Back to Journals » International Journal of General Medicine » Volume 18
Exploring Advances in Natural Plant Molecules for Allergic Rhinitis Immunomodulation in Vivo and in Vitro
Authors Li B, Dong B, Xie L, Li Y
Received 25 August 2024
Accepted for publication 20 December 2024
Published 1 February 2025 Volume 2025:18 Pages 529—565
DOI https://doi.org/10.2147/IJGM.S493021
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Vinay Kumar
Bingquan Li,1,* Boyang Dong,1,* Liangzhen Xie,2 Yan Li2
1Graduate School, Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China; 2Ear-Nose-Throat Department, The First Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Liangzhen Xie; Yan Li, Email [email protected]; [email protected]
Abstract: Allergic rhinitis (AR) is a prevalent allergic disease that imposes significant economic burdens and life pressures on individuals, families, and society, particularly in the context of accelerating globalization and increasing pathogenic factors. Current clinical therapies for AR include antihistamines, glucocorticoids administered via various routes, leukotriene receptor antagonists, immunotherapy, and several decongestants. These treatments have demonstrated efficacy in alleviating clinical symptoms and pathological states. However, with the growing awareness of AR and rising expectations for improvements in quality of life, these treatments have become associated with a higher incidence of side effects and an elevated risk of drug resistance. Furthermore, the development of AR is intricately associated with dysregulation of the immune system, yet the underlying pathogenetic mechanisms remain incompletely understood. In contrast, widely available natural plant molecules offer multiple targeting pathways that uniquely modify the typical pathophysiology of AR through immunomodulatory processes. This review presents a comprehensive analysis of both in vivo and in vitro studies on natural plant molecules that modulate immunity for treating AR. Additionally, we examine their specific mechanisms of action in animal models to provide new insights for developing safe and effective targeted therapies while guiding experimental and clinical applications against AR.
Keywords: natural plant molecules, allergic rhinitis, immunomodulation, therapeutic mechanism, research progress
Graphical Abstract:

Introduction
Allergic rhinitis (AR), a prevalent chronic inflammatory disease that affects the nasal mucosa of the upper respiratory tract, is categorized into two types: perennial and seasonal. This condition is characterized by recurrent episodes of sneezing, rhinorrhea, nasal congestion, and pruritus. In some patients, it may progress to allergic conjunctivitis, idiopathic dermatitis, allergic pharyngitis or eosinophilic esophagitis.1–3 The rapid advancement of the global economy coupled with accelerated industrialization and continuous changes in climate and environment has led to an increase in allergens and a rise in related risk factors, resulting in higher incidence rates of AR.4 Reports suggest that the global annual cost associated with AR is approximately $20 billion, with prevalence rates ranging from 10% to 40% worldwide and between 20% and 30% among adults; it exceeds 8.5% in school-age children and reaches up to 14% in adolescents in the Americas and Europe.5,6 The World Health Organization Working Group has recognized AR as a global health problem. This increasing prevalence poses a significant challenge to global health, having extensive implications for individuals as well as society at large. Therefore, there is an urgent need to identify safe and effective prevention methods along with control measures.
The pathogenesis of AR remains incompletely understood as it involves a variety of biological pathways primarily associated with regulating the immune system. The entry of allergens disrupts existing immune mechanisms and stimulates allergic reactions that damage the nasal mucosal structure and trigger an inflammatory response.7 Concurrently, it induces abnormal cellular secretion while compromising capillary integrity and impairing circulatory function. These processes can initiate various pathological reactions that may lead to allergic diseases through multiple pathways.8 Furthermore, studies have shown that the immune system transmits inflammatory signals to the central nervous system via both neuro-reflex and non-neuro-reflex mechanisms, activating specific brain regions and altering brain function. This understanding paves the way for targeted pharmacological interventions.9 Currently, in clinical practice, there are 15 commonly utilized antihistamines in Western medicine, including diphenhydramine, levocetirizine, desloratadine, and others; 6 types of glucocorticoids such as budesonide nasal spray, mometasone furoate, and fluticasone propionate; 4 varieties of decongestants including pseudoephedrine, oxymetazoline, and cylozolin; 8 different anti-leukotriene drugs like pranlukast, zafirlukast, and montelukast; along with 3 distinct forms of specific immunotherapy encompassing subcutaneous administration as well as sublingual and nasal immunotherapy.10 Since alternative therapies are excluded from these guidelines, treatment options remain limited. Additionally, other medications with targeted mechanisms are either undergoing clinical trials or have not yet reached the market. To date, many modern therapeutic strategies have failed to intervene effectively in the disease process and merely manage symptoms. As a result, identifying safe and effective drug-targeting pathways and developing multifunctional, multi-targeted drugs for AR within complex biological pathways present both challenges and opportunities.
In recent years, extensive studies have increasingly highlighted the unique therapeutic advantages of natural plant molecules in the treatment of AR. These molecules are characterized by their ability to target multiple pathways and channels while exhibiting minimal side effects, allowing them to influence relevant cells and mediators involved in immune regulation. Notably, the combination of specific monomers and components from Traditional Chinese Medicine (TCM) facilitates the development of personalized and precise therapeutic strategies. The herbs commonly utilized for the treatment of AR encompass Astragalus, Atractylodes macrocephala, Xanthium sibiricum, Divaricate saposhnikovia root, Mentha haplocalyx, Dark plum, and Magnoliae Flos. Their active constituents are frequently observed in curcumin, quercetin, as well as other polyphenols, flavonoids, and saponins.
However, due to the complexity of their pathways and mechanisms of action, the precise role of natural plant molecules in immunomodulation remains ambiguous. This paper aims to comprehensively explore the role of natural plant molecules in immunomodulating AR by systematically addressing the relationship between immunomodulation and the pathogenic aspects of AR, elucidating the specific mechanisms that affect its progression, and detailing the pathways through which natural plant molecules exert their immunomodulatory effects. The findings from this study will help identify new therapeutic targets for combating AR and provide valuable insights for future experimental studies and clinical applications.
Pathogenesis of AR
The pathogenesis of AR, a non-infectious disease, involves the activation of type 2 helper T cells (Th2 cells), which drive the IgE-mediated degranulation of mast cells and subsequent release of inflammatory mediators.11 Dendritic cells and macrophages within the body uptake allergens and inhibit Th1-type cytokine activity through antigenic Th2-type cytokines such as IL-4, IL-5, and IL-13, thereby inducing a series of immune responses. Additionally, cytokines such as IL-2 and INF-γ are involved in further contributing to these immune responses.12 AR progresses through two stages: sensitization and excitation.13 During the sensitization stage, initial exposure to allergens triggers a latency period of 1–2 weeks, during which protease activity compromises the barrier function of nasal mucosa and accelerates the production of pro-inflammatory factors. Concurrently, the nasal mucosal epithelium acts as the primary defense mechanism by extending dendritic cell projections beneath the mucosa upon invasion by allergens. This process exposes antigenic determinants to local lymph nodes, which synergistically stimulate signaling to naïve helper T cells (Th0 cells) and nasal mucosa CD4 cells, leading to their activation as Th2 cells and promoting the secretion of Th2 cytokines. Additionally, this activation results in increased expression of ICAM-1 by vascular endothelial cells. This activation sequence involves eosinophils, mast cells, and cytokines such as IL-4, IL-5, and IL-13. Cytokines play a pivotal role in the subsequent immune response by facilitating the differentiation of B cells into plasma cells and the synthesis of immunoglobulins that convert into specific IgE. This IgE binds to the surfaces of effector cells, such as mast cells and basophils, through the high-affinity receptor FcεRI in a sensitized state. Upon re-exposure to the same allergen, a patient with sensitized AR enters an excitation phase during which the allergen immediately binds to specific IgE, disrupting immune regulation. This phase consists of a rapid-onset and a delayed-onset phase. The rapid-onset phase occurs within seconds to six hours after allergen re-exposure and primarily involve effector cells like mast cells and basophils. During this phase, the allergen bridges two neighboring IgE molecules on effector cell surfaces, prompting cell degranulation and releasing numerous inflammatory mediators, including histamine, leukotrienes, prostaglandins, and platelet-activating factors. These mediators induce clinical symptoms such as nasal itching, sneezing, and a runny nose by binding to various receptors. The delayed phase occurs 12 to 24 hours after re-exposure primarily involving eosinophils as effector cells. Activated eosinophils release inflammatory mediators, including leukotrienes and eosinophil cationic proteins that contribute to symptoms like nasal congestion and a runny nose. During the activation phase, upon re-exposure to the allergen, it immediately binds to specific IgE on the surfaces of mast cells, triggering a pathological immune response (Figure 1).
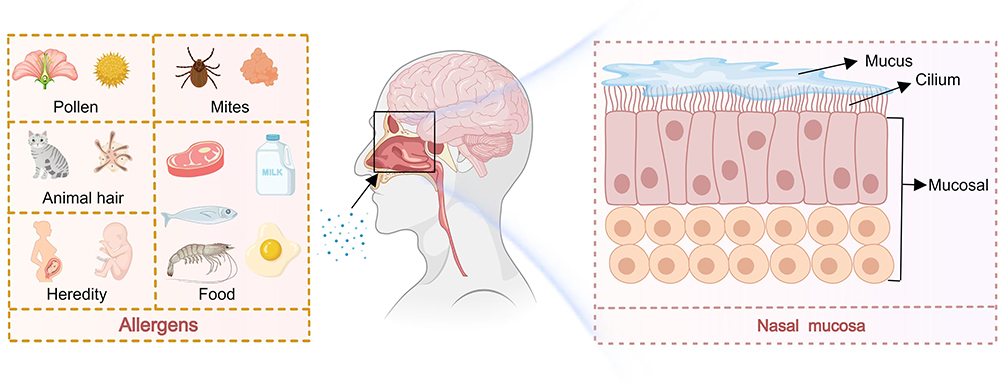 |
Figure 1 The pathogenesis of allergic rhinitis is that the allergen enters the nasal cavity. The allergen mainly comes from pollen, mites, animal fur, food and heredity, the mucosal cells produce pathological changes. |
Immunomodulatory Structure and Function
The concept of immunity can be categorized into two main types: physiological and pathological types. Animals are capable of distinguishing between two distinct categories of antigens: “self” and “non-self”. The body develops immune tolerance towards self-antigens while mounting rejection responses against non-self antigens. In healthy individuals, physiological immunity effectively detects and responds to pathogenic microorganisms through mechanisms such as phagocytosis, anti-inflammatory actions, and anti-infective responses, thereby maintaining physiological homeostasis.14 However, despite the immune system’s complex structure and numerous components, it can occasionally cause harm by triggering autoimmune diseases, hypersensitivity reactions, and inflammation in response to pathogen invasion. Upon encountering such invasion, the immune system rapidly mobilizes by differentiating immune cells that secrete and release molecules to activate the immune response.15 Subsequently, these immune cells secrete additional molecules that activate other bodily systems in a collaborative effort to eliminate pathogens and restore physiological stability. Modern immunology posits that the immune system of a healthy organism typically maintains stability through self-regulatory mechanisms. Nevertheless, when immune regulation is compromised, this can disrupt physiological balance and lead to disease. In Western medicine, immune-related diseases are addressed by administering exogenous substances to control disease factors, employing various drugs or surgical interventions to modulate the immune response and influence disease progression. Although these interventions often provide rapid relief, they primarily target symptoms rather than addressing the underlying causes of diseases, which may result in relapses and increased difficulty in managing the disease due to developed self-tolerance. As a result, Chinese medicine has gained attention from immunologists for its distinctive diagnostic and treatment approaches (Figure 2).
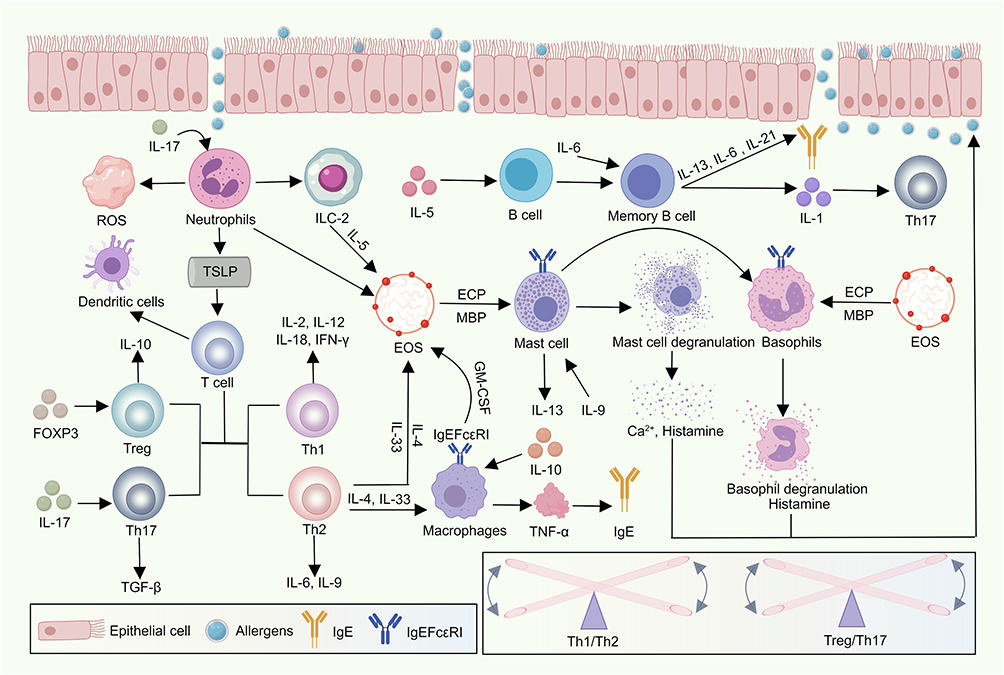 |
Figure 2 The Allergen invades the nasal mucosa, first into the neutrophil, triggering an oxidative stress response. It affects TSLP and releases Ilc-2. Ilc-2 release IL-5 factor under the activation of Allergen, make it act on B cells, and form memory B cells under the influence of IL-6, igE production and IL-1 cytokine production affect Th17 cell secretion. TSLP leads to imbalance of Th1, Th2, Treg and Th17 cells in T cells, and stimulates dendritic cell. Th2-releasing factors and IgE receptors act on macrophages to release IgE and other factors, il-33 and IL-4 factors released by Th2 Act together with IL-5 factors released by Ilc-2 on eosinophil granulocyte. The Eosinophil granulocyte release ECP, MBP and IgE receptors, which act on mast cells and basophil granulocyte. The mast cells degranulate to release calcium and histamine, and the basophil granulocyte degranulate to release histamine, which reacts with the nasal mucosa, initiate the corresponding inflammatory response. |
Granulocytes
Neutrophil
After differentiating and maturing in the bone marrow, neutrophils are released into either peripheral blood or tissues. In these locations, they carry out phagocytosis and degranulation functions that serve as the first line of innate immune defense.16 Stimulation by allergens on nasal mucosa results in a significant increase in mucosal neutrophils count along with T cells activation and enhanced migratory capacity for eosinophils.17 During the initial phase of AR, neutrophils, which are recruited prior to eosinophils, induce Th2-type inflammation in the airways through the formation of neutrophil extracellular traps (NETs), releasing inflammatory mediators.18 As AR progresses, the response to allergens is characterized by increased neutrophil chemotaxis, phagocytosis, and reactive oxygen species (ROS) production that contribute to oxidative stress. Furthermore, eosinophil cationic proteins (ECPs) are produced via an IgE-dependent mechanism, which further activates eosinophils and exacerbates inflammation.19
Basophil Granulocytes
Basophils, which mature in the bone marrow and are released into the peripheral blood, constitute only 0.5–1.0% of leukocytes.20 Despite their limited abundance, they rapidly synthesize Th2-type cytokines, thereby effectively supporting the immune response in AR.21 Basophils share several characteristics with mast cells and function as both Th2-inducing cells and antigen-presenting cells (APCs), primarily through the expression and engagement of the IgE receptor FcεRI on their cell surfaces.22 Upon exposure to allergens, allergen-specific IgE binds to basophils and cross-links with them, triggering the release of effector molecules such as histamine and leukotrienes. Notably, basophil activation is initiated by FcεRIα-mediated cross-linking via IgE antibodies independent of their presence on the basophil membrane. Basophils play a crucial role in the onset, development, and maintenance of AR as major effector cells that are highly sensitive to IgE-mediated activation.23 Research has demonstrated that basophils contribute to the pathogenesis of various allergic diseases, including AR. Clinically, the basophil activation test (BAT) aids in diagnosing AR and monitoring patient responses to therapy.24 However, the role of basophils in AR remains controversial, and further research is required to elucidate and clarify their involvement in the development and onset of AR.
Eosinophils
Eosinophils (EOS), derived from bone marrow hematopoietic stem cells, undergo differentiation and maturation within the bone marrow prior to being released into the peripheral blood.25 The development of EOS is influenced by various cytokines, including IL-5, IL-33, and granulocyte-macrophage colony-stimulating factor (GM-CSF).26 EOS serve as effector cells in natural immunity, along with neutrophils, participate in antibody-dependent cell-mediated cytotoxicity, resulting in tissue damage and inflammation. In allergic diseases, EOS go through multiple stages of development, adhesion, migration, and activation before being recruited to the site of inflammation.27 Mature EOS, regulated by IL-5, migrate from the bone marrow into the bloodstream. Following allergen stimulation, mast cells and endothelial cells at the site of inflammation secrete various cytokines that activate surface integrins (such as β1, β2, and β7) on EOS, thereby promoting their aggregation at the site. Once EOS adhere to epithelial cell ligands via integrins, a signaling cascade is activated, leading to the release of cytotoxic particles specific to EOS as they migrate toward the inflammation site, guided by the synergistic effects of chemokines.28 Additionally, activation products of eosinophils, including ECP and major basic protein (MBP), stimulate mast cells and basophils to release histamine, which increases vascular permeability and mucus secretion.29,30 This process ultimately damages the epithelium of respiratory mucosa and exacerbates local inflammatory responses.
Macrophages
Macrophages function as pivotal bridge cells between innate and adaptive immunity, playing a crucial role in coordinating immune responses. Upon pathogen invasion, macrophages present in the blood or tissues phagocytose and eliminate pathogens, induce cellular immune responses, and facilitate tissue repair and remodeling.31 Additionally, due to their inherent plasticity, macrophages exposed to diverse microenvironmental stimuli can differentiate into various subpopulations, predominantly M1 and M2-type macrophages.32 The roles of macrophages include: 1) providing antigenic information to lymphocytes; 2) producing oxygen free radicals and lymphokines, such as IL-1, which modulate inflammatory responses; 3) generating inflammatory mediators like arachidonic acid and platelet-activating factor; 4) modulating lymphocyte activation; and 5) exhibiting limited affinity for the IgE Fcε surface receptor. Macrophages play a crucial role in balancing Th1/Th2-type immune responses in AR.33 Cytokines and chemokines secreted by M2-type macrophages facilitate the differentiation of Th2 cells and the recruitment of EOS. Th2-type cytokines, such as IL-4 and IL-13, promote macrophage polarization towards the M2 phenotype through STAT6 activation. Conversely, M2 macrophages secrete various cytokines and chemokines including Arginase-1 (Arg-1), CCL17, and CCL24 to regulate eosinophil recruitment and mediate allergic inflammation.34 Furthermore, increased TGM2 expression in M2 macrophages enhances eosinophilic infiltration. Additionally, macrophages and classic T-lymphocyte immune mechanisms complement each other and regulate the progression and resolution of allergic diseases while also influencing the course of AR.35,36
Mast Cells
Mast cells (MCs) are the primary effector cells in type I hypersensitivity reactions, playing a crucial role in the activation and aggregation of neutrophils and EOS. Originating from the bone marrow, MCs circulate in the bloodstream in an immature form before migrating to various tissues.37 The degranulation of MCs is central to the pathophysiology of AR, with the secretory response varying significantly based on the size and distribution of FcεRI clusters.38 Immature MCs undergo maturation through IgE-induced histamine synthesis and cytokine release. Upon maturation, they produce and store numerous dense cytoplasmic granules, which are released in response to specific stimuli.39 The most common stimulus for this release is cross-linking of IgE molecules via multivalent antigen binding to the high-affinity IgE receptor, FcεRI. The binding of IgE to multiple FcεRI on the cell membrane initiates the phosphorylation of downstream proteins, activating signaling molecules and generating second messengers.40,41 This cascade results in the release of Ca2+ from the MC’s endoplasmic reticulum, leading to subsequent MC degranulation. Degranulation through the IgE-dependent pathway results in various symptoms, including damage to epithelial tissue, thickening of the basement membrane, hypertrophy of airway smooth muscles, and increased respiratory mucus.42,43
Dendritic Cells (DCs)
Dendritic cells (DCs) are derived from hematopoietic stem cells in the bone marrow. In the human nasal mucosa, DCs predominantly reside in the epithelium and lamina propria. They traverse the tightly connected epithelium through dendritic projections.44 Based on their origin, DCs can be classified into plasma cell-like dendritic cells (pDCs) and myeloid dendritic cells (mDCs).45 The two principal types of DCs found in blood and lymphoid tissues are conventional DCs (cDCs) and pDCs. Functioning as “transmitters” in the pathogenesis of AR, DCs by capture and present allergens to naive T cells, subsequently initiating a cascade of immune responses.46–48 Together with macrophages and B cells, DCs form the three primary populations of APCs. Unlike macrophages, which do not migrate to lymph nodes or activate T cells ex vivo, DCs possess the ability to transport antigens to draining lymph nodes, thereby initiating T cell activation. Only live, mature, and fully functional DCs that migrate to the lymph nodes can effectively stimulate a T cell response.49,50
B-Cells
B cells, which are essential for humoral immune defense and effective antigenic recognition, migrate back to local lymph nodes and serve as sources of allergen-specific IgE in AR.51 They can either mediate an immediate immune response by producing IgG or act as regulatory B cells in the absence of pathogenic entities.52 AR is categorized into two phases: sensitization and stimulation. During the sensitization phase, the initial exposure of the nasal mucosa to allergens involves a latency period of 1–2 weeks. During this time, protease activity impairs the barrier function of the nasal mucosa upon contact with the allergen and accelerates the production of pro-inflammatory factors. Patients with AR exhibit a significant increase in memory B cells, plasma cells, and CD19+CD24hiCD27+ Bregs, while naïve B cells are substantially reduced compared to healthy individuals.53 The initial increase in serum allergen-specific IgE characterizes the B cell response, likely representing a memory response triggered by exposure to allergens. Memory B cells, which become activated upon encountering antigens, maintain long-term sensitization or tolerance to allergens through rapid proliferation, somatic mutations, and class-switch recombination.54 Treg cells inhibit the functions of B cells and influence their proliferation, activation, and apoptosis. In AR patients, B cells have a notable impact on the magnitude, duration, and affinity of IgE production. The IL-21 receptor signaling in B cells is crucial for regulating IgE.55
T-Cells
T lymphocytes, crucial regulators of the cellular immune response, mature into two primary types: CD4+ T cells and CD8+ T cells. CD4+ T cells can be further subdivided into three subtypes based on their secretion of cytokines: Th0, Th1, and Th2.56 Th1 cells secrete cytokines such as IL-2, IL-12, and INF-γ that play a role in in humoral immunity and regulate the differentiation of cytotoxic T helper cells, thereby contributing to the pathogenesis of AR. In contrast, Th2 cells produce cytokines like IL-4 and IL-5, which activate MCs, EOS, and basophils. These immune cells accumulate at sites of inflammation where they degranulate and release inflammatory mediators to modulate AR.57 Regulatory T cells (Tregs), a specialized subtype of T lymphocytes characterized by the expression of CD4+, CD25+, and the transcription factor Foxp3, possess immunoregulatory functions exerting suppressive effects on immune responses. Thymic Treg cells (tTregs) interact with DCs to promote immune tolerance. Tregs can be categorized into two groups based on their origin: tTregs and induced Treg cells (iTregs), each playing distinct roles in immune regulation.58 In vivo, Tregs either differentiate directly in the thymus or develop from conventional CD4+ T cells in the periphery. Foxp3, a crucial regulator of Treg development and function, is essential for inducing and maintaining the Treg cell phenotype. Foxp3+ Treg cells sustain their suppressive functions both in vivo and in vitro after expansion, presenting potential therapeutic avenues for AR.59 The acetylation, phosphorylation, and ubiquitination of Foxp3 are integral to the development and functional maintenance of tTreg cells by influencing protein stability and DNA-binding capacity. Foxp3 collaborates with other transcription factors and proteins to shape the transcriptional profile and effector functions of Treg cells. Disrupting specific protein interactions with Foxp3 diminishes Treg cell functionality. Under specific environmental conditions, Treg cells can modulate the components of the Foxp3 protein complex to regulate inflammation associated with AR.60 The expression and stability of Foxp3 are crucial in maintaining the functionality of Treg cells. Treg cells exhibit both plasticity and instability, as activated Tregs can inhibit the interaction between T-cell responses and DCs. Post-translational modifications, including hyperacetylation, play a crucial role in regulating the plasticity and stability of Treg cells by modifying Foxp3. IL-2, a key cytokine for promoting T cell proliferation and differentiation, plays an indispensable role in the development, survival, and function of Foxp3+ Treg cells. Studies have demonstrated that naturally occurring Treg cells are characterized by their expression of Foxp3. The suppressive function of Treg cells is influenced not only by their expression of Foxp3 but also by the microenvironment. Tregs exert immunosuppressive effects through cell-contact mechanisms and secretion of inhibitory cytokines, while also influencing T cell function by regulating APCs.61 Foxp3 is currently recognized as the most reliable molecular marker for natural Treg cells, elucidating the mechanisms underlying Treg cell differentiation. Foxp3 collaborates with various transcription factors and proteins to shape the transcriptional profile and effector functions of regulatory T (Treg) cells.62 Disruption of specific protein interactions with Foxp3 impairs the functionality of Treg cells. Under certain environmental conditions, Treg cells can modulate the components of the Foxp3 protein complex to regulate inflammation associated with AR. The expression and stability of Foxp3 are crucial for maintaining Treg cell functionality. Treg cells exhibit both plasticity and instability; activated Tregs can inhibit the interaction between T-cell responses and DCs.63–65 Post-translational modifications of Foxp3, including hyperacetylation, play significant roles in regulating the plasticity and stability of Treg cells. Interleukin-2 (IL-2), a key cytokine for T-cell proliferation and differentiation, is indispensable for the development, survival, and function of Foxp3+ Treg cells. Numerous studies have demonstrated that naturally occurring Treg cells are characterized by the expression of Foxp3.66 The suppressive function of Treg cells is not only influenced by Foxp3 expression but also by the surrounding microenvironment. In addition to regulating APCs, Tregs exert their immune-suppressive effects through both cell-contact mechanisms and secretion of inhibitory cytokines. Currently recognized as the most reliable molecular marker for natural Treg cells, Foxp3 elucidates the underlying mechanisms involved in Treg cell differentiation.67 However, the instability in Foxp3 expression can significantly diminish their suppressive capacity, potentially disrupting immune tolerance and contributing to AR. The mechanisms governing Foxp3 stability and Treg cell integrity remain incompletely understood.68 Furthermore, understanding these regulatory mechanisms is crucial due to the impact of specific Treg cell populations on the severity of AR when exposed to allergens. Clinical trials have investigated the therapeutic potential of Treg cells for immune disorders, suggesting their prospective application in managing AR.69,70
Inflammatory Factor
IL-1
Interleukin-1 (IL-1), a lymphocyte-stimulating factor commonly found in the forms of IL-1α and IL-1β, is predominantly produced by macrophages.71 Additionally, almost all nucleated cells, including B cells and DCs, can produce IL-1. IL-1 plays a role in negative regulation of the inflammatory signaling pathway mediated by the IL-1 receptor/Toll-like receptor.72 Research indicates that IL-1R8 significantly negatively regulates Th17 cell differentiation, proliferation, and function by directly inhibiting the IL-1 signaling pathway.73 Immunomodulatory effects of IL-1 include: 1) synergistic effects with antigens to activate CD4+ T cells; 2) promotion of B cell growth and differentiation, enhancing CD4+ T cell activation and expression of the IL-2 receptor, leading to a 100-fold increase in the number of hemolytic vacuoles in splenocytes (PFC), indicating the role of IL-1 in promoting antibody production; 3) enhancement of antigen-presenting capacity for monocytes, macrophages, and other APCs; 4) synergy with IL-2 or interferon to enhance NK cell activity; and 5) attracting neutrophils, which subsequently release inflammatory mediators.74–76
IL-2
Interleukin-2 (IL-2), a cytokine produced by activated Th1 cells, primarily promotes the proliferation of killer T cells and NK cells, enhances the differentiation and proliferation of B cells, stimulates antibody production, and induces lymphokine-activated killer cells.77 IL-2 is crucial for the immune response; nonetheless, aging and various diseases can diminish IL-2 production, subsequently impacting the entire immune system. In order for IL-2 to exert its immunomodulatory effects, it must bind to the activated membrane interleukin-2 receptor (mIL-2R).78 The soluble interleukin-2 receptor (sIL-2R), a polypeptide released into the bloodstream by activated Th1 cells, competes with mIL-2R for IL-2 binding and inhibits the clonal expansion of activated T cells. The IL-2/IL-2R system plays a central role in regulating the body’s immune response.79
IL-3
Interleukin-3 (IL-3), an important member of the interleukin family and also referred to as multi-directional colony-stimulating factor (multi-CSF), is a cytokine produced by T lymphocytes.80 It stimulates the proliferation in the immune response-associated cells, including Th2 cells that secrete pivotal cytokines. IL-3 is crucial in the pathogenesis of AR as it stimulates the proliferation and differentiation of multipotent hematopoietic stem cells, MCs, EOS, and basophils.81,82 Furthermore, IL-3 induces not only multipotent hematopoietic stem cell and MCs proliferation and differentiation but also that of EOS and basophils.83 Due to its broad impact on various cell types, IL-3 is also referred to as a multi-colony stimulating factor. Under normal circumstances, IL-3 concentration is tightly regulated through negative feedback. However, in allergic conditions, activation of the IL-3 gene leads to abnormally high concentrations of this cytokine.84
IL-4
Interleukin-4 (IL-4) is a cytokine secreted by type II helper T cells (Th2 cells). As a hallmark of Th2 cells, IL-4 promotes the proliferation of activated B and T cells and facilitates the differentiation of CD4+ T cells into type II helper T cells.85 Additionally, it regulates humoral and adaptive immunity, induces class switching to IgE in B cell antibody production, and upregulates the expression of type II major histocompatibility complex.86 IL-4 plays critical roles in metabolic responses such as promoting the differentiation of type 2 helper T lymphocytes, inducing IgE production, upregulating cellular adhesion molecule 1 expression via the IgE receptor, facilitating eosinophil migration to the lungs, inhibiting apoptosis in T lymphocytes and promoting mucus hypersecretion.87 Furthermore, IL-4 enhances the adhesion of EOS and basophils to endothelial cells and inhibits the production of γ-interferon.88 Additionally, IL-4 prompts the proliferation, differentiation, and maturation of MCs. Upon activation, MCs release IL-4 into the extracellular microenvironment, which in turn activates T cells to produce more IL-4 and further initiates recruitment of inflammatory cells.89 The documented impact of IL-4 on MC proliferation, differentiation, and maturation is well-established. Moreover, it has been shown that IL-4 inhibits TGF-β-mediated FoxP3 expression, thereby preventing the conversion of naive T cells into Tregs. Allergenic individuals exhibit increased secretion of IL-4 following allergenic stimulation.90,91
IL-5
The multifunctional cytokine Interleukin-5 (IL-5) is produced by cells of the immune system. It is primarily produced by activated T cells and is particularly abundant in the Th2 cell subpopulation.92 IL-5, secreted by activated Th2 helper cells, possesses proliferative, chemotactic, and activating functions for EOS.93 When EOS infiltrate the nasal mucosa, they can release ECP and MBP, both of which are cytotoxic and neurotoxic.94 These proteins can damage the nasal mucosa and enhance glandular secretion, leading to symptoms such as nasal congestion, itching, runny nose, and sneezing. IL-5 stimulates the proliferation and differentiation of B cells while also promoting antibody production to enhance the humoral immune response. On one hand, IL-5’s activation of EOS synergizes with other cytokines to enhance their inflammatory infiltration and increase the expression of adhesion molecules.95 On the other hand, IL-5 prolongs the lifespan of EOS and alters their density from normal to low. While IL-5 activates EOS, these cells can also produce more IL-5 themselves, creating a feedback loop that exacerbates and prolongs AR.96,97
IL-6
Interleukin-6 (IL-6), a cytokine secreted by Th2 cells, is also produced by MCs, epithelial cells, and endothelial cells within the body. IL-6 exerts regulatory effects on the growth and differentiation of various cell types.98 The cytokine induces T-cell activation, growth, and differentiation, as well as promoting the differentiation of Th0 cells into Th17 cells and MC proliferation.99,100 In response to IL-6 stimulation, B-cells differentiate into mature plasma cells that subsequently secrete IgE. IL-6 plays a crucial role in the cascade of inflammatory mediators contributing to fibrosis and airway remodeling. The effects of IL-6 are dose-dependent: normal levels have beneficial effects, while excessive production leads to various inflammatory damages. Serum levels of IL-6 increase under conditions such as inflammation, infection, and certain tumors.101
IL-9
Interleukin-9 (IL-9), produced by Th2 cells, serves as an important factor in immune regulation and the inflammatory response.102 By enhancing MC survival and function, IL-9 promotes inflammation and triggers the release of additional inflammatory mediators. IL-9 acts on effector Th2 cells and memory T cells of the Th2 phenotype, which directly differentiate from them.103 In IL-9 cells, the transcription factor PU.1 activates Th9 cell differentiation while inhibiting Th2 cell differentiation. As a result, cytokine expression in Th2 cells is significantly reduced, whereas IL-9 expression is markedly increased.104,105
IL-10
Interleukin-10 (IL-10), derived from multiple cell types, is a multifunctional cytokine that regulates cellular growth and differentiation and plays a crucial role in inflammatory and immune responses.106 IL-10 exerts its influence on key functions of monocyte macrophages, including the release of immune mediators and antigen presentation. In brief, IL-10 inhibits the ability of monocyte macrophages to promote natural and specific immunity functions while enhancing cytostatic, immune tolerance-inducing, and scavenging activities.107 Indeed, IL-10 suppresses the secretion of inflammatory mediators by monocyte macrophages, thereby reducing the levels of TNF-α, IL-1β, IL-6, IL-8, G-CSF, and GM-CSF in response to LPS and IFN-γ.108 Furthermore, IL-10 promotes the release of anti-inflammatory factors, including IL-1 receptor antagonists and soluble TNF-α receptors. Thus, IL-10 significantly reduces the levels of key cytokines involved in innate immunity and inhibits antigen presentation by monocyte macrophages.109 Moreover, IL-10 suppresses the synthesis of IL-12, thereby hindering the Th1 immune response. The inhibitory effect of IL-10 on cellular factor production and antigen presentation is further enhanced by CD4+ T cell suppression. Additionally, IL-10 hampers IFN-γ production by Th1 cells, while a deficiency in IFN-γ enhances APC inactivation. IL-10 also decreases macrophage secretion of essential cytokine IL-23 required for Th17 cell-mediated immunity.110
IL-12
Interleukin-12 (IL-12), produced mainly by activated macrophages and DCs, is a significant immunomodulatory cytokine.111 This cytokine is crucial in promoting the secretion of Th1 cytokines, such as IFN-γ, which enhance immune responses of Th cells. Simultaneously, it inhibits the production of Th2 cytokines, like IL-4, that suppress immune responses of Th2 cells.112
IL-13
Interleukin-13 (IL-13), an immunomodulatory protein with inhibitory properties, shares both functional and structural similarities with IL-4. It is primarily produced by Th2 cells and MCs. As members of the Th2 gene family, they both utilize the same receptor chain and signaling system. In the absence of IL-4, IL-13 serves as a critical factor in inducing Th2 responses and is a major mediator of Th2 cell production.113 During the development of allergic diseases, such as AR, levels of IL-13 are significantly elevated. IL-13 induces mitosis in B-lymphocytes activated by IgM or anti-CD40 monoclonal antibody, contributing to their differentiation and antibody class switching. Additionally, it enhances the production of IgE, IgM, and IgG, while inhibiting apoptosis in B-cells.114 In the presence of activated CD4+ T cells or anti-CD40 monoclonal antibody, IL-13 autonomously induces B cells to synthesize IgE. IL-13 functions as a TH2 cell effector by inducing the synthesis and extracellular secretion of IgE by activated B lymphocytes. Additionally, it enhances the body’s response to mutagens, promotes cuprocyte proliferation and gland secretion, and exacerbates nasal cicatrization in AR.115,116 As a TH2 effector, IL-13 plays a pivotal role in the pathogenesis of AR through its regulation of Th1 and Th2 cell functions, promotion of IgE secretion by B cells, and localization of the response to mutagens.117
IL-17
Interleukin-17 (IL-17), a subset of pro-inflammatory cytokines, is mainly synthesized by activated CD4+ lymphocytes and stimulates T lymphocytes, particularly during the initial stages of inflammation, affecting a variety of cells and tissues.118 It induces mesenchymal stromal cells to secrete inflammatory and hematopoietic cytokines, such as IL-6 and IL-8. Furthermore, it prompts fibroblasts to facilitate the proliferation and differentiation of CD34+ hematopoietic progenitors, a process closely related to autoimmunity and tumorigenesis.119 Additionally, IL-17 amplifies inflammation by inducing the release of neutrophil chemokines.120
IL-18
Interleukin-18 (IL-18), a member of the serum IL-1 family, is a widely distributed pro-inflammatory factor with diverse biological activities. It is expressed in various cells including B-cells, T-cells, macrophages, osteoblasts, and keratinocytes.121 In AR patients, IL-18 can up-regulate IgE production and activate Th2 cells to produce cytokines. It synergizes with IL-12 to produce IFN-γ, which inhibits IL-4-induced IgE synthesis. This ultimately reduces serum IgE levels, decreased eosinophil aggregation in the nasal mucosa, and suppressed proliferation of Th2 cells. These actions result in decreased release of inflammatory mediators and airway hyperresponsiveness (AHR), ultimately reducing airway inflammation.122,123 Simultaneously, IL-18 promotes the secretion of IL-4 and IL-13 from T cells, MCs, and basophils, playing a dual regulatory role in Th1 and Th2 cytokine production as well as IgE synthesis. Furthermore, IL-18 enhances IFN-γ production through synergistic and autocrine effects during AR progression, inhibiting the synthesis of IgE induced by IL-4 while promoting Th1 differentiation and suppressing Th2 cells to affect the immune balance between Th1/Th2.124,125 It also activates Th2 cells to produce cytokines, contributing to the immune imbalance between Th1/Th2 associated with AR.126
IL-22
Interleukin-22 (IL-22), a member of the IL-10 cytokine family, is primarily synthesized by various immune cells, including Th17 and Th22 cells. IL-22 is crucial in maintaining the barrier function of the respiratory mucosa by promoting epithelial cell survival and repair, thereby enhancing resistance to mechanical and chemical damage.127 In AR, this function helps mitigate mucosal damage and strengthens defenses against external allergens.128 It collaborates with cytokines like IL-17 and may also be implicated in releasing inflammatory mediators that exacerbate the nasal mucosa’s inflammatory response, characterized by mucosal swelling and increased secretions.129
IL-33
The cytokine Interleukin-33 (IL-33), recently identified as tissue-derived, connects natural and adaptive immunity and promotes mucosal Th2 inflammatory responses; it is also referred to as an epithelial-derived alerting hormone. The ST2 receptor, specific to IL-33, becomes activated when IL-33 binds to ST2 on the surfaces of MCs, various immune cells (including Th2 and ILC2s), and DCs.130 This interaction recruits downstream signaling molecules that activate nuclear factor (NF)-κB and mitogen-activated protein kinase (MAPK), which mediate the release of inflammatory mediators from Th2 cytokines and promote allergic diseases.131 The ST2 receptor is prevalent among a variety of immune cells, including Th2 cells. Binding of IL-33 to the ST2 receptor forms an immune complex expressed in Th2 cells, promoting the release of inflammatory mediators such as IL-4, IL-5, and IL-13 while regulating the functions of basophils, MCs, EOS, and intrinsic lymphocytes.132
IFN-γ
Interferon-γ (IFN-γ), a characteristic cytokine secreted by Th1 cells, activates macrophages and neutrophils, inhibits the differentiation of Th0 cells into Th2 cells, and suppresses secretion, proliferation, and activation of Th2 cells. Consequently, it indirectly hampers type I allergic reactions.133 Inherently inflammatory in nature, Th1 cells can elicit an associated inflammatory response when allergen-specific Th1 cells are transferred to a patient’s lungs. Although this transfer does not directly induce AHR that exacerbates asthma and allergic reactions, IFN-γ is frequently detected at sites of allergic inflammation where it promotes inflammatory responses.134 Additionally, IFN-γ induces inducible nitric oxide synthase (iNOS) production in macrophages and promotes NO synthesis. It also stimulates microglial astrocytes to produce iNOS, potentially contributing to the development of certain allergic diseases.
TNF-α
Tumor necrosis factor-α (TNF-α), primarily produced by macrophages and monocytes, plays a crucial role in the pathogenesis of allergic diseases.135 In patients with AR, the nasal mucosa exhibits high levels of macrophages, monocytes, polymorphonuclear leukocytes, and MCs, all secreting TNF; consequently, elevated TNF concentrations are observed in their serum.136 TNF-α is produced and released by IgE-dependent activation of monocyte-macrophages, inducing the expression of adhesion molecules in epithelial cells. This cytokine also promotes the infiltration and activation of inflammatory cells, stimulates monocytes and T-cells to secrete IL-6 and IL-8, and directs Th cell differentiation towards Th2 phenotype. This cascade leads to an IgE-mediated type I hypersensitivity reaction, causing immune damage.137,138
TGF-β
Transforming growth factor-β (TGF-β) belongs to a superfamily that regulates cellular growth, differentiation, and metabolism. Secreted by various cells, TGF-β primarily governs the healing process of injuries and influences cell proliferation, differentiation, immune responses, and fibrosis.139 In terms of immune regulation, TGF-β1 has been discovered to chemotactically attract and regulate neutrophils, EOS, monocytes, and other inflammatory cells. This prompts their accumulation at the site of inflammation and triggers an inflammatory response.140 Additionally, TGF-β1 inhibits the differentiation of CD4+CD25− T cells into Treg cells while promoting a bias towards Th17 cell differentiation.141 These highly effective pro-inflammatory effectors enhance the recruitment of inflammatory cells through the release of mediators, thereby expanding and sustaining acute inflammatory responses in the body. Concerning immunosuppression mechanisms, TGF-β1 inhibits the maturation and differentiation processes in immune cells such as Th1 cells, Th2 cells, B cells, as well as innate lymphocytes, while reducing associated cytokine releases like IFN-γ and IL-2.142 The involvement of TGF-β1 further enhances the expression in the nasal mucosa, induces histopathological changes, and potentially leads to tissue remodeling within the nasal mucosa.143
T-Cell Levels
During the immune response, CD4+ T helper (Th) cells undergo several stages of differentiation. Initially, upon stimulation by external antigens and under the influence of various inducing factors, CD4+ T cells can differentiate into diverse subpopulations, including Th1, Th2, Th17, Th9, and Tregs. This process of differentiation is guided by the cytokines produced and the physiological functions of these cells. While operating independently as distinct T cell subpopulations, they also demonstrate interconnections.
Th1/Th2
When allergens invade the body, APCs process and present allergenic peptides to Th0 cells, initiating their differentiation into Th2 cells and resulting in a Th1/Th2 immune imbalance.144 Th2 cells facilitate the synthesis and secretion of IgE by MCs and promote IgE release from B cells through various cytokines. Concurrently, Th2 cells secrete cytokines such as IL-4 and IL-13, with IL-4 playing a crucial role in driving further differentiation from Th0 to Th2 and directly stimulating B cell production of IgE antibodies.145 In contrast, Th1 cells secrete IL-2 and interferon-γ (IFN-γ), both of which inhibit in vivo IgE production by blocking IL-4-induced IgE synthesis.146 In patients with AR, IgE binds to the IgE-FcεRI receptor on MCs, macrophages, and epithelial cells’ surfaces, sensitizing these cells. Upon re-exposure to the allergen, it crosslinks two adjacent surface-bound IgEs, triggering the release of inflammatory mediators, cytokines, and neuropeptides that affect the nasal mucosa, leading to various clinical symptoms of AR.147
Treg/Th17
Under normal physiological conditions, there is a dynamic equilibrium between the number and function of Th17 and Treg cells. Disruption of this balance can lead to the onset and exacerbation of various diseases, typically characterized by an increase in Th17 cells and a decrease in Treg cells. Th17 cells primarily contribute to tissue damage and inflammatory cell infiltration through secretion of IL-17, which induces the expression of metalloproteinases, chemokines, and pro-inflammatory cytokines.148 Treg cells, as a subset of T cells, inhibit and regulate the proliferation and activation of T lymphocytes, thereby exhibiting anti-inflammatory activity and maintaining immunological homeostasis. It is worth noting that a reduction in Treg cells is associated with several autoimmune diseases. In AR patients, Th17 cells exhibit enhanced pro-inflammatory effects compared to healthy individuals, resulting in elevated levels of cytokines such as IL-17, IL-25 (also known as IL-17E), and TNF-α.149 Additionally, Th17 cells stimulate other cell types to produce significant amounts of inflammatory cytokines, including IL-6, IL-8, and GM-CSF. Collectively, these cytokines exacerbate the inflammatory response, further worsening the clinical symptoms observed in AR.150
Natural Plant Molecules Modulate Pathways Associated With AR Immunomodulation
This paper initially discusses recent advancements in the immunomodulatory mechanisms associated with AR. Subsequently, we summarize and analyze experimental studies, recent reviews, and web-based pharmacological analyses pertaining to the utilization of natural plant molecules for AR treatment. We also provide a summary of the effectiveness and corresponding pathways by which these plant compounds modulate immunomodulation. The chemical structures of these plant compounds are shown in Figure 3.
Figure 3 Continued. Figure 3 Chemical structure of natural plant molecules.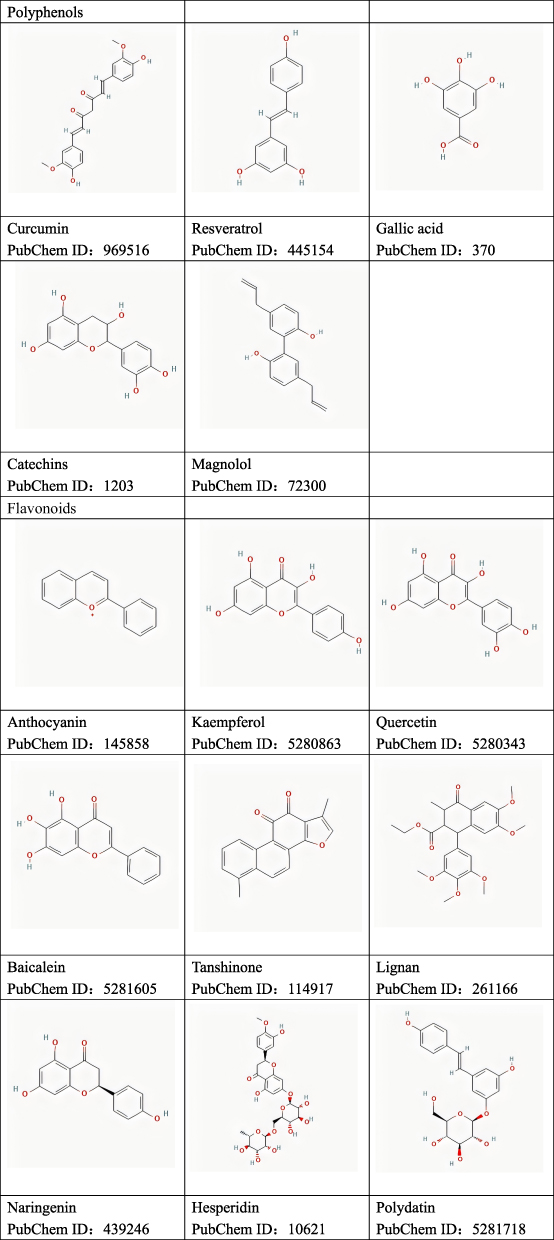
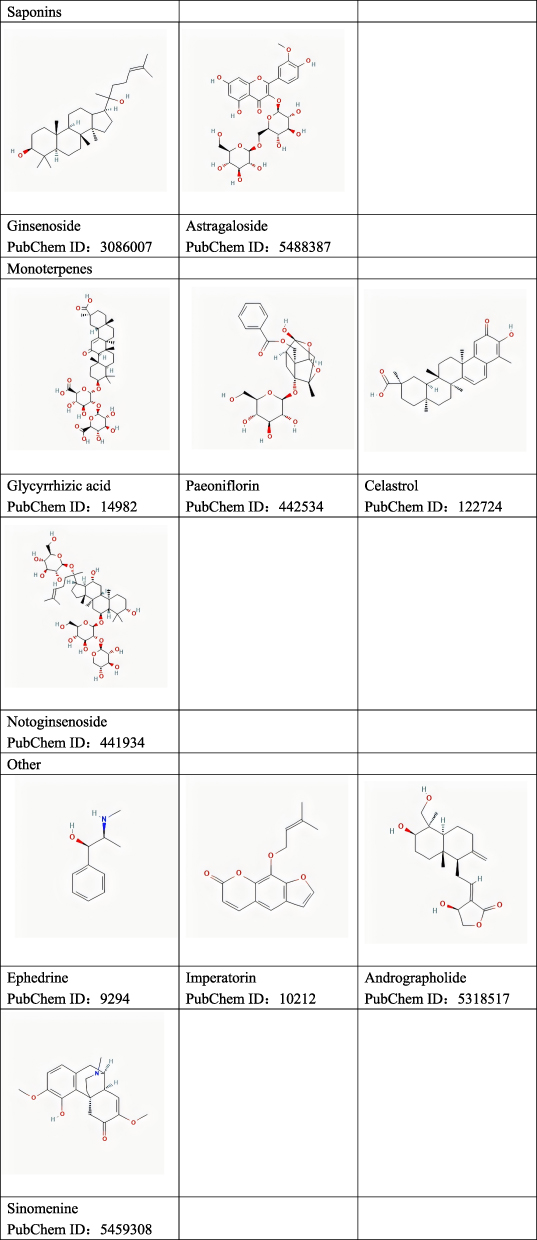
Polyphenols (Table 1)
Curcumin
Curcumin (Cur), the primary active compound found in turmeric, is a natural polyphenol with a medicinal history spanning over 4000 years. It is derived from the Curcuma longa plant and has been widely utilized in traditional Chinese medicine. In the past decade, there has been a surge in research focusing on the pharmacological activities and mechanisms of action of curcumin, attracting growing attention from the scientific community.162,163 Numerous studies have confirmed that curcumin possesses diverse properties, including anti-inflammatory, antioxidant, antibacterial, antifungal, antiprotozoal, antiviral, hepatic and renal protective, anti-fibrotic, anticancer, and anti-carcinogenic effects. Additionally, it exhibits anti-fibrotic and anticancer effects without causing significant toxic side effects.164,165 Recent findings suggest that curcumin can inhibit histamine release in animal models of type I and type IV allergic reactions, highlighting its potential as an anti-allergic agent. In a study conducted by Zhang Ning et al,151 curcumin was administered to OVA-sensitized mice, resulting in a reduction of immune cytokine levels through the inhibition of histamine release and specific IgE production in MCs. Furthermore, curcumin demonstrated its efficacy in alleviating nasal symptoms and improving the morphology of damaged nasal mucosa by attenuating histamine and specific IgE levels, reducing TNF-α expression, and inhibiting the release of inflammatory mediators through the stabilization of I-κBα in the cytosol, thus preventing PM-stimulated nuclear translocation of NF-κB in MCs. Under physiological conditions, the human body maintains redox homeostasis. Nevertheless, disruption of this balance caused by internal or external factors that increase free radical concentrations can lead to oxidative damage to biological macromolecules and various diseases.166 Furthermore, a study on human nasal fibroblasts demonstrated that curcumin reduced the levels of ROS induced by urban particulate matter (UPM), achieving this through the activation of the Nrf2/HO-1 pathway.152 It was observed that curcumin activated Nrf2 and inhibited extracellular signal-regulated kinase (ERK) in UPM-exposed fibroblasts, significantly increasing antioxidant enzymes like heme oxygenase-1 (HO-1) and superoxide dismutase (SOD2). Consequently, this alleviated oxidative stress and mitigated allergic nasal symptoms.
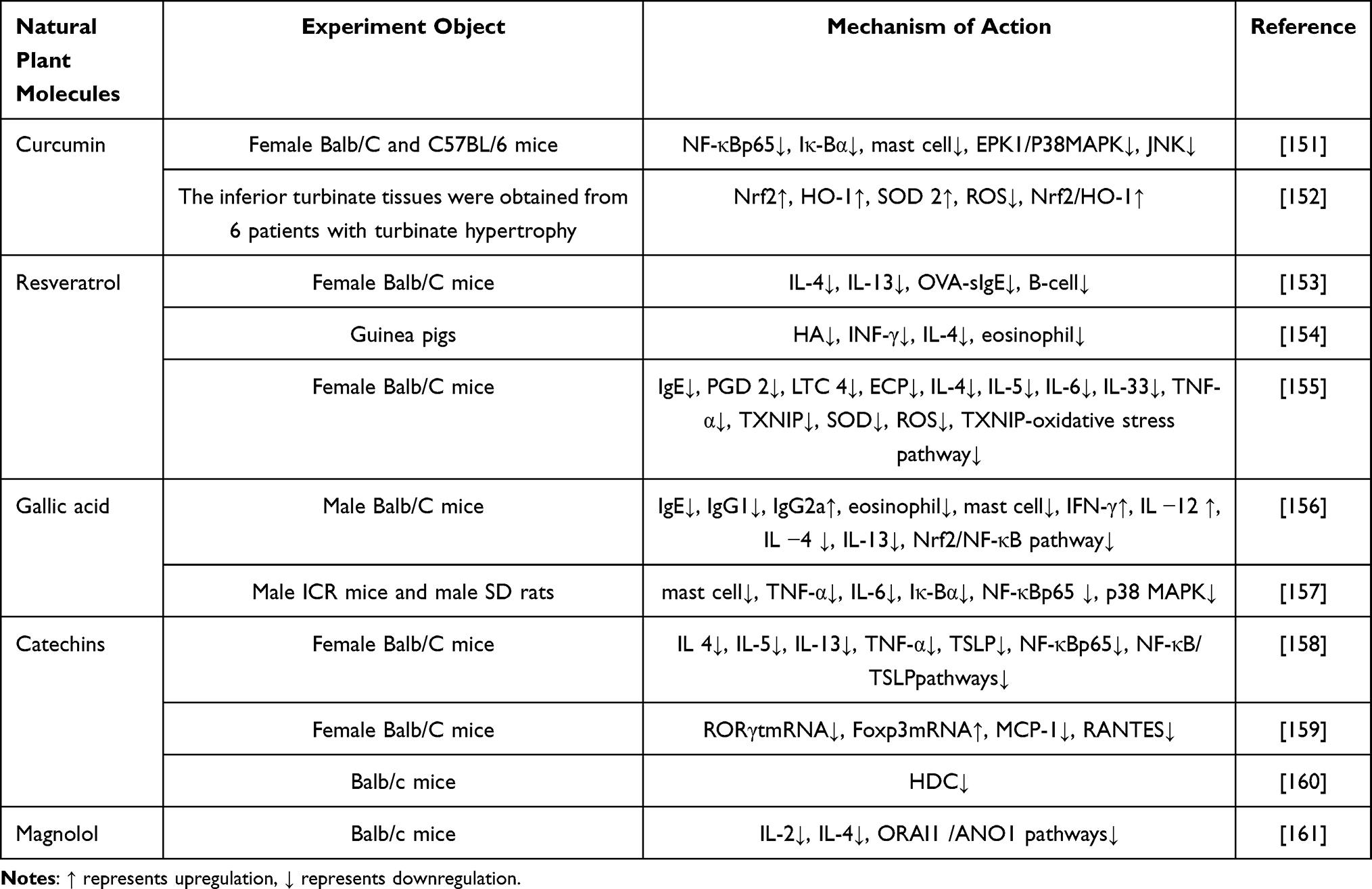 |
Table 1 The Immunomodulatory Effects and Mechanisms of Natural Plant Polyphenols on AR Were Summarized |
Resveratrol
Resveratrol is a naturally occurring polyphenol found in various herbs and foods, including tiger nuts, grapes, and soybeans. Currently, it is marketed as both a nutritional supplement and food additive. Due to its broad pharmacological effects, resveratrol exhibits potent properties in terms of anti-inflammatory, antioxidant, antiviral, hypolipidemic, hypoglycemic, and anticancer activities.167,168 It is anticipated to emerge as a novel therapeutic agent for disease prevention and treatment. Experiments conducted on different animal models have confirmed that resveratrol can repair damaged nasal mucosa cells by regulating the release of inflammatory factors and alleviating allergy symptoms through the reduction of oxidative stress.160,169 In a randomized controlled trial focusing on AR, it was demonstrated that resveratrol has the ability to regulate serum levels of inflammatory factors and T-lymphocyte subpopulations.170 This study further revealed that modifying the stoichiometry of resveratrol resulted in reduced infiltration of inflammatory cells,153 decreased serum levels of Th2 cytokines IL-4, IL-13, and OVA-sIgE; inhibited polarization of Th2 cells; regulated immune function; and altered Th2 polarization, while modifying the inflammatory response. Resveratrol not only suppresses the elevation of peripheral blood INF-γ but also mitigates histopathological changes in the nasal turbinate mucosa.154 It reduces eosinophil infiltration and inhibits glandular secretion. Additionally, resveratrol was found to decrease SOD activity by inhibiting the TXNIP-oxidative stress pathway,155 subsequently reducing MDA and ROS production. As a result, this led to the down-regulation of inflammatory mediators in splenic mononuclear cells and decreased levels of PGD2, LTC4, ECP, IL-4, IL-5, IL-6, IL-33, and TNF-α. However, further large-scale animal and human studies are necessary to elucidate and refine the mechanisms of action of resveratrol for potential therapeutic enhancement.
Gallic Acid
Gallic acid (GA) is a key constituent of several traditional Chinese herbal plants, including Pentaphyllum, Pawpaw Rhubarb, Cornu Cervi, and Myrtus communis. Previous studies have demonstrated that GA exhibits a range of properties, including antioxidant, anti-inflammatory, anti-allergic, antibacterial, antiviral, and antitumor effects with minimal toxicity to normal cells.171,172 These findings suggest the significant potential of GA in the field of medicine.173 Recently, Shenna et al conducted an experimental study on the effects of GA in mice with AR.156 The results revealed that GA modulates the Nrf2/NF-κB pathway which influences immunoglobulin levels (IgE, IgG1, and IgG2a), as well as alters nasal mucosa thickness. These outcomes indicate that GA can enhance immune function and augment resistance against pathogens in nasal mucosa while also modulating inflammatory responses. Sang-Hyun Kim et al demonstrated that GA reduces histamine release from MCs induced by compound 48/80 or immunoglobulin E (IgE),157 regulates cAMP and intracellular calcium levels, inhibits nuclear factor kB and p38 mitogen-activated protein kinase, and contributes to blocking the expression of pro-inflammatory factors. These actions mitigate in vivo allergic reactions, restore and enhance local immune function, strengthen the nasal mucosa’s resistance to pathogens, thereby reducing the incidence of AR.
Catechins
Catechins, classified as flavanols within the flavonoid family, are natural polyphenolic compounds found in various plants. Ongoing research has unveiled that catechins possess a range of pharmacological effects, including anti-inflammatory, antibacterial, antiviral, and antioxidant properties.174,175 Recent experimental studies utilizing catechins to treat AR in mice have demonstrated significant improvements in behavioral responses, reductions in leukocyte counts, inhibition of inflammatory cell infiltration, and alleviation of local inflammation symptoms.158 Furthermore, catechins have been shown to inhibit the NF-κB pathway and reduce the degradation of IκBα proteins while influencing upstream regulators of TSLP in epithelial cells. This modulation leads to decreased levels of TSLP and NF-κB p65 protein,176 suggesting that catechins can modify the response of nasal mucosa epithelial cells and provide protection to the nasal mucosa. Yan Yajie et al discovered that catechins reduce the levels of MCP-1 and nasal mucosa RORγt mRNA, while simultaneously increasing Foxp3 mRNA levels.159 This is associated with a decrease in inflammatory cell infiltration in the nasal mucosa. Additionally, catechins not only alleviate inflammatory symptoms and protect the nasal mucosa in mice with AR, but also restore the disrupted Th17/Treg balance, correct immune imbalances, and accelerate recovery from AR. Mohamed A. Morsy et al’s study demonstrated that catechins can inhibit histamine expression and reduce histidine decarboxylase (HDC) activity.160 Molecularly, catechins have been shown to exhibit greater activity than the standard HDC inhibitor, histidine methyl ester, positioning them as an effective alternative treatment.177
Magnolol
Magnolol (MG), a novel oleuropein compound derived from the traditional Chinese medicine magnolia, has been found to possess anti-inflammatory potential in previous in vivo and in vitro studies.178 The current study suggests that MG inhibits the nuclear translocation of activated NF-κB by reducing glial activation, peripheral blood cell infiltration, and the production of pro-inflammatory cytokines.179 It is now recognized that NF-κB, a protein with multiple transcriptional regulatory roles, serves as a central component in the complex network structure of allergic diseases such as AR and asthma. By regulating cytokines associated with inflammation, immunity, and inflammatory mediators, NF-κB plays a crucial role in inflammation and immune responses. Hong Thi Lam Phan et al conducted a study on AR treatment using MG,161 which demonstrated that MG (30 μM) inhibited anti-CD3-induced cell proliferation. Furthermore, it suppressed IL-2 production via ORAI1 channels in T lymphocytes and reduced IL-4 secretion through ATP-induced electrolyte transport mediated by ANO1 channels to correct the Th1/Th2 imbalance.
Flavonoids (Table 2)
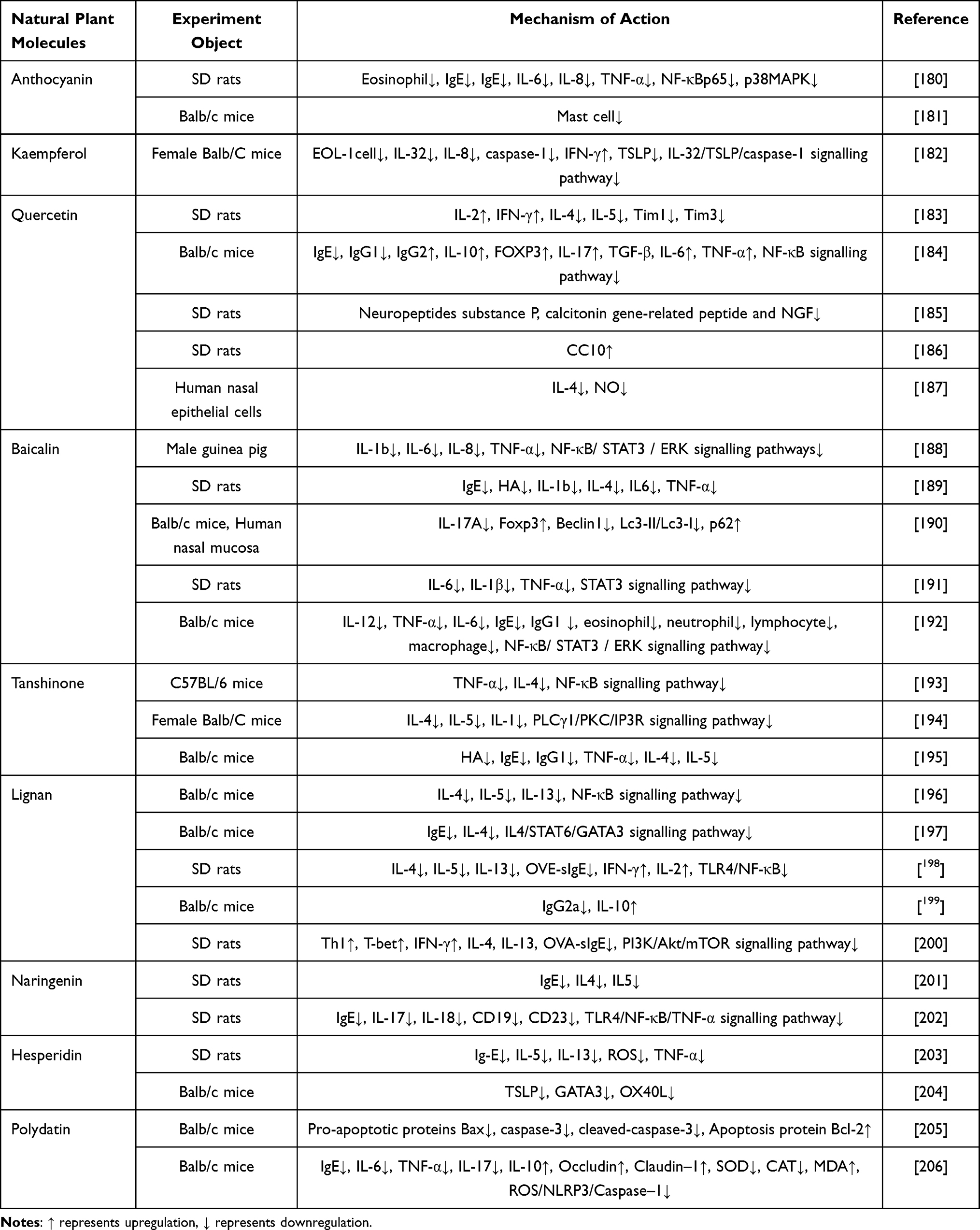 |
Table 2 To Summarize the Immunomodulatory Effects and Mechanisms of Flavonoids Natural Plant Molecules on AR |
Anthocyanin
Anthocyanin belongs to the class of polyphenolic flavonoids, which are secondary metabolites synthesized by higher plants. Anthocyanins are widely found in the kernels, skins, or seeds of various plants and contribute to the coloration of most angiosperm flowers, fruits, and leaves, ranging from orange-red to dark blue. Different plants exhibit varying sensitizing properties for anthocyanins. Being the most abundant and widely distributed flavonoid pigments, they absorb the longest wavelengths of light. Extensive studies on quercetin, catechins, and isoflavones with similar structures as anthocyanins may provide a basis for evaluating anthocyanins’ biological activities.207 Previous studies have demonstrated that anthocyanins possess anti-inflammatory, antibacterial, antioxidant, and anticancer properties.208 Recently, Hongli Hua et al examined the effects of anthocyanins in AR rats exposed to PM2.5.180 The researchers discovered that anthocyanins could restore the damaged epithelial structure of the nasal mucosa, resulting in a higher number of cilia, a more organized arrangement, and reduced infiltration of inflammatory cells as well as chemotaxis of goblet cells in the lamina propria. Additionally, anthocyanins were shown to decrease the levels of IgE, IL-6, IL-8, and TNF-α, while also reducing the expression of NF-κB p65 and phosphorylated p38MAPK proteins in the nasal mucosa, thus altering allergic behavioral symptoms in rats. Katsunori Yamaura et al found that anthocyanins alleviated itching symptoms in mice by inhibiting MC degranulation.181 Moreover, a separate investigation showed the potential therapeutic benefits of anthocyanins in restoring the Th17/Treg balance in autoimmune diseases.
Kaempferol
Kaempferol, a naturally occurring edible flavonoid derived primarily from the rhizome of the Kaempferia spp. ginger plant, is found in various vegetables and fruits and is renowned for its anti-inflammatory, antioxidant, and anticancer properties.209,210 Current studies indicate that EOS are immunomodulatory cells that interact with T and B lymphocytes. GM-CSF, in particular, serves as a critical survival and activating factor for hematopoietic cells, supporting the maturation of macrophages, EOS, and neutrophils. It is recognized as a pleiotropic cytokine with pro-inflammatory effects. Hyun-Aoh et al conducted an experimental study on the effects of kaempferol in an allergic rhinitis mouse model by assessing spleen weights,182 spleen indices, as well as changes in serum and nasal mucosal tissues. The results revealed that kaempferol demonstrated anti-allergic efficacy by inhibiting levels of new mediators such as IL-32 and TSLP, along with major mediators like IgE. Additionally, it was hypothesized that kaempferol could down-regulate histamine production while reducing inflammation-related protein levels and diminishing the infiltration of inflammatory cells. As a result, it may regulate both local and systemic allergic reactions while improving clinical allergy symptoms.
Quercetin
Quercetin is present in various parts of plants, including the stem bark, flowers, leaves, buds, seeds, and fruits. It mainly exists as glycosides and acts as a flavonol compound with a variety of biological activities.211 These activities encompass free radical scavenging antioxidant properties and antiallergenic effects that stimulate the immune system, exhibit antiviral activity, inhibit histamine release, and reduce pro-inflammatory cytokines.212 Extensive research has been conducted on quercetin’s role in multiple diseases from a pathogenesis perspective, and existing studies have shown that quercetin modulates Th1/Th2 and Treg/Th17 cytokine levels to alleviate AR pathogenesis.213 Peng et al demonstrated that quercetin can restore the cytokine balance of Th1/Th2 by activating the release of IL-2 and IFN-γ in Th1 cells and inhibiting the abnormal secretion of IL-4 and IL-5 in Th2 cells.183 Additionally, it can regulate Th1/Th2 cytokines at the gene level by suppressing the expression of Tim1 and Tim3 in the nasal mucous membrane tissues to change the original immune balance. Xia Ke et al demonstrated that quercetin reduced the serum levels of IgE,184 IgG1, and histamine, while increasing the level of IgG2. It also increased the percentage of Th17 cells, IL-10, and FOXP3, as well as elevated levels of IL-17, transforming growth factor-β, IL-6, and tumor necrosis factor-α. In addition, quercetin down-regulated COX-2, p-Tim3, and p-Tim2 cytokines in ovarian cancer cells, and it was also found to inhibit the expression of Tim1 and Tim3 in nasal mucosal tissues at the gene level to regulate Th1/Th2 cytokines and alter the original immunological balance state. Quercetin inhibited COX-2 expression along with p-IkBα and nuclear p65 expression, but upregulated ovalbumin-induced IkBαand cytoplasmic-65 levels, suggesting its inhibition on the nuclear transcription factor-κB pathway. The anti-allergic aspect of quercetin has been demonstrated by many scholars. As a plant extract, it serves as the main ingredient in many potential anti-allergic drugs, supplements, and fortified products. MisakoKashiwabara et al conducted a study to explored the modulatory effects of quercetin on neuropeptides from a neuroimmunological perspective. They administered quercetin through gavage using the distilled water purification technique.185 Their findings revealed that modulation of quercetin administration concentration inhibited changes in axon terminal calcium ion content after nasal stimulation. This inhibition subsequently reduced the release of neuropeptide substance P, calcitonin gene-related peptide, and nerve growth factor from nasal lavage fluid. Moreover, it simultaneously inhibited eosinophil degranulation and altered the ability of EOS to produce chemokines while modifying allergic responses. In a recent study, Amane Otaki et al utilized TNF-α derived from human nasal epithelial cells (HNEPCs) to stimulate the nasal mucosa of SD rats.186 The observation showed that quercetin enhances the upregulated ability of nasal cells to produce CC10, which effectively inhibits the chemotactic effects of phospholipase A2 and inflammatory cells, while downregulating the differentiation of Th2 cells. Consequently, this blocks allergen induction and reduces the inflammatory response, thus controlling the severity of AR. In addition, Nachi Ebihara et al187 employed nanotechnology to demonstrate that quercetin significantly inhibited IL-4-induced NO production in nasal mucosal epithelial cells by modulating their immune function. This inhibition is achieved through blocking STAT6 activation and iNOS mRNAs expression, leading to improved clinical symptoms of AR.
Baicalein
Baicalin, an active ingredient in the dried root extract of the traditional Chinese medicine plant Scutellaria baicalensis, is an antiviral, anti-inflammatory, and antiplatelet-coagulant agent with proven therapeutic efficacy in a variety of inflammation-induced diseases.214 In addition, baicalein can inhibit the generation of ROS by increasing the expression of Nrf2 protein. This protein plays an antioxidant role and inhibits oxidative damage; at the same time, it acts as an antagonist of 12-LOX’s specific target and reduces the programmed death of neuronal cells. It also attenuates overexpression of c-jun protein, and modulates the inflammatory state in AR distress syndrome, and mitigates response to oxidative stress.215 In animal cell experiments conducted by Yun-jiang Zhou, it was found that baicalin inhibited compound 48/80-induced release of histamine and b-hexosidase from HMC-1 MCs,188 demonstrating its protective effect on metamorphosis. Furthermore, it targeted the JAK2-STAT5 signaling pathway and inhibited the activation of JAK2, which subsequently suppressed the phosphorylation of downstream molecule STAT5. Consequently, STAT5 translocated to the nucleus where it regulated transcription of target genes encoding inflammatory mediators. These downstream molecules also translocated to the nucleus, regulated the transcription of target genes encoding inflammatory mediators, and blocked the production of IL-1β, IL-6, IL-8, and TNF-α by lipopolysaccharide (LPS)-stimulated HMC-1 MCs. In addition, baicalin was found to markedly attenuate the activation of IKKB and inhibit the phosphorylation of IkBɑ in the cytosol, thereby blocking nuclear translocation and suppressing NF-kB signaling in the lipopolysaccharide-stimulated HMC-1 MCs. This leads to inhibition of inflammation mediator production. Saizhen Chen et al, analyzed from a metabolomics perspective,189 discovered that baicalin intervention resulted in the alignment of the nasal mucosa and the reduction of eosinophil infiltration, as well as the inhibition of immunoglobulin E (IgE), histamine, IL-1β, IL-4, IL6, and TNF-α, which plays a protective role in preserving the integrity of the nasal mucosa. Jing Li et al through the human nasal mucosa and animal models, observed that baicalin decreased IL-17A expression,190 increased Foxp3 expression, reduced Beclin1 and Lc3-II/Lc3-I expression levels while increasing p62 expression level. They also found alterations in Treg/Th17 cell ratio due to baicalin treatment, leading to inhibition of Treg/Th17 cells’ activity. Meanwhile, they found that baicalin inhibited the release of toxic protein ECP from EOS cells, down-regulated cellular autophagy levels, which promoted the proliferation of Th17 cells while inhibiting Treg cell growth. These effects improved AR immune system response and controlled AR incidence. Liu Tao et al investigated the STAT3 signaling pathway and determined that baicalin inhibited p-STAT3 expression at inflammatory sites to reduce the secretion of serum IL-6, IL-1β and TNF-α.191 Also, they observed attenuated splenic injury during inflammation with baicalin treatment. In a recent study, Hu Qing et al,192 found that baicalein inhibited the activation of NF-κB/STAT3/ERK signaling pathway proteins, reduced the phosphorylation levels of ERK, NF-κBp65, and STAT3, inhibited inflammatory cell activity, alleviated inflammatory responses, accelerated the recovery of nasal mucosal tissues in mice, and improved the symptoms of AR.
Tanshinone
Tanshinone is a fat-soluble constituent of the dried roots and rhizomes of Salvia miltiorrhiza, which belongs to the Labiatae family. It exhibits a wide range of biological activities, with tanshinone being the most active fat-soluble constituent of Salvia miltiorrhiza. It possesses antioxidant, anti-inflammatory, anti-angiogenic, and anti-tumor properties.216 Recent domestic and international studies have also shown that tanshinone TIIA regulates the development, activation, and function of immune cells. It can participate in both innate and acquired immune response. TIIA promotes the reverse migration of neutrophils by inducing apoptosis and accelerating their loss of sensitivity to chemotaxis environments. This facilitates inflammation resolution and tissue repair. Additionally, it significantly reduces lipopolysaccharide-induced inflammation and tissue repair damage by reducing lipoprotein (TI) levels.217 In mice with acute lung injury induced by lipopolysaccharide, it effectively decreases neutrophil infiltration, myeloperoxidase activity, expression of inflammatory cytokines in bronchoalveolar lavage fluid while mitigating inflammatory factor-induced lung tissue damage. Earlier studies have shown that macrophages can be induced to polarize into the M1 type by up-regulating the P-STAT1/NF-KB signaling pathway, whereas TIIA effectively activates P-STAT6 phosphorylation and induces macrophages to polarize into the M2 type, thereby reducing the inflammatory response.218 It has been found that IgE-induced MC activation is negatively regulated by the Sirt1.LKB1.AMPK pathway. In vitro and ex vivo experimental studies have revealed that TIIA could inhibit IgE receptor (FcεRI)-mediated MC activation in vitro and MC-mediated allergic responses in vivo by activating the Sirt1.LKB1.AMPK pathway. Therefore, TIIA may become a new drug for allergic diseases mediated by MCs.219 Currently, it is believed that allergen-induced IgE antibodies binding to high-affinity FcεRI on the surface of MCs activate them, which triggers an intracellular signaling cascade reaction. Thus, targeting MCs may hold potential for treating AR. Huang Fuse et al investigated the effect of TIIA on MC-mediated AR through in vivo and ex vivo experiments.193 The results showed that TIIA could alleviate AR responses such as histamine release, IgE production, and vasodilatation in model mice. Additionally, it inhibited the degranulation of MCs and suppressed the activation of the NF-κB pathway to down-regulate the expression of inflammatory factors TNF-α and IL-4. These findings suggest that TIIA alleviates MC-mediated AR responses by inhibiting the activation of the NF-κB pathway and prevents immune diseases by regulating dendritic cell maturation. Research has also found that TIIA can slow down autoimmune diseases by inhibiting DC function and maturation, thereby affecting T-cell differentiation and leading to increased secretion of anti-inflammatory factors and decreased secretion of inflammatory factors in the body. TIIA can weaken the development of immune diseases by inhibiting DC-mediated adaptive immunity and acquired immunity. TIIA can regulate T lymphocytes in various immune diseases by inhibiting the activation of the NFκB pathway, thereby alleviating MC-mediated AR. TIIA is involved in the immune mechanism of many diseases by regulating T lymphocytes. By inhibiting inflammation, TIIA can regulate the balance of Th1/Th2 by significantly downregulating the levels of Th1 cytokines and upregulating the expression of Th2 cytokines in mice, thus regulating the balance of Th1/Th2. TIIA increases the ratio of CD3+, CD4+, and CD8+ T-lymphocyte subsets while decreasing inflammatory cytokine levels (IL-2, IL-4, INF-γ, TNF-α), and it also enhances the expression of anti-inflammatory cytokine IL-10 to boost immune response. In addition, Shouye Li found that Tan IIA could effectively inhibit the activation of the PLCγ1/PKC/IP3R pathway induced by IgE/Has in the nasal mucosa of mice with follicle-stimulated AR.194 It downregulated the expression of downstream inflammatory factors IL-4, IL-5, IL-1β, and α, as well as decreased intracellular calcium ion release and MC degranulation. Additionally, it inhibited tissue swelling, small blood vessel dilation, glandular hyperplasia, and acid granulocyte infiltration in mice. In a recent study conducted by Qing Chen et al, TIIA significantly reduced histamine,195 OVA-IgE, and OVA-IgG1 levels while inhibiting Th2 cytokine release (TNF-α, IL-4, and IL-5) in OVA-induced AR mice. Furthermore, it counteracted C48/80-induced histamine release and promoted MC degranulation. These findings suggest that TIIA could be used as a treatment for allergic diseases.
Lignan
Lignan is a natural compound of plant origin, typically a flavonoid, which is considered an effective anti-allergic component of Perilla frutescens. It plays an important role in various aspects of anti-inflammatory, antioxidant, and immune function enhancement by altering signaling pathways such as NF-κB, MAPK/AP-1, and JAK-STAT.220 Early studies have shown that it can improve the Th2 response in patients with allergic diseases by lowering the levels of IL-4, IL-5, and IL-13, with less infiltration of inflammatory cells in the upper respiratory tract (nasal mucosa).196 In addition, lignans can inhibit the release of mediators like leukotrienes and the expression of intercellular adhesion molecules induced by tumor necrosis factor-α in endothelial cells. They also inhibit the nuclear transcription factor-κB pathway and its related genes. The study conducted by Kai-Li Liang et al demonstrated that lignans effectively alleviate allergic symptoms,197 reduce serum IgE levels and mucus production, as well as suppress local and systemic IL-4 production in allergy-experienced mice. These findings suggest that lignans possess the ability to inhibit both TH2 polarization and TH2 cytokine production in human peripheral blood mononuclear cells (PBMCs); furthermore, lignans exhibit inhibitory effects on the release of mediators, such as tumor necrosis factor-α, from endothelial cells, along with their ability to suppress the nuclear transcription factor-κB pathway and its associated genes. Moreover, lignocaine treatment inhibited the IL4/STAT6/GATA3 signaling pathway in naïve CD4+ cells, leading to reduction in the expression of pSTAT6 and GATA3 and thereby alleviating nasal allergy symptoms. Lignans also inhibited ICaCC in CALU-3 cells, which are responsible for calcium-activated chloride currents but not CFTR currents in human respiratory epithelial CALU-3 cells. Additionally, inhibition of ANO1 activity reduced electrolytic secretion at the parietal membrane and helped alleviate the hypersecretory state of the nasal cavity in individuals with AR. Jin Huidong et al, in their study targeting the TLR4/NF-κB pathway,198 demonstrated that lignans could restore the imbalance of Th1/Th2 cytokines and ameliorate nasal mucosal inflammation by promoting the down-regulated levels of Th1-type cytokines IFN-γ and IL-2, while inhibiting the up-regulated Th2-type cytokines IL-4, IL-5, IL-13, OVE-sIgE, TLR4, and p65. Yuping Yang et al, from an AR pathogenesis perspective,199 found that it could enhance anti-OVA-IgE levels, autophagy-related factors (Beclin1, LC3II/LC3I), helper T-cells IL-17A expression, as well as flavic acid receptor-associated orphan nuclear receptor γt (RORγt) expression. These factors were shown to regulate h17 and Treg cell percentages while suppressing anti-OVA-IgG2a levels along with IL-10 and Foxp3 expression to improve inflammation levels. In a recent study, Wang Hui-Hui et al found that lignocaine intervention resulted in reduced eosinophil infiltration,200 orderly epithelial arrangement, decreased nasal mucosal cell exfoliation, and suppression of P-PI3K/PI3K, P-AKT/AKT, and mTOR protein levels. This suggests that it may reduce IL-4, IL-13, and OVA-sIgE by down-regulating PI3K/AKt/mTOR pathways. Furthermore, it reduces IL-4, IL-13, and OVA-sIgE levels, while significantly decreasing Th2 levels and GATA-3 protein levels. Conversely, it increases Th1 levels and T-bet protein level. These findings suggest that Lut may alleviate AR by suppressing inflammation through the regulation of Th1/Th2 balance.
Naringenin
Naringenin, a natural flavonoid derived from the seed coats of plants in the Lacertaceae family, exhibits anti-inflammatory, antioxidant, and antitumor properties. It is involved in regulating numerous metabolic and signaling pathways, including nuclear factor and inflammatory vesicle activation pathways. Ongoing research has increasingly brought naringenin to public attention in recent years.221,222 Earlier studies have demonstrated that naringenin enhances immunity and anti-inflammatory capacity by suppressing both inflammation and T-cell-mediated immune responses.223 Utilizing MC degranulation models and network pharmacological techniques, Yixuan Niu et al found that naringenin modulates AR through multiple targets (eg, AKT1, MAPK3, VEGFA) and pathways (eg, cancer, VEGF signaling).224 It significantly reduces P815 MC degranulation, increases IL-4 levels, decreases tumor necrosis factor-α levels, and improves inflammatory responses in AR. Furthermore, Abdulkadir Şahin et al discovered that naringenin improves the condition of damaged nasal mucosa in sensitized rats by realigning nasal mucosal epithelial cells,201 reducing inflammatory cell infiltration, and lowering serum total IgE levels as well as IL-4 and IL-5 levels to modify the inflammatory state. In cell experiments conducted by Niu Jinming et al, the TLR4/NF-κB/TNF-α signaling pathway was observed. It was demonstrated that naringenin inhibits the nuclear translocation of NF-κBp65 and suppresses TNF-α expression by downregulating TLR4 expression. This inhibition led to a reduction in the production of inflammatory mediators, such as IgE, and decreased the release of IL-17 and IL-18. Furthermore, naringenin attenuated CD1, CD9, and CD23 expression in the nasal mucosa, thereby interfering with the inflammatory response and mitigating pathological damage and symptoms in rats.202
Hesperidin
Hesperidin, a bioactive compound found in Chenpi, has demonstrated multiple pharmacological effects, including anti-cancer, cardiovascular and neuroprotective properties, anti-allergic asthma activity, anti-inflammatory actions, and immune regulation.225 In a previous study conducted by Korhan Kilic et al on OVA-sensitized AR rats,203 it was observed that hesperidin reduced serum levels of Ig-E, IL-5, IL-13, and total oxidative status (TOS), decreased the expression of TNF-α while enhancing the release of total antioxidant capacity (TAC). These findings underscore the antioxidant and anti-inflammatory characteristics of hesperidin along with its ability to regulate oxidative stress and improve allergic reactions. Li Wei et al investigated the signaling pathways influenced by hesperidin204 and noted its potential to regulate the Th2 immune response in AR. Hesperidin improved the pathological conditions of nasal mucosa and alleviated allergic reactions in mice by inhibiting the expression of TSLP, GATA3, and OX40L proteins and blocking the activation of the TSLP/OX40L/OX40 signaling pathway.
Polydatin
Polydatin, extracted from its rhizome, is a natural herbal component known for its anti-inflammatory, antioxidant, and antimotile effects. Reports suggest that polydatin has the potential to effectively treat allergic inflammatory diseases by inhibiting allergic inflammatory responses through targeting the Nrf2/HO-1 pathway, modulating the inflammatory responses of nasal mucosal epithelial cells, and reducing the release of inflammatory factors.226 In a study on mitochondrial autophagy conducted by Liu Siqi et al, it was observed that polydatin suppressed the expression of pro-apoptotic proteins (Bax, caspase-3, cleaved-caspase-3, and Cytochrome C) while enhancing the expression of anti-apoptotic protein Bcl-2 in AR model mice.205 This regulation resulted in upregulated mitochondrial kinases PINK1 and E3-ubiquitin ligase Parkin activation, leading to improved nasal mucosa thickness reduction in eosinophil secretion and alteration of allergy symptoms. Gao Qiong et al,227 in their study on oxidative stress responses, found that polydatin reduced serum IgE levels and pro-inflammatory factors (IL-6, TNF-α, IL-17), while increasing IL-10 levels and regulating the Th1/Th2 balance. Additionally, significant upregulation of polydatin and Claudin-1 in nasal mucosal tissues suggested enhanced barrier function and mucosal remodeling. Increases in SOD and CAT activities, along with decreased MDA levels, indicated a reduction in oxidative stress. The mode of action was linked to the modulation of the cellular pyroptosis pathway induced by ROS/NLRP3/Caspase-1, thereby alleviating the disease in AR model mice.206
Saponins (Table 3)
Ginsenoside
Ginsenoside, a ginseng extract, exhibits anti-inflammatory, immunomodulatory, anti-tumor, and anti-viral effects. Ginsenosides can be classified into three main types: proto-ginsenoside diols (eg, Rb1, Rb2, Rc, Rd, Rh2), proto-ginsenoside triols (eg, Re, Rf, Rg1, Rg2, Rh1), and oleanolic acid derivatives (eg, Ro, Rh3).235 Oh et al investigated the therapeutic effects of ginsenoside Rg1 using an ovalbumin-induced AR mouse model.228 They found that Rg1 potentially mitigates inflammation by decreasing levels of factors such as TSLP, IL-1β, IL-4, histamine, IgE, IgG1, EOS, and MC infiltration, while also inhibiting caspase-1 activity, receptor-interacting protein 2, IκB kinase-β, and NF-κB/RelA expression in activated human MCs-1 (HMCs-1), thereby alleviating allergy symptoms. Utilizing transcriptomics and metabolomics, Liu et al demonstrated that ginsenoside Rg3 exerted protective effects in mice by reducing the levels of IL-4, IL-13, IgE, IL-5, and MDA while increasing superoxide dismutase (SOD) in serum.229 Moreover, Rg3 influenced the expression of 12 genes and 8 metabolites, enhancing the protein expression of Nrf2 and HO-1 in the nasal mucosa of AR mice. In contrast, ML385 significantly inhibited the regulatory effects of the Nrf2/HO-1 pathway on oxidative stress modulation and protection in AR mice.
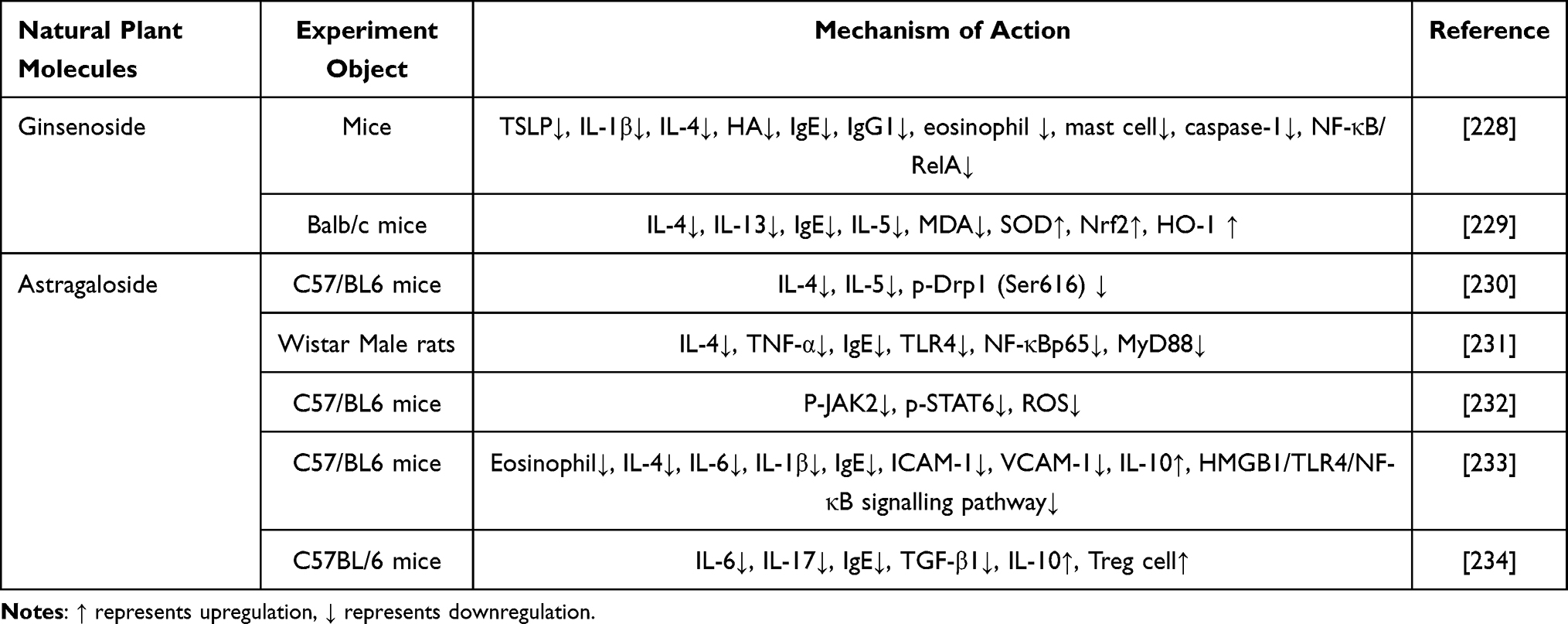 |
Table 3 The Immunomodulatory Effects and Mechanisms of Saponins From Natural Plants on AR Were Summarized |
Astragaloside
Astragaloside, a highly purified compound extracted from the legume species Astragalus membranaceus and Astragalus myrtillus, is renowned for its potent biological activity. It exhibits more than twice the effectiveness of standard astragaloside and over thirty times greater potency than Astragalus polysaccharides in terms of antiviral effects, despite its lower content. Astragaloside exhibits a wide range of pharmacological effects, including anti-inflammatory, anti-fibrotic, and immunomodulatory properties.230,236 In mitochondrial research conducted by Chen Xuqing et al, it was found that astragaloside reduced serum IL-4 and IL-5 levels.237 Furthermore, in OVA-sensitized mice models, it decreased the ratio of NKp46 cells expressing IL-4 and p-Drp1 (Ser616). Additionally, astragaloside inhibited NK2 differentiation in nasal mucosa NK cells, which restored mitochondrial morphology and substantially increased mitochondrial length. As a result, this alleviated nasal allergy symptoms as well as mucosal inflammation in mice. Further research by Ma Wenling on the mechanisms of astragaloside in treating AR revealed its ability to inhibit the TLR4/MyD88/NF-κB signaling pathway.231 In addition, it was found that astragaloside could modulate Th2 cytokine release and reduce pro-inflammatory factors such as IL-4 and TNF-α, thereby improving symptoms in AR rats. Observations of nasal mucosa tissue showed a more intact structure of the nasal mucosa epithelium in AR rats, with less shedding of cilia, fewer abnormal cup cells, and reduced eosinophil infiltration. Studies on nasal mucosa and spleen tissues in AR mice demonstrated that astragaloside intervention led to reduced levels of p-JAK2 and p-STAT6-positive cell counts, as well as ROS levels. Furthermore, it downregulated the ratios of p-JAK2/JAK2 and p-STAT6/STAT6, thus suppressing the JAK2/STAT6 signaling pathway and ameliorating AR inflammation.232 These findings suggest a potential target for precise treatment strategies for AR. Ying Tian et al discovered that a dose of 50.0 mg/kg of astragaloside modulated the HMGB1/TLR4/NF-κB signaling pathway,233 resulting in decreased protein expression and levels of inflammatory factors such as IL-4, IL-6, IL-1β, IgE, TNF-α, ICAM-1, and VCAM-1, while increasing IL-10 concentrations. From the perspective of T cell pathogenesis, Zhang Fei et al observed that astragaloside reduced the expression of IL-17, IL-6, and IL-1β in peripheral blood T17 cells and inhibited the proliferation of effector T cells through cytokine activation (eg, IL-10, TGF-β), thereby maintaining Th17/Treg cell balance and reducing AR severity.234
Monoterpenes (Table 4)
Glycyrrhizic Acid
Glycyrrhizic acid, a pentacyclic triterpene derivative, is a major active component of licorice with pharmacological effects that include anti-inflammatory, anti-allergic, and antioxidant activities. It inhibits oxidative stress and ROS release, reduces pro-inflammatory cytokine levels, and exhibits corticosteroid-like effects.250 Early studies have indicated that GA suppresses Th2-type cytokines (IL-4, IL-5, IL-13) and up-regulates the Th1-type cytokine interferon-γ (IFN-γ) in OVA-sensitized asthma and AR mouse models. This mechanism helps maintain a balance Th1/Th2 responses while mitigating inflammatory cell infiltration and airway inflammation.251 18 β-Glycyrrhetinic acid, an important metabolite derived from glycyrrhizic acid through hydrolysis, is also a pentacyclic triterpene with notable anti-inflammatory and antioxidative effects.238 Zhou Liuqing et al observed that glycyrrhizic acid reduced intestinal metaplasia effects by decreasing eosinophil counts in the intestinal walls of AR model rats.252 Dong Ming et al demonstrated that glycyrrhizic acid improved mitochondrial count,253 reduced cristae degradation, and decreased swelling, vacuolization, and autophagosome appearance in nasal mucosal epithelial cells. Research has demonstrated that 18β-Glycyrrhizic acid (18β-GA) can delay or even reverse pathological changes in nasal mucosal cilia,239 reduce OVA-sIgE levels, and inhibit the release of inflammatory mediators such as IL-1β, IL-6, IL-8, and TNF-α in AR rats, thereby alleviating allergy symptoms. Furthermore, 18β-GA downregulates proteins like CCL11, AQP1, AQP5, and TSLP in the nasal mucosa,240 thereby attenuating inflammation and allergic reactions. It also decreases MUC5AC expression to restore normal function of the ciliary transport system.241 Additionally, 18β-GA ameliorates ER expansion, vacuolization, and degranulation in the nasal mucosal cells of AR model rats.254 Research has investigated the mechanisms underlying AR and identified a direct involvement of the p50 subunit in Th2 cell differentiation.255 By inhibiting NF-κB activity and downregulating p50 expression, 18β-GA limits Th0 to Th2 differentiation while reducing IL-4 levels. It also suppresses transcription of genes associated with inflammatory factors via NF-κB inhibition, thus mitigating the inflammatory response in the nasal mucosa and demonstrating anti-inflammatory effects.256 In a pathway study, 18-β-glycyrrhizic acid sodium ameliorated nasal mucosal tissue lesions in young AR rats.242 It inhibited the secretion of IL-2 and IFN-γ, reduced the levels of IgE, IL-4, and IL-5, and restored Th1/Th2 cytokine ratios. Furthermore, it down-regulated ICAM-1 and AQP5 protein expression while inhibiting the activation of NF-κB and PI3K-AKt signaling pathways, demonstrating a significant protective efficacy in young AR rats. This treatment also conferred protective effects against AR in young mice.
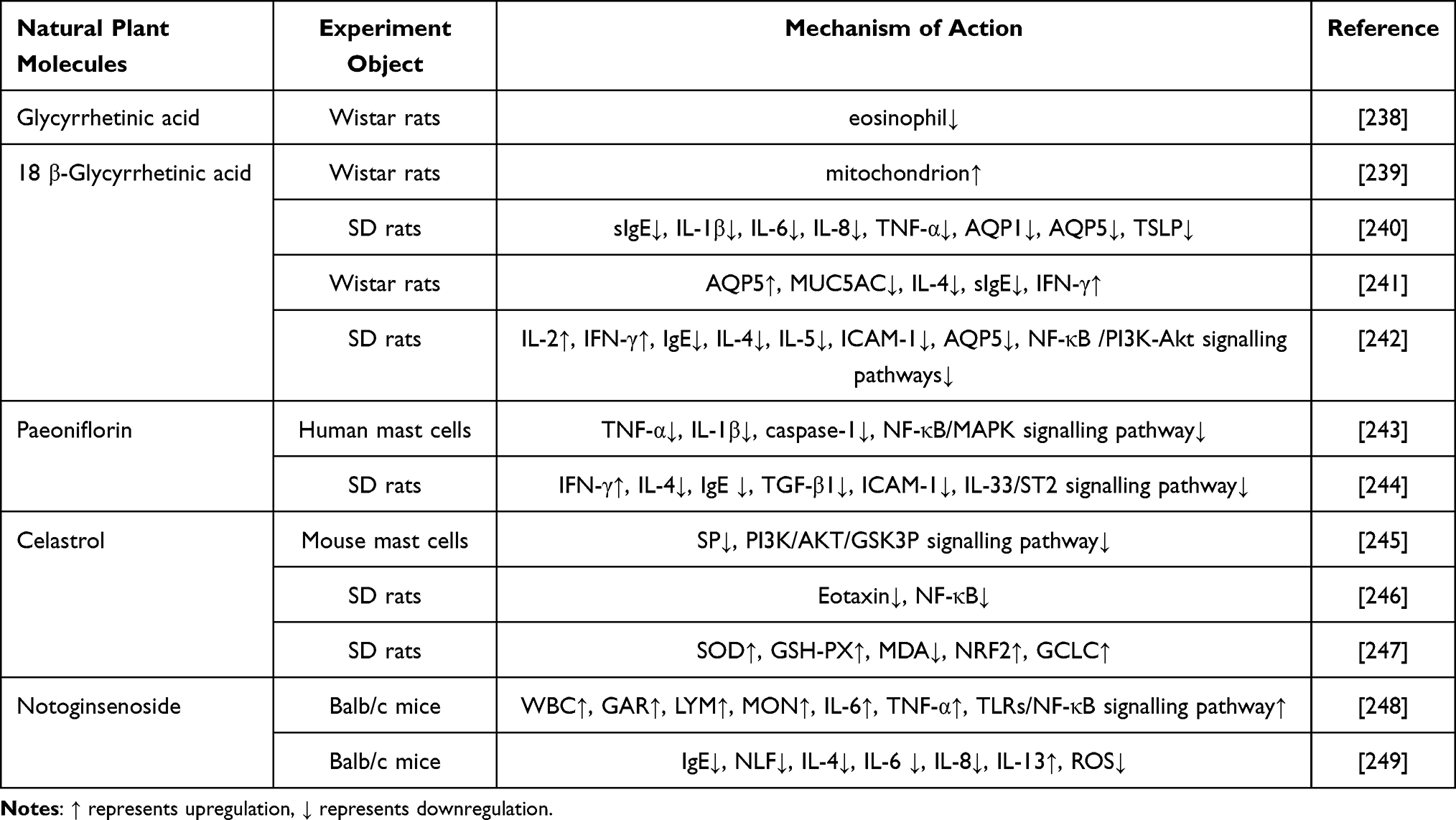 |
Table 4 The Immunomodulatory Effects and Mechanisms of Monoterpenoid Natural Plant Molecules on AR Were Summarized |
Paeoniflorin
Paeoniflorin, a monoterpene glycoside, is the primary bioactive compound found in Paeonia lactiflora. It is well-known for its anti-inflammatory, immunomodulatory, antioxidant, hepatoprotective, and lipid-lowering properties.257 In initial studies conducted by Guanghui Wang et al, it was observed that paeoniflorin effectively reduced the production of tumor necrosis factor-α and IL-1β by inhibiting the NF-κB/MAPK signaling pathway in PMACI-induced HMC-1 cells.243 This inhibition gave rise to a reduction in MC-mediated allergic inflammation and improvement in clinical symptoms. Furthermore, recent research conducted by Li Feng et al demonstrated that paeoniflorin modulated cellular immunity through the inhibition of the IL-33/ST2 pathway,244 leading to increased levels of IFN-γ and decreased levels of IL-4 and IgE. These actions also contributed to reduced expression of TGF-β1 and ICAM-1, thereby attenuating AR pathology and promoting nasal mucosal health.
Celastrol
Celastrol, a pentacyclic triterpenoid monomer, is an active ingredient itself, appearing as red crystals that are chemically stable and physiologically active in vivo. It is insoluble in water and possesses immunomodulatory and anti-inflammatory effects.258 Studies have shown that celastrol regulates the PI3K/AKT/GSK3β pathway to inhibit substance P-induced MC degranulation,245 thereby enhancing the survival of sensitized MCs, reducing cellular death, boosting immune response, and modulating symptoms of body metamorphosis. Li Ying et al investigated an OVA-sensitized AR rat model246 and discovered that celastrol modulates eotaxin expression by inhibiting NF-κB in the nasal mucosa, subsequently reducing EOS aggregation and activation to mitigate allergic pathogenesis. Cui Jing et al reported that pretreatment with celastrol increased SOD and GSH-PX levels,247 decreased MDA levels in the respiratory zone of nasal mucosa, elevated nuclear NRF2 expression and plasma GCLC protein levels, thus enhancing GSH content and antioxidant capacity of nasal mucosa in AR rats, while significantly improving the symptoms of metamorphic reactions.
Notoginsenoside
Notoginsenoside, the predominant chemical component in Panax ginseng, is primarily a dammarane-type tetracyclic triterpenoid saponin, comprising up to twelve percent of the plant’s content. It is known for its involvement in immune regulation, antitumor activity, antioxidant and anti-inflammatory effects, as well as cardiovascular protection.259,260 Gao Min’s initial study on cyclophosphamide-induced immunosuppressed mice demonstrated that notoginsenoside enhances immune cell defense by activating peripheral blood immune cells (WBC, GAR, LYM, MON),248 promoting lysozyme and cytokine (IL-6, TNF-α) secretion, and stimulating lymphocyte proliferation. Furthermore, it activates and regulates the TLRs/NF-κB signaling pathway by targeting proteins such as TLR-2, TLR-4, TRAF-6, TAK-1, IKKα, NF-κBp65, thereby enhancing macrophage vitality and phagocytosis, while increasing expression of NO, IL-6, and TNF-α, resulting in improved local and systemic immunoregulation. In a recent study, Zhang Yalin et al discovered that notoginsenoside reduced mitochondrial fission,261 inhibited serum OVA-specific IgE levels, and decreased cytokine (IL-4, IL-6, and IL-8) production in nasal lavage fluid (NLF). It also suppressed the expression of mitochondrial fission proteins DRP1 and ROS, hence reducing mitochondrial fission by inhibiting the TXNIP/NLRP3 signaling axis, enhancing mitochondrial integrity, and alleviating AR symptoms.
Other (Table 5)
Ephedrine
Ephedrine, an alkaloid derived from various ephedra plants, functions as a classical α- and β-adrenergic agonist, stimulating the central nervous system, elevating blood pressure, and dilating bronchi.266 Research has indicated that ephedrine modulates inflammation by upregulating the anti-inflammatory cytokine IL-10 while downregulating pro-inflammatory cytokines IL-1β, IL-6, IL-12, and TNF-α through the PI3K/Akt signaling pathway.249 In a mouse model of lipopolysaccharide-induced endotoxic shock, ephedrine was shown to promote immune homeostasis and exert anti-inflammatory effects by balancing cytokine production via TLR4 signaling. Recent studies have emphasized the enhanced therapeutic effects of combining ephedrine with other medications for the treatment of AR.267 In 1995, research revealed that ephedrine elicits a vasoconstrictive effect on nasal mucosal vessels, thereby optimizing the inspiratory flow rate.268 Additionally, Yang Ling et al observed that ephedrine suppresses COX2 expression in the nasal mucosa and downregulates mRNA and protein levels of thymic stromal lymphopoietin (TSLP) and OX40L, thus modulating the Th2 immune response in AR rats via the TSLP/OX40L pathway.262 This intervention significantly attenuated serum levels of IgE, IL-4, and IL-13, consequently modulating the inflammatory response in the nasal mucosa.
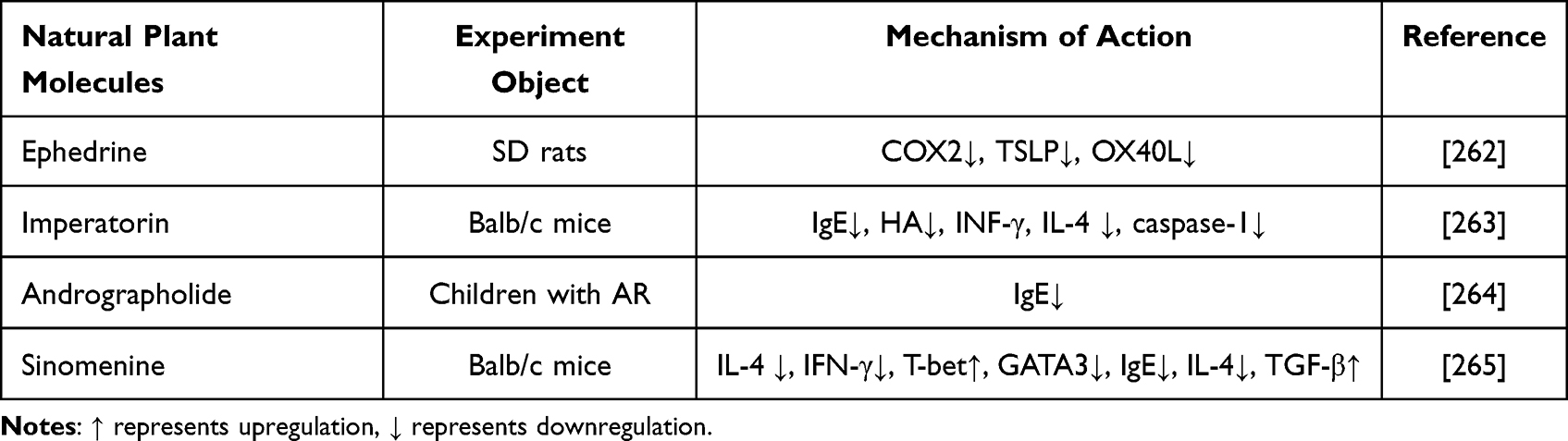 |
Table 5 The Immunomodulatory Effects and Mechanisms of Other Natural Plant Molecules on AR Were Summarized |
Imperatorin
Imperatorin is a principal pharmacological component in commonly utilized traditional Chinese medicines such as Serpentine, Angelica dahurica, Fenghuang, Angelica sinensis, Ming Dangshen, and Duhuo. It exhibits a broad spectrum of pharmacological effects, including analgesic, anti-inflammatory, antibacterial, and anti-tumor properties.269 Research has shown that imperatorin effectively inhibits MC activation and degranulation, thereby exerting a therapeutic effect on allergic inflammation.270 Imperatorin has been shown to significantly inhibit the degranulation of RBL-2H3 cells and reduce the release of histamine and various inflammatory markers, including IL-3, IL-4, IL-6, TNF-α, and COX-2, while promoting the expression of IFN-γ, thus modulating the cellular inflammatory response.271 In both in vivo and in vitro experiments using an OVA-induced AR mouse model, Hyun-AOh et al observed that ouabain effectively attenuated the elevated levels of IgE and histamine, while concurrently increasing interferon-γ levels.263 In addition, it substantially reduced the protein expression of IL-1β, macrophage inflammatory protein-2, intercellular adhesion molecule-1, and cyclooxygenase-2. Furthermore, imperatorin was found to inhibit the activation of MC, receptor-interacting protein 2 (RIP2), IκB kinase (IKK-β), NF-κB, and caspase-1, consequently leading to a significant reduction in IL-4 levels within splenic tissues. This mechanism ultimately enhances the body’s anti-inflammatory response.
Andrographolide
Andrographolide, a natural occurring active compound, is abundant in the plant Andrographis paniculata. It demonstrates potent anti-inflammatory and antioxidant properties by inhibiting the inflammatory response through blocking IKKγ phosphorylation, thereby preventing NF-κB activation and its subsequent nuclear translocation.272,273 In a study conducted by He Li et al, it was found that the administration of Andrographolide drops combined with Danxi Yupingfeng granules effectively reduced elevated serum levels of total immunoglobulin (Ig)E while also regulating the Th1/Th2 ratio to enhance immune function.264
Sinomenine
Sinomenine, a natural phytochemical derived from Cymbopogon, has been shown to possess anti-inflammatory and immunosuppressive properties.274 Initial studies have indicated that sinomenine inhibits the production of Th1 and Th2 cytokines, such as IL-4 and INF-γ, while simultaneously enhancing T-bet expression and attenuating GATA3 expression.275 This modulation alters the balance between Th1 and Th2 cells in the body, thereby regulating the overall Th1/Th2 equilibrium. Zhechen et al investigated the immunosuppressive effects of sinomenine in an OVA-induced AR model.265 Their findings revealed that this treatment reduced eosinophil infiltration, cuprocyte proliferation, and ciliated cell loss in the nasal epithelium. Additionally, it inhibited the expression of OVA-specific IgE and IL-4 levels while enhancing TGF-β secretion,276 ultimately regulating the expression of T-bet-a transcription factor critical for Th1 and Th2 differentiation, resulting in decreased overall inflammation along with an enhanced capacity for immune regulation.
Conclusion
Natural plant molecular sources of a wide range of medicinal ingredients mentioned above widely exist in the daily diet of vegetables and fruits. However, they are most abundant in many plants and herbs, and their extraction processes are more advanced. With the continuous advancement of research and the progress of science and technology, previously extracted components from food and medicinal herbs can now be isolated and purified into various forms such as volatile oils, capsules, and liquid injections, enabling diverse routes of administration into the human body. This facilitates enhanced absorption into the bloodstream, achieving optimal drug concentrations while minimizing inefficient utilization by the body’s metabolic processes. Although natural plant analysis has been extensively explored and studied, numerous undiscovered plant molecules persist due to technological limitations and limited understanding. For comprehensively investigated plant molecules, in-depth research should be conducted throughout disease onset and progression stages. Currently, traditional Chinese herbal medicine, particularly the focus of current research, exhibits significant potential in modern investigations into its active components. For thousands of years, the therapeutic effects of these natural products have been recognized through human medical experience, encompassing the use of natural animals and plants for medical treatment and diet therapy. Returning to their origins, our present emphasis lies on these plants and animals, especially herbs. These medicinal materials not only come from diverse sources, but also possess a wealth of medicinal ingredients. Some are even referred to as natural antioxidants that serve as crucial sources for novel drug development and effective means for disease prevention and treatment. In recent years, an increasing number of natural plant molecules have been found to exert important effects on AR, with immune regulation being a key underlying mechanism. Previous studies have unequivocally demonstrated a close association between AR pathogenesis and immune regulation. Several studies have suggested that changes in immune regulation within the body may influence the onset and progression of AR in a given population.277 If this hypothesis holds true, targeting immune cells and factors could potentially enhance the management of AR. Despite extensive research efforts and experimental evidence supporting the underlying mechanisms, limited attention has been directed towards pharmacological interventions specifically aimed at modulating immunity. Consequently, recent focus has shifted towards immunotherapy, gut microbiota intervention, and physical exercise.278 However, it is important to note that these approaches necessitate longer durations of treatment and patience for optimal benefits. At present, extensive research has been conducted in the field of nanotechnology to achieve targeted drug delivery and improve drug bioavailability, leading to significant advancements in nanomedicine and intranasal dosing.279–281 Nonetheless, breakthrough results are still lacking, bringing about a gradual shift of focus towards natural molecules.282 Although there is a lack of specific reports on the immunomodulatory effects of these plant molecules mentioned above, our study on their treatment of AR reveals that they alter disease progression by modulating the body’s immune capacity. These results suggest their potential for treating AR. Therefore, using natural plant molecules to regulate AR from an immune regulatory perspective may provide a new way to interfere the progression of AR and enhance cognitive function. In conclusion, it is indisputable that natural plant molecules have strong targeting abilities as AR therapeutic drugs. They can effectively penetrate the blood-brain barrier and provide inherent advantages in maintaining their integrity while regulating their functionality. Moreover, they effectively suppress the secretion of inflammatory factors and cells, modulate the allergic reactions, and ameliorate the pathological characteristics of AR by anti-inflammatory, anti-allergic mechanisms, and signal pathway regulation. These effects have been substantiated in animal models and cell-based experiments. Nevertheless, comprehensive investigations on the potential toxicological side effects associated with these plant molecules are lacking. Despite their relative reliability, toxicological assessments remain imperative. Moreover, further validation of the multi-pathway targeting effects of natural plant molecules on AR necessitates subsequent clinical trials. Furthermore, it is imperative to investigate the synergistic effects of multiple plant molecules rather than focusing solely on individual compounds. In addition, comprehensive considerations such as pharmacology, pharmacokinetics and pharmacogenetics should be taken into account when addressing issues related to dosage forms and metabolic timing. This multifaceted task presents various challenges, including trial design, trial period definition, ethical review and technical financial support. Therefore, concerted efforts are necessary to optimize the biological activity of natural plant molecules for AR immunoregulation and develop more efficacious therapeutic drugs.
Data Sharing Statement
All data are available in the manuscript and they are shown in the figures and tables.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported in part by the following grant: General project of The Natural Science Foundation of Heilongjiang Province, (Project No. LH2019H504).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Okubo K, Kurono Y, Ichimura K, et al. Japanese guidelines for allergic rhinitis 2020. Allergol Int. 2020;69(3):331–345. doi:10.1016/j.alit.2020.04.001
2. Greiner AN, Hellings PW, Rotiroti G, Scadding GK. Allergic rhinitis. Lancet. 2011;378(9809):2112–2122. doi:10.1016/s0140-6736(11)60130-x
3. Siddiqui ZA, Walker A, Pirwani MM, Tahiri M, Syed I. Allergic rhinitis: diagnosis and management. Br J Hosp Med. 2022;83(2):1–9. doi:10.12968/hmed.2021.0570
4. Wise SK, Damask C, Greenhawt M, et al. A synopsis of guidance for allergic rhinitis diagnosis and management from ICAR 2023. J Allergy Clin Immunol Pract. 2023;11(3):773–796. doi:10.1016/j.jaip.2023.01.007
5. Vlastos IM, Kalentakis Z, Doulaptsi M, Karatzanis A, Prokopakis EP. Multimorbidities in allergic rhinitis-current evidence from epidemiological studies, treatment trials, and molecular data. Curr Allergy Asthma Rep. 2023;23(2):133–140. doi:10.1007/s11882-022-01063-w
6. Stern J, Chen M, Fagnano M, Halterman JS. Allergic rhinitis co-morbidity on asthma outcomes in city school children. J Asthma. 2023;60(2):255–261. doi:10.1080/02770903.2022.2043363
7. Melvin TA, Ramanathan M. Role of innate immunity in the pathogenesis of allergic rhinitis. Curr Opin Otolaryngol Head Neck Surg. 2012;20(3):194–198. doi:10.1097/MOO.0b013e3283533632
8. Klimek L, Pfaar O. Allergische rhinitis. Immunologische und neurogene mechanismen [Allergic rhinitis. Immunological and neurogenic mechanisms]. Hno. 2011;59(12):1191–1197. doi:10.1007/s00106-011-2294-z
9. Wang DY, Clement P. Pathogenic mechanisms underlying the clinical symptoms of allergic rhinitis. Am J Rhinol. 2000;14(5):325–333. doi:10.2500/105065800781329483
10. Bernstein DI, Schwartz G, Bernstein JA. Allergic rhinitis: mechanisms and treatment. Immunol Allergy Clin N Am. 2016;36(2):261–278. doi:10.1016/j.iac.2015.12.004
11. Zhang Y, Lan F, Zhang L. Advances and highlights in allergic rhinitis. Allergy. 2021;76(11):3383–3389. doi:10.1111/all.15044
12. Baraniuk JN. Mechanisms of allergic rhinitis. Curr Allergy Asthma Rep. 2001;1(3):207–217. doi:10.1007/s11882-001-0007-5
13. Cohen B. Allergic rhinitis. Pediatr Rev. 2023;44(10):537–550. doi:10.1542/pir.2022-005618
14. Sakevych VD, Kutsenko NL, Mykytiuk MV, Kaĭdashev IP. [Clinical course and characteristics of cellular and humoral immunity in patients with allergic rhinitis]. Lik Sprava. 2014;(1–2):15–20. Slovenian.
15. Lara MC, Montes y Montes J, Carro C, et al. Rinitis alérgica e inmunidad celular [Allergic rhinitis and cellular immunity]. Alergia. 1985;32(3):65–67.
16. Borregaard N. Current concepts about neutrophil granule physiology. Curr Opin Hematol. 1996;3(1):11–18. doi:10.1097/00062752-199603010-00003
17. Zawrotniak M, Juszczak M, Mosio-Wójcik J, Rapala-Kozik M. Neutrophil extracellular traps in upper respiratory tract secretions: insights into infectious and allergic rhinitis. Front Immunol. 2023;14:1295921. doi:10.3389/fimmu.2023.1295921
18. Haixia W. The Expression of TLR9 in Blood Neutrophils and Monocytes of Allergic Rhinitis and Asthma [Master]. 2023.
19. Matsui M, Nagakubo D, Satooka H, Hirata T. A novel Siglec-F(+) neutrophil subset in the mouse nasal mucosa exhibits an activated phenotype and is increased in an allergic rhinitis model. Biochem Biophys Res Commun. 2020;526(3):599–606. doi:10.1016/j.bbrc.2020.03.122
20. Siracusa MC, Wojno ED, Artis D. Functional heterogeneity in the basophil cell lineage. Adv Immunol. 2012;115:141–159. doi:10.1016/b978-0-12-394299-9.00005-9
21. Nakanishi K. Basophils are potent antigen-presenting cells that selectively induce Th2 cells. Eur J Immunol. 2010;40(7):1836–1842. doi:10.1002/eji.201040588
22. Nakanishi K. Basophils as APC in Th2 response in allergic inflammation and parasite infection. Curr Opin Immunol. 2010;22(6):814–820. doi:10.1016/j.coi.2010.10.018
23. Siracusa MC, Kim BS, Spergel JM, Artis D. Basophils and allergic inflammation. J Allergy Clin Immunol. 2013;132(4):789–801; quiz788. doi:10.1016/j.jaci.2013.07.046
24. Caruso M, Cibella F, Emma R, et al. Basophil biomarkers as useful predictors for sublingual immunotherapy in allergic rhinitis. Int Immunopharmacol. 2018;60:50–58. doi:10.1016/j.intimp.2018.04.034
25. Naclerio RM, Baroody FM, Kagey-Sobotka A, Lichtenstein LM. Basophils and eosinophils in allergic rhinitis. J Allergy Clin Immunol. 1994;94(6 Pt 2):1303–1309. doi:10.1016/0091-6749(94)90346-8
26. Folci M, Ramponi G, Arcari I, Zumbo A, Brunetta E. Eosinophils as major player in type 2 inflammation: autoimmunity and beyond. Adv Exp Med Biol. 2021;1347:197–219. doi:10.1007/5584_2021_640
27. Klementsson H. Eosinophils and the pathophysiology of allergic rhinitis. Clin Exp Allergy. 1992;22(12):1058–1064. doi:10.1111/j.1365-2222.1992.tb00130.x
28. Milanese M, Ricca V, Canonica GW, Ciprandi G. Eosinophils, specific hyperreactivity and occurrence of late phase reaction in allergic rhinitis. Eur Ann Allergy Clin Immunol. 2005;37(1):7–10.
29. Akuthota P, Wang H, Weller PF. Eosinophils as antigen-presenting cells in allergic upper airway disease. Curr Opin Allergy Clin Immunol. 2010;10(1):14–19. doi:10.1097/ACI.0b013e328334f693
30. Sannohe S, Adachi T, Hamada K, et al. Upregulated response to chemokines in oxidative metabolism of eosinophils in asthma and allergic rhinitis. Eur Respir J. 2003;21(6):925–931. doi:10.1183/09031936.03.00028103a
31. Shapouri-Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
32. Siebeler R, de Winther MPJ, Hoeksema MA. The regulatory landscape of macrophage interferon signaling in inflammation. J Allergy Clin Immunol. 2023;152(2):326–337. doi:10.1016/j.jaci.2023.04.022
33. Gentek R, Molawi K, Sieweke MH. Tissue macrophage identity and self-renewal. Immunol Rev. 2014;262(1):56–73. doi:10.1111/imr.12224
34. Zhu X, Sun Y, Yu Q, Wang X, Wang Y, Zhao Y. Exosomal lncRNA GAS5 promotes M1 macrophage polarization in allergic rhinitis via restraining mTORC1/ULK1/ATG13-mediated autophagy and subsequently activating NF-кB signaling. Int Immunopharmacol. 2023;121:110450. doi:10.1016/j.intimp.2023.110450
35. Wen S, Li F, Tang Y, et al. MIR222HG attenuates macrophage M2 polarization and allergic inflammation in allergic rhinitis by targeting the miR146a-5p/TRAF6/NF-κB axis. Front Immunol. 2023;14:1168920. doi:10.3389/fimmu.2023.1168920
36. Luo M, Zhao F, Cheng H, Su M, Wang Y. Macrophage polarization: an important role in inflammatory diseases. Front Immunol. 2024;15:1352946. doi:10.3389/fimmu.2024.1352946
37. DeBruin EJ, Gold M, Lo BC, et al. Mast cells in human health and disease. Methods Mol Biol. 2015;1220:93–119. doi:10.1007/978-1-4939-1568-2_7
38. Amin K. The role of mast cells in allergic inflammation. Respir Med. 2012;106(1):9–14. doi:10.1016/j.rmed.2011.09.007
39. Pawankar R, Yamagishi S, Takizawa R, Yagi T. Mast cell-IgE-and mast cell-structural cell interactions in allergic airway disease. Curr Drug Targets Inflamm Allergy. 2003;2(4):303–312. doi:10.2174/1568010033484016
40. Peng HS, Zhu XH. [Study on the application of mast cells in the pathogenesis of allergic rhinitis]. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2018;32(2):157–160. Hawaiian. doi:10.13201/j.issn.1001-1781.2018.02.019
41. Liu R, Zhang Y, Wang Y, et al. Anti-inflammatory effect of dictamnine on allergic rhinitis via suppression of the LYN kinase-mediated molecular signaling pathway during mast cell activation. Phytother Res. 2023;37(9):4236–4250. doi:10.1002/ptr.7904
42. Pawankar R, Yamagishi S, Yagi T. Revisiting the roles of mast cells in allergic rhinitis and its relation to local IgE synthesis. Am J Rhinol. 2000;14(5):309–317. doi:10.2500/105065800781329582
43. Nian JB, Zeng M, Zheng J, et al. Epithelial cells expressed IL-33 to promote degranulation of mast cells through inhibition on ST2/PI3K/mTOR-mediated autophagy in allergic rhinitis. Cell Cycle. 2020;19(10):1132–1142. doi:10.1080/15384101.2020.1749402
44. Collin M, Ginhoux F. Human dendritic cells. Semin Cell Dev Biol. 2019;86:1–2. doi:10.1016/j.semcdb.2018.04.015
45. Colonna M, Trinchieri G, Liu YJ. Plasmacytoid dendritic cells in immunity. Nat Immunol. 2004;5(12):1219–1226. doi:10.1038/ni1141
46. Galicia G, Gommerman JL. Plasmacytoid dendritic cells and autoimmune inflammation. Biol Chem. 2014;395(3):335–346. doi:10.1515/hsz-2013-0213
47. Santana-de Anda K, Gómez-Martín D, Soto-Solís R, Alcocer-Varela J. Plasmacytoid dendritic cells: key players in viral infections and autoimmune diseases. Semin Arthritis Rheum. 2013;43(1):131–136. doi:10.1016/j.semarthrit.2012.12.026
48. Froidure A, Shen C, Pilette C. Dendritic cells revisited in human allergic rhinitis and asthma. Allergy. 2016;71(2):137–148. doi:10.1111/all.12770
49. Froidure A, Pilette C. Les cellules dendritiques humaines dans l’asthme et la rhinite allergique [Human dendritic cells in allergic asthma and rhinitis]. Med Sci. 2015;31(2):151–158. doi:10.1051/medsci/20153102011
50. Zheng R, Chen Y, Shi J, et al. Combinatorial IL-17RB, ST2, and TSLPR signaling in dendritic cells of patients with allergic rhinitis. Front Cell Dev Biol. 2020;8:207. doi:10.3389/fcell.2020.00207
51. Wang Y, Liu J, Burrows PD, Wang JY. B cell development and maturation. Adv Exp Med Biol. 2020;1254:1–22. doi:10.1007/978-981-15-3532-1_1
52. Weindel CG, Richey LJ, Bolland S, Mehta AJ, Kearney JF, Huber BT. B cell autophagy mediates TLR7-dependent autoimmunity and inflammation. Autophagy. 2015;11(7):1010–1024. doi:10.1080/15548627.2015.1052206
53. Fan K, Jin L, Yu S. Roles of regulatory B cells in the pathogenesis of allergic rhinitis. Allergol Immunopathol. 2022;50(5):7–15. doi:10.15586/aei.v50i5.615
54. Kim AS, Doherty TA, Karta MR, et al. Regulatory B cells and T follicular helper cells are reduced in allergic rhinitis. J Allergy Clin Immunol. 2016;138(4):1192–1195.e5. doi:10.1016/j.jaci.2016.03.017
55. Luo J, Guo H, Liu Z, et al. Analysis of peripheral B cell subsets in patients with allergic rhinitis. Allergy Asthma Immunol Res. 2018;10(3):236–243. doi:10.4168/aair.2018.10.3.236
56. Dong C. Cytokine regulation and function in T cells. Annu Rev Immunol. 2021;39:51–76. doi:10.1146/annurev-immunol-061020-053702
57. Khedri M, Samei A, Fasihi-Ramandi M, Taheri RA. The immunopathobiology of T cells in stress condition: a review. Cell Stress Chaperones. 2020;25(5):743–752. doi:10.1007/s12192-020-01105-0
58. Iinuma T, Kiuchi M, Hirahara K, et al. Single-cell immunoprofiling after immunotherapy for allergic rhinitis reveals functional suppression of pathogenic T(H)2 cells and clonal conversion. J Allergy Clin Immunol. 2022;150(4):850–860.e5. doi:10.1016/j.jaci.2022.06.024
59. Kaminuma O, Nishimura T, Saeki M, Mori A, Hiroi T. T cell-mediated nasal hyperresponsiveness in allergic rhinitis. Biol Pharm Bull. 2020;43(1):36–40. doi:10.1248/bpb.b18-01021
60. Wang XQ, Ke X, Shen Y, et al. Changes in circulating follicular helper T-cells in Chinese patients with allergic rhinitis. Acta Otolaryngol. 2016;136(2):199–204. doi:10.3109/00016489.2015.1093169
61. Ozdemir C, Akdis M, Akdis CA. T regulatory cells and their counterparts: masters of immune regulation. Clin Exp Allergy. 2009;39(5):626–639. doi:10.1111/j.1365-2222.2009.03242.x
62. Hassannia H, Abediankenari S, Ghaffari J. FOXP3 and TGF-β gene polymorphisms in allergic rhinitis. Iran J Immunol. 2011;8(4):218–225.
63. Yang Q, Li C, Wang W, et al. Infiltration pattern of gammadelta T cells and its association with local inflammatory response in the nasal mucosa of patients with allergic rhinitis. Int Forum Allergy Rhinol. 2019;9(11):1318–1326. doi:10.1002/alr.22421
64. Yu S, Han B, Liu S, et al. Derp1-modified dendritic cells attenuate allergic inflammation by regulating the development of T helper type1(Th1)/Th2 cells and regulatory T cells in a murine model of allergic rhinitis. Mol Immunol. 2017;90:172–181. doi:10.1016/j.molimm.2017.07.015
65. Tang L, Chen Y, Xiang Q, Xiang J, Tang Y, Li J. The association between IL18, FOXP3 and IL13 genes polymorphisms and risk of allergic rhinitis: a meta-analysis. Inflamm Res. 2020;69(9):911–923. doi:10.1007/s00011-020-01368-4
66. Ruan Y, Zhang Y, Zhang L. [Association between single-nucleotide polymorphisms of key genes in T regulatory cells signaling pathways and the efficacy of allergic rhinitis immune therapy]. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2016;51(1):34–42. Hawaiian. doi:10.3760/cma.j.issn.1673-0860.2016.01.006
67. Zhang G, Zhang D, Shi W, Sun P, Lin P. The impact of FOXP3 polymorphism on the risk of allergic rhinitis: a meta-analysis. Ann Hum Genet. 2017;81(6):284–291. doi:10.1111/ahg.12205
68. Zhang Y, Wang CS, Zhang L. [Correlation between FOXP3 gene polymorphisms and allergic rhinitis]. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2010;45(5):397–400. Hawaiian.
69. Coria-Ramírez É, Vargas-Camaño ME, Guido-Bayardo RL, et al. Concentraciones de linfocitos T CD4+ y FOXP3+ en pacientes con rinitis alérgica [Levels of CD4+ and FOXP3+ lymphocytes in patients with allergic rhinitis]. Rev Alerg Mex. 2013;60(1):5–10.
70. Mészáros G, Szalay B, Toldi G, et al. FoxP3+ regulatory T cells in childhood allergic rhinitis and asthma. J Investig Allergol Clin Immunol. 2009;19(3):238–240.
71. Sims JE, Smith DE. Regulation of interleukin-1 activity is enhanced by cooperation between the interleukin-1 receptor type II and interleukin-1 receptor accessory protein. Eur Cytokine Netw. 2003;14(2):77–81.
72. Tsang MS, Sun X, Wong CK. The role of new IL-1 family members (IL-36 and IL-38) in atopic dermatitis, allergic asthma, and allergic rhinitis. Curr Allergy Asthma Rep. 2020;20(8):40. doi:10.1007/s11882-020-00937-1
73. Heinig K, Sperandio M. Interleukin-1R8: balancing inflammation and hemostasis in platelets. Cardiovasc Res. 2016;111(4):307–309. doi:10.1093/cvr/cvw191
74. Massey WA, Randall TC, Kagey-Sobotka A, et al. Recombinant human IL-1 alpha and −1 beta potentiate IgE-mediated histamine release from human basophils. J Immunol. 1989;143(6):1875–1880. doi:10.4049/jimmunol.143.6.1875
75. Wang HR, Wei SZ, Song XY, et al. IL-1β and allergy: focusing on its role in allergic rhinitis. Mediators Inflamm. 2023;2023:1265449. doi:10.1155/2023/1265449
76. Joki-Erkkilä VP, Karjalainen J, Hulkkonen J, et al. Allergic rhinitis and polymorphisms of the interleukin 1 gene complex. Ann Allergy Asthma Immunol. 2003;91(3):275–279. doi:10.1016/s1081-1206(10)63530-2
77. Abbas AK, Trotta E, Simeonov RD, Marson A, Bluestone JA. Revisiting IL-2: biology and therapeutic prospects. Sci Immunol. 2018;3(25). doi:10.1126/sciimmunol.aat1482
78. Niederlova V, Tsyklauri O, Kovar M, Stepanek O. IL-2-driven CD8(+) T cell phenotypes: implications for immunotherapy. Trends Immunol. 2023;44(11):890–901. doi:10.1016/j.it.2023.09.003
79. Malek TR. The biology of interleukin-2. Annu Rev Immunol. 2008;26:453–479. doi:10.1146/annurev.immunol.26.021607.090357
80. Yang YC, Clark SC. Interleukin-3: molecular biology and biologic activities. Hematol Oncol Clin North Am. 1989;3(3):441–452. doi:10.1016/S0889-8588(18)30540-9
81. Varricchi G, Poto R, Marone G, Schroeder JT. IL-3 in the development and function of basophils. Semin Immunol. 2021;54:101510. doi:10.1016/j.smim.2021.101510
82. Podolska MJ, Grützmann R, Pilarsky C, Bénard A. IL-3: key orchestrator of inflammation. Front Immunol. 2024;15:1411047. doi:10.3389/fimmu.2024.1411047
83. Neel NF, Creasy BM, Rankin JN, et al. Absence of interleukin-3 does not affect the severity of local and systemic anaphylaxis but does enhance eosinophil infiltration in a mouse model of allergic peritonitis. Immunol Lett. 2004;95(1):37–44. doi:10.1016/j.imlet.2004.06.002
84. Durham SR, Ying S, Varney VA, et al. Cytokine messenger RNA expression for IL-3, IL-4, IL-5, and granulocyte/macrophage-colony-stimulating factor in the nasal mucosa after local allergen provocation: relationship to tissue eosinophilia. J Immunol. 1992;148(8):2390–2394. doi:10.4049/jimmunol.148.8.2390
85. Ho IC, Miaw SC. Regulation of IL-4 expression in immunity and diseases. Adv Exp Med Biol. 2016;941:31–77. doi:10.1007/978-94-024-0921-5_3
86. Yamanishi Y, Karasuyama H. Basophil-derived IL-4 plays versatile roles in immunity. Semin Immunopathol. 2016;38(5):615–622. doi:10.1007/s00281-016-0568-y
87. Hussain M, Borcard L, Walsh KP, et al. Basophil-derived IL-4 promotes epicutaneous antigen sensitization concomitant with the development of food allergy. J Allergy Clin Immunol. 2018;141(1):223–234.e5. doi:10.1016/j.jaci.2017.02.035
88. Ricci M, Matucci A, Rossi O. IL-4 as a key factor influencing the development of allergen-specific Th2-like cells in atopic individuals. J Investig Allergol Clin Immunol. 1997;7(3):144–150.
89. Dickensheets HL, Donnelly RP. Inhibition of IL-4-inducible gene expression in human monocytes by type I and type II interferons. J Leukoc Biol. 1999;65(3):307–312. doi:10.1002/jlb.65.3.307
90. Noval Rivas M, Burton OT, Oettgen HC, Chatila T. IL-4 production by group 2 innate lymphoid cells promotes food allergy by blocking regulatory T-cell function. J Allergy Clin Immunol. 2016;138(3):801–811.e9. doi:10.1016/j.jaci.2016.02.030
91. Widuri A. Correlación entre la expresión de interleucina 4 y la sensibilización a alérgenos en pacientes con rinitis alérgica [Correlation between interleukin 4 (IL-4) expression and allergen sensitization in allergic rhinitis patients]. Rev Alerg Mex. 2021;68(2):89–93. doi:10.29262/ram.v658i2.800
92. Takatsu K. Interleukin-5 and IL-5 receptor in health and diseases. Proc Jpn Acad Ser B Phys Biol Sci. 2011;87(8):463–485. doi:10.2183/pjab.87.463
93. Kouro T, Takatsu K. IL-5- and eosinophil-mediated inflammation: from discovery to therapy. Int Immunol. 2009;21(12):1303–1309. doi:10.1093/intimm/dxp102
94. Takatsu K, Kouro T, Nagai Y. Interleukin 5 in the link between the innate and acquired immune response. Adv Immunol. 2009;101:191–236. doi:10.1016/s0065-2776(08)01006-7
95. Takatsu K. Interleukin 5 and B cell differentiation. Cytokine Growth Factor Rev. 1998;9(1):25–35. doi:10.1016/s1359-6101(97)00034-8
96. Zabeau L, Gevaert P, Bachert C, Tavernier J. Interleukin-5, eosinophilic diseases and therapeutic intervention. Curr Drug Targets Inflamm Allergy. 2003;2(4):319–328. doi:10.2174/1568010033484043
97. Molfino NA, Gossage D, Kolbeck R, Parker JM, Geba GP. Molecular and clinical rationale for therapeutic targeting of interleukin-5 and its receptor. Clin Exp Allergy. 2012;42(5):712–737. doi:10.1111/j.1365-2222.2011.03854.x
98. Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014;6(10):a016295. doi:10.1101/cshperspect.a016295
99. Kang S, Narazaki M, Metwally H, Kishimoto T. Historical overview of the interleukin-6 family cytokine. J Exp Med. 2020;217(5):e20190347. doi:10.1084/jem.20190347
100. Tanaka T, Kishimoto T. The biology and medical implications of interleukin-6. Cancer Immunol Res. 2014;2(4):288–294. doi:10.1158/2326-6066.Cir-14-0022
101. Yang Y, Xiao J, Tang L, et al. Effects of IL-6 polymorphisms on individual susceptibility to allergic diseases: a systematic review and meta-analysis. Front Genet. 2022;13:822091. doi:10.3389/fgene.2022.822091
102. Deng Y, Wang Z, Chang C, Lu L, Lau CS, Lu Q. Th9 cells and IL-9 in autoimmune disorders: pathogenesis and therapeutic potentials. Hum Immunol. 2017;78(2):120–128. doi:10.1016/j.humimm.2016.12.010
103. Hauber HP, Bergeron C, Hamid Q. IL-9 in allergic inflammation. Int Arch Allergy Immunol. 2004;134(1):79–87. doi:10.1159/000078384
104. Li H, Rostami A. IL-9: basic biology, signaling pathways in CD4+ T cells and implications for autoimmunity. J Neuroimmune Pharmacol. 2010;5(2):198–209. doi:10.1007/s11481-009-9186-y
105. Soussi-Gounni A, Kontolemos M, Hamid Q. Role of IL-9 in the pathophysiology of allergic diseases. J Allergy Clin Immunol. 2001;107(4):575–582. doi:10.1067/mai.2001.114238
106. Saraiva M, O’Garra A. The regulation of IL-10 production by immune cells. Nat Rev Immunol. 2010;10(3):170–181. doi:10.1038/nri2711
107. Ouyang W, Rutz S, Crellin NK, Valdez PA, Hymowitz SG. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu Rev Immunol. 2011;29:71–109. doi:10.1146/annurev-immunol-031210-101312
108. Wei H, Li B, Sun A, Guo F. Interleukin-10 family cytokines immunobiology and structure. Adv Exp Med Biol. 2019;1172:79–96. doi:10.1007/978-981-13-9367-9_4
109. Nasiri R, Hirbod-Mobarakeh A, Movahedi M, et al. Gene polymorphisms of interleukin-10 and transforming growth factor beta in allergic rhinitis. Allergol Immunopathol. 2016;44(2):125–130. doi:10.1016/j.aller.2015.05.010
110. Golebski K, Layhadi JA, Sahiner U, et al. Induction of IL-10-producing type 2 innate lymphoid cells by allergen immunotherapy is associated with clinical response. Immunity. 2021;54(2):291–307.e7. doi:10.1016/j.immuni.2020.12.013
111. Trinchieri G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol. 2003;3(2):133–146. doi:10.1038/nri1001
112. Yin X, Ren XM, Wang JX, Xu O, Dong JH, Liu CM. [The progress of the IL-12 cytokine family in allergic rhinitis]. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2017;31(3):237–241. Hawaiian. doi:10.13201/j.issn.1001-1781.2017.03.019
113. de Vries JE. The role of IL-13 and its receptor in allergy and inflammatory responses. J Allergy Clin Immunol. 1998;102(2):165–169. doi:10.1016/s0091-6749(98)70080-6
114. de Vries JE, Zurawski G. Immunoregulatory properties of IL-13: its potential role in atopic disease. Int Arch Allergy Immunol. 1995;106(3):175–179. doi:10.1159/000236842
115. Scott TE, Lewis CV, Zhu M, et al. IL-4 and IL-13 induce equivalent expression of traditional M2 markers and modulation of reactive oxygen species in human macrophages. Sci Rep. 2023;13(1):19589. doi:10.1038/s41598-023-46237-2
116. Nur Husna SM, Md Shukri N, Mohd Ashari NS, Wong KK. IL-4/IL-13 axis as therapeutic targets in allergic rhinitis and asthma. PeerJ. 2022;10:e13444. doi:10.7717/peerj.13444
117. Campion NJ, Villazala-Merino S, Thwaites RS, et al. Nasal IL-13 production identifies patients with late-phase allergic responses. J Allergy Clin Immunol. 2023;152(5):1167–1178.e12. doi:10.1016/j.jaci.2023.06.026
118. Amin K, Issa SM, Ali KM, et al. Evidence for eosinophil and IL-17 mediated inflammation in allergic rhinitis. Clin Mol Allergy. 2020;18:6. doi:10.1186/s12948-020-00117-6
119. Wang M, Zhang W, Shang J, Yang J, Zhang L, Bachert C. Immunomodulatory effects of IL-23 and IL-17 in a mouse model of allergic rhinitis. Clin Exp Allergy. 2013;43(8):956–966. doi:10.1111/cea.12123
120. Quan SH, Zhang YL, Han DH, Iwakura Y, Rhee CS. Contribution of interleukin 17A to the development and regulation of allergic inflammation in a murine allergic rhinitis model. Ann Allergy Asthma Immunol. 2012;108(5):342–350. doi:10.1016/j.anai.2012.02.014
121. Yasuda K, Nakanishi K, Tsutsui H. Interleukin-18 in health and disease. Int J Mol Sci. 2019;20(3). doi:10.3390/ijms20030649
122. Li C, Xu Y, Luo X, Chen F. The effect of IL-18 on group 2 innate lymphoid cells in allergic rhinitis. Iran J Immunol. 2021;18(3):188–194. doi:10.22034/iji.2021.85982.1734
123. Sanders NL, Mishra A. Role of interleukin-18 in the pathophysiology of allergic diseases. Cytokine Growth Factor Rev. 2016;32:31–39. doi:10.1016/j.cytogfr.2016.07.001
124. Zhang M, Liu L. [The role of IL-18 in allergic rhinitis]. Lin Chuang Er Bi Yan Hou Ke Za Zhi. 2004;18(5):257–258. Afar.
125. Wang J, Shao H, Ye L, Zhang Y, Qin B. [Upregulation of IL-18 expression in blood CD4(+) Th2 cells of patients with allergic rhinitis]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2023;39(12):1100–1107. Wolof.
126. Kruse S, Kuehr J, Moseler M, et al. Polymorphisms in the IL 18 gene are associated with specific sensitization to common allergens and allergic rhinitis. J Allergy Clin Immunol. 2003;111(1):117–122. doi:10.1067/mai.2003.43
127. Dudakov JA, Hanash AM, van den Brink MR. Interleukin-22: immunobiology and pathology. Annu Rev Immunol. 2015;33:747–785. doi:10.1146/annurev-immunol-032414-112123
128. Tang J, Xiao P, Luo X, et al. Increased IL-22 level in allergic rhinitis significantly correlates with clinical severity. Am J Rhinol Allergy. 2014;28(6):197–201. doi:10.2500/ajra.2014.28.4088
129. Farfariello V, Amantini C, Nabissi M, et al. IL-22 mRNA in peripheral blood mononuclear cells from allergic rhinitic and asthmatic pediatric patients. Pediatr Allergy Immunol. 2011;22(4):419–423. doi:10.1111/j.1399-3038.2010.01116.x
130. Shakerian L, Kolahdooz H, Garousi M, et al. IL-33/ST2 axis in autoimmune disease. Cytokine. 2022;158:156015. doi:10.1016/j.cyto.2022.156015
131. Ding W, Zou GL, Zhang W, Lai XN, Chen HW, Xiong LX. Interleukin-33: its emerging role in allergic diseases. Molecules. 2018;23(7):1665. doi:10.3390/molecules23071665
132. Ramezani F, Babaie F, Aslani S, et al. The role of the IL-33/ST2 immune pathway in autoimmunity: new insights and perspectives. Immunol Invest. 2022;51(4):1060–1086. doi:10.1080/08820139.2021.1878212
133. Billiau A, Heremans H, Vermeire K, Matthys P. Immunomodulatory properties of interferon-gamma. An update. Ann N Y Acad Sci. 1998;856:22–32. doi:10.1111/j.1749-6632.1998.tb08309.x
134. Bayrak Degirmenci P, Aksun S, Altin Z, et al. Allergic rhinitis and its relationship with IL-10, IL-17, TGF-β, IFN-γ, IL 22, and IL-35. Dis Markers. 2018;2018:9131432. doi:10.1155/2018/9131432
135. Idriss HT, Naismith JH. TNF alpha and the TNF receptor superfamily: structure-function relationship(s). Microsc Res Tech. 2000;50(3):184–195. doi:10.1002/1097-0029(20000801)50:3<184::Aid-jemt2>3.0.Co;2-h
136. Wei X, Zhang Y, Fu Z, Zhang L. The association between polymorphisms in the MRPL4 and TNF-α genes and susceptibility to allergic rhinitis. PLoS One. 2013;8(3):e57981. doi:10.1371/journal.pone.0057981
137. Ciebiada M, Gorska-Ciebiada M, Gorski P. sICAM-1 and TNF-α in asthma and rhinitis: relationship with the presence of atopy. J Asthma. 2011;48(7):660–666. doi:10.3109/02770903.2011.604886
138. Gorska-Ciebiada M, Ciebiada M, Gorska MM, Gorski P, Grzelewska-Rzymowska I. Intercellular adhesion molecule 1 and tumor necrosis factor alpha in asthma and persistent allergic rhinitis: relationship with disease severity. Ann Allergy Asthma Immunol. 2006;97(1):66–72. doi:10.1016/s1081-1206(10)61372-5
139. Larson C, Oronsky B, Carter CA, et al. TGF-beta: a master immune regulator. Expert Opin Ther Targets. 2020;24(5):427–438. doi:10.1080/14728222.2020.1744568
140. Cerwenka A, Swain SL. TGF-beta1: immunosuppressant and viability factor for T lymphocytes. Microbes Infect. 1999;1(15):1291–1296. doi:10.1016/s1286-4579(99)00255-5
141. Mirzaei Y, Savari Z, Yazdani-Nafchi F, et al. The expression analysis of IL-6, IL-18, IL-21, IL-23, and TGF-β mRNA in the nasal mucosa of patients with allergic rhinitis. Afr Health Sci. 2022;22(1):630–640. doi:10.4314/ahs.v22i1.73
142. Wang M, Gu Z, Yang J, Zhao H, Cao Z. Changes among TGF-β1(+) Breg cells and helper T cell subsets in a murine model of allergic rhinitis with prolonged OVA challenge. Int Immunopharmacol. 2019;69:347–357. doi:10.1016/j.intimp.2019.01.009
143. Chen S, Jin YT, Zhu ZY, Wu LT, Xuan LH. [Acupoint application of herbal paste relieves symptoms and naso-mucosal inflammatory response by down-regulating serum TGF-β1 in allergic rhinitis rats]. Zhen Ci Yan Jiu. 2019;44(6):430–433. Hausa. doi:10.13702/j.1000-0607.180369
144. Bousquet J, Anto JM, Bachert C, et al. Allergic rhinitis. Nat Rev Dis Primers. 2020;6(1):96. doi:10.1038/s41572-020-00237-y
145. Mims JW. Allergic rhinitis. Facial Plast Surg Clin N Am. 2012;20(1):11–20. doi:10.1016/j.fsc.2011.10.002
146. Charfi A, Zainine R, Ben Ali S, et al. Rhinites allergiques [Allergic rhinitis]. Tunis Med. 2010;88(10):690–695.
147. Meng Y, Wang C, Zhang L. Recent developments and highlights in allergic rhinitis. Allergy. 2019;74(12):2320–2328. doi:10.1111/all.14067
148. Huang X, Chen Y, Zhang F, Yang Q, Zhang G. Peripheral Th17/Treg cell-mediated immunity imbalance in allergic rhinitis patients. Braz J Otorhinolaryngol. 2014;80(2):152–155. doi:10.5935/1808-8694.20140031
149. Guo LP, Yan M, Niu RB, et al. Role of Th2, Th17 and treg cells and relevant cytokines in pathogenesis of allergic rhinitis. Allergy Asthma Clin Immunol. 2024;20(1):40. doi:10.1186/s13223-024-00905-8
150. Tao B, Ruan G, Wang D, Li Y, Wang Z, Yin G. Imbalance of peripheral Th17 and regulatory T cells in children with allergic rhinitis and bronchial asthma. Iran J Allergy Asthma Immunol. 2015;14(3):273–279.
151. Dong-cai L, Peng W, Sheng L, Qiao-lian Y, Jian-pian L, Bao-hui C. Therapeutic effects of resveratrol on OVA-induced allergic rhinitis in mice and immune mechanisms. Chin Pharmacol Bull. 2021;37(02):215–220.
152. Choubey S, Varughese LR, Kumar V, Beniwal V. Medicinal importance of gallic acid and its ester derivatives: a patent review. Pharm Pat Anal. 2015;4(4):305–315. doi:10.4155/ppa.15.14
153. Acar M, Muluk NB, Yigitaslan S, et al. Can curcumin modulate allergic rhinitis in rats? J Laryngol Otol. 2016;130(12):1103–1109. doi:10.1017/s0022215116008999
154. Altıntoprak N, Kar M, Acar M, Berkoz M, Muluk NB, Cingi C. Antioxidant activities of curcumin in allergic rhinitis. Eur Arch Otorhinolaryngol. 2016;273(11):3765–3773. doi:10.1007/s00405-016-4076-4
155. Zhang N, Li H, Jia J, He M. Anti-inflammatory effect of curcumin on mast cell-mediated allergic responses in ovalbumin-induced allergic rhinitis mouse. Cell Immunol. 2015;298(1–2):88–95. doi:10.1016/j.cellimm.2015.09.010
156. Lv C, Zhang Y, Shen L. Preliminary clinical effect evaluation of resveratrol in adults with allergic rhinitis. Int Arch Allergy Immunol. 2018;175(4):231–236. doi:10.1159/000486959
157. Huimin M, Qianqian W, Yaling Y. Therapeutic effects of resveratrol on ovalbumin-induced allergic rhinitis in Guinea pigs. Lishizhen Med Mater Med Res. 2013;24(11):2642–2644.
158. Kim SH, Jun CD, Suk K, et al. Gallic acid inhibits histamine release and pro-inflammatory cytokine production in mast cells. Toxicol Sci. 2006;91(1):123–131. doi:10.1093/toxsci/kfj063
159. Pan Z, Zhou Y, Luo X, et al. Against NF-κB/thymic stromal lymphopoietin signaling pathway, catechin alleviates the inflammation in allergic rhinitis. Int Immunopharmacol. 2018;61:241–248. doi:10.1016/j.intimp.2018.06.011
160. Morsy MA, Patel SS, Bakrania A, et al. Ameliorative effect of a neoteric regimen of catechin plus cetirizine on ovalbumin-induced allergic rhinitis in rats. Life. 2022;12(6):820. doi:10.3390/life12060820
161. Phan HTL, Nam YR, Kim HJ, et al. In-vitro and in-vivo anti-allergic effects of magnolol on allergic rhinitis via inhibition of ORAI1 and ANO1 channels. J Ethnopharmacol. 2022;289:115061. doi:10.1016/j.jep.2022.115061
162. Wu S, Xiao D. Effect of curcumin on nasal symptoms and airflow in patients with perennial allergic rhinitis. Ann Allergy Asthma Immunol. 2016;117(6):697–702.e1. doi:10.1016/j.anai.2016.09.427
163. Thakare VN, Osama MM, Naik SR. Therapeutic potential of curcumin in experimentally induced allergic rhinitis in Guinea pigs. Int Immunopharmacol. 2013;17(1):18–25. doi:10.1016/j.intimp.2013.04.025
164. Moon PD, Han NR, Lee JS, et al. Effects of resveratrol on thymic stromal lymphopoietin expression in mast cells. Medicina. 2020;57(1):21. doi:10.3390/medicina57010021
165. Civelek M, Bilotta S, Lorentz A. Resveratrol attenuates mast cell mediated allergic reactions: potential for use as a nutraceutical in allergic diseases? Mol Nutr Food Res. 2022;66(15):e2200170. doi:10.1002/mnfr.202200170
166. Li Z, Du X, Li Y, et al. Pharmacokinetics of gallic acid and protocatechuic acid in humans after dosing with Relinqing (RLQ) and the potential for RLQ-perpetrated drug-drug interactions on organic anion transporter (OAT) 1/3. Pharm Biol. 2021;59(1):757–768. doi:10.1080/13880209.2021.1934039
167. Shana, Shuyu L, Xiaocheng Q, Jinfeng Z. Study on the anti-allergic effect of gallic acid in mice with allergic rhinitis. Chin J Clin Pharmacol. 2022;38(05):409–412.
168. Kim TE, Shin KH, Park JE, et al. Effect of green tea catechins on the pharmacokinetics of digoxin in humans. Drug Des Devel Ther. 2018;12:2139–2147. doi:10.2147/dddt.S148257
169. Rosen T. Green tea catechins: biologic properties, proposed mechanisms of action, and clinical implications. J Drugs Dermatol. 2012;11(11):e55–60.
170. Bolger GT, Pucaj K, Minta YO, Sordillo P. Relationship between the in vitro efficacy, pharmacokinetics and in vivo efficacy of curcumin. Biochem Pharmacol. 2022;205:115251. doi:10.1016/j.bcp.2022.115251
171. Kim JS, Oh JM, Choi H, et al. Activation of the Nrf2/HO-1 pathway by curcumin inhibits oxidative stress in human nasal fibroblasts exposed to urban particulate matter. BMC Complement Med Ther. 2020;20(1):101. doi:10.1186/s12906-020-02886-8
172. Robertson I, Wai Hau T, Sami F, et al. The science of resveratrol, formulation, pharmacokinetic barriers and its chemotherapeutic potential. Int J Pharm. 2022;618:121605. doi:10.1016/j.ijpharm.2022.121605
173. Tian B, Liu J. Resveratrol: a review of plant sources, synthesis, stability, modification and food application. J Sci Food Agric. 2020;100(4):1392–1404. doi:10.1002/jsfa.10152
174. Zhang W, Tang R, Ba G, Li M, Lin H. Anti-allergic and anti-inflammatory effects of resveratrol via inhibiting TXNIP-oxidative stress pathway in a mouse model of allergic rhinitis. World Allergy Organ J. 2020;13(10):100473. doi:10.1016/j.waojou.2020.100473
175. Fan Y, Piao CH, Hyeon E, et al. Gallic acid alleviates nasal inflammation via activation of Th1 and inhibition of Th2 and Th17 in a mouse model of allergic rhinitis. Int Immunopharmacol. 2019;70:512–519. doi:10.1016/j.intimp.2019.02.025
176. Zengfeng P. Catechin alleviates the Inflammation in Allergic Rhinitis by Inhibiting NF-κB/TSLP Signal Pathway [Master]. 2018.
177. Fu M, Fu S, Ni S, Zou L, Liu Y, Hong T. Anti-inflammatory effect of epigallocatechin gallate in a mouse model of ovalbumin-induced allergic rhinitis. Int Immunopharmacol. 2017;49:102–108. doi:10.1016/j.intimp.2017.05.030
178. Dai SY, Qin WX, Yu S, Li C, Yang YH, Pei YH. Honokiol and magnolol: a review of structure-activity relationships of their derivatives. Phytochemistry. 2024;223:114132. doi:10.1016/j.phytochem.2024.114132
179. Lee YC, Weng YS, Wang HY, et al. Magnolol induces apoptosis through extrinsic/intrinsic pathways and attenuates NF-κB/STAT3 signaling in non-small-cell lung cancer cells. Anticancer Res. 2022;42(8):3825–3833. doi:10.21873/anticanres.15873
180. Li J, Wu Y, Ma Y, et al. A UPLC-MS/MS method reveals the pharmacokinetics and metabolism characteristics of kaempferol in rats under hypoxia. Drug Metab Pharmacokinet. 2022;43:100440. doi:10.1016/j.dmpk.2021.100440
181. Periferakis A, Periferakis K, Badarau IA, et al. Kaempferol: antimicrobial properties, sources, clinical, and traditional applications. Int J Mol Sci. 2022;23(23). doi:10.3390/ijms232315054
182. Oh HA, Han NR, Kim MJ, Kim HM, Jeong HJ. Evaluation of the effect of kaempferol in a murine allergic rhinitis model. Eur J Pharmacol. 2013;718(1–3):48–56. doi:10.1016/j.ejphar.2013.08.045
183. Ebihara N, Takahashi K, Takemura H, Akanuma Y, Asano K, Sunagawa M. Suppressive effect of quercetin on nitric oxide production from nasal epithelial cells in vitro. Evid Based Complement Alternat Med. 2018;2018:6097625. doi:10.1155/2018/6097625
184. Li J, Lin X, Liu X, Ma Z, Li Y. Baicalin regulates Treg/Th17 cell imbalance by inhibiting autophagy in allergic rhinitis. Mol Immunol. 2020;125:162–171. doi:10.1016/j.molimm.2020.07.008
185. Tao L. Effects of Baicalein on a Model of Allergic Rhinitis and the Mechanism [Master]. 2020.
186. Qing H, Weiwei Y, Wenguang W, Lingyan P. Baicalein ameliorated inflammation in allergic rhinitis mice by inhibiting NF-κb stat3erk signaling pathway. Chin J Gerontol. 2024;44(01):165–170.
187. Wang Y, Jin W, Wang J. Tanshinone IIA regulates microRNA‑125b/foxp3/caspase‑1 signaling and inhibits cell viability of nasopharyngeal carcinoma. Mol Med Rep. 2021;23(5). doi:10.3892/mmr.2021.12010
188. Zhou YJ, Wang H, Sui HH, et al. Inhibitory effect of baicalin on allergic response in ovalbumin-induced allergic rhinitis Guinea pigs and lipopolysaccharide-stimulated human mast cells. Inflamm Res. 2016;65(8):603–612. doi:10.1007/s00011-016-0943-0
189. Li S, Li Z, Tan T, Dai S, Wu Y, Xu F. Tanshinone IIA improves degranulation of mast cells and allergic rhinitis induced by ovalbumin by inhibiting the PLCγ1/PKC/IP3R pathway. Hum Exp Toxicol. 2021;40(12_suppl):S702–s710. doi:10.1177/09603271211058884
190. Liang KL, Yu SJ, Huang WC, Yen HR. Luteolin attenuates allergic nasal inflammation via inhibition of Interleukin-4 in an allergic rhinitis mouse model and peripheral blood from human subjects with allergic rhinitis. Front Pharmacol. 2020;11:291. doi:10.3389/fphar.2020.00291
191. Jang TY, Jung AY, Kyung TS, Kim DY, Hwang JH, Kim YH. Anti-allergic effect of luteolin in mice with allergic asthma and rhinitis. Cent Eur J Immunol. 2017;42(1):24–29. doi:10.5114/ceji.2017.67315
192. Yongna J. The effect of tanshinone IIA on mast cell-mediated allergic rhinitis by modulating NF-ΚB pathway. J Wuhan Univ Med. 2023;45(23):3540–3544.
193. Hua H, Li F, Xi Y, Jiao W, Wen S, Tao Z. [To explore the therapeutic effect of myrtle oil, anthocyanin and hyaluronic acid in combination with topical application on allergic rhinitis in rats exposed to PM2.5]. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2020;34(8):719–725. Hawaiian. doi:10.13201/j.issn.2096-7993.2020.08.010
194. Alizadeh SR, Ebrahimzadeh MA. Quercetin derivatives: drug design, development, and biological activities, a review. Eur J Med Chem. 2022;229:114068. doi:10.1016/j.ejmech.2021.114068
195. Jafarinia M, Sadat Hosseini M, Kasiri N, et al. Quercetin with the potential effect on allergic diseases. Allergy Asthma Clin Immunol. 2020;16:36. doi:10.1186/s13223-020-00434-0
196. Wen RJ, Dong X, Zhuang HW, et al. Baicalin induces ferroptosis in osteosarcomas through a novel Nrf2/xCT/GPX4 regulatory axis. Phytomedicine. 2023;116:154881. doi:10.1016/j.phymed.2023.154881
197. Zhou YJ, Wang H, Sui HH, Li L, Zhou CL, Huang JJ. Inhibitory effect of baicalin on allergic response in ovalbumin-induced allergic rhinitis Guinea pigs and lipopolysaccharide-stimulated human mast cells. Inflamm Res. 2016;65(8):603–612. doi:10.1007/s00011-016-0943-0
198. Chen S, Chen G, Shu S, Xu Y, Ma X. Metabolomics analysis of baicalin on ovalbumin-sensitized allergic rhinitis rats. R Soc Open Sci. 2019;6(2):181081. doi:10.1098/rsos.181081
199. Huang H, Song C, Chang J. Synthesis and biological activity study of tanshinone derivatives: a literature and patent review. Curr Top Med Chem. 2020;20(28):2520–2534. doi:10.2174/1568026620666200922115109
200. Chen Q, Shao L, Li Y, et al. Tanshinone IIA alleviates ovalbumin-induced allergic rhinitis symptoms by inhibiting Th2 cytokine production and mast cell histamine release in mice. Pharm Biol. 2022;60(1):326–333. doi:10.1080/13880209.2022.2034894
201. Şahin A, Sakat MS, Kılıç K, et al. The protective effect of Naringenin against ovalbumin-induced allergic rhinitis in rats. Eur Arch Otorhinolaryngol. 2021;278(12):4839–4846. doi:10.1007/s00405-021-06769-7
202. Niu Y, She Z, Su C, Zhao Q, Wang S, Xiao B. The effects and the mechanisms of naringenin from Artemisia ordosica Krasch on allergic rhinitis based on mast cell degranulation model and network pharmacology. J Pharm Pharmacol. 2022;74(3):397–408. doi:10.1093/jpp/rgab166
203. Kilic K, Sakat MS, Yildirim S, et al. The amendatory effect of hesperidin and thymol in allergic rhinitis: an ovalbumin-induced rat model. Eur Arch Otorhinolaryngol. 2019;276(2):407–415. doi:10.1007/s00405-018-5222-y
204. Wei L, Shen N, Shuguang Z, et al. Effects of hesperidin on immune response and TSLP/OX40L/OX40 signaling pathway in mice with allergic rhinitis. J bovine immunol. 2022;38(09):753–760+767. doi:10.13431/j.cnki.immunol.j.20220105
205. Ding H, Lu X, Wang H, et al. NLRP3 inflammasome deficiency alleviates inflammation and oxidative stress by promoting PINK1/Parkin-mediated mitophagy in allergic rhinitis mice and nasal epithelial cells. J Asthma Allergy. 2024;17:717–731. doi:10.2147/JAA.S467774
206. Liu S, Wang C, Zhang Y, et al. Polydatin inhibits mitochondrial damage and mitochondrial ROS by promoting PINK1-Parkin-mediated mitophagy in allergic rhinitis. FASEB J. 2023;37(4):e22852. doi:10.1096/fj.202201231RR
207. de Ferrars RM, Czank C, Zhang Q, et al. The pharmacokinetics of anthocyanins and their metabolites in humans. Br J Pharmacol. 2014;171(13):3268–3282. doi:10.1111/bph.12676
208. Yamaura K, Ishiwatari M, Yamamoto M, Shimada M, Bi Y, Ueno K. Anthocyanins, but not anthocyanidins, from bilberry (Vaccinium myrtillus L.) alleviate pruritus via inhibition of mast cell degranulation. J Food Sci. 2012;77(12):H262–7. doi:10.1111/j.1750-3841.2012.02974.x
209. Rakha A, Umar N, Rabail R, et al. Anti-inflammatory and anti-allergic potential of dietary flavonoids: a review. Biomed Pharmacother. 2022;156:113945. doi:10.1016/j.biopha.2022.113945
210. Zhang Y, Fu K, Wang C, et al. Protective effects of dietary quercetin on cerebral ischemic injury: pharmacology, pharmacokinetics and bioavailability-enhancing nanoformulations. Food Funct. 2023;14(10):4470–4489. doi:10.1039/d2fo03122a
211. Ke X, Chen Z, Wang X, Kang H, Hong S. Quercetin improves the imbalance of Th1/Th2 cells and Treg/Th17 cells to attenuate allergic rhinitis. Autoimmunity. 2023;56(1):2189133. doi:10.1080/08916934.2023.2189133
212. Kashiwabara M, Asano K, Mizuyoshi T, Kobayashi H. Suppression of neuropeptide production by quercetin in allergic rhinitis model rats. BMC Complement Altern Med. 2016;16:132. doi:10.1186/s12906-016-1123-z
213. Otaki A, Furuta A, Asano K. Quercetin-induced enhancement of nasal epithelial cells’ ability to produce Clara cell 10-kD protein in vitro and in vivo. Medicines. 2023;10(4):28. doi:10.3390/medicines10040028
214. Zhong H, Li DY, Wang SY, et al. [Tanshinone IIa attenuates vascular calcification through inhibition of NF-κB and β-catenin signaling pathways]. Sheng Li Xue Bao. 2022;74(6):949–958. Dutch.
215. Zhao JY, Pu J, Fan J, et al. Tanshinone IIA prevents acute lung injury by regulating macrophage polarization. J Integr Med. 2022;20(3):274–280. doi:10.1016/j.joim.2022.01.006
216. Rebello CJ, Beyl RA, Lertora JJL, et al. Safety and pharmacokinetics of naringenin: a randomized, controlled, single-ascending-dose clinical trial. Diabetes Obes Metab. 2020;22(1):91–98. doi:10.1111/dom.13868
217. Liu Z, Niu X, Wang J. Naringenin as a natural immunomodulator against T cell-mediated autoimmune diseases: literature review and network-based pharmacology study. Crit Rev Food Sci Nutr. 2023;63(32):11026–11043. doi:10.1080/10408398.2022.2092054
218. Ming NJ, Liya C, Meimei W. Effects of Naringin on tlr4nf-kappa b-TNF-α signaling pathway in nasal mucosa of allergic rhinitis rats. Chin Lab Diagn. 2023;27(04):483–488.
219. Liu J, Zhou H, Song L, et al. Anthocyanins: promising natural products with diverse pharmacological activities. Molecules. 2021;26(13). doi:10.3390/molecules26133807
220. Wen Y, Wang Y, Zhao C, Zhao B, Wang J. The pharmacological efficacy of baicalin in inflammatory diseases. Int J Mol Sci. 2023;24(11):9317. doi:10.3390/ijms24119317
221. Fang X, Hu X. Advances in the synthesis of lignan natural products. Molecules. 2018;23(12):3385. doi:10.3390/molecules23123385
222. Wang L, Chen W, Jin H, et al. CXCL1/IGHG1 signaling enhances crosstalk between tumor cells and tumor-associated macrophages to promote MC-LR-induced colorectal cancer progression. Environ Pollut. 2024;351:124081. doi:10.1016/j.envpol.2024.124081
223. Yang Y, Wang L, Wang S, Wang Y, Du Y, Fan Y. Luteolin restored Treg/Th17 balance to ameliorate allergic rhinitis in a mouse model. Immunopharmacol Immunotoxicol. 2023;45(4):461–468. doi:10.1080/08923973.2023.2166527
224. Zeng W, Jin L, Zhang F, Zhang C, Liang W. Naringenin as a potential immunomodulator in therapeutics. Pharmacol Res. 2018;135:122–126. doi:10.1016/j.phrs.2018.08.002
225. Buzdağlı Y, Eyipınar CD, Kacı FN, Tekin A. Effects of hesperidin on anti-inflammatory and antioxidant response in healthy people: a meta-analysis and meta-regression. Int J Environ Health Res. 2023;33(12):1390–1405. doi:10.1080/09603123.2022.2093841
226. Karami A, Fakhri S, Kooshki L, Khan H. Polydatin: pharmacological mechanisms, therapeutic targets, biological activities, and health benefits. Molecules. 2022;27(19):6474. doi:10.3390/molecules27196474
227. Qing G, Di N. Effect and mechanism of polydatin on inflammatory response in mice with allergic rhinitis. Chin J Clin Pharmacol. 2023;39(20):2951–2955. doi:10.13699/j.cnki.1001-6821.2023.20.014
228. Oh HA, Seo JY, Jeong HJ, Kim HM. Ginsenoside Rg1 inhibits the TSLP production in allergic rhinitis mice. Immunopharmacol Immunotoxicol. 2013;35(6):678–686. doi:10.3109/08923973.2013.837061
229. Liu J, Yang N, Yi X, et al. Integration of transcriptomics and metabolomics to reveal the effect of ginsenoside Rg3 on allergic rhinitis in mice. Food Funct. 2023;14(5):2416–2431. doi:10.1039/d2fo03885d
230. Shi Z, Jiang J, Zhao D, Xie B, Li Y, Yu C. Effects of astragaloside IV on the pharmacokinetics of metoprolol in rats and its mechanism. Curr Drug Metab. 2022;23(2):131–136. doi:10.2174/1389200223666220128143133
231. WenLing M. To Investigate the Effect of Astragaloside IV on Allergic Rhinitis (ar) in Rats Based on TLR4/NF-κb/Myd88 Signaling Pathway [Master]. 2023.
232. Chen X, Ma H, Zhou L, Yan D, Liu S, Wu J. Preliminary study on the improvement effects of astragaloside IV on allergic rhinitis model mice. Chin Pharm. 2021;32(10):1187–1195.
233. Ying T, Fei Z, Chuanxin D. Effects of astragaloside IV on HMGB1TLR4NF-ΚB signaling pathway in allergic rhinitis mice. Mod immunol. 2021;41(01):50–55.
234. Ying ZFT. Astragaloside IV treatment of allergic rhinitis mice by balancingTh17/Treg cells and related cytokines. J Shenyang Pharm Univ. 2022;39(03):277–282. doi:10.14066/j.cnki.cn21-1349/r.2020.0351
235. Liu S, Chen W, Zhao Y, Zong Y, Li J, He Z. Research progress on effects of ginsenoside Rg2 and Rh1 on nervous system and related mechanisms. Molecules. 2023;28(23):7935. doi:10.3390/molecules28237935
236. Liang Y, Chen B, Liang D, et al. Pharmacological effects of astragaloside IV: a review. Molecules. 2023;28(16):6118. doi:10.3390/molecules28166118
237. Xuqing C, Yunlong Z, Huaan M, et al. Study on effect of astragaloside IV on mitochondrial dynamic balance and NK cell differentiation in mice with allergic rhinitis. Chin J Immunol. 2022;38(07):801–807.
238. Bai X, Wu Z, Hu Z. Effect of 18β-glycyrrhetinic acid on nasal mucosal remodeling in rats with allergic rhinitis. Chin J Clin Pharmacol. 2020;36(14):2103–2106. doi:10.13699/j.cnki.1001-6821.2020.14.054
239. Yang J, Xi K, Gui Y, et al. [The effect of 18β-sodium glycyrrhetinic acid on the nasal mucosa epithelial cilia in rat models of allergic rhinitis]. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2015;29(23):2060–2064. Hawaiian.
240. Yushui F, Yuanzheng F, Kaiming Y. 18β-glycyrrhetinic acid sodium ameliorates allergic rhinitis and nasal mucosa pathological changes and the balance mechanism of Th1/Th2 in young rats. West Med. 2021;33(11):1596–1601+1607.
241. Zhou YX, Gong XH, Zhang H, Peng C. A review on the pharmacokinetics of paeoniflorin and its anti-inflammatory and immunomodulatory effects. Biomed Pharmacother. 2020;130:110505. doi:10.1016/j.biopha.2020.110505
242. Li JL, Xi KH, Hou Y, et al. [Effects of 18β-glycyrrhetinic acid on the expression of CCL11, AQP1 and EOS in nasal mucosa of allergic rhinitis rats]. Sichuan Da Xue Xue Bao Yi Xue Ban. 2015;46(3):389–393. Danish.
243. Wang G, Cheng N. Paeoniflorin inhibits mast cell-mediated allergic inflammation in allergic rhinitis. J Cell Biochem. 2018;119(10):8636–8642. doi:10.1002/jcb.27135
244. Feng L, Wu Y, Fei JPX. To investigate the effect of paeoniflorin on immune balance in allergic rhinitis rats based on IL-33/ST2 pathway. Chin J Gerontol. 2024;44(12):3034–3038.
245. Ciyu Y. Effect of Tripterine on PI3K/AKT/GSK3 β Signaling Pathway in Substance p-sensitized Mast Cells [Master]. 2017.
246. Ying L, Xiangdong C. Effect of tripterine on expression of NF-κ b and Eotaxin in nasal mucosa of allergic rhinitis rats. J Clin Otolaryngol Head Neck Surg. 2012;26(20):943–945. doi:10.13201/j.issn.1001-1781.2012.20.011
247. Jing C, Hui L, Haijun W. Study on the mechanism of Tripterine in the treatment of allergic rhinitis in rats. J Clin Otolaryngol Head Neck Surg. 2014;28(08):550–553. doi:10.13201/j.issn.1001-1781.2014.08.013
248. Min G. Study on the Immunomodulatory Effect and Mechanism of Saponin Extract of Steamed Panax Notoginseng on Mice with Immunosuppression [Master]. 2022.
249. Ling Y, Liu J, Jiangping L, He L. Effect of ephedrine mediated TSLP/OX40L pathway in regulating Th2 type immune response in rats with allergic rhinitis. Chin J Immunol. 2022;38(03):319–323.
250. Liu Y, Sheng R, Fan J, Guo R. A review on structure-activity relationships of glycyrrhetinic acid derivatives with diverse bioactivities. Mini Rev Med Chem. 2022;22(15):2024–2066. doi:10.2174/1389557522666220126093033
251. Ma Y, Gui Y, Wang Y, et al. The effect of 18β-glycyrrhetinic acid on tight junctions of the nasal mucosa epithelial cells in rat models with allergic rhinitis. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2014;28(20):1590–1594. Chinese.
252. Li JL, Xi KH, Hou Y, et al. [Effects of 18β-glycyrrhetinic acid on the expression of CCL11, AQP1 and EOS in nasal mucosa of allergic rhinitis rats]. Sichuan Da Xue Xue Bao Yi Xue Ban. 2015;46(3):389–393. Chinese.
253. Yang GJ, Xi KH, Chen XW, et al. [Effect of 18-β glycyrrhetinic acid on the endoplasmic reticulum of nasal epithelial cells in allergic rhinitis model rats]. Zhongguo Zhong Xi Yi Jie He Za Zhi. 2015;35(5):578–582. Danish.
254. Chen K, Yang R, Shen FQ, Zhu HL. Advances in pharmacological activities and mechanisms of glycyrrhizic acid. Curr Med Chem. 2020;27(36):6219–6243. doi:10.2174/0929867325666191011115407
255. Fouladi S, Masjedi M, Ghasemi R, Hakemi GM, Eskandari N. The in vitro impact of glycyrrhizic acid on CD4+ T lymphocytes through OX40 receptor in the patients with allergic rhinitis. Inflammation. 2018;41(5):1690–1701. doi:10.1007/s10753-018-0813-8
256. Liuqing Z, Jing H, Kang P, et al. Observation of intestinal allergy reaction of sodium glycyrrhetinate in rats with allergic rhinitis. Ningxia Med J. 2019;41(08):728–729. doi:10.13621/j.1001-5949.2019.08.0728
257. Ma Y, Gui Y, Wang Y, et al. [The effect of 18β-glycyrrhetinic acid on tight junctions of the nasal mucosa epithelial cells in rat models with allergic rhinitis]. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2014;28(20):1590–1594. Hawaiian.
258. Shirai T, Nakai A, Ando E, et al. Celastrol suppresses humoral immune responses and autoimmunity by targeting the COMMD3/8 complex. Sci Immunol. 2023;8(81):eadc9324. doi:10.1126/sciimmunol.adc9324
259. Zheng D, Chu Y, Li S, et al. Enhancing effect of borneol on pharmacokinetics of ginsenoside Rb(1), ginsenoside Rg(1), and notoginsenoside R(1) in healthy volunteers after oral administration of compound Danshen dropping pills. Biomed Chromatogr. 2022;36(5):e5311. doi:10.1002/bmc.5311
260. Liu H, Yang J, Yang W, et al. Focus on notoginsenoside R1 in metabolism and prevention against human diseases. Drug Des Devel Ther. 2020;14:551–565. doi:10.2147/dddt.S240511
261. Zhang YL, Wang CY, Liu SQ, et al. Panax notoginseng saponin R1 attenuates allergic rhinitis through AMPK/DRP1 mediated mitochondrial fission. Chin Pharmacol Bull. 2023;39(03):512–519.
262. Shaikh WA. Ephedrine-saline nasal wash in allergic rhinitis. J Allergy Clin Immunol. 1995;96(5). doi:10.1016/s0091-6749(95)70257-1
263. Oh HA, Kim HM, Jeong HJ. Distinct effects of imperatorin on allergic rhinitis: imperatorin inhibits caspase-1 activity in vivo and in vitro. J Pharmacol Exp Ther. 2011;339(1):72–81. doi:10.1124/jpet.111.184275
264. Li H, Jianying C, Mingbin H. Effects of andrographolide driping pills combined with danxi yupingfeng granules on immunologic function of children with allergic rhinitis. Chin Pharm. 2017;28(05):643–645.
265. Chen Z, Tao ZZ, Zhou XH, Wu TT, Ye LF. Immunosuppressive effect of sinomenine in an allergic rhinitis mouse model. Exp Ther Med. 2017;13(5):2405–2410. doi:10.3892/etm.2017.4237
266. Zheng Y, Yang Y, Li Y, et al. Ephedrine hydrochloride inhibits PGN-induced inflammatory responses by promoting IL-10 production and decreasing proinflammatory cytokine secretion via the PI3K/Akt/GSK3β pathway. Cell Mol Immunol. 2013;10(4):330–337. doi:10.1038/cmi.2013.3
267. Gad MZ, Azab SS, Khattab AR, Farag MA. Over a century since ephedrine discovery: an updated revisit to its pharmacological aspects, functionality and toxicity in comparison to its herbal extracts. Food Funct. 2021;12(20):9563–9582. doi:10.1039/d1fo02093e
268. Zheng Y, Guo Z, He W, et al. Ephedrine hydrochloride protects mice from LPS challenge by promoting IL-10 secretion and inhibiting proinflammatory cytokines. Int Immunopharmacol. 2012;13(1):46–53. doi:10.1016/j.intimp.2012.03.005
269. Deng M, Xie L, Zhong L, Liao Y, Liu L, Li X. Imperatorin: a review of its pharmacology, toxicity and pharmacokinetics. Eur J Pharmacol. 2020;879:173124. doi:10.1016/j.ejphar.2020.173124
270. Wang C, Hou Y, Ge S, et al. Synthetic imperatorin derivatives alleviate allergic reactions via mast cells. Biomed Pharmacother. 2022;150:112982. doi:10.1016/j.biopha.2022.112982
271. Wang N, Wang J, Zhang Y, et al. Imperatorin ameliorates mast cell-mediated allergic airway inflammation by inhibiting MRGPRX2 and CamKII/ERK signaling pathway. Biochem Pharmacol. 2021;184:114401. doi:10.1016/j.bcp.2020.114401
272. Abu-Ghefreh AA, Canatan H, Ezeamuzie CI. In vitro and in vivo anti-inflammatory effects of andrographolide. Int Immunopharmacol. 2009;9(3):313–318. doi:10.1016/j.intimp.2008.12.002
273. Li J, Luo L, Wang X, Liao B, Li G. Inhibition of NF-kappaB expression and allergen-induced airway inflammation in a mouse allergic asthma model by andrographolide. Cell Mol Immunol. 2009;6(5):381–385. doi:10.1038/cmi.2009.49
274. Chen X, Lu C, Duan Y, Huang Y. Recent advancements in drug delivery of sinomenine, A disease-modifying anti-rheumatic drug. Pharmaceutics. 2022;14(12):2820. doi:10.3390/pharmaceutics14122820
275. Oh YC, Kang OH, Choi JG, et al. Anti-allergic effects of sinomenine by inhibition of prostaglandin D2 and leukotriene C4 in mouse bone marrow-derived mast cells. Immunopharmacol Immunotoxicol. 2011;33(2):266–270. doi:10.3109/08923973.2010.499914
276. Huang L, Dong Y, Wu J, et al. Sinomenine-induced histamine release-like anaphylactoid reactions are blocked by tranilast via inhibiting NF-κB signaling. Pharmacol Res. 2017;125(Pt B):150–160. doi:10.1016/j.phrs.2017.08.014
277. Bousquet J, Anto JM, Bachert C, et al. Allergic rhinitis. Nat Rev Dis Primers. 2020;6(1):95. doi:10.1038/s41572-020-00227-0
278. Ponda P, Carr T, Rank MA, Bousquet J. Nonallergic rhinitis, allergic rhinitis, and immunotherapy: advances in the last decade. J Allergy Clin Immunol Pract. 2023;11(1):35–42. doi:10.1016/j.jaip.2022.09.010
279. Tosca MA, Trincianti C, Naso M, Nosratian V, Ciprandi G. Treatment of allergic rhinitis in clinical practice. Curr Pediatr Rev. 2024;20(3):271–277. doi:10.2174/1573396320666230912103108
280. Klimek L, Sperl A, Becker S, Mösges R, Tomazic PV. Current therapeutical strategies for allergic rhinitis. Expert Opin Pharmacother. 2019;20(1):83–89. doi:10.1080/14656566.2018.1543401
281. Cardona V, Luengo O, Labrador-Horrillo M. Immunotherapy in allergic rhinitis and lower airway outcomes. Allergy. 2017;72(1):35–42. doi:10.1111/all.12989
282. Fan PS, Sun MJ, Qin D, Yuan CS, Chen XG, Liu Y. Nanosystems as curative platforms for allergic disorder management. J Mater Chem B. 2021;9(7):1729–1744. doi:10.1039/d0tb02590a
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.