotic signals were observed in the case of L. plantarum as compared to L. rhamnosus. Non-apoptotic signals are observed in the case of the L. acidophilus which is used as a reference control group. The observed apoptotic cells were with fragmented condensed nuclei as compared to the intact colour normal cells. After 24 hrs incubation, the L. plantarum treated HCT 115 cells show apoptotic shrunk cells with nuclear fragmentation pattern as compared to the L. rhamnosus. The other features observed in the case of the apoptotic cells are the formation of a micronucleus blebbing cell membrane with the shrinkage of the nucleus in L. plantarum. While L. rhamnosus shows a blebbing membrane with a micronucleus. Apoptosis is visualized by DAPI (Figure 4A–D).
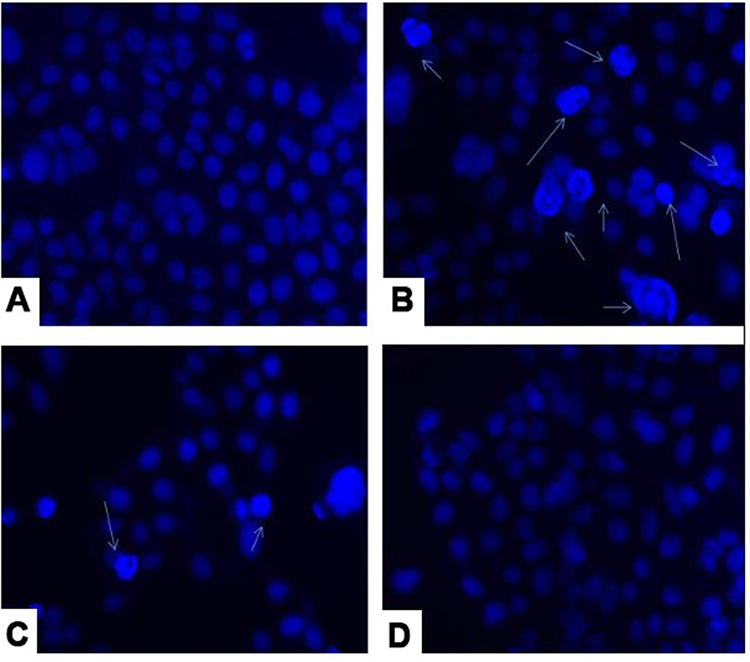 |
Figure 4 DAPI analysis of Lactobacillus strain (A) untreated cell line, (B) L. plantarum (C) L. rhamnosus and (D) L. acidophilus. |
Annexin V/PI Analysis of L. plantarum, L. rhamnosus, and L. acidophilus
HCT 115 cells are treated with the supernatant of L. plantarum, L. rhamnosus, and L. acidophilus for 24 hrs. Thus, mixed results of the fluorescent dots (annexin V+/PI-) are observed in the lower right quadrant indicating the early apoptosis phenomenon in the case of L. plantarum, and L. rhamnosus, while no such dots are observed in control L. acidophilus. Similarly mixed results of the fluorescent dots (annexin V+/PI-) are observed in the upper right quadrant indicating the late apoptosis phenomenon in cases of L. plantarum, and L. rhamnosus, while no such fluorescent dots are observed in control. The results revealed that about 57.5% of the early and 7.3% late apoptosis are observed on the HCT 115 cell lines by treatment with the L. plantarum supernatant (Figure 5A). In the case of the L. rhamnosus, the early and late apoptosis observed on the HCT 115 cell line was about 22.2% and 6.0% respectively (Figure 5B). The results also revealed that 0.0% and 0.1% of the early and late apoptosis are observed on the HCT 115 cell lines by treatment with the La supernatant, which was used as the reference standard control group in the study (Figure 5C). The necrotic dead count observed in the upper left quadrant in the case of L. plantarum, L. rhamnosus, and L. acidophilus were 1.7%, 6.4% and 0.7% respectively which are not affecting the overall study which was conducted. The pro-inflammatory agents are released due to the death of the cells by the necrosis phenomenon, while the death of the cells by apoptosis is the cascade event. It concludes that the death in the cell lines is due to the phagocytosis phenomenon, induced by the macrophages and cytokines due to the complementary system activation. The PI inclusion in the assay assesses the membrane integrity during the apoptosis process. The staining with the FITC-conjugated annexin V differentiates the category of death as necrosis or apoptosis (early or late). The current study proves that the L. plantarum exhibits as early and late apoptosis as compared with the L. rhamnosus with negligible necrosis events. Earlier studies have proven that L. plantarum SBL with a concentration of about 50 µg/mL after 24 hrs showed 20% and 51.5% early and late apoptosis on Hela cell lines.
 |
Figure 5 Annexin V/PI Analysis of L. plantarum, (L) rhamnosus, and L. acidophilus on HCT 115 Cells, (A) L. plantarum Treatment, Panel (B) L. rhamnosus Treatment, Panel (C) L. acidophilus Treatment (Control). |
Up/Down-Regulation of the Apoptotic Protein Expression of Lactobacillus Strains
The apoptotic proteins were investigated by the up-regulation and downregulation of the Bax, Bcl2 and beta-actin along with its expression ratio. Bax acts as pro-apoptotic proteins, it shows a significant increase (p<0.01) in the Bax of L. plantarum (treated) compared with the (untreated) control group. Similarly, there is a significant increase (p<0.05) in the Bax of L. rhamnosus compared with the control group. Thus increase in Bax is found higher in L. plantarum as compared to L. rhamnosus (Figure 6). Similarly Bcl2 acts as anti-apoptotic proteins, it shows a significant decrease (p<0.01) in the Bcl2 of L. plantarum compared with the control group. Similarly, there is a significant decrease (p<0.05) in the Bcl2 of L. rhamnosus compared with the control group. Thus, a decrease in Bcl2 expression is seen higher in L. plantarum as compared to L. rhamnosus. No significant changes are observed in the Beta-actin expression in case of L. plantarum and L. rhamnosus for both treated and untreated groups. This shows that L. plantarum shows significantly higher apoptotic protein expression properties as compared to L. rhamnosus. Apoptotic activities were reported by L. reuteri on HT 29 colon cancer cell lines by an increase in Bax and decrease in Bacl2. In a similar way, L. plantarum shows apoptotic activity by up-regulation of Bax and down-regulation of Bcl2.
 |
Figure 6 Up/Down-Regulation of Apoptotic Protein Expression of Lactobacillus Strains. Analysis of Bax (pro-apoptotic), Bcl2 (anti-apoptotic), and Beta-actin expression in treated and untreated groups. |
Discussion
Studying the cytotoxic effects of Lactobacillus strains on cancer cell lines is a major advancement in cancer research and treatment. This study aimed to investigate the the impact of three different Lactobacillus species (Lactobacillus plantarum, Lactobacillus rhamnosus, and Lactobacillus acidophilus) on HCT 115 colon cancer cells. We conducted several tests and examinations to understand the mechanics behind their ability to kill cells and to evaluate their potential as new options for cancer treatment. We used the SRB assay as the main method to evaluate cell viability and determine the cytotoxic effects of the Lactobacillus strains. This test, based on detecting cellular protein levels, provides information on cell viability by measuring their ability to bind the dye.25 We discovered that cell viability decreased in a dose-dependent manner after treatment with different Lactobacillus strains. Among them, L. plantarum showed the most antiproliferative effects compared to L. rhamnosus and L. acidophilus. This observation highlights the strong ability of L. plantarum to kill HCT 115 colon cancer cells, making it a good option for further investigation as a cancer treatment. This first report on the HCT15 cell line shows L. plantarum kills the colon cancer cells.
We used the SRB assay as the main method to evaluate cell viability and determine the cytotoxic effects of the lyophilized filtrates of Lactobacillus strains. The MTT assay measures the activity of mitochondrial enzymes in living cells to quantify cell viability by tracking the conversion of MTT into formazan crystals.26 Consistent with the SRB assay findings, we discovered that cell viability decreased in a dose-dependent manner after treatment with different Lactobacillus filtrates, especially L. plantarum. A similar kind of time and concentration-dependent MTT study was carried out on the L. plantarum DGK-17 with a dose of 25 to 250 μg, which showed the effective decline in the colon cancer cell line after 24 hrs.27 To the best of our knowledge, this is the first report on the HCT 115 cell line showing L. plantarum secretions’ cytotoxic effects on colon cancer cells.
We conducted further investigations, such as DAPI staining and Annexin V/propidium iodide (PI) analysis, to understand the processes of cell death caused by Lactobacillus strains.28 DAPI labelling allowed for the observation of nuclear structure, making it easier to identify characteristics of apoptosis such as nuclear condensation and disintegration. The results showed that cells treated with L. plantarum filtrates exhibited apoptotic features such as nuclear fragmentation and cell shrinkage, which are signs of programmed cell death. L. rhamnosus-treated cells showed reduced apoptotic signals, whereas La treatment did not trigger apoptosis, as indicated by the lack of apoptotic characteristics. L. plantarum mainly causes cell death by apoptosis, suggesting its promise as a focused treatment for cancer. An in-depth understanding of the apoptotic process was achieved by doing an Annexin V/PI analysis, which allows for the differentiation between the early and late stages of apoptosis. The study showed that L. plantarum and L. rhamnosus treatments induced both early and late apoptosis in HCT 115 cells, but L. acidophilus therapy resulted in modest apoptotic activity. This research offers additional proof of the particularity of L. plantarum-induced cell death and highlights its potential for use in cancer therapy. Furthermore, examination of apoptotic protein expression supported the apoptotic process responsible for the cytotoxic effects of L. plantarum.23–31
The pro-apoptotic protein Bax increased while the anti-apoptotic protein Bcl2 decreased in cells treated with L. plantarum filtrates, suggesting activation of the intrinsic apoptotic pathway. On the other hand, treatment with L. rhamnosus also resulted in higher Bax expression compared to L. plantarum, indicating distinct apoptotic signalling pathways amongst various Lactobacillus strains. Overall, our results highlight the strong inhibitory effects of L. plantarum filtrates on HCT 115 colon cancer cells by inducing apoptosis. The results provide important information on the molecular pathways responsible for the cytotoxic effects of Lactobacillus strains and highlight their potential as new therapeutic agents for treating cancer. Additional study is needed to clarify the precise signalling pathways involved in L. plantarum-induced apoptosis and to assess its effectiveness in both preclinical and clinical environments. Furthermore, investigating the possible synergistic impacts of Lactobacillus strains in combination with traditional cancer treatments could improve treatment results and lead to personalised cancer therapy strategies. Our focus on specific Lactobacillus strains may not fully represent the diverse probiotic landscape. Mechanistic intricacies of L. plantarum-induced apoptosis, though partially elucidated, require further exploration. The clinical relevance of our findings necessitates extensive research for safety, efficacy, and optimal dosage before considering L. plantarum filtrates as a therapeutic agent in cancer treatment. The study’s scope did not encompass the assessment of synergistic effects with traditional cancer treatments, suggesting avenues for future investigations into combination therapies for enhanced treatment outcomes.
Conclusions
The present study deals with the comparative cytotoxicity studies of Lactobacillus against the cancerous cell lines. The study shows that L. plantarum acts as an effective antiproliferative agent against cancer cell lines as compared to the L. rhamnosus and L. acidophilus. L. plantarum shows cell death by the apoptosis due to nuclear fragmentations. The current study shows that the L. plantarum exhibit the cell death mechanism as early and late apoptosis as compared with the L. rhamnosus with negligible necrosis events. The apoptotic protein expression studies show that L. plantarum kills the cancerous cell by up-regulation of Bax and downregulation of Bcl2. Thus, the in-vitro study shows the cancer killing tendency of the L. plantarum is by the combination of antiproliferative, and gene-protein regulation mechanisms.
Data Sharing Statement
The original contributions presented in the study are included in the article further inquiries can be directed to the corresponding author Sandip Patil ([email protected]).
Author Contributions
Luolin Wang and Zhenglei Xu contributed equally to this work and share the first authorship. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research was supported by the Guangdong Basic and Applied Basic Research Foundation (2021A1515110253), Researchers Supporting Project Number (RSPD2024R940), King Saud University, Riyadh, Saudi Arabia and Sanming Project of Medicine in Shenzhen (No. SZSM202211029).
Disclosure
Dr Abhinandan Patil reports grants from Inspire, Department of science and technology government of India, during the conduct of the study. The authors report no other conflicts of interest in this work.
References
1. Beniwal SS, Lamo P, Kaushik A, et al. Current status and emerging trends in colorectal cancer screening and diagnostics. Biosensors. 2023;13:926. doi:10.3390/bios13100926
2. Ferlay J, Colombet M, Soerjomataram I, et al. Cancer statistics for the year 2020: an overview. Int, J, Cancer. 2021;149:778–789. doi:10.1002/ijc.33588
3. Xie Y-H, Chen Y-X, Fang J-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct Target Ther. 2020;5:22. doi:10.1038/s41392-020-0116-z
4. Ma S-C, Zhang J-Q, Yan T-H, et al. Novel strategies to reverse chemoresistance in colorectal cancer. Cancer Med. 2023;12:11073–11096. doi:10.1002/cam4.5594
5. Basch CH, Hillyer GC, Jacques ET. News coverage of colorectal cancer on google news: descriptive study. JMIR Cancer. 2022;8:e39180. doi:10.2196/39180
6. Thursby E, Juge N. Introduction to the human gut microbiota. Biochem J. 2017;474:1823–1836. doi:10.1042/BCJ20160510
7. Śliżewska K, Markowiak-Kopeć P, Śliżewska W. The role of probiotics in cancer prevention. Cancers (Basel). 2020;13:20. doi:10.3390/cancers13010020
8. Gholipour F, Amini M, Baradaran B, Mokhtarzadeh A, Eskandani M. Anticancer properties of curcumin-treated Lactobacillus plantarum against the HT-29 colorectal adenocarcinoma cells. Sci Rep. 2023;13:2860. doi:10.1038/s41598-023-29462-7
9. Kaeid Sharaf L, Shukla G. Probiotics (Lactobacillus acidophilus and Lactobacillus rhamnosus GG) in Conjunction with Celecoxib (selective COX-2 inhibitor) Modulated DMH-Induced Early Experimental Colon Carcinogenesis. Nutr Cancer. 2018;70:946–955. doi:10.1080/01635581.2018.1490783
10. Bremnes RM, Dønnem T, Al-Saad S, et al. The role of tumor stroma in cancer progression and prognosis: emphasis on carcinoma-associated fibroblasts and non-small cell lung cancer. J Thorac Oncol. 2011;6:209–217. doi:10.1097/JTO.0b013e3181f8a1bd
11. Khedid K, Faid M, Mokhtari A, Soulaymani A, Zinedine A. Characterization of lactic acid bacteria isolated from the one humped camel milk produced in Morocco. Microbiol Res. 2009;164:81–91. doi:10.1016/j.micres.2006.10.008
12. Tannock GW, Tilsala-Timisjarvi A, Rodtong S, et al. Identification of lactobacillus isolates from the gastrointestinal tract, silage, and yoghurt by 16S-23S rRNA gene intergenic spacer region sequence comparisons. Appl Environ Microbiol. 1999;65:4264–4267. doi:10.1128/AEM.65.9.4264-4267.1999
13. Zee A, Deng DZQ, Adams M, et al. Sequencing Illumina libraries at high accuracy on the ONT MinION using R2C2. Genome Res. 2022;32:2092–2106. doi:10.1101/gr.277031.122
14. Malla MA, Dubey A, Kumar A, et al. Exploring the human microbiome: the potential future role of next-generation sequencing in disease diagnosis and treatment. Front Immunol. 2018;9:2868. doi:10.3389/fimmu.2018.02868
15. Malla MA, Dubey A, Yadav S, Kumar A, Hashem A, Abd Allah EF. Understanding and designing the strategies for the microbe-mediated remediation of environmental contaminants using omics approaches. Front Microbiol. 2018;9. doi:10.3389/fmicb.2018.01132
16. Aziz RK, Bartels D, Best AA, et al. The RAST Server: rapid annotations using subsystems technology. BMC Genomics. 2008;9(75). doi:10.1186/1471-2164-9-75
17. Kumar A, Vyas P, Malla M, Dubey D-A. Taxonomic and functional annotation of termite degraded butea monosperma (Lam.) Kuntze (Flame of the Forest). Open Microbiol J. 2019;13:154–163. doi:10.2174/1874285801913010154
18. Dubey A, Kumar A, Abd_Allah EF, Hashem A, Khan ML. Growing more with less: breeding and developing drought resilient soybean to improve food security. Ecol Indic. 2019;105:425–437. doi:10.1016/j.ecolind.2018.03.003
19. Tatusova T, DiCuccio M, Badretdin A, et al. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016;44:6614–6624. doi:10.1093/nar/gkw569
20. Salemi R, Vivarelli S, Ricci D, et al. Lactobacillus rhamnosus GG cell-free supernatant as a novel anti-cancer adjuvant. J Transl Med. 2023;21:195. doi:10.1186/s12967-023-04036-3
21. Botta C, Spyridopoulou K, Bertolino M, et al. Lactiplantibacillus plantarum inhibits colon cancer cell proliferation as function of its butyrogenic capability. Biomed Pharmacother. 2022;149:112755. doi:10.1016/j.biopha.2022.112755
22. Chuah L-O, Foo HL, Loh TC, et al. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement Altern Med. 2019;19:114. doi:10.1186/s12906-019-2528-2
23. Kang C-H, Kim J-S, Park HM, Kim S, Paek N-S. Antioxidant activity and short-chain fatty acid production of lactic acid bacteria isolated from Korean individuals and fermented foods. Biotech. 2021;3(11):217.
24. Munyenyembe K, Timmons C, Weiner AKM, Katz LA, Yan Y. DAPI staining and DNA content estimation of nuclei in uncultivable microbial eukaryotes (Arcellinida and Ciliates). Europ J Protistol. 2021;81:125840. doi:10.1016/j.ejop.2021.125840
25. Plessas S, Kiousi DE, Rathosi M, et al. Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains. Biomedicines. 2020;8:594. doi:10.3390/biomedicines8120594
26. El-Mokhtar MA, Hassanein KM, Ahmed AS, et al. Antagonistic activities of cell-free supernatants of lactobacilli against extended-spectrum β-lactamase producing Klebsiella pneumoniae and pseudomonas aeruginosa. Infect Drug Resist. 2020;13:543–552. doi:10.2147/IDR.S235603
27. Hiraishi N, Kanmura S, Oda K, et al. Extract of Lactobacillus plantarum strain 06CC2 induces JNK/p38 MAPK pathway-mediated apoptosis through endoplasmic reticulum stress in Caco2 colorectal cancer cells. Biochem Biophys Rep. 2019;20:100691. doi:10.1016/j.bbrep.2019.100691
28. Tiptiri-Kourpeti A, Spyridopoulou K, Santarmaki V, et al. Lactobacillus casei exerts anti-proliferative effects accompanied by apoptotic cell death and up-regulation of TRAIL in colon carcinoma cells. PLoS One. 2016;11:e0147960. doi:10.1371/journal.pone.0147960
29. Kim S, Lee HH, Choi W, et al. Anti-tumor effect of heat-killed bifidobacterium bifidum on human gastric cancer through Akt-p53-dependent mitochondrial apoptosis in xenograft models. Int J Mol Sci. 2022;23:9788. doi:10.3390/ijms23179788
30. Park J, Kwon M, Lee J, et al. Anti-cancer effects of lactobacillus plantarum L-14 cell-free extract on human malignant melanoma A375 cells. Molecules. 2020;25:3895. doi:10.3390/molecules25173895
31. Gao H, Li X, Chen X, et al. The functional roles of lactobacillus acidophilus in different physiological and pathological processes. J Microbiol Biotechnol. 2022;32:1226–1233. doi:10.4014/jmb.2205.05041
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.