Back to Journals » OncoTargets and Therapy » Volume 13
Exon Coverage Variations Between Cancer Tissues and Adjacent Non-Cancerous Tissues are Prognostic Factors in Gastric Cancer
Authors Pan X, Wang Y, Li C, Zhou Z, Zhong Y, Feng J, Lu J ![]()
Received 11 October 2019
Accepted for publication 2 December 2019
Published 7 January 2020 Volume 2020:13 Pages 61—70
DOI https://doi.org/10.2147/OTT.S234351
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
Xuan Pan,* Yajing Wang,* Chenchen Li,* Zhaofei Zhou, Yuejiao Zhong, Jifeng Feng, Jianwei Lu
Department of Medical Oncology, The Affiliated Cancer Hospital of Nanjing Medical University, Jiangsu Cancer Hospital, Jiangsu Institute of Cancer Research, Nanjing, Jiangsu 210009, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jifeng Feng; Jianwei Lu
Department of Medical Oncology, The Affiliated Cancer Hospital of Nanjing Medical University, Jiangsu Cancer Hospital, Jiangsu Institute of Cancer Research, 42 Baiziting Road, Nanjing, Jiangsu 210009, People’s Republic of China
Email [email protected]; [email protected]
Introduction: Gastric cancer is highly heterogeneous both clinically and pathologically and is one of the leading causes of cancer-related deaths worldwide. Genomic coverage variations, also known as copy number variations (CNVs), play a critical role in the carcinogenesis of gastric cancer. Many studies have demonstrated that DNA CNVs are important factors affecting the expression of protein-encoding genes in the gastric cancer genome.
Methods: Thirty gastric cancer patients from a Chinese population were enrolled. Genomic DNA was extracted from gastric cancer tissue and matched adjacent non-cancerous tissue from each patient. A panel of 1,021 genes including 3300 exons was designed and subjected to next-generation sequencing. Copy numbers of each gene and exon were calculated for each tissue. Coverage variations between gastric cancer tissue and matched adjacent non-cancerous tissue were also calculated, and we examined the correlation between overall survival of patients and coverage variation type for each exon.
Results: DNA from cancerous tissue and corresponding adjacent non-cancerous tissue were significantly different with respect to the pattern of gene copy number. Exon copy numbers were highly consistent among non-cancerous samples and confirmed that non-cancerous tissue contain diploid genomes. In contrast, the gene coverage pattern among cancerous tissue showed significant differences and confirmed that gastric cancer is a genetically heterogeneous disease. Numerous exon coverage variations were identified in gastric cancer tissue compared with matched, adjacent non-cancerous tissue. Overall survival between patients with and without coverage variations in regions of NOTCH2, NTRK3, ERBB2 and RERE exons exhibited significant differences. This is consistent with previous reports and indicates that these findings may have prognostic value.
Conclusion: Our results confirm that gastric cancer is a genetically heterogeneous disease. Exon coverage variations between cancer tissue and their adjacent non-cancerous tissue were shown to be associated with prognosis in gastric cancer.
Keywords: gastric cancer, next-generation sequencing, copy number variations, overall survival
Introduction
Gastric cancer is the fifth most common cancer and the third leading cause of cancer death worldwide.1 It is estimated that 27,510 new cases of gastric cancer will be diagnosed in 2019, accounting for approximately 15% of all malignancies.1 According to statistical data, 52% of the gastric cancer patients in the world reside in China.2 Despite improvements in treatment such as surgery, chemotherapy and radiotherapy, the prognosis for advanced gastric cancer remains dismal. The high mortality rate is primarily due to advanced stage diagnosis, tumor metastasis and the lack of effective drugs. Therefore, it is important to study the molecular basis of oncogenesis, development and metastasis of gastric cancer, in order to identify more sensitive detection methods, and effective therapeutic agents.
Gastric cancer is highly heterogeneous in both clinically and pathologically. As a complicated multistep process, the development of gastric cancer involves multiple signaling pathways with epigenetic and genetic variations,3–5 including copy number variations (CNVs), single-nucleotide variants (SNVs) and chromosomal translocations. These genetic alternations may cause the activation of pro-oncogenes and inactivation of tumor-suppressor genes, eventually leading to gastric tumorigenesis and progression.6 As a form of structural variation, the coverage variations of DNA sequences, genes and exons, which are known as copy number variations, are considered to be an important source of genetic diversity.7 It has been well recognized that CNV contributes to carcinogenesis in many human malignancies including gastric cancer, colorectal carcinoma, hepatocellular carcinoma and ovarian cancer.8–11 In the ToGA study, a randomized Phase III clinical trial, overall survival of patients with HER2 gene amplification was improved by trastuzumab combined with traditional chemotherapy in advanced gastric cancer.12 The results indicated that in gastric cancer, coverage variations may lead to the identification of therapeutic targets and improve diagnosis and treatment.13,14 Furthermore, the amplification of the PIK3CA, MET, MYC, and HER2 genes has also been confirmed to be associated with poor clinical outcome.8
A recent work based on 183 samples revealed that most gastric tumors carry one or more high-level CNVs encompassing known or putative therapeutic target genes, including HER2, TUBB3 and TOP2A.15 A large number of studies have found that loss of 1p36 and 9p21, as well as a gain of 3p22, 4q25, 8q24, 11p13 and 20q13 in gastric cancer. These regions are where many cancer-related genes are located, such as MYC and CTNNB1 are located.16,17 Gene copy number variations are important factors affecting gene expression, resulting in deregulation of various carcinogenic or anticancer pathways. Increasing evidence has also shown that the aberrant expression of miRNAs and lncRNAs are associated with genome coverage variations in gastric cancer.18,19 Since non-coding RNAs are key factor in carcinogenesis, further studies are warranted to evaluate the effect of CNV-associated non-coding RNAs in the diagnosis and prognosis of gastric cancer.
Array-based technologies for coverage variation detection have been used widely for more than a decade.20 However, the limitation of poor sensitivity and precision, low coverage and probe density restrict its application. In recent years, next-generation sequencing technologies21 have provided an efficient method to accurately detect coverage variations in whole genome with high resolution.22–24
In this study, we assessed the copy number of genes and exons from gastric cancer samples by using next-generation sequencing technology. Thirty gastric cancer patients in a Chinesec population were enrolled in this study. Genomic DNA was extracted from gastric cancer tissue along with matched adjacent non-cancerous tissue. A panel embracing 3,300 exons in 1,021 genes was designed and used to improve the sequencing effect. The gene copy number pattern was significantly different between cancer and paired non-cancerous tissues. Also, gene amplification and deletion were identified in cancerous tissue compared with their matched non-cancerous tissue. Additionally, we determined the prognostic value of NOTCH2, NTRK3, ERBB2, and RERE CNVs on overall survival (OS), indicating a significant role for CNVs in such a clinical application.
Materials and Methods
Ethics Statement
This study was approved by the Institutional Review Board of Jiangsu Cancer Hospital. Written informed consent from all patients was obtained.
Gastric Cancer Samples
Thirty patients from the Department of General Surgery, Jiangsu Cancer Hospital. who were diagnosed with gastric cancer were enrolled in 2013. Tumor tissue and paired adjacent non-cancerous tissue from each patient were collected. All histopathological diagnoses were reviewed independently by at least two senior pathologists.
DNA Extraction
Surgically excised human gastric cancer tissues were fixed at room temperature for 24 hrs with 10% formalin. Genomic DNA was isolated from fixed tumor tissue and matched non-cancerous tissue using a commercial DNA extraction kit (QIAamp FFPE Tissue kit, Qiagen, Hilden, Germany). DNA concentration was measured by Qubit fluorometer (ThermoFisher, Waltham, MA, USA) using Qubit dsDNA HS assay. DNA integrity was assessed on a 1% agarose gel and run along with a DL15000 DNA marker (ThermoFisher, Waltham, MA, USA). Approximately 200 ng of extracted DNA from each sample was loaded and samples containing DNA longer than 15 kbp passed the test.
Target Capture Sequencing
A panel embracing 3,300 exons of 1,017 genes was designed and used to capture target DNAs prior to sequencing. All of these genes are associated with gastric cancer and other cancer types, which include genes in the COSMIC database (http://cancer.sanger.ac.uk/cosmic), have been demonstrated to be mutated in gastric cancer, and reported to be oncogenes or tumor suppressor genes.25,26 Genes in the TCGA database (https://cancergenome.nih.gov/) which associated with other cancer types are also included. Biotinylated oligo probes (Roche NimbleGen, Inc., Madison, WI, USA) were custom designed and synthesized to capture all 3,300 exons and covered approximately 1.1 Mbp.
DNA libraries were constructed using the Illumina TruSeq DNA Library kit (Illumina, San Diego, LA, USA) according to the recommended protocol. For each sample, 1 µg of isolated DNA was initially sheared to approximately 250 bp using an ultrasonoscope. DNA fragments were then end-repaired, ligated to adapters, and amplified for 12 cycles in a thermal cycler. Customized probes were then used to capture the amplified libraries. Libraries were subsequently sequenced on a HiSeq3000 sequencing system (Illumina) in 2×150 bp paired-end mode.
Sequencing Data Analysis
To remove low-quality reads and reads containing adaptor sequences, a quality control was performed on the raw data. Using BWA (0.7.12-r1039),27 filtered reads were aligned to the human genome (GRCh37).
For each cancerous tissue and its matched adjacent non-cancerous tissue, read counts for each base was calculated using Samtools.28 The average coverage depth of each captured exon was then calculated and normalized to a same standard, two in each sample. We filtered the exons whose average coverage depth fell below 0.1. The heatmaps of all cancer tissues and all adjacent non-cancerous tissues were constructed using the R package Pheatmap, where red and blue indicates gain and loss, respectively. Cancer tissue and all adjacent non-cancerous tissue were analyzed by cluster analysis based on their exon coverage pattern. Next, the coverage depth ratio for each exon between cancer and paired normal samples was determined. A heatmap was generated based on the depth ratio by using the R package Pheatmap. The red and blue colors indicate exon gain and loss in the cancer tissue, respectively.
Overall survival (OS) curves were generated by using the R package survival based on Kaplan-Meier method. By using the log rank test, differences of P<0.05 were considered to be statistically significant. Patients were divided into three groups (Gain, Normal and Loss) by average coverage and based on the number of samples, the smallest group was merged with the adjacent group.
Results
Clinical Characteristics
The clinical characteristics of the patients in this study are listed in Table 1. Briefly, the enrolled gastric cancer patients consisted of 27 males and 3 females, and their age at diagnosis/surgery ranged from 32~79 years with a median age of 58.5 years. The follow-up time ranged from 6–50 months with a median time of 40 months. Gastric cancer tissues were obtained from three anatomical locations: 11 from the cardia, 12 from the body and 7 from the antrum. Based on the Lauren classification, 16 cases were classified as intestinal-type, 11 as diffuse-type, and 3 as mixed intestinal with diffuse histology. Based on the 7th National Comprehensive Cancer Network (NCCN) treatment guidelines, 2 samples were determined to be stage I, 4 samples were determined to be stage II, and the remaining 24 samples were determined to be stage III.
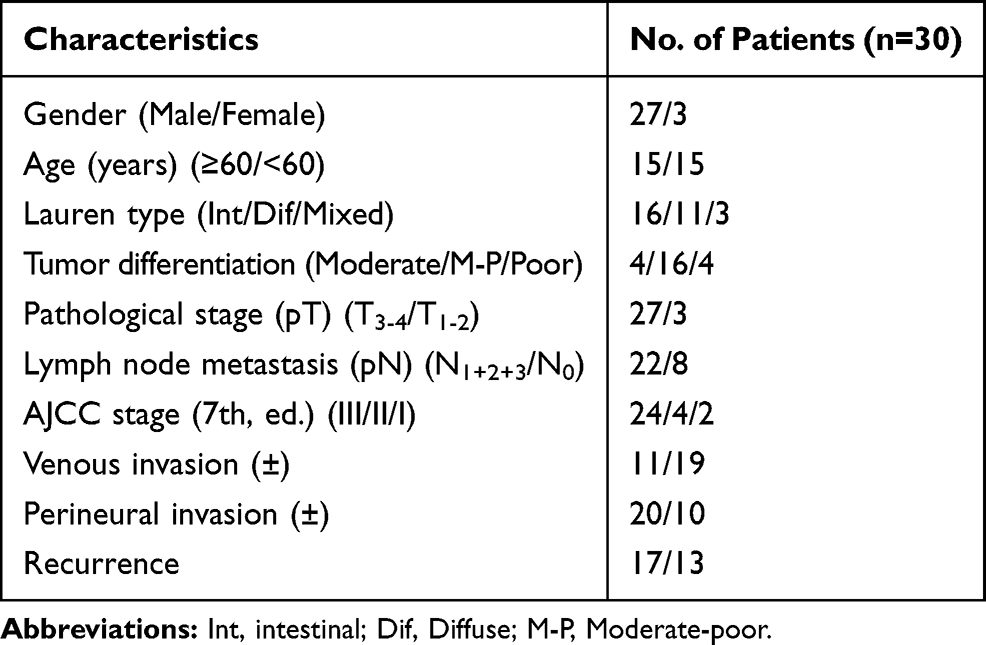 |
Table 1 Patients Clinicopathological Characteristics |
Capture and Sequencing Results
To improve the sequencing effort, a panel of 1,021 genes including 3,300 exons was investigated. A total of 30 cancerous and matched adjacent non-cancerous tissues were sequenced. The sequencing coverage depth of the cancer tissue ranged from 119.2-fold to 1303.4-fold, with the a median depth of 751.2-fold. The sequencing coverage depth of the adjacent non-cancerous tissue was lower than that of the cancer tissues with a median depth of 96.5-fold.
Overall Coverage Depth of Exons
Exon coverage depth was calculated and normalized. We filtered the exons with a coverage depth less than 0.1. The exon coverage pattern of non-cancerous tissues exhibited a high similarity among samples (Figure 1A). Because all the samples in this group represent normal tissue, they tend to have the same normal diploid genomes. The coverage variations among exons may be the result of different efficiencies of the capture probes and amplification bias. In contrast, the exon coverage pattern of the cancer tissue showed significant difference among samples (Figure 1B). Numerous incidents of copy number gain and loss, which consist of genes/exons involved in pathogenesis, progression, and drug resistance were observed in the cancerous samples compared with adjacent non-cancerous samples. The copy number of these genes/exons was approximately the same in the non-cancerous tissue. It has been demonstrated that gastric cancers are highly heterogeneous with respect to their genome. Different cancer tissues contain different genomic alterations, including single nucleotide variations, insertion, deletion and copy number variations. Even within one cancer tissue, the genomes among cells are considered to be heterogeneous. Therefore, the copy number patterns of normal and cancerous tissue demonstrate that the sequencing and analysis approaches are efficient and the results could provide sample genomic information.
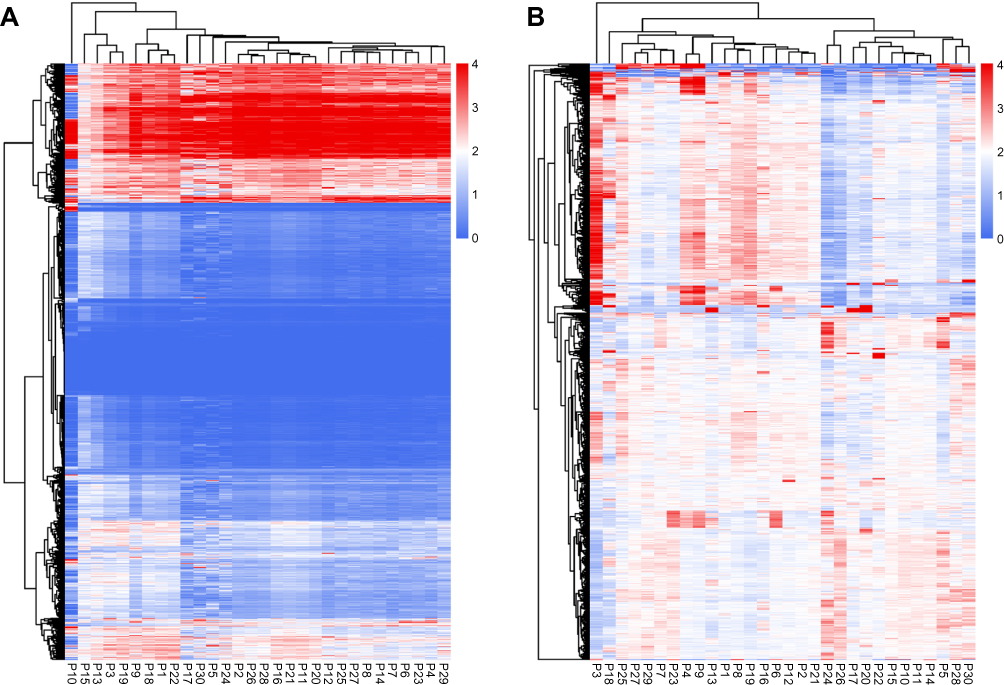 |
Figure 1 Heatmap of exon coverage. (A) Heatmap of exon coverage in adjacent non-cancerous normal tissues. (B) Heatmap of exon coverage in cancerous tissue. |
Based on the copy number pattern, we divided gastric cancer patients into two groups and one separate sample (P3). To investigate whether there was any difference between the two divided groups, we examined the clinical characteristics and overall survival difference. The clinicopathological characteristics of the two groups are shown in Table 2. The median age of group one was seven years younger than group two (56 vs 63). Meanwhile, based on Lauren classification, group one had a higher proportion of intestinal type disease compared with group two. More patients in group one had perineural invasion. With respect of overall survival, we found that there was a slight difference between the two groups (Figure 2), however, this was not significantly different.
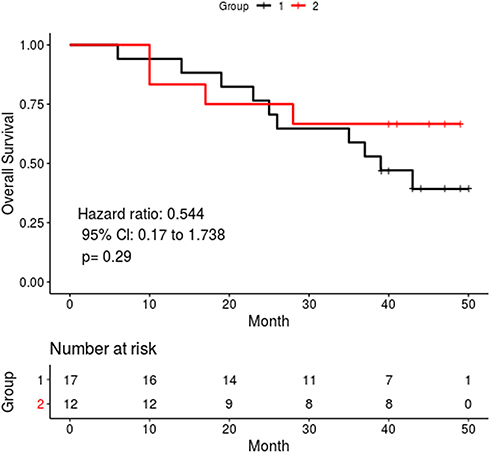 |
Figure 2 Overall survival curves of the two groups based on total exon coverage pattern. |
 |
Table 2 Patient Clinicopathological Characteristics of Group 1 and Group 2 |
Coverage Variations of Cancer Tissues Function as Prognostic Factor
To avoid the influence caused by different efficiencies of the capture probes and amplification bais, the coverage variation of cancerous tissue compared with adjacent non-cancerous normal tissue was investigated. The coverage depth ratio for each exon is shown in Figure 3. The exons exhibited different coverage ratios among the samples.
 |
Figure 3 Heatmap depicting the coverage ratio between cancer samples and their adjacent normal tissues. |
The relationship between coverage variations of the cancer tissue and prognosis of gastric cancer patients was further analyzed. We screened all of the 3300 exons which were designed to be captured and evaluated their potential as prognostic markers. Patients with a gain of the RERE_18 exon exhibited significantly decreased death hazard (Figure 4) compared with those without a gain (HR = 0.304, 95% CI = 0.105–0.885, P = 0.022).
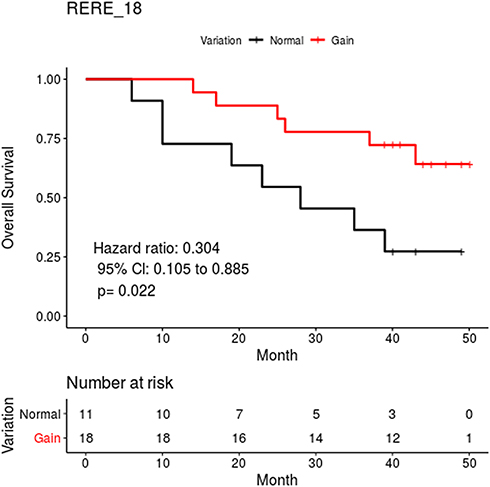 |
Figure 4 Overall survival curves of the two groups based on total RERE_18 coverage variations. |
Several NOTCH2 exons showed a correlation with overall survival. For example, patients with NOTCH2_13, NOTCH2_17, NOTCH2_19, NOTCH2_23, or NOTCH2_26 exon gains had a shorter overall survival than those without (Figure 5). However, a significant difference was only observed between the group with NOTCH2_13 exon gain and the normal group (HR=2.901, 95% CI = 0.997–8.446, P = 0.043).
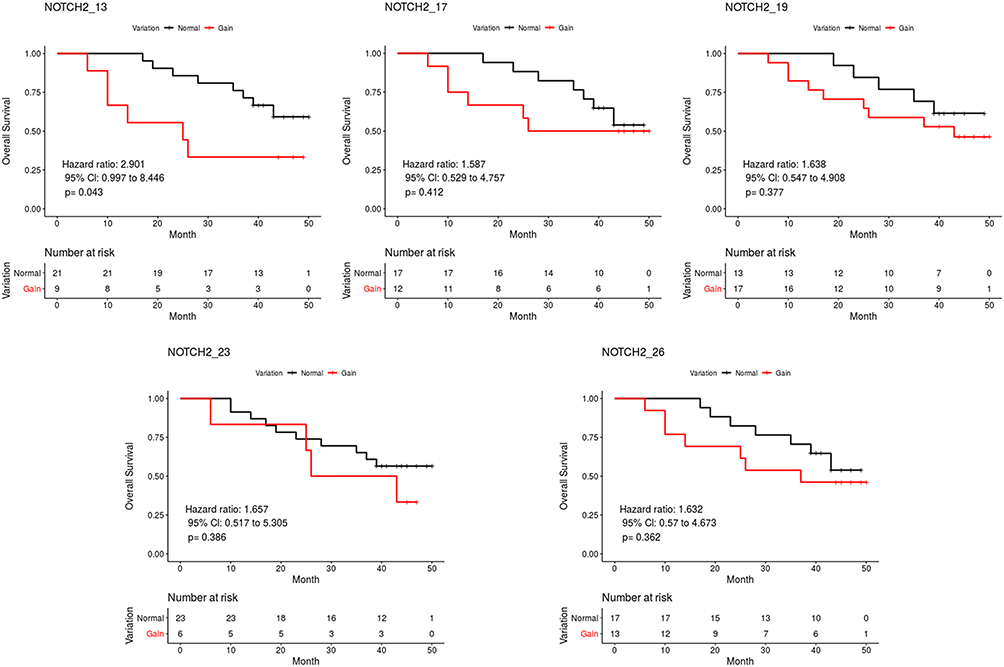 |
Figure 5 Overall survival curves of the two groups based on total coverage variations of NOTCH2. |
For NTRK3, patients with NTRK3_3, NTRK3_16, or NTRK3_19 exon gains had a shorter overall survival than those without a gain, while patients showing a loss of the NTRK3_4 or NTRK3_7 exons exhibited a longer overall survival than those without loss (Figure 6). No significant difference was found between the group with the gain of the NTRK3_19 exon (P = 0.104), loss of the NTRK3_4 exon (P = 0.112) and the normal group, respectively. The overall survival analysis also demonstrated an increased death hazard in the group with a gain of ERBB2_8,13,14,15,19,31 exons, and a decreased death hazards in the group showing loss of ERBB2_18,20,21,24,26 exons compared with the normal group. However, there was no statistically significant difference. Overall, these results demonstrated the prognostic value of coverage variations in gastric cancer.
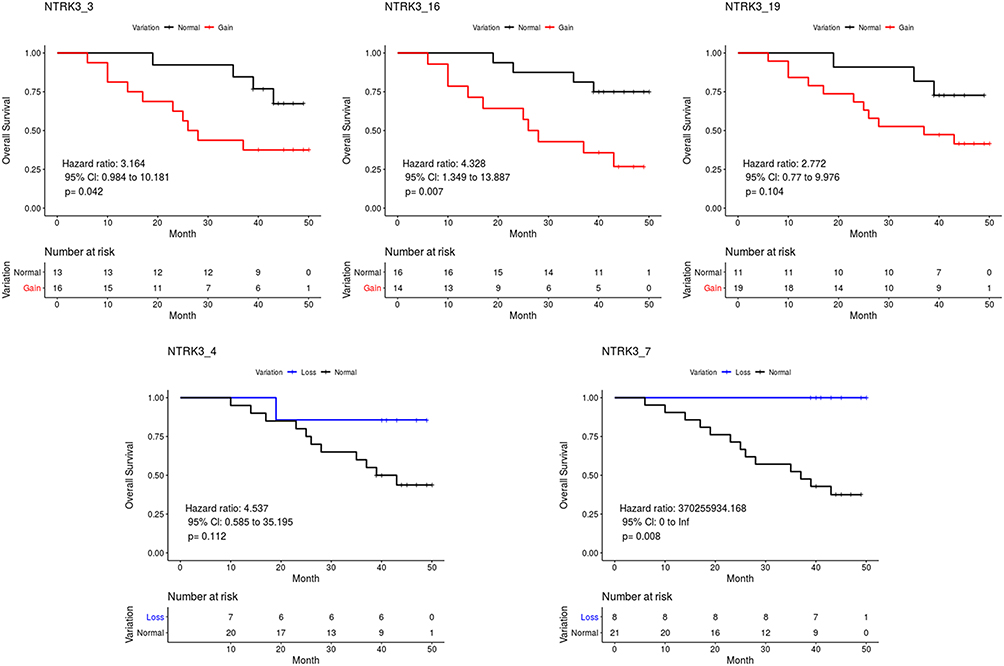 |
Figure 6 Overall survival curves of the two groups based on total coverage variations of NTRK3. |
Discussion
Gastric cancers represent a highly heterogeneous group of diseases among individuals and even among individual cells in one patient with different molecular and genetic characteristic. Despite advances in chemoradiotherapy and targeted therapy, the prognosis for gastric cancer remains poor. Genetic alterations, such as copy number variations, single nucleotide polymorphisms, and epigenetic alterations influence the expression of genes involved in different signaling pathways. This eventually leads to the initiation and development of gastric cancer.6 Gastric cancer may be divided into subtypes with different prognoses and recurrence rates according to their genetic characteristics. These approaches provide an opportunity to understand the molecular mechanisms of gastric cancer, which is urgently needed. As an important type of somatic mutation, copy number variation has been demonstrated to be critical in tumorigenesis and cancer prognosis.29–31 Copy number amplification often contributes to oncogene activation, while copy number deletion often contributes to tumor suppressor gene inactivation.
In the present study, we focused on gene copy number variations in 30 gastric cancer samples from a Chinese population using NGS technology. We evaluated the copy number of specific genes and exons in cancer tissue and adjacent normal tissue. Among non-cancerous tissue, the exon copy number was highly consistent among samples (Figure 1A). Meanwhile, different exons exhibited different coverage depth within one normal sample (Figure 1A). Normal tissues contain similar diploid genomes and therefore tend to generate similar copy number patterns among samples. However, different probes used in the sequencing library capture have different efficiencies, and this causes coverage differences among exons. In contrast, the exon coverage pattern among cancer tissues showed a significant difference (Figure 1B). As a genetically heterogeneous disease, gastric cancer tissue from different patients vary in gene and exon copy number. Therefore, a different coverage pattern was observed among cancer tissue.
To reveal the copy number variations from normal versus cancerous tissue, and to eliminate the influence caused by capture probes, we calculated the coverage depth ratios for specific exons. As shown in Figure 3, exons exhibited different coverage ratios within a sample and among samples. Further, we also analyzed the relationship between coverage variations of cancer tissue and the prognosis of gastric cancer patients by screening 3,300 exons in 1,021 genes. The overall survival of patients with a copy number gain for the NOTCH2_13, NTRK3_16 and NTRK3_3 exons was significantly shorter than those without exon gain. Meanwhile, the copy number gain of the RERE_18 exon exhibited a significantly decreased death hazard compared with those without exon gain. RERE is localized to PODs and colocalizes with the proapoptotic proteins PML and BAX.32 It functions as a tumor suppressor gene in neuroblastoma and colorectal carcinoma.33,34 Overexpression of RERE induces cell apoptosis in a caspase-dependent manner.32 The NOTCH signaling pathway regulates cancer stem cells, tumor growth, invasion and metastasis.35 In mammals, there are four NOTCH receptors (NOTCH 1–4) and five NOTCH ligands.36 The function of NOTCH2 signaling is cellular context-dependent and it may act ad both an oncogene and a tumor suppressor gene.37–39 Overexpression or gain-of-function mutations of NOTCH2 has been found in many cancer types. The expression of NOTCH2 in precancerous mucosal lesions has been reported to be significantly higher than in adjacent normal tissues, suggesting that NOTCH2 plays a vital role in the oncogenesis of gastric cancer.40 Moreover, another study has reported that gastric cancer patients with elevated NOTCH2 expression exhibited poor prognosis.41 Immunohistochemistry staining of gastric cancer specimens before and after undergoing chemotherapy has highlighted an important role for NOTCH2 in drug resistance. A significant increase in NOTCH2 expression was found in patients that exhibited disease regression following chemotherapy. This result has the potential to guide postoperative chemotherapy after neoadjuvant chemotherapy.42 In our study, patients with a gain of the NOTCH2_13 exon were found to have a significantly shorter overall survival, which is consistent with the reports described above.
Chromosomal rearrangements involving the NTRK family have been demonstrated to be oncogenic drivers in many tumor types.43 This may be due to functional gene fusions occurring during the chromosomal rearrangements. The existence of an NTRK fusion gene may predict sensitivity to selective TRK kinase inhibitors, such as Larotrectinib. NTRK3 expression was detected in the gastrointestinal epithelia,44 suggesting a role in gastrointestinal cells development. Takashi Kubo et al45 undertook a systematic screening of mutations in gastric cancer cell lines and primary gastric cancer samples with poorly differentiated histology. They found recurrent non-synonymous mutations in the NTRK3 gene. Coverage variations of NTRK3 could predict prognosis and chemosensitivity to platinum drugs in ovarian cancer.46 Moreover, NTRK3 has been reported to be a direct target of miR-497 in gastric cancer cell lines. In our study, we found that copy number gains of the NTRK3_16 and NTRK3_3 exons predict a poor prognosis in gastric cancer patients, while patients with an NTRK3_7 exon copy number loss had a longer overall survival. This is consistent with the role of NTRK3 described in previous studies.
Our results confirm that gastric cancer is a genetically heterogeneous disease. Coverage variations of some exons of the RERE, NOTCH2, and NTRK3 genes between cancer tissue and adjacent non-cancerous tissue were shown to be prognostic factors in gastric cancer. Further studies using a a larger sample size and longer follow-up times will be required to discover the roles of gene coverage variations in gastric cancer.
Acknowledgements
This work was funded by Jiangsu Provincial Medical Youth Talent QNRC2016645, Jiangsu Provincial Top Talent Scientific Research Project of “Six One Projects” for High Level Health Talents LGY2019076, and the Young Talents Program of Jiangsu Cancer Hospital QL201814. We thank Edanz Group for editing a draft of this manuscript.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
2. Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–E386. doi:10.1002/ijc.29210
3. Takeno SS, Leal MF, Lisboa LC, et al. Genomic alterations in diffuse-type gastric cancer as shown by high-resolution comparative genomic hybridization. Cancer Genet Cytogenet. 2009;190(1):1–7. doi:10.1016/j.cancergencyto.2008.09.007
4. Burbano RR, Assumpcao PP, Leal MF, et al. C-MYC locus amplification as metastasis predictor in intestinal-type gastric adenocarcinomas: CGH study in Brazil. Anticancer Res. 2006;26(4B):2909–2914.
5. Gigek CO, Chen ES, Calcagno DQ, Wisnieski F, Burbano RR, Smith MA. Epigenetic mechanisms in gastric cancer. Epigenomics. 2012;4(3):279–294. doi:10.2217/epi.12.22
6. Hudler P. Genetic aspects of gastric cancer instability. Sci World J. 2012;2012:761909. doi:10.1100/2012/761909
7. Nakamura Y. DNA variations in human and medical genetics: 25 years of my experience. J Hum Genet. 2009;54(1):1–8. doi:10.1038/jhg.2008.6
8. Liang L, Fang JY, Xu J. Gastric cancer and gene copy number variation: emerging cancer drivers for targeted therapy. Oncogene. 2016;35(12):1475–1482. doi:10.1038/onc.2015.209
9. Horpaopan S, Spier I, Zink AM, et al. Genome-wide CNV analysis in 221 unrelated patients and targeted high-throughput sequencing reveal novel causative candidate genes for colorectal adenomatous polyposis. Int J Cancer. 2015;136(6):E578–E589. doi:10.1002/ijc.29215
10. Despierre E, Moisse M, Yesilyurt B, et al. Somatic copy number alterations predict response to platinum therapy in epithelial ovarian cancer. Gynecol Oncol. 2014;135(3):415–422. doi:10.1016/j.ygyno.2014.09.014
11. Xu H, Zhu X, Xu Z, et al. Non-invasive analysis of genomic copy number variation in patients with hepatocellular carcinoma by next generation DNA sequencing. J Cancer. 2015;6(3):247–253. doi:10.7150/jca.10747
12. Bang YJ, Van Cutsem E, Feyereislova A, et al. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): a Phase 3, open-label, randomised controlled trial. Lancet. 2010;376(9742):687–697.
13. Xie L, Su X, Zhang L, et al. FGFR2 gene amplification in gastric cancer predicts sensitivity to the selective FGFR inhibitor AZD4547. Clin Cancer Res. 2013;19(9):2572–2583. doi:10.1158/1078-0432.CCR-12-3898
14. Okamoto W, Okamoto I, Arao T, et al. Antitumor action of the MET tyrosine kinase inhibitor crizotinib (PF-02341066) in gastric cancer positive for MET amplification. Mol Cancer Ther. 2012;11(7):1557–1564. doi:10.1158/1535-7163.MCT-11-0934
15. Labots M, Buffart TE, Haan JC, et al. High-level copy number gains of established and potential drug target genes in gastric cancer as a lead for treatment development and selection. Cell Oncol (Dordr). 2014;37(1):41–52. doi:10.1007/s13402-013-0162-4
16. Deng N, Goh LK, Wang H, et al. A comprehensive survey of genomic alterations in gastric cancer reveals systematic patterns of molecular exclusivity and co-occurrence among distinct therapeutic targets. Gut. 2012;61(5):673–684. doi:10.1136/gutjnl-2011-301839
17. Zhang D, Wang Z, Luo Y, et al. Analysis of DNA copy number aberrations by multiple ligation-dependent probe amplification on 50 intestinal type gastric cancers. J Surg Oncol. 2011;103(2):124–132. doi:10.1002/jso.21792
18. Hu Y, Wang J, Qian J, et al. Long noncoding RNA GAPLINC regulates CD44-dependent cell invasiveness and associates with poor prognosis of gastric cancer. Cancer Res. 2014;74(23):6890–6902. doi:10.1158/0008-5472.CAN-14-0686
19. Hu L, Wu Y, Tan D, et al. Up-regulation of long noncoding RNA MALAT1 contributes to proliferation and metastasis in esophageal squamous cell carcinoma. J Exp Clin Cancer Res. 2015;34:7. doi:10.1186/s13046-015-0123-z
20. Carter NP. Methods and strategies for analyzing copy number variation using DNA microarrays. Nat Genet. 2007;39(7 Suppl):S16–S21. doi:10.1038/ng2028
21. Metzker ML. Sequencing technologies - the next generation. Nat Rev Genet. 2010;11(1):31–46. doi:10.1038/nrg2626
22. Meyerson M, Gabriel S, Getz G. Advances in understanding cancer genomes through second-generation sequencing. Nat Rev Genet. 2010;11(10):685–696. doi:10.1038/nrg2841
23. Ley TJ, Mardis ER, Ding L, et al. DNA sequencing of a cytogenetically normal acute myeloid leukaemia genome. Nature. 2008;456(7218):66–72. doi:10.1038/nature07485
24. Ku CS, Loy EY, Salim A, Pawitan Y, Chia KS. The discovery of human genetic variations and their use as disease markers: past, present and future. J Hum Genet. 2010;55(7):403–415. doi:10.1038/jhg.2010.55
25. Chen K, Yang D, Li X, et al. Mutational landscape of gastric adenocarcinoma in Chinese: implications for prognosis and therapy. Proc Natl Acad Sci U S A. 2015;112(4):1107–1112. doi:10.1073/pnas.1422640112
26. Wang K, Yuen ST, Xu J, et al. Whole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancer. Nat Genet. 2014;46(6):573–582. doi:10.1038/ng.2983
27. Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics. 2009;25(14):1754–1760. doi:10.1093/bioinformatics/btp324
28. Li H, Handsaker B, Wysoker A, et al. The sequence alignment/map format and SAMtools. Bioinformatics. 2009;25(16):2078–2079. doi:10.1093/bioinformatics/btp352
29. Albertson DG, Collins C, McCormick F, Gray JW. Chromosome aberrations in solid tumors. Nat Genet. 2003;34(4):369–376. doi:10.1038/ng1215
30. Beroukhim R, Mermel CH, Porter D, et al. The landscape of somatic copy-number alteration across human cancers. Nature. 2010;463(7283):899–905. doi:10.1038/nature08822
31. Dancey JE, Bedard PL, Onetto N, Hudson TJ. The genetic basis for cancer treatment decisions. Cell. 2012;148(3):409–420. doi:10.1016/j.cell.2012.01.014
32. Waerner T, Gardellin P, Pfizenmaier K, Weith A, Kraut N. Human RERE is localized to nuclear promyelocytic leukemia oncogenic domains and enhances apoptosis. Cell Growth Differ. 2001;12(4):201–210.
33. Bader SA, Fasching C, Brodeur GM, Stanbridge EJ. Dissociation of suppression of tumorigenicity and differentiation in vitro effected by transfer of single human chromosomes into human neuroblastoma cells. Cell Growth Differ. 1991;2(5):245–255.
34. Tanaka K, Yanoshita R, Konishi M, et al. Suppression of tumourigenicity in human colon carcinoma cells by introduction of normal chromosome 1p36 region. Oncogene. 1993;8(8):2253–2258.
35. Tien AC, Rajan A, Bellen HJ. A Notch updated. J Cell Biol. 2009;184(5):621–629. doi:10.1083/jcb.200811141
36. Ranganathan P, Weaver KL, Capobianco AJ. Notch signalling in solid tumours: a little bit of everything but not all the time. Nat Rev Genet. 2011;11(5):338–351. doi:10.1038/nrc3035
37. Rohn JL, Lauring AS, Linenberger ML, Overbaugh J. Transduction of Notch2 in feline leukemia virus-induced thymic lymphoma. J Virol. 1996;70(11):8071–8080.
38. Fan X, Mikolaenko I, Elhassan I, et al. Notch1 and notch2 have opposite effects on embryonal brain tumor growth. Cancer Res. 2004;64(21):7787–7793. doi:10.1158/0008-5472.CAN-04-1446
39. Sriuranpong V, Borges MW, Ravi RK, et al. Notch signaling induces cell cycle arrest in small cell lung cancer cells. Cancer Res. 2001;61(7):3200–3205.
40. Sun Y, Gao X, Liu J, et al. Differential Notch1 and Notch2 expression and frequent activation of Notch signaling in gastric cancers. Arch Pathol Lab Med. 2011;135(4):451–458.
41. Wu X, Liu W, Tang D, et al. Prognostic values of four Notch receptor mRNA expression in gastric cancer. Sci Rep. 2016;6:28044. doi:10.1038/srep28044
42. Bauer L, Langer R, Becker K, et al. Expression profiling of stem cell-related genes in neoadjuvant-treated gastric cancer: a NOTCH2, GSK3B and beta-catenin gene signature predicts survival. PLoS One. 2012;7(9):e44566. doi:10.1371/journal.pone.0044566
43. Kummar S, Lassen UN. TRK inhibition: a new tumor-agnostic treatment strategy. Target Oncol. 2018;13(5):545–556. doi:10.1007/s11523-018-0590-1
44. Shibayama E, Koizumi H. Cellular localization of the Trk neurotrophin receptor family in human non-neuronal tissues. Am J Pathol. 1996;148(6):1807–1818.
45. Kubo T, Kuroda Y, Shimizu H, et al. Resequencing and copy number analysis of the human tyrosine kinase gene family in poorly differentiated gastric cancer. Carcinogenesis. 2009;30(11):1857–1864. doi:10.1093/carcin/bgp206
46. Ge L, Li N, Liu M, Xu NZ, Wang MR, Wu LY. Copy number variations of neurotrophic tyrosine receptor kinase 3 (NTRK3) may predict prognosis of ovarian cancer. Medicine. 2017;96(30):e7621. doi:10.1097/MD.0000000000007621
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.