Back to Journals » Journal of Inflammation Research » Volume 19
Exosomes Regulate Liver Diseases Through the TGF-β1/Smads Signaling Pathway and the Intervention of Traditional Chinese Medicine: A Narrative Review
Received 17 October 2025
Accepted for publication 7 January 2026
Published 30 January 2026 Volume 2026:19 572487
DOI https://doi.org/10.2147/JIR.S572487
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Fatih Türker
Guidong Huang,1 Zhipin Zhou2
1Department of Pharmacy, Maternal and Child Health Hospital of Guangxi Zhuang Autonomous Region, Nanning, People’s Republic of China; 2Department of Pharmacy, Liuzhou People’s Hospital, Liuzhou, People’s Republic of China
Correspondence: Zhipin Zhou, Department of Pharmacy, Liuzhou People’s Hospital, Liuzhou, 545006, People’s Republic of China, Email [email protected]
Abstract: Liver disease is a common chronic illness with increasing incidence and mortality. Exosomes are small membrane vesicles (30– 200 nm) containing complex RNAs and proteins that serve as mediators of intercellular communication. The role of exosomes in liver disease is complex and sometimes contradictory. In certain situations, they may suppress liver disease, whereas in others, they promote its progression. This duality raises important questions regarding the overall impact of these drugs on the treatment of liver disease. The Transforming Growth Factor-beta1 (TGF-β 1)/Smads signaling pathway plays a significant role in the occurrence and development of liver diseases, and exosomes mediate liver disease by regulating the TGF-β 1/Smads signaling pathway. Research has shown that Traditional Chinese Medicine (TCM) can exert liver-protective effects by influencing exosomes to modulate the TGF-β 1/Smads signaling pathway. Using keywords such as “exosomes”, “TGF-β 1/Smads signaling pathway”, “liver fibrosis”, “hepatocellular carcinoma”, “liver diseases”, and “traditional Chinese medicine”, this article reviews the literature from databases, including CNKI, PubMed, and SCOPUS. It summarizes how exosome-mediated regulation of the TGF-β 1/Smads signaling pathway affects the development of liver diseases and compiles relevant studies on the anti-liver disease effects of TCM through exosomal modulation of this pathway.
Keywords: exosome, TGF-β 1/Smads signaling pathway, liver disease, traditional Chinese medicine
The liver, as a central organ in material metabolism and energy conversion, is essential for maintaining bodily homeostasis. The incursion of various factors can lead to liver damage. The inflammatory infiltration resulting from liver damage induces alterations in liver cell structure and necrosis, which, in severe instances, may progress to hepatitis, liver fibrosis, and even liver cancer.1 It has been reported that chronic liver diseases account for 3.5% of the global mortality rate. Clearly, liver diseases have emerged as a significant global health threat.2 In light of this trend, the pursuit of a safe and effective therapeutic approach has emerged as a paramount concern within the field of biomedicine. TCM offers notable advantages in both the prevention and treatment of diseases, demonstrating significant therapeutic efficacy across various conditions. Exosomes and their components are pivotal in the pathogenesis and progression of diseases, primarily through the regulation of gene expression and cellular signal transduction. Current research indicates that the exosome-mediated transfer of RNA, miRNAs, proteins, and lipids is intricately linked to the onset and progression of diverse liver diseases.3 The TGF-β1/Smads signaling pathway is recognized as a critical pathway in liver pathologies. Modulating the release of exosomes via TCM and its active constituents may represent a novel therapeutic avenue for liver diseases, as illustrated in Figure 1.
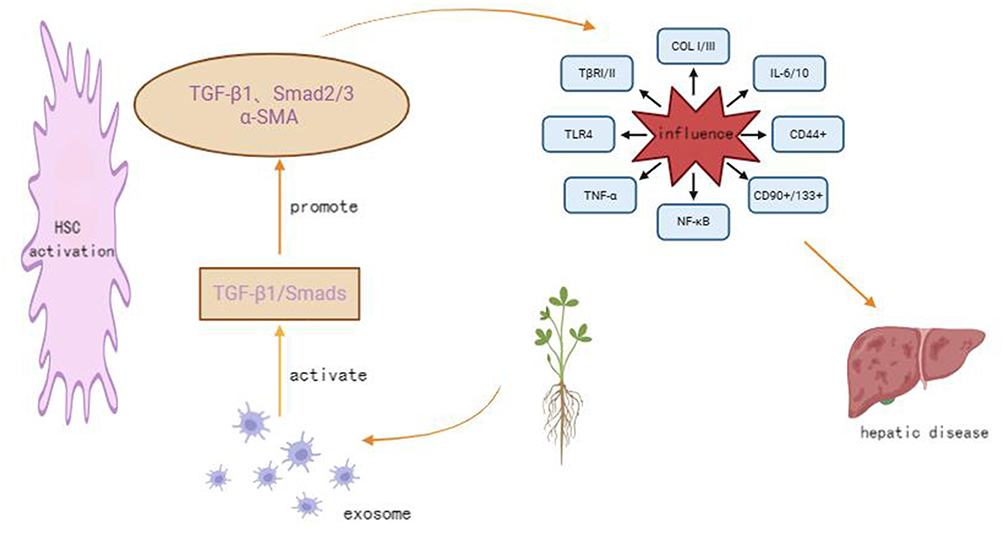 |
Figure 1 Shows that traditional Chinese medicine inhibits the activation of the TGF-β1/Smads signaling pathway by suppressing exosomes, thereby intervening in liver diseases. |
Exosomes
Exosomes are membrane-bound vesicles encapsulated within a phospholipid bilayer, with diameters ranging from 30 to 200 nm. They are secreted by nearly all cell types and are present in all biological fluids.4 Exosome secretion occurs under both physiological and pathological conditions, and their formation is intricately linked to the multivesicular endosomal pathway. The biogenesis of exosomes involves two endocytic processes. Initially, the cell membrane encapsulates surface proteins and soluble proteins from the extracellular environment, forming an early endosome. Subsequently, the cell membrane selectively encapsulates intracellular molecules, undergoing another endocytic process to form intracellular multivesicular bodies. These bodies may then fuse with lysosomes or the cell membrane, releasing numerous small vesicles known as exosomes,5 as illustrated in Figure 2. Exosomes facilitate the transport of their contents to the cytoplasm of target cells through three mechanisms: direct targeting of cell receptor signals, fusion with the target cell’s cytoplasmic membrane, and internalization into the target cell, thereby enabling intercellular material exchange.6,7 As mediators of intercellular communication, exosomes, when internalized by recipient cells, deliver various bioactive substances, including DNA, mRNA, lipids, miRNAs, and functional proteins. These substances play a crucial role in intercellular communication and can alter the phenotype of recipient cells through a paracrine mechanism mediated by exosomes.8
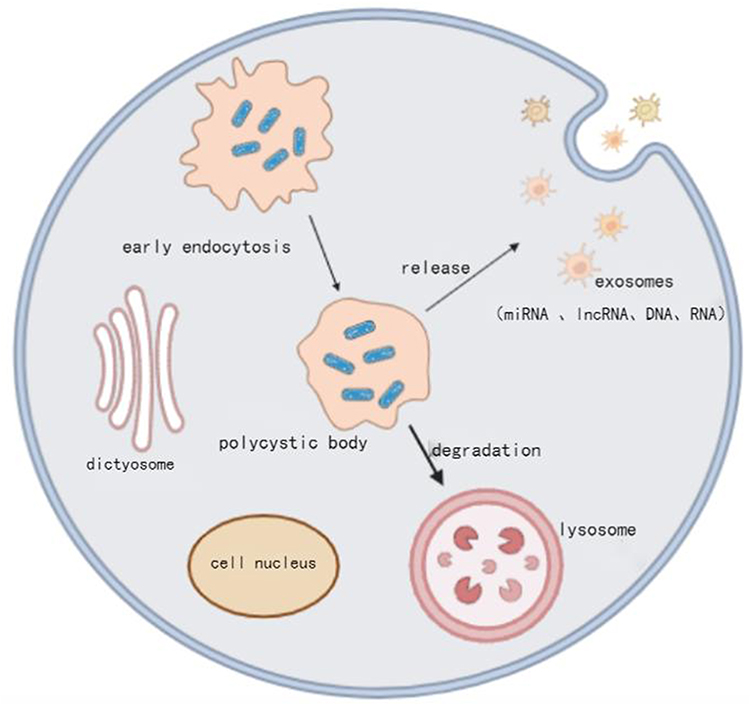 |
Figure 2 The Formation Process of Exosomes. |
Exosomes perform their physiological functions upon binding to target cells. Exosomes originating from various cellular sources exhibit specificity in their interactions with target cells, thereby performing distinct physiological roles. They can modulate a range of pathological and physiological processes, including immune responses, inflammatory reactions and immune evasion mechanisms. The discovery of exosomes was first made by Pan and Johnstone in 1983 within the reticular cells of sheep.9 Initially, exosomes were perceived as cellular mechanisms for waste excretion. However, Raposo et al highlighted their involvement in antigen presentation and adaptive immune responses,10 which subsequently drew significant research attention to exosomes. Further investigations have demonstrated that exosomes play a role in tumor immune evasion and chronic inflammatory responses.11 Additionally, exosomes harbor valuable biomarkers that are of substantial significance in clinical disease treatment.
TGF-β1/Smads Signaling Pathway
The TGF-β1/Smads signaling pathway is the mechanism through which TGF-β1 transmits signals via the Smad protein family to exert its biological effects. This pathway comprises the TGF-β superfamily and Smad proteins. Intracellularly, TGF-β1 exists in its inactive form. Upon binding to the type II receptor (TβRII) on the cell membrane, extracellular TGF-β1 activates this receptor. The activated TβRII subsequently recruits the TβRI, forming a heterotrimer consisting of the TβRII, ligand, and TβRI. Concurrently, the TβRII phosphorylates the kinase domain of the TβRI, which in turn phosphorylates Smad2 and Smad3. Phosphorylated Smad2/Smad3 (p-Smad2/Smad3) complexes with Smad4 to form a heterotrimer that translocates into the cell nucleus, thereby initiating transcription. Only p-Smad3 can transmit the TGF-β1 stimulation signal from the cytoplasm to the nucleus, thereby exerting biological effects. Smad7 functions as an inhibitory factor by counteracting the TGF-β1 signaling pathway and preventing the binding of Smad4 to Smad2/3. As an inhibitor of TGF-β1 signal transduction, Smad7 can bind to TβRI, thereby obstructing the phosphorylation of Smad2/3.12 This is illustrated in Figure 3.
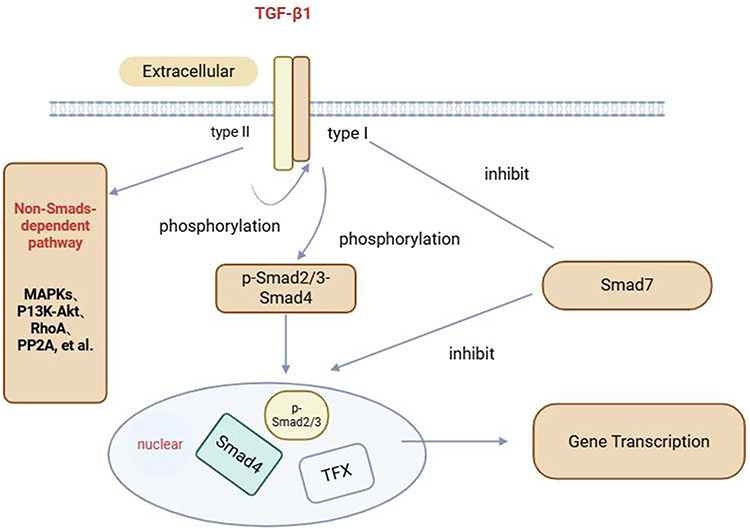 |
Figure 3 Schematic diagram of the TGF-β1/Smads signaling pathway. |
TGF-β is integral to maintaining the equilibrium of early embryonic development and adult physiological processes. It is intricately associated with epithelial-mesenchymal transition (EMT), extracellular matrix (ECM) deposition, and the formation of cancer-associated fibroblasts.12 The TGF-β signaling pathway is significantly implicated in the progression of liver diseases, spanning from the initial inflammatory response due to chronic liver injury to the activation of hepatic stellate cells (HSC) that secrete ECM, leading to liver fibrosis and eventually to the advanced stages of liver cirrhosis and liver cancer.13 Given its role in HSC activation and ECM deposition, TGF-β is recognized as the primary pro-fibrotic cytokine,14 with TGF-β1 being its predominant isoform. Smads, which serve as kinase substrates for TGF-β receptors, are involved in intracellular TGF-β signal transduction and regulate the ECM through nuclear transcription.15 The TGF-β1/Smads signaling pathway is pivotal in HSC activation and the progression of chronic liver diseases. Liver cells, including parenchymal, Kupffer, and sinusoidal endothelial cells, produce TGF-β1 via paracrine or HSC autocrine mechanisms, which in turn activates HSC, resulting in liver inflammation and damage. This process promotes ECM production via the TGF-β1/Smads signaling pathway, thereby contributing to the development of chronic liver diseases.16
Exosomes Mediating the TGF-β1/Smads Signaling Pathway
Exosomes Mediating the TGF-β1/Smads Signaling Pathway and Liver Fibrosis
Liver fibrosis is a cyclical process characterized by continuous hepatic damage, resulting in hepatocyte injury, necrosis, and alterations in the hepatic microenvironment. The underlying mechanisms are complex and multifaceted in nature. Pro-inflammatory factors, apoptotic bodies of hepatocytes, growth factors mediated by endothelial cells, and reactive oxygen species generated by damaged hepatocytes and macrophages can perpetually activate HSC, leading to excessive ECM deposition and the development of liver fibrosis.17 Liu et al demonstrated that exosomes can modulate the TGF-β1/Smads signaling pathway, thereby contributing to the pathogenesis of liver fibrosis.18 TGF-β1 is a pivotal factor in promoting HSC activation, and the TGF-β1/Smads signaling pathway is critical for ECM expression and production. Exosomes not only facilitate HSC activation by acting as pro-fibrotic agents but also exert an anti-fibrotic effect by targeting TGF-β rece.
Exosomes Mediate TGF-β1/Smads Signaling Pathway to Promote Liver Fibrosis
Exosomes from various sources have been shown to activate the TGF-β1/Smads signaling pathway, thereby activating HSC and facilitating liver fibrosis. Specifically, exosomes derived from macrophages containing NEAT1 have been observed to elevate the levels of TGF-β1 and phosphorylated Smad2/3 in LX-2 cells, leading to HSC activation. Further investigations have revealed that NEAT1 contributes to HSC activation through the phagocytosis of miRNA-342, which subsequently induces the downstream Sp1/TGF-β1/Smad signaling pathway, thereby promoting liver fibrosis.19 Additionally, research has identified that miRNA-33 from Schistosoma japonicum egg-derived exosomes enhances the expression of α-SMA, COLI α1, and COLIII α1 in LX-2 cells via the TGF-β1 pathway, thus advancing liver fibrosis progression.20 In HSC expressing activated TGF-β1, exosomal miRNA-497 was upregulated. Studies have demonstrated that exosomal miRNA-497 targets Smad7, facilitating the transduction of the TGF-β1/Smad signaling pathway, which increases the level of phosphorylated Smad2/3, promotes HSC activation, and contributes to liver fibrosis.21 TβRIII serves as a negative regulator of TGF-β signaling and is directly targeted by sja-miRNA-2162 in hematopoietic stem cells. An increase in the secretion of sja-miRNA-2162 from parasites or plants acts on TβRIII to promote liver fibrosis.22 Furthermore, miRNA-139-5p of the lncRNA NEAT1 sponge can inhibit the expression of β-catenin, which interacts with SOX9 to promote the activation of hematopoietic stem cells. SOX9 can bind to the TGF-β1 promoter, thereby accelerating the transcriptional activity of TGF-β1 and promoting liver fibrosis.23
Exosomes Mediate the TGF-β1/Smads Signaling Pathway for Anti-Liver Fibrosis
TGF-β1 is the primary factor responsible for ECM accumulation, and disruption of the signal transduction involving TGF-β1 can effectively prevent tissue fibrosis.24 The TGF-β receptors, TβRI and TβRII, are the principal proteins that mediate the transport of TGF-β1 from the extracellular environment into the cells. Exosomes can exert anti-liver fibrosis effects by targeting TβRI/II. In TGF-β1-activated LX-2 cells, exosomes derived from human embryonic stem cells (HESC) carrying miRNA-6766-3p target TβRII and inhibit the TGF-β1/Smads pathway, thereby reducing COL I and α-SMA expression, attenuating HSC activation, and inhibiting liver fibrosis.25 Exosomes derived from mesenchymal stem cells (MSC) transport Let-7b to HSC, downregulating TβRI expression, inhibiting COL I synthesis, and alleviating liver fibrosis.26 In a thioacetamide-induced liver cirrhosis rat model, exosomes derived from HESC increased the levels of anti-inflammatory cytokines TGF-β1 and interleukin (IL)-10, while reducing the levels of tumor necrosis factor (TNF)-α and IL-2, thereby inhibiting the inflammatory response and promoting liver recovery.27 Exosomes derived from MSC inhibit the TGF-β1 signaling pathway, reduce p-Smad2, α-SMA, and COL I levels, inhibit HSC activation, and consequently reduce ECM deposition and liver fibrosis.28,29 Li et al found that exosomes derived from human umbilical cord MSC not only improved inflammation and collagen deposition in the liver induced by CCl4 fibrosis but also inhibited TGF-β1-induced EMT in HL7702 cells. This mechanism involves the inhibition of the TGF-β1/Smad2 signaling pathway, reducing the levels of COL I/III and ECM markers, E/N-cadherin and vimentin-positive cells.30 TGF-β1 can induce mouse HSC activation, and exosomes derived from MSC miRNA-181-5p inhibit HSC activation, reduce the production of pro-fibrotic proteins COL I and α-SMA, and thereby alleviate liver collagen deposition and improve liver fibrosis.31 Exosomes derived from MSCs carrying miRNA-618 have an inhibitory effect on the liver fibrosis model induced by TGF-β1 activation of LX-2 cells, reducing the activated phenotype of liver fibrosis.32 Exosomes released by natural killer cells can inhibit TGF-β1-induced activation of HSC, reduce collagen deposition and hepatocyte necrosis, and reverse liver fibrosis.33 Similarly, studies have shown that exosomes derived from natural killer cell miRNA-223 inhibit TGF-β1-induced activation of HSC and prevent liver fibrosis.34 Wang et al demonstrated that exosomes derived from Schistosoma japonicum eggs, Sja-miRNA-71a, can improve liver fibrosis. This mechanism involves directly targeting the 4D signaling protein and inhibiting the TGF-β1/Smad/IL-13/signal transducer and activator of transcription (STAT)-6 pathway, thereby reducing the release of α-SMA and COL I.35 In summary, exosomes from various sources primarily inhibit HSC activation by blocking the TGF-β1/Smad signaling pathway and reducing the release of collagen and α-SMA, thereby exerting anti-liver fibrosis/chronic liver disease effects.
Exosomes Mediate TGF-β1/Smads Signaling Pathway and Liver Cancer
Liver cancer represents an advanced stage of chronic liver disease and ranks as the third most lethal cancer in clinical practice. The global incidence of liver cancer is progressively increasing.36 Recent advancements in understanding the regulatory mechanisms of cellular vesicle transport have highlighted the significance of exosomes in tumor invasion and metastasis. Deng Jinghuan et al have confirmed that exosomes present in the serum of patients with metastatic liver cancer significantly contribute to the epithelial-mesenchymal transition of cancer cells via the TGF-β1/Smad3 signaling pathway.37 Similarly, Li Wenhua et al demonstrated that exosomes secreted by Huh7 liver cancer cells through autocrine mechanisms upregulate the TGF-β1/Smads pathway, thereby influencing liver cancer metastasis. This elucidates the developmental mechanisms of liver cancer and offers novel therapeutic approaches for hepatocellular carcinoma.38 Given the pivotal role of tumor-derived exosomes in the migration and invasion of receptor cells, exosomes may emerge as novel therapeutic and prognostic targets for patients with liver cancer.
Exosomes Mediate TGF-β1/Smads Signaling Pathway to Promote Liver Cancer
Exosomes modulate the TGF-β1/Smads signaling pathway, facilitating the formation of cancer stem cells and EMT, thereby promoting liver cancer progression. Research indicates that miRNA-126, originating from hepatoblastoma cells, enhances the expression of TGF-β1 and IL-6 in bone marrow MSCs, significantly increasing the proportion of CD44+, CD90+, and CD133+ cells, ultimately inducing the differentiation of bone marrow MSCs into cancer stem cells, thus promoting liver cancer development.39 Exosomes related to endoplasmic reticulum stress, specifically HSP70 in liver cancer cells, markedly upregulate TGF-β1 expression, stimulate the secretion of IL-6/10 by macrophages, and elevate TLR4 expression in macrophages, thereby promoting M2 polarization of macrophages.40 SBF2-AS1, a long non-coding RNA (lncRNA) with oncogenic properties, functions as a competitive endogenous RNA for miRNA-140-5p. TβRI has been identified as a direct target of miRNA-140-5p, and the enforced expression of SBF2-AS1 augments TβRI expression in liver cancer cells, thereby promoting liver cancer progression.41 Exosomes derived from various liver cancer cell sources can activate the TGF-β1/Smad signaling pathway to induce EMT.42
Exosomes Mediate TGF-β1/Smads Signaling Pathway to Inhibit Liver Cancer
Exosomes are instrumental in facilitating the TGF-β1/Smads signaling pathway, thereby exerting anti-hepatocarcinogenic effects and enhancing drug sensitivity in hepatocellular carcinoma cells. Exosomes derived from camel colostrum have been demonstrated to downregulate the expression of inflammation-related genes, such as TGF-β1, TNF-α, and NF-κB, as well as genes associated with angiogenesis in HepaRG liver cancer cells, thus exhibiting both anti-cancer and anti-angiogenic properties.43 In the context of TGF-β1-induced EMT in Hep3B cells, exosomes enriched with hsa-miRNA-374a-5p have been observed to inhibit the proliferation, migration, and invasion of liver cancer cells.44 Additionally, it has been reported that long non-coding RNA can inhibit the TGF-β1 signaling pathway in liver cancer cells, thereby enhancing their sensitivity to therapeutic agents.45 Zhang et al identified that exosomal miRNA-663a functions as a tumor suppressor, playing a crucial role in inhibiting the proliferation, invasion, and tumorigenesis of liver cancer by modulating TGF-β1, both in vitro and in vivo.46 Furthermore, lncRNA SBF2-AS1 was markedly upregulated in liver cancer, whereas miRNA-361-5p was significantly downregulated. miRNA-361-5p acts as a negative regulator of TGF-β1 in liver cancer, and the downregulation of lncRNA SBF2-AS1 modulates miRNA-361-5p to intervene in the TGF-β1 signaling pathway, thereby inhibiting the proliferation and migration of liver cancer cells.47
Exosomal TGF-β1/Smads Signaling Pathway and Related Chronic Liver Diseases
Liver diseases can be broadly divided into two main categories: infectious and noninfectious. Non-infectious liver diseases include metabolic, autoimmune, drug-induced, alcoholic, genetic, and oncological conditions. As the largest metabolic organ in the body, the liver is vulnerable to oxidative stress, damage, and inflammatory reactions due to the accumulation of metabolites or the invasion of toxic substances during metabolic processes. Excessive inflammatory infiltration may lead to hepatitis, which can progressively advance to fibrosis/cirrhosis and potentially liver cancer.48 Therefore, inhibiting oxidative stress and inflammatory responses is a crucial strategy for preventing and treating chronic liver diseases. Research indicates that the overexpression of miRNA-374a significantly reduces TGF-β1 levels, decreases α-SMA and COLI levels, inhibits the activation of HSC, and prevents the onset of related liver diseases.49 Ma et al discovered that hepatocyte exosomes stimulated by thyroid-stimulating hormone modulate hepatic triglyceride accumulation in mice via the TGF-β1/fatty acid triglyceride lipase axis, thereby reducing the likelihood of metabolic liver diseases.50
Exosomes Mediate TGF-β1/Smads Signaling Pathway to Promote Liver Injury
Exosomes have been recognized for their role in regulating protein factor expression by activating the TGF-β1/Smads signaling pathway, which in turn leads to HSC activation and worsens liver injury. In the context of Hepatitis C Virus (HCV) infection, liver cells release exosomes enriched with miRNA-19a, which targets the suppressor of cytokine signaling 3 (SOCS3) in HSC. This action activates the STAT3-mediated TGF-β1 signaling pathway, thereby facilitating HCV progression.51 Kim et al demonstrated that during HCV replication, there is an increased release of exosomes containing miRNA-192, which boosts the expression of TGF-β1, COL I α1, and α-SMA in HSC, thus activating the TGF-β1 signaling pathway and promoting HSC activation.52 Furthermore, hepatocytes that secrete exosomes with TGF-β1 contribute to the expansion of regulatory T cells within the follicle during HCV infection, thereby impairing the antiviral immune response and exacerbating HCV infection.53 Additionally, exosomes derived from adipocytes have been shown to disrupt the TGF-β1 signaling pathway, leading to an imbalance in collagen and ECM deposition, which promotes the progression of non-alcoholic liver disease.54 Collectively, these studies highlight the significant role of exosomes in the pathogenesis of chronic liver diseases via the TGF-β1 pathway, suggesting a promising avenue for future research on liver disease development and treatment.
Exosomes Mediate TGF-β1/Smads to Improve Liver Injury
In a model of liver injury induced by schistosomiasis, exosomes derived from MSC significantly reduced liver granuloma formation and decreased the expression of Smad1/3 and proline, thereby offering a protective effect on the liver. In vitro studies have demonstrated that MSC-derived extracellular vesicles (MSC-EVs) inhibit the activation of HSC triggered by TGF-β1, lower the levels of α-SMA and Smad1/3, and consequently alleviate liver injury.55 In mouse models of liver injury and liver tumors induced by CCl4, exosomes from human umbilical cord mesenchymal stem cells reduced TGF-β1 levels in liver cells, improved liver injury, and inhibited tumor development.56 Zheng et al showed that exosomes derived from dendritic cells enhanced the expression of anti-inflammatory cytokines TGF-β1 and IL-10 by modulating the balance between regulatory T cells (Tregs) and Th17 cells and downregulating IL-17, thereby ameliorating liver ischemia-reperfusion injury.57 In a non-alcoholic steatohepatitis (NASH) rat model, MSC-derived exosomes significantly decreased TGF-β1 levels, reduced the release of inflammatory cytokines TNF-α, IL-1β, and IL-6, diminished liver inflammation, and improved NASH.58
Mediated Regulation of TGF-β1/Smads Signaling Pathway by Exosomes by Traditional Chinese Medicine
Traditional Chinese Medicine Formula
Jianpi Huayu Decoction, a traditional Chinese medicinal formulation, is employed in the treatment of various malignant tumors, including liver cancer. Both in vivo and in vitro studies have demonstrated that Jianpi Huayu Decoction downregulates the expression of miRNA-23a-3p in exosomes, thereby inhibiting the TGF-β1/Smads signaling pathway and obstructing the proliferation, migration, invasion, and EMT of liver cancer.59 Feng Kunliang et al utilized TGF-β1 to stimulate MHCC97H cells, inducing an EMT model of liver cancer cells, and discovered that Jianpi Huayu Formula intervened in the migration, invasion, and EMT of MHCC97H cells via exosomes.60 Jianpi Xiaoai Formula is known for its detoxification properties, ability to dispel wind and dampness, and promotion of diuresis and swelling reduction, and is extensively used in the clinical treatment of advanced cancers. Yang Xiao et al identified that exosomes derived from colon cancer cells could up-regulate the expression of TGF-β1 in hepatic Kupffer cells through macrophage migration inhibitory factor (MIF), while Jianpi Xiaoai Formula containing drug serum could inhibit MIF expression and reduce TGF-β1 expression in hepatic Kupffer cells, thereby preventing liver metastasis of cancer cells.61 Astragalus Decoction, a classic Chinese medicinal formula, has been shown to significantly reduce liver fibrosis. Research indicates that Lnc-C18orf26-1 is upregulated in TGF-β1-activated HSC and competitively binds to miRNA-663a. Astragalus Decoction increases miRNA-663a expression in HSC and reduces lnc-C18orf26-1 expression. Recent studies have shown that Astragalus Decoction suppresses COLIα2, α-SMA, TGF-β1, TβRI, and p-Smad2 expression in HSC. The reversal of these effects by miRNA-663a inhibitors implies that Astragalus Decoction may partially inhibit HSC proliferation and activation by modulating the lnc-c18orf26-1/miRNA-663a/TGF-β1/TβRI/p-Smad2 axis.62 Xue Sanjiashan is acknowledged for its capacity to enhance circulation, eliminate stasis, and clear heat while nourishing the blood. Zhang et al demonstrated that Xue Sanjiashan upregulates miRNA-29b-3p expression and inhibits the expression of VEGFA, PDGA, and TGF-β1, which is advantageous for mitigating pathological angiogenesis in liver tissue.63 Zhan Jie et al identified that Danshao Huaxian Capsule upregulates miRNA-21 and downregulates Smad7 expression, suggesting potential preventive effects on liver fibrosis induced by arsenic exposure in rats.64 The anti-liver-disease effects of traditional Chinese medicine compounds through exosome-mediated modulation of the TGF-β1/Smad signaling pathway are summarized in Table 1.
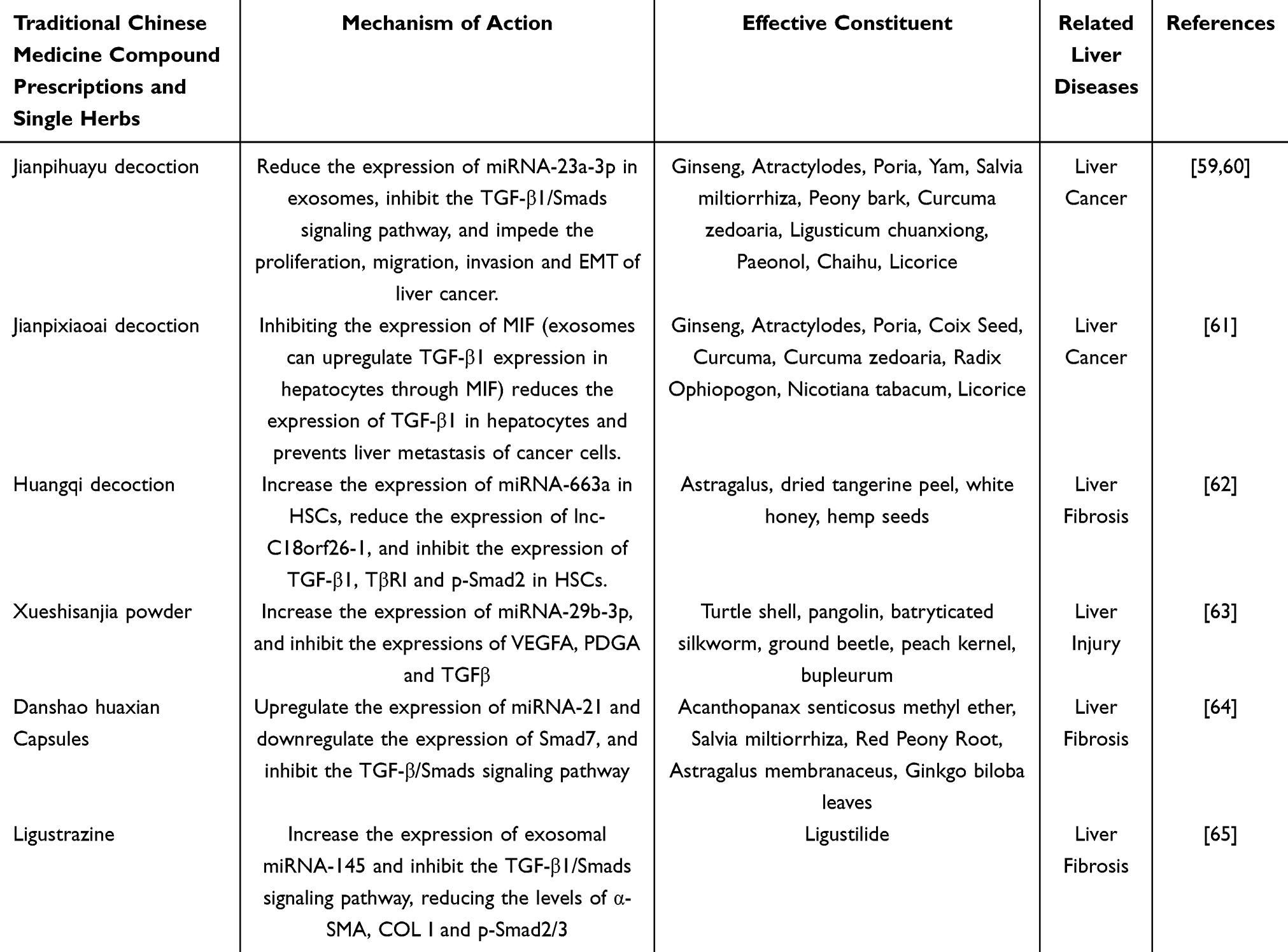 |
Table 1 The Mechanism by Which Traditional Chinese Medicine Compound Formulas and Single Chinese Herbs Exert Anti-Liver Disease Effects Through Exosomes by Inhibiting the TGF-β1/Smads Signaling Pathway |
Effective Components of Traditional Chinese Medicine
Emodin, an anthraquinone derivative, exhibits significant anticancer properties in various cancer types. In co-culture experiments involving M2 macrophages and liver cancer cell lines (HepG2 and Huh7), M2 macrophages facilitated the proliferation and migration of these cells. However, emodin counteracted this effect by downregulating exosomal miRNA-26a, thereby inhibiting the expression of TGF-β1 and reversing these outcomes.66 Rhodiola rosea is characterized by its ability to promote blood circulation, arrest bleeding, clear heat, eliminate dampness, detoxify, and reduce swelling, making it a common therapeutic agent in the clinical management of liver diseases. Exosomes loaded with rhodiola acid were found to decrease the mRNA levels of TGF-β1 and Smad4, while upregulating the epithelial marker E-cadherin and downregulating the mesenchymal marker vimentin, thereby inhibiting the EMT in liver cancer.67 Curcumin, a yellow pigment derived from turmeric rhizomes, is known for its lipid-lowering and antitumor effects. It modulates the miRNA-21/TIMP3 signaling pathway to deactivate the TGF-β1/smad3 signaling pathway, thereby inhibiting the proliferation and apoptosis of HepG2 and HCCLM3 cells and exerting anti-liver cancer effects.68 Chlorogenic acid, also known as coffee tannic acid, possesses antioxidant, tumor-preventive, antibacterial, and antiviral properties. Wang et al reported that chlorogenic acid reduces protein levels of connective tissue growth factor, p-Smad1/2/3, TβRI, and α-SMA through the interaction of exosomal miRNA-21/IL-13/Smad7 signaling, while increasing Smad7 levels, thus inhibiting liver fibrosis induced by LX-2 cells and schistosomiasis-infected mice.69 Similarly, studies have demonstrated that chlorogenic acid, by blocking the regulation of the TGF-β1/Smad7 signaling pathway by exosomal miRNA-21, reduces serum levels of α-SMA, COL I, and TGF-β1, thereby ameliorating liver fibrosis.70 Methyl salicic acid, an active component of Salix alba, exhibits protective effects against liver injury in mice. In vitro experiments revealed that methyl salicic acid inhibits HSC activation and reduces collagen accumulation. Further in vivo studies confirmed that methyl salicylate modulates the ERK and TGF-β1/Smads signaling pathways mediated by exosomal miRNA-21, thereby reducing collagen production and inflammatory factor release and inhibiting the progression of liver fibrosis.71 Tannic acacia extract has been shown to inhibit the TGF-β1/Smads signaling pathway regulated by exosomal miRNA-21, reducing the levels of α-SMA, COL I, and p-Smad2/3, and exerting hepatoprotective effects.72 Oxymatrine is frequently used in the clinical treatment of chronic liver diseases. It reduces the expression of exosomal miRNA-195 and α-SMA induced by TGF-β1 in HSCs, upregulates Smad7 expression, and prevents liver fibrosis.73 Dalbergine, an active component found in plants such as sandalwood, blueberry, grape, and guava palm, possesses anticancer, anti-inflammatory, and antioxidant properties. In vitro experiments demonstrated that dalbergine reduced the expression of fibroblast-specific protein 1 and vimentin during EMT while increasing E-cadherin levels. In vivo studies have indicated that dalbergine downregulates liver extracellular exosomal miRNA-34a, inhibits TGF-β1/Smad signaling, and prevents EMT in liver cells during liver fibrosis.74 Cinnamaldehyde, a compound derived from cinnamon, exhibits extensive pharmacological activity. It targets the downregulation of its expression by exosomal miRNA-17-5p, upregulates Smad7 expression, thereby interfering with the TGF-β1/Smads pathway, inhibiting ECM deposition, and protecting against thioacetamide-induced liver fibrosis in mice.75 Schisandrin B exhibits various biological activities, including hepatoprotective, anti-inflammatory, and anticancer effects. It upregulates the expression of exosomal miRNA-101-5p, thereby obstructing the TGF-β1 signaling pathway and inhibiting liver fibrosis in rats.76 Polygonum cuspidatum saponin, an active component of Polygonum cuspidatum rhizomes, exerts protective effects in patients with liver fibrosis and in experimental animal models. It promotes the expression of exosomal miR-203 by inhibiting the nuclear translocation of zinc finger E-box binding homeobox 1 (ZEB1) and intervenes in the TGF-β1/Smads pathway of survivin protein activation, thereby ameliorating fructose-induced liver fibrosis in rats.77 Glycyrrhetinic acid modulates the miRNA-663a/TGF-β1/Smads signaling pathway, downregulates the expressions of TGF-β1, TβRI, α-SMA, Smad4, and p-Smad2/3 in HSC, and prevents HSC activation.78 The anti-liver-disease effects of the active ingredients of traditional Chinese medicine are mediated by exosomes through the TGF-β1/Smads signaling pathway, as illustrated in Table 2.
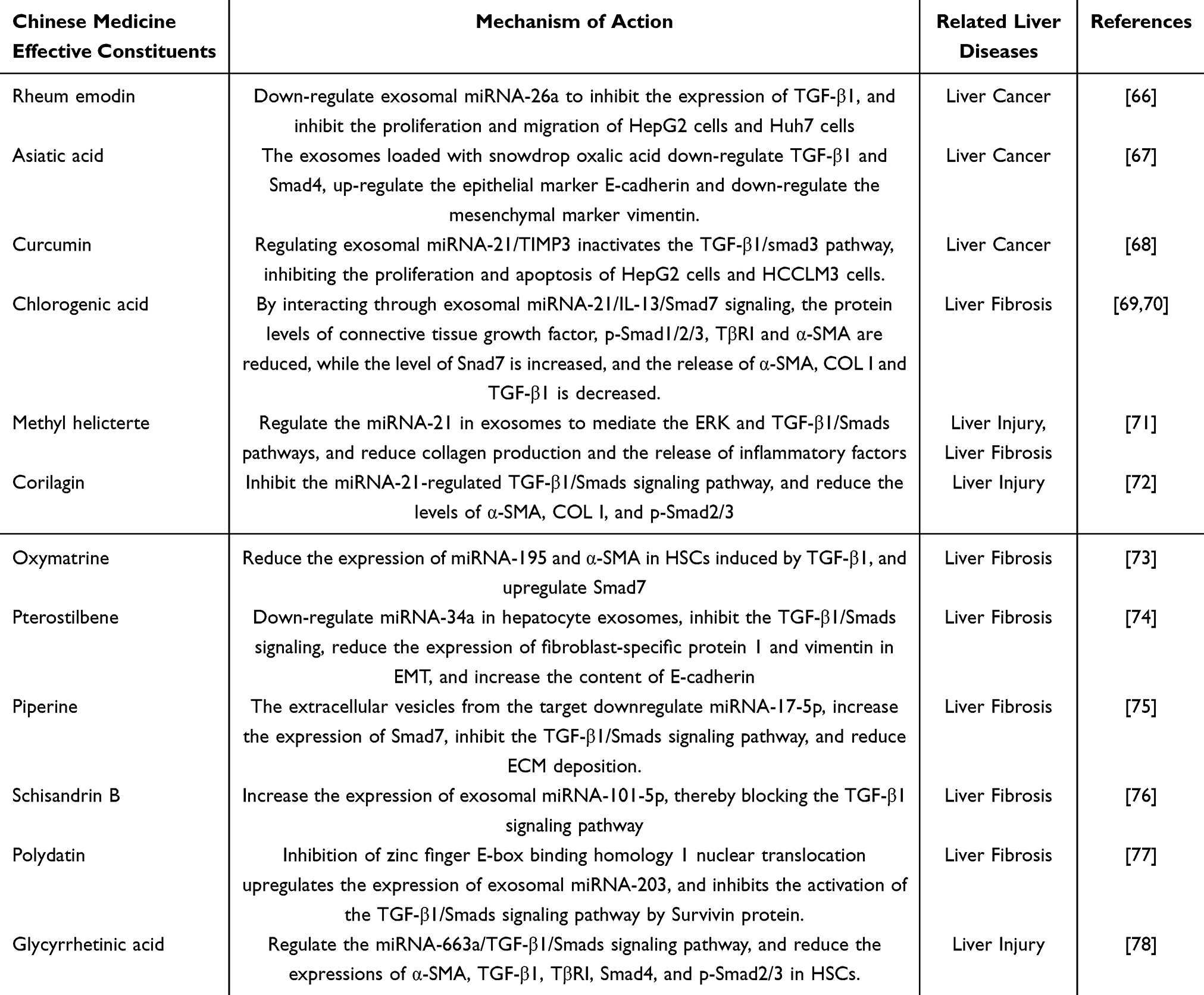 |
Table 2 The Mechanism by Which the Active Components of Traditional Chinese Medicine Exert Anti-Liver Disease Effects Through Inhibiting the TGF-β1/Smads Signaling Pathway via Exosomes |
Single Chinese Herb
Ligustrazine, a compound extracted from the rhizome of Ligusticum chuanxiong, is primarily used to treat ischemic diseases. In a study conducted by Qiu et al, ligustrazine was administered to a rat model of liver fibrosis induced by ethanol-induced biliary atresia and to an HSC model activated by TGF-β1. The findings indicated that ligustrazine enhances the expression of exosomal miRNA-145 and inhibits the TGF-β1/Smad signaling pathway. This results in a reduction in α-SMA, COL I, and p-Smad2/3 protein levels, thereby reversing liver fibrosis.65 The effects of individual Chinese herbs on liver disease, mediated through exosomes via the TGF-β1/Smad signaling pathway, are summarized in Table 1.
Summary and Outlook
Liver diseases continue to pose a significant threat to human health owing to their complex and diverse pathogenesis, presenting a formidable challenge to the medical community. Among these, the TGF-β1/Smad signaling pathway is intricately linked to the activation of HSC and liver diseases, making it a potential target for therapeutic intervention. Exosomes from various sources contribute to the onset and progression of liver diseases by modulating the TGF-β1/Smad signaling pathway. These exosomes can target and regulate the antagonist Smad7 protein, thereby influencing the TGF-β1/Smads signaling pathway, or they can target and bind to the upstream receptors TβRI/II to mediate this pathway. Additionally, exosomes can modulate the expression of downstream Smad2/3/4 proteins to disrupt TGF-β1 signal transduction or interfere with the TGF-β1/Smad signaling pathway through other regulatory factors. The pathogenesis of liver diseases involves multiple molecules and signaling pathways, with the TGF-β1/Smads signaling pathway being a crucial pro-fibrotic pathway regulated by one or more exosomes. Notably, most exosomes play a dual role in either promoting or inhibiting liver diseases. This review aims to synthesize the existing evidence to elucidate the regulatory mechanisms of exosome-mediated TGF-β1/Smad signaling pathways in liver diseases. In the contexts of liver fibrosis, hepatocellular carcinoma, and liver injury, exosomes mediate the TGF-β1/Smads pathway by regulating the expression of related factors such as Smad2/3/4 and TβRI/II, thereby inhibiting the release of α-SMA and COL I/III and mitigating inflammatory responses.
Current investigations into the role of exosomes in liver cancer, particularly in the context of the TGF-β1/Smads signaling pathway, are limited to a few studies. Exosomes are intricately associated with liver cancer progression. On the one hand, although exosomes do not directly regulate liver cancer promotion via the TGF-β1/Smads signaling pathway, they can induce modifications in cancer-related factors by modulating this pathway, thereby facilitating the development of liver cancer. On the other hand, Exosomes inhibit liver cancer progression by directly suppressing the expression of liver cancer-associated factors or enhancing drug sensitivity. Furthermore, in the context of liver injury, exosomes derived from damaged liver cells are known to alter factors associated with the TGF-β1/Smad signaling pathway, potentially stimulating HSC activation or exacerbating existing liver conditions. Conversely, exosomes derived from MSCs have been shown to ameliorate liver injury by inhibiting the expression of inflammatory cells and factors.
Investigating the specific mechanisms of exosomes within the TGF-β1/Smads signaling pathway presents significant challenges due to their dual nature. Exosomes derived from the same cell type can have opposing effects. For example, NEAT1 exosomes from macrophages activate HSC by elevating TGF-β1 and p-Smad2/3 levels, thereby promoting liver fibrosis.19 Conversely, exosomes from macrophages secrete anti-inflammatory factors IL-10 and TGF-β, which inhibit liver injury.79 Similarly, miRNA-142 and miRNA-223 exosomes from macrophages inhibit the proliferation of hepatocellular carcinoma when transferred to hepatoma cells.80 Furthermore, the same exosomes can exhibit contrasting effects when derived from different sources. In NAFLD mice, miRNA-21 expression is reduced, and following miRNA-21 knockout, there is an increase in lipid levels, fibrosis, and TGF-β1 levels.81 In TGF-β1-induced HSC, miRNA-21-5p expression is upregulated, enhancing the expression of the pro-fibrotic genes α-SMA and COLIα1 by inhibiting Smad7 expression.82 Additionally, variations in exosome isolation methods and purity are also evident. Exosomes derived in vivo and those prepared in vitro differ fundamentally in their origin, composition, function, and application, leading to variations in the results. In vivo, it is challenging to ascertain whether a single exosome acts independently or interacts with multiple exosomes. Consequently, comprehensive research on the molecular mechanisms of exosomes, purification methods, and in vivo effects is urgently needed.
TCM offers notable advantages in the treatment of chronic liver diseases, primarily because of its active ingredients, which often exhibit hepatoprotective and hepatolytic properties. TCM emphasizes a holistic approach and syndrome differentiation in disease treatment, exerting therapeutic effects across multiple levels and targets. This comprehensive perspective helps overcome the limitations of single-efficacy treatments. Currently, scholarly attention is increasingly focused on the impact of TCM on exosomes and their regulation of the TGF-β1/Smad signaling pathway in ameliorating liver diseases. Although research on the regulation of the TGF-β1/Smad signaling pathway by TCM through exosomes remains limited, it presents a promising avenue for further investigation, potentially offering novel insights into the treatment of liver disease with TCM. Future advancements in bioinformatics, high-throughput sequencing, and metabolomics are expected to facilitate the screening and prediction of disease-drug-target associations related to exosomes. This will enhance our understanding of the molecular mechanisms underlying the effects of exosomes, TCM active ingredients, and their anti-liver disease properties. By drawing on network pharmacology methodologies, a comprehensive and systematic exosome/TGF-β1/Smads signaling pathway molecular network can be established. This will enable the effective application of research findings and the exploration of new research questions, thereby broadening research perspectives and contributing significantly to the treatment and study of liver disease. In conclusion, strengthening research on the mechanisms by which TCM regulates the TGF-β1/Smad signaling pathway through exosomes provides a foundation for the clinical application of TCM in the treatment of liver disease.
Data Sharing Statement
The datasets generated during and/or analyzed during the current study are available from both authors on reasonable request.
Acknowledgments
The author expresses his gratitude to Dr. Deng Jiasheng for his valuable suggestions on the entire paper.
Author Contributions
All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Dr. Guidong Huang: conceptualization, formal analysis, investigation, writing original draft.
Dr. Zhipin Zhou: writing review & editing, funding acquisition, resources, supervision.
Funding
The Guangxi Natural Science Foundation Project (2021GXNSFAA075020) funded the entire paper.
Disclosure
The authors have no conflicts of interest to disclose.
References
1. Meirelles LDS, Marson RF, Solari MIG, et al. Are liver pericytes just precursors of myofibroblasts in hepatic diseases? insights from the crosstalk between perivascular and inflammatory cells in liver injury and repair. Cells. 2020;9(1):188. doi:10.3390/cells9010188
2. Asrani SK, Devarbhavi H, Eaton J, et al. Burden of liver diseases in the world. J Hepatol. 2019;70(1):151–14. doi:10.1016/j.jhep.2018.09.014
3. Kostallari E, Valainathan S, Biquard L, et al. Role of extracellular vesicles in liver diseases and their therapeutic potential. Adv Drug Deliv Rev. 2021;175:113816. doi:10.1016/j.addr.2021.05.026
4. Wu P, Zhang B, Ocansey DKW, et al. Extracellular vesicles: a bright star of nanomedicine. Biomaterials. 2021;269:120467. doi:10.1016/j.biomaterials.2020.120467
5. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
6. Keerthikumar S, Chisanga D, Ariyaratne D, et al. ExoCarta: a web-based compendium of exosomal cargo. J Mol Biol. 2016;428(4):688–692. doi:10.1016/j.jmb.2015.09.019
7. Skotland T, Hessvik NP, Sandvig K, et al. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J Lipid Res. 2019;60(1):9–18. doi:10.1194/jlr.R084343
8. Wortzel I, Dror S, Kenific CM, et al. Exosome-mediated metastasis: communication from a distance. Dev Cell. 2019;49(3):347–360. doi:10.1016/j.devcel.2019.04.011
9. Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor[J]. Cell. 1983;33(3):967–978. doi:10.1016/0092-8674(83)90040-5
10. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi:10.1084/jem.183.3.1161
11. Robbins PD, Dorronsoro A, Booker CN. Regulation of chronic inflammatory and immune processes by extracellular vesicles. J Clin Invest. 2016;126(4):1173–1180. doi:10.1172/JCI81131
12. Liu S, Ren J, Ten Dijke P. Targeting TGFβ signal transduction for cancer therapy. Signal Transduct Target Ther. 2021;6(1):8. doi:10.1038/s41392-020-00436-9
13. Fabregat I, Moreno-Càceres J, Sánchez A, et al. TGF-β signalling and liver disease. FEBS J. 2016;283(12):2219–2232. doi:10.1111/febs.13665
14. Dewidar B, Meyer C, Dooley S, et al. TGF-β in hepatic stellate cell activation and liver fibrogenesis-updated 2019. Cells. 2019;8(11):1419. doi:10.3390/cells8111419
15. Lang Q, Liu Q, Xu N, et al. The antifibrotic effects of TGF-β1 siRNA on hepatic fibrosis in rats. Biochem Biophys Res Commun. 2011;409(3):448–453. doi:10.1016/j.bbrc.2011.05.023
16. Friedman SL. Mechanisms of hepatic fibrogenesis. Gastroenterology. 2008;134(6):1655–1669. doi:10.1053/j.gastro.2008.03.003
17. Trivedi P, Wang S, Friedman SL. The power of plasticity-metabolic regulation of hepatic stellate cells. Cell Metab. 2021;33(2):242–257. doi:10.1016/j.cmet.2020.10.026
18. Liu Y, Zheng Y, Yang Y, et al. Exosomes in liver fibrosis: the role of modulating hepatic stellate cells and immune cells, and prospects for clinical applications. Front Immunol. 2023;14:1133297. doi:10.3389/fimmu.2023.1133297
19. Shu B, Zhang RZ, Zhou YX, et al. METTL3-mediated macrophage exosomal NEAT1 contributes to hepatic fibrosis progression through Sp1/TGF-β1/Smad signaling pathway. Cell Death Discov. 2022;8(1):266. doi:10.1038/s41420-022-01036-y
20. Wang Y, Gong W, Zhou H, et al. A novel miRNA from egg-derived exosomes of schistosoma japonicum promotes liver fibrosis in murine schistosomiasis. Front Immunol. 2022;13:860807. doi:10.3389/fimmu.2022.860807
21. Zhou QY, Yang HM, Liu JX, et al. MicroRNA-497 induced by Clonorchis sinensis enhances the TGF-β/Smad signaling pathway to promote hepatic fibrosis by targeting Smad7. Parasit Vectors. 2021;14(1):472. doi:10.1186/s13071-021-04972-3
22. WHe X, Wang Y, Fan X, et al. A schistosome miRNA promotes host hepatic fibrosis by targeting transforming growth factor beta receptor III. J Hepatol. 2020;72(3):519–527. doi:10.1016/j.jhep.2019.10.029
23. Wang Q, Wei S, Li L, et al. miR-139-5p sponged by LncRNA NEAT1 regulates liver fibrosis via targeting β-catenin/SOX9/TGF-β1 pathway. Cell Death Discov. 2021;7(1):243. doi:10.1038/s41420-021-00632-8
24. Walton KL, Johnson KE, Harrison CA. Targeting TGF-β mediated SMAD signaling for the prevention of fibrosis. Front Pharmacol. 2017;8:461. doi:10.3389/fphar.2017.00461
25. Wang N, Li X, Zhong Z, et al. 3D hESC exosomes enriched with miR-6766-3p ameliorates liver fibrosis by attenuating activated stellate cells through targeting the TGFβRII-SMADS pathway. J Nanobiotechnol. 2021;19(1):437. doi:10.1186/s12951-021-01138-2
26. Mengjie C, Jiali Q, Yifei C, et al. HucMSC-ex regulates TGF-βR_1 through Let-7b to inhibit collagen synthesis in hepatic stellate cells. J Jiangsu Univ. 2018;28(06):478–482.
27. Mardpour S, Hassani SN, Mardpour S, et al. Extracellular vesicles derived from human embryonic stem cell-MSCs ameliorate cirrhosis in thioacetamide-induced chronic liver injury. J Cell Physiol. 2018;233(12):9330–9344. doi:10.1002/jcp.26413
28. Jang YJ, An SY, Kim JH. Identification of MFGE8 in mesenchymal stem cell secretome as an anti-fibrotic factor in liver fibrosis. BMB Rep. 2017;50(2):58–59. doi:10.5483/BMBRep.2017.50.2.012
29. An SY, Jang YJ, Lim HJ, et al. Milk fat globule-egf factor 8, secreted by mesenchymal stem cells, protects against liver fibrosis in mice. Gastroenterology. 2017;152(5):1174–1186. doi:10.1053/j.gastro.2016.12.003
30. Li T, Yan Y, Wang B, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013;22(6):845–854. doi:10.1089/scd.2012.0395
31. Qu Y, Zhang Q, Cai X, et al. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J Cell Mol Med. 2017;21(10):2491–2502. doi:10.1111/jcmm.13170
32. Sun C, Shi C, Duan X, et al. Exosomal microRNA-618 derived from mesenchymal stem cells attenuate the progression of hepatic fibrosis by targeting Smad4. Bioengineered. 2022;13(3):5915–5927.
33. Wang L, Wang Y, Quan J. Exosomes derived from natural killer cells inhibit hepatic stellate cell activation and liver fibrosis. Hum Cell. 2020;33(3):582–589. doi:10.1007/s13577-020-00371-5
34. Wang L, Wang Y, Quan J. Exosomal miR-223 derived from natural killer cells inhibits hepatic stellate cell activation by suppressing autophagy. Mol Med. 2020;26(1):81. doi:10.1186/s10020-020-00207-w
35. Wang L, Liao Y, Yang R, et al. Sja-miR-71a in Schistosome egg-derived extracellular vesicles suppresses liver fibrosis caused by schistosomiasis via targeting semaphorin 4D. J Extracell Vesicles. 2020;9(1):1785738. doi:10.1080/20013078.2020.1785738
36. Konyn P, Ahmed A, Kim D. Current epidemiology in hepatocellular carcinoma. Expert Rev Gastroenterol Hepatol. 2021;15(11):1295–1307. doi:10.1080/17474124.2021.1991792
37. Jinghuan D, Guodong L, Jing Z, et al. The influence of serum exosomes-mediated TGF-β pathway on epithelial-mesenchymal transition of HepG2 cells in advanced metastatic liver cancer. Chin J Oncol Prevention Treatment. 2023;30(02):84–92.
38. Wenhua L, Qianwen W, Xiaofang W, et al. Autocrine exosomes from liver cancer cells regulate their own cellular behaviors through TGF-β1. Chin J Immunol. 2023;39(07):1442–1445.
39. Hu Y, Zai H, Jiang W, et al. miR-126 in extracellular vesicles derived from hepatoblastoma cells promotes the tumorigenesis of hepatoblastoma through inducing the differentiation of BMSCs into cancer stem cells. J Immunol Res. 2021;2021:6744715. doi:10.1155/2021/6744715
40. Wei H, Manman Z, Luolu F, et al. Endoplasmic reticulum stress-induced hepatoma cells promote macrophage M2 polarization through exosome-Toll-like receptor 4 signaling pathway. Chin J Pharmacol. 2021;37(07):1008–1015.
41. Li Y, Liu G, Li X, et al. Long non-coding RNA SBF2-AS1 promotes hepatocellular carcinoma progression through regulation of miR-140-5p-TGFBR1 pathway. Biochem Biophys Res Commun. 2018;503(4):2826–2832. doi:10.1016/j.bbrc.2018.08.047
42. Qu Z, Feng J, Pan H, et al. Exosomes derived from HCC cells with different invasion characteristics mediated EMT through TGF-β/Smad signaling pathway. Onco Targets Ther. 2019;12:6897–6905. doi:10.2147/OTT.S209413
43. El-Kattawy AM, Algezawy O, Alfaifi MY, et al. Therapeutic potential of camel milk exosomes against HepaRG cells with potent apoptotic, anti-inflammatory, and anti-angiogenesis effects for colostrum exosomes. Biomed Pharmacother. 2021;143:112220. doi:10.1016/j.biopha.2021.112220
44. Lin Q, Zhou CR, Bai MJ, et al. Exosome-mediated miRNA delivery promotes liver cancer EMT and metastasis. Am J Transl Res. 2020;12(3):1080–1095.
45. Takahashi K, Yan IK, Kogure T, et al. Extracellular vesicle-mediated transfer of long non-coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio. 2014;4:458–467. doi:10.1016/j.fob.2014.04.007
46. Zhang C, Chen B, Jiao A, et al. miR-663a inhibits tumor growth and invasion by regulating TGF-β1 in hepatocellular carcinoma. BMC Cancer. 2018;18(1):1179.
47. Wu YH, Yu B, Chen WX, et al. Downregulation of lncRNA SBF2-AS1 inhibits hepatocellular carcinoma proliferation and migration by regulating the miR-361-5p/TGF-β1 signaling pathway. Aging. 2021;13(15):19260–19271. doi:10.18632/aging.203248
48. Filliol A, Schwabe RF. FoxM1 Induces CCl2 secretion from hepatocytes triggering hepatic inflammation, injury, fibrosis, and liver cancer. Cell Mol Gastroenterol Hepatol. 2020;9(3):555–556. doi:10.1016/j.jcmgh.2020.01.002
49. Yuan Q, Zhang Z, Hu X, et al. miR-374a/Myc axis modulates iron overload-induced production of ROS and the activation of hepatic stellate cells via TGF-β1 and IL-6. Biochem Biophys Res Commun. 2019;515(3):499–504. doi:10.1016/j.bbrc.2019.05.152
50. Ma S, Wang Y, Fan S, et al. TSH-stimulated hepatocyte exosomes modulate liver-adipose triglyceride accumulation via the TGF-β1/ATGL axis in mice. Lipids Health Dis. 2025;24(1):81. doi:10.1186/s12944-025-02509-6
51. Devhare PB, Sasaki R, Shrivastava S, et al. Exosome-mediated intercellular communication between hepatitis c virus-infected hepatocytes and hepatic stellate cells. J Virol. 2017;91(6):e02225–16.
52. Kim JH, Lee CH, Lee SW. Exosomal transmission of microrna from hcv replicating cells stimulates transdifferentiation in hepatic stellate cells. Mol Ther Nucleic Acids. 2019;14:483–497.
53. Cobb DA, Kim OK, Golden-Mason L, et al. Hepatocyte-derived exosomes promote T follicular regulatory cell expansion during hepatitis C virus infection. Hepatology. 2018;67(1):71–85. doi:10.1002/hep.29409
54. Koeck ES, Iordanskaia T, Sevilla S, et al. Adipocyte exosomes induce transforming growth factor beta pathway dysregulation in hepatocytes: a novel paradigm for obesity-related liver disease. J Surg Res. 2014;192(2):268–275. doi:10.1016/j.jss.2014.06.050
55. Dong L, Pu Y, Chen X, et al. hUCMSC-extracellular vesicles downregulated hepatic stellate cell activation and reduced liver injury in S. japonicum-infected mice. Stem Cell Res Ther. 2020;11(1):21. doi:10.1186/s13287-019-1539-8
56. Jiang W, Tan Y, Cai M, et al. Human umbilical cord MSC-derived exosomes suppress the development of CCl4-induced liver injury through antioxidant effect. Stem Cells Int. 2018;2018:6079642.
57. Zheng L, Li Z, Ling W, et al. Exosomes derived from dendritic cells attenuate liver injury by modulating the balance of treg and th17 cells after ischemia reperfusion. Cell Physiol Biochem. 2018;46(2):740–756. doi:10.1159/000488733
58. Ohara M, Ohnishi S, Hosono H, et al. Extracellular vesicles from amnion-derived mesenchymal stem cells ameliorate hepatic inflammation and fibrosis in rats. Stem Cells Int. 2018;2018:3212643.
59. Xie CF, Feng KL, Wang JN, et al. Jianpi Huayu decoction inhibits the epithelial-mesenchymal transition of hepatocellular carcinoma cells by suppressing exosomal miR-23a-3p/Smad signaling. J Ethnopharmacol. 2022;294:115360.
60. Kunliang F, Qinglian C, Chunfeng X, et al. The healthy spleen and detoxifying formula affects the migration, invasion and epithelial-mesenchymal transition of hepatoma cells through exosomes in vitro. New Drugs Trad Chin Med Clin Pharmacol. 2021;32(12):1745–1751.
61. Xiao Y, Luo J, Yongmin L, et al. Study on the intervention of jianpi xiaoai formula on the regulation of hepatic kupffer cell tgf-β1 expression by exosomal mif in colon cancer cells. J Beijing Univ Chin Med. 2022;45(02):184–192.
62. Dong BS, Liu FQ, Yang WN, et al. Huangqi Decoction, a compound Chinese herbal medicine, inhibits the proliferation and activation of hepatic stellate cells by regulating the long noncoding RNA-C18orf26-1/microRNA-663a/transforming growth factor-β axis. J Integr Med. 2023;21(1):47–61. doi:10.1016/j.joim.2022.11.002
63. Zhang T, Yang Y, Wang B, et al. XSSJS inhibits hepatic fibrosis by promoting the miR-29b-3p/VEGFA axis in vitro and in vivo. Biosci Rep. 2022;42(2):BSR20212241.
64. Jie Z, Jun W, Xueke Z. Expression of miRNA-21 and Smad7 in liver tissues of rats with liver fibrosis caused by arsenic exposure and drug intervention. World Chinese Digest. 2014;22(34):5249–5257.
65. Qiu JL, Zhang GF, Chai YN, et al. Ligustrazine attenuates liver fibrosis by targeting miR-145 mediated transforming growth factor-β/smad signaling in an animal model of biliary atresia. J Pharmacol Exp Ther. 2022;381(3):257–265. doi:10.1124/jpet.121.001020
66. Yin J, Zhao X, Chen X, et al. Emodin suppresses hepatocellular carcinoma growth by regulating macrophage polarization via microRNA-26a/transforming growth factor beta 1/protein kinase B. Bioengineered. 2022;13(4):9548–9563. doi:10.1080/21655979.2022.2061295
67. Chen T, Zhu C, Wang X, et al. asiatic acid encapsulated exosomes of hepatocellular carcinoma inhibit epithelial-mesenchymal transition through transforming growth factor beta/smad signaling pathway. J Biomed Nanotechnol. 2021;17(12):2338–2350. doi:10.1166/jbn.2021.3208
68. Li J, Wei H, Liu Y, et al. Curcumin inhibits hepatocellular carcinoma via regulating miR-21/TIMP3 Axis. Evid Based Complement Alternat Med. 2020;2020:2892917. doi:10.1155/2020/2892917
69. Wang Y, Yang F, Xue J, et al. Antischistosomiasis liver fibrosis effects of chlorogenic acid through IL-13/miR-21/Smad7 signaling interactions in vivo and in vitro. Antimicrob Agents Chemother. 2017;61(2):e01347–16. doi:10.1128/AAC.01347-16
70. Yang F, Luo L, Zhu ZD, et al. Chlorogenic Acid Inhibits Liver Fibrosis by Blocking the miR-21-Regulated TGF-β1/Smad7 Signaling Pathway in Vitro and in Vivo. Front Pharmacol. 2017;8:929. doi:10.3389/fphar.2017.00929
71. Huang Q, Zhang X, Bai F, et al. Methyl helicterte ameliorates liver fibrosis by regulating miR-21-mediated ERK and TGF-β1/Smads pathways. Int Immunopharmacol. 2019;66:41–51. doi:10.1016/j.intimp.2018.11.006
72. Zhou X, Xiong J, Lu S, et al. Inhibitory effect of corilagin on mir-21-regulated hepatic fibrosis signaling pathway. Am J Chin Med. 2019;47(7):1541–1569. doi:10.1142/S0192415X19500794
73. Song LY, Ma YT, Fang WJ, et al. Inhibitory effects of oxymatrine on hepatic stellate cells activation through TGF-β/miR-195/Smad signaling pathway. BMC Complement Altern Med. 2019;19(1):138. doi:10.1186/s12906-019-2560-2
74. Song L, Chen TY, Zhao XJ, et al. Pterostilbene prevents hepatocyte epithelial-mesenchymal transition in fructose-induced liver fibrosis through suppressing miR-34a/Sirt1/p53 and TGF-β1/Smads signalling. Br J Pharmacol. 2019;176(11):1619–1634. doi:10.1111/bph.14573
75. Abdelhamid AM, Selim A, Zaafan MA. The hepatoprotective effect of piperine against thioacetamide-induced liver fibrosis in mice: the involvement of mir-17 and tgf-β/Smads pathways. Front Mol Biosci. 2021;8:754098. doi:10.3389/fmolb.2021.754098
76. Cuiqiong W, Chao X, Xinling F, et al. Schisandrin B suppresses liver fibrosis in rats by targeting miR-101-5p through the TGF-β signaling pathway[J]. Artif Cells Nanomed Biotechnol. 2020;48(1):473–478. doi:10.1080/21691401.2020.1717507
77. Zhao X, Yang Y, Yu H, et al. Polydatin inhibits ZEB1-invoked epithelial-mesenchymal transition in fructose-induced liver fibrosis. J Cell Mol Med. 2020;24(22):13208–13222. doi:10.1111/jcmm.15933
78. Guo Y XX, Dong WN, Dong B-S, et al. Glycyrrhetinic Acid -induced mir-663a alleviates hepatic stellate cell activation by attenuating the TGF- β /smad signaling pathway. Evid Based Complement Alternat Med. 2020;2020:3156267. doi:10.1155/2020/3156267
79. Shapouri-Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
80. Aucher A, Rudnicka D, Davis DM. MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation. J Immunol. 2013;191(12):6250–6260. doi:10.4049/jimmunol.1301728
81. Jagtap U, Quan A, Ono Y, et al. miR-21: a therapeutic target for delaying severe liver disease and hepatocellular carcinoma in high-fat-diet-fed mice. bioRxiv. 2024;613915.
82. Li W, Yu X, Chen X, et al. HBV induces liver fibrosis via the TGF-β1/miR-21-5p pathway. Exp Ther Med. 2021;21;2:169.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Post-Translational Modifications of Histones and Non-Histones in Liver Disease and Traditional Chinese Medicine Treatment: A Narrative Review
Xie Z, Deng Y, Zhang X, Chen J, Deng J, Fang Y, Ye X, Zhou Z
Pharmacogenomics and Personalized Medicine 2026, 19:555701
Published Date: 3 February 2026