Back to Journals » International Journal of Nanomedicine » Volume 19
Evolving Tumor Characteristics and Smart Nanodrugs for Tumor Immunotherapy
Authors Sun W ![]() , Xie S
, Xie S ![]() , Liu SF, Hu X, Xing D
, Liu SF, Hu X, Xing D
Received 15 December 2023
Accepted for publication 11 April 2024
Published 1 May 2024 Volume 2024:19 Pages 3919—3942
DOI https://doi.org/10.2147/IJN.S453265
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lijie Grace Zhang
Wenshe Sun,1– 3,* Shaowei Xie,4,* Shi Feng Liu,1,* Xiaokun Hu,1 Dongming Xing1,2
1The Affiliated Hospital of Qingdao University, Qingdao, 266071, People’s Republic of China; 2Qingdao Cancer Institute, Qingdao University, Qingdao, 266071, People’s Republic of China; 3Medical Science and Technology Innovation Center, Shandong First Medical University and Shandong Academy of Medical Sciences, Jinan, 250117, People’s Republic of China; 4Department of Ultrasound, Ren Ji Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200127, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaokun Hu; Dongming Xing, The Affiliated Hospital of Qingdao University, Qingdao, 266071, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Typical physiological characteristics of tumors, such as weak acidity, low oxygen content, and upregulation of certain enzymes in the tumor microenvironment (TME), provide survival advantages when exposed to targeted attacks by drugs and responsive nanomedicines. Consequently, cancer treatment has significantly progressed in recent years. However, the evolution and adaptation of tumor characteristics still pose many challenges for current treatment methods. Therefore, efficient and precise cancer treatments require an understanding of the heterogeneity degree of various factors in cancer cells during tumor evolution to exploit the typical TME characteristics and manage the mutation process. The highly heterogeneous tumor and infiltrating stromal cells, immune cells, and extracellular components collectively form a unique TME, which plays a crucial role in tumor malignancy, including proliferation, invasion, metastasis, and immune escape. Therefore, the development of new treatment methods that can adapt to the evolutionary characteristics of tumors has become an intense focus in current cancer treatment research. This paper explores the latest understanding of cancer evolution, focusing on how tumors use new antigens to shape their “new faces”; how immune system cells, such as cytotoxic T cells, regulatory T cells, macrophages, and natural killer cells, help tumors become “invisible”, that is, immune escape; whether the diverse cancer-associated fibroblasts provide support and coordination for tumors; and whether it is possible to attack tumors in reverse. This paper discusses the limitations of targeted therapy driven by tumor evolution factors and explores future strategies and the potential of intelligent nanomedicines, including the systematic coordination of tumor evolution factors and adaptive methods, to meet this therapeutic challenge.
Keywords: smart nanomedicine, tumor evolution, immune cells, fibroblasts
Graphical Abstract:

Introduction
Cancer development is driven by an environment that fosters cellular evolution and genetic instability.1 Key factors contributing to this instability include acidosis, hypoxia, and the presence of reactive oxygen species, which are highly selective forces that induce genetic mutations.2 Therefore, malignant cancers have emerged as dynamic evolutionary entities composed of cells adapted to survive in various microhabitats. This adaptation helps prevent the onset of drug resistance. Cytotoxic cancer therapies, such as radiotherapy, exert significant evolutionary pressure on surviving cells, accelerating their rate of mutation and evolution. Nanomedicines designed to target specific components of the tumor microenvironment (TME) have shown promise owing to their enhanced selectivity, permeability, and drug-release capabilities. These properties enable effective targeting of the TME, thereby improving therapeutic outcomes.3 TME-responsive nanomedicines can be broadly categorized into two groups, namely 1) those targeting tumor-cell specific markers such as surface receptors and kinases (such as HER2, EGFR, and VEGFR), which are crucial for tumor cell differentiation, proliferation, and invasion;4 and 2) those targeting the unique physiological and pathological characteristics of tumor tissues, including acidic pH, high osmotic pressure, and elevated redox state.5,6
The application of responsive nanodrugs in tumor therapy has significantly expanded in recent years. For example, pH-responsive nanodrugs leverage the acidic extracellular environment of tumors to precisely release drugs into the TME, thereby inhibiting tumor cell proliferation.7,8 Similarly, thermally responsive nanodrugs utilize the elevated temperatures of invasive tumor tissues for localized drug release,9 while enzyme-responsive nanodrugs exploit specific TME enzymes for targeted drug delivery.6,10
However, the effectiveness of microenvironment-responsive nanodrugs in tumor therapy is limited by the tumor cell evolution, which can lead to mutations that alter receptor expression, thereby diminishing the therapeutic effect of these nanodrugs and contributing to TME heterogeneity.11 The complexity of the TME varies not only across different types of tumors but also within the same tumor, affecting the efficacy of certain nanodrugs.12 Moreover, the presence of multiple drug-resistant proteins expressed by cells within the TME can further reduce the effectiveness of responsive nanodrugs.12–15
Distinguishing the TME Components
Distinguishing between components of the TME related to tumor cells and those related to immune cells is crucial to overcome these challenges. The TME is a complex network that includes not only tumor cells, but also a variety of immune cells, such as cancer-associated fibroblasts (CAFs) and natural killer (NK) cells, which play significant roles in tumor progression and immune escape. By understanding the distinct roles and interactions of these components within the TME, more effective nanomedicine strategies that target both tumor and immune cell dynamics can be developed (Figure 1).
 |
Figure 1 The war between tumor evolution and intelligent Nanomedicines. This article summarizes the trajectory of tumor evolution from four aspects: mutation generates new antigens, giving the tumor a “new look”; immune regulation disorder, promoting an anti-inflammatory environment, and “hiding” the tumor; CAFs provide support and coordination for the tumor, can the future awaken anti-tumor immunity and attack the tumor? The irregular changes in tumor blood vessels meet the nutritional needs of tumor evolution. Based on this, targeted intelligent nano drugs are developed to lead tumor “degeneration”. |
Recent advancements in the understanding of tumor evolution and role of the immune system within the TME have paved the way for the development of smart nanomedicines. These innovative therapies are designed to exploit the unique characteristics of the tumor and its microenvironment, offering a more targeted and effective approach to cancer treatment. Among these, nanomedicines that specifically target immune components of the TME have shown excellent potential.16 These therapies focus on the modulation of immune cells, such as T cells, NK cells, and macrophages, to enhance the natural immune response of the body to tumors.16
Targeting the Immune Landscape
Tumor-associated macrophages (TAMs) and the broader immune landscape within the TME play critical roles in cancer progression and response to therapy. TAMs, which can exhibit a spectrum of activation states, from pro-inflammatory (M1) to immunosuppressive (M2), often contribute to tumor progression and immune escape. The M2 phenotype is frequently associated with tumor promotion, supporting tumor growth through mechanisms such as angiogenesis promotion, anti-tumor immunity suppression, and facilitation of tumor cell migration and invasion. Given their significant role, targeting TAMs using smart nanodrugs to modulate their activity is a promising strategy for enhancing the efficacy of cancer immunotherapy. By shifting the balance towards a more pro-inflammatory phenotype or by inhibiting the immunosuppressive functions of TAMs, smart nanomedicines can potentially disrupt tumor-promoting interactions within the TME.17–20
Concurrently, the immune landscape within the TME, encompassing a variety of immune cells, can either suppress tumor growth or be co-opted by the tumor to promote its survival and spread. This complex interplay highlights the necessity of developing smart nanomedicines designed to disrupt tumor-promoting interactions and stimulate effective anti-tumor immune responses. For example, nanomedicines that target CAFs aim to disrupt the supportive environment they provide to tumors, thereby inhibiting tumor growth and metastasis.21 Similarly, nanomedicines that modulate the activity of T cells, NK cells, and macrophages have been designed to enhance their tumor-killing capabilities.16
Advanced Nanomaterials
Advanced nanomaterials play crucial roles in next-generation cancer therapies. These materials are engineered to respond to specific signals within the TME such as pH, temperature, and enzyme activity changes. This responsiveness enables the precise delivery of therapeutic agents to the tumor site, minimizes side effects, and improves treatment efficacy. Furthermore, these advanced nanomaterials can be designed to simultaneously target multiple components of the TME, offering a multifaceted approach for cancer therapy.12–15
Future Directions
The future of cancer treatment lies in the development of therapies that can adapt to the evolving nature of tumors (Figure 2). By leveraging the latest advancements in nanotechnology and our growing understanding of the TME, researchers are working towards creating smart nanomedicines that can outpace tumor evolution.22 These therapies not only target the tumor cells themselves but also modulate the surrounding immune environment to prevent immune escape and promote sustained anti-tumor responses. In conclusion, distinguishing between the different components of the TME and targeting them with smart nanomedicines is a promising strategy for the development of more effective cancer treatments.16,21 By focusing on the unique interactions within the TME, including those involving immune cells, researchers can develop therapies that are both adaptive and capable of overcoming the challenges posed by tumor heterogeneity and evolution.22–24
 |
Figure 2 Visual Representation of Tumor Progression and Evolution. This figure illustrates the dynamic process of tumor development from early to advanced stages, highlighting key events and interactions within the TME. Starting from the left, normal cells undergo genetic mutations, leading to the initial formation of a tumor. As the tumor grows, it exhibits increased heterogeneity due to further mutations and selective pressures from the TME. Key features depicted include the generation of new antigens giving the tumor a “new look”, immune regulation disorder allowing the tumor to evade immune surveillance, the supportive role of CAFs in tumor growth and progression, and the development of new blood vessels through tumor angiogenesis to meet the nutritional demands of the growing tumor. This comprehensive overview underscores the complexity of tumor evolution and the challenges in targeting cancer effectively. |
Neoantigen “Traps” for Tumors
Tumor neoantigens (TNAs) refer to a class of mutant proteins that appear in cancer cells, but differ from normal cells.25,26 TNAs play two roles in tumor progression. On one hand, TNAs give tumor cells a certain competitive advantage that may promote tumor development and progression.27 However, TNAs are usually immunogenic; therefore, they can trigger the immune system to attack tumors, making them important targets for tumor immunotherapy.28 Immunotherapy using TNAs can identify, capture, and destroy cancer cells, thus improving their therapeutic efficacy.29,30 Several types of nanovaccines and nanomedicines that target TNAs have been explored.
TNAs and Emerging Tumor Immunotherapy
Neoantigens are created by changes in the DNA of individual cancer cells and are important targets for cancer immunotherapy because they can activate certain immune cells, such as CD8+ T cells, to attack tumors.31–36 Neoantigens are typically identified using whole-exome sequencing and computer modeling, and various algorithm combinations are used to predict their immunogenicity.28,37 Various approaches, including the use of tumor cell fragments, nucleic acids, and synthetic peptide vaccines, have been used to create vaccines targeting neoantigens.38 Additionally, nanoplatform vaccines based on tumor-specific TMEs can provide personalized treatment while overcoming delivery barriers in vivo, realizing targeted delivery, enhancing drug efficacy, and reducing adverse drug reactions (Figure 3).
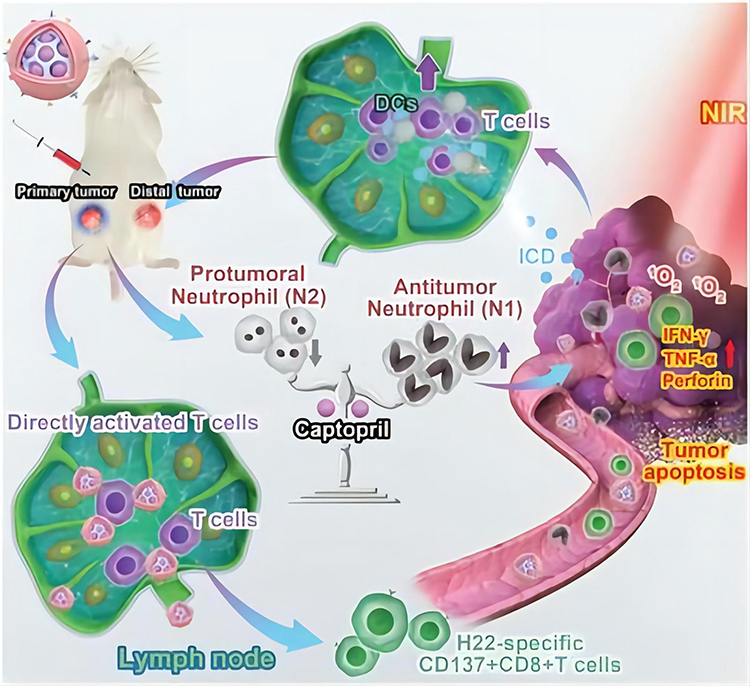 |
Figure 3 Schematic representation of the computer-based prediction of neoantigen nanovaccine efficiency. Computational analysis has been used to predict a new, specific antigen for H22 liver cancer cells, and the results were verified experimentally. As a result, a new type of nano-vaccine that could activate DC was developed. This vaccine not only targets tumor tissues through photodynamic therapy (PDT) and enhances the release of tumor-associated antigens (TAA) but also directly activates CD8+ T cells and induces lymphocyte homing. Additionally, the vaccine featured a tumor acid-triggered captopril release function that helped reduce the pre-tumor N2 phenotype and further enhanced the immune response, significantly prolonging the survival of mice carrying H22. Note: Reprinted from Wang Y, Zhao Q, Zhao B, et al. Remodeling tumor-associated neutrophils to enhance dendritic cell-based HCC neoantigen nano-vaccine efficiency. Adv Sci. 2022;9(11):e2105631. © 2022 The Authors. Advanced Science published by Wiley-VCH GmbH.39 |
Principle of New Antigen Vaccines
Individual-specific non-self proteins produced by non-synonymous mutations in tumor cells have strong immunogenicity and lack expression in healthy tissues, making them promising targets for immunotherapy against tumors.40 Forty years ago, it was hypothesized that tumor-specific antigens exist on tumor cells and can be recognized and bound by HLA molecules, activating specific T cells, and inducing anti-tumor immunity.41 However, traditional cloning methods are expensive and incapable of accurately identifying TNAs, making them unsuitable for use as tumor vaccines. Sequencing technologies for whole genomes and exomes are developing rapidly, enabling thousands of tumor-related genes to be efficiently and inexpensively identified. Current research is focusing on neoantigens that T cells can specifically recognize, which differ from traditional tumor-associated antigens (TAAs).42,43 Neoantigens, as compared to TAAs, are more immunogenic and have a stronger affinity for major histocompatibility complex (MHC) molecules, making them more resistant to central immune tolerance. Monach et al reported that TNAs can be used in cancer immunotherapy.44 Approximately 95% of tumor mutations consist of point mutations, whereas the remaining 5% consist of indels and frameshift mutations.45,46 Insertion, deletion, and frameshift mutations cause more obvious changes in the sequence and spatial organization of amino acids, and the mutated peptides have a stronger affinity for MHC molecules, making them more easily recognized by T cells as neoantigens.47 Owing to the immunogenicity of various tumors and decreased immune function of patients, approximately 1–2% of T cells spontaneously recognize endogenous neoantigens.48 Therefore, the design of multifunctional and specific vaccines based on the acquisition of efficient neoantigens is an effective strategy for tumor immunotherapy.
Multifunctional Nanovaccine Design Strategy
Multifunctional nanovaccines employ nanomaterials to transport various components, such as antigens, adjuvants, and immunomodulators, to target specific tumor markers and evolutionary traits for effective prevention or treatment (Table 1). These vaccines are designed by selecting stable nanocarriers such as polymers, liposomes, metal nanoparticles, and quantum dots. Antigens are attached to these carriers using chemical or biological methods, considering factors such as stability and immunogenicity.31 Potent adjuvants can be incorporated to enhance the immune response. Additionally, the physical and chemical attributes of the nanomaterials, including their shape, size, surface characteristics, and charge, can be adjusted to suit different diseases.32
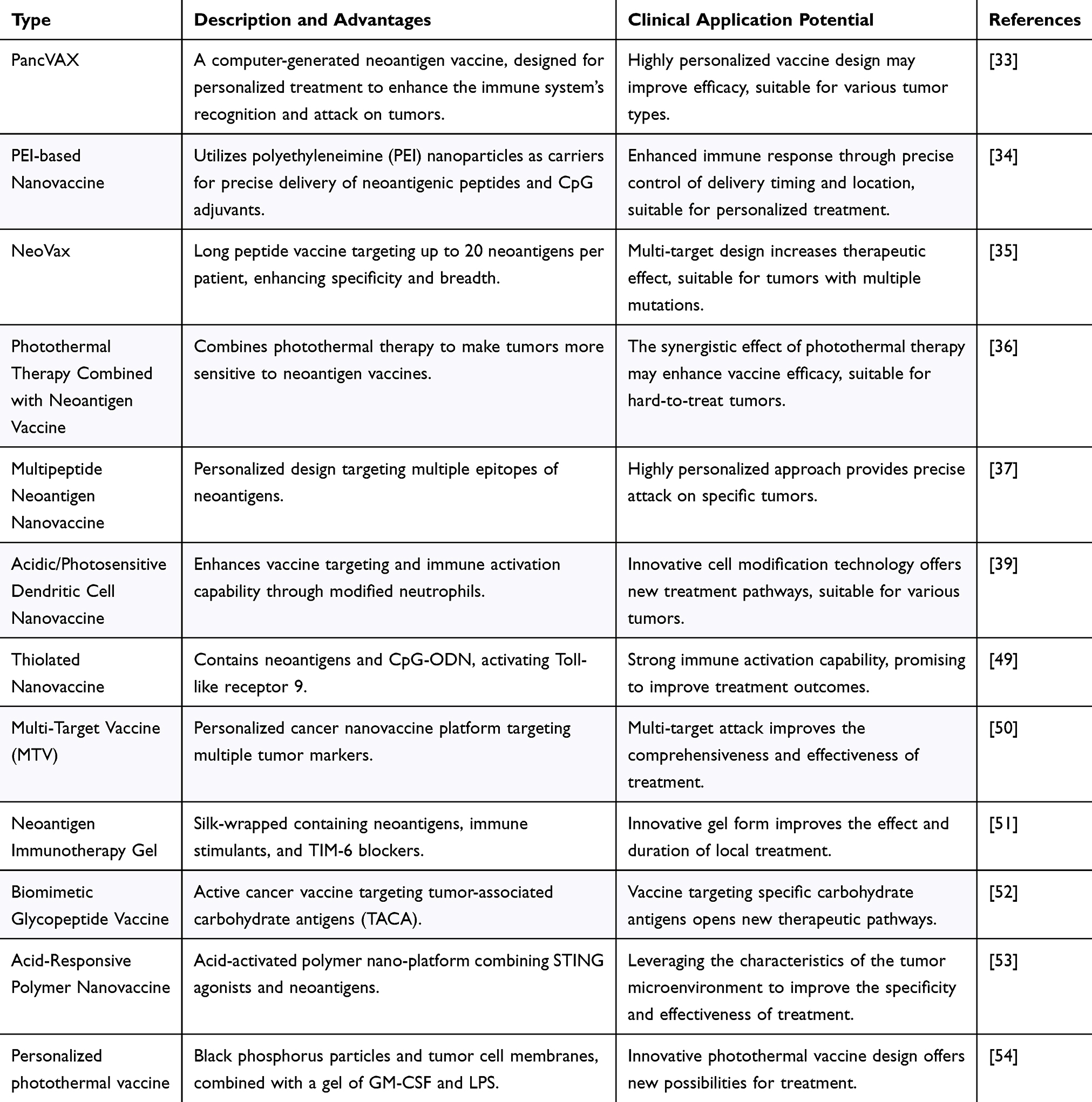 |
Table 1 Summary of Nano-Neoantigen Strategies |
Researchers are advancing neoantigen-targeted cancer vaccines and therapies to stimulate immune responses against tumors.33 Techniques such as whole-exome and ribonucleic acid (RNA) sequencing, along with predictive algorithms, are used to create personalized vaccines such as PancVAX for pancreatic cancer, which shows promise when combined with adjuvants such as ADU-V16.34 Novel vaccine platforms are being tested for the coordinated delivery of neoantigenic peptides and adjuvants, with some showing sustained immune responses in melanoma patients.35 Photothermal therapy can reportedly enhance the effectiveness of neoantigenic vaccines by altering the TME.36,37
However, the low expression of TNAs poses a challenge for immune recognition. Combined treatments, such as chemotherapy and immune checkpoint inhibitors, can increase neoantigen expression. Gene-editing technologies, such as CRISPR-Cas9, may also enhance the visibility of tumor cells to the immune system. Multifunctional platforms, such as nanoSTING-vax and DNA nanodevices, are being developed to improve vaccine delivery.38 Individualized nanovaccine platforms have been used in adjuvant cancer immunotherapy to prevent postoperative recurrence.40 Other innovative approaches include microbial-based immunotherapy and injectable nanocomposite hydrogels for tumor treatment.41,42,55 Clinical trials are ongoing, with some focusing on combining chemotherapy with immune checkpoint inhibitors to induce immunogenic tumor cell death and enhance neoantigen presentation. Despite progress, challenges, such as patient variability, tumor cell evasion, and immune suppression, remain. New strategies aim to direct tumor evolution, limit tumor escape, and promote progression using multifunctional nanovaccines.56
Immune Cell Interaction with Tumors
The immune system normally identifies and eliminates abnormal cells, including cancer cells.43 However, tumor cells can sometimes avoid detection and destruction by manipulating immune responses, leading to more aggressive cancers.57 Understanding the complex interactions between immune and tumor cells is crucial for developing new cancer treatments. The relationship between the immune system and tumor cells affects tumor growth, spread, and its response to treatment. The effect depends on the type, number, and condition of the immune cells involved. Although immune cells such as NK cells, T cells, and macrophages can attack and remove cancer cells, tumors can evolve to escape this immune response, which is a key challenge in cancer therapy. To counteract immune escape by tumors, therapies aim to boost the anti-tumor activity of the immune system, including immune checkpoint inhibitors and drugs that overcome tumor resistance to immune cell attack.44–46 Artificial intelligence, particularly machine learning, is also being explored to predict and enhance the interactions between immune cells and tumors for better treatment outcomes.
Recent advancements in nanomedicine have focused on modulating immune cells to improve tumor clearance using strategies targeting various immune cells such as killer T cells, regulatory T cells, NK cells, and macrophages. Immunotherapy remains a prominent cancer treatment method.47
Nanotherapy to Activate Killer T Cells
Killer T cells are key immune cells that recognize and attack tumor cells. However, owing to complex tumor-immune interactions, killer T cells are always in an inactivated state.48 The factors involved in killer T cell failure are complex, and recent studies have focused on several aspects (Figure 4), namely 1) the immune-checkpoint molecules regulate the T cell immune response by conveying signals between them and the tumor cells. Immune checkpoint molecules normally prevent the development of autoimmune diseases and excessive immune responses.58 However, when tumor cells develop cancer, they use these immune checkpoint molecules to inhibit the action of killer T cells, thus evading immune attack. 2) Tumor-associated cytokines: tumor cells can release substances such as TGF-β and IL-10, which can suppress the activity of immune cells.43 These factors also promote immune tolerance, allowing tumor cells to survive and grow in vivo. 3) T cell subpopulation imbalance: T cell subpopulation imbalance contributes to immune inactivation during cancer development. For example, a deficiency in CD4+ helper T cells may lead to a decrease in the tumor immune response.59 Moreover, the function and number of CD8+ killer T cells may be suppressed, leading to tumor immune escape. 4) Immune escape mechanisms: several mechanisms help tumor cells evade T cell immune attacks.60 For example, MHC molecules are often downregulated or absent in tumor cells, thereby inhibiting T cell recognition and attack. Tumor cells may also express variants of immune antigens, thus avoiding recognition by the immune system or the release of immunosuppressive agents that inhibit T cell activity.61
 |
Figure 4 The CAR-T cells can only be activated when in contact with tumor cells that express the antigen and when light is present. This figure illustrates the conditional activation of CAR-T cells, highlighting the requirement for both antigen expression by tumor cells and the presence of light for CAR-T cell activation. This dual-dependency ensures targeted and controlled therapeutic action against cancer cells, minimizing off-target effects and enhancing treatment specificity. Note: Nguyen NT, Huang K, Zeng H, et al. Nano-optogenetic engineering of CAR T cells for precision immunotherapy with enhanced safety. Nat Nanotechnol. 2021;16(12):1424–1434. Springer Nature.62 |
Several therapies targeting killer T cells have received FDA approval and successfully completed clinical trials for cancer in recent years. For example, chimeric-antigen-receptor (CAR)-T cell therapy effectively treats tumors by modifying the T cells of patients into CAR-T cells capable of recognizing and attacking cancer cells.63 Cancer cells are attacked using immune checkpoint therapy by blocking T cell immune checkpoints, which has achieved remarkable results in a variety of cancer treatments.64,65 Additionally, T cell clonal expansion techniques can increase the number and activity of immune cells by expanding the T cell population in vitro; this approach has been widely used in tumor immunotherapy.66 However, these therapies have limitations. Although CAR-T cell therapy can be used to treat a variety of hematological tumors, it is less effective for solid tumors.63 Second, killer T cell therapy is associated with a higher prevalence of side effects such as fever, nausea, and vomiting; in severe cases, the therapy may provoke an immune factor storm that can lead to organ failure.67 Some cancer cells may become resistant to activated killer T cell therapy, which may reduce its efficacy. Researchers have addressed these limitations by developing more comprehensive, precise, and effective treatments, and are exploring new targets to better exploit the benefits of activated killer T cell therapy while reducing risks and providing more options for cancer treatment (Table 2).
 |
Table 2 Summary of Nanomaterials Used for T-Cell Regulation |
One study used gold nanoparticles and computed tomography imaging to track tumor-specific T cells in vivo, providing a new monitoring method for cancer immunotherapy.68 A research team developed a method to deliver drugs using TCR signaling-responsive nanoparticles, which significantly improved tumor-specific T cell therapy.69 Other studies using a novel artificially designed cytokine have successfully expanded liver cancer-specific T cells and achieved effective tumor clearance.70 To improve the specificity of immunotherapy, another research group developed a fluoropolymer nanoparticle-based cancer vaccine that significantly enhanced the responsiveness of the immune system to specific tumor antigens.71 Owing to the development of engineered cell technologies that expand the disease treatment scope, a research team successfully treated genetic diseases using nanoparticle-delivered CRISPR/Cas9 technology, achieving behavioral recovery in a mouse model.72
Unlike most previous studies, Tian et al provided evidence that sequential changes at the injection site (sequential site-directed inoculation) significantly enhanced the immunogenicity of DNA vaccines developed in a previous study.73 Guo et al developed a “backpack” aptamer to coordinate the effectiveness of anti-tumor vaccines,75 and Song et al created a multivalent peptide-based hydrogel cancer vaccine without additional adjuvants.76 This vaccine closely mimics the immune function, effectively delivers antigenic epitopes, facilitates antigen presentation, and triggers a broad-spectrum anti-tumor CD8+ T cell response. Consequently, B16 tumor growth significantly decreased. These studies exploited the evolutionary properties of tumor cells to develop a multifunctional, specific T cell activation strategy.
Restriction of Tregs to Enhance Immunotherapy
Tregs are an important class of regulatory T cells whose main role is to maintain immune tolerance and suppress autoimmune responses; however, their role in tumor immunity is more complex.56 On one hand, Tregs can inhibit the activity of tumor-specific T cells, thus attenuating the recognition and clearance of tumor antigens and suppressing the anti-tumor immune response.77 Additionally, Tregs may inhibit tumor growth by regulating macrophage functions and releasing immunosuppressive factors to prevent tumor spread and growth.78 Immunotherapeutic strategies targeting Tregs have recently been widely studied and applied.79 The current immunotherapeutic strategies mainly involve Treg-targeting antibodies and small-molecule drugs that inhibit Treg function, combined with immune cell therapy using Tregs and tumor-specific T cells.77,80 Although Tregs are not responsible for direct tumor clearance, immunotherapy-targeting Tregs have recently received increasing attention because the specificity of Treg immune regulation is well understood. However, a deeper understanding of the role of Tregs in tumor immunity is needed to more effectively balance anti-tumor immune effects with excessive systemic immune activation.
Recent studies have demonstrated the vital role of the TGF-β signaling pathway in regulating the development and differentiation of Tregs.81 Additionally, PD-1 inhibitors may enhance the effects of immunotherapy by reducing their suppressive effect on Tregs.82 Il-33 also promotes the differentiation and activation of Tregs, thereby increasing the number and activity of immune cells.83 These findings provide important clues to further investigate the role of regulatory Tregs, indicating that the promotion of immunotherapy by regulatory Tregs requires the synergistic secretion of multiple immune checkpoint inhibitors to achieve better therapeutic effects.
Nanomedicine can enhance Treg function and offer potential treatments for tumors and autoimmune diseases. Studies have shown that nanoparticles can regulate Tregs and MDSCs, boost T cell responses, and extend survival in cancer models.84 For example, nanoparticles have been used to co-deliver checkpoint inhibitors and cytokines or combined with CD28-binding aptamers to activate T cells while suppressing Tregs, resulting in significant tumor growth inhibition.85 Nanoliposome-delivered curcumin effectively suppresses Tregs and boosts CD8+ T cell responses in melanoma and breast cancer.62 Additionally, nanocarriers have been developed for the targeted delivery of immunosuppressive drugs to modulate Tregs in skin grafts and autoantigens in multiple sclerosis models.86 These advances highlight the potential of nanotechnology-enhanced immunotherapy for the treatment of immune-related diseases with greater efficacy and precision.56
Research Advances in NK Cells and Nanotherapy
NK cells are key immune cells capable of attacking virus-infected and tumor cells, and are gaining attention in tumor immunotherapy research.87 NK cells function by 1) recognizing and destroying tumor cells without T-cell involvement via interactions between their surface receptors and MHC-I molecules; 2) producing immune-modulating factors, such as IFN-γ and TNF-α, to promote tumor cell death; and 3) this contributes to the creation of an anti-tumor immune memory for quicker and more effective responses to recurring tumor cells. Research has focused on understanding the NK cell immune surveillance mechanisms, immune memory formation, and their applications in cell-based immunotherapy, thereby offering new perspectives for the development of cancer immunotherapy strategies.88 Recent advances in the study of regulatory NK cells have focused on the following three aspects (Figure 5):
 |
Figure 5 The natural killer cells (NKs) are the body’s first line of defense against tumors and viruses. This study demonstrates a nanobiology-based drug-delivery system that enhances NK cytotoxicity by inhibiting intracellular inhibitory checkpoints in the TME. Note: Adapted from Biber G, Sabag B, Raiff A, et al. Modulation of intrinsic inhibitory checkpoints using nano-carriers to unleash NK cell activity. EMBO Mol Med. 2022;14(1):e14073. Creative Commons.89 |
- NK cell activation is linked to surface receptors, which are influenced by the local tissue environment and other cells. Modifying the TME can enhance the NK cell efficacy against tumors.75,90,91
- The anti-tumor effects of NK cells vary with the tumor localization and type. Biochemical agents such as peptidase inhibitors can increase cytotoxicity in certain cancers.92,93
- In addition to killing tumors, NK cells help regulate inflammation and augment tumor immunotherapy by interacting with T cells. Research on NK cells provides valuable insights into the immune system and development of new nanotherapies.94,95 Researchers have continued to explore and optimize methods to regulate NK cells and apply them clinically in oncology and viral infections (Table 3). Among these methods, the use of cytokines (such as IL-2 and IL-15) or the activation of specific T cell subsets (such as Vγ9Vδ2-T cells) to regulate the function and number of NK cells has become an important research direction.96–98 The exploration of related mechanisms and methods has also driven important progress in the study of nanomedicines capable of regulating NK cells. One study used single-cell analysis to examine how immunosuppression affects NK cells in different states,99 demonstrating that immunosuppression decreases the number and function of NK cells; however, the NK cell function could be partially restored by nanomedicines. This study provides a useful reference for further research and application of nanomedicines in the modulation of NK cells. Recent studies have shown that the metal ions (manganese and copper ions) present in the TME affect immunocompetence.100 In conclusion, these findings provide strong support for the design of strategies to regulate the TME and NK cells.101

Table 3 Summary of Nanomaterials That Modulate NK Cells
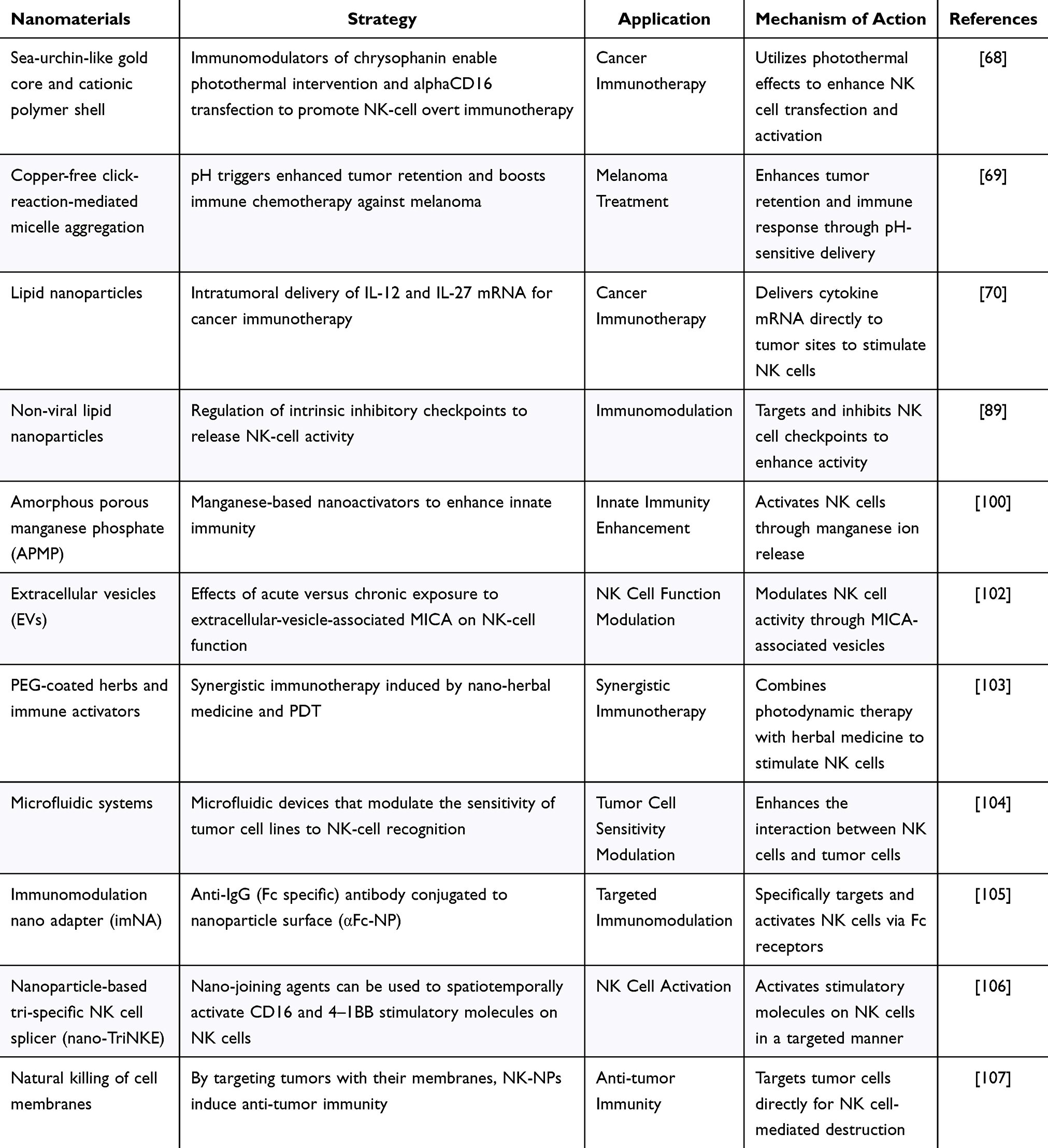
Nanotechnology enhances drug bioavailability and targeting, thereby improving the efficacy of NK cell cancer immunotherapies. For example, CD16-targeted liposomal nanoparticles were developed to specifically deliver inhibitory peptides to NK cells, thereby boosting their cytotoxic activity.108 These nanoparticles not only increased the NK cell-killing capacity but also activated NK cells by incorporating inflammatory factors. Nanocapsules containing Interleukin-15 demonstrated improved therapeutic effects against breast cancer.76 Smart nanosystems that release drugs in response to the pH of the TME have been designed to target NK cells more effectively.109 Additionally, microenvironment-responsive nanoparticles and microfluidic devices have been used to modulate NK cell activity and enhance immunotherapy against cancers, such as melanoma, by altering the surface expression of MHC-I molecules on cancer cells.110 These advancements highlight the role of the TME pH, metal ion concentration, and mechanical stress in the creation of innovative nano-based NK cell activators.
Although progress has been made in the use of nanomedicine to modulate NK cells for cancer therapy, several challenges remain, namely 1) NK cells are limited in number and can exhibit reduced activity because of tumor-derived factors, necessitating strategies to enhance their viability;111 2) NK cells lack T cell specificity, which can lead to difficulties in distinguishing cancer cells from normal cells. Future nanocarriers should be designed to improve tumor-specific recognition;112 3) Unlike T cells, NK cells do not have a memory response to tumor cells, hindering the prevention of tumor recurrence. Combining NK cell therapy with cytotoxic T cells may be beneficial;113 and 4) The TME inhibits NK cell responses through various suppressive factors. Current nanosystems require further development to more effectively modulate the TME to support NK cell activity.75
Development of Anti-Tumor Immune Effects and Drug-Carrier Properties of TMAs
M2 macrophages in the TME aid cancer progression, whereas M1 macrophages can fight tumors. Macrophages are also key to targeted drug delivery and are being explored for cell therapy to improve solid tumor treatment. The CD47-SIRPα signaling pathway, which tumors exploit to evade phagocytosis, is a current focus, with therapies targeting this pathway to enhance macrophage activity and antigen presentation.17–20 Mechanistic research is targeting various factors to modulate macrophage phenotypes in the TME, including inhibiting CCL2 to reduce M2 polarization and using molecules such as PRL-3, IRF8, and miR-181b/CCR2 to promote M1 polarization and tumor attack.114–116 Gut microbes and metabolites such as zinc gluconate also influence macrophage function, and novel strategies such as TAK1 activation are being studied to enhance macrophage-mediated tumor destruction.117 Additionally, macrophages are being investigated as potential immune checkpoint inhibitors, and findings suggest that macrophage activation can be increased by inhibiting checkpoint molecules, such as PD-1 and IL-10.118,119 These findings suggest that macrophages are promising therapeutic targets for cancer treatment (Figure 6).
 |
Figure 6 Illustration depicting the release of siRNA@M-/PTX-CA-OMV drugs triggered by pH change and its effects in vivo, including tumor cell death, blocking communication between tumor cells and macrophages, deprivation of glucose in endothelial tip cells, reconstruction of pericyte walls, and inhibition of metastasis. Note: Adapted from Guo Q, Li X, Zhou W, et al. Sequentially triggered bacterial outer membrane vesicles for macrophage metabolism modulation and tumor metastasis suppression. ACS Nano. 2021;15(8):13826–13838. Copyright © 2021 American Chemical Society.120 |
Nanomedical research is increasingly focusing on macrophages in cancer therapy, aiming to shift their polarization from a tumor-promoting M2 phenotype to a tumor-fighting M1 phenotype.121 Song et al used manganese dioxide nanoparticles to direct macrophages towards the M1 phenotype, thereby reducing tumor size.122 Ramesh et al developed double inhibitor-loaded nanoparticles (DNTs) that blocked the CSF1R and SHP2 pathways, effectively converting M2 to M1 macrophages and enhancing phagocytosis.123 These DNTs outperformed the single-drug treatments. Wang et al reported a nanodelivered drug that inhibits breast cancer growth by inducing M1 macrophage conversion and modulating inflammatory cytokines.124 Small RNA delivery to macrophages has also been explored, with miR-155-loaded nanoparticles reducing TAMs and siRNA-loaded nanoparticles targeting KRAS mutations in tumor cells and macrophages, thereby improving therapeutic outcomes. Qin et al designed a pH-sensitive drug delivery system capable of targeting the acidic TME, which released paclitaxel in response to the tumor pH and increased glycolysis in M2 macrophages.124
Recent advances in nanotechnology have resulted in the development of multifunctional nanocarriers and drug combinations for targeted drug delivery. Research has shown that nanodrugs can target both macrophages and cancer cells, release drugs in response to environmental stimuli and modulate immune responses to enhance therapy (Table 4). Novel nanocarriers have been used to deliver anti-tumor drugs and induce macrophage polarization to assist in tumor immunotherapy.125 Additionally, nanoparticle/bacterial complexes and immune-stimulating agents have been employed to target and reprogram TAMs from an immunosuppressive to a pro-inflammatory state.126 The potential of M1-like macrophages as antigen-presenting cells in anti-tumor immunity has also been explored using specific nanoparticles designed to promote this conversion.127 These innovations underscore the potential of nanotechnology to improve the efficacy of cancer immunotherapy.
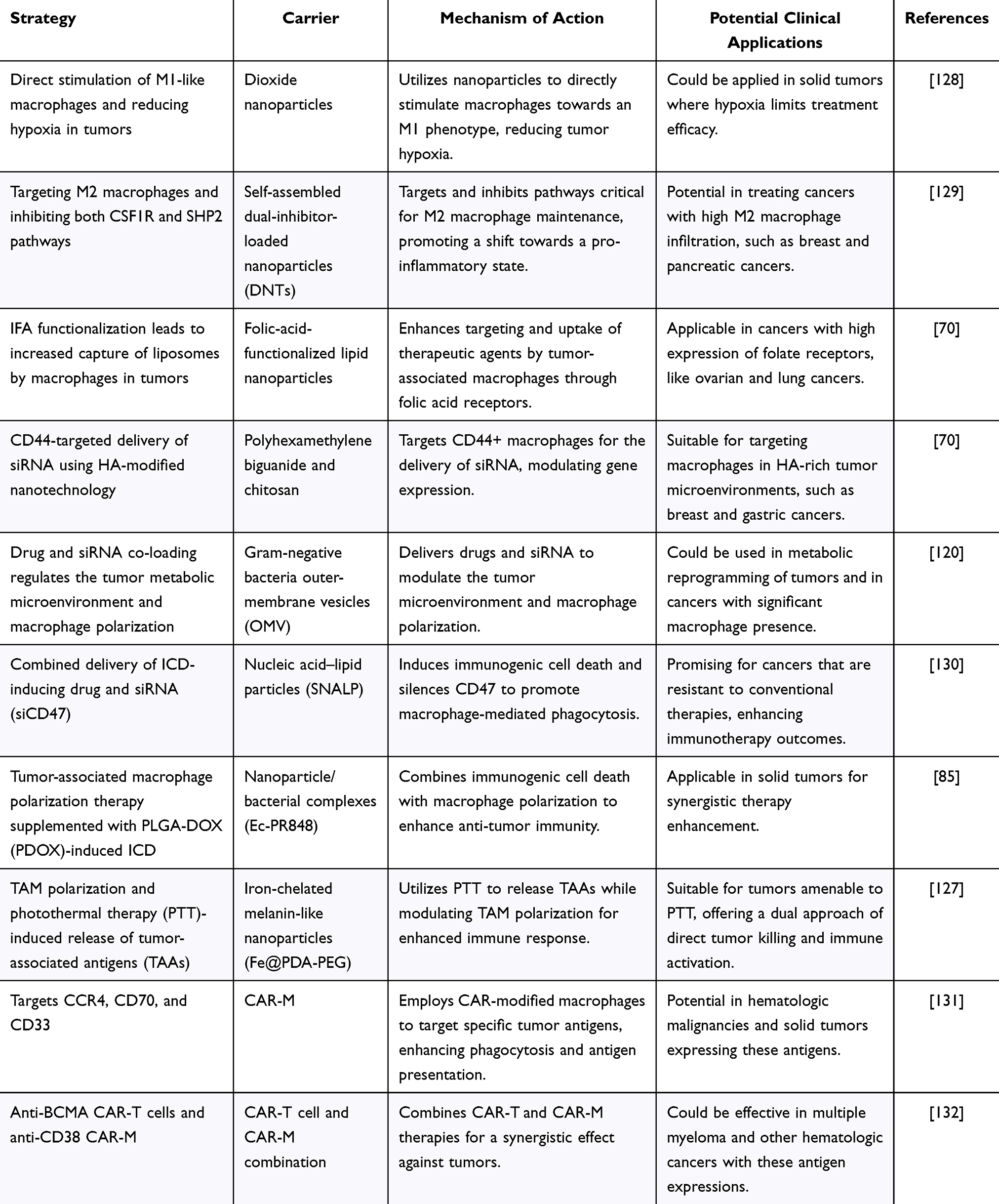 |
Table 4 Summary of Macrophages as Therapeutic Targets and Vector Strategies |
CAR-T cells are currently successfully used to treat solid tumors and hematologic cancers. Access to solid tumors is a stumbling block for T cell therapy. Macrophage-based cell therapy is expected to overcome this limitation as mononuclear phagocytes continue to be transferred into tumors, and CAR-modified myeloid cell (CAR-M) therapy is gaining attention.131 CAR-M cell therapies are rapidly developing and are mainly based on CAR structures in human bone marrow or hematopoietic stem cells, which are genetically edited and transplanted into patients. Current research is focused on identifying more specific targets and refining the dose and treatment regimens for CAR-M therapy. Among these, CAR-M cell therapies targeting anti-CCR4, -CD70, and -CD33 have achieved remarkable therapeutic effects in solid tumors and leukemia.133–135 Dual CAR-T and CAR-M therapies have demonstrated complementary advantages. Moreover, dual-specific CD38 and BCMA CAR-T cells have been shown to be effective and safe for multiple myeloma (MM) with relapses and refractory disease.136 Using different CAR cells for both targets is reportedly highly effective, and simultaneous application of anti-BCMA CAR-T cells and anti-CD38 CAR-M cells for MM can effectively inhibit MM growth and prevent the emergence of multidrug resistance.137
CAR-M therapy has the advantages of high targeting ability, significant therapeutic effects, and better tolerability. However, CAR-M therapy still faces many challenges. For example, the production and processing of CAR-M is immature and presents potential safety risks and the therapeutic effects of CAR-M are still limited, which may cause severe side effects.138 Therefore, CAR-M therapy must be more comprehensively studied to improve its efficacy and safety, and further clinical studies must be conducted to realize its clinical application.
Macrophages are an integral part of the TME and interact with cancer cells, stroma, and immune cells in complex ways. Macrophages play a dual role in the efficacy of current therapeutic modalities, including chemotherapy and immunotherapy.139 TAMs are major drivers of T cell checkpoint blockade and mediate resistance to immunotherapy. Although the direct targeting of bone marrow monocytes faces challenges, ongoing trials targeting TAMs in combination with other therapies are promising. The main challenge in developing TAM immunobiological therapy lies in the diversity and plasticity of mononuclear phagocytes in tumors.140 Identifying and targeting specific subpopulations may offer new strategies for the selective depletion and suppression of tumor-promoting cells. Beyond the CD47-SIRPα axis, various molecular targets for regulating myelomonocyte function have been identified, including CSF1R and CCR2, which are crucial for the survival, proliferation, differentiation, and recruitment of myelomonocytes to the TME. Additionally, TAM receptors, such as Tyro3, Axl, and MerTK, play roles in the clearance of apoptotic cells and immune regulation.141 These insights open new avenues for smart nanomedicines to specifically modulate myelomonocyte function and enhance therapeutic outcomes in cancer therapy. Complementary approaches for immunotherapy are required to fully harness the potential of TAM-targeted therapies.142,143 The attraction of monocytes to tumor tissues may be useful for developing cellular therapies for solid tumors. However, the selection of receptors, circulating precursor replenishment, survival time in the tumor environment, functional orientation, and spatial localization within the tumor tissue must still be fully understood.144 Clinical trials may offer an opportunity to address these critical questions for CAR-M therapy development.
Defective Nature of Fibroblasts as Therapeutic Tumor Targets and Their Multipotency as Adjuvants
CAFs are cells found in primary and metastatic cancers that participate in cancer progression by interacting with other cells in the TME.46 CAFs produce extracellular matrix components and undergo epigenetic modifications leading to the production of cellular factors, metabolites, and exosomes. These factors modulate tumor angiogenesis, immune escape, and its metabolism.145 Most clinical trials targeting CAFs failed and may have accelerated cancer progression.146 The characteristics of and interactions between CAFs and other cells change dynamically as cancer progresses.147 Further research is required to understand the bimodal functions of the different CAF subtypes to develop new diagnostic and therapeutic approaches.148
Recent Research Investigating the Influence of CAFs on the TME
Cancer development, progression, and metastasis lead to dynamic changes in the host tissue and formation of a complex tumor stroma known as the TME (Figure 7). The tumor stroma in certain cancer types, such as pancreatic and breast cancer, develops because of connective tissue proliferation, resulting in abundant fibrous and/or connective tissues. The key tumor stroma elements, including angiogenesis, immune response, metabolism, hypoxia, and fibroblasts, have been studied as potential therapeutic targets.149,150 Recent studies using single-cell RNA sequencing (scRNA-seq) of different cancers revealed that CAFs are highly heterogeneous mesenchymal spectrum cells with various recognized functions.151–155 Ongoing research is investigating whether CAFs form a distinct class of mesenchymal cells with distinct functions. Similar to fibroblasts in non-malignant tissue or wound healing, CAFs have essential features such as the ability to secrete phenotypes and synthesize the ECM. CAFs may also have altered epigenetic and transcriptional profiles.152 However, their roles in cancer progression remain unclear. Preclinical studies suggest that fibroblast activation is a host response and defense mechanism against tumors.153 CAFs can create a physical barrier to the ECM via encapsulation, which is a connective molding response.153 However, tumor growth is supported by the hijacking of wound healing mechanisms by cancer cells following tumor formation.154 Therefore, the functional heterogeneity of CAFs may result in fibroblasts supporting or suppressing tumors according to the context.
 |
Figure 7 Schematic of CAF-specific imaging. A peptide-based NIR probe was developed to selectively react with fibroblast-activating protein alpha (FAP-α) and form nanofibers specifically on the surface of cancer-associated fibroblasts (CAFs). The probe utilized an assembly/aggregation-induced retention (AIR) effect, which resulted in enhanced accumulation and retention of the probe around the tumor. After administration, the tumor showed a 5.48-fold increase in signal intensity compared with the non-aggregated control molecule at five hours. Additionally, the selective assembly of the probe resulted in higher signal intensities in tumors than in the liver and kidneys, with four- and five-fold differences, respectively. Note: Zhao XX, Li LL, Zhao Y, et al. In situ self-assembled nanofibers precisely target cancer-associated fibroblasts for improved tumor imaging. Angew Chem Int Ed Engl. 2019;58(43):15287–15294. © 2019 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim.156 |
CAFs are Important Contributors to and Target of Tumor Drug Resistance
Through their coordination of multiple functions, CAFs play an important role in tumor progression, both within the TME and throughout the host, and despite the fact that CAFs disrupt the adhesion of cancer cells and promote invasion and metastasis, cancer cells can survive with CAF adhesion.153 CAFs also produce a series of cytokines, chemokines, and other effector molecules, which are involved in the anti-tumor immunity mediated by immune cells.155 CAFs recruit and suppress immunosuppressive cells while limiting cytokine production and effector immune cells [136]. Direct attachment of cancer cells to CAFs may impart resistance by increasing the survival signals within tumor cells.157 CAFs also help suppress immunity through ECM remodeling by regulating matrix metalloproteinases, collagen, polysaccharides, and fibronectin.158 Finally, CAFs overexpress surface immune checkpoint molecules that inhibit apoptosis, thereby contributing to their function and activation.
Treatment Strategies for CAFs
Researchers have identified a variety of mechanisms by which CAFs promote tumor growth. Some therapeutic strategies, such as those targeting hyaluronic acid, have alarmingly not been sufficiently effective in clinical trials or, in some cases, have shortened patient survival.157,159 These unexpected results highlight the importance of considering CAF heterogeneity for tumor promotion and neoplasia and are important for targeting specific subpopulations of CAFs to develop effective therapeutic interventions. Standardizing and reprogramming CAFs using substances such as vitamin D or carbotriol has been demonstrated to inhibit their tumor-promoting functions and improve pancreatic cancer treatment outcomes in preclinical models.158 JAK inhibitors can reportedly reprogram CAF isoforms, resulting in increased ECM deposition and tumor growth inhibition.158 A more comprehensive understanding of gene expression and the function of different fibroblast populations in normal and cancerous tissues is needed to design new strategies to effectively reprogram CAFs to a “normal” or inactive state.
Based on small-molecule screening results, scriptaid inhibited CAF differentiation through Histone Deacetylases 1, 3, and 8.158 Scriptaid reduced cell stiffness and contraction, inhibited ECM secretion, reduced the number of CAFs, and retarded tumor growth. CAF and Pancreatic Ductal Adenocarcinoma accelerate their migration and invasion via the CXCL-CXCR2 axis.158 Although preclinical studies have demonstrated the advantages of targeting only CAF, targeting both CAF compartments and tumors may improve the outcomes of clinical trials. A new dendrimer linked to the mTOR inhibitor rapamycin effectively reduced VEGF expression and mTOR signaling in both fibroblasts and prostate cancer cells, which resulted in a decrease in fibroblast-mediated progression and metastasis of prostate tumors.160 Dual delivery of PT-100 and oxaliplatin chemotherapy in a preclinical trial was the only treatment that improved the survival rate of mice as it decreased the colon tumor growth, CAF content, pre-tumor immune cell recruitment, and angiogenesis. Neutralizing tGF-β in a mouse model shifted the CAF kinetics, significantly reducing the activity and frequency of myCAF subpopulations, while promoting the development of a fibroblast population characterized by a strong response to interferon and an amplified immunomodulatory profile.161
Progress of Nanoagents in CAF Imaging and Treatment
Inhibitors that specifically target CAFs have yet to be approved, largely because of the lack of precision in targeting CAF-specific pathways. Some direct targeting strategies, such as inhibiting CAF signaling pathways or depleting the general CAF population,145 have been explored; however, these may also affect other cells that share similar pathways. However, this approach may deplete other cells via similar signaling pathways. Furthermore, the depletion of the entire CAF population, rather than specific subpopulations, eliminates both the pre-tumor and anti-tumor CAF subpopulations.162 Therefore, researchers have used nanodelivery systems to accurately transport drugs for CAF imaging, offering enhanced stability, solubility, and bioavailability with lower systemic toxicity. Researchers found that CAFs can be imaged using multifunctional nanoparticles combined with fluorescent probes.163 Localization and tracking of CAFs in humans can reportedly be achieved by using magnetic nanoparticles for magnetic resonance imaging techniques.164
Nanodelivery systems that target CAFs to improve the effectiveness of anticancer drugs have significantly advanced in recent years (Table 5). These systems use active drugs that disrupt the function of CAFs and help drugs accumulate within the tumor core.165 Various nanoparticles have been explored, including those targeting FAP, Hh-activated active targeting, angiotensin II receptor, sigma receptor, SPARC, tenascin, PDGFR, M6P/IGFII receptor, CXCR4, and integrin αVβ3.165 Biodegradable polymeric NPs loaded with twisted α-invertin and coated with the CREKA peptide have also been developed to regulate the TME by blocking CAF activation and enhancing CAF uptake.166 Similarly, cyclopamine, which depletes CAFs, was encapsulated in polymeric micelles along with paclitaxel to improve therapeutic efficacy.167 Enzymes overexpressed on CAF membranes, such as albumin nanoparticles of PTX encapsulated into CAP-modified liposomes that are responsive to FAP-α, have also been used to optimize drug delivery.168 These developments offer promising avenues for improving cancer treatments by targeting CAFs.
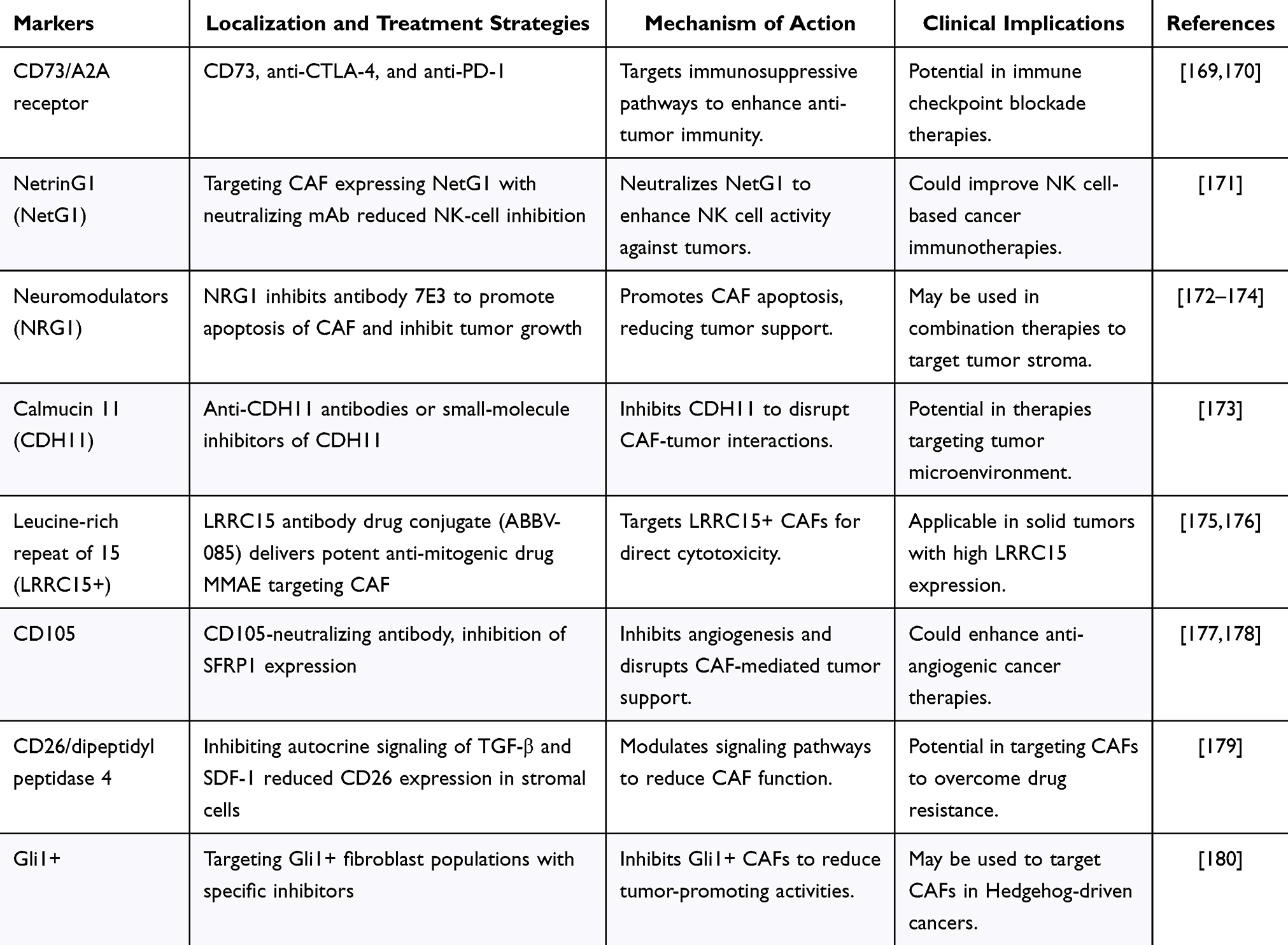 |
Table 5 Summary of CAF-Specific Targets |
Nanocarriers produced from hybrid nanomaterials, such as gold core and silver shell nanoparticles (Ag@AuNPs), have been shown to inhibit CAFs by promoting tumor growth, leading to reduced metastasis in animal models.181 These nanoparticles alter gene expression related to cancer invasion and metastasis, effectively immobilizing CAFs by inducing lipid production.182 However, targeting CAFs is challenging because of their diversity and the difficulty in identifying specific markers and subpopulations. Single-cell genomics advances may improve our understanding of CAF heterogeneity.183 Personalized therapies based on CAF phenotypes can minimize side effects and enhance patient outcomes. Despite this progress, issues such as precise targeting, material toxicity, and immune responses hamper the development of nanomaterials for CAF-targeted therapies.
Summary and Outlook
This article explores the intricate landscape of tumor evolution and the pivotal role that smart nanomedicines play in current and future cancer therapy paradigms. The unique physiological characteristics of tumors, including weak acidity, low oxygen content, and the upregulation of specific enzymes, present challenges and opportunities for targeted cancer treatments. Smart nanomedicines with the ability to adapt and respond to the TME have emerged as a promising avenue for overcoming the limitations of traditional therapies and addressing the complex heterogeneity of tumors. The development of smart nanomedicines has been driven by a deeper understanding of the TME and tumor evolution. These advanced therapeutic agents are designed to exploit the unique features of the TME and offer improved selectivity, permeability, and drug-release properties. Recent advancements have highlighted the potential of targeting the tumor cell surface receptors, physiological and pathological characteristics of tumor tissues, and dynamic interactions within the TME. However, the evolution of the tumor characteristics and adaptability of cancer cells continue to pose significant challenges, underscoring the need for ongoing research and innovation.
The smart nanomedicines and cancer therapy field is poised for transformative advancements. The future of cancer treatment is likely to be characterized by a more personalized approach, leveraging the latest advances in genomic sequencing and bioinformatics to tailor therapies to the tumor profile of an individual. This precision medicine approach requires a deeper understanding of tumor heterogeneity and the development of nanomedicines capable of adapting to the evolving TME landscape. Innovative research and development efforts are crucial to overcome the current limitations of smart nanomedicines. These efforts include exploring new materials and delivery systems, enhancing the targeting capabilities of nanomedicines, and developing strategies to circumvent drug resistance. Additionally, integrating smart nanomedicines with other therapeutic modalities, such as immunotherapy and targeted therapy, offers the potential for synergistic effects that could significantly improve treatment outcomes. The integration of artificial intelligence and machine learning technologies into the design and optimization of nanomedicines presents another exciting frontier. These technologies can aid in predicting the behavior of nanomedicines within the TME, optimizing drug delivery, and identifying novel therapeutic targets.
In conclusion, the smart nanomedicine and cancer therapy field is at a critical juncture with the potential to revolutionize cancer treatment. Continued exploration and innovation are essential to unlock the full potential of smart nanomedicines in combating the complex and dynamic nature of tumors. A multidisciplinary approach, combining insights from nanotechnology, oncology, immunology, and computational biology, will be key to advancing the field and achieving the ultimate goal of effective, personalized cancer therapy.
Funding
This work was financially supported by the National Natural Science Foundation of China (82203069, 82272002, 81901747) and Natural Science Foundation of Shanghai, China (22ZR1438500).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Roehlen N, Muller M, Nehme Z, et al. Treatment of HCC with claudin-1-specific antibodies suppresses carcinogenic signaling and reprograms the tumor microenvironment. J Hepatol. 2023;78(2):343–355. doi:10.1016/j.jhep.2022.10.011
2. Cillo AR, Kurten CHL, Tabib T, et al. Immune landscape of viral- and carcinogen-driven head and neck cancer. Immunity. 2020;52(1):183–199 e9. doi:10.1016/j.immuni.2019.11.014
3. Cheng Z, Li M, Dey R, Chen Y. Nanomaterials for cancer therapy: current progress and perspectives. J Hematol Oncol. 2021;14(1):85. doi:10.1186/s13045-021-01096-0
4. Peng S, Xiao F, Chen M, Gao H. Tumor-microenvironment-responsive nanomedicine for enhanced cancer immunotherapy. Adv Sci. 2022;9(1):e2103836. doi:10.1002/advs.202103836
5. Yang G, Xu L, Chao Y, et al. Hollow MnO(2) as a tumor-microenvironment-responsive biodegradable nano-platform for combination therapy favoring antitumor immune responses. Nat Commun. 2017;8(1):902. doi:10.1038/s41467-017-01050-0
6. Jia X, Zhang Y, Zou Y, et al. Dual Intratumoral Redox/Enzyme-Responsive NO-releasing nanomedicine for the specific, high-efficacy, and low-toxic cancer therapy. Adv Mater. 2018;30(30):e1704490. doi:10.1002/adma.201704490
7. Wu Y, Chen S, Zhu J. Hydrogen bond-mediated supramolecular polymeric nanomedicine with pH/Light-responsive methotrexate release and synergistic chemo-/photothermal therapy. Biomacromolecules. 2022;23(10):4230–4240. doi:10.1021/acs.biomac.2c00717
8. Liu Y, Jin J, Xu H, et al. Construction of a pH-responsive, ultralow-dose triptolide nanomedicine for safe rheumatoid arthritis therapy. Acta Biomater. 2021;121:541–553. doi:10.1016/j.actbio.2020.11.027
9. Gao H, Bi Y, Wang X, et al. Near-infrared guided thermal-responsive nanomedicine against orthotopic superficial bladder cancer. ACS Biomater Sci Eng. 2017;3(12):3628–3634. doi:10.1021/acsbiomaterials.7b00405
10. Wang X, Sun B, Ye Z, et al. Enzyme-Responsive COF-based thiol-targeting nanoinhibitor for curing bacterial infections. ACS Appl Mater Interfaces. 2022;14(34):38483–38496. doi:10.1021/acsami.2c08845
11. Sa P, Sahoo SK, Dilnawaz F. Responsive role of nanomedicine in the tumor microenvironment and cancer drug resistance. Curr Med Chem. 2022. doi:10.2174/0929867329666220922111336
12. Mendes BB, Sousa DP, Conniot J, Conde J. Nanomedicine-based strategies to target and modulate the tumor microenvironment. Trends Cancer. 2021;7(9):847–862. doi:10.1016/j.trecan.2021.05.001
13. Iyer AK, Singh A, Ganta S, Amiji MM. Role of integrated cancer nanomedicine in overcoming drug resistance. Adv Drug Deliv Rev. 2013;65(13–14):1784–1802. doi:10.1016/j.addr.2013.07.012
14. Erin N, Grahovac J, Brozovic A, Efferth T. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist Updat. 2020;53:100715. doi:10.1016/j.drup.2020.100715
15. Manabe T, Bivona TG. Remodeling of the tumor/tumor microenvironment ecosystem during KRAS G12C inhibitor clinical resistance in lung cancer. J Clin Invest. 2022;132(4). doi:10.1172/JCI156891
16. Liu HH, Zhang ZY, Song L, Gao J, Liu YF. Lipid metabolism of cancer stem cells. Oncol Lett. 2022;23(4). doi:10.3892/ol.2022.13239
17. Chen D, Zhang X, Li Z, Zhu B. Metabolic regulatory crosstalk between tumor microenvironment and tumor-associated macrophages. Theranostics. 2021;11(3):1016–1030. doi:10.7150/thno.51777
18. Liu Y, Tiruthani K, Wang M, et al. Tumor-targeted gene therapy with lipid nanoparticles inhibits tumor-associated adipocytes and remodels the immunosuppressive tumor microenvironment in triple-negative breast cancer. Nanoscale Horiz. 2021;6(4):319–329. doi:10.1039/d0nh00588f
19. Lan Q, Lai W, Zeng Y, et al. CCL26 Participates in the PRL-3-induced promotion of colorectal cancer invasion by stimulating tumor-associated macrophage infiltration. Mol Cancer Ther. 2018;17(1):276–289. doi:10.1158/1535-7163.MCT-17-0507
20. Qin S, LaRosa G, Campbell JJ, et al. Expression of monocyte chemoattractant protein-1 and interleukin-8 receptors on subsets of T cells: correlation with transendothelial chemotactic potential. Eur J Immunol. 1996;26(3):640–647. doi:10.1002/eji.1830260320
21. Wang YC, Tsai SH, Chen MH, et al. Mineral nanomedicine to enhance the efficacy of adjuvant radiotherapy for treating osteosarcoma. ACS Appl Mater Interfaces. 2022;14(4):5586–5597. doi:10.1021/acsami.1c21729
22. Gillies RJ, Verduzco D, Gatenby RA. Evolutionary dynamics of carcinogenesis and why targeted therapy does not work. Nat Rev Cancer. 2012;12(7):487–493. doi:10.1038/nrc3298
23. van der Meel R, Sulheim E, Shi Y, Kiessling F, Mulder WJM, Lammers T. Smart cancer nanomedicine. Nat Nanotechnol. 2019;14(11):1007–1017. doi:10.1038/s41565-019-0567-y
24. Shi Y, Lammers T. Combining nanomedicine and immunotherapy. Acc Chem Res. 2019;52(6):1543–1554. doi:10.1021/acs.accounts.9b00148
25. Peng M, Mo Y, Wang Y, et al. Neoantigen vaccine: an emerging tumor immunotherapy. Mol Cancer. 2019;18(1):128. doi:10.1186/s12943-019-1055-6
26. Zhang Z, Lu M, Qin Y, et al. Neoantigen: a new breakthrough in tumor immunotherapy. Front Immunol. 2021;12:672356. doi:10.3389/fimmu.2021.672356
27. Blass E, Ott PA. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat Rev Clin Oncol. 2021;18(4):215–229. doi:10.1038/s41571-020-00460-2
28. Lybaert L, Lefever S, Fant B, et al. Challenges in neoantigen-directed therapeutics. Cancer Cell. 2023;41(1):15–40. doi:10.1016/j.ccell.2022.10.013
29. Musetti S, Huang L. Nanoparticle-mediated remodeling of the tumor microenvironment to enhance immunotherapy. ACS Nano. 2018;12(12):11740–11755. doi:10.1021/acsnano.8b05893
30. Zhang Y, Zhang Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol Immunol. 2020;17(8):807–821. doi:10.1038/s41423-020-0488-6
31. Zhang QL, Hong S, Dong X, et al. Bioinspired nano-vaccine construction by antigen pre-degradation for boosting cancer personalized immunotherapy. Biomaterials. 2022;287:121628. doi:10.1016/j.biomaterials.2022.121628
32. Huang D, Wu T, Lan S, Liu C, Guo Z, Zhang W. In situ photothermal nano-vaccine based on tumor cell membrane-coated black phosphorus-Au for photo-immunotherapy of metastatic breast tumors. Biomaterials. 2022;289:121808. doi:10.1016/j.biomaterials.2022.121808
33. Kinkead HL, Hopkins A, Lutz E, et al.. Combining STING-based neoantigen-targeted vaccine with checkpoint modulators enhances antitumor immunity in murine pancreatic cancer. JCI Insight. 2018;3(20). doi:10.1172/jci.insight.122857
34. Park KS, Nam J, Son S, Moon JJ. Personalized combination nano-immunotherapy for robust induction and tumor infiltration of CD8(+) T cells. Biomaterials. 2021;274:120844. doi:10.1016/j.biomaterials.2021.120844
35. Hu Z, Leet DE, Allesoe RL, et al. Personal neoantigen vaccines induce persistent memory T cell responses and epitope spreading in patients with melanoma. Nat Med. 2021;27(3):515–525. doi:10.1038/s41591-020-01206-4
36. Nam J, Son S, Park KS, Moon JJ. Photothermal therapy combined with neoantigen cancer vaccination for effective immunotherapy against large established tumors and distant metastasis. Adv Ther. 2021;4:8.
37. Sha H, Liu Q, Xie L, et al. Case report: pathological complete response in a lung metastasis of phyllodes tumor patient following treatment containing peptide neoantigen nano-vaccine. Front Oncol. 2022;12:800484. doi:10.3389/fonc.2022.800484
38. Shae D, Baljon JJ, Wehbe M, et al. Co-delivery of peptide neoantigens and stimulator of interferon genes agonists enhances response to cancer vaccines. ACS Nano. 2020;14(8):9904–9916. doi:10.1021/acsnano.0c02765
39. Wang Y, Zhao Q, Zhao B, et al. Remodeling tumor-associated neutrophils to enhance dendritic cell-based HCC neoantigen nano-vaccine efficiency. Adv Sci. 2022;9(11):e2105631. doi:10.1002/advs.202105631
40. Liu Q, Chu Y, Shao J, et al. Benefits of an immunogenic personalized neoantigen nanovaccine in patients with high-risk gastric/gastroesophageal junction cancer. Adv Sci (Weinh). 2022;10(1):e2203298. doi:10.1002/advs.202203298
41. Selvanesan BC, Chandra D, Quispe-Tintaya W, et al. Listeria delivers tetanus toxoid protein to pancreatic tumors and induces cancer cell death in mice. Sci Transl Med. 2022;14(637):eabc1600. doi:10.1126/scitranslmed.abc1600
42. Zhang W, Shi Y, Li H, et al. In situ injectable nano-complexed hydrogel based on chitosan/dextran for combining tumor therapy via hypoxia alleviation and TAMs polarity regulation. Carbohydr Polym. 2022;288:119418. doi:10.1016/j.carbpol.2022.119418
43. Sellars MC, Wu CJ, Fritsch EF. Cancer vaccines: building a bridge over troubled waters. Cell. 2022;185(15):2770–2788. doi:10.1016/j.cell.2022.06.035
44. Cheng K, Zhao R, Li Y, et al. Bioengineered bacteria-derived outer membrane vesicles as a versatile antigen display platform for tumor vaccination via Plug-and-Display technology. Nat Commun. 2021;12(1):2041. doi:10.1038/s41467-021-22308-8
45. Younis NK, Roumieh R, Bassil EP, Ghoubaira JA, Kobeissy F, Eid AH. Nanoparticles: attractive tools to treat colorectal cancer. Semin Cancer Biol. 2022;86(Pt 2):1–13. doi:10.1016/j.semcancer.2022.08.006
46. Mao X, Xu J, Wang W, et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021;20(1):131. doi:10.1186/s12943-021-01428-1
47. Zhang N, Zhang H, Wu W, et al. Machine learning-based identification of tumor-infiltrating immune cell-associated lncRNAs for improving outcomes and immunotherapy responses in patients with low-grade glioma. Theranostics. 2022;12(13):5931–5948. doi:10.7150/thno.74281
48. Oh DY, Fong L. Cytotoxic CD4(+) T cells in cancer: expanding the immune effector toolbox. Immunity. 2021;54(12):2701–2711. doi:10.1016/j.immuni.2021.11.015
49. Zhang D, Lin Z, Wu M, et al. Cytosolic delivery of thiolated neoantigen nano-vaccine combined with immune checkpoint blockade to boost anti-cancer T cell immunity. Adv Sci. 2021;8(6):2003504. doi:10.1002/advs.202003504
50. Mohsen MO, Vogel M, Riether C, et al. Targeting mutated plus germline epitopes confers pre-clinical efficacy of an instantly formulated cancer nano-vaccine. Front Immunol. 2019;10:1015. doi:10.3389/fimmu.2019.01015
51. Zhao Q, Wang Y, Zhao B, et al. Neoantigen immunotherapeutic-gel combined with TIM-3 blockade effectively restrains orthotopic hepatocellular carcinoma progression. Nano Lett. 2022;22(5):2048–2058. doi:10.1021/acs.nanolett.1c04977
52. Reuven EM, Leviatan Ben-Arye S, Yu H, et al. Biomimetic glyconanoparticle vaccine for cancer immunotherapy. ACS Nano. 2019;13(3):2936–2947. doi:10.1021/acsnano.8b07241
53. Zhou L, Hou B, Wang D, et al. Engineering polymeric prodrug nanoplatform for vaccination immunotherapy of cancer. Nano Lett. 2020;20(6):4393–4402. doi:10.1021/acs.nanolett.0c01140
54. Ye X, Liang X, Chen Q, et al. Surgical tumor-derived personalized photothermal vaccine formulation for cancer immunotherapy. ACS Nano. 2019;13(3):2956–2968. doi:10.1021/acsnano.8b07371
55. Kanemura H, Hayashi H, Tomida S, et al. The tumor immune microenvironment and frameshift neoantigen load determine response to PD-L1 blockade in extensive-stage SCLC. JTO Clin Res Rep. 2022;3(8):100373. doi:10.1016/j.jtocrr.2022.100373
56. Badr MY, Halwani AA, Odunze U, et al. The topical ocular delivery of rapamycin to posterior eye tissues and the suppression of retinal inflammatory disease. Int J Pharm. 2022;621:121755. doi:10.1016/j.ijpharm.2022.121755
57. Dumauthioz N, Labiano S, Romero P. Tumor resident memory T Cells: new players in immune surveillance and therapy. Front Immunol. 2018;9:2076. doi:10.3389/fimmu.2018.02076
58. Zhou G, Sprengers D, Boor PPC, et al. Antibodies against immune checkpoint molecules restore functions of tumor-infiltrating T cells in hepatocellular carcinomas. Gastroenterology. 2017;153(4):1107–1119 e10. doi:10.1053/j.gastro.2017.06.017
59. Kim HD, Song GW, Park S, et al. Association between expression level of PD1 by tumor-infiltrating CD8(+) T cells and features of hepatocellular carcinoma. Gastroenterology. 2018;155(6):1936–1950 e17. doi:10.1053/j.gastro.2018.08.030
60. Lv H, Lv G, Chen C, et al. NAD(+) metabolism maintains inducible PD-L1 expression to drive tumor immune evasion. Cell Metab. 2021;33(1):110–127 e5. doi:10.1016/j.cmet.2020.10.021
61. Guo W, Gao H, Li H, et al. Self-assembly of a multifunction DNA Tetrahedron for Effective Delivery of Aptamer PL1 and Pcsk9 siRNA potentiate immune checkpoint therapy for colorectal cancer. ACS Appl Mater Interfaces. 2022;14(28):31634–31644. doi:10.1021/acsami.2c06001
62. Nguyen NT, Huang K, Zeng H, et al. Nano-optogenetic engineering of CAR T cells for precision immunotherapy with enhanced safety. Nat Nanotechnol. 2021;16(12):1424–1434. doi:10.1038/s41565-021-00982-5
63. Sterner RC, Sterner RM. CAR-T cell therapy: current limitations and potential strategies. Blood Cancer J. 2021;11(4):69. doi:10.1038/s41408-021-00459-7
64. Zhang L, Su HL, Wang HL, et al. Tumor chemo-radiotherapy with rod-shaped and spherical gold nano probes: shape and active targeting both matter. Theranostics. 2019;9(7):1893–1908. doi:10.7150/thno.30523
65. Topalian SL, Taube JM, Anders RA, Pardoll DM. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat Rev Cancer. 2016;16(5):275–287. doi:10.1038/nrc.2016.36
66. Tran E, Robbins PF, Lu YC, et al. T-cell transfer therapy targeting mutant KRAS in cancer. N Engl J Med. 2016;375(23):2255–2262. doi:10.1056/NEJMoa1609279
67. Ma S, Li X, Wang X, et al. Current progress in CAR-T cell therapy for solid tumors. Int J Biol Sci. 2019;15(12):2548–2560. doi:10.7150/ijbs.34213
68. Meir R, Shamalov K, Betzer O, et al. Nanomedicine for cancer immunotherapy: tracking cancer-specific T-cells in vivo with gold nanoparticles and CT imaging. ACS Nano. 2015;9(6):6363–6372. doi:10.1021/acsnano.5b01939
69. Tang L, Zheng Y, Melo MB, et al. Enhancing T cell therapy through TCR-signaling-responsive nanoparticle drug delivery. Nat Biotechnol. 2018;36(8):707–716. doi:10.1038/nbt.4181
70. Zakeri N, Hall A, Swadling L, et al. Characterisation and induction of tissue-resident gamma delta T-cells to target hepatocellular carcinoma. Nat Commun. 2022;13(1):1372. doi:10.1038/s41467-022-29012-1
71. Xu J, Lv J, Zhuang Q, et al. A general strategy towards personalized nanovaccines based on fluoropolymers for post-surgical cancer immunotherapy. Nat Nanotechnol. 2020;15(12):1043–1052. doi:10.1038/s41565-020-00781-4
72. Lee B, Lee K, Panda S, et al. Nanoparticle delivery of CRISPR into the brain rescues a mouse model of fragile X syndrome from exaggerated repetitive behaviours. Nat Biomed Eng. 2018;2(7):497–507. doi:10.1038/s41551-018-0252-8
73. Tian X, Zhang Y, He Z, et al. Successive site translocating inoculation improved T cell responses elicited by a DNA vaccine encoding SARS-CoV-2 S protein. Front Immunol. 2022;13:875236. doi:10.3389/fimmu.2022.875236
74. Song H, Su Q, Nie Y, et al. Supramolecular assembly of a trivalent peptide hydrogel vaccine for cancer immunotherapy. Acta Biomater. 2023;158:535–546. doi:10.1016/j.actbio.2022.12.070
75. Bald T, Krummel MF, Smyth MJ, Barry KC. The NK cell-cancer cycle: advances and new challenges in NK cell-based immunotherapies. Nat Immunol. 2020;21(8):835–847. doi:10.1038/s41590-020-0728-z
76. Wege AK, Weber F, Kroemer A, Ortmann O, Nimmerjahn F, Brockhoff G. IL-15 enhances the anti-tumor activity of trastuzumab against breast cancer cells but causes fatal side effects in humanized tumor mice (HTM). Oncotarget. 2017;8(2):2731–2744. doi:10.18632/oncotarget.13159
77. Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017;27(1):109–118. doi:10.1038/cr.2016.151
78. Knochelmann HM, Dwyer CJ, Bailey SR, et al. When worlds collide: th17 and Treg cells in cancer and autoimmunity. Cell Mol Immunol. 2018;15(5):458–469. doi:10.1038/s41423-018-0004-4
79. Ohue Y, Nishikawa H.Regulatory T (Treg) cells in cancer: can Treg cells be a new therapeutic target?. Cancer Sci. 2019;110(7):2080–2089. doi:10.1111/cas.14069
80. Tanaka A, Sakaguchi S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019;49(8):1140–1146. doi:10.1002/eji.201847659
81. Liu G, Bi Y, Xue L, et al. Dendritic cell SIRT1-HIF1alpha axis programs the differentiation of CD4+ T cells through IL-12 and TGF-beta 1. Proc Natl Acad Sci U S A. 2015;112(9):E957–65. doi:10.1073/pnas.1420419112
82. Buder-Bakhaya K, Hassel JC. Biomarkers for clinical benefit of immune checkpoint inhibitor treatment-a review from the melanoma perspective and beyond. Front Immunol. 2018;9:1474. doi:10.3389/fimmu.2018.01474
83. Stier MT, Zhang J, Goleniewska K, et al. IL-33 promotes the egress of group 2 innate lymphoid cells from the bone marrow. J Exp Med. 2018;215(1):263–281. doi:10.1084/jem.20170449
84. Ponzetti M, Ucci A, Puri C, et al. Effects of osteoblast-derived extracellular vesicles on aggressiveness, redox status and mitochondrial bioenergetics of MNNG/HOS osteosarcoma cells. Front Oncol. 2022;12:983254. doi:10.3389/fonc.2022.983254
85. Wei B, Pan J, Yuan R, et al. Polarization of tumor-associated macrophages by nanoparticle-loaded Escherichia coli combined with immunogenic cell death for cancer immunotherapy. Nano Lett. 2021;21(10):4231–4240. doi:10.1021/acs.nanolett.1c00209
86. Zhu Y, Shen R, Vuong I, et al. Multi-step screening of DNA/lipid nanoparticles and co-delivery with siRNA to enhance and prolong gene expression. Nat Commun. 2022;13(1):4282. doi:10.1038/s41467-022-31993-y
87. Liu S, Galat V, Galat Y, Lee YKA, Wainwright D, Wu J. NK cell-based cancer immunotherapy: from basic biology to clinical development. J Hematol Oncol. 2021;14(1):7. doi:10.1186/s13045-020-01014-w
88. Wu SY, Fu T, Jiang YZ, Shao ZM. Natural killer cells in cancer biology and therapy. Mol Cancer. 2020;19(1):120. doi:10.1186/s12943-020-01238-x
89. Biber G, Sabag B, Raiff A, et al. Modulation of intrinsic inhibitory checkpoints using nano-carriers to unleash NK cell activity. EMBO Mol Med. 2022;14(1):e14073. doi:10.15252/emmm.202114073
90. Gong Y, Klein Wolterink RGJ, Wang J, Bos GMJ, Germeraad WTV. Chimeric antigen receptor natural killer (CAR-NK) cell design and engineering for cancer therapy. J Hematol Oncol. 2021;14(1):73. doi:10.1186/s13045-021-01083-5
91. Terren I, Orrantia A, Vitalle J, Zenarruzabeitia O, Borrego F. NK cell metabolism and tumor microenvironment. Front Immunol. 2019;10:2278. doi:10.3389/fimmu.2019.02278
92. Lopez-Soto A, Gonzalez S, Smyth MJ, Galluzzi L. Control of Metastasis by NK Cells. Cancer Cell. 2017;32(2):135–154. doi:10.1016/j.ccell.2017.06.009
93. Sawyer AJ, Garand M, Chaussabel D, Feng CG. Transcriptomic profiling identifies neutrophil-specific upregulation of cystatin F as a marker of acute inflammation in humans. Front Immunol. 2021;12:634119. doi:10.3389/fimmu.2021.634119
94. Chen C, Ai QD, Chu SF, Zhang Z, Chen NH. NK cells in cerebral ischemia. Biomed Pharmacother. 2019;109:547–554. doi:10.1016/j.biopha.2018.10.103
95. Shimasaki N, Jain A, Campana D. NK cells for cancer immunotherapy. Nat Rev Drug Discov. 2020;19(3):200–218. doi:10.1038/s41573-019-0052-1
96. Jacobs SK, Wilson DJ, Kornblith PL, Grimm EA. Interleukin-2 and autologous lymphokine-activated killer cells in the treatment of malignant glioma. Preliminary report. J Neurosurg. 1986;64(5):743–749. doi:10.3171/jns.1986.64.5.0743
97. Xiang Z, Liu Y, Zheng J, et al. Targeted activation of human Vgamma9Vdelta2-T cells controls Epstein-Barr virus-induced B cell lymphoproliferative disease. Cancer Cell. 2014;26(4):565–576. doi:10.1016/j.ccr.2014.07.026
98. Bergmann C, Strauss L, Zeidler R, Lang S, Whiteside TL. Expansion of human T regulatory type 1 cells in the microenvironment of cyclooxygenase 2 overexpressing head and neck squamous cell carcinoma. Cancer Res. 2007;67(18):8865–8873. doi:10.1158/0008-5472.CAN-07-0767
99. Chen YP, Yin JH, Li WF, et al. Single-cell transcriptomics reveals regulators underlying immune cell diversity and immune subtypes associated with prognosis in nasopharyngeal carcinoma. Cell Res. 2020;30(11):1024–1042. doi:10.1038/s41422-020-0374-x
100. Hou L, Tian C, Yan Y, Zhang L, Zhang H, Zhang Z. Manganese-based nanoactivator optimizes cancer immunotherapy via enhancing innate immunity. ACS Nano. 2020;14(4):3927–3940. doi:10.1021/acsnano.9b06111
101. Voli F, Valli E, Lerra L, et al. Intratumoral copper modulates PD-L1 expression and influences tumor immune evasion. Cancer Res. 2020;80(19):4129–4144. doi:10.1158/0008-5472.CAN-20-0471
102. Vulpis E, Loconte L, Peri A, et al. Impact on NK cell functions of acute versus chronic exposure to extracellular vesicle-associated MICA: dual role in cancer immunosurveillance. J Extracell Vesicles. 2022;11(1):e12176. doi:10.1002/jev2.12176
103. Wu X, Yang H, Chen X, et al. Nano-herb medicine and PDT induced synergistic immunotherapy for colon cancer treatment. Biomaterials. 2021;269:120654. doi:10.1016/j.biomaterials.2021.120654
104. Perozziello G, La Rocca R, Cojoc G, et al. Microfluidic devices modulate tumor cell line susceptibility to NK cell recognition. Small. 2012;8(18):2886–2894. doi:10.1002/smll.201200160
105. Jiang CT, Chen KG, Liu A, et al. Immunomodulating nano-adaptors potentiate antibody-based cancer immunotherapy. Nat Commun. 2021;12(1):1359. doi:10.1038/s41467-021-21497-6
106. Au KM, Park SI, Wang AZ. Trispecific natural killer cell nanoengagers for targeted chemoimmunotherapy. Sci Adv. 2020;6(27):eaba8564. doi:10.1126/sciadv.aba8564
107. Deng G, Sun Z, Li S, et al. Cell-membrane immunotherapy based on natural killer cell membrane coated nanoparticles for the effective inhibition of primary and abscopal tumor growth. ACS Nano. 2018;12(12):12096–12108. doi:10.1021/acsnano.8b05292
108. Lin X, Li F, Gu Q, et al. Gold-seaurchin based immunomodulator enabling photothermal intervention and alphaCD16 transfection to boost NK cell adoptive immunotherapy. Acta Biomater. 2022;146:406–420. doi:10.1016/j.actbio.2022.04.029
109. Deng M, Guo R, Zang S, et al. pH-triggered copper-free click reaction-mediated micelle aggregation for enhanced tumor retention and elevated immuno-chemotherapy against melanoma. ACS Appl Mater Interfaces. 2021;13(15):18033–18046. doi:10.1021/acsami.1c02567
110. Liu JQ, Zhang C, Zhang X, et al. Intratumoral delivery of IL-12 and IL-27 mRNA using lipid nanoparticles for cancer immunotherapy. J Control Release. 2022;345:306–313. doi:10.1016/j.jconrel.2022.03.021
111. Myers JA, Miller JS. Exploring the NK cell platform for cancer immunotherapy. Nat Rev Clin Oncol. 2021;18(2):85–100. doi:10.1038/s41571-020-0426-7
112. Fang F, Xie S, Chen M, et al. Advances in NK cell production. Cell Mol Immunol. 2022;19(4):460–481. doi:10.1038/s41423-021-00808-3
113. Terren I, Orrantia A, Astarloa-Pando G, Amarilla-Irusta A, Zenarruzabeitia O, Borrego F. Cytokine-induced memory-like NK cells: from the basics to clinical applications. Front Immunol. 2022;13:884648. doi:10.3389/fimmu.2022.884648
114. Zhu P, Lu T, Wu J, et al. Gut microbiota drives macrophage-dependent self-renewal of intestinal stem cells via niche enteric serotonergic neurons. Cell Res. 2022;32(6):555–569. doi:10.1038/s41422-022-00645-7
115. Zhang R, Feng L, Dong Z, et al. Glucose & oxygen exhausting liposomes for combined cancer starvation and hypoxia-activated therapy. Biomaterials. 2018;162:123–131. doi:10.1016/j.biomaterials.2018.02.004
116. Zhang D, Xu Z, Tao T, et al. Retraction notice to” Modification of TAK1 by O-linked N-acetylglucosamine facilitates TAK1 activation and promotes M1 macrophage polarization. Cell Signal. 2017;36(36):267. doi:10.1016/j.cellsig.2017.05.016
117. Genova C, Dellepiane C, Carrega P, et al. Therapeutic implications of tumor microenvironment in lung cancer: focus on immune checkpoint blockade. Front Immunol. 2021;12:799455. doi:10.3389/fimmu.2021.799455
118. Jiao S, Subudhi SK, Aparicio A, et al. Differences in tumor microenvironment dictate T helper lineage polarization and response to immune checkpoint therapy. Cell. 2019;179(5):1177–1190 e13. doi:10.1016/j.cell.2019.10.029
119. Kubelka CF, Ruppel A, Gemsa D, Krammer PH. In vivo activation of macrophages by T cell-derived lymphokines: killing of tumor cells and schistosomula of Schistosoma mansoni. Immunobiology. 1986;171(4–5):311–319. doi:10.1016/S0171-2985(86)80063-8
120. Guo Q, Li X, Zhou W, et al. Sequentially triggered bacterial outer membrane vesicles for macrophage metabolism modulation and tumor metastasis suppression. ACS Nano. 2021;15(8):13826–13838. doi:10.1021/acsnano.1c05613
121. Czimmerer Z, Halasz L, Daniel B, et al. The epigenetic state of IL-4-polarized macrophages enables inflammatory cistromic expansion and extended synergistic response to TLR ligands. Immunity. 2022;55(11):2006–2026 e6. doi:10.1016/j.immuni.2022.10.004
122. Wang H, Ding T, Guan J, et al. Interrogation of folic acid-functionalized nanomedicines: the regulatory roles of plasma proteins reexamined. ACS Nano. 2020;14(11):14779–14789. doi:10.1021/acsnano.0c02821
123. Fei Y, Wang Z, Huang M, et al. MiR −155 regulates M2 polarization of hepatitis B virus-infected tumour-associated macrophages which in turn regulates the malignant progression of hepatocellular carcinoma. J Viral Hepat. 2023;30:417–426. doi:10.1111/jvh.13809
124. Tirella A, Kloc-Muniak K, Good L, et al. CD44 targeted delivery of siRNA by using HA-decorated nanotechnologies for KRAS silencing in cancer treatment. Int J Pharm. 2019;561:114–123. doi:10.1016/j.ijpharm.2019.02.032
125. Yang Y, Liu X, Ma W, et al. Light-activatable liposomes for repetitive on-demand drug release and immunopotentiation in hypoxic tumor therapy. Biomaterials. 2021;265:120456. doi:10.1016/j.biomaterials.2020.120456
126. Li K, Lin C, He Y, et al. Engineering of cascade-responsive nanoplatform to inhibit lactate efflux for enhanced tumor chemo-immunotherapy. ACS Nano. 2020;14(10):14164–14180. doi:10.1021/acsnano.0c07071
127. Rong L, Zhang Y, Li WS, Su Z, Fadhil JI, Zhang C. Iron chelated melanin-like nanoparticles for tumor-associated macrophage repolarization and cancer therapy. Biomaterials. 2019;225:119515. doi:10.1016/j.biomaterials.2019.119515
128. Song M, Liu T, Shi C, Zhang X, Chen X. Bioconjugated manganese dioxide nanoparticles enhance chemotherapy response by priming tumor-associated macrophages toward m1-like phenotype and attenuating tumor hypoxia. ACS Nano. 2016;10(1):633–647. doi:10.1021/acsnano.5b06779
129. Ramesh A, Kumar S, Nandi D, Kulkarni A. CSF1R- and SHP2-inhibitor-loaded nanoparticles enhance cytotoxic activity and phagocytosis in tumor-associated macrophages. Adv Mater. 2019;31(51):e1904364. doi:10.1002/adma.201904364
130. Abdel-Bar HM, Walters AA, Lim Y, et al. An ”eat me” combinatory nano-formulation for systemic immunotherapy of solid tumors. Theranostics. 2021;11(18):8738–8754. doi:10.7150/thno.56936
131. Li W, Wang F, Guo R, Bian Z, Song Y. Targeting macrophages in hematological malignancies: recent advances and future directions. J Hematol Oncol. 2022;15(1):110. doi:10.1186/s13045-022-01328-x
132. Liu M, Liu J, Liang Z, et al. CAR-macrophages and CAR-T cells synergistically kill tumor cells in vitro. Cells. 2022;11(22):3692. doi:10.3390/cells11223692
133. Chang DK, Peterson E, Sun J, et al. Anti-CCR4 monoclonal antibody enhances antitumor immunity by modulating tumor-infiltrating Tregs in an ovarian cancer xenograft humanized mouse model. Oncoimmunology. 2016;5(3):e1090075. doi:10.1080/2162402X.2015.1090075
134. Sauer T, Parikh K, Sharma S, et al. CD70-specific CAR T cells have potent activity against acute myeloid leukemia without HSC toxicity. Blood. 2021;138(4):318–330. doi:10.1182/blood.2020008221
135. Albinger N, Pfeifer R, Nitsche M, et al. Primary CD33-targeting CAR-NK cells for the treatment of acute myeloid leukemia. Blood Cancer J. 2022;12(4):61. doi:10.1038/s41408-022-00660-2
136. Schepisi G, Gianni C, Palleschi M, et al. The new frontier of immunotherapy: chimeric antigen receptor T (CAR-T) Cell and Macrophage (CAR-M) therapy against breast cancer. Cancers (Basel). 2023;15(5):1597. doi:10.3390/cancers15051597
137. Tang Y, Yin H, Zhao X, et al. High efficacy and safety of CD38 and BCMA bispecific CAR-T in relapsed or refractory multiple myeloma. J Exp Clin Cancer Res. 2022;41(1):2. doi:10.1186/s13046-021-02214-z
138. Chen Y, Yu Z, Tan X, et al. CAR-macrophage: a new immunotherapy candidate against solid tumors. Biomed Pharmacother. 2021;139:111605. doi:10.1016/j.biopha.2021.111605
139. Vitale I, Manic G, Coussens LM, Kroemer G, Galluzzi L. Macrophages and metabolism in the tumor microenvironment. Cell Metab. 2019;30(1):36–50. doi:10.1016/j.cmet.2019.06.001
140. Rothlin CV, Carrera-Silva EA, Bosurgi L, Ghosh S. TAM receptor signaling in immune homeostasis. Annu Rev Immunol. 2015;33:355–391. doi:10.1146/annurev-immunol-032414-112103
141. Lemke G, Rothlin CV. Immunobiology of the TAM receptors. Nat Rev Immunol. 2008;8(5):327–336. doi:10.1038/nri2303
142. Logtenberg MEW, Scheeren FA, Schumacher TN. The CD47-SIRPalpha Immune Checkpoint. Immunity. 2020;52(5):742–752. doi:10.1016/j.immuni.2020.04.011
143. Veillette A, Chen J. SIRPalpha-CD47 immune checkpoint blockade in anticancer therapy. Trends Immunol. 2018;39(3):173–184. doi:10.1016/j.it.2017.12.005
144. Klichinsky M, Ruella M, Shestova O, et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat Biotechnol. 2020;38(8):947–953. doi:10.1038/s41587-020-0462-y
145. Pereira BA, Vennin C, Papanicolaou M, et al. CAF subpopulations: a new reservoir of stromal targets in pancreatic cancer. Trends Cancer. 2019;5(11):724–741. doi:10.1016/j.trecan.2019.09.010
146. Bhattacharjee S, Hamberger F, Ravichandra A, et al.. Tumor restriction by type I collagen opposes tumor-promoting effects of cancer-associated fibroblasts. J Clin Invest. 2021;131(11). doi:10.1172/JCI146987
147. Nallasamy P, Nimmakayala RK, Karmakar S, et al. Pancreatic tumor microenvironment factor promotes cancer stemness via SPP1-CD44 axis. Gastroenterology. 2021;161(6):1998–2013 e7. doi:10.1053/j.gastro.2021.08.023
148. Fiori ME, Di Franco S, Villanova L, Bianca P, Stassi G, De Maria R. Cancer-associated fibroblasts as abettors of tumor progression at the crossroads of EMT and therapy resistance. Mol Cancer. 2019;18(1):70. doi:10.1186/s12943-019-0994-2
149. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
150. Kalluri R. Basement membranes: structure, assembly and role in tumour angiogenesis. Nat Rev Cancer. 2003;3(6):422–433. doi:10.1038/nrc1094
151. Dominguez CX, Muller S, Keerthivasan S, et al. Single-Cell RNA sequencing reveals stromal evolution into LRRC15(+) myofibroblasts as a determinant of patient response to cancer immunotherapy. Cancer Discov. 2020;10(2):232–253. doi:10.1158/2159-8290.CD-19-0644
152. Bernard V, Semaan A, Huang J, et al. Single-cell transcriptomics of pancreatic cancer precursors demonstrates epithelial and microenvironmental heterogeneity as an early event in neoplastic progression. Clin Cancer Res. 2019;25(7):2194–2205. doi:10.1158/1078-0432.CCR-18-1955
153. Elyada E, Bolisetty M, Laise P, et al. Cross-species single-cell analysis of pancreatic ductal adenocarcinoma reveals antigen-presenting cancer-associated fibroblasts. Cancer Discov. 2019;9(8):1102–1123. doi:10.1158/2159-8290.CD-19-0094
154. Hosein AN, Huang H, Wang Z, et al.. Cellular heterogeneity during mouse pancreatic ductal adenocarcinoma progression at single-cell resolution. JCI Insight. 2019;5:16.
155. Peng J, Sun BF, Chen CY, et al. Single-cell RNA-seq highlights intra-tumoral heterogeneity and malignant progression in pancreatic ductal adenocarcinoma. Cell Res. 2019;29(9):725–738. doi:10.1038/s41422-019-0195-y
156. Zhao XX, Li LL, Zhao Y, et al. In situ self-assembled nanofibers precisely target cancer-associated fibroblasts for improved tumor imaging. Angew Chem Int Ed Engl. 2019;58(43):15287–15294. doi:10.1002/anie.201908185
157. Zhang G, He Y, Liu Y, Du Y, Yang C, Gao F. Reduced hyaluronan cross-linking induces breast cancer malignancy in a CAF-dependent manner. Cell Death Dis. 2021;12(6):586. doi:10.1038/s41419-021-03875-6
158. Kim DJ, Dunleavey JM, Xiao L, et al. Suppression of TGFbeta-mediated conversion of endothelial cells and fibroblasts into cancer associated (myo)fibroblasts via HDAC inhibition. Br J Cancer. 2018;118(10):1359–1368. doi:10.1038/s41416-018-0072-3
159. Luo X, Fong ELS, Zhu C, et al. Hydrogel-based colorectal cancer organoid co-culture models. Acta Biomater. 2021;132:461–472. doi:10.1016/j.actbio.2020.12.037
160. Kozakowski N, Hartmann C, Klingler HC, et al. Immunohistochemical expression of PDGFR, VEGF-C, and proteins of the mToR pathway before and after androgen deprivation therapy in prostate carcinoma: significant decrease after treatment. Target Oncol. 2014;9(4):359–366. doi:10.1007/s11523-013-0298-1
161. Harryvan TJ, Hawinkels L, Mini-tumor W, et al. A novel pancreatic cancer mini-tumor model to study desmoplasia and myofibroblastic cancer-associated fibroblast differentiation. Gastro Hep Adv. 2022;1(4):678–681. doi:10.1016/j.gastha.2022.04.019
162. Liu G, Sun Z, Fu Z, Ma L, Wang X. Temperature sensing and bio-imaging applications based on polyethylenimine/CaF(2) nanoparticles with upconversion fluorescence. Talanta. 2017;169:181–188. doi:10.1016/j.talanta.2017.03.054
163. Karthickraja D, Kumar GA, Sardar DK, et al. Fabrication of Nd(3+) and Yb(3+) doped NIR emitting nano fluorescent probe: a candidate for bioimaging applications. Mater Sci Eng C Mater Biol Appl. 2021;125:112095. doi:10.1016/j.msec.2021.112095
164. Rizvi S, Gores GJ. Pathogenesis, diagnosis, and management of cholangiocarcinoma. Gastroenterology. 2013;145(6):1215–1229. doi:10.1053/j.gastro.2013.10.013
165. Kieffer Y, Hocine HR, Gentric G, et al. Single-cell analysis reveals fibroblast clusters linked to immunotherapy resistance in cancer. Cancer Discov. 2020;10(9):1330–1351. doi:10.1158/2159-8290.CD-19-1384
166. Feng J, Xu M, Wang J, et al. Sequential delivery of nanoformulated alpha-mangostin and triptolide overcomes permeation obstacles and improves therapeutic effects in pancreatic cancer. Biomaterials. 2020;241:119907. doi:10.1016/j.biomaterials.2020.119907
167. Zhao J, Wang H, Hsiao CH, et al. Simultaneous inhibition of hedgehog signaling and tumor proliferation remodels stroma and enhances pancreatic cancer therapy. Biomaterials. 2018;159:215–228. doi:10.1016/j.biomaterials.2018.01.014
168. Yu Q, Qiu Y, Li J, et al. Targeting cancer-associated fibroblasts by dual-responsive lipid-albumin nanoparticles to enhance drug perfusion for pancreatic tumor therapy. J Control Release. 2020;321:564–575. doi:10.1016/j.jconrel.2020.02.040
169. Young A, Ngiow SF, Barkauskas DS, et al. Co-inhibition of CD73 and A2AR Adenosine signaling improves anti-tumor immune responses. Cancer Cell. 2016;30(3):391–403. doi:10.1016/j.ccell.2016.06.025
170. Allard B, Pommey S, Smyth MJ, Stagg J. Targeting CD73 enhances the antitumor activity of anti-PD-1 and anti-CTLA-4 mAbs. Clin Cancer Res. 2013;19(20):5626–5635. doi:10.1158/1078-0432.CCR-13-0545
171. Francescone R, Barbosa Vendramini-Costa D, Franco-Barraza J, et al. Netrin G1 promotes pancreatic tumorigenesis through cancer-associated fibroblast-driven nutritional support and immunosuppression. Cancer Discov. 2021;11(2):446–479. doi:10.1158/2159-8290.CD-20-0775
172. Ogier C, Colombo PE, Bousquet C, et al. Targeting the NRG1/HER3 pathway in tumor cells and cancer-associated fibroblasts with an anti-neuregulin 1 antibody inhibits tumor growth in pre-clinical models of pancreatic cancer. Cancer Lett. 2018;432:227–236. doi:10.1016/j.canlet.2018.06.023
173. Peran I, Dakshanamurthy S, McCoy MD, et al. Cadherin 11 promotes immunosuppression and extracellular matrix deposition to support growth of pancreatic tumors and resistance to gemcitabine in mice. Gastroenterology. 2021;160(4):1359–1372 e13. doi:10.1053/j.gastro.2020.11.044
174. Zhang Z, Karthaus WR, Lee YS, et al. Tumor microenvironment-derived NRG1 promotes antiandrogen resistance in prostate cancer. Cancer Cell. 2020;38(2):279–296 e9. doi:10.1016/j.ccell.2020.06.005
175. Purcell JW, Tanlimco SG, Hickson J, et al. LRRC15 is a novel mesenchymal protein and stromal target for antibody-drug conjugates. Cancer Res. 2018;78(14):4059–4072. doi:10.1158/0008-5472.CAN-18-0327
176. Cui J, Dean D, Wei R, Hornicek FJ, Ulmert D, Duan Z. Expression and clinical implications of leucine-rich repeat containing 15 (LRRC15) in osteosarcoma. J Orthop Res. 2020;38(11):2362–2372. doi:10.1002/jor.24848
177. Kato M, Placencio-Hickok VR, Madhav A, et al. Heterogeneous cancer-associated fibroblast population potentiates neuroendocrine differentiation and castrate resistance in a CD105-dependent manner. Oncogene. 2019;38(5):716–730. doi:10.1038/s41388-018-0461-3
178. Hutton C, Heider F, Blanco-Gomez A, et al. Single-cell analysis defines a pancreatic fibroblast lineage that supports anti-tumor immunity. Cancer Cell. 2021;39(9):1227–1244 e20. doi:10.1016/j.ccell.2021.06.017
179. Mezawa Y, Daigo Y, Takano A, et al. CD26 expression is attenuated by TGF-beta and SDF-1 autocrine signaling on stromal myofibroblasts in human breast cancers. Cancer Med. 2019;8(8):3936–3948. doi:10.1002/cam4.2249
180. Helms EJ, Berry MW, Chaw RC, et al. Mesenchymal lineage heterogeneity underlies nonredundant functions of pancreatic cancer-associated fibroblasts. Cancer Discov. 2022;12(2):484–501. doi:10.1158/2159-8290.CD-21-0601
181. Saw PE, Chen J, Song E. Targeting CAFs to overcome anticancer therapeutic resistance. Trends Cancer. 2022;8(7):527–555. doi:10.1016/j.trecan.2022.03.001
182. Papait A, Romoli J, Stefani FR, et al. Fight the Cancer, Hit the CAF!. Cancers (Basel). 2022;14(15):3570. doi:10.3390/cancers14153570
183. Wu F, Yang J, Liu J, et al. Signaling pathways in cancer-associated fibroblasts and targeted therapy for cancer. Signal Transduct Target Ther. 2021;6(1):218. doi:10.1038/s41392-021-00641-0
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.