Back to Journals » Journal of Multidisciplinary Healthcare » Volume 18
Exoskeleton Robot Gait Training and Its Impact on the Gut Microbiota-Brain Axis in Incomplete Spinal Cord Injury Patients: A Narrative Review of Rehabilitation Mechanisms
Received 30 May 2025
Accepted for publication 18 September 2025
Published 6 October 2025 Volume 2025:18 Pages 6411—6430
DOI https://doi.org/10.2147/JMDH.S543841
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Zhixia Zhang,1 Wei Huang2
1Health and Elderly Care Department, Shandong Institute of Commerce and Technology, Jinan City, Shandong Province, People’s Republic of China; 2Medical Department, Zaozhuang Vocational College, Zaozhuang City, Shandong Province, People’s Republic of China
Correspondence: Zhixia Zhang, Health and Elderly Care Department, Shandong Institute of Commerce and Technology, No. 4516, Tourism Road, Jinan City, Shandong Province, 250103, People’s Republic of China, Email [email protected]
Abstract: Exoskeleton robot-assisted gait training represents a significant advancement in neurorehabilitation for patients with incomplete spinal cord injury (iSCI). While its efficacy in improving motor function is increasingly documented, emerging evidence suggests these interventions may exert therapeutic effects through previously unrecognized physiological pathways involving the gut microbiota-brain axis. This review synthesizes current evidence regarding the bidirectional relationship between exoskeleton-based locomotor training and alterations in gut microbiome composition and function in the context of iSCI. Following spinal cord injury, significant dysbiosis occurs, characterized by reduced microbial diversity and altered taxonomic representation, which correlates with neuroinflammation, autonomic dysfunction, and impaired recovery. Exoskeleton-mediated gait rehabilitation appears to partially restore microbial homeostasis through multiple mechanisms, including autonomic nervous system regulation, altered intestinal transit time, modified intestinal barrier integrity, and immunomodulation. These microbiome modifications potentially facilitate neuroplasticity and functional recovery through microbiota-derived metabolites that traverse the blood-brain barrier or communicate via vagal afferents. The integration of metagenomic analysis with functional neuroimaging and detailed autonomic assessment in prospective studies represents a critical research direction. This emerging perspective extends beyond biomechanical rehabilitation, suggesting a comprehensive neurobiological effect that includes modulation of the microbiota-gut-brain axis, with significant implications for optimizing therapeutic strategies for individuals with incomplete spinal cord injury.
Keywords: exoskeleton robot, incomplete spinal cord injury, gut microbiota-brain axis, neuroplasticity, rehabilitation
Introduction
Incomplete spinal cord injury (iSCI), characterized by partial preservation of sensory and/or motor function below the neurological level of injury, affects approximately 65–75% of the spinal cord injury population and presents unique rehabilitative opportunities.1 The preservation of neural pathways provides a substrate for functional recovery that can be optimized through targeted neurorehabilitative interventions. Robotic exoskeleton technology represents a paradigm shift in rehabilitation strategies for patients with iSCI, enabling systematic, reproducible, and prolonged gait training that exceeds the capabilities of conventional therapy in repetition volume and consistency.2
While the mechanical advantages of exoskeleton-assisted gait training are well-established, a nascent body of evidence suggests these interventions may exert therapeutic effects through previously unrecognized physiological pathways. Of particular interest is the potential influence on the gut microbiota-brain axis, a bidirectional communication network linking intestinal microbial communities with central nervous system function.3 Emerging research indicates that spinal cord injury induces substantial alterations in gut microbiome composition and metabolic activity, characterized by decreased microbial diversity, altered taxonomic representation, and modified functional capacity.4,5 These dysbiotic patterns have been associated with systemic inflammation, immune dysregulation, and impaired neurological recovery.6
The convergence of exoskeleton rehabilitation, gut microbiome science, and neurological recovery following SCI presents a compelling opportunity to explore novel mechanistic frameworks. This narrative review aims to systematically examine the bidirectional relationship between exoskeleton-based locomotor training and alterations in gut microbiome composition and function in the context of incomplete spinal cord injury.
Pathophysiology of Incomplete Spinal Cord Injury and Gastrointestinal Dysfunction
Neurological Consequences of Incomplete Spinal Cord Injury
Incomplete spinal cord injury represents a complex pathophysiological cascade initiated by primary mechanical trauma and perpetuated by secondary injury mechanisms that disrupt ascending and descending neural pathways with variable preservation of axonal connectivity.1 The American Spinal Injury Association (ASIA) Impairment Scale classifies incomplete injuries as grades B, C, and D, reflecting progressive degrees of motor and sensory preservation. This neurological preservation forms the substrate for potential functional recovery through activity-dependent neuroplasticity and reorganization of spared neural circuits.7 The neurological consequences extend beyond sensorimotor deficits to encompass autonomic dysregulation affecting multiple organ systems. Disruption of descending autonomic pathways results in varying degrees of sympathetic and parasympathetic dysfunction, manifesting as cardiovascular instability, thermoregulatory impairment, and visceral organ dysfunction.8 The extent of autonomic dysfunction correlates with lesion level and completeness, with cervical and high thoracic injuries typically producing more profound autonomic impairments due to interruption of sympathetic outflow from the thoracolumbar spinal cord (T1-L2).9
Gastrointestinal Dysfunction Following Spinal Cord Injury
Gastrointestinal dysfunction represents a significant consequence of spinal cord injury, affecting up to 80% of patients and substantially impacting quality of life.10 These complications result from disruption of the coordinated neural control of digestive functions, encompassing both autonomic and somatic nervous system components.11
Following SCI, the disruption of descending autonomic pathways results in altered sympathetic and parasympathetic input to the gastrointestinal tract, leading to dysmotility, altered secretory function, and impaired sphincter control.11 Upper motor neuron lesions (above T12) typically result in reduced colonic motility and prolonged transit time due to loss of coordinated parasympathetic control, while lower motor neuron lesions may cause reduced contractile function and sphincter hypotonia.11,12 Beyond motility disorders, SCI induces significant alterations in gastrointestinal mucosal integrity and barrier function. Experimental models demonstrate increased intestinal permeability associated with tight junction protein disruption and mucosal inflammation.5,13 This barrier dysfunction facilitates bacterial translocation and systemic inflammation, potentially exacerbating neuroinflammation and impeding neurological recovery.5
Neurogenic Bowel Dysfunction and Its Classification
Neurogenic bowel dysfunction (NBD) represents a constellation of gastrointestinal symptoms following SCI, including constipation, incontinence, abdominal pain, and bloating, which significantly impact quality of life and social participation.14 NBD is typically classified into two patterns based on the predominant pathophysiology: (1) Upper Motor Neuron Bowel Syndrome: Resulting from lesions above the conus medullaris (T12), characterized by increased colonic transit time, hyperreflexic rectal wall, increased anal tone, and loss of voluntary sphincter control.15 (2) Lower Motor Neuron Bowel Syndrome: Associated with lesions at the conus medullaris or cauda equina, characterized by areflexic bowel, reduced colonic motility, and decreased anal tone.15 The clinical management of NBD typically involves a multifaceted approach incorporating dietary modifications, pharmacological interventions, and bowel training programs.14 However, the efficacy of these conventional approaches is often suboptimal, highlighting the need for novel therapeutic strategies addressing the underlying pathophysiological mechanisms, including potential modulation of the gut microbiota.
The Gut Microbiota-Brain Axis: Structure and Function
Composition and Function of the Gut Microbiota
The human gastrointestinal tract harbors a complex microbial ecosystem comprising approximately 1013–1014 microorganisms, collectively termed the gut microbiota, which encompasses a diverse array of bacteria, archaea, fungi, viruses, and protozoa.16 Bacterial communities predominate, with the Firmicutes and Bacteroidetes phyla typically constituting 90% of the intestinal microbiota, accompanied by members of the Proteobacteria, Actinobacteria, Verrucomicrobia, and Fusobacteria phyla in varying proportions.17 The gut microbiome, encompassing the collective genomes of the microbiota, contains approximately 3.3 million non-redundant genes, exceeding the human genome by a factor of 150.18 These functions include fermentation of indigestible dietary components, synthesis of essential vitamins and amino acids, metabolism of xenobiotics, development and regulation of the immune system, and maintenance of intestinal barrier integrity.19 Microbial metabolic activities generate diverse bioactive compounds that influence host physiology at both local and systemic levels. Short-chain fatty acids (SCFAs), primarily acetate, propionate, and butyrate, represent principal metabolites produced through bacterial fermentation of dietary fiber.20 These SCFAs serve as energy substrates for colonocytes, regulate intestinal pH and oxygen tension, modulate immune function, and influence systemic metabolism through interaction with G-protein-coupled receptors.20,21 Beyond SCFAs, the gut microbiota synthesizes and modifies numerous compounds with neuroactive properties, including neurotransmitters (eg, γ-aminobutyric acid, serotonin, dopamine), neurotransmitter precursors (eg, tryptophan metabolites), and neuropeptides, which may influence central nervous system function through various pathways.22
Communication Pathways of the Gut-Brain Axis
The gut-brain axis represents a bidirectional communication network integrating the central nervous system, the autonomic nervous system, the enteric nervous system, and the gut microbiota through neural, immune, endocrine, and metabolic pathways.23 This complex interorgan communication system enables coordinated responses to environmental challenges and maintains homeostasis across multiple physiological domains. The vagus nerve constitutes a primary neural communication pathway within the gut-brain axis, transmitting signals bidirectionally between the gastrointestinal tract and the central nervous system.24 Vagal afferents detect various mechanical and chemical stimuli within the gastrointestinal tract, including microbially-derived metabolites such as SCFAs, which can activate vagal signaling through interaction with enteroendocrine cells.24,25 The gut-associated lymphoid tissue (GALT) represents the largest immune compartment in the body and serves as a critical interface between the microbiota and the host immune system.26 Intestinal microbiota and their metabolites modulate local immune responses through interaction with pattern recognition receptors on intestinal epithelial cells and immune cells, influencing cytokine production, T cell differentiation, and regulatory T cell development.26
The gut microbiota interacts with the host endocrine system through multiple mechanisms, including modulation of enteroendocrine cell function, metabolism of steroid hormones, and influence on the hypothalamic-pituitary-adrenal (HPA) axis.27 Enteroendocrine cells secrete various peptide hormones in response to luminal factors, including microbially-derived metabolites.28 These hormones regulate digestive functions and may influence central nervous system activity through endocrine, paracrine, or neural mechanisms. Microbially-derived metabolites represent significant mediators of gut-brain communication, with SCFAs serving as prototypical examples.20 These SCFAs can influence central nervous system function through multiple mechanisms, including direct action on central nervous system cells following passage across the blood-brain barrier, modulation of systemic immune responses affecting neuroinflammation, stimulation of enteroendocrine cells to release hormones that act centrally, and activation of vagal afferents either directly or indirectly through interaction with enteroendocrine cells.29
Gut Microbiota in Neurological Health and Disease
The gut microbiota plays a fundamental role in neurodevelopment and maintenance of neurological function, as evidenced by studies in germ-free animals demonstrating altered brain development, neurotransmitter systems, and behavior.30 During critical developmental periods, the gut microbiota contributes to the establishment of normal blood-brain barrier integrity, microglial maturation, myelination patterns, and synaptic pruning.30,31 Alterations in gut microbiota composition and function have been associated with numerous neurological and psychiatric conditions, including Parkinson’s disease, Alzheimer’s disease, multiple sclerosis, autism spectrum disorders, depression, and anxiety disorders.3 While causal relationships remain to be fully established, mechanistic studies suggest several pathways through which gut dysbiosis may contribute to neurological pathology, including increased intestinal permeability leading to systemic inflammation and neuroinflammation, altered production of neuroactive microbial metabolites, immune system dysregulation affecting central nervous system function, modulation of the enteric nervous system with consequences for vagal signaling, and endocrine disruption affecting stress responses and brain function.3 Targeting the gut microbiota through dietary interventions, probiotics, prebiotics, and fecal microbiota transplantation has shown promise in ameliorating certain neurological and psychiatric symptoms in both preclinical models and clinical studies.32 These findings collectively suggest a bidirectional relationship between gut microbiota and neurological health, with significant implications for understanding and treating neurological disorders, potentially including spinal cord injury.
Gut Microbiota Alterations Following Spinal Cord Injury
Characterization of Post-SCI Dysbiosis
Spinal cord injury induces profound alterations in gut microbiota composition and function, collectively termed dysbiosis, documented in both animal models and human patients.4–6 These microbiome perturbations manifest rapidly following injury and persist chronically, suggesting relevance to both acute pathophysiological processes and long-term complications. In rodent models of SCI, 16S rRNA gene sequencing analyses reveal consistent patterns of dysbiosis characterized by reduced alpha diversity (species richness and evenness), altered beta diversity (community composition differences), and taxonomic shifts including decreased abundance of Bacteroidetes and increased abundance of Firmicutes at the phylum level.5,6 At finer taxonomic resolution, SCI induces significant reductions in beneficial bacterial genera such as Lactobacillus, Bifidobacterium, and Bacteroides, with concurrent increases in potentially pathogenic taxa including Clostridium, Enterococcus, and certain Proteobacteria.5,6
Human studies corroborate these findings, with cross-sectional analyses demonstrating significant differences in microbiota composition between SCI patients and healthy controls.4 Microbiome alterations in human SCI include reduced diversity, increased Firmicutes/Bacteroidetes ratio, and enrichment of opportunistic pathogens, paralleling observations in experimental models.4 Furthermore, metagenomic analyses indicate alterations in microbial functional capacity following SCI, with decreased abundance of genes involved in SCFA production, anti-inflammatory functions, and intestinal barrier maintenance.33 The extent and nature of dysbiosis appear to correlate with injury characteristics, with more severe dysbiosis observed in complete compared to incomplete injuries and in cervical/high thoracic injuries compared to lower thoracic/lumbar lesions. This association with injury level and completeness suggests that disruption of descending autonomic pathways, particularly sympathetic innervation to the gastrointestinal tract, may contribute significantly to post-SCI dysbiosis.
Mechanisms Contributing to Post-SCI Dysbiosis
Multiple interrelated mechanisms contribute to the development and maintenance of gut dysbiosis following spinal cord injury. The autonomic nervous system provides critical regulation of gastrointestinal function through sympathetic and parasympathetic pathways, which are disrupted following SCI. Following injury, particularly with lesions above T6, disruption of descending autonomic pathways results in sympathetic dysregulation manifesting as either reduced sympathetic tone or uncoordinated sympathetic outflow during autonomic dysreflexia episodes.8 These autonomic disturbances alter the intestinal microenvironment through changes in motility, transit time, mucus secretion, and epithelial permeability, thereby modifying ecological niches within the gastrointestinal tract and selecting for altered microbial communities.34
Neurogenic bowel dysfunction following SCI typically manifests as reduced colonic motility and prolonged transit time, particularly with upper motor neuron lesions.14 This decreased intestinal transit provides extended substrate availability for microbial fermentation and allows more extensive bacterial colonization of the proximal gastrointestinal tract, potentially contributing to small intestinal bacterial overgrowth and alterations in community structure throughout the intestine.35
SCI induces significant intestinal barrier dysfunction characterized by epithelial tight junction disruption, increased permeability, and mucosal inflammation.5,13 This barrier compromise allows translocation of bacterial products and even viable bacteria across the intestinal epithelium, triggering local and systemic inflammatory responses.5 The resulting inflammation further compromises barrier function through cytokine-mediated effects on tight junction proteins and epithelial cell viability, creating a positive feedback loop that perpetuates barrier dysfunction and bacterial translocation.36 Following SCI, both local intestinal immunity and systemic immune function undergo significant alterations characterized by initial hyperinflammation followed by immunosuppression.37 These immunological disturbances modify host-microbiota interactions, potentially contributing to dysbiosis through altered selective pressures on microbial communities. Specifically, changes in antimicrobial peptide production, altered mucus layer characteristics, and dysregulated IgA secretion following SCI may permit expansion of opportunistic pathogens and reduction of beneficial symbionts.5,13 Following SCI, patients frequently experience alterations in dietary intake, physical activity, and medication use, all of which significantly impact gut microbiota composition and function.38 Reduced dietary diversity, increased consumption of processed foods, and inadequate fiber intake common among SCI patients may reduce substrates for beneficial microbial fermentation and promote dysbiosis.39 Physical inactivity, an inevitable consequence of motor impairment following SCI, may further contribute to microbiome alterations.40,41
Consequences of Post-SCI Dysbiosis for Neurological Recovery
Emerging evidence suggests that gut dysbiosis following SCI may not merely represent a secondary consequence of injury but may actively contribute to neurological pathophysiology and influence recovery trajectories through various mechanisms.5,42 Gut dysbiosis following SCI is associated with intestinal barrier dysfunction and increased translocation of microbial products such as lipopolysaccharide (LPS) into the circulation, triggering systemic inflammation.5 This systemic inflammatory state may exacerbate neuroinflammation through multiple pathways, including direct transport of inflammatory mediators across the compromised blood-spinal cord barrier, activation of peripheral immune cells that subsequently migrate to the injury site, and stimulation of neural inflammatory pathways through vagal afferents.5
The gut microbiota produces numerous metabolites with neuroactive properties, including neurotransmitters, neurotransmitter precursors, and SCFAs, which may influence neuronal function and neuroplasticity.22 Dysbiosis following SCI alters the production of these neuroactive compounds, potentially impacting neurological recovery processes.6 SCFAs, particularly butyrate, exhibit neuroprotective and neuroregenerative properties through mechanisms including histone deacetylase (HDAC) inhibition, microglial phenotype modulation, and blood-brain barrier integrity enhancement.43 Bidirectional interactions between the gut microbiota and the autonomic nervous system are well-established, with experimental evidence demonstrating that gut microbial communities can modulate both sympathetic and parasympathetic activity through various mechanisms including stimulation of vagal afferents, production of neurotransmitters, and regulation of neuroendocrine responses.24 Dysbiosis following SCI may exacerbate autonomic dysfunction through these pathways, potentially contributing to cardiovascular instability, thermoregulatory impairment, and further gastrointestinal dysfunction.44 Neuroplasticity represents a fundamental mechanism underlying functional recovery following incomplete spinal cord injury.7 Emerging evidence suggests that the gut microbiota may significantly influence neuroplasticity through multiple pathways, including modulation of microglial function, regulation of brain-derived neurotrophic factor (BDNF) expression, and production of metabolites affecting synaptic plasticity.30 Given that activity-dependent plasticity of spared neural circuits represents a primary therapeutic target in incomplete SCI rehabilitation, microbiota-mediated influences on neuroplasticity may significantly impact recovery potential and response to rehabilitative interventions, including exoskeleton-assisted gait training.
Exoskeleton Robot Technology for SCI Rehabilitation
Evolution and Classification of Rehabilitation Exoskeletons
Robotic exoskeletons for neurorehabilitation represent the culmination of decades of technological advancement in robotics, materials science, control systems, and rehabilitation engineering. The evolution of these devices has progressed from passive orthoses providing structural support to sophisticated powered systems capable of generating coordinated, task-specific movements with adaptive control algorithms.45 Rehabilitation exoskeletons can be classified according to multiple parameters, including actuation mechanism (passive, active, or hybrid), structural configuration (tethered or untethered), control strategy (position control, impedance control, assistance-as-needed, or bioelectrical signal-based control), and clinical application (rehabilitation or assistive purposes).2,45,46 Contemporary rehabilitation exoskeletons for SCI patients include stationary systems such as the Lokomat (Hocoma AG, Switzerland), which incorporates a treadmill and body-weight support system, and overground walking systems such as the Ekso (Ekso Bionics, USA), ReWalk (ReWalk Robotics, Israel), and HAL (Cyberdyne, Japan), which enable locomotion through various environments.2
Biomechanical and Neurophysiological Principles of Exoskeleton Training
The therapeutic efficacy of exoskeleton-assisted gait training derives from integration of fundamental biomechanical and neurophysiological principles that collectively promote functional recovery through structured, task-specific practice aligned with neuroplasticity mechanisms. Exoskeletons facilitate high-volume, consistent repetition of gait patterns, leveraging principles of motor learning and activity-dependent neuroplasticity.47 This repetitive, task-specific practice promotes reorganization of spared neural circuits and strengthening of synaptic connections involved in locomotion, potentially enhancing functional connectivity within the spinal cord and between spinal and supraspinal centers. Locomotor recovery following incomplete SCI relies significantly on activation and modulation of spinal locomotor networks, including central pattern generators (CPGs) capable of generating rhythmic motor output with appropriate afferent input.7 Exoskeleton training provides structured, rhythmic sensory input to these spinal networks through cyclical loading, proprioceptive signaling, and cutaneous feedback during gait phases.48 This patterned afferent stimulation may “awaken” dormant spinal circuits and promote their reorganization through activity-dependent plasticity mechanisms. Exoskeleton systems enable precise control of biomechanical loading parameters, allowing progressive challenge to neuromuscular systems according to established principles of exercise physiology and neural adaptation.49 Graduated reduction in robotic assistance, incremental increases in walking speed or distance, and systematic modification of support parameters facilitate progressive overload necessary for continued adaptation and functional improvement.
In addition to promoting neurological recovery, exoskeleton training provides mechanical benefits including reduction of spasticity through rhythmic stretching, prevention of contractures through maintenance of joint range of motion, and mitigation of secondary orthopedic complications associated with immobility.50 These mechanical effects complement neuroplasticity-targeted interventions, creating a comprehensive rehabilitation approach addressing multiple dimensions of post-SCI disability.
Clinical Evidence for Exoskeleton Training Efficacy in Incomplete SCI
A substantial body of evidence supports the efficacy of exoskeleton-assisted gait training in improving functional outcomes following incomplete spinal cord injury, though methodological heterogeneity and limitations necessitate cautious interpretation of current findings.51,52 Randomized controlled trials comparing exoskeleton training to conventional physical therapy in iSCI patients demonstrate significant improvements in various functional measures, including walking speed, endurance, independence, and lower extremity motor scores.51,52 A systematic review and meta-analysis by Nam et al (2017)53 found significant benefits of robotic-assisted gait training on walking independence (odds ratio 3.39, 95% CI 1.50–7.69) and walking capacity (standardized mean difference 0.59, 95% CI 0.23–0.96) compared to conventional therapy in subacute SCI patients. The efficacy of exoskeleton training appears to vary with patient characteristics and injury parameters, with patients with American Spinal Injury Association (ASIA) Impairment Scale C and D classifications (motor incomplete) typically demonstrating greater functional improvements than those with ASIA A and B classifications (motor complete).53
Beyond ambulatory function, exoskeleton training yields improvements in secondary health measures including spasticity reduction, pain modulation, and enhanced cardiovascular, pulmonary, and metabolic parameters.50 Neurophysiological studies utilizing transcranial magnetic stimulation, electroencephalography, and functional magnetic resonance imaging demonstrate that exoskeleton training induces measurable neuroplastic changes, including enhanced corticospinal excitability, modified cortical activation patterns, and strengthened functional connectivity between motor centers.54 While current evidence supports the efficacy of exoskeleton training in iSCI rehabilitation, significant research gaps remain, including optimal protocol parameters, long-term outcome sustainability, and cost-effectiveness compared to conventional interventions.51 Furthermore, the mechanisms underlying observed improvements, particularly regarding non-motor outcomes and systemic physiological effects, remain incompletely understood, highlighting the need for mechanistic investigations exploring novel pathways such as the gut microbiota-brain axis.
Effects of Exoskeleton Training on Gut Microbiota in Spinal Cord Injury
Evidence of Exercise-Induced Microbiome Modulation
Physical exercise has emerged as a significant modulator of gut microbiota composition and function, with potential implications for exoskeleton-assisted rehabilitation in SCI patients. Systematic reviews of both animal and human studies demonstrate consistent exercise-induced alterations in microbial communities, though specific changes vary with exercise modality, intensity, duration, and host characteristics.40,41 In healthy individuals, longitudinal studies of exercise training programs reveal increased microbial diversity, modified community composition, and beneficial taxonomic shifts including enrichment of Faecalibacterium, Roseburia, Akkermansia, and Bifidobacterium species associated with anti-inflammatory and metabolic benefits.40,41 These microbial changes correlate with improved metabolic parameters, reduced inflammatory markers, and enhanced intestinal barrier function. While various activity-based rehabilitation paradigms demonstrate beneficial effects on recovery following SCI, exoskeleton-assisted gait training offers unique mechanistic advantages for gut microbiome modulation that distinguish it from alternative interventions. Unlike stationary modalities such as functional electrical stimulation (FES) cycling, upper extremity ergometry, or body-weight supported treadmill training with partial unloading, exoskeleton-mediated locomotion provides comprehensive physiological stimulation through several integrated pathways: (1) Orthostatic hemodynamic regulation: The sustained upright positioning during exoskeleton ambulation uniquely challenges baroreceptor reflexes and induces splanchnic blood flow redistribution, potentially enhancing mesenteric perfusion and intestinal mucosal oxygenation in ways that supine or seated exercise modalities cannot replicate. This orthostatic challenge may restore diurnal autonomic rhythms disrupted by chronic recumbency, thereby normalizing circadian-regulated gut microbial oscillations; (2) Mechanical transduction pathways: The rhythmic ground reaction forces and physiological joint loading patterns during overground exoskeleton walking generate mechanotransductive signals throughout the musculoskeletal system, stimulating mechanoreceptors, proprioceptors, and visceral afferents more comprehensively than non-weight-bearing or partially unloaded activities. These mechanical signals may influence intestinal motility patterns and transit time through somatovisceral reflexes; (3) Cortico-autonomic integration: The cognitive demands of exoskeleton-assisted walking, requiring active participation in motor planning, postural control, and locomotor pattern generation, engage cortical and subcortical networks that interface with descending autonomic pathways. This volitional component, absent in passive or electrically-stimulated movements, may facilitate stronger top-down modulation of gastrointestinal function through prefrontal-hypothalamic-brainstem circuits; (4) Neuroendocrine modulation: The psychological and motivational benefits associated with restoration of upright ambulation may attenuate stress-related hypothalamic-pituitary-adrenal (HPA) axis dysregulation, reducing cortisol-mediated intestinal permeability and inflammation that contribute to dysbiosis. Furthermore, the achievement of locomotor milestones during exoskeleton training may enhance therapeutic alliance and treatment adherence, indirectly supporting gut health through improved nutritional intake and reduced psychological distress; and (5) Metabolic-microbial coupling: The higher metabolic demands of exoskeleton-assisted overground walking compared to stationary or supported exercises may alter systemic and local metabolic substrates available to gut microbiota, potentially selecting for beneficial metabolically-active species while suppressing opportunistic pathogens that thrive in the metabolically-compromised post-SCI environment. The mechanisms underlying exercise-mediated microbiome alterations include transit time modification, bile acid metabolism changes, immune modulation, myokine signaling, and autonomic regulation.41,55–58 While limited research has directly examined the effects of exoskeleton training on gut microbiota in SCI patients, the demonstrated impact of exercise on microbiome composition in both healthy individuals and various clinical populations provides a mechanistic framework for understanding potential exoskeleton-induced microbiome modulation in this context.
Autonomic Effects of Exoskeleton Training and Implications for Gut Function
Exoskeleton-assisted gait training induces significant modulation of autonomic nervous system function, with potential implications for gut microbiota through alterations in gastrointestinal physiology and the intestinal microenvironment.50,59 In SCI patients, particularly those with high thoracic or cervical lesions disrupting supraspinal control of sympathetic outflow, cardiovascular and autonomic responses to orthostatism and exercise are often impaired.59 Exoskeleton training, by imposing orthostatic stress and facilitating functional lower extremity movement, challenges and potentially reconditions these autonomic responses through several mechanisms. The training provides orthostatic conditioning through repeated exposure to upright posture, potentially enhancing baroreflex sensitivity and vasomotor control.50 It modifies splanchnic blood flow patterns through gravitational effects and muscle pump activation, potentially influencing intestinal perfusion, mucosal oxygenation, and consequently the physicochemical environment experienced by gut microbiota. Weight-bearing, postural transitions, and cyclical loading during exoskeleton walking generate visceral afferent signaling from abdominal and pelvic organs, potentially modulating autonomic reflex arcs affecting gastrointestinal function.59 Regular exoskeleton training may favorably shift sympathovagal balance, potentially reducing sympathetic hyperactivity and enhancing parasympathetic tone characteristic of chronic SCI.59 This autonomic rebalancing could beneficially impact gut motility, secretion, and immune function, creating conditions more favorable for beneficial microbial communities. Limited research has directly examined autonomic adaptations to exoskeleton training in relation to gut function and microbiota composition. However, studies documenting improvements in cardiovascular parameters, thermoregulation, and autonomic symptomatology following robotic locomotor training provide a foundation for hypothesizing corresponding benefits for gastrointestinal autonomic function and, consequently, gut microbiota homeostasis.50,59
Immunomodulatory Effects of Exoskeleton Training
Exoskeleton-assisted gait training induces significant immunomodulatory effects that may influence gut microbiota composition and function through various pathways involving neuroimmune interactions, systemic inflammatory regulation, and local intestinal immune modulation.12,59 SCI typically induces a complex immunological disturbance characterized by initial hyperinflammation followed by chronic immunosuppression, with implications for susceptibility to infections, wound healing, and neurological recovery.37 Regular physical activity, including that facilitated by exoskeleton technology, may beneficially modulate this dysregulated immune function through several mechanisms. Skeletal muscle contraction during exoskeleton-assisted walking stimulates production of myokines with anti-inflammatory properties, including interleukin-6 (IL-6), IL-10, and IL-1 receptor antagonist (IL-1ra).57 Regular exercise has been shown to expand and enhance the function of regulatory T cells, which play critical roles in maintaining intestinal immune homeostasis and preventing inappropriate inflammation that could disrupt microbial communities.60
Physical activity modifies stress hormone profiles, including cortisol and catecholamines, which significantly influence immune function and intestinal physiology.56 Regular exercise enhances mucosal immune function, including secretory IgA production and antimicrobial peptide expression, which regulate host-microbiota interactions and maintain appropriate microbial community boundaries.56 The mechanical stimulation and altered hemodynamics associated with upright positioning and locomotion during exoskeleton training may enhance gut-associated lymphoid tissue (GALT) activity and development, potentially improving its regulatory influence on gut microbial communities.61 While specific research examining immunological adaptations to exoskeleton training in relation to gut microbiota in SCI patients remains limited, the documented immunomodulatory effects of physical activity in other populations provide a mechanistic framework for understanding potential benefits in this context.
Metabolic and Endocrine Influences on Gut Microbiota
Exoskeleton-assisted gait training induces significant metabolic and endocrine adaptations that may influence gut microbiota composition and function through various pathways involving altered substrate availability, hormone signaling, and metabolic regulation.50 SCI typically results in adverse metabolic consequences including reduced energy expenditure, altered body composition with increased adiposity, impaired glucose tolerance, and dyslipidemia, creating an unfavorable metabolic environment that may contribute to gut dysbiosis.50 The metabolic effects of exoskeleton-mediated locomotion on gut microbiota modulation remain an emerging area of investigation, with current evidence derived from heterogeneous study designs and varied outcome measures. While Gorgey et al (2017)50 provided preliminary observations in a limited case series (n=4) demonstrating increased energy expenditure (mean increase: 1.74 ± 0.48 kcal/min above resting) and enhanced activity levels during exoskeleton ambulation, these findings require cautious interpretation given the small sample size and absence of control comparisons. We explicitly acknowledge these methodological limitations while recognizing the value of preliminary data in guiding future research directions. More robust evidence emerges from complementary investigations employing larger cohorts and controlled designs. Kressler et al (2014)62 conducted a prospective intervention study (n=32) examining metabolic adaptations to robotic-assisted locomotor training in chronic SCI, demonstrating statistically significant improvements in insulin sensitivity assessed via homeostatic model assessment of insulin resistance (HOMA-IR; baseline: 3.8 ± 2.1 vs post-intervention: 2.4 ± 1.3, p<0.01), though this investigation utilized tethered robotic systems rather than mobile exoskeletons. The translational relevance to untethered exoskeleton systems requires further validation. Additionally, Astorino et al (2013)63 employed a randomized controlled design (n=13) to evaluate activity-based therapy incorporating weight-bearing and stepping components analogous to exoskeleton locomotion, reporting significant improvements in oral glucose tolerance test responses (area under the curve reduction: 18.4%, p=0.03) and fasting insulin concentrations (reduction: 22.7%, p=0.02), suggesting that the upright, weight-bearing characteristics inherent to exoskeleton training may confer metabolic benefits relevant to gut microbiome modulation.
Regular exoskeleton training may theoretically enhances insulin sensitivity and glucose tolerance through increased glucose transport protein (GLUT4) expression, enhanced insulin signaling, and improved muscular glucose utilization.50 These metabolic improvements could potentially reduce intestinal glucose availability, theoretically limiting the growth of opportunistic pathogens that preferentially utilize simple sugars while promoting beneficial bacterial species adapted to more complex carbohydrate fermentation. However, direct causal relationships between exoskeleton-induced metabolic improvements and specific alterations in gut microbial taxonomy remain empirically unestablished, representing a critical knowledge gap requiring systematic investigation. Exoskeleton-assisted activity may beneficially modulate adipokine production, including reduced leptin and increased adiponectin levels, which influence inflammatory states and energy homeostasis.50 Regular physical activity modifies bile acid metabolism, potentially altering the intestinal bile acid profile which serves as both a nutrient source and regulator for certain microbial species.41 Exercise influences the secretion of various gastrointestinal hormones, including glucagon-like peptide-1 (GLP-1), peptide YY (PYY), and ghrelin, which regulate motility, secretion, and appetite.64 By normalizing these hormonal signals, exoskeleton training may improve gastrointestinal function and create conditions more favorable for beneficial microbial communities. Critical evaluation of the current evidence reveals substantial methodological limitations: (1) The paucity of studies directly examining exoskeleton-specific effects on metabolic-microbiome interactions; (2) Heterogeneity in device specifications, training protocols, and outcome measures limiting cross-study comparisons; (3) Absence of mechanistic investigations employing multi-omics approaches to elucidate causal pathways; and (4) Lack of longitudinal data characterizing temporal dynamics of metabolic-microbiome coupling during rehabilitation trajectories. These limitations underscore the preliminary nature of current knowledge while highlighting opportunities for rigorous future investigation employing adequately powered randomized controlled trials with integrated metabolomic and metagenomic profiling.
Mechanistic Pathways Linking Exoskeleton Training, Gut Microbiota, and Neurological Recovery
Microbiota-Derived Metabolites in Neuroplasticity and Neuroprotection
The gut microbiota produces numerous metabolites with neuroactive properties that may influence neuroplasticity and neuroprotection following spinal cord injury, providing a potential mechanistic link between exoskeleton-induced microbiome modulation and enhanced neurological recovery.43 However, it is critical to establish that these microbial metabolites function as modulatory rather than primary determinants of neuroplasticity, complementing endogenously synthesized neurotransmitters within the central nervous system. These microbial metabolites can access the central nervous system through various routes, including crossing the blood-brain barrier, activating vagal afferents, and modulating immune cells that subsequently migrate to the central nervous system.3
Short-chain fatty acids (SCFAs), particularly butyrate, propionate, and acetate, represent the most extensively studied microbial metabolites with neuroactive properties.43 These SCFAs exert multiple effects on neurological function and recovery, including histone deacetylase (HDAC) inhibition, microglial phenotype modulation, and blood-spinal cord barrier protection.43,65 Quantitative analysis reveals that while plasma SCFA concentrations typically range from 50–150 μM in healthy individuals, these levels decrease by approximately 30–40% following SCI-induced dysbiosis, potentially creating suboptimal conditions for neuroplastic processes rather than completely blocking them.66,67 Butyrate promotes oligodendrocyte differentiation and enhances expression of myelin-related genes, potentially facilitating remyelination of spared axons following incomplete SCI.68
Beyond SCFAs, the gut microbiota produces or modifies numerous other compounds with potential implications for neurological recovery. Regarding neurotransmitter production, it is essential to clarify the mechanistic constraints: while enteric bacteria synthesize gamma-aminobutyric acid (GABA), serotonin, and dopamine, these peripherally-produced neurotransmitters exhibit limited blood-brain barrier permeability. For instance, approximately 90% of the body’s serotonin is synthesized by enterochromaffin cells under microbial influence; however, this peripheral pool cannot directly cross the blood-brain barrier due to its hydrophilic nature. Instead, microbially-influenced peripheral neurotransmitters may indirectly modulate central nervous system function through: (1) Vagal afferent activation, with mechanistic studies demonstrating that bacterial-derived GABA can activate vagal signaling at concentrations as low as 10 μM;69 (2) Modulation of circulating tryptophan availability, as gut bacteria compete for tryptophan metabolism, potentially affecting the 1–3% of dietary tryptophan that crosses the blood-brain barrier for central serotonin synthesis;70 and (3) Influence on enteric nervous system function, which contains approximately 500 million neurons and can modulate ascending neural signaling independent of direct neurotransmitter translocation.22
The mechanistic relationship between altered microbial metabolite production and neuroplasticity following SCI requires nuanced interpretation. Rather than suggesting a binary “blocking” effect, current evidence supports a model of graduated modulation wherein dysbiosis-associated metabolite alterations create a neurochemical milieu that may be suboptimal but not prohibitive for neuroplastic processes. Specifically, reduced SCFA production following SCI may: (1) Diminish HDAC inhibition, resulting in decreased histone acetylation and potentially reduced expression of neuroplasticity-related genes (estimated 15–25% reduction based on in vitro studies);71 (2) Shift microglial phenotypes toward a more pro-inflammatory state, as butyrate concentrations below 0.5 mM fail to adequately suppress microglial activation;72 and (3) Compromise blood-spinal cord barrier integrity, as propionate has been shown to enhance barrier function through upregulation of tight junction proteins at physiological concentrations (50–100 μM).73
Furthermore, the temporal dynamics of metabolite-neuroplasticity interactions remain incompletely characterized. Acute post-injury phases may exhibit heightened sensitivity to metabolite fluctuations due to concurrent neuroinflammatory processes, while chronic phases may demonstrate adaptive compensation mechanisms that partially mitigate metabolite deficiencies. Exoskeleton training, by beneficially modulating gut microbiota composition and function, may enhance the production of these neuroprotective and neuroplasticity-promoting metabolites, thereby augmenting the direct neurological benefits of the locomotor activity itself. This synergistic effect should be conceptualized as optimization of the neurochemical environment for plasticity rather than reversal of a complete blockade, acknowledging that neuroplasticity following incomplete SCI involves complex interactions between multiple signaling systems of which microbial metabolites represent one modulatory component. This synergistic effect may be particularly relevant in incomplete SCI, where preserved neural pathways provide a substrate for both activity-dependent plasticity and responsiveness to microbiota-derived neuroactive compounds.
Neuroinflammatory Modulation Through the Gut-Brain Axis
Neuroinflammation represents a critical determinant of secondary injury progression and functional recovery following SCI, with both detrimental and reparative roles depending on timing, magnitude, and cellular phenotypes involved.37 The gut microbiota significantly influences neuroinflammatory processes through various pathways, providing another potential mechanism linking exoskeleton-induced microbiome modulation with enhanced neurological recovery.74 SCI typically induces a dysregulated inflammatory response characterized by initial hyperinflammation followed by chronic, low-grade inflammation that may impede recovery.37 Gut dysbiosis following SCI contributes to this inflammatory dysregulation through several mechanisms, including intestinal barrier disruption, Toll-like receptor activation, T cell differentiation regulation, and microglial phenotype modulation.5,74–76 Exoskeleton training may beneficially modulate these neuroinflammatory pathways through microbiome-mediated mechanisms. Exercise promotes the expansion of beneficial bacterial taxa that produce anti-inflammatory metabolites and enhance intestinal barrier function.40 It increases the abundance of SCFA-producing bacteria and enhances fermentation capacity, potentially increasing production of these anti-inflammatory metabolites.40 Exoskeleton training may enhance vagal tone, activating the cholinergic anti-inflammatory pathway that suppresses systemic inflammation through α7 nicotinic acetylcholine receptor signaling on immune cells.24 Regular physical activity normalizes stress hormone profiles, which significantly influence both gut microbiota composition and inflammatory responses.56 The integration of these microbiome-mediated anti-inflammatory mechanisms with the direct neurological benefits of exoskeleton training represents a potential synergistic approach to optimizing recovery following incomplete SCI.
Autonomic Regulation and Neuroendocrine Pathways
The autonomic nervous system represents a critical bidirectional communication pathway within the gut-brain axis, with significant implications for both neurological recovery following SCI and gut microbiota regulation.24 Exoskeleton training may beneficially modulate these autonomic and neuroendocrine pathways, providing another mechanistic link between locomotor rehabilitation, microbiome health, and improved neurological outcomes. SCI disrupts descending autonomic pathways, resulting in varying degrees of sympathetic and parasympathetic dysfunction depending on injury level and completeness.8 This autonomic dysregulation contributes to gut dysbiosis through altered gastrointestinal motility, secretion, and blood flow, while simultaneously impairing neurological recovery through dysregulated stress responses and neuroinflammation.5,8 Exoskeleton training may ameliorate autonomic dysfunction through several neuromodulatory mechanisms, including vagal tone enhancement, sympathetic adaptation, hypothalamic-pituitary-adrenal (HPA) axis regulation, and enteric nervous system stimulation.56,58,59 The gut microbiota, in turn, significantly influences autonomic and neuroendocrine function through various mechanisms, including vagal afferent activation, neurotransmitter production, and stress hormone metabolism.22,24,77 The bidirectional relationship between autonomic function, gut microbiota, and neurological recovery creates a complex network of interactions that may be beneficially modulated by exoskeleton training. By simultaneously enhancing autonomic regulation through locomotor activity and promoting beneficial microbiota through altered gastrointestinal physiology, exoskeleton therapy may create a positive feedback loop that optimizes conditions for neurological recovery following incomplete SCI.
Integrative Model: The Exoskeleton-Gut-Brain Axis in iSCI Recovery
Based on the preceding mechanistic discussions, we propose an integrative model conceptualizing the “Exoskeleton-Gut-Brain Axis” as a complex, bidirectional communication network through which exoskeleton-assisted gait training may enhance neurological recovery following incomplete spinal cord injury via gut microbiota modulation. This model encompasses multiple interacting pathways that collectively influence both central and peripheral aspects of recovery. The mechanical-microbial pathway involves exoskeleton training providing repetitive, task-specific locomotor activation that directly stimulates neuroplastic mechanisms while simultaneously altering gastrointestinal mechanical function through postural changes, visceral organ movement, and altered hemodynamics. These mechanical changes modify the intestinal microenvironment, selecting for altered microbial communities that may produce metabolites supporting neural recovery and plasticity.
The pelvic floor neuromuscular complex represents a critical yet underinvestigated component of the exoskeleton-gut-brain axis, warranting systematic integration into our conceptual framework. During exoskeleton-assisted locomotion, the coordinated activation patterns of hip extensors, abductors, and trunk stabilizers facilitate synergistic recruitment of pelvic floor musculature through several mechanisms: (1) Myofascial force transmission via the thoracolumbar fascia and its continuity with the endopelvic fascia generates passive tension in pelvic floor structures during gait cycles, with electromyographic studies demonstrating 15–30% maximal voluntary contraction in levator ani muscles during normal walking patterns; (2) Neural overflow phenomena, wherein volitional activation of proximal hip musculature during exoskeleton-assisted stepping triggers involuntary co-activation of pelvic floor muscles through shared sacral nerve root innervation (S2-S4), potentially facilitating motor relearning in neurologically impaired pelvic floor circuits; (3) Intra-abdominal pressure modulation during weight-bearing phases of gait creates rhythmic loading-unloading cycles on pelvic floor structures, providing proprioceptive feedback that may enhance anorectal sensation and external anal sphincter recruitment patterns critical for continence control; and (4) Biomechanical coupling between lumbopelvic rhythm and pelvic floor activation, wherein the cyclical anterior-posterior pelvic tilt during exoskeleton ambulation facilitates differential activation of superficial versus deep pelvic floor muscle layers, potentially improving selective motor control necessary for coordinated defecation reflexes.
The implications for neurogenic bowel dysfunction management are multifaceted: Enhanced pelvic floor neuromuscular function may improve voluntary sphincter control, reducing fecal incontinence episodes reported in 75% of SCI patients; Rhythmic pelvic floor activation during gait may facilitate colorectal motility through mechanical stimulation of enteric nervous system mechanoreceptors, potentially reducing colonic transit time which increases by 2–4 fold following SCI; Improved pelvic floor proprioception may enhance anorectal sensory discrimination, critical for appropriate defecation timing in patients with disrupted supraspinal control; and Strengthened pelvic floor musculature may augment intra-abdominal pressure generation necessary for effective evacuation, addressing the paradoxical combination of constipation and incontinence characteristic of neurogenic bowel.
The autonomic-microbial pathway involves exoskeleton-mediated challenges to autonomic reflexes during upright posture and locomotion promoting autonomic adaptation and enhanced regulation, potentially improving sympathovagal balance and visceral organ function. This autonomic modulation alters gastrointestinal secretory, immunological, and vascular parameters, creating conditions more favorable for beneficial microbial communities that produce metabolites enhancing autonomic regulation through vagal afferent stimulation and neuroendocrine effects. The neuroimmune-microbial pathway involves physical activity and stress reduction associated with successful exoskeleton locomotion modulating neuroimmune interactions, potentially reducing neuroinflammation and systemic inflammatory tone. This immunomodulation influences gut barrier function and microbial community selection, favoring anti-inflammatory microbes that further dampen inflammatory responses through metabolite production and immune cell phenotype modification.
Within this integrated model (Figure 1), temporal dynamics represent critical considerations, with different pathways potentially predominating at various phases post-injury and during the rehabilitation process. Acute phase responses (0–4 weeks post-initiation) may demonstrate heightened mechanosensitivity and inflammatory modulation; subacute adaptations (4–12 weeks) likely involve metabolic reprogramming and microbial community restructuring; while chronic phase consolidation (>12 weeks) may establish stable neuroimmune-metabolic equilibria supporting sustained functional improvements. This integrative model provides a comprehensive framework for understanding how exoskeleton training may influence neurological recovery through previously unrecognized gut-brain interactions, complementing the established direct neurophysiological benefits of locomotor training.
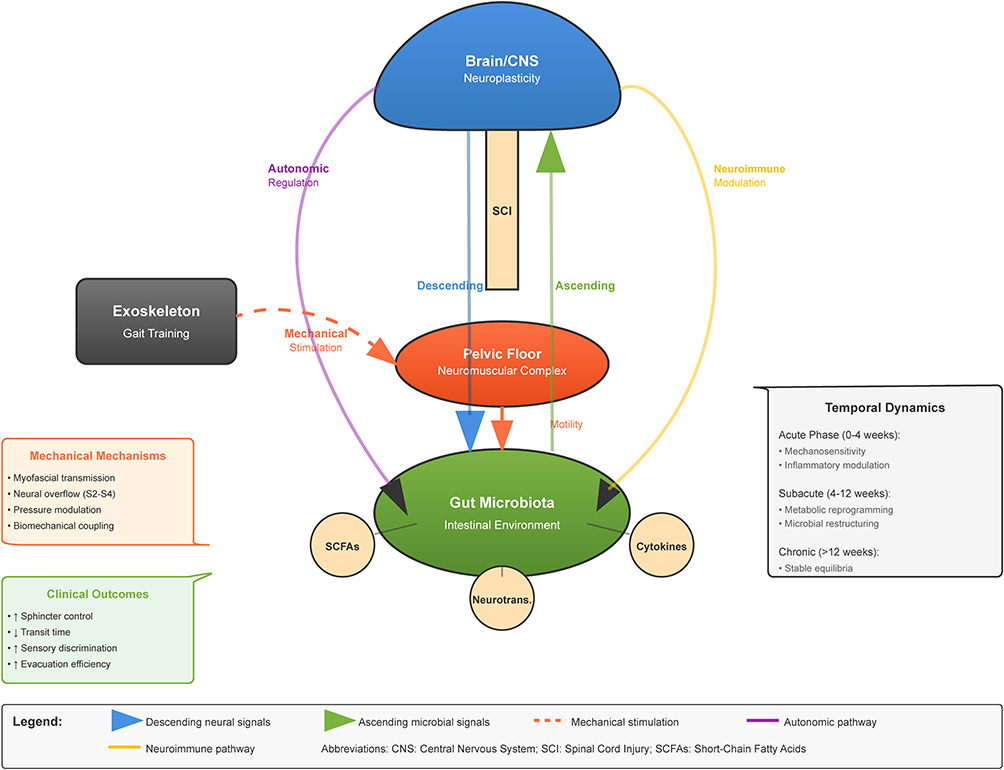 |
Figure 1 Integrative Model of the Exoskeleton-Gut-Brain Axis in Incomplete Spinal Cord Injury Rehabilitation. |
Clinical Implications and Therapeutic Applications
Potential for Integrated Rehabilitation Approaches
Recognition of the potential influence of exoskeleton training on the gut microbiota-brain axis offers significant opportunities for developing integrated rehabilitation approaches that simultaneously target locomotor function, autonomic regulation, and microbial homeostasis to optimize recovery following incomplete spinal cord injury. Integration of targeted nutritional strategies with exoskeleton training represents a promising approach to synergistically modulate the gut microbiota in support of neurological recovery. Prebiotic supplementation with fermentable dietary fibers can selectively promote the growth of beneficial bacterial species, particularly those producing SCFAs with neuroprotective and anti-inflammatory properties.78 Combining prebiotics with exoskeleton training may enhance microbial responses to exercise and augment production of neuroactive metabolites supporting recovery. Specific probiotic strains, particularly those with demonstrated neurological benefits (psychobiotics), may complement exoskeleton-induced microbiome changes and accelerate restoration of beneficial microbial communities disrupted by SCI. Anti-inflammatory dietary patterns characterized by high polyphenol content, omega-3 fatty acids, and diverse plant fibers support both beneficial microbiota composition and neurological health.79 Implementing such dietary approaches alongside exoskeleton rehabilitation may create synergistic effects on neuroinflammation, oxidative stress, and neuroplasticity mechanisms. Strategic implementation of intermittent fasting or time-restricted feeding may enhance microbiome diversity and SCFA production while upregulating neuroprotective pathways through mechanisms including autophagy, reduced oxidative stress, and enhanced mitochondrial function.41 Strategies specifically targeting autonomic regulation may complement exoskeleton training to optimize both neurological recovery and gut microbial health. Structured breathing exercises incorporated before or after exoskeleton sessions may enhance vagal tone, improve sympathovagal balance, and reduce stress responses, potentially benefiting both neurological recovery and gut function.80 Non-invasive stimulation of vagal afferents through transcutaneous vagus nerve stimulation may enhance the autonomic benefits of exoskeleton training, potentially improving both neurological outcomes and gut microbial homeostasis through activation of the cholinergic anti-inflammatory pathway.81 The psychological dimensions of SCI recovery significantly influence both neurological outcomes and gut microbiota through neuroendocrine and immune pathways, warranting specific attention within integrated rehabilitation approaches. Structured mindfulness practices integrated with exoskeleton rehabilitation may reduce stress hormones, inflammatory markers, and psychological distress that negatively impact both neurological recovery and gut microbial health.82 Addressing catastrophizing, fear-avoidance behaviors, and other maladaptive psychological responses to SCI may reduce chronic stress states that promote gut dysbiosis and neuroinflammation.83
Biomarkers for Monitoring Rehabilitation Effects on the Gut-Brain Axis
Effective implementation of integrated rehabilitation approaches targeting the exoskeleton-gut-brain axis requires appropriate biomarkers for monitoring intervention effects, personalizing treatment protocols, and predicting outcomes. Several promising biomarker categories warrant consideration for monitoring microbiome-related effects of rehabilitation interventions. Taxonomic profiles through regular assessment of fecal microbiota composition using 16S rRNA sequencing or shotgun metagenomic approaches may track changes in microbial diversity and abundance of key bacterial taxa associated with neurological recovery.84 Beyond taxonomic identification, characterization of microbial functional capacity through metagenomic, metatranscriptomic, or metabolomic approaches may reveal changes in metabolic pathways relevant to neurological recovery.84 Quantification of neuroactive microbial metabolites in fecal, serum, or cerebrospinal fluid samples, particularly SCFAs, tryptophan metabolites, and neurotransmitter-related compounds, may provide direct evidence of functional microbiome alterations relevant to neurological recovery.43 Assessment of intestinal barrier function through biomarkers such as zonulin, lipopolysaccharide binding protein (LBP), soluble CD14, and intestinal fatty acid binding protein (I-FABP) may indicate improvements in barrier integrity associated with beneficial microbiome changes and reduced bacterial translocation.79
Monitoring pro-inflammatory and anti-inflammatory cytokines in serum or cerebrospinal fluid may reveal immunomodulatory effects of integrated rehabilitation approaches affecting both gut health and neurological recovery. Quantification of regulatory T cell populations and function may indicate enhanced immunoregulatory capacity associated with beneficial microbiome alterations and improved neurological outcomes. Heart rate variability analysis, including time-domain, frequency-domain, and non-linear metrics, provides a non-invasive window into autonomic function that may reflect both central adaptations to rehabilitation and gut-brain interaction improvements.80 Salivary or hair cortisol measurements may indicate neuroendocrine normalization associated with integrated rehabilitation approaches targeting the gut-brain axis.56 Advanced neuroimaging techniques including functional magnetic resonance imaging, diffusion tensor imaging, and magnetic resonance spectroscopy may reveal structural and functional neural adaptations associated with integrated exoskeleton and gut-brain axis interventions.85 Quantitative electroencephalographic parameters reflecting cortical activity, connectivity, and plasticity may indicate central nervous system adaptations that correlate with both functional improvements and gut-brain axis modulation.54
Personalized Rehabilitation Strategies Based on Microbiome Profiles
The substantial interindividual variation in gut microbiome composition, even among patients with similar injury characteristics, suggests potential for personalized rehabilitation approaches targeting the exoskeleton-gut-brain axis based on individual microbiome profiles and their predicted functional implications. Analysis of baseline microbiome composition may enable stratification of SCI patients into subgroups likely to respond differently to exoskeleton rehabilitation or requiring specific complementary interventions. Patients could be categorized based on the degree of microbial community disruption relative to healthy controls, with more severe dysbiosis potentially warranting more aggressive intervention.84 Assessment of the abundance and activity of SCFA-producing bacteria may identify patients with reduced capacity for producing these neuroprotective metabolites, potentially benefiting from specific dietary interventions alongside exoskeleton training.43 Microbiome signatures associated with compromised intestinal barrier function may identify patients at higher risk for bacterial translocation and systemic inflammation, warranting targeted interventions to restore barrier integrity.79 Microbial community configurations associated with enhanced pro-inflammatory potential may identify patients at higher risk for detrimental neuroinflammation, potentially benefiting from more intensive anti-inflammatory approaches. Based on microbiome profiling and associated biomarkers, personalized intervention protocols might include precision probiotic selection based on individual deficiencies identified through microbiome analysis, targeted prebiotic prescription based on identified deficiencies in microbial metabolic functions,78 and personalized dietary recommendations addressing individual microbiome characteristics.79 Exoskeleton training parameters including session frequency, duration, intensity, and progression could be tailored based on integrated assessment of both neurological and microbiome characteristics.53 Complementary autonomic training based on autonomic assessment in relation to microbiome profiles could complement exoskeleton sessions to optimize vagal tone and sympathovagal balance.80 True personalization requires not only baseline tailoring but ongoing adjustment based on individual responses. Regular reassessment of microbiome composition and function alongside neurological and functional parameters would enable dynamic adjustment of intervention protocols based on individual response patterns and trajectories.84 Integrated monitoring of both microbiome and neurological parameters may identify when recovery plateaus occur and suggest specific modifications to overcome them.53
Research Challenges and Future Directions
Methodological Challenges in Gut-Brain Axis Research
Investigating the relationships between exoskeleton training, gut microbiota, and neurological recovery following incomplete spinal cord injury presents substantial methodological challenges. Variation in sample collection procedures, including timing relative to meals and medications, stool consistency, storage conditions, and processing delays, can significantly impact microbiome analysis results.84 Standardized protocols incorporating immediate freezing, consistent collection timing, and detailed metadata documentation are essential for obtaining reliable, comparable results across studies. Different microbiome characterization methods, including 16S rRNA gene sequencing, shotgun metagenomic sequencing, metatranscriptomics, and metabolomics, provide complementary but distinct insights with varying depth, breadth, and functional resolution.84 Integration of multiple analytical approaches within single studies would provide more comprehensive understanding of both taxonomic and functional microbial changes associated with rehabilitation interventions. Numerous factors influence gut microbiota composition, including diet, medication use (particularly antibiotics), bowel management routines, and environmental exposures, creating substantial potential for confounding in rehabilitation studies.84 Detailed documentation of these variables, stratified randomization, and statistical approaches for controlling confounders are essential for isolating intervention effects. The bidirectional nature of gut-brain interactions creates challenges in determining whether observed microbiome changes represent causes or consequences of neurological and functional alterations.3 Interventional designs with temporal sequencing of changes, mechanistic biomarker assessment, and potentially gnotobiotic animal models would help elucidate directional relationships. The gut-brain axis encompasses multiple interacting systems—neural, immune, endocrine, and metabolic—each with distinct regulatory mechanisms and response dynamics, creating challenges for isolating specific pathways mediating observed effects.3 Multimodal assessment approaches incorporating simultaneous measurement of neural, immune, endocrine, and metabolic parameters would enable more comprehensive pathway mapping and interaction analysis.
Future Research Priorities
The emerging field of exoskeleton-gut-brain axis interactions in SCI rehabilitation presents numerous opportunities for transformative research that could significantly advance both mechanistic understanding and clinical applications. Systematic investigation of specific neural, immune, endocrine, and metabolic pathways mediating interactions between exoskeleton training, gut microbiota, and neurological recovery represents a critical research priority.3 Characterization of optimal timing relationships between rehabilitation initiation, microbiome modulation, and neurological recovery potential would inform more effective intervention sequencing.6 Systematic investigation of relationships between exoskeleton training parameters (intensity, duration, frequency), microbiome alterations, and neurological outcomes would enable more precise intervention prescription.53 Elucidation of specific mechanisms by which microbiota-derived metabolites access and influence the central nervous system following SCI, particularly considering blood-spinal cord barrier disruption and repair dynamics, represents an important mechanistic question.43 Comparative analysis of different rehabilitation modalities regarding their impact on gut microbiota and associated neurological outcomes would clarify whether observed effects are exoskeleton-specific or generalizable across mobility interventions.51 Development and validation of integrated assessment protocols combining microbiome analysis, metabolomics, immunological profiling, autonomic assessment, and detailed neurological/functional evaluation would enable more comprehensive characterization of gut-brain interactions in the rehabilitation context.84 Advancement of technologies enabling simultaneous real-time monitoring of gut and neurological parameters during rehabilitation sessions would provide unprecedented insights into immediate gut-brain interactions during exoskeleton use.24 Design and implementation of pragmatic clinical trials evaluating integrated rehabilitation approaches targeting both locomotor function and gut microbiota in real-world clinical settings would assess translational potential and implementation challenges.53 Systematic assessment of complementary interventions (eg, probiotics, prebiotics, dietary modifications) as adjuncts to exoskeleton rehabilitation would determine optimal integrated approaches for enhancing both microbial and neurological outcomes.
Translational Considerations
The translation of research findings regarding exoskeleton-gut-brain interactions into clinical practice requires careful consideration of various practical, ethical, and healthcare system factors. Integration of gut microbiome analysis and targeted interventions with exoskeleton rehabilitation would require additional resources, including specialized equipment, expertise, and time, potentially limiting implementation in resource-constrained settings.86 Management of combined rehabilitation and microbiome modulation protocols adds complexity to patient care, potentially increasing cognitive load for healthcare providers and adherence challenges for patients.51 Effective implementation requires coordination between rehabilitation specialists, gastroenterologists, dietitians, microbiologists, and potentially other specialists, creating logistical and communication challenges within traditionally siloed healthcare systems.86 Patient preferences regarding rehabilitation modalities, dietary modifications, and supplement use vary substantially and significantly impact adherence and satisfaction.83 The additional burden of microbiome-targeted interventions beyond already demanding rehabilitation protocols requires careful assessment and minimization.83 Novel integrated approaches incorporating microbiome modulation might generate unrealistic outcome expectations among patients eager for enhanced recovery, necessitating balanced communication about potential benefits, limitations, and uncertainties.83 Current healthcare reimbursement models may not adequately cover integrated approaches combining rehabilitation with microbiome assessment and intervention, particularly given limited large-scale efficacy evidence.52 Effective gut-brain targeted rehabilitation requires coordination across care transitions (inpatient rehabilitation to outpatient settings to home) that are often fragmented in current healthcare systems.86 Implementation of integrated gut-brain rehabilitation approaches requires expanding knowledge and skills among rehabilitation professionals regarding microbiome science, nutritional interventions, and relevant assessment techniques.86 Establishing appropriate quality metrics for integrated gut-brain rehabilitation approaches would enable performance monitoring, comparative effectiveness assessment, and continuous improvement activities.52
Conclusion: Toward an Integrated Neurorehabilitation Paradigm
The convergence of exoskeleton robot technology, gut microbiome science, and neurological rehabilitation represents a significant frontier in the management of incomplete spinal cord injury. This review has synthesized emerging evidence suggesting that beyond its direct biomechanical effects, exoskeleton-assisted gait training may influence neurological recovery through previously underrecognized mechanisms involving the gut microbiota-brain axis. Incomplete spinal cord injury induces significant gut dysbiosis characterized by reduced microbial diversity, altered taxonomic representation, and modified metabolic capacity, contributing to systemic inflammation, autonomic dysfunction, and potentially impaired neurological recovery. Concurrently, the gut microbiota produces numerous neuroactive compounds that influence central nervous system function through various neural, immune, endocrine, and metabolic pathways, creating a bidirectional communication network that may significantly impact recovery trajectories. Exoskeleton-assisted gait training, through its effects on autonomic function, immune regulation, and metabolic activity, appears capable of beneficially modulating the gut microbiome, potentially enhancing the production of neuroprotective and neuroplasticity-promoting microbial metabolites. These microbiome alterations may complement the direct neurophysiological benefits of locomotor training through multiple mechanisms, including reduction of neuroinflammation, enhancement of neurotrophic factor expression, promotion of favorable neuroendocrine profiles, and optimization of neuroenergetic metabolism.
The integrated “exoskeleton-gut-brain axis” model proposed in this review offers a framework for understanding these complex interactions and developing novel therapeutic approaches that simultaneously target both locomotor function and microbial homeostasis. Such integrated approaches might include combinations of exoskeleton training with targeted nutritional strategies, probiotic/prebiotic supplementation, autonomic regulation techniques, and stress reduction methods, creating synergistic effects on both microbiome composition and neurological recovery.
While substantial research challenges remain in fully elucidating the mechanisms and optimizing the implementation of gut-brain targeted rehabilitation approaches, the potential for transformative impact on patient outcomes warrants intensive investigation in this emerging field. Future research priorities should include mechanistic pathway delineation, critical window identification, dose-response characterization, development of integrated assessment protocols, and implementation of pragmatic clinical trials evaluating combined approaches in real-world settings.
The exploration of gut microbiota-brain axis modulation through exoskeleton-assisted gait training represents a promising new frontier in neurorehabilitation science, extending our understanding beyond traditional biomechanical paradigms to encompass the complex biological networks mediating neurological recovery. This integrated perspective offers the potential for developing more comprehensive, mechanistically-targeted rehabilitation approaches that maximize recovery potential for individuals with incomplete spinal cord injury.
Data Sharing Statement
All data generated or analyzed in this study are included in the present manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure
The authors have no conflicts of interest to declare for this work.
References
1. Ahuja CS, Wilson JR, Nori S. et al. Traumatic spinal cord injury. Nature Reviews Disease Primers. 2017;3(1):17018. doi:10.1038/nrdp.2017.18
2. Esquenazi A, Talaty M, Jayaraman A. Powered Exoskeletons for Walking Assistance in Persons with Central Nervous System Injuries: a Narrative Review. PM R J inj funct rehabilitat. 2017;9(1):46–62. doi:10.1016/j.pmrj.2016.07.534
3. Cryan JF, O’Riordan KJ, Cowan CSM, et al. The Microbiota-Gut-Brain Axis. Physiol Rev. 2019;99(4):1877–2013. doi:10.1152/physrev.00018.2018
4. Gungor B, Adiguzel E, Gursel I, Yilmaz B, Gursel M. Intestinal Microbiota in Patients with Spinal Cord Injury. PLoS One. 2016;11(1):e0145878. doi:10.1371/journal.pone.0145878
5. Kigerl KA, Hall JC, Wang L, Mo X, Yu Z, Popovich PG. Gut dysbiosis impairs recovery after spinal cord injury. J Exp Med. 2016;213(12):2603–2620. doi:10.1084/jem.20151345
6. O’Connor G, Jeffrey E, Madorma D, et al. Investigation of Microbiota Alterations and Intestinal Inflammation Post-Spinal Cord Injury in Rat Model. J Neurotrauma. 2018;35(18):2159–2166. doi:10.1089/neu.2017.5349
7. Dietz V, Fouad K. Restoration of sensorimotor functions after spinal cord injury. Brain. 2014;137(Pt 3):654–667. doi:10.1093/brain/awt262
8. Weaver LC, Fleming JC, Mathias CJ, Krassioukov AV. Disordered cardiovascular control after spinal cord injury. Handbook Clin Neurol. 2012;109:213–233.
9. West CR, Bellantoni A, Krassioukov AV. Cardiovascular function in individuals with incomplete spinal cord injury: a systematic review. Topics Spinal Cord Injury Rehab. 2013;19(4):267–278. doi:10.1310/sci1904-267
10. Tate DG, Forchheimer M, Rodriguez G, et al. Risk Factors Associated With Neurogenic Bowel Complications and Dysfunction in Spinal Cord Injury. Arch Phys Med Rehabil. 2016;97(10):1679–1686. doi:10.1016/j.apmr.2016.03.019
11. Ebert E. Gastrointestinal involvement in spinal cord injury: a clinical perspective. JGLD. 2012;21(1):75–82.
12. Hayes HB, Jayaraman A, Herrmann M, Mitchell GS, Rymer WZ, Trumbower RD. Daily intermittent hypoxia enhances walking after chronic spinal cord injury: a randomized trial. Neurology. 2014;82(2):104–113. doi:10.1212/01.WNL.0000437416.34298.43
13. Sun J, Wang F, Li H, et al. Neuroprotective Effect of Sodium Butyrate against Cerebral Ischemia/Reperfusion Injury in Mice. Biomed Res Int. 2015;2015:395895. doi:10.1155/2015/395895
14. Emmanuel A, Mahmoud SA, El Moshy S. Neurogenic bowel dysfunction. F1000Research. 2019;8:8. doi:10.12688/f1000research.17047.1
15. Krassioukov A, Eng JJ, Claxton G, Sakakibara BM, Shum S. Neurogenic bowel management after spinal cord injury: a systematic review of the evidence. Spinal Cord. 2010;48(10):718–733. doi:10.1038/sc.2010.14
16. Sender R, Fuchs S, Milo R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016;14(8):e1002533. doi:10.1371/journal.pbio.1002533
17. Rinninella E, Raoul P, Cintoni M, et al. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms. 2019;7(1):14. doi:10.3390/microorganisms7010014
18. Qin J, Li R, Raes J, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59–65. doi:10.1038/nature08821
19. Rowland I, Gibson G, Heinken A, et al. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. 2018;57(1):1–24. doi:10.1007/s00394-017-1445-8
20. Morrison DJ, Preston T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016;7(3):189–200. doi:10.1080/19490976.2015.1134082
21. Tan J, McKenzie C, Potamitis M, Thorburn AN, Mackay CR, Macia L. The role of short-chain fatty acids in health and disease. Adv Immunol. 2014;121:91–119.
22. Wall R, Cryan JF, Ross RP, Fitzgerald GF, Dinan TG, Stanton C. Bacterial neuroactive compounds produced by psychobiotics. Adv Exp Med Biol. 2014;817:221–239.
23. Carabotti M, Scirocco A, Maselli MA, Severi C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. 2015;28(2):203–209.
24. Bonaz B, Bazin T, Pellissier S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front Neurosci. 2018;12:49. doi:10.3389/fnins.2018.00049
25. Browning KN, Travagli RA. Central nervous system control of gastrointestinal motility and secretion and modulation of gastrointestinal functions. Compr Physiol. 2014;4(4):1339–1368. doi:10.1002/j.2040-4603.2014.tb00587.x
26. Thaiss CA, Zmora N, Levy M, Elinav E. The microbiome and innate immunity. Nature. 2016;535(7610):65–74. doi:10.1038/nature18847
27. Neuman H, Debelius JW, Knight R, Koren O. Microbial endocrinology: the interplay between the microbiota and the endocrine system. FEMS Microbiol Rev. 2015;39(4):509–521. doi:10.1093/femsre/fuu010
28. Martin AM, Sun EW, Rogers GB, Keating DJ. The Influence of the Gut Microbiome on Host Metabolism Through the Regulation of Gut Hormone Release. Front Physiol. 2019;10:428. doi:10.3389/fphys.2019.00428
29. Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. 2019;16(8):461–478.
30. Luczynski P, Ka MN, Oriach CS, Clarke G, Dinan TG, Cryan JF. Growing up in a Bubble: using Germ-Free Animals to Assess the Influence of the Gut Microbiota on Brain and Behavior. Int J Neuropsychopharmacol. 2016;19(8):pyw020. doi:10.1093/ijnp/pyw020
31. Hoban AE, Stilling RM, Ryan FJ, et al. Regulation of prefrontal cortex myelination by the microbiota. Transl Psychiatry. 2016;6(4):e774. doi:10.1038/tp.2016.42
32. Liu RT, Walsh RFL, Sheehan AE. Prebiotics and probiotics for depression and anxiety: a systematic review and meta-analysis of controlled clinical trials. Neurosci Biobehav Rev. 2019;102:13–23. doi:10.1016/j.neubiorev.2019.03.023
33. Wallace DJ, Sayre NL, Patterson TT, Nicholson SE, Hilton D, Grandhi R. Spinal cord injury and the human microbiome: beyond the brain-gut axis. Neurosurgical Focus. 2019;46(3):E11. doi:10.3171/2018.12.FOCUS18206
34. Cervi AL, Lukewich MK, Lomax AE. Neural regulation of gastrointestinal inflammation: role of the sympathetic nervous system. Autonomic Neurosci. 2014;182:83–88. doi:10.1016/j.autneu.2013.12.003
35. Quigley EMM. Gut microbiome as a clinical tool in gastrointestinal disease management: are we there yet? Nat Rev Gastroenterol Hepatol. 2017;14(5):315–320. doi:10.1038/nrgastro.2017.29
36. König J, Wells J, Cani PD, et al. Human Intestinal Barrier Function in Health and Disease. Clin Transl Gastroenterol. 2016;7(10):e196. doi:10.1038/ctg.2016.54
37. Schwab JM, Zhang Y, Kopp MA, Brommer B, Popovich PG. The paradox of chronic neuroinflammation, systemic immune suppression, autoimmunity after traumatic chronic spinal cord injury. Exp Neurol. 2014;258:121–129. doi:10.1016/j.expneurol.2014.04.023
38. David LA, Maurice CF, Carmody RN, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505(7484):559–563. doi:10.1038/nature12820
39. Levine AM, Nash MS, Green BA, Shea JD, Aronica MJ. An examination of dietary intakes and nutritional status of chronic healthy spinal cord injured individuals. Paraplegia. 1992;30(12):880–889. doi:10.1038/sc.1992.165
40. Allen JM, Mailing LJ, Niemiro GM, et al. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med Sci Sports Exercise. 2018;50(4):747–757. doi:10.1249/MSS.0000000000001495
41. Mailing LJ, Allen JM, Buford TW, Fields CJ, Woods JA. Exercise and the Gut Microbiome: a Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exer Sport Sci Rev. 2019;47(2):75–85. doi:10.1249/JES.0000000000000183
42. Myers SA, Gobejishvili L, Saraswat Ohri S, et al. Following spinal cord injury, PDE4B drives an acute, local inflammatory response and a chronic, systemic response exacerbated by gut dysbiosis and endotoxemia. Neurobiol Dis. 2019;124:353–363. doi:10.1016/j.nbd.2018.12.008
43. Silva YP, Bernardi A, Frozza RL. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol. 2020;11:25. doi:10.3389/fendo.2020.00025
44. Furness JB, Callaghan BP, Rivera LR, Cho HJ. The enteric nervous system and gastrointestinal innervation: integrated local and central control. Adv Exp Med Biol. 2014;817:39–71.
45. Vélez-Guerrero MA, Callejas-Cuervo M, Mazzoleni S. Artificial Intelligence-Based Wearable Robotic Exoskeletons for Upper Limb Rehabilitation: a Review. Sensors. 2021;21(6):1.
46. Sawicki GS, Beck ON, Kang I, Young AJ. The exoskeleton expansion: improving walking and running economy. J Neuroeng Rehabil. 2020;17(1):25. doi:10.1186/s12984-020-00663-9
47. Edgerton VR, Roy RR. Robotic training and spinal cord plasticity. Brain Res Bull. 2009;78(1):4–12. doi:10.1016/j.brainresbull.2008.09.018
48. Hofstoetter US, Krenn M, Danner SM, et al. Augmentation of Voluntary Locomotor Activity by Transcutaneous Spinal Cord Stimulation in Motor-Incomplete Spinal Cord-Injured Individuals. Artif Organs. 2015;39(10):E176–186. doi:10.1111/aor.12615
49. Hornby TG, Reisman DS, Ward IG, et al. Clinical Practice Guideline to Improve Locomotor Function Following Chronic Stroke, Incomplete Spinal Cord Injury, and Brain Injury. JNPT. 2020;44(1):49–100.
50. Gorgey AS, Wade R, Sumrell R, Villadelgado L, Khalil RE, Lavis T. Exoskeleton Training May Improve Level of Physical Activity After Spinal Cord Injury: a Case Series. Topics Spinal Cord Injury Rehab. 2017;23(3):245–255. doi:10.1310/sci16-00025
51. Cheung EYY, TKW Ng, KK Kelvin, Kwan RLC, Cheing GLY. Robot-Assisted Training for People With Spinal Cord Injury: a Meta-Analysis. Arch Phys Med Rehabil. 2017;98(11):2320–2331.e2312. doi:10.1016/j.apmr.2017.05.015
52. Holanda LJ, Silva PMM, Amorim TC, Lacerda MO, Simão CR, Morya E. Robotic assisted gait as a tool for rehabilitation of individuals with spinal cord injury: a systematic review. J Neuroeng Rehabil. 2017;14(1):126. doi:10.1186/s12984-017-0338-7
53. Nam KY, Kim HJ, Kwon BS, Park JW, Lee HJ, Yoo A. Robot-assisted gait training (Lokomat) improves walking function and activity in people with spinal cord injury: a systematic review. J Neuroeng Rehabil. 2017;14(1):24. doi:10.1186/s12984-017-0232-3
54. Chisholm AE, Malik RN, Blouin JS, Borisoff J, Forwell S, Lam T. Feasibility of sensory tongue stimulation combined with task-specific therapy in people with spinal cord injury: a case study. J Neuroeng Rehabil. 2014;11(1):96. doi:10.1186/1743-0003-11-96
55. Mach N, Fuster-Botella D. Endurance exercise and gut microbiota: a review. J Sport Health Sci. 2017;6(2):179–197. doi:10.1016/j.jshs.2016.05.001
56. Campbell JP, Turner JE. Debunking the Myth of Exercise-Induced Immune Suppression: redefining the Impact of Exercise on Immunological Health Across the Lifespan. Front Immunol. 2018;9:648. doi:10.3389/fimmu.2018.00648
57. Pedersen BK. Physical activity and muscle-brain crosstalk. Nat Rev Endocrinol. 2019;15(7):383–392. doi:10.1038/s41574-019-0174-x
58. Saint-Georges-Chaumet Y, Edeas M. Microbiota-mitochondria inter-talk: consequence for microbiota-host interaction. Pathog Dis. 2016;74(1):ftv096. doi:10.1093/femspd/ftv096
59. West CR, Crawford MA, Laher I, Ramer MS, Krassioukov AV. Passive Hind-Limb Cycling Reduces the Severity of Autonomic Dysreflexia After Experimental Spinal Cord Injury. Neurorehabil Neural Repair. 2016;30(4):317–327. doi:10.1177/1545968315593807
60. Weinhold M, Shimabukuro-Vornhagen A, Franke A, et al. Physical exercise modulates the homeostasis of human regulatory T cells. J Allergy Clin Immunol. 2016;137(5):1607–1610.e1608. doi:10.1016/j.jaci.2015.10.035
61. Tilg H, Zmora N, Adolph TE, Elinav E. The intestinal microbiota fuelling metabolic inflammation. Nat Rev Immunol. 2020;20(1):40–54. doi:10.1038/s41577-019-0198-4
62. Kressler J, Cowan RE, Bigford GE, Nash MS. Reducing cardiometabolic disease in spinal cord injury. Phys Med Rehab Clin North Am. 2014;25(3):573–604. doi:10.1016/j.pmr.2014.04.006
63. Astorino TA, Harness ET, Witzke KA. Effect of chronic activity-based therapy on bone mineral density and bone turnover in persons with spinal cord injury. Eur J Appl Physiol. 2013;113(12):3027–3037. doi:10.1007/s00421-013-2738-0
64. Hazell TJ, Islam H, Townsend LK, Schmale MS, Copeland JL. Effects of exercise intensity on plasma concentrations of appetite-regulating hormones: potential mechanisms. Appetite. 2016;98:80–88. doi:10.1016/j.appet.2015.12.016
65. Hoyles L, Snelling T, Umlai UK, et al. Microbiome-host systems interactions: protective effects of propionate upon the blood-brain barrier. Microbiome. 2018;6(1):55. doi:10.1186/s40168-018-0439-y
66. Li JM, Yu R, Zhang LP, et al. Dietary fructose-induced gut dysbiosis promotes mouse hippocampal neuroinflammation: a benefit of short-chain fatty acids. Microbiome. 2019;7(1):98. doi:10.1186/s40168-019-0713-7
67. Kratsman N, Getselter D, Elliott E. Sodium butyrate attenuates social behavior deficits and modifies the transcription of inhibitory/excitatory genes in the frontal cortex of an autism model. Neuropharmacology. 2016;102:136–145. doi:10.1016/j.neuropharm.2015.11.003
68. Chen T, Noto D, Hoshino Y, Mizuno M, Miyake S. Butyrate suppresses demyelination and enhances remyelination. J Neuroinflammation. 2019;16(1):165. doi:10.1186/s12974-019-1552-y
69. Bravo JA, Forsythe P, Chew MV, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA. 2011;108(38):16050–16055. doi:10.1073/pnas.1102999108
70. Jaglin M, Rhimi M, Philippe C, et al. Indole, a Signaling Molecule Produced by the Gut Microbiota, Negatively Impacts Emotional Behaviors in Rats. Front Neurosci. 2018;12:216. doi:10.3389/fnins.2018.00216
71. Braniste V, Al-Asmakh M, Kowal C, et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Trans Med. 2014;6(263):263ra158. doi:10.1126/scitranslmed.3009759
72. O’Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res. 2015;277:32–48. doi:10.1016/j.bbr.2014.07.027
73. Noakes M, Keogh JB, Foster PR, Clifton PM. Effect of an energy-restricted, high-protein, low-fat diet relative to a conventional high-carbohydrate, low-fat diet on weight loss, body composition, nutritional status, and markers of cardiovascular health in obese women. Am J Clin Nutr. 2005;81(6):1298–1306. doi:10.1093/ajcn/81.6.1298
74. Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci. 2017;20(2):145–155. doi:10.1038/nn.4476
75. Heiman ML, Greenway FL. A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol Metabol. 2016;5(5):317–320. doi:10.1016/j.molmet.2016.02.005
76. Erny D, Hrabě de Angelis AL, Jaitin D, et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. 2015;18(7):965–977. doi:10.1038/nn.4030
77. Foster JA, Rinaman L, Cryan JF. Stress & the gut-brain axis: regulation by the microbiome. Neurobiol Stress. 2017;7:124–136. doi:10.1016/j.ynstr.2017.03.001
78. Carlson AL, Xia K, Azcarate-Peril MA, et al. Infant Gut Microbiome Associated With Cognitive Development. Biol Psychiatry. 2018;83(2):148–159. doi:10.1016/j.biopsych.2017.06.021
79. Ghosh TS, Rampelli S, Jeffery IB, et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: the NU-AGE 1-year dietary intervention across five European countries. Gut. 2020;69(7):1218–1228. doi:10.1136/gutjnl-2019-319654
80. Russo MA, Santarelli DM, O’Rourke D. The physiological effects of slow breathing in the healthy human. Breathe. 2017;13(4):298–309. doi:10.1183/20734735.009817
81. Guiraud D, Andreu D, Bonnet S, et al. Vagus nerve stimulation: state of the art of stimulation and recording strategies to address autonomic function neuromodulation. J Neural Eng. 2016;13(4):041002. doi:10.1088/1741-2560/13/4/041002
82. Tang YY, Hölzel BK, Posner MI. The neuroscience of mindfulness meditation. Nat Rev Neurosci. 2015;16(4):213–225. doi:10.1038/nrn3916
83. Craig A, Guest R, Tran Y, Middleton J. Cognitive Impairment and Mood States after Spinal Cord Injury. J Neurotrauma. 2017;34(6):1156–1163. doi:10.1089/neu.2016.4632
84. Allaband C, McDonald D, Vázquez-Baeza Y, et al. Microbiome 101: studying, Analyzing, and Interpreting Gut Microbiome Data for Clinicians. Clin Gastroenterol Hepatol. 2019;17(2):218–230. doi:10.1016/j.cgh.2018.09.017
85. Mateo S, Di Rienzo F, Bergeron V, Guillot A, Collet C, Rode G. Motor imagery reinforces brain compensation of reach-to-grasp movement after cervical spinal cord injury. Front Behav Neurosci. 2015;9:234. doi:10.3389/fnbeh.2015.00234
86. Morawietz C, Moffat F. Effects of locomotor training after incomplete spinal cord injury: a systematic review. Arch Phys Med Rehabil. 2013;94(11):2297–2308. doi:10.1016/j.apmr.2013.06.023
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.