Back to Journals » Drug Design, Development and Therapy » Volume 20
Evidence Based on Sleep Disorder Exacerbating Postoperative Pain via Kynureninase
Authors Huang N, Chen Y, Yang X, Tao C, Zhang Y, Yang Y, Shu B, Duan G ![]() , Chen Y
, Chen Y
Received 11 October 2025
Accepted for publication 19 January 2026
Published 5 February 2026 Volume 2026:20 565401
DOI https://doi.org/10.2147/DDDT.S565401
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leonidas D. Panos
Niannian Huang,* Yongqin Chen,* Xinqing Yang, Chengkun Tao, Yifan Zhang, Yi Yang, Bin Shu, Guangyou Duan, Yuanjing Chen
Department of Anaesthesiology, The Second Affiliated Hospital, Chongqing Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Guangyou Duan, Email [email protected] Yuanjing Chen, Email [email protected]
Background: Surgery is a frequently employed intervention for patients; however, 30– 80% of patients report moderate to severe pain after surgery. Postoperative pain is affected by a variety of factors, with sleep disturbance being a critical modulating factor that exacerbates postoperative pain intensity. Kynureninase (KYNU), a key hydrolase in the kynurenine pathway that produces quinolinic acid, may link sleep disturbances to pain sensitization; however, the underlying mechanisms remain unclear.
Methods: We obtained skin and blood samples from 40 abdominal surgery patients and performed whole-transcriptome sequencing. Through analysis of the intersecting gene sets between patients with adequate and inadequate analgesia, as well as between those with sleep disorder and normal sleep, we identified genes significantly linked to both sleep disorders and postoperative pain. We established an animal model by making a hind paw plantar incision in mice subjected to chronic sleep deprivation. We then performed behavioral tests, including the tail flick test and von Frey filaments, measured the expression of differentially expressed genes via real-time quantitative polymerase chain reaction and immunofluorescence, and validated the results using pharmacological and calcium imaging methods.
Results: We found that sleep deprivation greatly exacerbated postoperative pain, and KYNU was notably higher in the skin, blood, and dorsal root ganglion (DRG). Repeated intraperitoneal injections of carbidopa, a KYNU modulator, reduced the severity of pain induced by sleep deprivation and abnormal excitability of DRG neurons. Furthermore, in preclinical models, carbidopa treatment effectively reduced both inflammatory and neuropathic pain.
Conclusion: We conclude that the upregulated expression of KYNU is associated with the exacerbation of postoperative pain due to sleep deprivation and identified the compound carbidopa as possessing significant analgesic potential.
Keywords: postoperative pain, sleep disorders, kynureninase, neural excitability, carbidopa
Introduction
The global surgical burden is estimated to involve approximately 310 million procedures annually.1 Despite advances in perioperative care, 30–80% of patients still report moderate-to-severe postoperative pain.2,3 Inadequate postoperative pain management not only hinders functional recovery and adversely affects health-related quality of life but may also contribute to the development of persistent postsurgical pain syndrome, thereby imposing a significant socioeconomic burden.4 Contemporary perioperative analgesic strategies rely heavily on opioids and other systemic agents, but their use is frequently limited by adverse effects such as respiratory depression, gastrointestinal dysmotility, and the risk of prolonged opioid use and dependence.5,6 Consequently, elucidating the molecular mechanisms that exacerbate postoperative hyperalgesia is critical for identifying novel, non-opioid therapeutic targets.
Among the various established risk factors for poor postoperative pain outcomes, sleep disturbance is a critical modulating factor that exacerbates postoperative pain intensity. Clinical evidence indicates a substantial association between sleep disorders and insufficient postoperative pain management. A systematic review and meta-analysis of 29 cohort studies with 16,104 participants found that preoperative significant insomnia symptoms were linked to moderate to severe pain on the first postoperative day (odds ratio 2.69, 95% confidence interval 2.03–3.57, P < 0.0001).7 Numerous studies have demonstrated that preoperative sleep disorders not only exacerbate acute postoperative pain and hinder functional recovery, but also significantly elevate the risk of developing chronic postsurgical pain syndrome.8–10 While the clinical association is well-established, the molecular bridge translating sleep loss into pain hypersensitivity remains incompletely characterized.
Emerging evidence implicates the kynurenine pathway (KP) of tryptophan metabolism as a key mediator in this process. Approximately 90–95% of dietary tryptophan is degraded via the KP to generate a spectrum of bioactive metabolites, including kynurenic acid and quinolinic acid (QUIN).11,12 QUIN is a selective NMDA receptor agonist with well-established neurotoxic and pronociceptive properties; elevated QUIN levels have been linked to hyperalgesia, neuropathic pain, and excitotoxic neuronal injury.13–15 Within this pathway, KYNU is a pyridoxal-5’-phosphate-dependent hydrolase that catalyzes the conversion of kynurenine and 3-hydroxykynurenine (3-HK) to anthranilic acid and 3-hydroxyanthranilic acid (3-HAA), positioning it upstream of QUIN biosynthesis.11,16 KYNU is upregulated in several inflammatory conditions, including psoriasis, where increased expression correlates with local immune activation.16 In neuropathic pain models, enhanced expression of KP enzymes such as kynurenine 3-monooxygenase (KMO), KYNU, and 3-hydroxyanthranilate 3,4-dioxygenase has been observed in central neurons, accompanied by an increased QUIN-to-kynurenine ratio and pain- and depression-like behaviors.17–19 These findings support the concept that increased KYNU activity can shift KP flux toward QUIN and other neurotoxic metabolites, thereby augmenting neuronal excitability and facilitating central sensitization. Nevertheless, the contribution of KYNU to the link between sleep disturbance and postoperative pain has not been elucidated.
Carbidopa is widely used in combination with levodopa as a peripheral aromatic L-amino acid decarboxylase inhibitor in the treatment of Parkinson’s disease. Beyond this established indication, biochemical and pharmacological studies have shown that carbidopa can inhibit tryptophan 2,3-dioxygenase and KYNU, thereby altering KP flux and the profile of downstream metabolites.20,21 In addition, pharmacological targeting of KYNU with carbidopa has been reported to attenuate tissue damage and inflammatory responses in models of radiation-induced intestinal injury, highlighting KYNU inhibition as a feasible strategy to modulate stress- and inflammation-driven KP activation.22 Although these findings support the rationale for repurposing carbidopa as a KYNU inhibitor, its efficacy in modulating sleep-induced pain via this mechanism has not been explored.
To investigate this, we combined clinical transcriptomics with basic mechanistic research. We hypothesized that sleep disturbance exacerbates postoperative pain through the upregulation of KYNU and subsequent NMDA-mediated neuronal excitability, and that this process is reversible via carbidopa treatment. We performed whole-transcriptome sequencing on tissues from patients with sleep disorders to validate KYNU upregulation. In animal models, we utilized sleep deprivation paradigms to assess the impact of KYNU activity on pain thresholds. We investigated the cellular mechanisms by assessing the effects of KYNU modulation on neuronal excitability using calcium imaging in primary neurons. Finally, we evaluated the therapeutic potential of repurposing carbidopa to modulate KYNU and attenuate sleep deprivation-induced postoperative hyperalgesia. These findings aim to clarify the mechanistic link between sleep disturbance and postoperative pain and to establish KYNU as a potential therapeutic target for optimizing perioperative analgesia.
Methods
Participants and Ethics
This cross-sectional study was conducted in accordance with the Declaration of Helsinki and was approved by the Institutional Ethics Committee of the Second Affiliated Hospital of Chongqing Medical University (approval ID: 2022–93). Written informed consent was obtained from all the patients. We screened 40 patients who underwent abdominal surgery and were eligible for inclusion. The inclusion criteria comprised: aged ≥65 years, comprehends informed consent, and no cognitive impairment. Patients who declined to sign the consent forms were excluded.
Clinical Procedures
One day before surgery, participants were instructed to use the visual analog scale (VAS) at rest (ranging from 0 to 10, where 0 indicates no pain and 10 signifies the worst and most intolerable pain) and complete the Pittsburgh Sleep Quality Index (PSQI). For the clinical transcriptomic analysis, peripheral blood samples were collected from older patients (≥ 65 years) undergoing elective abdominal surgery. Blood was drawn after induction of anesthesia. Preoperatively, patients were classified into a chronic insomnia group or a normal sleep group according to predefined clinical criteria. Postoperatively, pain intensity was assessed using the VAS during postoperative days 1–3; patients who reported an VAS ≥ 4 at any time point were classified as the postoperative pain group, whereas those with VAS < 4 throughout this period were classified as the no‑pain group. Each patient therefore contributed to two comparisons: (1) sleep disturbance (SD) versus normal sleep (NS) and (2) inadequate analgesia (IA) versus adequate analgesia (AA). For the SD_vs_NS comparison, 25 samples were included in the SD group and 15 in the NS group. For the IA_vs_AA comparison, 11 samples were included in the IA group and 29 in the AA group. The sample size was determined by the number of eligible patients with high‑quality RNA within the study period and is comparable to previous exploratory RNA‑seq studies in similar clinical settings; no formal a priori power calculation was performed because this analysis was designed as a hypothesis‑generating discovery study.
RNA Extraction, Library Preparation, Sequencing and Data Processing
Total RNA was extracted from whole blood using TRIzol reagent (Invitrogen, USA) according to the manufacturer’s instructions. RNA purity and integrity were assessed by NanoPhotometer spectrophotometry and an Agilent 2100 Bioanalyzer. For each sample, 1 µg of total RNA was used to generate sequencing libraries with the NEBNext Ultra RNA Library Prep Kit for Illumina (New England Biolabs, USA), followed by poly(A)+ mRNA enrichment, cDNA synthesis, adaptor ligation and PCR amplification. Indexed libraries were pooled and sequenced on an Illumina NovaSeq platform to obtain 150 bp paired‑end reads.
Raw reads were quality‑filtered with fastp (v0.20.0), and clean reads were aligned to the human reference genome using HISAT2. Gene‑level read counts were obtained with featureCounts, and FPKM values were estimated with StringTie for downstream differential expression analysis and visualization.
Differential Expression and Multiple-Testing Correction
Differentially expressed genes (DEGs) between SD and NS, and between IA and AA, were identified using DESeq2 in R on the raw count matrix. Lowly expressed genes were filtered out according to DESeq2 recommendations before model fitting. Genes were considered differentially expressed if they met both of the following criteria: (i) P value < 0.05 and (ii) log2 fold change>0. These thresholds were used to generate the volcano plots shown in the main figures. Overlaps of DEGs between different comparisons (eg, SD_vs_NS and IA_vs_AA) were visualized using Venn diagrams based on the same DEG definition.
Animals
Male C57BL/6J mice, aged 8 to 10 weeks, were procured from SLNAC Experimental Animal Technology Co. in Shanghai, China. The mice were maintained under standard laboratory conditions, which included a 12-hour light/dark cycle, and were given unrestricted access to food and water. Animal procedures followed the ARRIVE Guidelines 2.0 and the Guide for the Care and Use of Laboratory Animals (8th edition, National Research Council, 2011). The study protocol was approved by the Animal Research Committee of the Second Affiliated Hospital of Chongqing Medical University (approval no. IACUC-SAHCQMU-2024-00168). Euthanasia was performed in accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals.
Chronic Sleep Deprivation (CSD)
Following a previous methodology,23 we kept the mice awake by periodically introducing or replacing novel objects, such as blocks, balls, and tunnels, to engage their natural exploratory behavior. The sleep intervention was initiated at the onset of the light cycle (zeitgeber time 0, ZT0) and continued for 6 h (ZT0-6). Following the sleep intervention, all new objects were removed, enabling the mice to rest and sleep freely in their usual housing environments. The sleep deprivation protocol was systematically implemented over 7 consecutive days, consistently during the same time period (ZT0-6). During the experiment, the behavioral states of the mice were continuously monitored to ensure that they remained awake without displaying significant stress responses.
Plantar Incision (PINC)
A PINC was made as previously described.24 Briefly, mice were anesthetized with 1.5–2% isoflurane, and the plantar skin of the right hind paw was sterilized with povidone-iodine followed by 75% ethanol. A 0.5 cm longitudinal incision was made on the plantar surface. The plantar muscles were exposed, separated, and incised longitudinally. Subsequently, the plantar muscle was elevated for 10s using microcurved forceps and repeated every 5 min for a total duration of 1 h. Following completion of the procedure, the muscle and skin were sutured with 6–0 suture material.
Spared Nerve Injury (SNI) Model
We performed a model of neuropathic pain using SNI as previously described.25 Mice were anesthetized with 2% isoflurane. After shaving and disinfecting with povidone-iodine and 75% ethanol, a small incision was made on the lateral left mid-thigh to access the sciatic nerve and its three terminal branches by separating the muscle layers. The tibial and common peroneal nerves were ligated and cut, and the sural nerve was preserved. Following the surgical procedure, the muscle layer and skin were sutured, and the animals were placed on a heated pad to facilitate recovery from anesthesia.
Inflammatory Pain Mode
The carrageenan (Car) model was established as previously described.26 In brief, mice received an intraplantar injection of 5 μL λ-carrageenan (1% w/v, Sigma-Aldrich, the United States) in the right hind paw.
Von Frey Test
Paw withdrawal threshold (PWT) was determined using the up-down method with a series of von Frey filaments, as previously described.27,28 Briefly, the PWT was evaluated using von Frey filaments (0.04, 0.07, 0.16, 0.4, 0.6, 1.0, 1.4, 2.0, and 4.0 g) applied perpendicularly to the plantar surface of the right hind paw for 5 s, with sufficient force to bend the filament slightly. Testing began with a 0.6 g filament using the up-down method. If a positive response (such as rapid paw withdrawal, licking, or shaking) was observed, a subsequent lower-force filament was used. If there was no positive response, the next higher-force filament was tested. After the initial positive response, four effective measurements were taken consecutively at 10s intervals to avoid sensitization. The PWT was calculated using MATLAB software.
Tail Flick Test
Thermal nociceptive thresholds were assessed utilizing the tail flick test, wherein the tail was submerged in water maintained at a temperature of 50°C±0.5°C. The latency period was recorded at the onset of the rapid tail-flick response. A cutoff time of 10s was implemented to prevent potential permanent damage to the tail. Each mouse underwent three measurements spaced at 10 min intervals, and the mean of these three measurements was calculated to determine the pain threshold.
Drug Administration
Carbidopa (CBA) is a non-specific KYNU inhibitor.20,21 CBA (Selleck, United States, 28860–95-9) was diluted in 8.33% DMSO +30% PEG300 +5% Tween80+saline and administered intraperitoneally at a dosage of 30 mg/kg29,30 following a PINC. Two hours after injection, we tested the PWT of the mice.
Immunofluorescence
The L4-L6 DRG were harvested and subsequently fixed in 4% paraformaldehyde at 4°C overnight, and then dehydrated in 30% sucrose for two days. The samples were embedded with optimal cutting temperature compound, and a 20-μm frozen section was created using a cryostat. (RWD, FS800). The sections were washed with phosphate buffered saline (PBS), blocked with 10% goat serum, perforated with 0.5%Triton x-100 at 37°C for 1h, and then incubated with primary antibodies at 4°C overnight. The primary antibodies were used at a 1:500 dilution: rabbit anti-KYNU antibody (Proteintech, United States, 11796-1-AP) and guinea pig anti-NeuN antibody (Sigma-Aldrich, United States, ABN90). The sections were washed with PBS, followed by immunostaining with secondary antibodies for 2 h. Goat anti-Rabbit 488 (Invitrogen, United States, A11034, 1:1000) and Goat anti-Guinea Pig 550 (Invitrogen, United States, SA5-10095) were used as secondary antibodies. Sections were rewashed with PBS and counterstained using DAPI Fluoromount-G (Southern Biotech, 0100–20). Finally, images were captured using a confocal microscope (Olympus, Tokyo, Japan, SpinSR) and analyzed using ImageJ software.
RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
RNA was extracted from the skin and white blood cells using the Eastep Super Total RNA Extraction Kit (Promega, United States, LS1040). The mRNA was reverse-transcribed using the Transcriptor First Strand cDNA-synthesis Kit (Roche, Switzerland, 4897030001). RT-qPCR was performed using a SYBR Green Taq Mix kit (Roche, Switzerland, 06402712001) and quantified using an RT-qPCR system (Bio-Rad, United States). Relative gene expression levels were determined using the comparative cycle threshold method, with GAPDH serving as the reference housekeeping gene for data normalization. The primers used were as follows:31,32 KYNU, F: GGGTTCCAGGTTGGGATGAG, R: GGGCTAACGCTGAGTGCTAA; GAPDH, F: GGCTGCCCAGAACATCAT, R: CGGACACATTGGGGGTAC.
DRG Neuron Culture
Primary DRG cells were extracted as previously described.33 The mice were euthanized, and the DRG from the L4-L6 segments were harvested and placed in a 35-mm tissue culture dish. The tissue was subjected to enzymatic digestion using 3 mg/mL collagenase (Gibco, United States, 17104019) for 30 min at 37°C, followed by treatment with 1.25% trypsin (Gibco, United States, 15090046) for an additional 30 min. The digested tissues were carefully washed with neurobasal medium (Gibco, 10888022) and mechanically dissociated. The cells were seeded on laminin-coated coverslips in 35-mm tissue culture dishes and incubated at 37°C in a 5% CO2 atmosphere. The neurobasal medium used for maintenance was supplemented with 2% B27 (Gibco, 17504044), 1% glutamine (Gibco, 35050061), and 0.5% penicillin-streptomycin.
Calcium Imaging
Cells were washed three times with Ca2+- and Mg2+-free Hanks’ Balanced Salt Solution (HBSS) after 24 h incubation, and then incubated with 4 μM of Fluo4-AM (Yeasen, China, 40704ES772) in the dark for 30 min. Subsequently, the cells were washed three times with HBSS and incubated for a minimum of 20 min prior to imaging. External stimulation was performed using a transient confocal dish containing 2 mM CaCl2. Calcium ion (Ca2+) imaging was performed before, during, and after application of the chemical in the bath. All imaging experiments were performed using a confocal microscope (Olympus, Japan, SpinSR).
Statistical Analysis
Statistical analyses were conducted using SPSS Statistics version 27.0 (IBM, New York, United States) and GraphPad Prism 10 (GraphPad Software, La Jolla, CA, United States). Clinical data are presented as mean ± standard deviation (SD), while preclinical experimental data are expressed as mean ± standard error of the mean (SEM). Categorical variables are presented as frequencies and percentages [n (%)]. Data normality was assessed using the Shapiro–Wilk test. Homogeneity of variance was evaluated using Levene’s test. For normally distributed data with equal variances, comparisons between two groups were performed using Unpaired t t-test, and comparisons among multiple groups were analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. For pain thresholds assessed at multiple time points, two-way repeated measures ANOVA was employed, followed by Tukey’s post hoc test for multiple comparisons. Categorical variables were compared using chi-square test or Fisher’s exact test as appropriate. When data did not meet the assumptions of normality or homogeneity of variance, appropriate non-parametric tests were used: Mann–Whitney U-test for two-group comparisons and Kruskal–Wallis test followed by Dunn’s post hoc test for multiple group comparisons. A P value < 0.05 was considered statistically significant.
Results
Preoperative Sleep Disturbances Exacerbated Postoperative Pain
A flow diagram of the study is shown in Figure 1. We screened a total of 60 patients for eligibility, of whom 40 were enrolled. Ultimately, 40 patients completed the follow-up and were included in the final analysis. Patients with PSQI scores > 5 were classified as having sleep disorders.34 In this study, the prevalence of sleep disorders was approximately 60% (25/40). The demographic data and perioperative characteristics of the patients are shown in Table 1. There were no significant differences in other characteristics between individuals with and without sleep disorders before surgery, except for the body mass index (P = 0.038) and PSQI (P < 0.001). Subsequently, we analyzed the postoperative pain scores at rest, as shown in Figure 2A, which revealed that patients with sleep disorders exhibited elevated pain scores following surgery. Furthermore, the prevalence of moderate to severe pain is 32% among patients with sleep disorders and 20% in those with normal sleep patterns (Figure 2B). While daily VAS scores showed no significant group differences at individual time points (Figure 2C), the maximum VAS (Figure 2A) and area under curve (AUC) (Figure 2D) were significantly higher in the sleep disorder group. This suggests that sleep disturbances primarily affect peak pain intensity and cumulative pain burden rather than pain levels at specific time points. The discrepancy may reflect higher inter-individual variability in daily VAS scores or transient pain spikes captured by maximum VAS but averaged out in daily assessments.
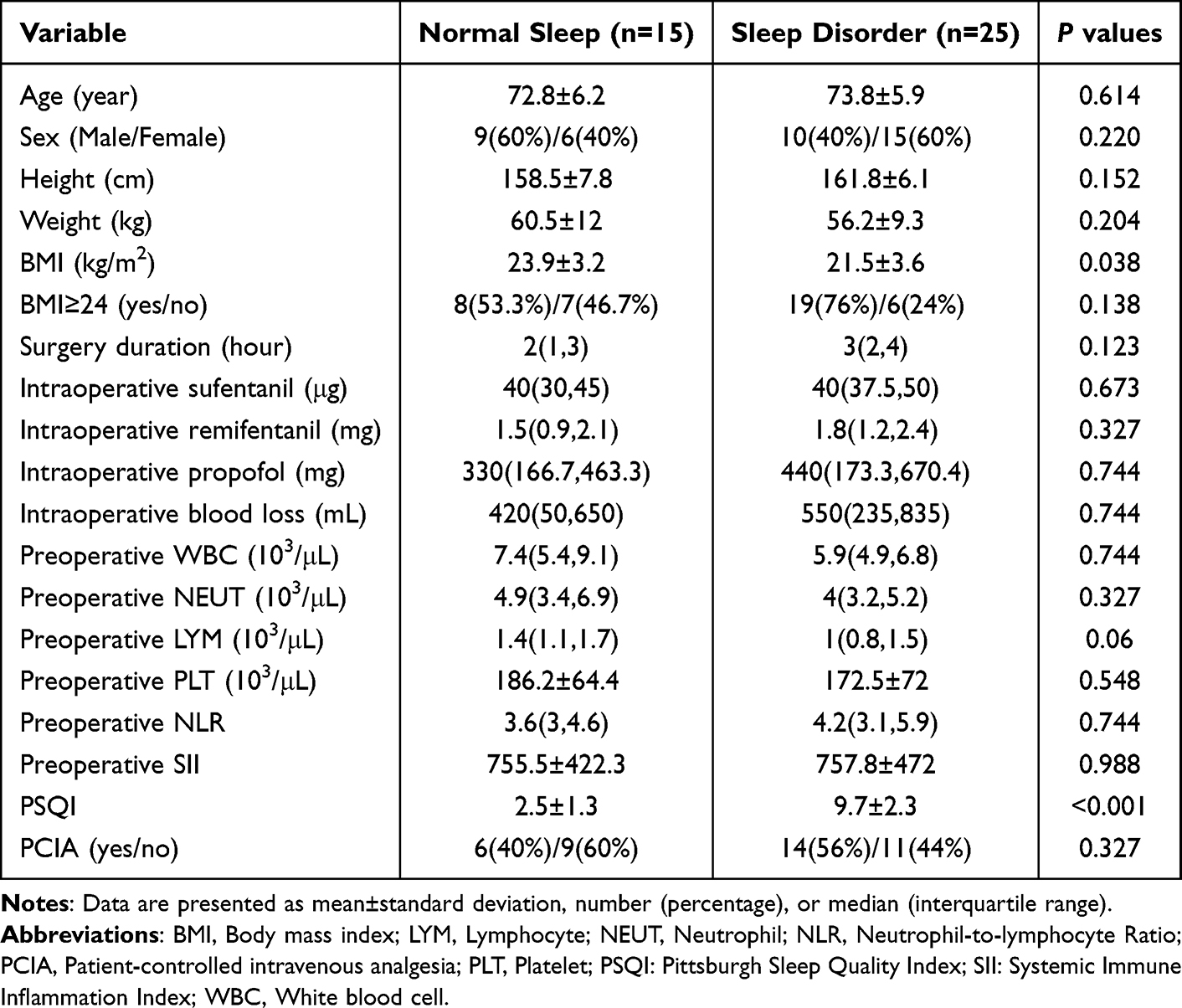 |
Table 1 Demographic Data and Perioperative Characteristics of The Included Patients |
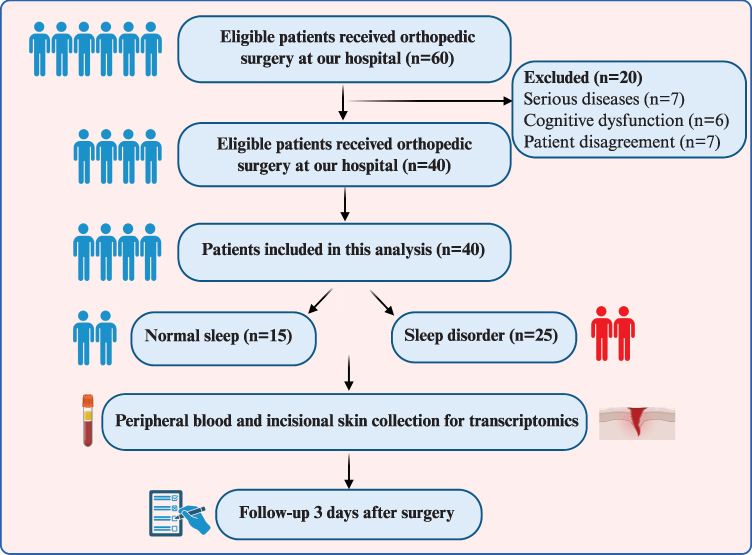 |
Figure 1 Flow chart of the study. |
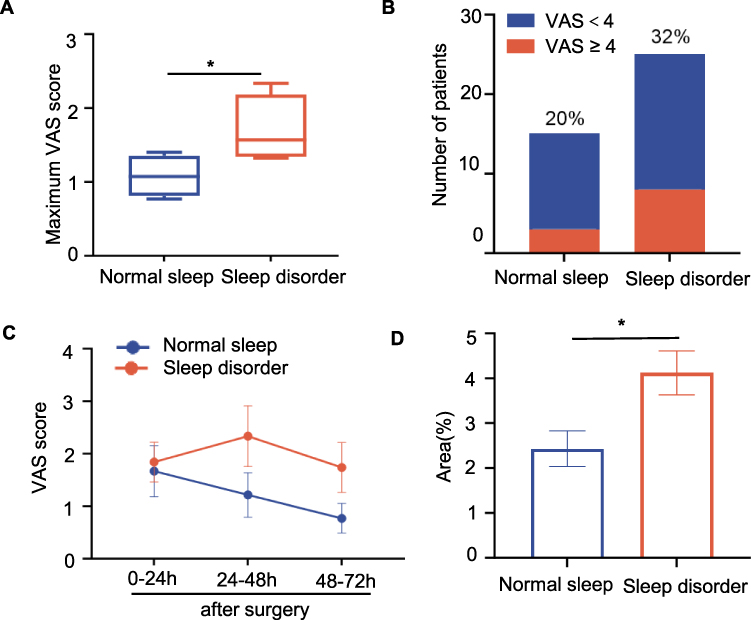 |
Figure 2 Preoperative sleep disturbances exacerbate postoperative pain. (A) Maximum VAS scores were significantly higher in patients with sleep disorders compared to those with normal sleep (P = 0.031, Unpaired t-test). (B) The proportion of patients experiencing moderate-to-severe pain (VAS ≥ 4) showed a numerical trend toward higher incidence in the sleep disorder group (32% vs 20%, P = 0.329, Fisher’s exact test). (C and D) VAS scores on postoperative days 1–3. Two-way mixed ANOVA showed no significant main effect of group (P = 0.1609) or time (P = 0.2540), and no group × time interaction (P = 0.3453), Šídák’s multiple comparisons test showed no significant differences at individual time points. AUC of VAS scores was significantly higher in the sleep disorder group (P = 0.0108, Unpaired t-test). Data are mean ± SEM. *P < 0.05. Abbreviations: AUC, area under the curve; n.s, not significant; VAS, Visual Analog Scale. |
Sleep Deprivation Increased Pain Sensitivity, Aggravated Postoperative Pain, and Prolonged Pain Recovery in Mice
Our preliminary clinical findings showed that patients with sleep disorder experienced severe postoperative pain. To confirm this, we performed animal experiments to evaluate the effects of sleep deprivation on basal and postoperative pain sensitivity (Figure 3A). Compared with the control group, mice exposed to 6 h of daily sleep deprivation exhibited a significant decrease in tail-flick thresholds at both 5 and 7 days (Figure 3B), as well as a reduction in mechanical pain thresholds at 7 days (Figure 3C). Conversely, a regimen of 2 h of daily sleep deprivation did not yield statistically significant effects on thermal or mechanical pain thresholds. Six hours of daily sleep deprivation for 7 consecutive days had the greatest effect on pain thresholds and caused no significant difference in body weight in mice (Figure S1), so we adopted this protocol for all future experiments.
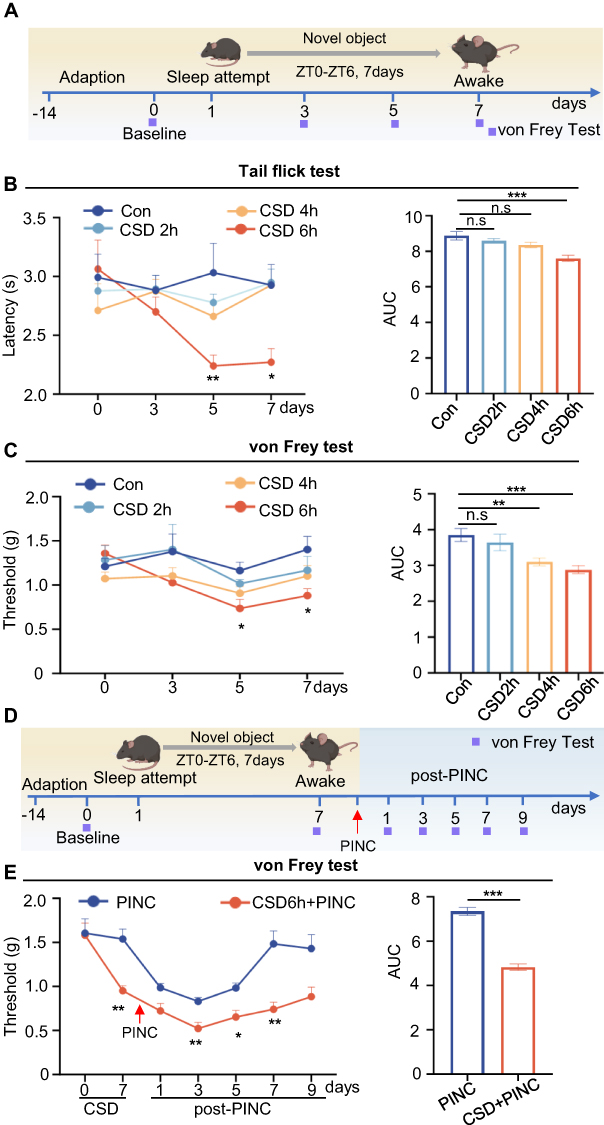 |
Figure 3 Sleep deprivation increases pain sensitivity, aggravates postoperative pain, and prolongs pain recovery in mice. (A) Schematic for CSD and behavioral testing timeline. (B) Thermal nociceptive threshold was measured by tail flick test. Two-way RM ANOVA revealed a significant treatment × time interaction (P = 0.0041, Greenhouse-Geisser corrected). Tukey’s post-hoc tests: CSD 6h vs control on days 5 (P = 0.0024) and 7 (P = 0.0426); CSD6h vs CSD 2h (P = 0.0055) and CSD 4h (P = 0.0029) on day 7. AUC analysis confirmed the dose-dependent effect (P < 0.001, one-way ANOVA): Con vs CSD 6h (P < 0.001). Con: n = 7 mice; CSD 2h: n = 7 mice; CSD 4h: n = 8 mice; CSD 6h: n = 8 mice. (C) Mechanical nociceptive threshold measured using von Frey filaments. Two-way RM ANOVA revealed significant main effects of treatment (P = 0.0309) and time (P = 0.0075, Greenhouse-Geisser corrected). Tukey’s post-hoc tests showed CSD 6h vs control on days 5 (P = 0.0385) and 7 (P = 0.0490). AUC analysis confirmed the hyperalgesic effect (P < 0.001, one-way ANOVA): Con vs CSD 6h (P < 0.001), Con vs CSD 4h (P = 0.0071). Sample sizes as in (B). (D) Schematic for PINC with or without prior CSD. (E) Mechanical nociceptive threshold in PINC model. Two-way RM ANOVA revealed significant main effects of treatment (P < 0.001), time (P < 0.001, Greenhouse-Geisser corrected), and treatment × time interaction (P = 0.0012). Šídák’s test showed CSD+PINC vs PINC on CSD day 7 (P = 0.0012), post-PINC days 3 (P = 0.0065), 5 (P = 0.0129), and 7 (P = 0.0017). AUC analysis confirmed that CSD+PINC mice showed significantly lower AUC (P < 0.001, Unpaired t-test). n = 14 mice per group. Data are mean±SEM. *P < 0.05, **P < 0.01, ***P < 0.001 (vs control in (B and C); vs PINC in (E)). Abbreviations: AUC, area under curve; Con, control; CSD, chronic sleep deprivation; PINC, plantar incision. |
To investigate the impact of sleep disturbance on postoperative pain, we developed an incisional pain model in mice subjected to sleep deprivation and measured the PWT at multiple time points (Figure 3D). As shown in Figure 3E, sleep deprivation significantly reduced postoperative pain at 3, 5, and 7 days after surgery. Additionally, mice in the normal sleep group exhibited recovery from pain approximately 7 days post-surgery, whereas those in the sleep deprivation group did not achieve baseline pain thresholds even 9 days post-surgery. This part of our study confirmed that sleep disturbance led to a reduction in pain thresholds, exacerbation of postoperative pain, and extension of recovery duration.
KYNU Expression Was Increased in Both the Skin and Blood of Patients with Sleep Disorders
To explore the mechanism by which sleep disorders worsen pain, we first divided patients into four groups based on normal sleep (NS) or sleep disorders (SD) and adequate analgesia (AA) or inadequate analgesia (IA) (Figure 4A), and then collected skin and blood samples for transcriptomic analysis. To obtain more comprehensive biological information, we identified differentially expressed genes with P < 0.05. Compared with patients with NS, those with SD exhibited 1,037 and 738 differentially expressed genes in the peripheral blood and skin tissues, respectively, including 365 upregulated and 307 downregulated genes in the blood, and 481 upregulated and 257 downregulated genes in the skin (Figure 4B). Compared with the IA group, the AA group demonstrated 1,499 and 1,186 differentially expressed genes in the peripheral blood and skin tissues, respectively, including 1,278 upregulated and 221 downregulated genes in the blood, and 209 upregulated and 977 downregulated genes in the skin (Figure 4C).
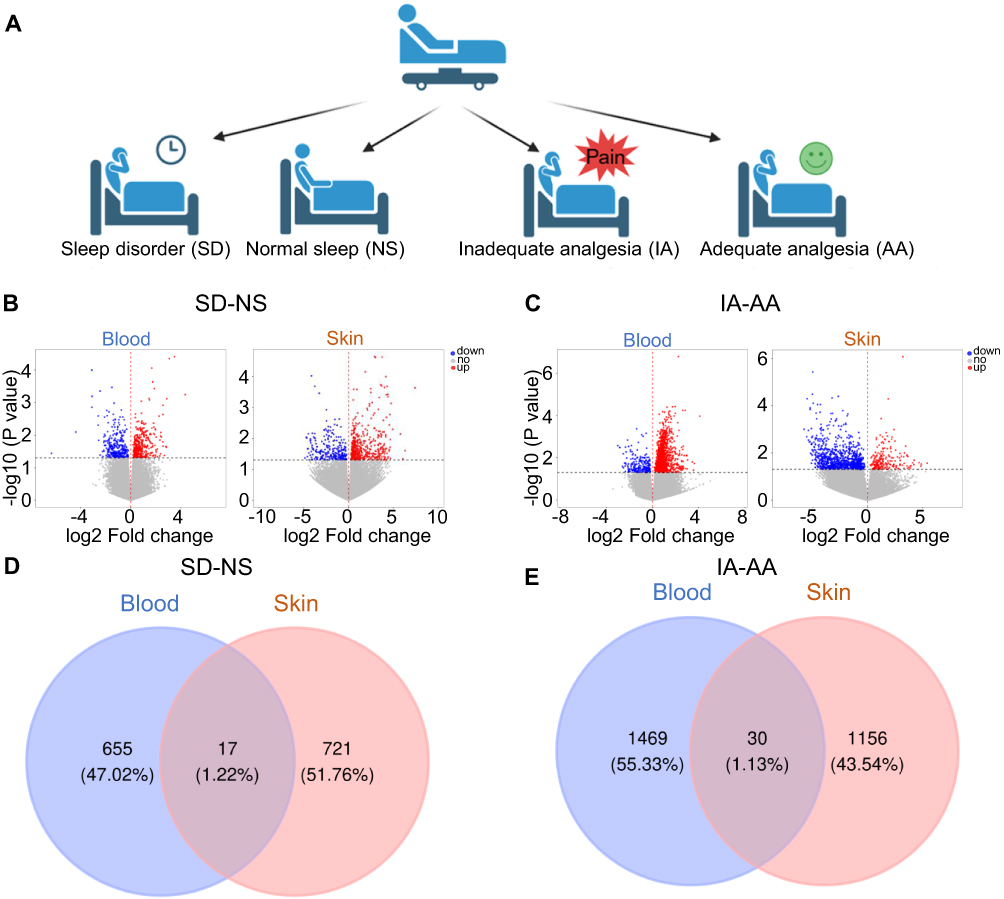 |
Figure 4 KYNU expression is increased in both the skin and blood. (A) Grouping information of the study participants. (B) Gene expression volcano map of the skin and blood in SD vs NS groups. (C) Gene expression volcano map of the skin and blood in AA vs IA groups. (D) Venn analysis identifies differential gene co-expression between the skin and blood in SD vs NS groups. (E) Venn analysis identifies differential gene co-expression between the skin and blood in AA vs IA groups. Abbreviations: AA, adequate analgesia; KYNU, kynureninase; NS, normal sleep; IA, inadequate analgesia; SD, sleep disorder. |
Furthermore, we performed a Venn diagram analysis of the differentially expressed genes in the peripheral blood and skin tissues. The results revealed that in the comparison between patients with NS and those with SD, 17 differentially expressed genes were simultaneously detected in both peripheral blood and skin, with eight genes consistently upregulated and five consistently downregulated in both tissues, whereas the remaining showed inconsistent expression trends between the different tissues (Figure 4D and Table 2). In the comparison between the IA and AA groups, 30 differentially expressed genes were simultaneously detected in both peripheral blood and skin, with 11 genes consistently upregulated and six consistently downregulated in both tissues, whereas the remaining showed inconsistent expression trends between different tissues (Figure 4E and Table 3). Notably, KYNU was the only gene that showed differential expression in both the SD and NS comparison and the IA and AA comparison, with upregulation detected in both contexts. Subsequently, a correlation analysis was performed to examine the relationship between KYNU expression levels and PSQI as well as postoperative maximum and cumulative pain scores. These findings indicate a significant positive correlation between KYNU expression levels in the skin and each of these variables (Figure 5A–C). In contrast, the expression level of KYNU in the blood does not demonstrate a significant correlation with these parameters (Figure 5D–F). Based on these findings, we hypothesized that KYNU may be a critical molecular link between sleep disorders and pain.
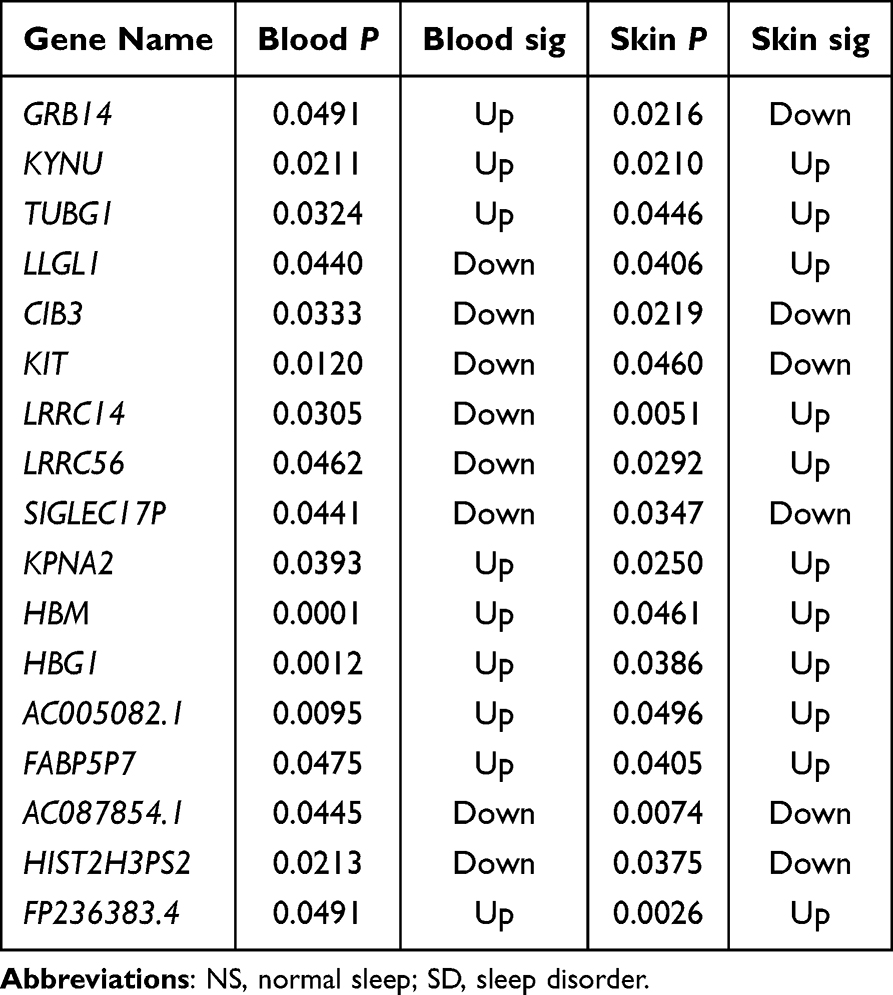 |
Table 2 Differential Gene Expression in Skin and Blood of SD Vs NS Patients |
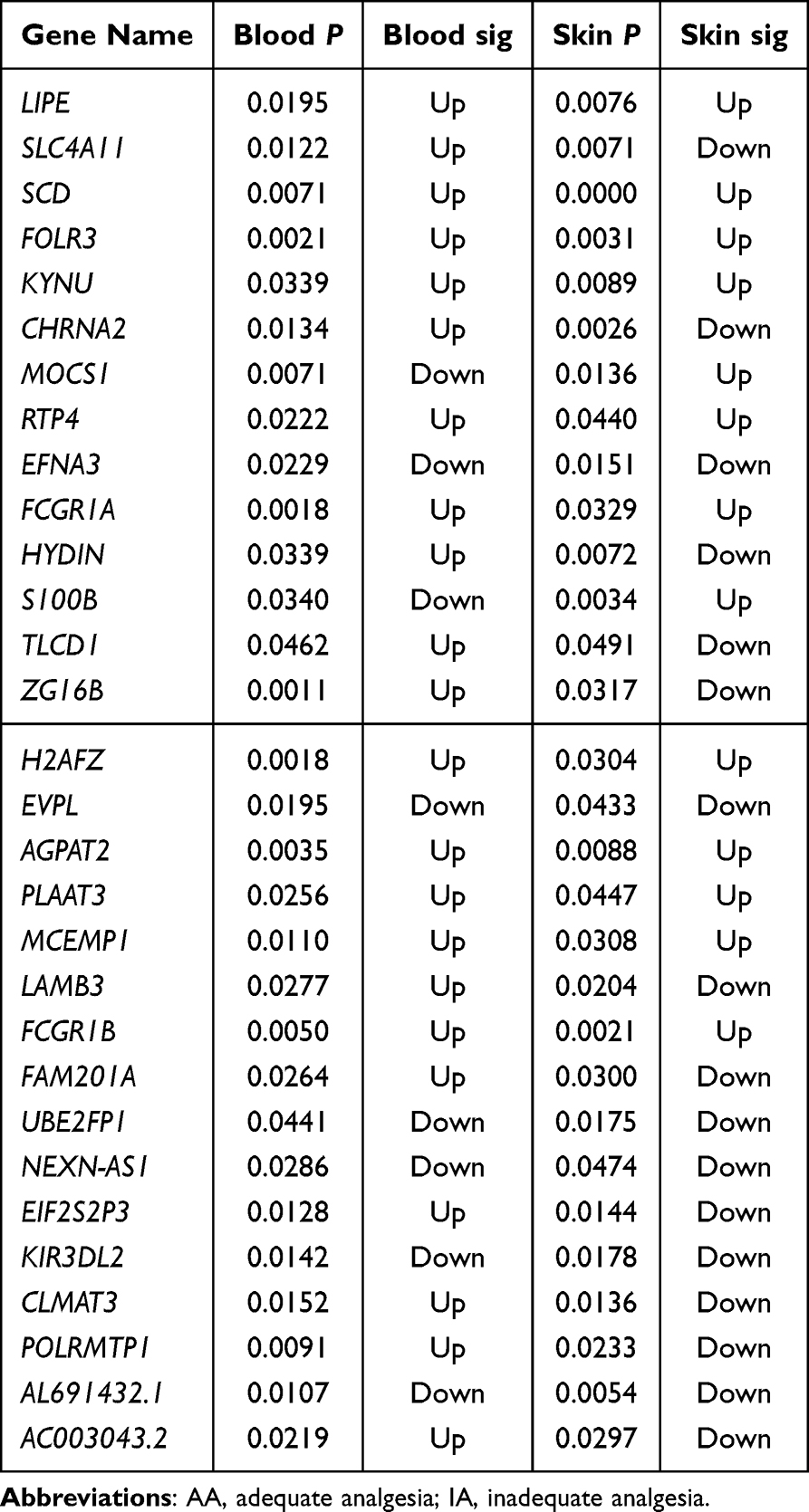 |
Table 3 Differential Gene Expression in Skin and Blood of IA Vs AA Patients |
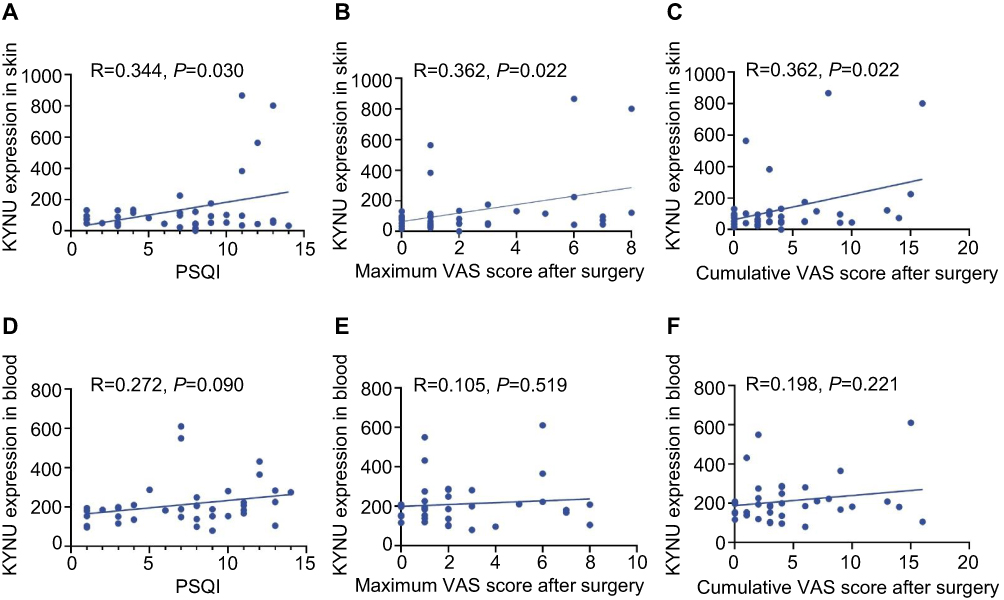 |
Figure 5 Correlation analysis between KYNU expression and sleep quality and postoperative pain outcomes. Skin KYNU expression showed significant positive correlations with (A) PSQI scores (R = 0.344, P = 0.030), (B) maximum VAS scores (R = 0.362, P = 0.022), and (C) cumulative VAS scores (R = 0.362, P = 0.022). Blood KYNU expression showed no significant correlations with (D) PSQI scores (R = 0.272, P = 0.090) or (E) maximum VAS scores (R = 0.198, P = 0.221), but correlated with (F) cumulative VAS scores (R = 0.362, P = 0.022). Pearson correlation analysis, n = 40 patients. Dashed line represents linear regression fit. Each dot represents one patient. Abbreviations: KYNU, kynureninase; PSQI, Pittsburgh Sleep Quality Index; VAS, Visual Analogue Scale. |
KYNU Expression Was Upregulated in the Skin, Blood Cells, and DRG, Which Concurrently Increased Neuronal Excitability in Mice Subjected to Sleep Deprivation
Clinical transcriptomic analyses have demonstrated increased KYNU expression in individuals with sleep disorders and suboptimal postoperative pain management. Through basic research, we further explored how KYNU might contribute to the worsening of postoperative pain due to sleep disorders (Figure 6A). Compared with the normal sleep group, mice with CSD showed significantly upregulated KYNU mRNA expression levels in the blood and skin tissues (Figure 6B and C), consistent with our clinical findings. The DRG, formed by clusters of primary sensory neuron cell bodies, is responsible for transmitting peripheral sensory information (such as pain, temperature, and touch) to the central nervous system and plays a crucial role in the conduction and integration of pain signals.35 We measured KYNU expression levels in the DRG following sleep deprivation using RT-qPCR. The findings indicated a significant upregulation of KYNU expression in the DRG as a result of sleep deprivation (Figure 6D).
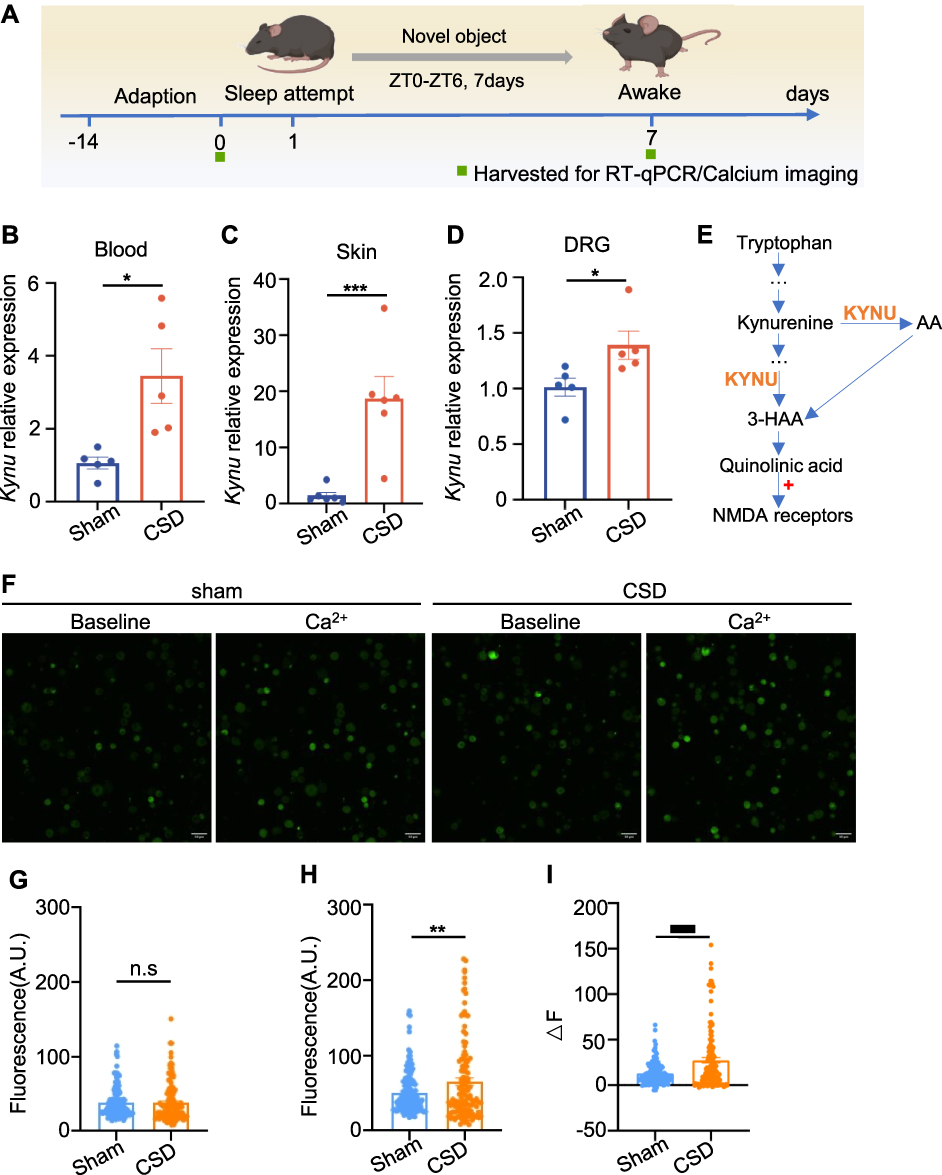 |
Figure 6 KYNU expression is increased in the skin, blood cells, and DRG, leading to an increased neuronal excitability in sleep-deprived mice. (A) Schematic for establishing a CSD mouse model and tissue collection. (B) KYNU increased in the CSD mice blood (P = 0.0142, Unpaired t test, n = 5 per group). (C) KYNU increased in CSD mice skin (P = 0.0022, Mann–Whitney test, n = 6 per group). (D) KYNU increased in CSD mice DRG (P = 0.0159, Mann–Whitney test, n = 5 per group). (E) The KP of tryptophan degradation. (F) Fluorescent calcium imaging of DRG neurons. (G) The fluorescence intensity prior to 2mM Ca2+ stimulation (P = 0.8895, Unpaired t test, n > 100 cells from 6 mice). (H) The fluorescence intensity after 2mM Ca2+ stimulation (P = 0.0065, Unpaired t test, n > 100 cells from 6 mice). (I) The differential fluorescence intensity between the pre- and post-stimulation states (P <0.001, Unpaired t test, n > 100 cells from 6 mice). Error bars represent mean±SEM. Scale bar, 50 μm. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: 3-HAA, 3-hydroxyanthranilic acid; AA, Anthranilic acid; CSD, chronic sleep deprivation; DRG, dorsal root ganglia; KYNU, kynureninase; NMDA, N-methyl-D-aspartate; n.s, not significant; SEM, standard error of the mean. |
KYNU is a key enzyme in the metabolic pathway that converts tryptophan to quinolinic acid and catalyzes the transformation of tryptophan derivatives into quinolinic acid11,36 (Figure 6E). Quinolinic acid is an endogenous agonist of the N-methyl-D-aspartate (NMDA) receptor, and its binding to this receptor enhances neuronal excitability.37 Calcium imaging is widely used to assess neuronal activity. To evaluate the alterations in neuronal excitability, we used calcium imaging, as illustrated in Figure 6F–I. Compared with the normal sleep group, mice subjected to sleep deprivation exhibited no significant difference in baseline calcium signal intensity (Figure 6G). However, upon stimulation with 2 mM Ca2+, the amplitude of the calcium signal in the sleep deprivation group was significantly higher than that the control group (Figure 6H). These findings indicate that sleep deprivation markedly enhances neuronal sensitivity and excitability in response to stimulation. Subsequently, we assessed KYNU protein expression levels in the DRG on the first postoperative day using immunofluorescence staining. The results showed that compared with the surgery-only group, DRG neurons in the sleep deprivation plus surgery group exhibited significantly increased KYNU expression (Figure 7). These findings suggest that sleep disorders not only upregulate basal KYNU expression levels in the DRG but also amplify surgery-induced KYNU expression. This indicates that KYNU may serve as a pivotal molecular target connecting sleep disorders with postoperative pain.
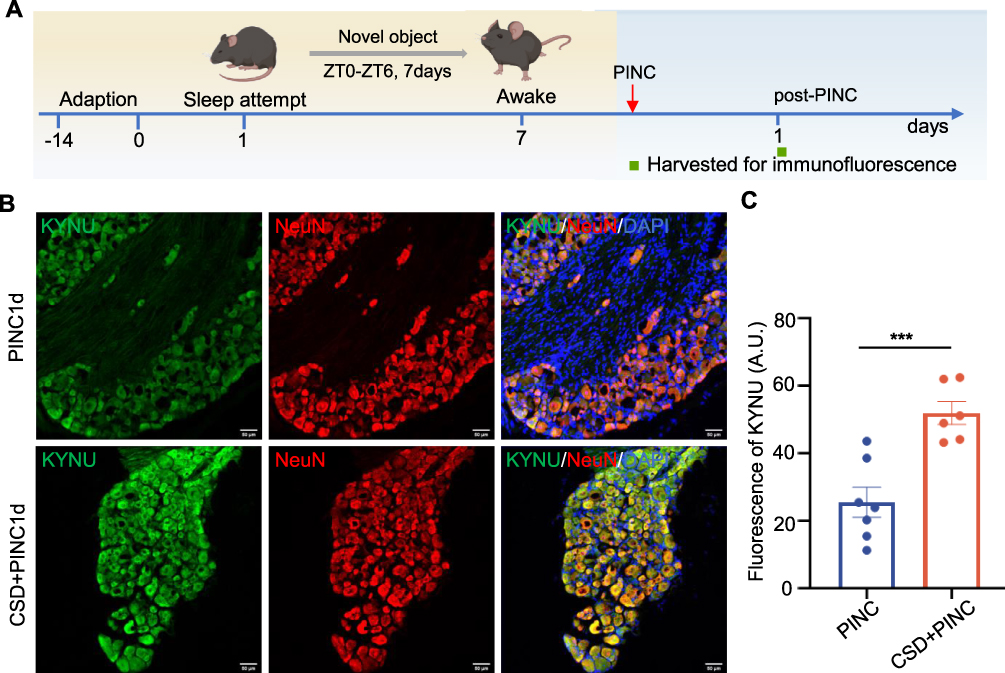 |
Figure 7 KYNU expression is increased in the DRG of mice subjected to CSD and PINC. (A) Schematic for inducing PINC surgery following CSD for 7 days and subsequent tissue collection. (B) Representative immunofluorescence images of the L4-L5 DRG sections showing KYNU (green) and NeuN (red) with DAPI (blue) (Scale bars = 50 μm). (C) CSD+PINC mice showed significantly higher KYNU fluorescence intensity compared to PINC alone (P < 0.001, Unpaired t test). Error bars represent mean±SEM (PINC: n = 7 mice, CSD+PINC: n = 6 mcie). *** P < 0.001 for PINC vs CSD+PINC. Abbreviations: CSD, chronic sleep deprivation; KYNU, kynureninase; PINC, plantar incision. |
Carbidopa Alleviated Postoperative Pain and Decreased Neuronal Excitability
To determine the functional role of KYNU in the exacerbation of pain associated with sleep disorders, we administered the CBA via intraperitoneal injection and subsequently evaluated pain responses. We evaluated the analgesic effects of CBA in a basic postoperative pain model (Figure 8A). First, we determined that carbidopa reaches peak alleviate pain threshold effect at approximately 2 h post-injection in mice (Figure S2). Therefore, all behavioral testing and tissue collection were performed 2 h after carbidopa or vehicle administration to capture the period of optimal target engagement. Compared with the vehicle control group, the CBA-treated group exhibited significantly reduced mechanical hyperalgesia on postoperative days 1 and 3 (Figure 8B). Subsequently, we evaluated the analgesic effects of CBA using a model of sleep deprivation combined with postoperative pain (Figure 8C). As shown in Figure 8D, CBA treatment significantly increased the mechanical pain thresholds on postoperative days 1, 2, and 3. These findings indicate that KYNU modulation can significantly alleviate postoperative pain in mice, confirming that KYNU plays a crucial role in increasing pain sensitivity in sleep disorders.
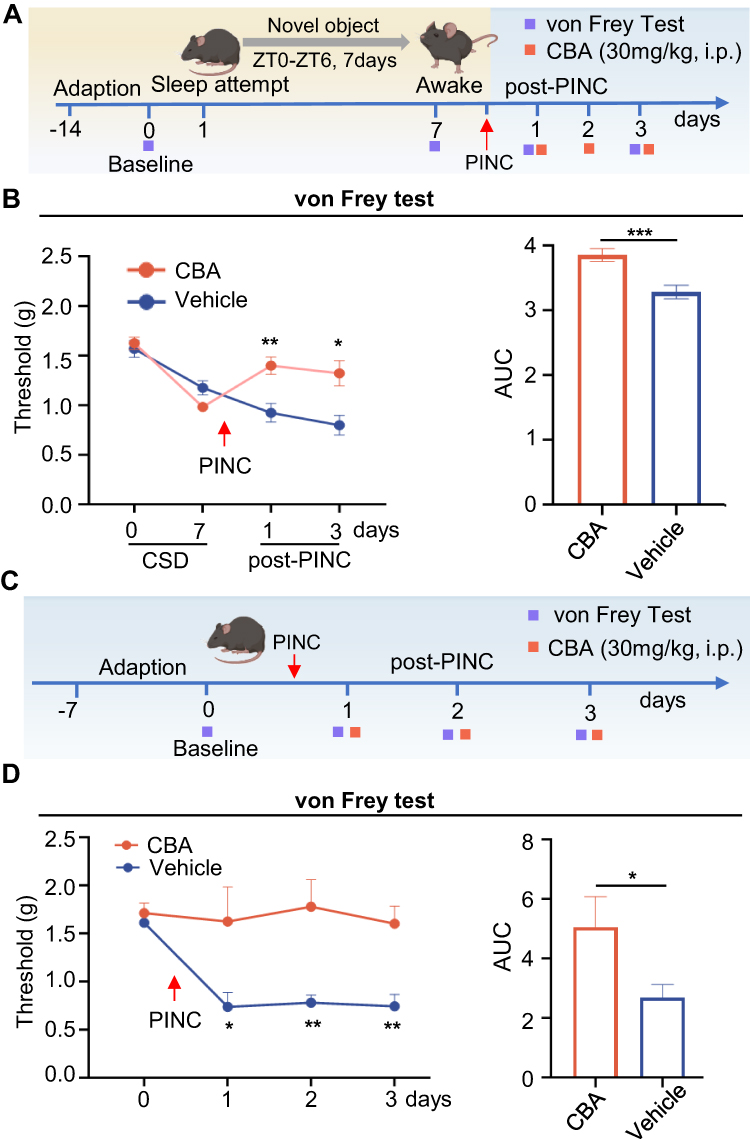 |
Figure 8 CBA alleviates postoperative pain in the PINC model. (A) Experimental timeline for Panel B. Mice underwent 7 days of CSD followed by PINC surgery. CBA (30 mg/kg, i.p.) or vehicle was administered 30 min before PINC and daily for 3 postoperative days. (B) Two-way RM ANOVA revealed significant main effects of treatment (P = 0.0011), time (P < 0.001, Greenhouse-Geisser corrected), and treatment × time interaction (P < 0.001, Greenhouse-Geisser corrected), Tukey’s post-hoc tests showed no significant differences at baseline or after CSD (day 7) between groups (both P > 0.05), but CBA significantly attenuated hyperalgesia on post-PINC day 1 (P = 0.007) and day 3 (P = 0.0189). AUC analysis confirmed treatment effects (P < 0.001, Unpaired t test; CBA: n=10 mice, vehicle: n=9 mice). (C) Experimental timeline for Panel D. PINC was performed on day 0. CBA (30 mg/kg, i.p.) or vehicle was administered 30 min before PINC and daily for 3 days. (D) Two-way RM ANOVA revealed significant main effects of treatment (P = 0.0019), time (P = 0.0227, Greenhouse-Geisser corrected), and a significant treatment × time interaction (P = 0.0318, Greenhouse-Geisser corrected), Tukey’s post-hoc tests showed no significant difference at baseline (P > 0.05), but CBA significantly attenuated hyperalgesia on all post-PINC days (day 1, P = 0.0432; day 2, P = 0.0073; day 3, P = 0.0017). AUC analysis confirmed treatment effects (P = 0.0238, Unpaired t test). n = 9 mice per group. Data are presented as mean±SEM. * P < 0.05, ** P < 0.01, *** P < 0.001 for CBA vs vehicle at the same timepoint. Abbreviations: AUC, area under the curve; CBA, carbidopa; CSD, chronic sleep deprivation; PINC, plantar incision; i.p., intraperitoneal injection. |
Calcium imaging techniques were employed to investigate the effect of KYNU modulation on the excitability of DRG neurons. As illustrated in Figure 9, compared with the surgery-only group, sleep deprivation significantly increased the excitability of postoperative DRG neurons, whereas the administration of CBA significantly reduced neuronal excitability. These findings indicated that KYNU modulation alleviated postoperative pain by reducing the abnormal excitability of DRG neurons.
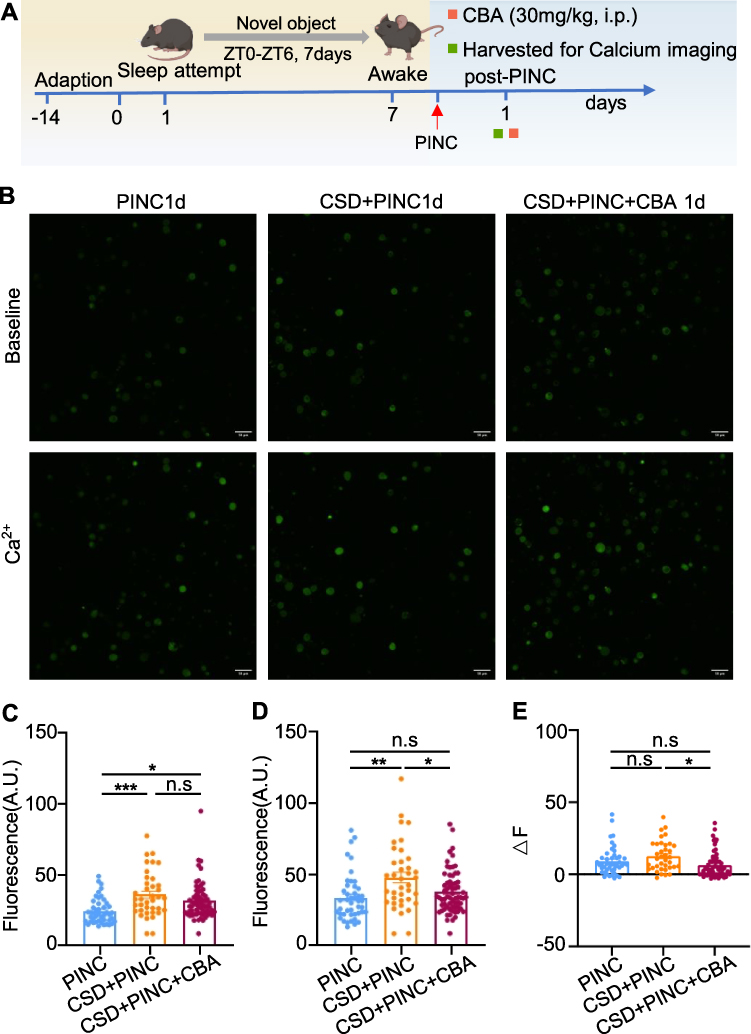 |
Figure 9 CBA decreases neuronal excitability. (A)Schematic for inducing PINC following CSD and subsequent tissue collection. (B) Fluorescent calcium imaging of the DRG neurons. (C)The fluorescence intensity prior to 2 mM Ca2+ stimulation (P < 0.001, n > 38 cells from 6 mice). (D) The fluorescence intensity after 2mM Ca2+ stimulation (n > 38 cells from 6 mice). (E) The differential fluorescence intensity between the pre- and post-stimulation states (P = 0.0080, n > 38 cells from 6 mice). The data were analyzed using the one-way ANOVA flowed by Tukey’s multiple comparisons test. Error bars represent mean±SEM. Scale bar, 50 μm. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: CBA, carbidopa; CSD, chronic sleep deprivation; PINC, plantar incision; n.s, not significant; i.p., intraperitoneal injection; SEM, standard error of the mean. |
Carbidopa Alleviated Inflammatory and Neuropathic Pain
Previous research findings have indicated that KYNU modulation significantly reduces postoperative pain in mice, confirming the efficacy of CBA as an analgesic agent (Figure 8). Furthermore, we evaluated the efficacy of CBA in inflammatory and neuropathic pain models. First, we evaluated the analgesic effects of CBA in Car model (Figure 10A). Compared with the vehicle control group, the CBA-treated group exhibited significantly reduced mechanical hyperalgesia on post-Car days 1, 2 and 3 (Figure 10B). Subsequently, we evaluated the analgesic effects of CBA in the SNI model (Figure 10C). As shown in Figure 10D, CBA treatment significantly increased the mechanical pain thresholds on post SNI days 7, 10, and 14. These findings indicated that CBA exhibited significant analgesic effects in both inflammatory and neuropathic pain models, suggesting its considerable potential as an analgesic agent.
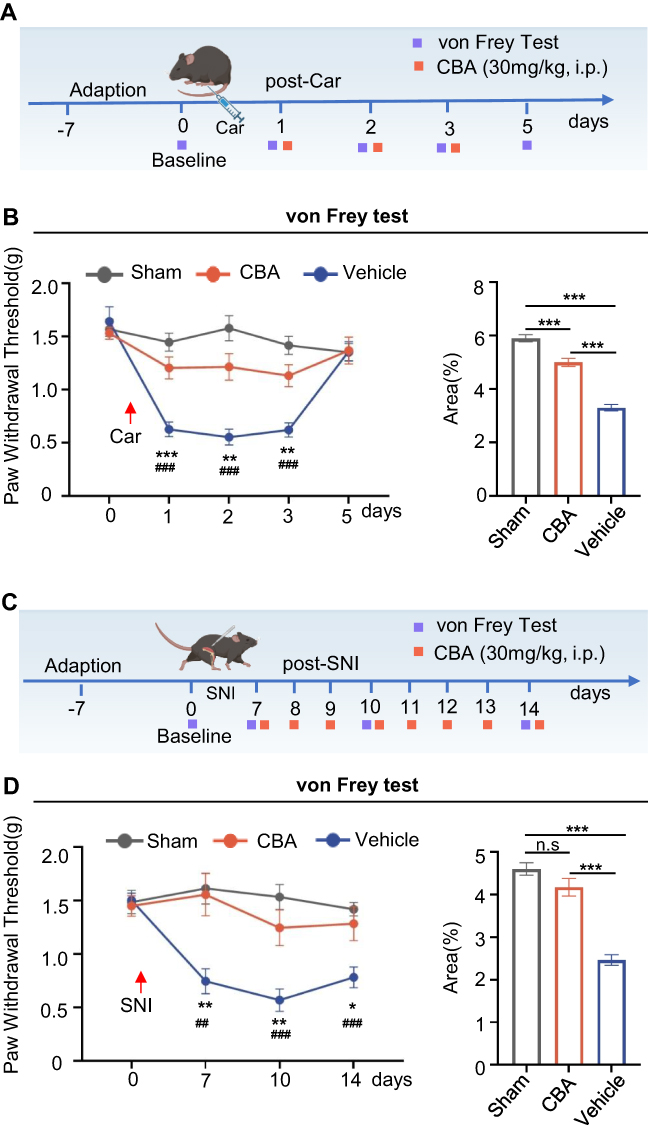 |
Figure 10 CBA reduces mechanical hyperalgesia in inflammatory and neuropathic pain. (A) Experimental timeline for the inflammatory pain model. Car (5 μL of 1% in saline) or saline (Sham) was injected into the right hind paw on day 0. CBA (30 mg/kg, i.p.) or vehicle was administered 30 min before Car injection and daily for 3 days thereafter. (B) Two-way RM ANOVA revealed significant main effects of treatment (P < 0.001), time (P < 0.001, Greenhouse-Geisser corrected), and treatment × time interaction (P < 0.001, Greenhouse-Geisser corrected), Tukey’s post-hoc tests: CBA vs vehicle on days 1 (P < 0.001), 2 (P = 0.0012), and 3 (P = 0.0024); no difference on day 5 (P > 0.05). Sham differed from vehicle on days 1–3 (all P < 0.001). AUC analysis confirmed treatment effects (P < 0.001, one-way ANOVA): CBA vs Vehicle (P < 0.001), CBA vs sham (P < 0.001), Sham vs Vehicle (P < 0.001). (C) Timeline for neuropathic pain model. SNI performed on day 0. CBA (30 mg/kg, i.p.) was administered daily from days 7–14 post-surgery. (D) Two-way RM ANOVA revealed significant main effects of treatment (P < 0.001), time (P = 0.0107, Greenhouse-Geisser corrected), and a significant treatment × time interaction (P = 0.0114, Greenhouse-Geisser corrected), Tukey’s post-hoc tests: CBA vs vehicle on day 7 (P = 0.0085), day 10 (P = 0.0097), and day 14 (P = 0.0421). Sham differed from vehicle on all days (day 7: P = 0.0011; day 10: P < 0.001; day 14: P < 0.001). AUC analysis confirmed treatment effects (P < 0.001, one-way ANOVA): CBA vs Vehicle (P < 0.001), CBA vs sham (P > 0.05), Sham vs Vehicle (P < 0.001). Data are presented as mean ± SEM. Sham: n = 8 mice; vehicle: n = 8 mice, CBA: n = 10 mice. *P < 0.05, **P < 0.01, ***P < 0.001 for CBA vs vehicle; ##P < 0.01, ###P < 0.001 for sham vs vehicle. Abbreviations: AUC, area under curve; Car, carrageenan; CBA, carbidopa; n.s, not significant; i.p., intraperitoneal injection; SNI, spared nerve injury. |
Discussion
In this study, we combined clinical observations with basic research to confirm that preoperative sleep disturbances worsen postoperative pain. Our study demonstrated significant KYNU upregulation in the skin, blood, and DRG. Furthermore, the modulation of KYNU activity mitigated sleep deprivation-induced postoperative pain by reducing neuronal excitability. To the best of our knowledge, this is the first study to explore the role of KYNU in sleep disorders and postoperative pain, suggesting that KYNU may serve as a potential therapeutic target for alleviating the exacerbation of postoperative pain associated with sleep disorders.
Surgery is a common intervention for these patients. Approximately 30–80% of surgical patients report experiencing acute pain after surgical procedures.2,3 Approximately 10% of patients experience severe and intractable chronic postsurgical pain one year after surgery.38–40 Understanding the factors that contribute to exacerbated postsurgical pain may enable us to predict which patients are likely to experience delayed recovery, thereby providing an opportunity for effective interventions in the management of postsurgical pain. Recent research indicates that among the various factors influencing postsurgical pain, preoperative sleep disturbance is the most significant predictor of pain in surgical patients.41,42 Patients with primary insomnia exhibit reduced pain thresholds, and sleep deprivation further intensifies pain in patients with rheumatoid arthritis.43,44 Our clinical observations revealed that preoperative sleep disorders exacerbated postoperative pain. To mimic clinical conditions, the present study established a preclinical animal model of chronic sleep disturbance (CSD), in which naïve mice were subjected to CSD for 6 h daily for 7 days before surgery. Our data showed that 6 h daily for 7 days of CSD aggravated basal and postoperative pain hypersensitivity and prolonged the duration of postoperative pain. These findings are consistent with previous literature.23,45,46
To explore how sleep disorders increase pain hypersensitivity, we collected the skin and blood samples from patients in a clinical study for transcriptomic analysis. Analysis of gene sets from patients with sleep disturbances and postoperative pain revealed that KYNU, a key hydrolase of kynurenine in the KP, was significantly upregulated in the skin and blood. KP plays an important role in the pathogenesis of many autoimmune and neurodegenerative disorders and pain.11,47,48 Numerous enzymes, including indoleamine 2,3-dioxygenase, monoamine oxidase, kynurenine aminotransferase, kynurenine 3-monooxygenase (KMO), and KYNU, play a crucial role in KP regulation. It has been established that KYNU is aberrantly expressed in various diseases, including psoriasis, tumors, depression, and neurodegenerative disorders.16,20,49 The current results confirmed that KYNU expression was increased in the skin and blood of individuals with sleep disorders. Furthermore, KYNU expression was significantly upregulated in the skin, blood, and DRG of mice subjected to sleep deprivation. Similarly, in a previous study, KYNU mRNA levels were upregulated in the spinal cord following chronic constriction injury (CCI) in mice.50 However, a previous study conducted in rats demonstrated that KYNU mRNA levels remained unchanged in both the spinal cord and DRG at different time points after CCI.51 This divergence is likely attributable to variations in species and animal models, necessitating further investigation in subsequent experiments.
Kynurenine is subsequently metabolized to quinolinic acid (QUIN) by multiple enzymes, including KMO, KYNU, and 3-hydroxyanthranilate oxidase.11,52,53 QUIN, a selective endogenous NMDA receptor agonist, is involved in the development of neuropathy.54,55 Notably, QUIN can induce persistent damage and facilitate the influx of substantial quantities of Ca2+ into the neurons and astrocytes.14 Calcium ions, which serve as pivotal intracellular signaling molecules, are integral to neuronal function.56,57 Calcium imaging technology is crucial for investigating neuronal excitability and activity.58–60 Previous research has shown that sleep disturbances exacerbate pain perception by increasing neuronal excitability, resulting in the abnormal transmission of pain signals and consequently intensifying the experience of pain.23,61 In the current study, KYNU expression was increased in individuals with sleep disorders and in sleep-deprived mice. We hypothesized that KYNU enhances neuronal excitability through the action of QUIN, thereby increasing pain sensitivity. As anticipated, neuronal excitability was enhanced after sleep deprivation. Moreover, repeated intraperitoneal injections of CBA, a non-specific KYNU inhibitor, decreased pain sensitivity and neuronal excitability. In addition, the analgesic effect of CBA was confirmed in both the inflammatory and neuropathic pain models. Nonetheless, future investigations are essential to determine whether KYNU directly contributes to pain associated with sleep disorders by elevating QUIN levels.
The current study has several limitations. Although it offers valuable insights, it is important to acknowledge the potential limitations inherent in its design. First, this was a single-center study involving patients undergoing abdominal surgery. The generalizability of the findings derived from single-center studies may be constrained by variations in patient demographics, surgical methodologies, and postoperative management practices across different centers. Second, the limited sample size can contribute to heightened variability in the research findings. Third, the role of enzymes regulating the KP in pain has been investigated in other pain models.50,51 For the first time, we have investigated the role of the KP in the exacerbation of pain associated with sleep disorders. This study employed transcriptomic and Wien analyses to identify differential molecules, specifically focusing on KYNU. Consequently, this investigation exclusively assessed KYNU expression, while the alterations in other enzymes within the KP were not addressed. Fourth, this study utilized calcium imaging technology and pharmacological methods to demonstrate that KYNU enhances pain perception by increasing neuronal excitability. However, the specific mechanisms involved remain unexplored, and future research should focus on this aspect. Fifth, we employed an established experimental approach by providing novel objects in the mouse cage, as noted in previous research.23 During the sleep deprivation period, we did not observe obvious body weight loss and classical stress-related behaviors including excessive grooming, barbering (whisker or fur trimming), stereotypic behaviors, aggression, or reduced exploratory activity. However, a limitation of this study is the absence of measurements for indicators that can reflect stress, such as blood pressure, heart rate, and plasma corticosterone levels. Additionally, the study has a limitation of short-term follow-up, which we acknowledge and discuss. The research focused on acute postoperative pain within the first week after surgery, as this period represents the peak of clinical burden. However, emerging evidence suggests that sleep disturbances may contribute to the development of chronic postsurgical pain.62 Future studies with extended follow-up (3–6 months) are needed to determine whether preoperative sleep disturbances and KYNU pathway activation predict the transition from acute to chronic postoperative pain. Moreover, such studies could explore whether early interventions with KYNU inhibitors might prevent pain chronification.
In conclusion, our clinical research indicates that patients with preoperative sleep disorders tend to experience heightened postoperative pain, a phenomenon potentially associated with the upregulation of KYNU expression. Using a mouse model, we found that sleep deprivation greatly increased in severity after paw plantar incision, and KYNU was notably higher in the skin, blood, and DRG. Repeated intraperitoneal injections of CBA, a KYNU modulator, decreased pain severity and abnormal DRG neuronal excitability caused by sleep deprivation. Moreover, CBA showed strong pain-relieving effects in both inflammatory and neuropathic pain models. Thus, KYNU is a potentially significant contributor to the exacerbation of postoperative pain in patients with sleep disorders, while the mechanism of carbidopa’s action requires further investigation. Carbidopa emerged in our preclinical model as one possible modulator of this pathway and may represent a potential therapeutic target worthy of further investigation. However, substantial additional work is required before any clinical translation can be considered. Overall, our findings highlight KYNU as a key link between sleep disturbance and postoperative pain, offering new insights into how sleep issues can worsen postsurgical pain.
Clinical Perspectives
- Patients with preoperative sleep disorders are known to experience more severe and prolonged postoperative pain, but the molecular mechanisms underlying this relationship remain unclear, prompting this study to explore potential therapeutic targets.
- By integrating transcriptomic analyses of human skin and blood samples with a mouse model of sleep deprivation and postoperative pain, we identified kynureninase (KYNU) as a key mediator linking sleep disorders and heightened postoperative pain. Pharmacological modulation of KYNU with carbidopa reduced pain severity and neuronal hyperexcitability in both sleep-deprived and broader pain models.
- Our findings highlight KYNU as a critical molecular driver of postoperative pain exacerbated by sleep deprivation and demonstrate the therapeutic potential of carbidopa for managing inflammatory, neuropathic, and postoperative pain, offering novel insights into pain management strategies.
Abbreviations
AA, adequate analgesia; CBA, carbidopa; CSD, chronic sleep deprivation; DRG, dorsal root ganglia; IA, inadequate analgesia; KMO, kynurenine 3-monooxygenase; KP; kynurenine pathway; KYNU, kynureninase; NMDA, N-methyl-D-aspartate; NS, normal sleep; PINC, plantar incision; PSQI, Pittsburgh Sleep Quality Index; PWT, paw withdrawal threshold; QUIN, quinolinic acid; RT-qPCR, Real-time quantitative polymerase chain reaction; SD, sleep disorder; VAS, Visual Analogue Scale.
Data Sharing Statement
The data that support the findings of this study are available upon reasonable request from the corresponding authors, Guangyou Duan (E-mail: [email protected]) or Yuanjing Chen (E-mail: [email protected]).
Acknowledgments
We express our gratitude to the participants for their selfless contributions to this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82571388 and 82171210).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Dobson GP. Trauma of major surgery: a global problem that is not going away. Int J Surg. 2020;81:47–23. doi:10.1016/j.ijsu.2020.07.017
2. Meissner W, Zaslansky R. A survey of postoperative pain treatments and unmet needs. Best Pract Res. 2019;33:269–286. doi:10.1016/j.bpa.2019.10.003
3. Apfelbaum JL, Chen C, Mehta SS, Gan ATJ. Postoperative pain experience: results from a national survey suggest postoperative pain continues to be undermanaged. Anesthesia Analgesia. 2003;97:534–540. doi:10.1213/01.ANE.0000068822.10113.9E
4. Carley ME, Chaparro LE, Choinière M, et al. Pharmacotherapy for the prevention of chronic pain after surgery in adults: an updated systematic review and meta-analysis. Anesthesiology. 2021;135:304–325. doi:10.1097/ALN.0000000000003837
5. de Boer HD, Detriche O, Forget P. Opioid-related side effects: postoperative ileus, urinary retention, nausea and vomiting, and shivering. A review of the literature. Best Pract Res Clin Anaesthesiol. 2017;31:499–504. doi:10.1016/j.bpa.2017.07.002
6. Kianian S, Bansal J, Lee C, Zhang K, Bergese SD. Perioperative multimodal analgesia: a review of efficacy and safety of the treatment options. Anesthesiol Perioper Sci. 2024;2:9. doi:10.1007/s44254-023-00043-1
7. Niklasson A, Finan PH, Smith MT, et al. The relationship between preoperative sleep disturbance and acute postoperative pain control: a systematic review and meta-analysis. Sleep Med Rev. 2025;79:102014. doi:10.1016/j.smrv.2024.102014
8. Wu H, Su W, Huang S, Xiao Y, Lu L. Correlation between pre-operative sleep disturbance and post-operative pain in patients with rotator cuff tear. Front Integr Neurosci. 2022;16:942513. doi:10.3389/fnint.2022.942513
9. Bjurström MF, Irwin MR, Bodelsson M, Smith MT, Mattsson-Carlgren N. Preoperative sleep quality and adverse pain outcomes after total Hip arthroplasty. Eur J Pain. 2021;25:1482–1492. doi:10.1002/ejp.1761
10. Coronado RA, Pennings JS, Master H, et al. The combined influence of sleep disturbance and depression on 12-month outcomes after lumbar spine surgery. Spine. 2024;49:1339–1347. doi:10.1097/BRS.0000000000005000
11. Schwarcz R, Stone TW. The kynurenine pathway and the brain: challenges, controversies and promises. Neuropharmacology. 2017;112:237–247. doi:10.1016/j.neuropharm.2016.08.003
12. Ciapała K, Mika J, Rojewska E. The kynurenine pathway as a potential target for neuropathic pain therapy design: from basic research to clinical perspectives. Int J Mol Sci. 2021;22:11055. doi:10.3390/ijms222011055
13. Pocivavsek A, Schwarcz R, Erhardt S. Neuroactive kynurenines as pharmacological targets: new experimental tools and exciting therapeutic opportunities. Pharmacol Rev. 2024;76:978–1008. doi:10.1124/pharmrev.124.000239
14. Lugo-Huitrón R, Ugalde Muñiz P, Pineda B, Pedraza-Chaverrí J, Ríos C, Pérez-de la Cruz V. Quinolinic acid: an endogenous neurotoxin with multiple targets. Oxid Med Cell Longev. 2013;2013:104024. doi:10.1155/2013/104024
15. Behan WM, McDonald M, Darlington LG, Stone TW. Oxidative stress as a mechanism for quinolinic acid-induced hippocampal damage: protection by melatonin and deprenyl. Br J Pharmacol. 1999;128:1754–1760. doi:10.1038/sj.bjp.0702940
16. Wang M, Wang Y, Zhang M, et al. Kynureninase contributes to the pathogenesis of psoriasis through pro‐inflammatory effect. J Cell Physiol. 2022;237:1044–1056. doi:10.1002/jcp.30587
17. Auyeung A, Wang HC, Aravagiri K, Knezevic NN. Kynurenine pathway metabolites as potential biomarkers in chronic pain. Pharm. 2023;16:681. doi:10.3390/ph16050681
18. Harden JL, Lewis SM, Lish SR, et al. The tryptophan metabolism enzyme L-kynureninase is a novel inflammatory factor in psoriasis and other inflammatory diseases. J Allergy Clin Immunol. 2016;137:1830–1840. doi:10.1016/j.jaci.2015.09.055
19. Maganin AG, Souza GR, Fonseca MD, et al. Meningeal dendritic cells drive neuropathic pain through elevation of the kynurenine metabolic pathway in mice. J Clin Invest. 2022;132. doi:10.1172/JCI153805
20. Badawy AA-B, Bano S. Tryptophan metabolism in rat liver after administration of tryptophan, kynurenine metabolites, and kynureninase inhibitors. Int J Tryptophan Res. 2016;
21. Badawy AA-B, Bano S, Steptoe A. Tryptophan in alcoholism treatment II: inhibition of the rat liver mitochondrial low km aldehyde dehydrogenase activity, elevation of blood acetaldehyde concentration and induction of aversion to alcohol by combined administration of tryptophan and benserazide. Alcohol Alcoholism. 2011;46:661–671. doi:10.1093/alcalc/agr135
22. Liu Q, Ling Z, Zhu Y, et al. Targeting kynureninase attenuates radiation-induced intestinal injury via MAPK signaling suppression. Mediators Inflamm. 2025;2025:7023259. doi:10.1155/mi/7023259
23. Alexandre C, Latremoliere A, Ferreira A, et al. Decreased alertness due to sleep loss increases pain sensitivity in mice. Nat Med. 2017;23:768–774. doi:10.1038/nm.4329
24. You S, Xu F, Zhu X, et al. Effect of intraoperative noise on postoperative pain in surgery patients under general anesthesia: evidence from a prospective study and mouse model. Int J Surg. 2023;109:3872–3882. doi:10.1097/JS9.0000000000000672
25. Xu Z, Xie W, Feng Y, et al. Positive interaction between GPER and β-alanine in the dorsal root ganglion uncovers potential mechanisms: mediating continuous neuronal sensitization and neuroinflammation responses in neuropathic pain. J Neuroinflammation. 2022;19:164. doi:10.1186/s12974-022-02524-9
26. van der Vlist M, Raoof R, Willemen HLDM, et al. Macrophages transfer mitochondria to sensory neurons to resolve inflammatory pain. Neuron. 2022;110:613–626.e9. doi:10.1016/j.neuron.2021.11.020
27. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53:55–63. doi:10.1016/0165-0270(94)90144-9
28. Shu B. Opioid infusions at different times of the day produce varying degrees of opioid-induced hyperalgesia. Br J Anaesth. 2023;131:1072–1081. doi:10.1016/j.bja.2023.08.039
29. Kaakkola S, Wurtman RJ. Effects of catechol-O-methyltransferase inhibitors and L-3,4-dihydroxyphenylalanine with or without carbidopa on extracellular dopamine in rat striatum. J Neurochem. 1993;60:137–144. doi:10.1111/j.1471-4159.1993.tb05831.x
30. Nicholas AP. Levodopa-induced hyperactivity in mice treated with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Mov Disord. 2007;22:99–104. doi:10.1002/mds.21235
31. Cheng D, Qin Z-S, Zheng Y, et al. Minocycline, a classic antibiotic, exerts psychotropic effects by normalizing microglial neuroinflammation-evoked tryptophan-kynurenine pathway dysregulation in chronically stressed male mice. Brain Behav Immun. 2023;107:305–318. doi:10.1016/j.bbi.2022.10.022
32. Mo L, Jing H, Du X, et al. Goat and cow milk differ in altering the microbiota composition and neurotransmitter levels in insomnia mouse models. Food Funct. 2023;14:6526–6540. doi:10.1039/d3fo00797a
33. Zhang Q, Ren Y, Mo Y, et al. Inhibiting Hv1 channel in peripheral sensory neurons attenuates chronic inflammatory pain and opioid side effects. Cell Res. 2022;32:461–476. doi:10.1038/s41422-022-00616-y
34. Kawaratani H, Miyaaki H, Hiraoka A, et al. The usefulness of the athens insomnia scale for evaluating sleep disturbance in patients with chronic liver disease comparing with pittsburgh sleep quality index and epworth sleepiness scale. Med. 2022;58:741. doi:10.3390/medicina58060741
35. Esposito MF, Malayil R, Hanes M, Deer T. Unique characteristics of the dorsal root ganglion as a target for neuromodulation. Pain Med. 2019;20:S23–30. doi:10.1093/pm/pnz012
36. Liu D, Liang C-H, Huang B, et al. Tryptophan metabolism acts as a new anti-ferroptotic pathway to mediate tumor growth. Adv Sci. 2023:10e2204006. doi:10.1002/advs.202204006
37. Stone TW, Perkins MN. Quinolinic acid: a potent endogenous excitant at amino acid receptors in CNS. Eur J Pharmacol. 1981;72:411–412. doi:10.1016/0014-2999(81)90587-2
38. Katz J, Seltzer Z. Transition from acute to chronic postsurgical pain: risk factors and protective factors. Expert Rev Neurother. 2009;9:723–744. doi:10.1586/ern.09.20
39. Kehlet H, Jensen TS, Woolf CJ. Persistent postsurgical pain: risk factors and prevention. Lancet. 2006;367:1618–1625. doi:10.1016/S0140-6736(06)68700-X
40. Perkins FM, Kehlet H. Chronic pain as an outcome of surgery. A review of predictive factors. Anesthesiology. 2000;93:1123–1133. doi:10.1097/00000542-200010000-00038
41. Yu S, Xiong Y, Lu G, Xiong X. Effects of preoperative sleep disorders on anesthesia recovery and postoperative pain in patients undergoing laparoscopic gynecological surgery under general anesthesia. Mediators Inflammation. 2022;2022:7998104. doi:10.1155/2022/7998104
42. Ni Y, Yu M, Liu C. Sleep disturbance and cognition in the elderly: a narrative review. Anesthesiol Perioper Sci. 2024;2:26. doi:10.1007/s44254-024-00066-2
43. Irwin MR, Olmstead R, Carrillo C, et al. Sleep loss exacerbates fatigue, depression, and pain in rheumatoid arthritis. Sleep. 2012;35:537–543. doi:10.5665/sleep.1742
44. Haack M, Scott-Sutherland J, Santangelo G, Simpson NS, Sethna N, Mullington JM. Pain sensitivity and modulation in primary insomnia. Eur J Pain. 2012;16:522–533. doi:10.1016/j.ejpain.2011.07.007
45. Xue J, Li H, Xu Z, et al. Paradoxical sleep deprivation aggravates and prolongs incision-induced pain hypersensitivity via BDNF signaling-mediated descending facilitation in rats. Neurochem Res. 2018;43:2353–2361. doi:10.1007/s11064-018-2660-2
46. Wang P-K, Cao J, Wang H, et al. Short-Term sleep disturbance–induced stress does not affect basal pain perception, but does delay postsurgical pain recovery. J Pain. 2015;16:1186–1199. doi:10.1016/j.jpain.2015.07.006
47. Savitz J. The kynurenine pathway: a finger in every pie. Mol Psychiatry. 2020;25:131–147. doi:10.1038/s41380-019-0414-4
48. Mecs L, Tuboly G, Nagy E, Benedek G, Horvath G. The peripheral antinociceptive effects of endomorphin-1 and kynurenic acid in the rat inflamed joint model. Anesth Analg. 2009;109:1297–1304. doi:10.1213/ane.0b013e3181b21c5e
49. Xu H, Hu C, Wang Y, et al. Glutathione peroxidase 2 knockdown suppresses gastric cancer progression and metastasis via regulation of kynurenine metabolism. Oncogene. 2023;42:1994–2006. doi:10.1038/s41388-023-02708-4
50. Ciapała K, Pawlik K, Ciechanowska A, Mika J, Rojewska E. Effect of pharmacological modulation of the kynurenine pathway on pain-related behavior and opioid analgesia in a mouse model of neuropathic pain. Toxicol Appl Pharmacol. 2023;461:116382. doi:10.1016/j.taap.2023.116382
51. Rojewska E, Ciapała K, Piotrowska A, Makuch W, Mika J. Pharmacological inhibition of indoleamine 2,3-dioxygenase-2 and kynurenine 3-monooxygenase, enzymes of the kynurenine pathway, significantly diminishes neuropathic pain in a rat model. Front Pharmacol. 2018;9:724. doi:10.3389/fphar.2018.00724
52. Chiarugi A, Carpenedo R, Molina MT, Mattoli L, Pellicciari R, Moroni F. Comparison of the neurochemical and behavioral effects resulting from the inhibition of kynurenine hydroxylase and/or kynureninase. J Neurochem. 1995;65:1176–1183. doi:10.1046/j.1471-4159.1995.65031176.x
53. Guidetti P, Amori L, Sapko MT, Okuno E, Schwarcz R. Mitochondrial aspartate aminotransferase: a third kynurenate-producing enzyme in the mammalian brain. J Neurochem. 2007;102:103–111. doi:10.1111/j.1471-4159.2007.04556.x
54. Guillemin GJ. Quinolinic acid, the inescapable neurotoxin. FEBS J. 2012;279:1356–1365. doi:10.1111/j.1742-4658.2012.08485.x
55. Collins S, Sigtermans MJ, Dahan A, Zuurmond WWA, Perez RSGM. NMDA receptor antagonists for the treatment of neuropathic pain. Pain Med. 2010;11:1726–1742. doi:10.1111/j.1526-4637.2010.00981.x
56. Pikor D, Hurła M, Słowikowski B, et al. Calcium ions in the physiology and pathology of the central nervous system. Int J Mol Sci. 2024;25:13133. doi:10.3390/ijms252313133
57. Martiszus BJ, Tsintsadze T, Chang W, Smith SM. Enhanced excitability of cortical neurons in low-divalent solutions is primarily mediated by altered voltage-dependence of voltage-gated sodium channels. Elife. 2021;10e67914. doi:10.7554/eLife.67914
58. Ross WN. Understanding calcium waves and Sparks in central neurons. Nat Rev Neurosci. 2012;13:157–168. doi:10.1038/nrn3168
59. Anderson M, Zheng Q, Dong X. Investigation of pain mechanisms by calcium imaging approaches. Neurosci Bull. 2018;34:194–199. doi:10.1007/s12264-017-0139-9
60. Li N, Liu B, Wu W, et al. Upregulation of transcription factor 4 downregulates NaV1.8 expression in DRG neurons and prevents the development of rat inflammatory and neuropathic hypersensitivity. Exp Neurol. 2020;327:113240. doi:10.1016/j.expneurol.2020.113240
61. Guo M, Wu Y, Zheng D, et al. Preoperative acute sleep deprivation causes postoperative pain hypersensitivity and abnormal cerebral function. Neurosci Bull. 2022;38:1491–1507. doi:10.1007/s12264-022-00955-1
62. Varallo G. Sleep disturbances and sleep disorders as risk factors for chronic postsurgical pain: a systematic review and meta-analysis. Sleep Med Rev. 2022;63.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.