Back to Journals » OncoTargets and Therapy » Volume 12
Epigenetics mechanisms mediate the miR-125a/BRMS1 axis to regulate invasion and metastasis in gastric cancer
Authors Xiong J, Tu Y, Feng Z ![]() , Li D, Yang Z, Huang Q, Li Z, Cao Y, Jie Z
, Li D, Yang Z, Huang Q, Li Z, Cao Y, Jie Z
Received 28 March 2019
Accepted for publication 17 August 2019
Published 12 September 2019 Volume 2019:12 Pages 7513—7525
DOI https://doi.org/10.2147/OTT.S210376
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Carlos E Vigil
Jianbo Xiong,1,* Yi Tu,2,* Zongfeng Feng,1,* Daojiang Li,3 Zhouwen Yang,1 Qiuxia Huang,4 Zhengrong Li,1 Yi Cao,1 Zhigang Jie1
1Department of Gastrointestinal Surgery, First Affiliated Hospital, Nanchang University, Nanchang 330006, Jiangxi Province, People’s Republic of China; 2Department of Pathology, First Affiliated Hospital, Nanchang University, Nanchang 330006, Jiangxi Province, People’s Republic of China; 3Department of General Surgery, Zhongnan Hospital of Wuhan University, Wuhan 430071, Hubei Province, People’s Republic of China; 4Department of Nursing, First Affiliated Hospital, Nanchang University, Nanchang 330006, Jiangxi Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhigang Jie; Yi Cao
Department of Gastrointestinal Surgery, First Affiliated Hospital, Nanchang University, No.17 Yongwai Zheng Road, Nanchang 330006, Jiangxi Province, People’s Republic of China
Tel +86 7 918 869 2522
Email [email protected]; [email protected]
Purpose: Altered expression of breast cancer metastasis suppressor 1 (BRMS1), is a tumor suppressor, which is found in many types of cancers, including gastric cancer (GC), but the mechanism by which BRMS1 inhibits invasion and metastasis in GC is unknown. The aim of the study was to investigate the molecular mechanisms of miR-125a/BRMS1 in GC.
Materials and methods: The expression of BRMS1 and miR-125a were detected by quantitative real-time PCR (qRT-PCR) and analyzed by bioinformatics. BSP and MSP were used to detecte the methylation status of miR-125a and BRMS1 which was treated by 5-Aza or not. Western Blot and qRT-PCR were used to analyze the expression of BRMS1 and EZH2. Transwell was performed to explore the invasion and metastasis ability of GC cells. The nude mice were used for the tumor formation assay.
Results: BRMS1 may be regulated by copy number variation (CNV), methylation and miR-125a-5p. As one of the essential components of PRC2, EZH2 is an important regulatory factor resulting in the low expression of miR-125a. An epigenetic mechanism mediates the miR-125a/BRMS1 axis to inhibit the invasion and metastasis of GC cells. In vivo experiments, it is also showed that BRMS1 is involved in invasion and metastasis but not the proliferation in GC.
Conclusion: These studies shed light on the mechanism of BRMS1 inhibition of GC invasion and metastasis and the development of new drugs targeting the miR-125a/BRMS1 axis, which will be a promising therapeutic strategy for GC and other human cancers.
Keywords: gastric cancer, miR-125a/BRMS1 axis, epigenetic, EZH2, invasion and metastasis
Introduction
As one of the most severe health conditions worldwide, gastric cancer (GC) remains the fourth most common cancer and the third leading cause of cancer-related mortality worldwide.1,2 According to the latest Chinese cancer data statistics report, the new cancer cases and crude incidence of GC is 30.0/105, behind the lung cancer and female breast cancer, and the cancer deaths and crude mortality rate of GC is 21.48/105, following the lung cancer and live cancer.3 Although recent advances have made in diagnosis, surgical resection, chemotherapy and radiation therapy, the prognosis of patients with advanced GC remains rather poor due to the recurrence and metastasis.4 Therefore, identification of the appropriate molecular biomarkers and elucidation of the potential molecular mechanisms are needed for GC therapy.
BRMS1 (breast cancer metastasis suppressor 1) was discovered using a differential comparison of parental MDA-MB-435 cells, and the neo11/435 clone led to the discovery of the BRMS1 chromosomal locus and showed suppressed metastatic potential in the lungs and regional lymph nodes without preventing primary tumor growth, as shown by Jabed Seraj and colleagues in 2000.5,6 This molecule has been reported to suppress metastasis not only in breast cancer, but also in non-small cell lung cancer, ovarian cancer, melanoma, rectal cancer and so on.7 Detection BRMS1 expression in primary GC tissues and metastatic GC tissues by immunohistonchemistry (IHC) and construction of a xenograft model, we found that BRMS1 was markedly stronger in the primary tumor compared with metastasis gastric tumor, which can further suppress the metastasis of GC cells in nude mice but did not inhibit the growth of gastric tumors.8 For the past years, our team have found that the expression of BRMS1 was regulated by miR-125a-5p, which is low expressed in GC and further affects the invasion and metastasis of GC cells in vitro.9 A previous report showed that the expression of BRMS1 can significantly inhibit the adhesion, migration, invasion and metastasis of mouse forestomach carcinoma GC cells, which is related to the NF-κB signal pathway.10 However, the mechanism responsible for the downregulation of miR-125a expression in GC is unknown. In addition to the noncoding RNA miR-125a discussed in our previous study,9 is there any other potential mechanism leading to the low expression of the tumor suppressor BRMS1 in GC?
Epigenetics is defined as the study of changes in gene function that are mitotically and/or meiotically heritable and that do not involve a change in the sequence of DNA.11 Epigenetic modifications play a critical role in the development of pediatric and adult cancers, contributing to the cumulative changes observed as normal cells undergo malignant transformation.12 The most common mammalian epigenetic modifications are DNA methylation, histone modification, chromatin structure and transcriptional activities. Both DNA methylation and histone modification are associated with silencing critical tumor suppressor genes and activating oncogenes involved in cancer.13 Researchers have reported that BRMS1 was downregulated by DNA methylation in breast cancer, non-small cell lung cancer and cervical cancer,14 but the mechanism of its low expression in GC remains unclear. Moreover, genomic DNA copy number variation (CNV) is an important type of genetic alternation observed in tumor cells, and it contributes to tumor evolution by altering the expression of genes within the region.15 Our previous study has found that both of BRMS1 and miR-125a-5p were low expressed in GC tissues and BRMS1 is a potential target gene of miR-125a-5p in GC.9 However, the frequency as well as the relevance of BRMS1 and miR-125a in GC has yet to be determined. Many studies have shown that the enhancer of zeste homolog 2 (EZH2) is an epigenetic modifier, which has been reported that overexpressed in GC tissues and promoted GC proliferation and metastasis.16,17 As one of the enzymatic subunit of the polycomb repressive complex 2 (PRC2), EZH2 has been reported to silence the expression of miR-125a and miR-320c in multiple myeloma.18,19 Whether the miR-125a expression will be inhibited by EZH2 in GC is unknown.
In this study, we evaluated the potential mechanism of miR-125a and BRMS1 in GC and found that BRMS1 may be regulated by CNV, methylation and miR-125a-5p. The histone methylation-modified protein EZH2 is an important regulatory factor that results in the low expression of miR-125a. Epigenetic changes mediate the miR-125a/BRMS1 axis to inhibit the invasion and metastasis of GC cells. In vivo experiments also showed that BRMS1 is involved in invasion and metastasis but not proliferation in GC.
Materials and methods
Patients and tissue samples
Matched GC and adjacent normal gastric tissues were acquired from 31 patients who underwent primary surgical resection at the First Affiliated Hospital of Nanchang University, China, from March 2016 to December 2017. All of the specimens were evaluated by two pathologists according to the American Joint Committee on Cancer (AJCC) TNM staging classification for carcinoma of the stomach (7th ed. 2010). No local or systemic treatments were administered to these patients before surgery. All of the involved patients provided informed consent and a signed agreement. The study protocol was approved by the Ethics Committee of the First Affiliated Hospital of Nanchang University (Nanchang, China).
Cell culture
The human GC cell lines SGC-7901, MKN-28 and BGC-823 (ATCC, Manassas, VA, USA) and the human normal gastric epithelial cell line GES-1 (Shanghai Institute for Life Science, Chinese Academy of Sciences, Shanghai, China) were cultured in RPMI 1640 (Solarbio, Beijing, China) supplemented with 10% fetal bovine serum (Biological industries, USA) and incubated at 37 °C, 5% CO2, and saturated humidity.
Cell transfection
The human miR-125a plasmids, miR-control plasmids, anti-miR-125a and anti-miR-control were constructed as previously described.9 The full-length fragments of BRMS1 were amplified, which were inserted into a pEX-4 vector to generate a BRMS1 overexpression plasmid (GenePharma, Shanghai, China), with empty pEX-4 vectors serving as a negative control. The siRNA small-interfering (si)RNA (#S1 sequence, 5ʹ-GGAAUAAGUACGGAAUGUGA-3ʹ) that targets the BRMS1 gene was synthesized by Guangzhou RiboBio Co., Ltd., (Guangzhou, China) as previously described.9,20 The target siRNA for EZH2 (GenePharma, Shanghai, China) was as follows: shEZH2#1: 5ʹ-CCAACACAAGTCATCCCATTA-3ʹ; shEZH2#2: 5ʹ-TATGATGGTTAACGGTGATCA-3ʹ.
Total RNA extraction and quantitative real-time polymerase chain reaction
Total RNA was extracted from the cultured cells and frozen tissues using TRIzol reagent (Invitrogen, Karlsruhe, Germany) following the manufacturer’s protocol. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed to amplify the targets using a LightCycler® 480 real-time PCR system (Roche Diagnostics, Basel, Switzerland) with specific mRNA/miRNA primers and SYBR® Premix Ex Taq™ II reagents (TaKaRa Bio, Inc.) according to the manufacturer’s protocol. The specific primers used are presented in Table 1. The thermocycler conditions were as follows: 94 °C for 4 min and then 35 cycles of 94°C for 20 sec, 60 °C for 30 sec, and 72 °C for 30 sec.
 |
Table 1 All primers and sequences used in the present study |
DNA extraction, methylation-specific PCR (MSP) and bisulfite sequencing PCR (BSP)
Genomic DNA from patient samples and cell lines was isolated using an Easypure Genomic DNA kit (TransGen Biotech, Beijing, China). We conducted a bisulfite conversion and purification using the EpiTect Bisulfite (QIAGEN, CA) with the procedure according to the product manual. Methylation-specific PCR (MSP) and bisulfite sequencing PCR (BSP) were performed and analyzed as previously described.21,22
5-Aza-2ʹ-deoxyazacytidine demethylation treatment
To block DNA methylation, SGC-7901, MKN-28 and BGC-823 cells were treated with 5-Aza-2ʹ-deoxyazacytidine (Sigma-Aldrich, St Louis, MO) as previously described.22,23
Protein extraction and Western blot
Protein extraction and cell lysates were prepared using RIPA protein extraction reagent (Beyotime, Beijing, China) supplemented with a protease inhibitor cocktail (Transgen Biotech, China). GAPDH (ZSJQB Co., Ltd., Beijing) was used as a control. BRMS1 (ab134968) and EZH2 (ab186006) were purchased from Abcam.
Transwell assay
The cell migration and invasion abilities were detected by a transwell Boyden chamber (8 um pore size, Corning Life Science, USA). The Matrigel was added to the upper chamber for the invasion assay and without Matrigel for migration assays. The experimental procedures refer to the previous studies.24,25
Animal experiments
Female athymic BALB/c nude mice (4 weeks old) were used for the tumor formation assay as previously described.8 Female BALB/c nude mice (age, four weeks, weight, >15g) were purchased from Hunan SJA Laboratory Animal Co., Ltd (Changsha, China), which were randomly divided into four groups (n=5) to establish GC xenografts. Then, the mice were injected subcutaneously or inoculated peritoneally with 6×106 gastric cells in 200ul PBS. Tumor length (L) and width (W) were measured every week. Tumor volume was evaluated using the following formula: tumor volume =LW2π/6. All mice were sacrificed after 30 days, and the peritoneal metastatic nodules were measured. All animal experiments were approved by the Animal Ethics Committee and conducted by the official recommendations for the Care and Use Laboratory Animals of First Affiliated Hospital, Nanchang University.
ChIP
We performed chromatin immunoprecipitation (ChIP) using the EZ ChIPTM Chromatin immunoprecipitation Kit (Millipore, Bedford, MA) for cell line samples according to the previous studies.26,27
Bioinformatics methods
TCGA stomach adenocarcinoma dataset (STAD) was downloaded from cBioportal (http://www.cbioportal.org/). The transcriptional regulatory networks of miRNAs and protein-coding genes from ChIP-seq data were analyzed by ChIPBase (http://rna.sysu.edu.cn/chipbase/index.php). We used the online tool MethPrimer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi) to predict the gene CpG island. The predicted targets of miR-125a and their target sites were analyzed using microRNA, TargetScan and PicTar, and the methylation status of each DNA clone sequence was analyzed comprehensively and comparatively by CpG viewer, QUMA or BiQ analyzer. The Pol II promoter sequences im primary sequence data was searched by Promoter Scan (http://www-bimas.cit.nih.gov/molbio/proscan/).
Statistical analysis
Statistical analyses were performed using Graphpad Prism 7.0 software. Experimental results are presented using means ± SD. Independent-samples Student's t-test or one-way ANOVA were used to analyze the possible differences between groups. The association between miR-125a expression and CNV were analyzed by Spearman’s test (r; P-value). P-values < 0.05 was considered statistically significant.
Results
Evaluation of the potential mechanism of mir-125a and BRMS1 low expression in GC
Chromosomal structural variation and epigenetic modification are two of the main mechanisms leading to abnormally expression of genes.28 In a previous study, we have found that both miR-125a and BRMS1 were expressed at low levels in GC,9 but the mechanism underlying their low expression remains unknown. In this study, we speculate that copy number alternations (CNAs or CNV) or deletion may contribute to the low expression of miR-125a and BRMS1. However, we did not find a significant correlation between CNV and miR-125a expression when we downloaded the CNV and mRNA expression data from TCGA stomach carcinoma dataset (P=0.9442) (Figure 1A). Next, we used 5-Aza-dc to treat SGC7901 and MKN28 and found that the expression of miR-125a and its target gene BRMS1 was altered (Figure 1B and C). Therefore, we hypothesized that the promoter methylation may be the major cause of the low expression of miR-125a. Notably, BRMS1 expression changed much more than miR-125a when we added 5-Aza-dc to SGC7901 and MKN28 cells (Figure 1B and C). To rule out the ability of the demethylating agent 5-Aza-dc to alter the expression of miR-125a which may lead to BRMS1 expression changes, we added anti-miR-125a and found that BRMS1 still presented high expression (Figure 1D). Based on the above analysis, in addition to being regulated by miR-125a, we suspect that BRMS1 may be regulated by other mechanisms such as methylation. To further confirm our conjecture, we analyzed the data from TCGA stomach carcinoma dataset and found that BRMS1 may be dually regulated by CNV(r=0.52, P<0.0001) and promoter methylation (r=−0.21, P<0.0001) (Figure 1E and F).
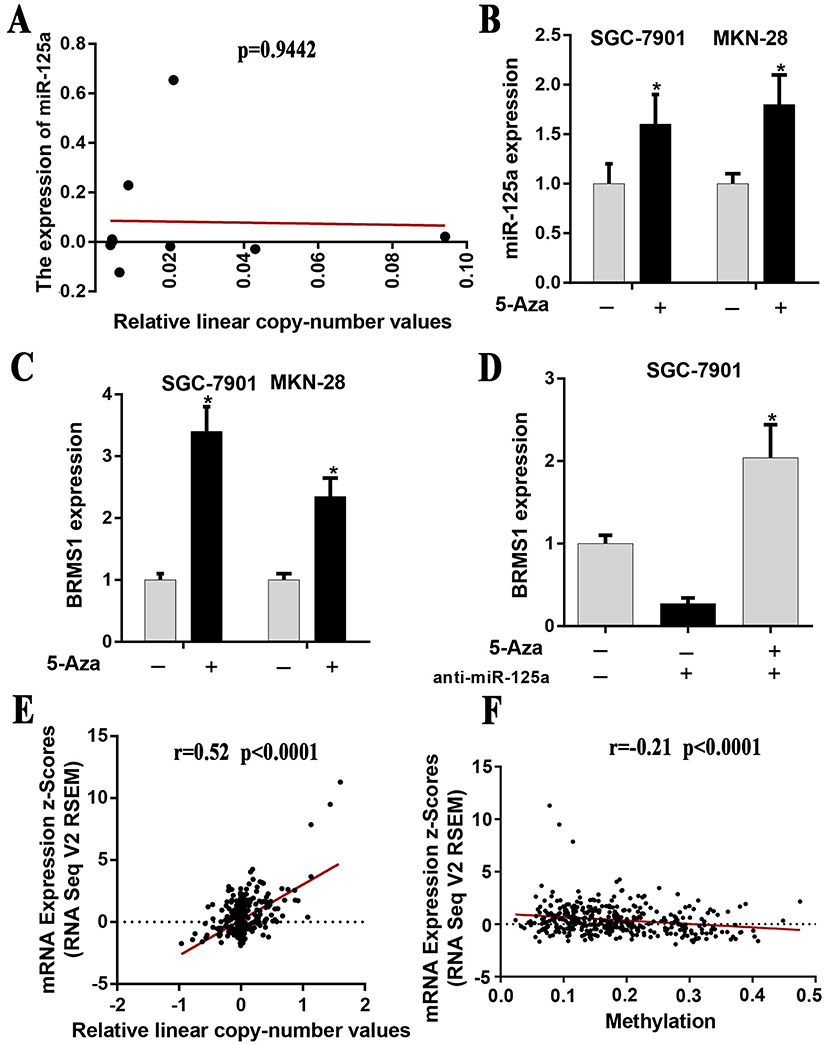 |
Figure 1 Evaluation of the potential mechanism of miR-125a and BRMS1 low expression in GC. (A) The relationship between CNV and miR-125a expression is not significant (P=0.9442). The data were downloaded from TCGA stomach carcinoma dataset. (B) Treatment with 5-Aza significantly increased miR-125a expression compared with untreated cells of SGC-7901 and MKN-28. (C) Treatment with 5-Aza significantly increased BRMS1 expression compared with untreated cells of SGC-7901 and MKN-28. (D) BRMS1 mRNA expression was analyzed by qRT-PCR after treatment with 5-Aza and/or anti-miR-125a in SGC-7901 cells. (E and F) The relationship between BRMS1 expression and CNV/DNA methylation presented a significant correlation (BRMS1 vs CNV: r=0.52, P<0.0001; BRMS1 vs DNA methylation: r=−0.21, P<0.0001). The data were downloaded from TCGA stomach carcinoma dataset. *P<0.05. |
Promoter methylation is not the primary mechanism leading to the low expression of miR-125a
Increasing numbers of studies have revealed that changes in DNA methylation of promoter associated CpG dinucleotides are an important mechanism leading to the dysregulation of miRNAs in cancers.29 Using the online tool MethPrimer30 with the default criteria, we found that miR-125a presented one clear CpG island in its upstream chromosomal sequence (Figure 2A). To further confirm whether CpG island hypermethylation of the miR-125a promoter may suppress its expression, we carried out bisulfite sequencing (BSP) of CpG islands in the GES-1, SGC-7901 and MKN-28 cell lines (Figure 2B). By statistical analysis, the methylation level between GES-1 and SGC-7901, MKN-28 showed no significant difference (P>0.05) (Figure 2C). To test whether the hypermethylation of miR-125a also occurs in primary GC tissues, primary GC tissues and paired para-carcinoma tissues were used to examine DNA methylation status and expression patterns of miR-125a by BSP and qRT-PCR analysis. The qRT-PCR results showed that the expression levels of miR-125a in the 3 of 4 primary tumors were reduced compared with those of the para-carcinoma tissues (P<0.05) (Figure 2D). The methylation level of miR-125a in the primary GC was the same as that in the para-carcinoma tissues, but there was no statistically significant difference (P=0.1671) (Figure 2E and F). All of the results suggest that promoter methylation is not the primary mechanism leading to the low expression of miR-125a.
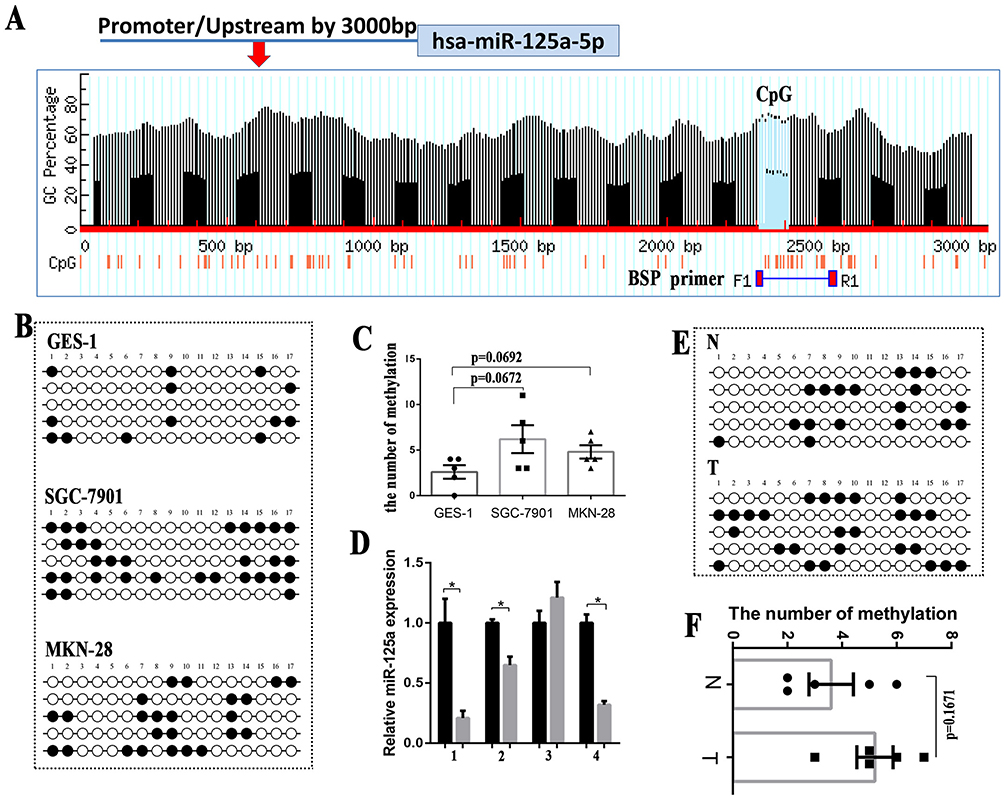 |
Figure 2 Promoter methylation is not the primary mechanism leading to the low expression of miR-125a. (A) Bioinformatic prediction of one CpG island upstream of miR-125a. (B) The miR-125a methylation status of GES-1, SGC-7901 and MKN-28 cells was analyzed by BSP. (C) The methylation level between GES-1 and SGC-7901 or GES-1 and MKN-28 showed no significant difference (P>0.05). (D) The qRT-PCR results showed that the expression levels of miR-125a in the 3 of 4 primary tumors were reduced compared with the paracarcinoma tissues (P<0.05). (E) The miR-125a methylation status in 4 paired GC tissues and para-carcinoma tissues was analyzed by BSP. (F) The statistical analysis showed no significant difference in the number of methylation between the GC tissues and paracarcinoma tissues(P=0.1671). *P<0.05. |
EZH2 is an important molecule affecting the low expression of miR-125a
In addition to promoter methylation, histone methylation may be another important mechanism for the silencing of tumor suppressor genes, including microRNA.29,31 It has been reported that EZH2 inhibition downregulated the expression of miR-125a and miR-320c in multiple myeloma.18,19 Therefore, we speculate whether there is a correlation between miR-125a and EZH2 in GC. As was shown in Figure 3A, there was a negative correlation between EZH2 and miR-125a in GC from TCGA Pan-Cancer (r=−0.216, P=4.06x10−6)32 which was analyzed by ChIPbase.33 Our test results in 31 paired GC tissues analyzed by qPCR also confirmed the conclusion (P<0.05). To further explore whether miR-125a targets EZH2 and downregulates its expression, we searched for it in popular miRNA target databases (microRNA.org, TargetScan, PITA) but did not find a target relationship between them (Figure 3B). As shown in Figure 3C, EZH2 expression did not present a significant change regardless of whether we added miR-125a mimics or anti-miR-125a (P>0.05)(Figure 3C). All the results implied that miR-125a may not be upstream of EZH2 and regulate its expression. Therefore, is EZH2 located in the upstream of miR-125a and does EZH2 regulate its expression?
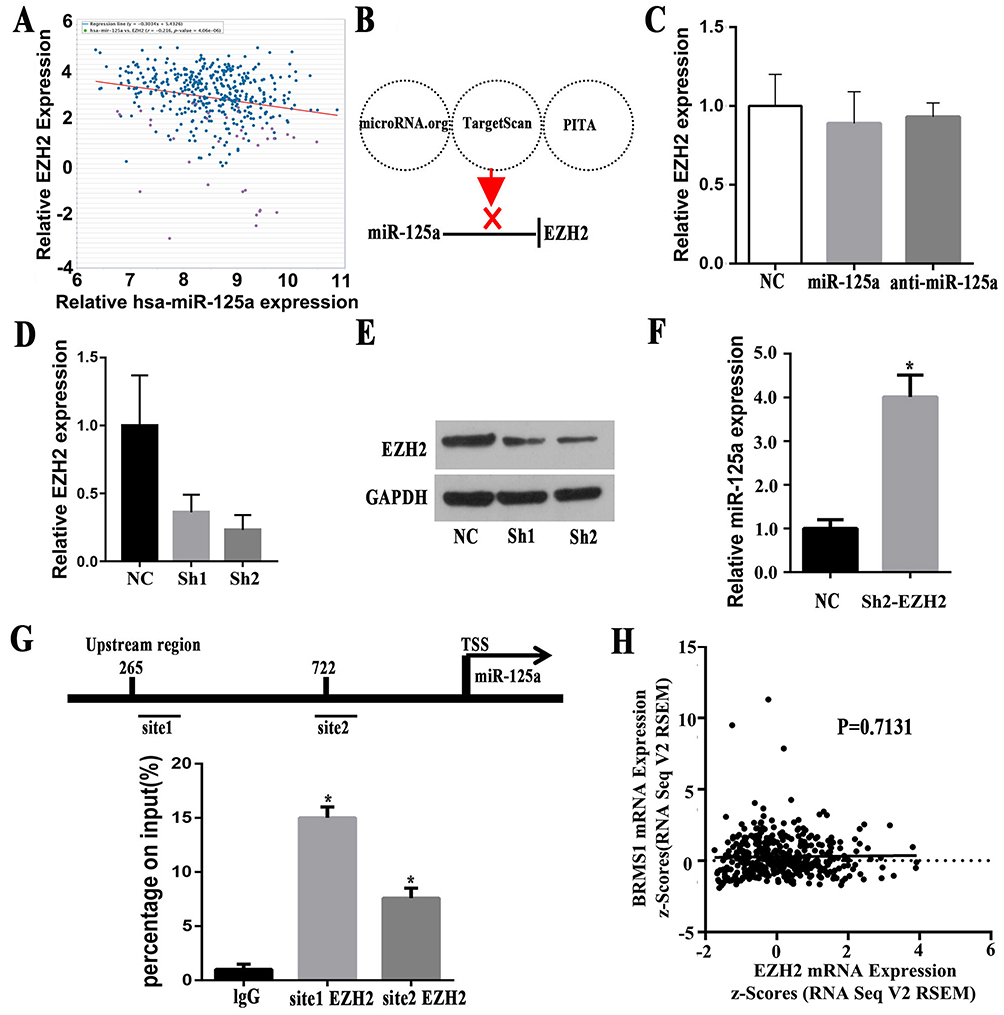 |
Figure 3 EZH2 is an important molecule affecting the low expression of miR-125a. (A) The expression of EZH2 and miR-125a in GC from TCGA Pan-Cancer, which was analyzed by ChIPbase (r=−0.216, P=4.06x10−6). (B) MiR-125a did no target EZH2, as shown by microRNA.org, TargetScan and PITA database analysis. (C) EZH2 expression did not present a significant change with/without miR-125a mimics or anti-miR-125a (P>0.05) (Figure 3C). (D and E) qRT-PCR and Western blot analysis of EZH2 expression in SGC-7901 which was stably transduced with lentiviral vectors carrying EZH2-specific small hairpin RNAs (shEZH2). (F) Decreasing the expression of EZH2 results in a markedly improved the expression of miR-125a compared with the NC group. (G) Bioinformatic prediction of two sites indicates possible enrichment of EZH2 in the promoter region of miR-125a, and ChIP assays found that the miR-125a promoter region was enriched for EZH2 in GC. (H) Bioinformatic analysis showed that there is no significant correlation between EZH2 and BRMS1 expression (P=0.7131). The data were downloaded from TCGA stomach carcinoma dataset. *P<0.05. |
To further confirm this speculation, two RNAi sequences targeting EZH2 were synthesized and packed into lentiviruses and successfully interfered with the expression of EZH2, as shown by qRT-PCR and Western blots (Figure 3D and E). We selected the sh2 with the higher interference efficiency for the subsequent experiments. As shown in Figure 3F, decreasing the expression of EZH2 markedly improved the expression of miR-125a compared with the NC group (P<0.05), which suggests that EZH2 may regulate miR-125a expression. The EZH2 enrichment region on the promoter region of miR-125a was also found on the Promoter Scan dababase.32 To confirm that EZH2 binds to histone methylation in the promoter region of miR-125a, we performed a ChIP assay and found that the miR-125a promoter region was enriched in EZH2 in GC (Figure 3G). Notably, there was no significant correlation between EZH2 and BRMS1 expression by analyze the data downloaded from TCGA database (P=0.7131) (Figure 3H), which implies that BRMS1 may be mainly regulated by promoter methylation, CNV and miR-125a-5p.
Promoter methylation affects the expression of BRMS1
The above analysis suggests that promoter methylation may affect the expression of BRMS1. We predicted the CpG island 1000 bp above the BRMS1 transcription initiation site by MethPrimer30 and found that the CpG island is mainly concentrated in the 800 bp above the transcription start site (Figure 4A). By comparison, we confirmed that the promoter of BRMS1, which was analyzed by Promoter Scan,34 lies in the range of the CpG islands. To further investigate the relationship between BRMS1 and DNA promoter methylation, we carried out a BSP test in MKN-28, SGC-7901 and GES-1 cells. The results showed higher levels of DNA methylation in MKN-28 and SGC-7901 cell lines than GES-1; however, BRMS1 expression was lower in the MKN-28 and SGC-7901 cell lines than the GES-1 cell line (P<0.05), in which only limited CpG island methylation was found (Figure 4B and C). To explore whether methylation was also present in the GC tissues, we performed MSP experiments in the 31 paired GC tissues and paracarcinoma tissues. The methylation level in the GC tissues was higher than that in the para-carcinoma tissues (51.6% vs 16.1%), which is the same as the results in the cell lines (Figure 4D). Two paired GC tissues were selected for further BSP testing, and the results also showed significant methylation in GC tissues compared with the paracarcinoma tissues (Figure 4E and F).
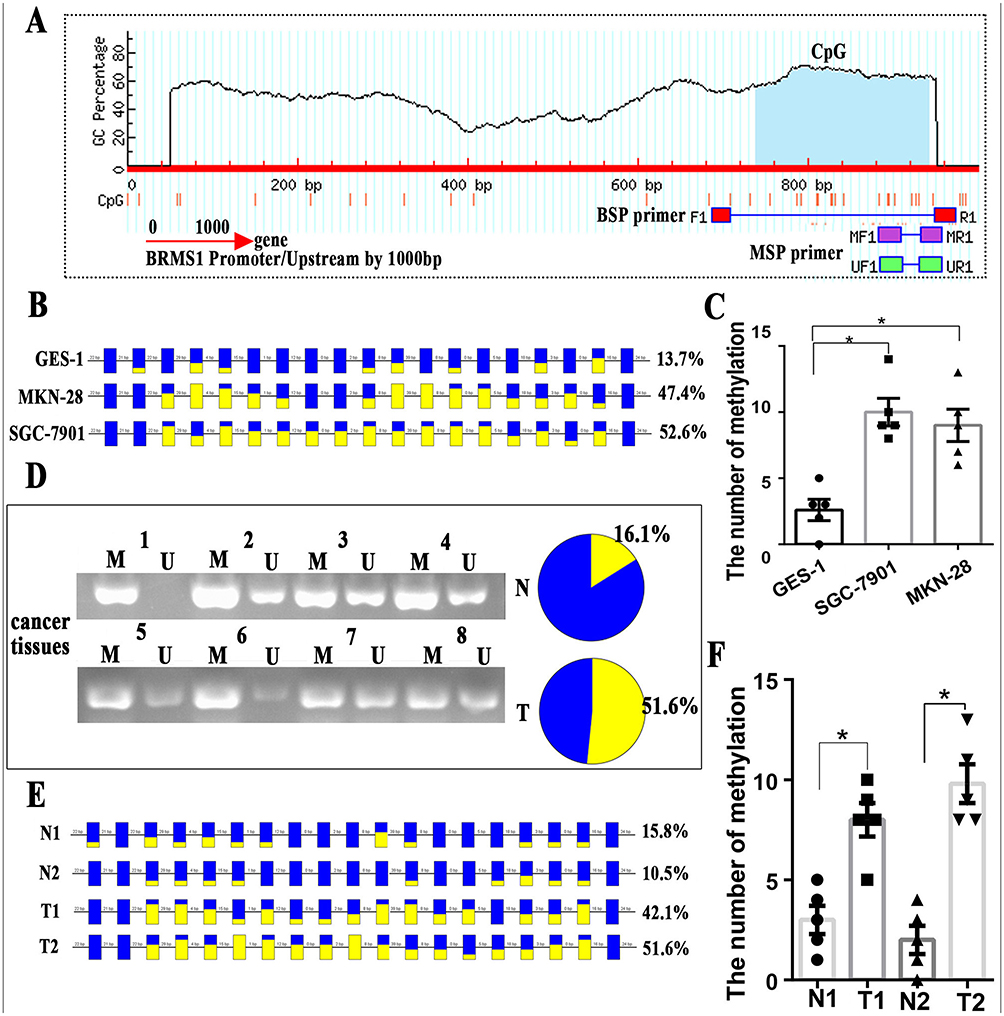 |
Figure 4 Promoter methylation affects the expression of BRMS1. (A) Bioinformatic prediction of one CpG island upstream of BRMS1, which is mainly concentrated 300 bp above the transcription start site. (B) The bisulfite sequencing of the BRMS1 upstream region in MKN-28, SGC-7901 and GES-1 cell lines analyzed by BiQ Analyzer. (C) The number of methylations was higher in MKN-28 and SGC-7901 than in GES-1. (D) Representative MSP analyses for BRMS1 methylation in GC tissues. U, unmethylated state; M, methylated state. (E) The bisulfite sequencing of the BRMS1 upstream region in 2 paired GC tissues and paracarcinoma tissues analyzed by BiQ Analyzer. Each square represents a CpG site. For the figure B and E, methylated CpG dinucleotides are represented by yellow squares whereas unmethylated CpG sites are represented by blue squares. If the methylation state of a CpG site could not be defined, it is represented as not present. (F) The number of methylations was higher in GC tissues than in para-carcinoma tissues. *P<0.05. |
Epigenetic regulation of the miR-125a/BRMS1 axis
Based on the above findings, we speculate that EZH2 may regulate the miR-125a/BRMS1 axis, which is also influenced by DNA methylation, as shown in Figure 5A. To further evaluate the role of EZH2 in the miR-125a/BRMS1 axis, we knocked down EZH2 expression in SGC-7901 cells by interference lentiviruses and found that not only was miR-125a expression changed, but also the target gene BRMS1 expression increased. Interestingly, BRMS1 expression was partly reversed when we added miR-125a inhibitor to inhibit miR-125a expression (Figure 5B and C). Subsequently, the 5-Aza-dc was added to SGC-7901, and we found that BRMS1 expression increased, along with decreased cell invasion (Figure 5D).
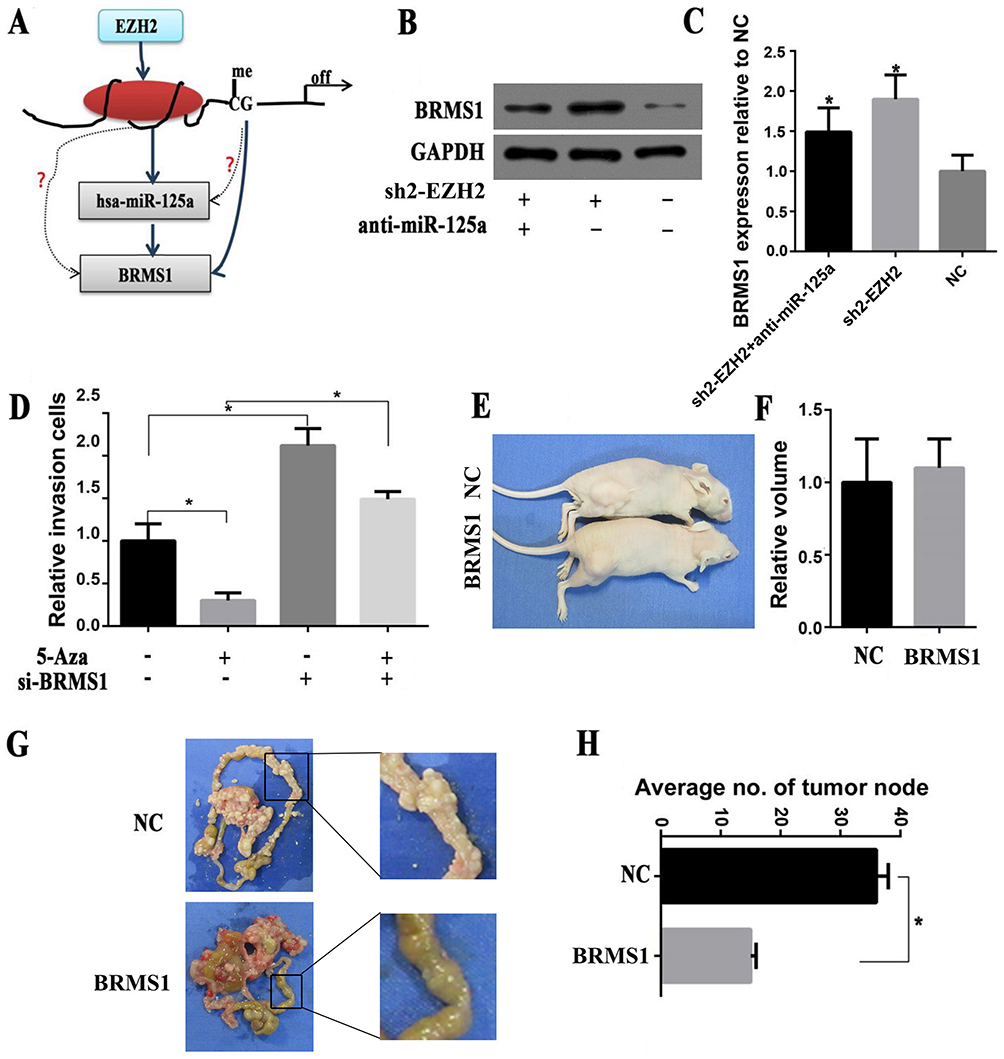 |
Figure 5 Epigenetic regulation of the miR-125a/BRMS 1 axis to promote GC invasion and metastasis in vitro and in vivo. (A) Working model for the regulation of the miR-125a/BRMS1 axis by EZH2 in GC. (B and C) Western blot analysis of the expression of BRMS1 treated with/without Sh2-EZH2/anti-miR-125a. (D) Invasion assay analysis of the invasive cells in SGC-7901 cells treated with/without 5-Aza/siBRMS1. (E and F) Representative images of metastatic tumors in nude mice inoculated with NC/shBRMS1. The volume of metastatic tumors between NC and shBRMS1 was no significantly different (P>0.05). (G and H) Representative images of metastatic tumors in the mesentery and intestine of nude mice inoculated with NC/shBRMS1. The number of tumor nodes was significantly lower in the siBRMS1 group than in the NC group (P<0.05). *P<0.05. |
BRMS1 is involved in GC invasion and metastasis rather than proliferation in vivo
In a previous study, we have investigated the role of BRMS1 in GC invasion and metastasis in vitro.8 To further confirm this conclusion, we selected the nude mice for in vivo experiments and found that overexpression of BRMS1 did not affect the proliferation of GC cells compared with the control group (P>0.05) (Figure 5E and F). However, there were fewer tumor nodes compared to the control group when we upregulated BRMS1 expression in GC cells and inoculate them in the mice peritoneal (P<0.05) (Figure 5G and H).
Discussion
As a tumor suppressor gene discovered over a decade ago, BRMS1 has been reported to suppress metastasis not only in breast cancer but also in hepatocellular carcinoma, non-small cell lung cancer, ovarian cancer and melanoma.6,7 Recently, we and others have demonstrated that BRMS1 expression was markedly stronger in the primary GC compared with metastasis GC and improve BRMS1 expression reduced the numbers of metastasis tumors in xenograft model.8,10 BRMS1 also can be recognized as a biomarker to predict the platinum-resistance for the patients of GC.35 Our previous research also revealed that both miR-125a-5p and BRMS1 were low expression in GC tissues and miR-125a may act as a tumor suppressor by targeting BRMS1, which can further suppress metastasis of GC cells,9 but the mechanisms for these genes are unknown.
Epigenetics plays an important role in the silencing of critical tumor suppressor genes and the activation of oncogenes leading to carcinogenesis, which is not accompanied by modification in the DNA sequence.13 Various biochemical pathways that are essential for tumorigenesis are regulated by epigenetic phenomena, such as histone modifications, nucleosome remodeling, DNA methylation and miRNA mediated targeting of various genes.36,37 However, somatic CNAs of genes are widespread genetic events in cancers that may result in the activation of oncogenes and inactivation of tumor-suppressor genes.38 To evaluate the relationship between CNAs and the expression of miR-125a, we analyzed the data downloaded from TCGA stomach carcinoma dataset, but there was no significant difference between them. DNA methylation is a common epigenetic alternation and plays a key role in the silencing of numerous cancer-related genes, thereby affecting a number of vital cellular processes, including the cell cycle checkpoint, apoptosis, signal transduction, cell adhesion and angiogenesis.39 Considering the role of DNA methylation in tumorigenesis, we treated the SGC-7901 and BGC-823 cell lines with 5-Aza-dc and found that BRMS1 expression changed much more than miR-125a. BRMS1 expression remained high when miR-125a inhibitors were added to cell lines treated with 5-Aza-dc. These results showed that the low expression of BRMS1 may not only be regulated by miR-125a but also by DNA methylation and CNV. The bioinformatics analyses from TCGA also confirmed our findings.
DNA methylation of the promoter CpG island has been shown to be a key mediator underlying the downregulation of microRNA expression.31 The bioinformatics results from MethPrimer implied that the promoter of miR-125a contains CpG islands. To explore whether this CpG island will be methylated and play a key role in the low expression of miR-125a, BSP analysis was performed in GC tissues and cell lines. The results showed that there were no significantly difference for the methylation status of miR-125a in GC cancer tissues and cell lines. Although some reports have shown that low expression of miR-125a is associated with DNA methylation in colorectal cancer, multiple myeloma and so on,40,41 the same phenomenon was not observed in our studies. Some reasons may explain our results. First, only 4 paired GC tissues were tested in this study. More paired GC tissues will be tested in future studies to illuminate the relationship between DNA methylation and miR-125a expression. Second, we only selected MKN-28, SGC-7901 and GES-1 cell lines in this study. The miR-125a promoter methylation level in MKN-28 and SGC-7901 cells may not increase as in other GC cell lines. In conclusion, DNA promoter methylation may not be the primary cause of the low expression of miR-125a.
Histone modifications can lead to activation or repression of gene expression, depending on the type and position of attached functional groups and amino acid residues that are modified.42,43 EZH2, also called histone lysine methyltransferase (HKMT), was cloned and identified as a gene belonging to the Polycomb group of genes, which epigenetically silences the expression of some tumor suppressor genes (TSGs).44 High expression of EZH2 was associated with poor prognosis in GC.45,46 Knockdown of EZH2 by siRNA has been demonstrated to inhibit GC cell proliferation, colony formation, migration and invasion.16 Considering the critical role of EZH2 in GC, we analyzed data of the correlation between EZH2 and miR-125a in data from the Chipbase datasets and by qPCR analyses of 31 paired GC tissues. The results showed that there is a negative correlation between them. However, the bioinformatics results did not identify EZH2 as a potential miR-125a target. Moreover, overexpression or inhibition of miR-125a did not influence EZH2 expression. All these results implied that EZH2 may not be regulated by miR-125a. Although miRNAs regulate multiple target genes by sequence-specific interactions with the miRNA response elements (MRE) on the 3ʹ untranslated regions (3ʹ-UTR) of related mRNA targets,47,48 we did not find evidence for a direct effect. However, downregulation of EZH2 significantly increased miR-125a expression and the ChIP assay indicated that the miR-125a promoter region was enriched in EZH2 in GC. The results showed that EZH2 may be located in upstream of miR-125a and mediate miR-125a gene expression. The previous studies also reported that EZH2 could regulate the methylation of microRNAs, including miR-200b/a/429, miR-34a, miR-361 and so on.49–51 For this study, we have revealed the targeted relationship in which EZH2 regulates miR-125a expression, which is vrified the previous research18. In brief, the above studies elucidated the regulatory relationships between histone EZH2 and microRNA.
In addition, we did not find a correlation between EZH2 and BRMS1 by bioinformatics analysis, which means that EZH2 may not regulate BRMS1 expression, but DNA promoter methylation, CNV and miR-125a may be the primary cause for the low expression of BRMS1. Previous studies have reported that BRMS1 is downregulated by DNA methylation in breast, non-small cell lung and cervical cancer and others.14,52,53 However, there is no evidence that promoter methylation is associated with low expression of BRMS1 in GC in previous studies. In this study, to confirm the above conjecture, BSP and MSP analyses were performed in GC cell lines and GC tissues, which showed that the promoter methylation level of BRMS1 is higher in GC cell lines (MKN-28 and SGC-7901) and GC tissues than GES-1 and the paired normal gastric tissues. The results obtained in this study combined with our observations indicated that EZH2 may regulate the DNA promoter of miR-125a, while BRMS1 may be mainly regulated by promoter methylation but not EZH2.
In a previous study, we have confirmed that BRMS1 is a potential target gene of miR-125a.9 Based on previous reports and the foregoing content of this study, we speculate that the miR-125a/BRMS1 axis may be regulated by epigenetics. Knockdown of EZH2 expression altered the expression of miR-125a, and the expression of the target gene BRMS1 was increased. Interestingly, BRMS1 was partly reversed when we added anti-miR-125a to decrease miR-125a expression. Different combinations of 5-Aza and siBRMS1 showed that the miR-125a/BRMS1 axis was regulated by epigenetics, which further influenced GC metastasis and invasion. To further explore whether similar phenomena can occur in vivo, BRMS1 low/high expression cell lines were administered to nude mice. These results showed that increased BRMS1 expression did not affect the proliferation of GC cells but suppressed metastasis of tumor cells, which further verifies the previous studies.54–56
Conclusion
In summary, we have confirmed that EZH2 regulates the expression of miR-125a, while BRMS1 may be mainly regulated by promoter methylation, CNV and miR-125a-5p in GC. An epigenetic mechanism mediates the miR-125a/BRMS1 axis to inhibit the invasion and metastasis of GC cells. These findings help illuminate the mechanism of BRMS1 inhibition of GC invasion and metastasis and contribution to the development of new drugs targeting the miR-125a/BRMS1 axis, which will be a promising therapeutic strategy for GC and other human cancers.
Ethics approval and consent to participate
The research protocol was reviewed and approved by the Ethical Committee and Institutional Review Board of the First Affiliated Hospital of Nanchang University, and written informed consent was obtained from each patient included in the study.
Acknowledgments
This work was supported by the National Science Foundation of China (no. 81460373 and 81860428), the Education Department of Jiangxi Province Science and Technology Research Projects (No. GJJ160246), the “Talent 555 Project” of Jiangxi Province, People’s Republic of China, the Science and Technology Plan of Health Commission of Jiangxi Province (No.20191026), the Spark Promotion Plan of Grassroots Health Appropriate Technology of Health Commission of Jiangxi Province (No.20198012) and the Graduate Student Innovation Special Fund Project of Jiangxi Province (No. YC2018-S112).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
2. Allemani C, Matsuda T, Di Carlo V, et al. Global surveillance of trends in cancer survival 2000-14 (CONCORD-3): analysis of individual records for 37 513 025 patients diagnosed with one of 18 cancers from 322 population-based registries in 71 countries. Lancet. 2018;391(10125):1023–1075. doi:10.1016/S0140-6736(17)33326-3
3. Chen W, Sun K, Zheng R, et al. Cancer incidence and mortality in China, 2014. Chin J Cancer Res. 2018;30(1):1–12. doi:10.21147/j.issn.1000-9604.2018.01.01
4. Song Z, Wu Y, Yang J, Yang D, Fang X. Progress in the treatment of advanced gastric cancer. Tumour Biol. 2017;39(7):1010428317714626. doi:10.1177/1010428317714626.
5. Seraj MJ, Samant RS, Verderame MF, Welch DR. Functional evidence for a novel human breast carcinoma metastasis suppressor, BRMS1, encoded at chromosome 11q13. Cancer Res. 2000;60(11):2764–2769.
6. Welch DR, Manton CA, Hurst DR. Breast cancer metastasis suppressor 1 (BRMS1): robust biological and pathological data, but still enigmatic mechanism of action. Adv Cancer Res. 2016;132:111–137. doi:10.1016/bs.acr.2016.05.003
7. Kodura MA, Souchelnytskyi S. Breast carcinoma metastasis suppressor gene 1 (BRMS1): update on its role as the suppressor of cancer metastases. Cancer Metastasis Rev. 2015;34(4):611–618. doi:10.1007/s10555-015-9583-z
8. Jie Z, Chen X, Li Z, et al. Effect of breast-cancer metastasis suppressor 1 (BRMS1) on growth and metastasis of human gastric cancer cells in vivo. Chinese Ger J Clin Oncol. 2012;11(10):592–597. doi:10.1007/s10330-011-0920-6
9. Cao Y, Tan S, Tu Y, et al. MicroRNA-125a-5p inhibits invasion and metastasis of gastric cancer cells by targeting BRMS1 expression. Oncol Lett. 2018;15(4):5119–5130. doi:10.3892/ol.2018.7983
10. Guo XL, Wang YJ, Cui PL, et al. Effect of BRMS1 expression on proliferation, migration and adhesion of mouse forestomach carcinoma. Asian Pac J Trop Med. 2015;8(9):724–730. doi:10.1016/j.apjtm.2015.07.020
11. Hirst M, Marra MA. Epigenetics and human disease. Int J Biochem Cell Biol. 2009;41(1):136–146. doi:10.1016/j.biocel.2008.09.011
12. Mummaneni P, Shord SS. Epigenetics and oncology. Pharmacotherapy. 2014;34(5):495–505. doi:10.1002/phar.2014.34.issue-5
13. Puneet KHR, Kumari S, Tiwari S, Khanna A, Narayan G. Epigenetic mechanisms and events in gastric cancer-emerging novel biomarkers. Pathol Oncol Res. 2018;24(4):757–770. doi:10.1007/s12253-018-0410-z
14. Panagopoulou M, Lambropoulou M, Balgkouranidou I, et al. Gene promoter methylation and protein expression of BRMS1 in uterine cervix in relation to high-risk human papilloma virus infection and cancer. %A Panagopoulou M. Tumour Biol. 2017;39(4):1010428317697557. doi:10.1177/1010428317697557
15. Fan B, Dachrut S, Coral H, et al. Integration of DNA copy number alterations and transcriptional expression analysis in human gastric cancer. PLoS One. 2012;7(4):e29824. doi:10.1371/journal.pone.0029824
16. Gan L, Xu M, Hua R, et al. The polycomb group protein EZH2 induces epithelial-mesenchymal transition and pluripotent phenotype of gastric cancer cells by binding to PTEN promoter. J Hematol Oncol. 2018;11(1):9. doi:10.1186/s13045-017-0547-3
17. Xu J, Wang Z, Lu W, et al. EZH2 promotes gastric cancer cells proliferation by repressing p21 expression. Pathol Res Pract. 2019;215(6):152374. doi:10.1016/j.prp.2019.03.003
18. Alzrigat M, Jernberg-Wiklund H. The miR-125a and miR-320c are potential tumor suppressor microRNAs epigenetically silenced by the polycomb repressive complex 2 in multiple myeloma. RNA Dis. 2017;4:2.
19. Alzrigat M, Parraga AA, Agarwal P, et al. EZH2 inhibition in multiple myeloma downregulates myeloma associated oncogenes and upregulates microRNAs with potential tumor suppressor functions. Oncotarget. 2017;8(6):10213–10224. doi:10.18632/oncotarget.14378
20. Wu Y, Jiang W, Wang Y, et al. Breast cancer metastasis suppressor 1 regulates hepatocellular carcinoma cell apoptosis via suppressing osteopontin expression. PLoS One. 2012;7(8):e42976. doi:10.1371/journal.pone.0042976
21. Ma G, Liu H, Hua Q, et al. KCNMA1 cooperating with PTK2 is a novel tumor suppressor in gastric cancer and is associated with disease outcome. Mol Cancer. 2017;16(1):46. doi:10.1186/s12943-017-0613-z
22. Lin X, Li F, Xu F, et al. Aberration methylation of miR-34b was involved in regulating vascular calcification by targeting Notch1. Aging (Albany NY). 2019;11(10):3182–3197. doi:10.18632/aging.101973
23. Luo M, Li Y, Shi X, et al. Aberrant methylation of EYA4 promotes epithelial-mesenchymal transition in esophageal squamous cell carcinoma. Cancer Sci. 2018;109(6):1811–1824. doi:10.1111/cas.13615
24. Dai H, Shen K, Yang Y, et al. PUM1 knockdown prevents tumor progression by activating the PERK/eIF2/ATF4 signaling pathway in pancreatic adenocarcinoma cells. Cell death & disease. 2019;10(8):595.
25. Wu J, Long Z, Cai H, Yu S, Liu X. Homeobox B7 accelerates the cancer progression of gastric carcinoma cells by promoting epithelial-mesenchymal transition (EMT) and activating Src-FAK pathway. OncoTargets and therapy. 2019;12:3743-3751.
26. Niu Y, Ma F, Huang W, et al. Long non-coding RNA TUG1 is involved in cell growth and chemoresistance of small cell lung cancer by regulating LIMK2b via EZH2. Mol Cancer. 2017;16(1):5. doi:10.1186/s12943-016-0575-6
27. Qi M, Hu J, Cui Y, et al. CUL4B promotes prostate cancer progression by forming positive feedback loop with SOX4. Oncogenesis. 2019;8(3):23. doi:10.1038/s41389-019-0131-5
28. Nebbioso A, Tambaro FP, Dell’Aversana C, Altucci L. Cancer epigenetics: moving forward. PLoS Genet. 2018;14(6):e1007362. doi:10.1371/journal.pgen.1007664
29. Morales S, Monzo M, Navarro A. Epigenetic regulation mechanisms of microRNA expression. Biomol Concepts. 2017;8(5–6):203–212. doi:10.1515/bmc-2017-0024
30. Li LC, Dahiya R. MethPrimer: designing primers for methylation PCRs. Bioinformatics. 2002;18(11):1427–1431. doi:10.1093/bioinformatics/18.11.1427
31. Ramassone A, Pagotto S, Veronese A, Visone R. Epigenetics and microRNAs in cancer. Int J Mol Sci. 2018;19(2):459. doi:10.3390/ijms19020459
32. Vivian J, Rao A, Nothaft FA, et al. Rapid and efficient analysis of 20,000 RNA-seq samples with Toil. bioRxiv. 2016;062497.
33. Zhou K-R, Liu S, Sun W-J, et al. ChIPBase v2.0: decoding transcriptional regulatory networks of non-coding RNAs and protein-coding genes from ChIP-seq data. Nucleic Acids Res. 2017;45(D1):D43–D50. doi:10.1093/nar/gkw965
34. Prestridge DS. Predicting Pol II promoter sequences using transcription factor binding sites. J Mol Biol. 1995;249(5):923–932. doi:10.1006/jmbi.1995.0349
35. Pan J, Xiang Z, Dai Q, Wang Z, Liu B, Li C. Prediction of platinum-resistance patients of gastric cancer using bioinformatics. J Cell Biochem. 2019;120(8):13478–13486. doi:10.1002/jcb.28621
36. Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150(1):12–27. doi:10.1016/j.cell.2012.06.013
37. Biswas S, Rao CM. Epigenetics in cancer: fundamentals and beyond. Pharmacol Ther. 2017;173:118–134. doi:10.1016/j.pharmthera.2017.02.011
38. Beroukhim R, Mermel CH, Porter D, et al. The landscape of somatic copy-number alteration across human cancers. Nature. 2010;463:899. doi:10.1038/nature08700
39. Suzuki H, Maruyama R, Yamamoto E, Kai M. DNA methylation and microRNA dysregulation in cancer. Mol Oncol. 2012;6(6):567–578. doi:10.1016/j.molonc.2012.07.007
40. Bi C, Chung TH, Huang G, et al. Genome-wide pharmacologic unmasking identifies tumor suppressive microRNAs in multiple myeloma. Oncotarget. 2015;6(28):26508–26518. doi:10.18632/oncotarget.4769
41. Chen H, Xu Z. Hypermethylation-associated silencing of miR-125a and miR-125b: a potential marker in colorectal cancer. Dis Markers. 2015;2015:345080. doi:10.1155/2015/105358
42. Kurdistani SK. Histone modifications in cancer biology and prognosis. Prog Drug Res. 2011;67:91–106.
43. Nowacka-Zawisza M, Wisnik E. DNA methylation and histone modifications as epigenetic regulation in prostate cancer (Review). Oncol Rep. 2017;38(5):2587–2596. doi:10.3892/or.2017.5972
44. Pan Y-M, Wang C-G, Zhu M, et al. STAT3 signaling drives EZH2 transcriptional activation and mediates poor prognosis in gastric cancer. Mol Cancer. 2016;15(1):79. doi:10.1186/s12943-016-0561-z
45. Matsukawa Y, Semba S, Kato H, Ito A, Yanagihara K, Yokozaki H. Expression of the enhancer of zeste homolog 2 is correlated with poor prognosis in human gastric cancer. Cancer Sci. 2006;97(6):484–491. doi:10.1111/j.1349-7006.2006.00203.x
46. Cai GH, Wang K, Miao Q, Peng YS, Chen XY. Expression of polycomb protein EZH2 in multi-stage tissues of gastric carcinogenesis. J Dig Dis. 2010;11(2):88–93. doi:10.1111/j.1751-2980.2010.00420.x
47. Li G, Li L, Sun Q, et al. MicroRNA-3200-5p promotes osteosarcoma cell invasion via suppression of BRMS1. Mol Cells. 2018;41(6):523–531. doi:10.14348/molcells.2018.2200
48. Sun F, Yu M, Yu J, et al. miR-338-3p functions as a tumor suppressor in gastric cancer by targeting PTP1B. Cell Death Dis. 2018;9(5):522. doi:10.1038/s41419-018-0611-0
49. Ning X, Shi Z, Liu X, et al. DNMT1 and EZH2 mediated methylation silences the microRNA-200b/a/429 gene and promotes tumor progression. Cancer Lett. 2015;359(2):198–205. doi:10.1016/j.canlet.2015.01.005
50. Kwon H, Song K, Han C, et al. Epigenetic silencing of miRNA-34a in human cholangiocarcinoma via EZH2 and DNA methylation: impact on regulation of notch pathway. Am J Pathol. 2017;187(10):2288–2299. doi:10.1016/j.ajpath.2017.06.014
51. Ihira K, Dong P, Xiong Y, et al. EZH2 inhibition suppresses endometrial cancer progression via miR-361/Twist axis. Oncotarget. 2017;8(8):13509–13520. doi:10.18632/oncotarget.14586
52. Chimonidou M, Kallergi G, Georgoulias V, Welch DR, Lianidou ES. Breast cancer metastasis suppressor-1 promoter methylation in primary breast tumors and corresponding circulating tumor cells. Mol Cancer Res. 2013;11(10):1248–1257. doi:10.1158/1541-7786.MCR-13-0096
53. Balgkouranidou I, Chimonidou M, Milaki G, et al. Breast cancer metastasis suppressor-1 promoter methylation in cell-free DNA provides prognostic information in non-small cell lung cancer. Br J Cancer. 2014;110(8):2054–2062. doi:10.1038/bjc.2014.104
54. Smith PW, Liu Y, Siefert SA, Moskaluk CA, Petroni GR, Jones DR. Breast cancer metastasis suppressor 1 (BRMS1) suppresses metastasis and correlates with improved patient survival in non-small cell lung cancer. Cancer Lett. 2009;276(2):196–203. doi:10.1016/j.canlet.2008.11.024
55. Sheng XJ, Zhou DM, Liu Q, Lou SY, Song QY, Zhou YQ. BRMS1 inhibits expression of NF-kappaB subunit p65, uPA and OPN in ovarian cancer cells. Eur J Gynaecol Oncol. 2014;35(3):236–242.
56. Ventura BV, Quezada C, Maloney SC, et al. Expression of the metastasis suppressor BRMS1 in uveal melanoma. Ecancermedicalscience. 2014;8:410.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.