Back to Journals » OncoTargets and Therapy » Volume 11
Experimental study of inhibitory effects of diallyl trisulfide on the growth of human osteosarcoma Saos-2 cells by downregulating expression of glucose-regulated protein 78
Authors Zhang Y ![]() , Xie WP, Zhang YK
, Xie WP, Zhang YK ![]() , Chen YQ, Wang DL, Li G, Guan DH
, Chen YQ, Wang DL, Li G, Guan DH
Received 6 September 2017
Accepted for publication 28 October 2017
Published 9 January 2018 Volume 2018:11 Pages 271—277
DOI https://doi.org/10.2147/OTT.S150933
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Samir Farghaly
Yue Zhang,1,* Wen-Peng Xie,1,* Yong-Kui Zhang,2 Yi-Qiang Chen,3 Dong-Li Wang,2 Gang Li,2 Dong-Hui Guan2
1First Clinical Medical College, Shandong University of Traditional Chinese Medicine, Jinan, People’s Republic of China; 2Department of Orthopedics, Affiliated Hospital of Shandong University of Traditional Chinese Medicine, Jinan, People’s Republic of China; 3Department of Orthopedics, The First People’s Hospital of Taian City, Taian, People’s Republic of China
*These authors contributed equally to the paper
Background: Diallyl trisulfide (DATS) is a natural organic sulfur compound isolated from garlic that has good anticancer activity according to many previous reports. There are many studies pointing out that DATS can downregulate expression of the glucose-regulated protein 78 (GRP78), which is associated with poor prognosis and drug resistance in various types of human cancers. However, it remains unknown whether DATS has the same effect on human osteosarcoma cells. This study attempted to clarify the potential molecular mechanisms of the action of DATS in human osteosarcoma Saos-2 cells.
Methods: We used an inverted phase microscope and immunofluorescent staining to observe the morphological changes of Saos-2 cells after being cultured in different concentrations of DATS (0, 25, 50, and 100 µM) for 24 h, or for four time periods (24, 48, 72, and 96 h) in the same DATS concentration (50 µM). Quantitative real-time polymerase chain reaction and Western blot were used to detect the expression level of GRP78 mRNA and proteins in Saos-2 cells. GRP78 expression was suppressed in Saos-2 cells by utilizing small-interfering RNA, and the cells were subsequently used to study the anti-proliferative effects of DATS treatment.
Results: The expression level of GRP78 mRNA and proteins was significantly downregulated due to the increased concentration and effective times of DATS (P<0.05). In addition, there were significant associations between GRP78 silencing and cell proliferation (P<0.05) of DATS treatment.
Conclusion: These results indicate that DATS inhibits the growth of human osteosarcoma Saos-2 cells by downregulating the expression of GRP78.
Keywords: diallyl trisulfide, osteosarcoma, Saos-2, glucose-regulated protein 78
Introduction
Osteosarcoma is a primary malignant tumor of bones that originates from mesenchymal tissue, frequently occurring in young adults and adolescents.1 The 5-year survival rate without metastases for osteosarcoma patients has been increased from 10%–20% to 60%–70% due to the application of neoadjuvant chemotherapy.2–6 Although neoadjuvant chemotherapy is effective in prolonging patient survival,7,8 it often results in the acquisition of drug resistance and the occurrence of side effects such as myelosuppression, toxicity of the kidneys, heart and nervous system, and gastrointestinal reactions. Therefore, it is necessary to develop molecule-targeted drugs with a good curative effect for patients with osteosarcoma.
GRP78, also known as immunoglobulin heavy chain binding protein, is a molecular chaperone located in the endoplasmic reticulum (ER). It is a member of the 70 kDa heat shock (HSP70) protein family9 and has the function of facilitating protein folding, assembly and transport, and regulating ER stress signaling.10 During ER stress resulting from an external stimulus such as cytotoxic chemical stimulation, ultraviolet radiation, or cancers, cells initiate an unfolded protein response (UPR), which employs GRP78 as a necessary regulator to maintain the homeostasis of the ER.11,12 Recent studies showed that an overexpression of GRP78 was detected in several cancers such as hepatocellular carcinoma,13 gastric cancer,14 endometrial carcinoma,15 ovarian serous cystadenocarcinoma,16 prostate cancer,17 lung cancer,18 neuroblastoma,19 mouth cancer,20 colorectal cancer,21 and ureter tumors.22 Likewise, they showed that inhibiting expression of GRP78 can promote the apoptosis of tumor cells and slow down the growth and metastasis of tumor cells through their in vivo or in vitro experiments. Xia et al23 reported that suppressing expression of GRP78 and Akt decreased P-gp expression and mitigated chemoresistance in human osteosarcoma. Our previous study similarly implied that there was a direct association between overexpression of GRP78 and osteosarcoma growth and metastasis.24 Because of its importance for the survival of cancer cells, GRP78 could be used as an anticancer drug target.
Diallyl trisulfide (DATS) has been reported to possess anticancer activities. However, its cancer growth inhibitory effects and molecular mechanisms in human osteosarcoma cells have not been well and fully studied. Our previous proteomic study showed that DATS has the ability to suppress cell proliferation of Saos-2 cells by blocking cell cycle progression and inducing apoptosis in a dose- and time-dependent manner.25 In addition, DATS-sensitive proteins, including 18 downregulated proteins and nine upregulated proteins, were detected. Li et al26 implied that DATS inhibited proliferation, invasion, and angiogenesis of osteosarcoma cells by switching on suppressor microRNAs and inactivating Notch-1 signaling. Wang et al27 indicated that DATS induced osteosarcoma cell apoptosis through reactive oxygen species-mediated downregulation of the PI3K/Akt pathway. However, the exact underlying molecular mechanisms are still unclear. Now the challenge is how to elucidate the antineoplastic mechanisms of DATS, which may be conducive to the development of chemotherapy for osteosarcoma. In this study, we aimed to observe the inhibitory effects of DATS on human osteosarcoma Saos-2 cells and to investigate the expression of GRP78 in osteosarcoma Saos-2 cells after they were treated with different concentrations of DATS for various time periods.
Materials and methods
Materials
DATS (98% pure) was purchased from LKT Laboratories (Minneapolis, MN, USA). DMEM media and fetal bovine serum (FBS) were obtained from Gibco BRL (Grand Island, NY, USA). The rabbit anti-human polyclonal GRP78 antibody and mouse anti-human polyclonal β-actin were purchased from AbCam (Cambridge, UK). The fluorescein isothiocyanate (FITC)-labeled goat anti-rabbit immunoglobulin G [IgG; heavy and light (H+L) chain] was obtained from EarthOx Life Sciences (Millbrae, CA, USA). HRP-conjugated goat anti-rabbit and rat anti-mouse IgG were obtained from SCBT (Santa Cruz, CA, USA). Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and synthesized into cDNA using the PrimScript RT reagent kit (TaKaRa, Dalian, People’s Republic of China). The Bradford protein assay kit was purchased from Bio-Rad Laboratories (Hercules, CA, USA). The primers were GRP78 forward: 5′-CGTCCTATGTCGCTTCACT-3′, reverse: 5′-TGTCTTTGTTTGCCCACCTC-3′.
Cell culture
Human osteosarcoma Saos-2 cell lines were obtained from the Institute of Biochemistry and Cell Biology at the Chinese Academy of Sciences (Shanghai, People’s Republic of China). Cells were cultured in DMEM medium supplemented with 10% FBS at 37°C in humidified air containing 5% CO2. The cells were used in the logarithmic phase of growth throughout the experiment.
Immunofluorescent staining
Different concentrations of DATS (0, 25, 50, and 100 μM) were cultured for 24 h, and four time periods (24, 48, 72, and 96 h) in the same concentration of DATS (50 μM). Saos-2 cells growing on 24-well culture plates were washed three times in PBS and fixed in PBS containing 4% formaldehyde for 30 min. Then the cells were washed twice in PBS and incubated for 30 min in 0.1% Triton X-100 in PBS. Next, the cells were blocked with 200 μL 10% normal goat serum (diluted with PBS) per well and incubated for 30 min at room temperature. Then, they were incubated with 200 μL primary anti-GRP78 antibody (dilution, 1:1,000) per well in a moist chamber at 4°C overnight. The cells were then washed three times with PBS and incubated with the goat anti-rabbit IgG/FITC antibody (dilution, 1:200) at room temperature for 30 min. Following this they were blocked with glycerol, followed by incubation with DAPI for 5 min at room temperature.
The samples were analyzed using a Leica DM4000B microscope (Leica Microsystems GmbH, Wetzlar, Germany), and images were captured using the Image Pro Plus image analysis system 7.0 (Media Cybernetics, Inc., Rockville, MD, USA) in order to detect the expression level. Cells in which the cytoplasm was stained green were considered to be GRP78 positive cells.
Quantitative real-time polymerase chain reaction (qRT-PCR)
All qRT-PCRs were performed using SYBR Premix Ex Taq II (TaKaRa) and measured in a LightCyler 480 system (Roche, Basel, Switzerland). The levels of β-actin were used as an internal control. All of the reactions were run in triplicate. All data output from the qRT-PCR experiments were analyzed by using the 2−ΔΔCT method (cells were cultured in the same way as mentioned in the “Immunofluorescent staining” section).
Western blot analysis
Protein was extracted from cell lines using RIPA buffer containing phenylmethanesulfonyl fluoride. A bicinchoninic acid protein assay kit (Beyotime, Haimen, People’s Republic of China) was used to confirm the total protein concentration. Protein samples were separated by SDS-PAGE and transferred to polyvinyl membranes. Membranes were incubated overnight at 4°C with the primary anti-GRP78 antibody (dilution, 1:1,000), followed by the secondary antibody, HRP-conjugated goat anti-rabbit IgG (dilution, 1:20,000). The formula of the expression level of GRP78 protein was as follows: expression level of GRP78 protein = value of GRP78 protein in samples/value of β-actin in samples (cells were cultured in the same way as mentioned in the “Immunofluorescent staining” section).
Signals were observed by using a chemiluminescent detection kit (Amersham Pharmacia Biotech, Piscataway, NJ, USA). The antibody against β-actin functioned as an internal reference. Typhoon PhosphorImager (GE Healthcare, Piscataway, NJ, USA) was used to measure the quality of the protein.
siRNA and cell transfections
Saos-2 cells were grown to 70% confluence in 60-mm culture plates and washed twice with PBS. The cells were transfected for 48 h with 100 nM SMART pool siRNAs specific to non-targeting siRNA (scrambled siRNA) or GRP78 mRNA using Lipofectamine 2000 reagent (Thermo Fisher Scientific, Waltham, MA, USA) in serum-free DMEM, according to the manufacturer’s protocols.
MTT assay
Saos-2 cells were cultured in a 96-well culture plate with 5,000 cells/well. Cells were then exposed to scr-siRNA or GRP78-siRNA. The cells were treated with 50 μM DATS for 24 h, and then a cell survival assay was performed.
Statistical analysis
The data were collected and shown as mean ± SD of at least three independent experiments. Statistical analysis was performed by one-way ANOVA followed by a post hoc comparison using the Bonferroni test. P<0.05 was considered to be statistically different.
Results
Immunofluorescent staining
GRP78 was mainly located in the endoplasmic reticulum (ER). Cells in which ER was stained green were considered to be GRP78 positive cells. Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; blue). Fluorescence intensity of GRP78 in Saos-2 cells cultured in different concentrations of DATS for 24 h or in the same concentration of DATS (50 μM) for four time periods is shown in Figure 1.
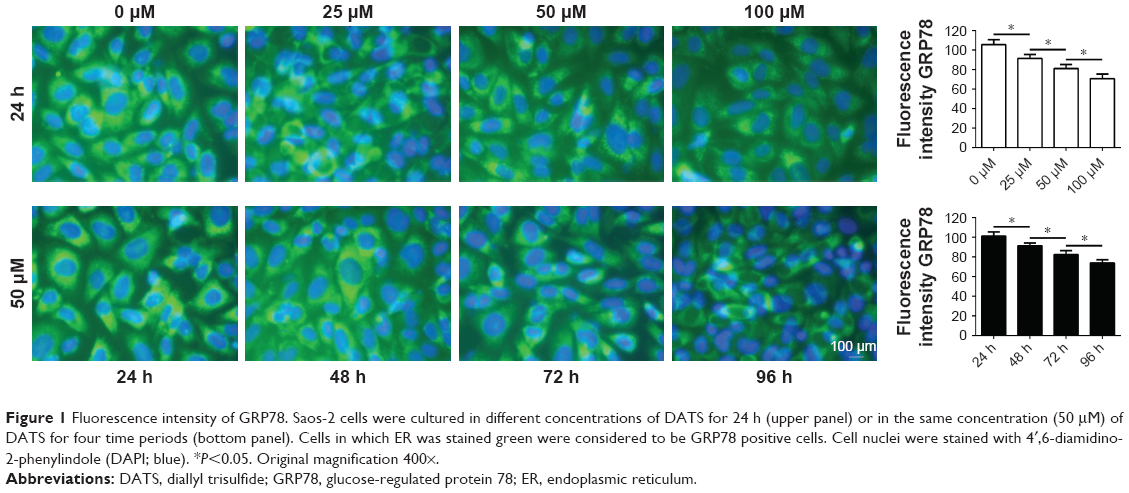 | Figure 1 Fluorescence intensity of GRP78. Saos-2 cells were cultured in different concentrations of DATS for 24 h (upper panel) or in the same concentration (50 μM) of DATS for four time periods (bottom panel). Cells in which ER was stained green were considered to be GRP78 positive cells. Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; blue). *P<0.05. Original magnification 400×. |
qRT-PCR
GRP78 mRNA was identified in all samples. Relative expression of GRP78 mRNA in Saos-2 cells cultured in different concentrations of DATS for 24 h or in the same concentration of DATS (50 μM) for four time periods is shown in Figure 2.
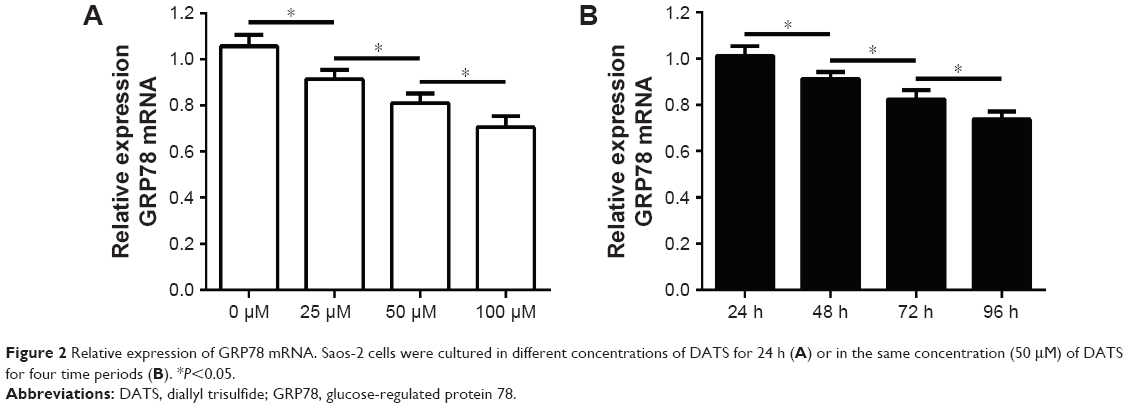 | Figure 2 Relative expression of GRP78 mRNA. Saos-2 cells were cultured in different concentrations of DATS for 24 h (A) or in the same concentration (50 μM) of DATS for four time periods (B). *P<0.05. |
Western blot analysis
GRP78 protein was expressed in all samples. The relative intensity of GRP78 protein in Saos-2 cells cultured in different concentrations of DATS for 24 h or in the same concentration of DATS (50 μM) for four time periods is shown in Figure 3.
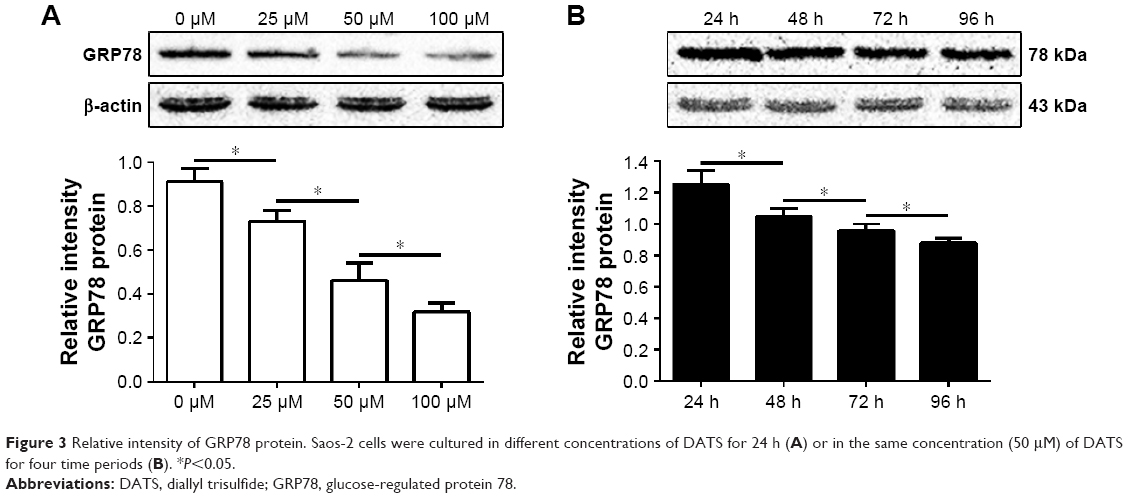 | Figure 3 Relative intensity of GRP78 protein. Saos-2 cells were cultured in different concentrations of DATS for 24 h (A) or in the same concentration (50 μM) of DATS for four time periods (B). *P<0.05. |
GRP78 silencing and cell proliferation
Before evaluating potential antiproliferative effects of DATS on cell proliferation, the efficiency of GRP78 silencing was verified by Western blot analysis, which demonstrated a significant reduction of GRP78 expression in cells transfected with GRP78-siRNA (Figure 4). To assess the anti-proliferative effects of DATS on cell proliferation, MTT assay was performed, and there were significant associations between GRP78 silencing and cell proliferation of DATS treatment (Figure 4).
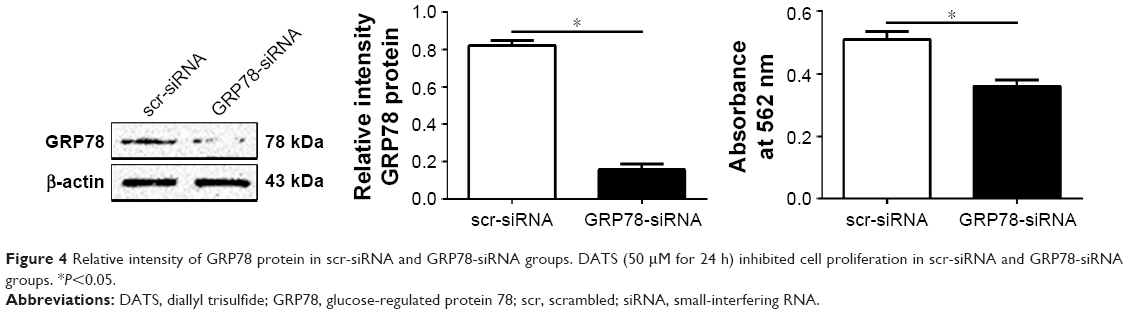 | Figure 4 Relative intensity of GRP78 protein in scr-siRNA and GRP78-siRNA groups. DATS (50 μM for 24 h) inhibited cell proliferation in scr-siRNA and GRP78-siRNA groups. *P<0.05. |
Discussion
DATS has a significant anticancer effect on multiple cancers, as demonstrated in previous epidemiological and experimental studies. It blocks the tumor cell proliferation cycle and triggers cell death by inducing apoptosis. These studies were mainly about malignancies such as prostate cancer,28 gastric cancer,29 lung cancer,30 breast cancer,31 and colon cancer,32 while research on the effect and mechanism of DATS against osteosarcoma was rare. Our previous study showed that DATS could induce cell apoptosis in a dose- and time-dependent manner through regulating expression of some specific proteins.25 In addition, a direct association between over-expression of GRP78 and osteosarcoma growth and metastasis came to light through our other previous study.24 Therefore, we continued to conduct experimental studies in order to elucidate the mechanism and target of DATS on growth of human osteosarcoma on the basis of our previous research results.
In this study, we proposed and demonstrated the hypothesis, for the first time, that DATS could damage the normal structure of cells and inhibit the growth of osteosarcoma by downregulating expression of GRP78. Furthermore, our results showed that DATS downregulated the expression level of GRP78 in a dose- and time-dependent manner. Over-expression of GRP78 is known to correlate with the risk of tumor progression and metastasis, as reported in many articles.13–22 Combining these reports and our previous results, we believed that DATS might inhibit the growth of osteosarcoma by downregulating expression of GRP78. We verified our hypothesis by a series of experiments such as qRT-PCR and Western blot. Our experimental results are consistent with our hypothesis, so we report, for the first time, that DATS can inhibit the growth of osteosarcoma cells by downregulating expression of GRP78. This is the possible mechanism and molecular target of DATS in the treatment of osteosarcoma. Therefore, downregulating expression of GRP78 with DATS treatment could be a possible therapeutic target for better management of osteosarcoma growth.
GRP78 is associated with tumor cell proliferation, apoptosis resistance, and metastasis, and plays an important role in the occurrence and apoptosis of tumors.33 It can regulate several cancer-associated inflammatory cytokines, such as interleukin-6 (IL-6),34 macrophage migration inhibitory factor (MIF),35 transformed growth factor β (TGF-β),36 and interleukin-10 (IL-10).37 In addition, over-expression of GRP78 in poorly perfused solid tumors usually results from the ER stress pathway activated by glucose deprivation, hypoxia, and acidosis.10 Therefore, downregulation or inhibition of GRP78 expression can promote apoptosis of tumor cells and prevent tumor growth and metastasis. Although some previous investigations found that DATS could inhibit the growth of osteosarcoma cells, the mechanism and target were not clear. Inspired by this, we conducted an in-depth study, in order to investigate the mechanism and target of DATS in the treatment of osteosarcoma cells.
In our experiment, the inhibitory effects on the cell growth curve and the morphological changes of Saos-2 cells were observed in a dose- and time-dependent manner after treatment with DATS. Simultaneously, the expression level of GRP78 was downregulated in a dose- and time-dependent manner. As previously stated, we have shown for the first time that DATS can inhibit the growth of osteosarcoma cells by downregulating expression of GRP78.
Conclusion
This study revealed the novel finding of the mechanism and molecular targets of DATS on the growth of osteosarcoma cells. These results indicate that DATS inhibits the growth of human osteosarcoma Saos-2 cells by downregulating expression of GRP78. The inhibitory effects of DATS on GRP78 expression of osteosarcoma cells provide a basis for the clinical use of DATS and supply a novel alternative molecular target for the treatment of osteosarcoma. However, further in-depth preclinical experiments using relevant animal models are required.
Acknowledgment
The authors thank Dr Edward C Mignot, Shandong University, for linguistic advice.
This study was funded by the National Science Foundation of Shandong Province (number ZR2014HQ034) and the Medical Science Development Project of Shandong Province (number 2014WS0432).
Disclosure
The authors report no conflicts of interest in this work.
References
Wang H, Sun N, Li X, Li K, Tian J, Li J. Diallyl trisulfide induces osteosarcoma cell apoptosis through reactive oxygen species-mediated downregulation of the PI3K/Akt pathway. Oncol Rep. 2016;35(6):3648–3658. | ||
Ferrari S, Serra M. An update on chemotherapy for osteosarcoma. Expert Opin Pharmacother. 2015;16(18):2727–2736. | ||
Bielack S, Kempf-Bielack B, Von Kalle T, et al. Controversies in childhood osteosarcoma. Minerva Pediatr. 2013;65(2):125–148. | ||
Bielack SS, Hecker-Nolting S, Blattmann C, Kager L. Advances in the management of osteosarcoma. F1000Res. 2016;5:2767. | ||
Petrilli AS, de Camargo B, Filho VO, et al. Results of the Brazilian Osteosarcoma Treatment Group Studies III and IV: prognostic factors and impact on survival. J Clin Oncol. 2006;24(7):1161–1168. | ||
Wang W, Yang J, Wang Y, et al. Survival and prognostic factors in Chinese patients with osteosarcoma: 13-year experience in 365 patients treated at a single institution. Pathol Res Pract. 2017;213(2):119–125. | ||
Vetterlein MW, Wankowicz SAM, Seisen T, et al. Neoadjuvant chemotherapy prior to radical cystectomy for muscle-invasive bladder cancer with variant histology. Cancer. 2017;123(22):4346–4355. | ||
Guo J, Glass JO, McCarville MB, et al. Assessing vascular effects of adding bevacizumab to neoadjuvant chemotherapy in osteosarcoma using DCE-MRI. Br J Cancer. 2015;113(9):1282–1288. | ||
Xia YZ, Yang L, Xue GM, et al. Combining GRP78 suppression and MK2206-induced Akt inhibition decreases doxorubicin-induced P-glycoprotein expression and mitigates chemoresistance in human osteosarcoma. Oncotarget. 2016;7(35):56371–56382. | ||
Li Z, Li Z. Glucose regulated protein 78: a critical link between tumor microenvironment and cancer hallmarks. Biochim Biophys Acta. 2012;1826(1):13–22. | ||
Kim SR, Kim DI, Kang MR, et al. Endoplasmic reticulum stress influences bronchial asthma pathogenesis by modulating nuclear factor κB activation. J Allergy Clin Immunol. 2013;132(6):1397–1408. | ||
Gray MJ, Mhawech-Fauceglia P, Yoo E, et al. AKT inhibition mitigates GRP78 (glucose-regulated protein) expression and contribution to chemoresistance in endometrial cancers. Int J Cancer. 2013;133(1):21–30. | ||
Chen WT, Zhu G, Pfaffenbach K, Kanel G, Stiles B, Lee AS. GRP78 as a regulator of liver steatosis and cancer progression mediated by loss of the tumor suppressor PTEN. Oncogene. 2014;33(42):4997–5005. | ||
Zhang J, Jiang Y, Jia Z, et al. Association of elevated GRP78 expression with increased lymph node metastasis and poor prognosis in patients with gastric cancer. Clin Exp Metastasis. 2006;23(7–8):401–410. | ||
Calì G, Insabato L, Conza D, et al. GRP78 mediates cell growth and invasiveness in endometrial cancer. J Cell Physiol. 2014;229(10):1417–1426. | ||
Fan LM, Su J, Dong H, Wei M, Cui MH. [Expression and role of glucose-regulated protein 78 in ovarian serous adenocarcinoma]. Zhonghua Yi Xue Za Zhi. 2013;93(9):659–662. Chinese. | ||
Wu CT, Wang WC, Chen MF, et al. Glucose-regulated protein 78 mediates hormone-independent prostate cancer progression and metastasis through maspin and COX-2 expression. Tumour Biol. 2014;35(1):195–204. | ||
Koomägi R, Mattern J, Volm M. Glucose-related protein (GRP78) and its relationship to the drug-resistance proteins P170, GST-pi, LRP56 and angiogenesis in non-small cell lung carcinomas. Anticancer Res. 1999;19(5B):4333–4336. | ||
Hsu WM, Hsieh FJ, Jeng YM, et al. GRP78 expression correlates with histologic differentiation and favorable prognosis in neuroblastic tumors. Int J Cancer. 2005;113:920–927. | ||
Lin CY, Chen WH, Liao CT, et al. Positive association of glucose-regulated protein 78 during oral cancer progression and the prognostic value in oral precancerous lesions. Head Neck. 2010;32(8):1028–1039. | ||
Li Z, Zhang L, Zhao Y, et al. Cell-surface GRP78 facilitates colorectal cancer cell migration and invasion. Int J Biochem Cell Biol. 2013;45(5):987–994. | ||
Park CH, Choi MS, Ha JY, Kim BH, Park CH, Kim CI. Effect of overexpression of glucose-regulated protein 78 and bcl-2 on recurrence and survival in patients with ureter tumors. Korean J Urol. 2013;54(10):671–676. | ||
Xia YZ, Yang L, Xue GM, et al. Combining GRP78 suppression and MK2206-induced Akt inhibition decreases doxorubicin-induced P-glycoprotein expression and mitigates chemoresistance in human osteosarcoma. Oncotarget. 2016;7(35):56371–56382. | ||
Zhang Y, Li N, Wang D, Chen Y, Li G. Expression and significance of glucose-regulated protein 78 in human osteosarcoma. Oncol Lett. 2015;9(5):2268–2274. | ||
Zhang YK, Zhang XH, Li JM, Sun DS, Yang Q, Diao DM. A proteomic study on a human osteosarcoma cell line Saos-2 treated with diallyl trisulfide. Anticancer Drugs. 2009;20(8):702–712. | ||
Li Y, Zhang J, Zhang L, Si M, Yin H, Li J. Diallyl trisulfide inhibits proliferation, invasion and angiogenesis of osteosarcoma cells by switching on suppressor microRNAs and inactivating of Notch-1 signaling. Carcinogenesis. 2013;34(7):1601–1610. | ||
Wang H, Sun N, Li X, Li K, Tian J, Li J. Diallyl trisulfide induces osteosarcoma cell apoptosis through reactive oxygen species-mediated downregulation of the PI3K/Akt pathway. Oncol Rep. 2016;35(6):3648–3658. | ||
Borkowska A, Knap N, Antosiewicz J. Diallyl trisulfide is more cytotoxic to prostate cancer cells PC-3 than to noncancerous epithelial cell line PNT1A: a possible role of p66Shc signaling axis. Nutr Cancer. 2013;65(5):711–717. | ||
Pan Y, Lin S, Xing R, et al. Epigenetic upregulation of metallothionein 2A by diallyl trisulfide enhances chemosensitivity of human gastric cancer cells to docetaxel through attenuating NF-κB activation. Antioxid Redox Signal. 2016;24(15):839–854. | ||
Li W, Tian H, Li L, et al. Diallyl trisulfide induces apoptosis and inhibits proliferation of A549 cells in vitro and in vivo. Acta Biochim Biophys Sin (Shanghai). 2012;44(7):577–583. | ||
Kim SH, Kaschula CH, Priedigkeit N, Lee AV, Singh SV. Forkhead box Q1 is a novel target of breast cancer stem cell inhibition by diallyl trisulfide. J Biol Chem. 2016;291(26):13495–13508. | ||
Lai KC, Hsu SC, Yang JS, Yu CC, Lein JC, Chung JG. Diallyl trisulfide inhibits migration, invasion and angiogenesis of human colon cancer HT-29 cells and umbilical vein endothelial cells, and suppresses murine xenograft tumour growth. J Cell Mol Med. 2015;19(2):474–484. | ||
Ramsay RG, Ciznadija D, Mantamadiotis T, Anderson R, Pearson R. Expression of stress response protein glucose regulated protein-78 mediated by c-Myb. Int J Biochem Cell Biol. 2005;37(6):1254–1268. | ||
Hori O, Matsumoto M, Kuwabara K, et al. Exposure of astrocytes to hypoxia/reoxygenation enhances expression of glucose-regulated protein 78 facilitating astrocyte release of the neuroprotective cytokine interleukin 6. J Neurochem. 1996;66(3):973–979. | ||
Vera PL, Wang X, Bucala RJ, Meyer-Siegler KL. Intraluminal blockade of cell-surface CD74 and glucose regulated protein 78 prevents substance P-induced bladder inflammatory changes in the rat. PLoS One. 2009;4(6):e5835. | ||
Oida T, Weiner HL. Overexpression of TGF-β 1 gene induces cell surface localized glucose-regulated protein 78-associated latency-associated peptide/TGF-β. J Immunol. 2010;185(6):3529–3535. | ||
Chen YY, Zheng MZ, Lv PP, Hu L, Wang LL, Shen YL. Hydrogen peroxide regulates glucose-regulated protein 78 expression via a cyclooxygenase-2 dependent mechanism. J Biochem Mol Toxicol. 2010;24(5):279–285. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.