Back to Journals » Journal of Asthma and Allergy » Volume 16
Epidemiology and Immunopathogenesis of Virus Associated Asthma Exacerbations
Authors Bakakos A ![]() , Sotiropoulou Z, Vontetsianos A, Zaneli S, Papaioannou AI
, Sotiropoulou Z, Vontetsianos A, Zaneli S, Papaioannou AI ![]() , Bakakos P
, Bakakos P ![]()
Received 14 July 2023
Accepted for publication 16 September 2023
Published 26 September 2023 Volume 2023:16 Pages 1025—1040
DOI https://doi.org/10.2147/JAA.S277455
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Amrita Dosanjh
Agamemnon Bakakos, Zoi Sotiropoulou, Angelos Vontetsianos, Stavroula Zaneli, Andriana I Papaioannou, Petros Bakakos
1st University Department of Respiratory Medicine, National and Kapodistrian University of Athens, Athens, 11527, Greece
Correspondence: Agamemnon Bakakos, 1st University Department of Respiratory Medicine, National and Kapodistrian University of Athens, Kononos 11 Street, Athens, 11634, Greece, Tel +306972455781, Email [email protected]
Abstract: Asthma is a common airway disease, affecting millions of people worldwide. Although most asthma patients experience mild symptoms, it is characterized by variable airflow limitation, which can occasionally become life threatening in the case of a severe exacerbation. The commonest triggers of asthma exacerbations in both children and adults are viral infections. In this review article, we will try to investigate the most common viruses triggering asthma exacerbations and their role in asthma immunopathogenesis, since viral infections in young adults are thought to trigger the development of asthma either right away after the infection or at a later stage of their life. The commonest viral pathogens associated with asthma include the respiratory syncytial virus, rhinoviruses, influenza and parainfluenza virus, metapneumovirus and coronaviruses. All these viruses exploit different molecular pathways to infiltrate the host. Asthmatics are more prone to severe viral infections due to their unique inflammatory response, which is mostly characterized by T2 cytokines. Unlike the normal T1 high response to viral infection, asthmatics with T2 high inflammation are less potent in containing a viral infection. Inhaled and/or systematic corticosteroids and bronchodilators remain the cornerstone of asthma exacerbation treatment, and although many targeted therapies which block molecules that viruses use to infect the host have been used in a laboratory level, none has been yet approved for clinical use. Nevertheless, further understanding of the unique pathway that each virus follows to infect an individual may be crucial in the development of targeted therapies for the commonest viral pathogens to effectively prevent asthma exacerbations. Finally, biologic therapies resulted in a complete change of scenery in the treatment of severe asthma, especially with a T2 high phenotype. All available data suggest that monoclonal antibodies are safe and able to drastically reduce the rate of viral asthma exacerbations.
Keywords: asthma, virus, exacerbation, pathogenesis, lungs
Introduction
Asthma is a common and heterogeneous disease that affects millions of people on a global basis. Although most cases are mild, there are patients who suffer from severe asthma and experience various exacerbations during their life. These exacerbations can usually be limited and controlled with medication, but some can even prove lethal. Therefore, the causes of asthma exacerbations need to be addressed and if possible, prevented.1 There is a great variety of contributing factors responsible for asthma exacerbations, characterized as episodes of coughing, chest tightness, wheezing and shortness of breath. Respiratory viral infections play an important role in the asthma immunopathogenesis and are associated to the induction of asthma exacerbations.2 Specifically, in 70% of adult patients suffering from an asthma exacerbation, viral or bacterial infections were observed,3 while according to multiple clinical studies, up to 45% of cases of adult-onset asthma occur after an acute infection of upper or lower respiratory tract.4
According to a prospective study including 105.519 people followed for up to 23 years, regardless of smoking history, those experiencing any infection recorded a higher risk of developing asthma. Early asthma patients with no smoking history, had a significantly higher risk of contracting any infection (hazard ratio [HR], 1.65; 95% confidence interval [CI], 1.40–1.94), pneumonia (HR, 2.44; 95% CI, 1.92–3.11), or any non-respiratory tract infection (HR, 1.36; 95% CI, 1.11–1.67); the results were similar in early asthma patients with smoking history. In addition, people who never smoked but had any history of asthma, also, had a significantly higher risk of infection (HR, 1.44; 95% CI, 1.24–1.66) and pneumonia (HR, 1.99; 95% CI, 1.62–2.44).5
Multiple pathogens, up to this point, have been associated with asthma onset and disease exacerbations. Precisely, Respiratory syncytial virus (RSV) and rhinovirus (RV) have been linked to asthma onset later in life,6 while RSV (2–20%), parainfluenza virus (PIV) (2–11%), RV (44–88%)7 and several other viruses, such as coronavirus (CoV), Influenza virus (IAV), enterovirus (EV), metapneumovirus (MPV), adenovirus (AdV), bocavirus (BoV) have been described to lead in asthma exacerbations and worsening of wheezing.8 Additionally, it is crucial to note that 10% of these cases have a coinfection with another virus, which is typically RV.2 In this review, we will try to summarize the epidemiology and immunopathogenesis of the commonest viruses associated with asthma exacerbations and discuss treatment options for viral asthma exacerbations.
Respiratory Syncytial Virus (RSV)
The most frequent cause of severe lower respiratory tract infection (LRTI) in newborns and young children is RSV, which normally infects people by the age of 2 and can lead to recurrent infections throughout one’s lifetime.9 RSV is a pneumovirus in the Paramyxoviridae family and is a single-stranded enveloped RNA virus with 2 major antigenic groups, A and B.10 It is transmitted via large droplets and secretions from infected patients. RSV is responsible for the majority of bronchiolitis cases especially in infants during the age of 3–6 months and almost all children have RSV infection by the age of 2.11 The peak prevalence is during midwinter. Studies support that both mild and severe RSV bronchiolitis may be followed by recurrent wheezing for several years.12–14
An annual epidemic of RSV-related infection peaks in January or February in the United States, and the age at which RSV infection carries the greatest risk of morbidity is 50 years old.15 Severe early RSV infection might cause allergic sensitization and asthma. Sigurs et al16 examined 13-year-old children (92 control individuals and 46 RSV-infected). In the RSV-infected group, they found 43% prevalence of asthma and recurrent wheeze and 39% prevalence of allergic rhinoconjunctivitis, respectively, compared to 8% and 15% in the control participants. In a different study by Schauer et al,17 it was demonstrated that 33% of RSV-infected infants tested positive for immunoglobulin E (IgE) antibodies as opposed to just 2.3% of the children in the control group. They came to the conclusion that severe RSV-induced bronchiolitis in the first year of life is a significant risk factor for wheezing and being sensitive to common allergens in the year after.18 Consequently, RSV infection is a significant risk factor for the future development of asthma, particularly in people who are genetically predisposed to allergic disease. Moreover, RSV infection is also a pivotal factor of asthma worsening and exacerbations, not only in childhood but also in adult asthma.18
Murine studies indicate that prior sensitization to individual RSV surface proteins followed three weeks later by RSV infection can induce polarized cytokine responses that follow broad type 1 and type 2 repertoires.19,20 Normally, Th1 responses with IFN-γ and IL-2 production are seen with viral disease. The pulmonary histologic findings in mice previously immunized with the inactivated RSV vaccine are more consistent with a T2 high response, an ineffective immune response to viral pathogens.21 Mice with T2 high response developed severe disease with eosinophilia, whereas those with Th1 responses had reduced immunopathology and enhanced viral clearance.22
Primarily, RSV targets nasal epithelial cells that release pro-inflammatory cytokines and recruit immune cells, such as monocytes, macrophages and dendritic cells (DCs).23 According to current literature RSV-caused asthma is closely related to the atopic constitution since it is characterized by a T2 dominant immune response.24 The suggested mechanisms are the following: RSV stimulates the T cell responses to inhalant allergens, by triggering the local T2 cytokine at the airway mucosa; promotes eosinophils recruitment at lesional sites in the airway mucosa; and generates a T2-polarized RSV-specific immunological memory, which, following a RSV reinfection, leads to intense infiltrates of eosinophils and Th2 cells secreting interleukin IL-4 in the lung tissue.25–29 Finally, RSV makes the airways abnormally susceptible to the RSV-caused proinflammatory effects by upregulating NK-1 receptor gene expression and, thereby, increasing the synthesis of the substance P and the density of its receptors on target immune cells, including lymphocytes, macrophages, mast cells, and endothelial cells.24
Recently, two RSV vaccines were approved by FDA for adults over 60 years old, marking a new era, in which immunization against RSV could significantly reduce the susceptibility of patients to lower respiratory tract infection due to RSV. A Phase 3 trial to achieve this was conducted by Papi et al,30 where researchers targeted the RSV F glycoprotein, whose role is crucial for viral fusion and infiltration of host-cells. The vaccine contains a form of this protein in its prefusion state, and immunization is achieved by exposing epitopes, which are then targeted by neutralizing antibodies. 25,000 adults over 60 years old were recruited and randomized in a 1:1 scale to either the vaccine or placebo with a median follow-up period of 6.7 months. Only 7 adults in the vaccine arm were infected by RSV compared to 47 in the placebo group. Additionally, only 1 patient in the vaccination group suffered from a severe RSV infection in contrast to the placebo group, with 17 patients suffering from a severe infection. These results yield a vaccine efficacy of 82.6% and an even greater efficacy of 94.1% against severe infection.30 Thus, vaccination has now been approved by FDA for all adults over 60 years old as a single dose regimen.
The second vaccine approved for RSV immunization in adults over 60 years old, achieves immunization with a similar logic. It contains a bivalent F glycoprotein in a stabilized prefusion conformation for RSV subgroups A and B. The phase 3 trial interim analysis included a total of almost 35,000 patients, also randomized in a 1:1 ratio to either vaccine or placebo. Only 11 vaccinated people were infected by RSV in comparison to 33 in the placebo group, yielding a vaccine efficacy of 66.7%. As for the secondary outcome of severe RSV LRTI, 2 vaccinated and 14 patients who received placebo suffered from it, which in turn yields an efficacy against severe RSV infection of 85.7%.31
Additionally, the same vaccine was used in another phase 3 clinical trial, which recruited pregnant women at 24 through 36 weeks of gestation and vaccinated them in order to prevent RSV infection in the newborns. At the interim analysis of over 7000 participants the vaccine exerted an efficacy of 81.8% for the first 90 days after birth, which slightly dropped to 69.4% in the 180 days follow-up period. The FDA approved the bivalent RSV vaccine for pregnant women at 32 through 36 weeks of gestation.32 Last but not least, it should be underlined that all vaccines currently approved have an excellent safety profile with no issues raised whatsoever.30–32
Rhinovirus
Rhinovirus is the most prevalent member of the Picornaviridae family of RNA viruses, known as Enterovirus (EV), and has been linked with both the onset and worsening of asthma. There are three major clades that RV can be divided, RV-A, RV-B, and RV-C. RV-C has been found to have the strongest correlation with the severity of asthma exacerbations out of these 3 variables.2 With regard to 60% of all virus-induced exacerbations, RV is the leading viral pathogen.33
In addition to RV, additional EV have been found in people who are experiencing an asthma attack, and these viruses may be responsible for the development or worsening of asthma. Numerous studies have noted a connection between an increased frequency of EV infections and the emergence of asthma.34,35 One study analyzed patients retrospectively over a ten-year period (January 2000 to December 2011) and studied the connection between EV infection and asthma. Patients with a documented EV infection had an incidence of asthma that was 1.48 times higher than those without.36 Quantitative studies, however, found no differences between participants with mild asthma and healthy individuals regarding the frequency of lower airway infections or viral loads in sputum.37,38
RVs can infect ciliated cells in the nasal epithelium but may also infect other nasal cells. Intercellular adhesion molecule-1 (ICAM-1) is the receptor that most rhinovirus serotypes use to enter into human cells.39,40 In asthmatic patients, ICAM-1 is upregulated in the lower airway epithelium and may justify the increased susceptibility of those individuals to RV infection.40,41 Although, RVs usually affect the upper respiratory epithelium, their isolation from bronchoscopic samples indicates that they can also infect the lower respiratory tract.42,43
In vitro infection of cell lines and primary bronchial epithelial cells (BECs) shows that RV infection leads to the release of the pro- inflammatory mediators IL-6, IL-8, TNF-a, and IL-1b as well as RANTES and granulocyte-macrophage colony stimulating factor (GM-CSF).44,45 Spurrell et al46 in their study also suggest that increased levels of RANTES and IP-10 have been shown to correlate with in vivo virus replication. RANTES serves as a chemoattractant for eosinophils,47 and GM-CSF is a potent activator of eosinophil survival and adhesion molecule expression, and is a co-factor for eosinophil superoxide production and de-granulation.16,48–50 It has been shown that the combination of RV infection and direct exposure to allergens cause epithelial cell production of IL-25 and IL-33 in the airways, mediators involved in T2 type inflammation and remodeling.51 After transmission, RV adheres to the ICAM-1 receptor and enters the BECs which is followed by the formation of viral RNA and virions. These events elicit activation of nuclear factor kappa B (NFkB) which, in turn, leads to the production of pro-inflammatory cytokines. In non-asthmatic patients, these cytokines induce apoptosis and lysis of the infected cells limiting virus transmission to healthy cells. In contrast, in asthmatic patients, there is no early activation of apoptosis and as a result, neighboring cells become infected, enhancing the release of pro-inflammatory cytokines responsible for airway inflammation.11,52
Parainfluenza Virus
Human parainfluenza viruses (PIVs) are a significant contributing factor of respiratory illness in both children and adults, causing a great variety of clinical symptoms, such as colds, croup, bronchiolitis, and pneumonia. Seasonal PIV epidemics cause a large burden of disease in children, accounting for 40% of lower respiratory tract infections (LRTI) leading to hospitalization in children and 75% of croup cases.53 The Paramyxoviridae family of single-stranded, enclosed RNA viruses includes parainfluenza viruses. Serotypes 1 through 4 refer to the four main serotypes of PIV, while PIV4 is further separated into two genera (HPIV4a and HPIV4b).54 Adult illnesses are often not severe, though asthmatics may have airway hyperresponsiveness from the production of cytokines and chemokines. More specifically, in a study evaluating 19 adult asthmatics suffering from PIV 3-induced asthma exacerbation, it was reported that PIV3 infection was linked to a substantial rise in sputum cysteinyl leukotrienes (cysLTs).55
Influenza Virus
Influenza virus presenting both as a seasonal infection and sporadic pandemics is one of the most often found viruses during asthma exacerbations, causing an average of 3.1 million annual inpatient days and 31.4 million outpatient visits.2,56,57 The prevalence of influenza viruses can reach up to 20% in infants presenting wheezing and 20% to 25% in adults experiencing acute asthma exacerbations during flu seasons, while the detection rate of influenza virus in patients with asthma exacerbations is only approximately 10% worldwide.58–60
Asthma was the most prevalent comorbidity among patients during the 2009 influenza pandemic (H1N1), accounting for 22% to 29% of all hospitalized patients with influenza.61 Most differences between the pandemic influenza and seasonal influenza were that pH1N1 were linked to a higher incidence of pneumonia (46% vs 40%) and a greater need for intensive care admission (22% vs 16%). Children with asthma accounted for 44% of hospitalized children with influenza.62 It has yet to be elucidated whereas people with asthma contract the influenza virus more frequently compared to the general population. Asthmatics have been demonstrated to have decreased type 1 interferon (IFN) responses to RV infection.57,63 Further to this, children with asthma were twice as likely to contract H1N1 influenza during the 2009–2010 season, according to a US study that tracked participants’ symptoms and weekly nose samples.64 In any case, it is widely acknowledged that the presence of asthma increases the risk of developing a severe disease due to influenza virus; hence, people with asthma have been designated as a population that needs immunization on a priority basis.65
Metapneumovirus
The single-stranded RNA virus known as human MPV, which is a member of the Pneumoviridae subfamily, exhibits many of the same signs and symptoms as RSV. It normally peaks between December and April, in the late winter and early spring and wheezing, pneumonia, and mild, self-limiting acute upper respiratory tract infections are among the commonest symptoms of hMPV infection.57,66 MPV’s contribution to asthma attacks varies depending on the demographic characteristics of each population. Approximately 5% of asthmatics experiencing an exacerbation have MPV detected globally, compared to a substantially greater percentage of people with MPV LRTI who have been diagnosed with asthma.60,66 Children with MPV LRTI were included in a US study in which 14% to 33% had an asthma diagnosis or a history of wheezing.67
Covid-19 (SARS-CoV-2)
During these last years of COVID 19 pandemic, caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), there was major concern about patients with asthma and the possibility of a greater susceptibility to SARS-CoV-2 infection and disease severity. It is now known that Angiotensin-converting enzyme (ACE-2) receptor is the major receptor responsible for SARS-CoV-2 entry into human cells. As a result, when ACE2 receptor is over expressed, especially in nasal epithelial cells and type II alveolar epithelial cells, the patients’ lungs are possibly exposed to a greater risk of contracting SARS-CoV-2, increasing at the same time disease severity.68–70
According to early epidemiological research, asthmatics had a relatively low incidence of COVID-19 and did not appear to be at a higher risk of contracting SARS-CoV-2. However, there was a wide range in the reported COVID-19 prevalence among asthmatics around the world. Early studies in China have not reported an increased prevalence of asthma among patients with Covid-19 infection71–74 although data from European areas revealed great discrepancy among different countries.75
In respect to disease severity, asthma is not considered as an independent risk factor for worse outcomes, like other comorbidities (obesity, cardiovascular disease and diabetes)76 and in the systemic meta-analysis of Liu et al77 asthma was not linked with COVID-19 severity and mortality. Finally, another large study in Korea which included over 7.000 patients, initially documented a statistically significant difference in mortality rate for asthmatics with COVID-19, but after adjustment with other comorbidities, asthma was not found to be a standalone risk factor for worse Covid-19 outcomes.78
An interesting hypothesis even claims that asthma might act protectively against SARS-COV-2 infection. As already mentioned, the virus uses the ACE-2 receptor to enter the airway epithelial cells. In asthmatic patients under maintenance treatment with inhaled corticosteroids, there is a down-regulation of the ACE-2 receptors, which implies a possible protective effect of the cornerstone asthma treatment against SARS-COV-2.79 The role of ICS in COVID infection has been debated during the course of the pandemic. In a systematic review and meta-analysis recently published, ICS either as monotherapy or combined with inhaled bronchodilators did not increase the risk of infection from SARS-COV-2.80 Although initial results for inhaled ciclesonide hinted that it could shorten the viral shedding duration in patients infected from COVID,81 so far its use is not encouraged in the general population. Two open-label Phase 2 trials with budesonide yielded some positive results, however its use was not proposed in the guidelines by the National Institute for Health and Care Excellence (NICE).82,83 Nevertheless, asthmatics under maintenance treatment with ICS should continue using them as prescribed, since stopping ICS greatly increases the risk of asthma exacerbations.84
Additionally, it has been also hypothesized that the repeated airway injury occurring in asthmatic patients due to their exposure to allergens, viruses and other environmental triggers might in turn be the reason that their inflammatory response to infection from SARS-COV-2 was less severe compared to healthy subjects who do not express this “immune tolerance”.85 Finally, it should be noted that most asthmatics suffer from mucus hypersecretion, rich in glycoproteins and substances, such as MUC5AC covering most of the lower respiratory tract. Thus, mucus could possibly act as a shield and prevent the virus from infecting the airways, as it was shown in vitro in mice with an overexpression of MUC5AC, in which the viral load of the influenza virus was much lower compared to their counterparts with normal mucus expression.86
Immunopathogenesis of Viral Infections in Asthma
As it has already been mentioned, respiratory viruses are the most frequent causative agents of disease in humans, with significant impact on morbidity and mortality worldwide.87 Viral infection of the respiratory tract can result in a variety of specific syndromes such as the common cold, pharyngitis, tracheobronchitis, croup, bronchiolitis or pneumonia. Such syndromes may be superimposed on a background of pre-existing chronic respiratory diseases, such as asthma.88 The most common viral asthma triggers and their basic features are summarized in Table 1.
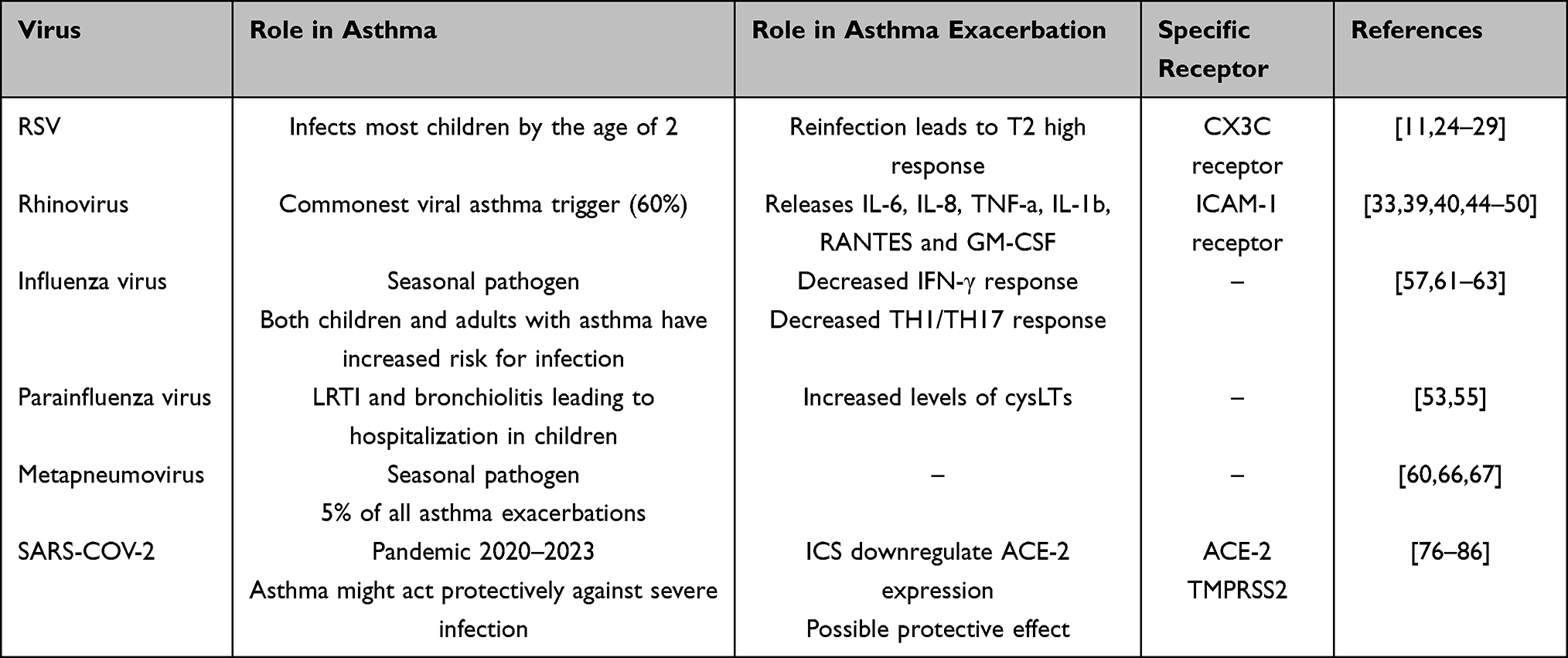 |
Table 1 The Commonest Asthma Viral Triggers, Their Role in the Development and Exacerbation of Asthma and the Molecular Pathway via Which They Trigger an Asthma Exacerbation |
Asthma is characterized by chronic airway inflammation, mucus hypersecretion, smooth muscle contraction and infiltration of inflammatory cells like T-lymphocytes and eosinophils in the airway mucosa, which all lead to variable airflow obstruction. Some of the risk factors associated with asthma development or its exacerbations are exposure to environmental allergens, tobacco smoke, familial predisposition, atopic characteristics (ie, increased blood eosinophilic count), gastroesophageal reflux disease, obesity and viral respiratory tract infections. Nevertheless, the exact etiology of asthma and its exacerbations is complex and not fully clarified.89
The association between asthma development and/or exacerbations and viral respiratory tract infections has been suggested, but a causal relation has never been documented. Although, the association of viral infections and exacerbations of asthma is clearly defined, the role of viral infections in the etiology of asthma itself is more controversial.90 Respiratory infections by IAV, RSV, RV, PIV and MPV have all been implicated in the development of asthma as well as exacerbations.91 Several studies suggest that repeated respiratory viral infections during early age are associated with an increased risk of developing asthma in later childhood. Illi et al92 studied the association between different types of early childhood infections and the subsequent development of asthma. In this birth control study, 1314 children were followed from birth to the age of 7 years. At each follow-up parents filled questionnaires mainly focused on asthmatic and atopic symptoms as well as the child’s diseases. The main result of the study was that repeated LRTIs were strongly associated with the diagnosis of asthma. Legg et al20 provided two possible explanations for the induction of asthma due to viral infections during childhood. The first one is that there is an excess type 2 and deficient type 1 T helper (Th1) cell response. The second one is that viral infections at early-stage result in airway inflammation and remodeling predisposing to wheezing.20 However, these remain only possible explanations and have not been proven to be the only mechanism of asthma development.
Susceptibility to Viral Infections
Viral infections influence the immune system in completely different ways at different time points in the asthma pathogenesis and as a result they can have a great influence on the course of asthma.89 The impact of viral infections on the respiratory tract system depends on the host’s immunity system. Most individuals infected by respiratory viruses remain asymptomatic or express mild symptoms and the infection is usually self-limited due to the host’s adequate immune response. However, if the immune system fails to provide the necessary protection, the airway function could be seriously damaged and develop clinical signs of disease. Factors that determine the clinical outcome of an exposure to a respiratory virus may depend on the virus itself (type, virulence, dose), the host (age, genetic susceptibility, immune status) and the environment (temperature, humidity, pollution).93 According to Folkerts et al,94 respiratory viruses can cause increased airway responsiveness in humans and in animals, especially if they achieve to colonize the lower airways. This rarely occurs because in immunocompetent individuals the infection is self-restricted. This contrasts with subjects with underlying diseases such as asthma, who may be more susceptible to viral infections due to abnormalities that occur in innate immune response. More specifically, antiviral responses have been characterized by a Th1 phenotypic response with raised levels of IFN-γ and recruitment of CD8 cells. On the other hand, Th2 response thought to be dominant in most asthmatics, with increased levels of IL-4, IL-5 and IL-13 and so these patients are less effective in clearing the viruses.11 To summarize, in asthmatic patients viruses may take advantage of a deficient antiviral response and spread to the lower airways, which are predisposed to develop exaggerated constrictive responses to nonspecific stimuli.41,93
An interesting theory has been developed for allergic asthma and viral infections. It is known that plasmacytoid dendritic cells (pDCs) execute a major role in the host defense against viral infections. In allergic asthma, IgE receptor is overexpressed in the surface of pDCs and has been shown that the higher the IgE plasma levels are, the higher the expression of its receptor is too leading to a decreased antiviral activity of these cells. The “cross-linking” theory suggests that reducing the circulating IgE can in turn reduce the expression of its receptor on pDCs and this can booster the anti-viral activity of pDCs and lead to the reduction of viral exacerbations of asthma.95
Although viruses are the most common asthma exacerbation triggers, bacterial superinfections can often deteriorate the clinical status of a patient. The most common bacteria implicated in asthma are atypical pathogens (most importantly Mycoplasma pneumoniae and Chlamydia pneumoniae), S. pneumoniae and H. influenzae.3,96 Bacterial co-infections may influence both exacerbation risk and severity. Moreover, asthmatic patients are in a higher risk of developing pneumococcal pneumonia than healthy individuals.97 Therefore, especially in an inpatient setting, clinicians should remain vigilant and treat any superimposed bacterial infections.
Severe Asthma Viral Exacerbations
Asthma is divided into mild, moderate and severe based on the treatment required to maintain asthma control. Mild asthmatics rely on low dose ICS and beta-2 agonists as needed, moderate asthmatics require higher doses of ICS and LABA, while severe asthmatics cannot achieve control even with high dose ICS and LABA treatment, thus requiring special treatment. Asthma exacerbations according to GINA are characterized as “episodes of progressive increase in shortness of breath, cough, wheezing, or chest tightness, or some combination of these symptoms, accompanied by decreases in expiratory airflow that can be quantified by measurement of lung function compared to patients’ baseline and are associated with increased morbidity and mortality”.98
The severity of the exacerbation can be assessed through serial measurements of forced expiratory volume during the 1st second (FEV1) and peak expiratory flow (PEF).51 PEF values below 50% of patient’s best or predicted value indicate acute severe asthma. Clinically, patients suffering from a severe asthma exacerbation cannot complete phrases and prefer to sit leaning forward. Moreover, they may be agitated with increased respiratory rate (>30 breaths/min), use accessory muscles and present with tachycardia (>110 beats/min) and low oxygen saturation (<90%) without the concomitant use of oxygen therapy.99
In established asthma, viral infections are a frequent cause of asthma exacerbation.89 Upper respiratory viral infections are responsible for 80–85% of asthma exacerbations during childhood.100,101 In adults, rates of virus detection have varied with studies, but they remain the single most prevalent trigger for acute asthma.11 In both populations, rhinovirus was the most frequently identified virus responsible for triggering of asthma.33 Several studies have demonstrated that the risk for hospital admission due to asthma exacerbation is higher in patients with the combination of sensitization, current exposure to high levels of sensitizing allergens and the presence of viral infection.102,103 The hyper-responsiveness of the lower airway may be explained by the deficient IFN-β response to viral infection.11
The complex virus–host interaction in asthmatics is characterized by impaired host immunity due to destruction of epithelium and tight junctions, augmented cell death and deficient Th1 response and IFN-γ production in combination with enhanced viral entry through increased expression of viral receptors and production of proinflammatory cytokines and interleukins.104,105 Several studies have demonstrated that RV-A, RV-B and RV-C subtypes adhere to the bronchial epithelium through receptors including intercellular adhesion protein 1 (ICAM-1), low-density lipoprotein receptors (LDLR) and cadherin-related family member 3 (CDHR3), while RSV attaches to CX3CR1.106–108
In acute severe asthma, patients may present with increased levels of neutrophils and eosinophils in their blood tests, and this mixed population of immune cells can be used as a biomarker for severe disease, poor response to corticosteroids and consequently worse prognosis.109 Furthermore, the presence of neutrophils in the airways is associated with increased bronchial hyperresponsiveness regardless of the number of eosinophils.110
Patients with a severe asthma attack present with serious gas exchange abnormalities. Hypoxemia occurs mainly due to V/Q mismatch and in advanced stages, hypercapnia may develop due to respiratory muscle fatigue and alveolar hypoventilation. During an acute exacerbation, severe bronchospasm leads to increased airway resistance, which is associated with reduced airflow and premature airway closure.111 Premature small airways’ closure results into dynamic hyperinflation, air trapping and increased respiratory load. If the patient continues to be unresponsive to treatment, accessory muscles are activated to perform the work of breathing. The respiratory load increases even more leading to more severe respiratory failure and even respiratory arrest.
Another important factor for further deterioration is the hemodynamic instability, which results from the elevated intrathoracic pressures. High intrathoracic pressures lead to a reduced preload and an increased afterload of the right ventricle.112 The hemodynamic instability in combination with the unresolving dynamic hyperinflation can make a patient drowsy with reduced or even absent respiratory sounds, which may be a sign of imminent respiratory arrest. Hence, identification and immediate treatment of a severe asthma exacerbation is vital as it may reduce morbidity and mortality.
Viral Asthma Exacerbations: Treatment Options
Currently, bronchodilation therapies and corticosteroids (administered both orally and inhaled) are the mainstay of treatment of acute asthma exacerbations and act by relaxing airway smooth muscle and reducing inflammation, respectively. Inhaled corticosteroids (ICS) exert their anti-inflammatory effect through the interaction of the glucocorticoid receptor (GR) with negative glucocorticoid response element (GRE) sites and through binding of pro-inflammatory transcription factors (eg, AP-1, NF-κB).113 Long-acting β2-adrenergic receptor agonists (LABA) have been shown to enhance the anti-proliferative and anti-inflammatory effects of ICS and also synergistically inhibit the production of immunopathogenic chemokines by RV-infected cells.114 According to Jackson et al,105 the combination of LABA (salmeterol and formoterol) with ICS (fluticasone and budesonide) can inhibit the production of several chemokines (CXCL8, CXCL10 and CCL5) involved in inflammation and remodeling-associated growth factors (FGF and VEGF) upon RV infection. As a result, the virus-induced inflammation and remodeling are halted and this may prevent further exacerbations.
Oral corticosteroids (OCS) offer a more systematic anti-inflammatory efficiency in the setting of an acute exacerbation. Not only they manage to reduce the risk of relapse by more than 50%, they also reduce the risk of hospitalization by 60% compared to beta-2-agonists.115 So far, short courses of oral corticosteroids (no more than 5–7 days) without a dose tapering are suggested in asthma exacerbations with a severe clinical presentation.116 However, even short-term treatment with oral corticosteroids can be harmful and their side-effects must not be neglected. Their use should be solely reserved for severe asthma exacerbations, while mild to moderate exacerbations should focus on steroid-sparing therapies.117
Other available treatments reversing bronchoconstriction and reducing inflammation are anticholinergics, such as long-acting muscarinic receptor antagonists (LAMA). They inhibit the action of acetylcholine, which causes secretion of mucus and bronchoconstriction. LAMA augment the bronchodilatory activity of short-acting β2-adrenergic receptor agonists (SABA) and they have been proved beneficial for patients with severe asthma exacerbations.118,119
Phosphodiesterase (PDE) inhibitors and in particular theophylline, up-regulate histone deacetylase 2-expression whose deficiency is implicated in the resistance of some patients to steroids.120 Moreover, theophylline has anti-inflammatory and brochodilatory effects and it can be added in a combination therapy of SABA and ICS in acute severe asthma. However, its application is limited due to many side effects and interactions with other common drugs and for this reason its use cannot be suggested.121,122 Roflumilast, another PDE-4 inhibitor has been tested in asthma and trials show that its use can be equivalent to 400mcg of beclomethasone dipropionate (medium strength ICS dose for asthma).123 Roflumilast exerts anti-inflammatory effects and also attenuates allergen-induced bronchoconstriction in asthmatics. Nevertheless, its use is limited in asthma and it is not indicated in the treatment of an acute disease exacerbation.124
Leukotriene receptor antagonists constitute another class of drugs targeting inflammatory pathways. Leukotrienes are inflammatory lipid mediators produced by the arachidonic acid pathway (AA) in activated immune cells and they are separated in two classes: cysteinyl leukotrienes and LTB4. Increased levels of CysLT cause airway smooth muscle contraction and increased vascular permeability through binding to the CysLT receptors (mainly CysLT1R) in the airways.125 Montelukast, which is a CysLT1R antagonist has shown promising results in ameliorating lung function in patients with moderate to severe viral asthma exacerbations.126–128
For many asthmatics, the aforementioned approaches may be adequate to control symptoms and reduce the impact of viral infections, however a subgroup of patients with more severe asthma requires more treatment options to reduce the burden of viral exacerbations and the overall morbidity and mortality.129 The mechanisms of viral entry, replication, and the production of cytokines in parallel with the deficient immune response of patients with asthma, constitute potential therapeutic targets.
Bronchial epithelial cells are infected by viruses. Using treatments to enhance anti-viral immunity to block or even prevent infection is challenging and more extensive research is needed in this area. Nevertheless, inflammatory cytokines that can activate type-2 innate lymphoid cells (ILC2) and Th2 cells, which produce type-2 cytokines, could be targeted by specific drugs.130 ILC2 and Th2 cells in the lung produce high levels of IL-5 and IL-13.130,131 Type-2 cytokines play a major role in mediating allergic airways inflammation in asthma. IL-4 and IL-13 are necessary for IgE production while IL-5 is responsible for recruitment and maturation of eosinophils. Increased numbers of eosinophils are associated with virus-induced asthma exacerbations and in particular, sputum eosinophilia correlates with asthma exacerbation severity.132 Increased secretion of mucus and airway hyperreactivity (AHR) is stimulated by IL-13. Type-2 cytokines also have an impact on anti-viral immunity. IL-4 and IL-13 can inhibit RV-16 induced interferon production and virus replication.133 Since RV is the commonest cause of asthma exacerbations, it is important to investigate whether by blocking IL-4 and IL-13, RV-induced exacerbations of asthma could be prevented.128
Several monoclonal antibodies targeting interleukins have been or are currently being investigated in clinical trials. Pascolizumab which is a humanized anti-IL-4 monoclonal antibody (mAb) had limited effect in treating asthma and was never approved134,135 while there is no evidence that blocking IL-4 had any protection against virus induced asthma exacerbations. Tralokinumab and lebrikizumab, two humanized anti-IL-13 mAbs although at the beginning showed promising results by improving lung function and reducing the need for β2 agonist administration and the overall incidence of asthma exacerbations they finally failed to provide efficacy in the treatment of asthma.136,137 Dupilumab which binds the common IL-4/IL-13 receptor has been approved for use in patients with severe asthma as it seems to improve asthma control and significantly reduce exacerbations by all causes.138 Especially in patients with comorbid chronic rhinosinusitis with nasal polyps, dupilumab has shown excellent results and it can be a first-line choice in the setting of severe nasal polyps disease and moderate to severe asthma, since it has shown similar results with anti-IL-5 biologics.139
Anti-IL-5 mAbs including mepolizumab, reslizumab and anti-IL-5 receptor alpha mAb benralizumab have been approved for patients with asthma and high eosinophil levels and T2 inflammation.140 All anti-IL-5 agents have shown excellent results in reducing all-cause asthma exacerbations, including viral exacerbations, both in randomized and real-world trials. A possible mechanism of action against viruses could be that IL-5-induced airway eosinophilia suppresses TLR-7 expression and antiviral IFN secretion, causing severe inflammation. IL-5 and eosinophil levels are known to increase in the airways of patients infected by rhinovirus and higher levels of either of them have been related with increased severity of viral infections.141
IgE-mediated activation of FcµR1 on mast cells and basophils causes the release of inflammatory cytokines and induces inflammation. Omalizumab is a monoclonal antibody that binds and neutralizes IgE thus preventing activation of FcµR1142 and consequently, reduces inflammation. It effectively decreases asthma exacerbations and improves symptom control, while it allows the reduction of high ICS doses in severe asthmatics.143 Two studies have investigated whether omalizumab could be used as prophylaxis and reported significant exacerbation reductions some which were caused by viruses.144,145 Patients under anti-IgE treatment do not express a seasonal exacerbation pattern, like most asthmatics do, with increased exacerbations during the winter and spring and much fewer exacerbations in the summer. In a post hoc analysis of the ICATA study145 evaluating the use of omalizumab in children suffering from severe asthma, it was shown that while patients in the placebo group experienced a seasonal surge in asthma exacerbations (9% in the winter and 8.1% during spring), those randomized in the treatment arm maintained steady and low exacerbation rates in all seasons (4.3% during the winter and 4.2% in spring).145 This comes in accord with the theory of anti-IgE treatment enhancing anti-viral immunity.
Moreover, Garcia et al146 have demonstrated that the use of omalizumab can reduce the levels of the IgE receptor on pDCs and strengthen the anti-viral response of these cells. The reduction of both circulating IgE and of its receptor on pDCs leads to the reduction of the rate of viral asthma exacerbations through an innate immunity path rather than the well-known allergic path omalizumab treatment mainly targets.95
Type-2-promoting cytokines such as IL-25 and IL-33 (which are part of the alarmins and stimulate ILC2 cells) are associated with viral replication in bronchial epithelium, type-2 responses and inflammation in asthma exacerbations.147 IL-25 is an IL-17 family member and is a main regulator in T2 high responses and in type-2 cytokine production.148 A study showed that by blocking IL-25 receptor with anti-IL-17RB mAb in a mouse model with RV-induced asthma exacerbation, it prevented virus induced allergic inflammation and type-2 cytokine production.149 IL-33 expression is also linked with increased type-2 cytokine production and asthma exacerbation severity. Blocking IL-33 in vitro with an anti-ST2 mAb, resulted in inhibition of type-2 cytokines production by T cells from RV-infected asthmatic BECs.150 Some positive results on the anti-IL-33 biologic itepekimab were derived from a Phase II trial recently published, where it was compared to treatment with placebo, dupilumab and a combination of itepekimab and dupilumab was also used. Although it failed to surpass the results of dupilumab, itepekimab showed a better efficacy than placebo treatment and it could possibly be a future choice for patients with lower baseline blood eosinophils than the other biologics currently approved for severe asthma.151 Targeting alarmins can be a future scope of research, however so far, the only alarmin with a positive phase 3 trial is thymic stromal lymphopoietin (TSLP), with the biologic tezepelumab.152,153 Blockage of TSLP production from the epithelial cells reduces production of T2 high cytokines, such as IL-5 and IL-13 and since viruses can trigger TSLP production, treatment with an anti-TSLP agent could decrease viral asthma exacerbations through an innate immunity pathway.154
As mentioned above, inhibiting different steps in the viral replication procedure and enhancing anti-viral immunity are other potential therapeutic targets. Toll-like receptors (TLRs) are responsible for the activation of innate immune cells and the expression of anti-viral interferons in response to infections. Specifically, TLR7 and TLR9 have been implicated in asthma treatment. In IL-5 induced airways eosinophilia, TLR7 expression is reduced and it is inversely correlated with the level of eosinophils in the sputum.155 TLR7 can promote anti-viral immunity and cause bronchodilation in certain doses. Thus, TLR7 agonists are already tested in clinical trials and could constitute potential treatment options for virus-induced asthma exacerbations. Regarding TLR9, it is expressed in the airway epithelium and detects bacterial and viral CpG-DNA.156 In a previous study, TLR9 agonist treatments during an allergen challenge, decreased type-2 cytokine production, airway hyperresponsiveness and lung eosinophilia.157 However, more studies are required to examine the effects of TLR9 agonists in asthma.
Anti-viral type I interferons have immunomodulatory and anti-proliferative properties. Type I IFNs signal through JAK-1 and protein-tyrosine kinase (Tyk)-2, which phosphorylates STAT-1 and STAT2 to form a heterodimer before translocating into the nucleus and binding to promoters of interferon stimulated genes (ISGs).158 Many ISGs are involved in inhibiting viral replication.159 IFNs can be of vital importance in viral asthma exacerbations as they can improve control of viral infection and suppress type-2 driven allergic airways disease.
ICAM-1 is responsible for the recruitment and activation of cells expressing its natural ligands. It is expressed on epithelial cells, white blood cells and endothelial cells and is natural receptor for 90% of RV-species.40 Targeting ICAM-1 could be a potential treatment for viral induced asthma. Administration of anti-human ICAM-1 successfully prevented HRV16-induced exacerbation of allergic airway inflammation and airway hyper-responsiveness in mice.160 However, since data are limited, randomized controlled clinical trials are required to be carried out on humans to check its efficacy.
To summarize, the use of all biologics currently approved for severe T2 high asthma has led to a great reduction of all-cause exacerbations, including virus induced. Each category of biologics exerts its own anti-viral effects and it would be fascinating to further explore the anti-viral properties off all monoclonal antibodies approved for severe asthma treatment.161
Conclusions
Viral infections are the commonest triggers of asthma exacerbations in both children and adults, with the main causative agents being RSV and RV, respectively. Especially in predisposed children, viral infections might even be the trigger for development of asthma. In adults, viral infections can lead to severe exacerbations, which in some cases are life threatening. Therefore, understanding the mechanisms behind each type of viral infection in asthmatic patients and the differences in the type of inflammatory response compared to non-asthmatic individuals is crucial. So far, all data suggest that there is no special treatment targeting specific viruses in a molecular level, although the inhibition of several pathways has shown promising results in laboratory experiments in animal models. The mainstay of asthma treatment remains inhaled bronchodilators and corticosteroids. All biological therapies currently approved for asthma mainly target T2 inflammation and manage to reduce the rate of viral asthma exacerbations by restricting the otherwise uncontrolled T2 response to stimuli, such as allergens, pollutants and viruses. Further research on specific viruses could possibly yield new therapeutic options targeting viruses on a molecular level and add more treatment options for asthmatic patients in the event of a virus-associated asthma exacerbation.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Busse WW, Lemanske RF, Gern JE. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet. 2010;376(9743):826–834. doi:10.1016/S0140-6736(10)61380-3
2. Resiliac J, Grayson MH. Epidemiology of infections and development of asthma. Immunol Allergy Clin North Am. 2019;39(3):297–307. doi:10.1016/j.iac.2019.03.001
3. Iikura M, Hojo M, Koketsu R, et al. The importance of bacterial and viral infections associated with adult asthma exacerbations in clinical practice. PLoS One. 2015;10(4):e0123584. doi:10.1371/journal.pone.0123584
4. Hahn DL. Infectious asthma: a reemerging clinical entity? J Fam Pract. 1995;41(2):153–157.
5. Helby J, Nordestgaard BG, Benfield T, Bojesen SE. Asthma, other atopic conditions and risk of infections in 105 519 general population never and ever smokers. J Intern Med. 2017;282(3):254–267. doi:10.1111/joim.12635
6. Rossi GA, Colin AA. Infantile respiratory syncytial virus and human rhinovirus infections: respective role in inception and persistence of wheezing. Eur Respir J. 2015;45(3):774–789. doi:10.1183/09031936.00062714
7. Jackson DJ, Gangnon RE, Evans MD, et al. Wheezing rhinovirus illnesses in early life predict asthma development in high-risk children. Am J Respir Crit Care Med. 2008;178(7):667–672. doi:10.1164/rccm.200802-309OC
8. Tregoning JS, Schwarze J. Respiratory viral infections in infants: causes, clinical symptoms, virology, and immunology. Clin Microbiol Rev. 2010;23(1):74–98. doi:10.1128/CMR.00032-09
9. Mohapatra SS, Boyapalle S. Epidemiologic, experimental, and clinical links between respiratory syncytial virus infection and asthma. Clin Microbiol Rev. 2008;21(3):495–504. doi:10.1128/CMR.00054-07
10. Jartti T, Gern JE. Role of viral infections in the development and exacerbation of asthma in children. J Allergy Clin Immunol. 2017;140(4):895–906. doi:10.1016/j.jaci.2017.08.003
11. Wark PA, Gibson PG. Asthma exacerbations 3: pathogenesis. Thorax. 2006;61(10):909–915. doi:10.1136/thx.2005.045187
12. Sigurs N, Bjarnason R, Sigurbergsson F, Kjellman B. Respiratory syncytial virus bronchiolitis in infancy is an important risk factor for asthma and allergy at age 7. Am J Respir Crit Care Med. 2000;161(5):1501–1507. doi:10.1164/ajrccm.161.5.9906076
13. Pullan CR, Hey EN. Wheezing, asthma, and pulmonary dysfunction 10 years after infection with respiratory syncytial virus in infancy. Br Med J. 1982;284(6330):1665–1669. doi:10.1136/bmj.284.6330.1665
14. Stein RT, Sherrill D, Morgan WJ, et al. Respiratory syncytial virus in early life and risk of wheeze and allergy by age 13 years. Lancet. 1999;354(9178):541–545. doi:10.1016/S0140-6736(98)10321-5
15. Parrott RH, Kim HW, Arrobio JO, et al. Epidemiology of respiratory syncytial virus infection in Washington, D.C. II. Infection and disease with respect to age, immunologic status, race and sex. Am J Epidemiol. 1973;98(4):289–300. doi:10.1093/oxfordjournals.aje.a121558
16. Sigurs N, Gustafsson PM, Bjarnason R, et al. Severe respiratory syncytial virus bronchiolitis in infancy and asthma and allergy at age 13. Am J Respir Crit Care Med. 2005;171(2):137–141. doi:10.1164/rccm.200406-730OC
17. Schauer U, Hoffjan S, Bittscheidt J, et al. RSV bronchiolitis and risk of wheeze and allergic sensitisation in the first year of life. Eur Respir J. 2002;20(5):1277–1283. doi:10.1183/09031936.02.00019902
18. Falsey AR, Hennessey PA, Formica MA, Cox C, Walsh EE. Respiratory syncytial virus infection in elderly and high-risk adults. N Engl J Med. 2005;352(17):1749–1759. doi:10.1056/NEJMoa043951
19. Alwan WH, Openshaw PJ. Distinct patterns of T- and B-cell immunity to respiratory syncytial virus induced by individual viral proteins. Vaccine. 1993;11(4):431–437. doi:10.1016/0264-410X(93)90284-5
20. Legg JP, Hussain IR, Warner JA, Johnston SL, Warner JO. Type 1 and type 2 cytokine imbalance in acute respiratory syncytial virus bronchiolitis. Am J Respir Crit Care Med. 2003;168(6):633–639. doi:10.1164/rccm.200210-1148OC
21. Domachowske JB, Rosenberg HF. Respiratory syncytial virus infection: immune response, immunopathogenesis, and treatment. Clin Microbiol Rev. 1999;12(2):298–309. doi:10.1128/CMR.12.2.298
22. Alwan WH, Kozlowska WJ, Openshaw PJ. Distinct types of lung disease caused by functional subsets of antiviral T cells. J Exp Med. 1994;179(1):81–89. doi:10.1084/jem.179.1.81
23. Russell CD, Unger SA, Walton M, Schwarze J. The human immune response to respiratory syncytial virus infection. Clin Microbiol Rev. 2017;30(2):481–502. doi:10.1128/CMR.00090-16
24. Manti S, Piedimonte G. An overview on the RSV-mediated mechanisms in the onset of non-allergic asthma. Front Pediatr. 2022;10:998296.
25. Culley FJ, Pollott J, Openshaw PJ. Age at first viral infection determines the pattern of T cell-mediated disease during reinfection in adulthood. J Exp Med. 2002;196(10):1381–1386. doi:10.1084/jem.20020943
26. Varga SM, Wang X, Welsh RM, Braciale TJ. Immunopathology in RSV infection is mediated by a discrete oligoclonal subset of antigen-specific CD4(+) T cells. Immunity. 2001;15(4):637–646. doi:10.1016/S1074-7613(01)00209-6
27. Openshaw P, Murphy EE, Hosken NA, et al. Heterogeneity of intracellular cytokine synthesis at the single-cell level in polarized T helper 1 and T helper 2 populations. J Exp Med. 1995;182(5):1357–1367. doi:10.1084/jem.182.5.1357
28. Sigurs N, Aljassim F, Kjellman B, et al. Asthma and allergy patterns over 18 years after severe RSV bronchiolitis in the first year of life. Thorax. 2010;65(12):1045–1052. doi:10.1136/thx.2009.121582
29. Piedimonte G. RSV infections: state of the art. Cleve Clin J Med. 2015;82(11 Suppl 1):S13–S18. doi:10.3949/ccjm.82.s1.03
30. Papi A, Ison MG, Langley JM, et al. Respiratory syncytial virus prefusion F protein vaccine in older adults. N Engl J Med. 2023;388(7):595–608. doi:10.1056/NEJMoa2209604
31. Walsh EE, Perez Marc G, Zareba AM, et al. Efficacy and safety of a bivalent RSV prefusion F vaccine in older adults. N Engl J Med. 2023;388(16):1465–1477. doi:10.1056/NEJMoa2213836
32. Kampmann B, Madhi SA, Munjal I, et al. Bivalent prefusion F vaccine in pregnancy to prevent RSV illness in infants. N Engl J Med. 2023;388(16):1451–1464. doi:10.1056/NEJMoa2216480
33. Kistler A, Avila PC, Rouskin S, et al. Pan-viral screening of respiratory tract infections in adults with and without asthma reveals unexpected human coronavirus and human rhinovirus diversity. J Infect Dis. 2007;196(6):817–825. doi:10.1086/520816
34. Moss RB. Enterovirus 68 infection--association with asthma. J Allergy Clin Immunol Pract. 2016;4(2):226–228. doi:10.1016/j.jaip.2015.12.013
35. Foster CB, Coelho R, Brown PM, et al. A comparison of hospitalized children with enterovirus D68 to those with rhinovirus. Pediatr Pulmonol. 2017;52(6):827–832. doi:10.1002/ppul.23661
36. Yeh JJ, Lin CL, Hsu WH. Effect of enterovirus infections on asthma in young children: a national cohort study. Eur J Clin Invest. 2017;47(12):e12844. doi:10.1111/eci.12844
37. DeMore JP, Weisshaar EH, Vrtis RF, et al. Similar colds in subjects with allergic asthma and nonatopic subjects after inoculation with rhinovirus-16. J Allergy Clin Immunol. 2009;124(2):245–252, 252 e241–243. doi:10.1016/j.jaci.2009.05.030
38. Mosser AG, Vrtis R, Burchell L, et al. Quantitative and qualitative analysis of rhinovirus infection in bronchial tissues. Am J Respir Crit Care Med. 2005;171(6):645–651. doi:10.1164/rccm.200407-970OC
39. Jacobs SE, Lamson DM, St George K, Walsh TJ. Human rhinoviruses. Clin Microbiol Rev. 2013;26(1):135–162. doi:10.1128/CMR.00077-12
40. Greve JM, Davis G, Meyer AM, et al. The major human rhinovirus receptor is ICAM-1. Cell. 1989;56(5):839–847. doi:10.1016/0092-8674(89)90688-0
41. Wark PA, Johnston SL, Bucchieri F, et al. Asthmatic bronchial epithelial cells have a deficient innate immune response to infection with rhinovirus. J Exp Med. 2005;201(6):937–947. doi:10.1084/jem.20041901
42. Papadopoulos NG, Bates PJ, Bardin PG, et al. Rhinoviruses infect the lower airways. J Infect Dis. 2000;181(6):1875–1884. doi:10.1086/315513
43. Gern JE, Galagan DM, Jarjour NN, Dick EC, Busse WW. Detection of rhinovirus RNA in lower airway cells during experimentally induced infection. Am J Respir Crit Care Med. 1997;155(3):1159–1161. doi:10.1164/ajrccm.155.3.9117003
44. Terajima M, Yamaya M, Sekizawa K, et al. Rhinovirus infection of primary cultures of human tracheal epithelium: role of ICAM-1 and IL-1beta. Am J Physiol. 1997;273(4):L749–L759. doi:10.1152/ajplung.1997.273.4.L749
45. Mosser AG, Brockman-Schneider R, Amineva S, et al. Similar frequency of rhinovirus-infectible cells in upper and lower airway epithelium. J Infect Dis. 2002;185(6):734–743. doi:10.1086/339339
46. Spurrell JC, Wiehler S, Zaheer RS, Sanders SP, Proud D. Human airway epithelial cells produce IP-10 (CXCL10) in vitro and in vivo upon rhinovirus infection. Am J Physiol Lung Cell Mol Physiol. 2005;289(1):L85–L95. doi:10.1152/ajplung.00397.2004
47. Kameyoshi Y, Dorschner A, Mallet AI, Christophers E, Schroder JM. Cytokine RANTES released by thrombin-stimulated platelets is a potent attractant for human eosinophils. J Exp Med. 1992;176(2):587–592. doi:10.1084/jem.176.2.587
48. Sedgwick JB, Quan SF, Calhoun WJ, Busse WW. Effect of interleukin-5 and granulocyte-macrophage colony stimulating factor on in vitro eosinophil function: comparison with airway eosinophils. J Allergy Clin Immunol. 1995;96(3):375–385. doi:10.1016/S0091-6749(95)70057-9
49. Lopez AF, Williamson DJ, Gamble JR, et al. Recombinant human granulocyte-macrophage colony-stimulating factor stimulates in vitro mature human neutrophil and eosinophil function, surface receptor expression, and survival. J Clin Invest. 1986;78(5):1220–1228. doi:10.1172/JCI112705
50. Nagata M, Sedgwick JB, Bates ME, Kita H, Busse WW. Eosinophil adhesion to vascular cell adhesion molecule-1 activates superoxide anion generation. J Immunol. 1995;155(4):2194–2202. doi:10.4049/jimmunol.155.4.2194
51. Kostakou E, Kaniaris E, Filiou E, et al. Acute severe asthma in adolescent and adult patients: current perspectives on assessment and management. J Clin Med. 2019;8(9):1283. doi:10.3390/jcm8091283
52. Corne JM, Marshall C, Smith S, et al. Frequency, severity, and duration of rhinovirus infections in asthmatic and non-asthmatic individuals: a longitudinal cohort study. Lancet. 2002;359(9309):831–834. doi:10.1016/S0140-6736(02)07953-9
53. Branche AR, Falsey AR. Parainfluenza virus infection. Semin Respir Crit Care Med. 2016;37(4):538–554. doi:10.1055/s-0036-1584798
54. Henrickson KJ. Parainfluenza viruses. Clin Microbiol Rev. 2003;16(2):242–264. doi:10.1128/CMR.16.2.242-264.2003
55. Matsuse H, Kondo Y, Saeki S, et al. Naturally occurring parainfluenza virus 3 infection in adults induces mild exacerbation of asthma associated with increased sputum concentrations of cysteinyl leukotrienes. Int Arch Allergy Immunol. 2005;138(3):267–272. doi:10.1159/000088728
56. Molinari NA, Ortega-Sanchez IR, Messonnier ML, et al. The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine. 2007;25(27):5086–5096. doi:10.1016/j.vaccine.2007.03.046
57. Coverstone AM, Wang L, Sumino K. Beyond respiratory syncytial virus and rhinovirus in the pathogenesis and exacerbation of asthma: the role of metapneumovirus, bocavirus and influenza virus. Immunol Allergy Clin North Am. 2019;39(3):391–401. doi:10.1016/j.iac.2019.03.007
58. Teichtahl H, Buckmaster N, Pertnikovs E. The incidence of respiratory tract infection in adults requiring hospitalization for asthma. Chest. 1997;112(3):591–596. doi:10.1378/chest.112.3.591
59. Heymann PW, Carper HT, Murphy DD, et al. Viral infections in relation to age, atopy, and season of admission among children hospitalized for wheezing. J Allergy Clin Immunol. 2004;114(2):239–247. doi:10.1016/j.jaci.2004.04.006
60. Zheng XY, Xu YJ, Guan WJ, Lin LF. Regional, age and respiratory-secretion-specific prevalence of respiratory viruses associated with asthma exacerbation: a literature review. Arch Virol. 2018;163(4):845–853. doi:10.1007/s00705-017-3700-y
61. Jain S, Kamimoto L, Bramley AM, et al. Hospitalized patients with 2009 H1N1 influenza in the United States, April-June 2009. N Engl J Med. 2009;361(20):1935–1944. doi:10.1056/NEJMoa0906695
62. Dawood FS, Kamimoto L, D’Mello TA, et al. Children with asthma hospitalized with seasonal or pandemic influenza, 2003–-2009. Pediatrics. 2011;128(1):e27–e32. doi:10.1542/peds.2010-3343
63. Contoli M, Message SD, Laza-Stanca V, et al. Role of deficient type III interferon-lambda production in asthma exacerbations. Nat Med. 2006;12(9):1023–1026. doi:10.1038/nm1462
64. Kloepfer KM, Olenec JP, Lee WM, et al. Increased H1N1 infection rate in children with asthma. Am J Respir Crit Care Med. 2012;185(12):1275–1279. doi:10.1164/rccm.201109-1635OC
65. Grohskopf LA, Blanton LH, Ferdinands JM, et al. Prevention and control of seasonal influenza with vaccines: recommendations of the advisory committee on immunization practices - United States, 2022–23 influenza season. MMWR Recomm Rep. 2022;71(1):1–28. doi:10.15585/mmwr.rr7101a1
66. Williams JV, Harris PA, Tollefson SJ, et al. Human metapneumovirus and lower respiratory tract disease in otherwise healthy infants and children. N Engl J Med. 2004;350(5):443–450. doi:10.1056/NEJMoa025472
67. Agapov E, Sumino KC, Gaudreault-Keener M, Storch GA, Holtzman MJ. Genetic variability of human metapneumovirus infection: evidence of a shift in viral genotype without a change in illness. J Infect Dis. 2006;193(3):396–403. doi:10.1086/499310
68. Bakakos A, Krompa A. Asthma in the era of SARS CoV-2 virus. J Asthma. 2022;59(8):1501–1508. doi:10.1080/02770903.2021.1941093
69. Yao Y, Wang H, Liu Z. Expression of ACE2 in airways: implication for COVID-19 risk and disease management in patients with chronic inflammatory respiratory diseases. Clin Exp Allergy. 2020;50(12):1313–1324. doi:10.1111/cea.13746
70. Hoffmann M, Kleine-Weber H, Schroeder S, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–280 e278. doi:10.1016/j.cell.2020.02.052
71. Wu Z, McGoogan JM. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: summary of a report of 72 314 cases from the Chinese center for disease control and prevention. JAMA. 2020;323(13):1239–1242. doi:10.1001/jama.2020.2648
72. Li X, Xu S, Yu M, et al. Risk factors for severity and mortality in adult COVID-19 inpatients in Wuhan. J Allergy Clin Immunol. 2020;146(1):110–118. doi:10.1016/j.jaci.2020.04.006
73. Zhang JJ, Dong X, Cao YY, et al. Clinical characteristics of 140 patients infected with SARS-CoV-2 in Wuhan, China. Allergy. 2020;75(7):1730–1741. doi:10.1111/all.14238
74. Zhang JJ, Cao YY, Dong X, et al. Distinct characteristics of COVID-19 patients with initial rRT-PCR-positive and rRT-PCR-negative results for SARS-CoV-2. Allergy. 2020;75(7):1809–1812. doi:10.1111/all.14316
75. Adir Y, Saliba W, Beurnier A, Humbert M. Asthma and COVID-19: an update. Eur Respir Rev. 2021;30(162):210152. doi:10.1183/16000617.0152-2021
76. Williamson EJ, Walker AJ, Bhaskaran K, et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature. 2020;584(7821):430–436. doi:10.1038/s41586-020-2521-4
77. Liu S, Cao Y, Du T, Zhi Y. Prevalence of comorbid asthma and related outcomes in COVID-19: a systematic review and meta-analysis. J Allergy Clin Immunol Pract. 2021;9(2):693–701. doi:10.1016/j.jaip.2020.11.054
78. Choi YJ, Park JY, Lee HS, et al. Effect of asthma and asthma medication on the prognosis of patients with COVID-19. Eur Respir J. 2021;57(3):2002226. doi:10.1183/13993003.02226-2020
79. Peters MC, Sajuthi S, Deford P, et al. COVID-19-related genes in sputum cells in asthma. Relationship to demographic features and corticosteroids. Am J Respir Crit Care Med. 2020;202(1):83–90. doi:10.1164/rccm.202003-0821OC
80. Chen CH, Chen CY, Lai CC, et al. The association between inhaled corticosteroid and the risks of SARS-COV-2 infection: a systematic review and meta-analysis. J Infect Public Health. 2023;16(5):823–830. doi:10.1016/j.jiph.2023.03.019
81. Song JY, Yoon JG, Seo YB, et al. Ciclesonide inhaler treatment for mild-to-moderate COVID-19: a randomized, open-label, phase 2 trial. J Clin Med. 2021;10(16):3545. doi:10.3390/jcm10163545
82. Ramakrishnan S, Nicolau DV Jr, Langford B, et al. Inhaled budesonide in the treatment of early COVID-19 (STOIC): a phase 2, open-label, randomised controlled trial. Lancet Respir Med. 2021;9(7):763–772. doi:10.1016/S2213-2600(21)00160-0
83. Yu LM, Bafadhel M, Dorward J, et al. Inhaled budesonide for COVID-19 in people at high risk of complications in the community in the UK (PRINCIPLE): a randomised, controlled, open-label, adaptive platform trial. Lancet. 2021;398(10303):843–855. doi:10.1016/S0140-6736(21)01744-X
84. Halpin DMG, Singh D, Hadfield RM. Inhaled corticosteroids and COVID-19: a systematic review and clinical perspective. Eur Respir J. 2020;55(5):2001009. doi:10.1183/13993003.01009-2020
85. Farne H, Singanayagam A. Why asthma might surprisingly protect against poor outcomes in COVID-19. Eur Respir J. 2020;56(6):2003045. doi:10.1183/13993003.03045-2020
86. Ehre C, Worthington EN, Liesman RM, et al. Overexpressing mouse model demonstrates the protective role of Muc5ac in the lungs. Proc Natl Acad Sci U S A. 2012;109(41):16528–16533. doi:10.1073/pnas.1206552109
87. Boncristiani HF, Criado MF, Arruda E. Respiratory viruses. Ency Microbiol. 2009;500–5189. doi:10.1016/B978-012373944-5.00314-X
88. Message SD, Johnston SL. The immunology of virus infection in asthma. Eur Respir J. 2001;18(6):1013–1025. doi:10.1183/09031936.01.00228701
89. van Rijt LS, van Kessel CH, Boogaard I, Lambrecht BN. Respiratory viral infections and asthma pathogenesis: a critical role for dendritic cells? J Clin Virol. 2005;34(3):161–169. doi:10.1016/j.jcv.2005.07.002
90. Oliver BG, Robinson P, Peters M, Black J. Viral infections and asthma: an inflammatory interface? Eur Respir J. 2014;44(6):1666–1681. doi:10.1183/09031936.00047714
91. Hansbro NG, Horvat JC, Wark PA, Hansbro PM. Understanding the mechanisms of viral induced asthma: new therapeutic directions. Pharmacol Ther. 2008;117(3):313–353.
92. Illi S, von Mutius E, Lau S, et al. Early childhood infectious diseases and the development of asthma up to school age: a birth cohort study. BMJ. 2001;322(7283):390–395. doi:10.1136/bmj.322.7283.390
93. Dakhama A, Lee YM, Gelfand EW. Virus-induced airway dysfunction: pathogenesis and biomechanisms. Pediatr Infect Dis J. 2005;24(11 Suppl):S159–S169, discussion S166–S157. doi:10.1097/01.inf.0000188155.46381.15
94. Folkerts G, Busse WW, Nijkamp FP, Sorkness R, Gern JE. Virus-induced airway hyperresponsiveness and asthma. Am J Respir Crit Care Med. 1998;157(6 Pt 1):1708–1720. doi:10.1164/ajrccm.157.6.9707163
95. Lommatzsch M, Korn S, Buhl R, Virchow JC. Against all odds: anti-IgE for intrinsic asthma? Thorax. 2014;69(1):94–96. doi:10.1136/thoraxjnl-2013-203738
96. Ackland J, Watson A, Wilkinson TMA, Staples KJ. Interrupting the conversation: implications for crosstalk between viral and bacterial infections in the asthmatic airway. Front Allergy. 2021;2:738987. doi:10.3389/falgy.2021.738987
97. Talbot TR, Hartert TV, Mitchel E, et al. Asthma as a risk factor for invasive pneumococcal disease. N Engl J Med. 2005;352(20):2082–2090. doi:10.1056/NEJMoa044113
98. Levy ML, Bacharier LB, Bateman E, et al. Key recommendations for primary care from the 2022 Global Initiative for Asthma (GINA) update. NPJ Prim Care Respir Med. 2023;33(1):7. doi:10.1038/s41533-023-00330-1
99. Fergeson JE, Patel SS, Lockey RF. Acute asthma, prognosis, and treatment. J Allergy Clin Immunol. 2017;139(2):438–447. doi:10.1016/j.jaci.2016.06.054
100. Rakes GP, Arruda E, Ingram JM, et al. Rhinovirus and respiratory syncytial virus in wheezing children requiring emergency care: igE and eosinophil analyses. Am J Respir Crit Care Med. 1999;159(3):785–790. doi:10.1164/ajrccm.159.3.9801052
101. Johnston SL, Pattemore PK, Sanderson G, et al. Community study of role of viral infections in exacerbations of asthma in 9–11 year old children. BMJ. 1995;310(6989):1225–1229. doi:10.1136/bmj.310.6989.1225
102. Murray CS, Poletti G, Kebadze T, et al. Study of modifiable risk factors for asthma exacerbations: virus infection and allergen exposure increase the risk of asthma hospital admissions in children. Thorax. 2006;61(5):376–382. doi:10.1136/thx.2005.042523
103. Green RM, Custovic A, Sanderson G, Hunter J, Johnston SL, Woodcock A. Synergism between allergens and viruses and risk of hospital admission with asthma: case-control study. BMJ. 2002;324(7340):763. doi:10.1136/bmj.324.7340.763
104. Georas SN, Rezaee F. Epithelial barrier function: at the front line of asthma immunology and allergic airway inflammation. J Allergy Clin Immunol. 2014;134(3):509–520. doi:10.1016/j.jaci.2014.05.049
105. Jackson DJ, Johnston SL. The role of viruses in acute exacerbations of asthma. J Allergy Clin Immunol. 2010;125(6):1178–1187; quiz 1188–1179. doi:10.1016/j.jaci.2010.04.021
106. Johnson SM, McNally BA, Ioannidis I, et al. Respiratory syncytial virus uses CX3CR1 as a receptor on primary human airway epithelial cultures. PLoS Pathog. 2015;11(12):e1005318. doi:10.1371/journal.ppat.1005318
107. Basnet S, Palmenberg AC, Gern JE. Rhinoviruses and their receptors. Chest. 2019;155(5):1018–1025. doi:10.1016/j.chest.2018.12.012
108. Bochkov YA, Watters K, Ashraf S, et al. Cadherin-related family member 3, a childhood asthma susceptibility gene product, mediates rhinovirus C binding and replication. Proc Natl Acad Sci U S A. 2015;112(17):5485–5490. doi:10.1073/pnas.1421178112
109. Moore WC, Hastie AT, Li X, et al. Sputum neutrophil counts are associated with more severe asthma phenotypes using cluster analysis. J Allergy Clin Immunol. 2014;133(6):1557–1563 e1555. doi:10.1016/j.jaci.2013.10.011
110. Douwes J, Gibson P, Pekkanen J, Pearce N. Non-eosinophilic asthma: importance and possible mechanisms. Thorax. 2002;57(7):643–648. doi:10.1136/thorax.57.7.643
111. Rodriguez-Roisin R. Acute severe asthma: pathophysiology and pathobiology of gas exchange abnormalities. Eur Respir J. 1997;10(6):1359–1371. doi:10.1183/09031936.97.10061359
112. Pepe PE, Marini JJ. Occult positive end-expiratory pressure in mechanically ventilated patients with airflow obstruction: the auto-PEEP effect. Am Rev Respir Dis. 1982;126(1):166–170. doi:10.1164/arrd.1982.126.1.166
113. Barnes PJ. How corticosteroids control inflammation: Quintiles prize lecture 2005. Br J Pharmacol. 2006;148(3):245–254. doi:10.1038/sj.bjp.0706736
114. Edwards MR, Johnson MW, Johnston SL. Combination therapy: synergistic suppression of virus-induced chemokines in airway epithelial cells. Am J Respir Cell Mol Biol. 2006;34(5):616–624. doi:10.1165/rcmb.2005-0385OC
115. Rowe BH, Spooner C, Ducharme FM, Bretzlaff JA, Bota GW. Early emergency department treatment of acute asthma with systemic corticosteroids. Cochrane Database Syst Rev. 1996;2001(1):CD002178.
116. Krishnan JA, Davis SQ, Naureckas ET, Gibson P, Rowe BH. An umbrella review: corticosteroid therapy for adults with acute asthma. Am J Med. 2009;122(11):977–991. doi:10.1016/j.amjmed.2009.02.013
117. Price D, Castro M, Bourdin A, Fucile S, Altman P. Short-course systemic corticosteroids in asthma: striking the balance between efficacy and safety. Eur Respir Rev. 2020;29(155):190151. doi:10.1183/16000617.0151-2019
118. Griffiths B, Ducharme FM. Combined inhaled anticholinergics and short-acting beta2-agonists for initial treatment of acute asthma in children. Cochrane Database Syst Rev. 2013;8:CD000060. doi:10.1002/14651858.CD000060.pub2
119. Rodrigo GJ, Castro-Rodriguez JA. Anticholinergics in the treatment of children and adults with acute asthma: a systematic review with meta-analysis. Thorax. 2005;60(9):740–746. doi:10.1136/thx.2005.047803
120. Cosio BG, Mann B, Ito K, et al. Histone acetylase and deacetylase activity in alveolar macrophages and blood mononocytes in asthma. Am J Respir Crit Care Med. 2004;170(2):141–147. doi:10.1164/rccm.200305-659OC
121. Celis P, Rada G. High-dose inhaled corticosteroids or addition of theophylline in patients with poorly controlled asthma? Medwave. 2015;15(Suppl 2):e6224. doi:10.5867/medwave.2015.6224
122. BinMahfouz H, Borthakur B, Yan D, George T, Giembycz MA, Newton R. Superiority of combined phosphodiesterase PDE3/PDE4 inhibition over PDE4 inhibition alone on glucocorticoid- and long-acting beta2-adrenoceptor agonist-induced gene expression in human airway epithelial cells. Mol Pharmacol. 2015;87(1):64–76. doi:10.1124/mol.114.093393
123. Lipworth BJ. Phosphodiesterase-4 inhibitors for asthma and chronic obstructive pulmonary disease. Lancet. 2005;365(9454):167–175. doi:10.1016/S0140-6736(05)17708-3
124. Bardin P, Kanniess F, Gauvreau G, Bredenbroker D, Rabe KF. Roflumilast for asthma: efficacy findings in mechanism of action studies. Pulm Pharmacol Ther. 2015;35(Suppl):S4–S10. doi:10.1016/j.pupt.2015.08.006
125. Laidlaw TM, Boyce JA. Cysteinyl leukotriene receptors, old and new; implications for asthma. Clin Exp Allergy. 2012;42(9):1313–1320. doi:10.1111/j.1365-2222.2012.03982.x
126. Camargo CA Jr, Gurner DM, Smithline HA, et al. A randomized placebo-controlled study of intravenous montelukast for the treatment of acute asthma. J Allergy Clin Immunol. 2010;125(2):374–380. doi:10.1016/j.jaci.2009.11.015
127. Seymour ML, Gilby N, Bardin PG, et al. Rhinovirus infection increases 5-lipoxygenase and cyclooxygenase-2 in bronchial biopsy specimens from nonatopic subjects. J Infect Dis. 2002;185(4):540–544. doi:10.1086/338570
128. Message SD, Laza-Stanca V, Mallia P, et al. Rhinovirus-induced lower respiratory illness is increased in asthma and related to virus load and Th1/2 cytokine and IL-10 production. Proc Natl Acad Sci U S A. 2008;105(36):13562–13567. doi:10.1073/pnas.0804181105
129. Tay H, Wark PA, Bartlett NW. Advances in the treatment of virus-induced asthma. Expert Rev Respir Med. 2016;10(6):629–641. doi:10.1080/17476348.2016.1180249
130. Barlow JL, Bellosi A, Hardman CS, et al. Innate IL-13-producing nuocytes arise during allergic lung inflammation and contribute to airways hyperreactivity. J Allergy Clin Immunol. 2012;129(1):191–198 e191–194. doi:10.1016/j.jaci.2011.09.041
131. Bartemes KR, Kephart GM, Fox SJ, Kita H. Enhanced innate type 2 immune response in peripheral blood from patients with asthma. J Allergy Clin Immunol. 2014;134(3):671–678 e674. doi:10.1016/j.jaci.2014.06.024
132. Jatakanon A, Lim S, Barnes PJ. Changes in sputum eosinophils predict loss of asthma control. Am J Respir Crit Care Med. 2000;161(1):64–72. doi:10.1164/ajrccm.161.1.9809100
133. Contoli M, Ito K, Padovani A, et al. Th2 cytokines impair innate immune responses to rhinovirus in respiratory epithelial cells. Allergy. 2015;70(8):910–920. doi:10.1111/all.12627
134. Steinke JW. Anti-interleukin-4 therapy. Immunol Allergy Clin North Am. 2004;24(4):599–614, vi. doi:10.1016/j.iac.2004.06.008
135. Hart TK, Blackburn MN, Brigham-Burke M, et al. Preclinical efficacy and safety of pascolizumab (SB 240683): a humanized anti-interleukin-4 antibody with therapeutic potential in asthma. Clin Exp Immunol. 2002;130(1):93–100. doi:10.1046/j.1365-2249.2002.01973.x
136. Hanania NA, Noonan M, Corren J, et al. Lebrikizumab in moderate-to-severe asthma: pooled data from two randomised placebo-controlled studies. Thorax. 2015;70(8):748–756. doi:10.1136/thoraxjnl-2014-206719
137. Piper E, Brightling C, Niven R, et al. A phase II placebo-controlled study of tralokinumab in moderate-to-severe asthma. Eur Respir J. 2013;41(2):330–338. doi:10.1183/09031936.00223411
138. Wenzel S, Ford L, Pearlman D, et al. Dupilumab in persistent asthma with elevated eosinophil levels. N Engl J Med. 2013;368(26):2455–2466. doi:10.1056/NEJMoa1304048
139. Bakakos A, Schleich F, Bakakos P. Biological therapy of severe asthma and nasal polyps. J Pers Med. 2022;12(6):976. doi:10.3390/jpm12060976
140. Ortega HG, Liu MC, Pavord ID, et al. Mepolizumab treatment in patients with severe eosinophilic asthma. N Engl J Med. 2014;371(13):1198–1207. doi:10.1056/NEJMoa1403290
141. Efthimiou J, Poll C, Barnes PJ. Dual mechanism of action of T2 inhibitor therapies in virally induced exacerbations of asthma: evidence for a beneficial counter-regulation. Eur Respir J. 2019;54(1):1802390. doi:10.1183/13993003.02390-2018
142. MacGlashan DW Jr, Bochner BS, Adelman DC, et al. Down-regulation of Fc(epsilon)RI expression on human basophils during in vivo treatment of atopic patients with anti-IgE antibody. J Immunol. 1997;158(3):1438–1445. doi:10.4049/jimmunol.158.3.1438
143. Walker S, Monteil M, Phelan K, Lasserson TJ, Walters EH. Anti-IgE for chronic asthma in adults and children. Cochrane Database Syst Rev. 2006;2:CD003559. doi:10.1002/14651858.CD003559.pub3
144. Teach SJ, Gill MA, Togias A, et al. Preseasonal treatment with either omalizumab or an inhaled corticosteroid boost to prevent fall asthma exacerbations. J Allergy Clin Immunol. 2015;136(6):1476–1485. doi:10.1016/j.jaci.2015.09.008
145. Busse WW, Morgan WJ, Gergen PJ, et al. Randomized trial of omalizumab (anti-IgE) for asthma in inner-city children. N Engl J Med. 2011;364(11):1005–1015. doi:10.1056/NEJMoa1009705
146. Chanez P, Contin-Bordes C, Garcia G, et al. Omalizumab-induced decrease of FcxiRI expression in patients with severe allergic asthma. Respir Med. 2010;104(11):1608–1617. doi:10.1016/j.rmed.2010.07.011
147. Mahmutovic-Persson I, Akbarshahi H, Bartlett NW, et al. Inhaled dsRNA and rhinovirus evoke neutrophilic exacerbation and lung expression of thymic stromal lymphopoietin in allergic mice with established experimental asthma. Allergy. 2014;69(3):348–358. doi:10.1111/all.12329
148. Ballantyne SJ, Barlow JL, Jolin HE, et al. Blocking IL-25 prevents airway hyperresponsiveness in allergic asthma. J Allergy Clin Immunol. 2007;120(6):1324–1331. doi:10.1016/j.jaci.2007.07.051
149. Beale J, Jayaraman A, Jackson DJ, et al. Rhinovirus-induced IL-25 in asthma exacerbation drives type 2 immunity and allergic pulmonary inflammation. Sci Transl Med. 2014;6(256):256ra134. doi:10.1126/scitranslmed.3009124
150. Jackson DJ, Makrinioti H, Rana BM, et al. IL-33-dependent type 2 inflammation during rhinovirus-induced asthma exacerbations in vivo. Am J Respir Crit Care Med. 2014;190(12):1373–1382. doi:10.1164/rccm.201406-1039OC
151. Wechsler ME, Ruddy MK, Pavord ID, et al. Efficacy and safety of Itepekimab in patients with moderate-to-severe asthma. N Engl J Med. 2021;385(18):1656–1668. doi:10.1056/NEJMoa2024257
152. Menzies-Gow A, Corren J, Bourdin A, et al. Tezepelumab in adults and adolescents with severe, uncontrolled asthma. N Engl J Med. 2021;384(19):1800–1809. doi:10.1056/NEJMoa2034975
153. Corren J, Parnes JR, Wang L, et al. Tezepelumab in adults with uncontrolled asthma. N Engl J Med. 2017;377(10):936–946. doi:10.1056/NEJMoa1704064
154. Calderon AA, Dimond C, Choy DF, et al. Targeting interleukin-33 and thymic stromal lymphopoietin pathways for novel pulmonary therapeutics in asthma and COPD. Eur Respir Rev. 2023;32(167):220144. doi:10.1183/16000617.0144-2022
155. Hatchwell L, Collison A, Girkin J, et al. Toll-like receptor 7 governs interferon and inflammatory responses to rhinovirus and is suppressed by IL-5-induced lung eosinophilia. Thorax. 2015;70(9):854–861. doi:10.1136/thoraxjnl-2014-205465
156. Greene CM, McElvaney NG. Toll-like receptor expression and function in airway epithelial cells. Arch Immunol Ther Exp. 2005;53(5):418–427.
157. Kline JN, Waldschmidt TJ, Businga TR, et al. Modulation of airway inflammation by CpG oligodeoxynucleotides in a murine model of asthma. J Immunol. 1998;160(6):2555–2559. doi:10.4049/jimmunol.160.6.2555
158. Borden EC, Sen GC, Uze G, et al. Interferons at age 50: past, current and future impact on biomedicine. Nat Rev Drug Discov. 2007;6(12):975–990. doi:10.1038/nrd2422
159. Schoggins JW. Interferon-stimulated genes: roles in viral pathogenesis. Curr Opin Virol. 2014;6:40–46. doi:10.1016/j.coviro.2014.03.006
160. Traub S, Nikonova A, Carruthers A, et al. An anti-human ICAM-1 antibody inhibits rhinovirus-induced exacerbations of lung inflammation. PLoS Pathog. 2013;9(8):e1003520. doi:10.1371/journal.ppat.1003520
161. McGregor MC, Krings JG, Nair P, Castro M. Role of biologics in asthma. Am J Respir Crit Care Med. 2019;199(4):433–445. doi:10.1164/rccm.201810-1944CI
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A Comparison of the Effectiveness of Asthma Medications on Asthma Exacerbations in Real World National Cohort
Park HJ, Jeon S, Lee HS, Kim BY, Chae YJ, Kim GO, Park JW, Lee JH
Journal of Asthma and Allergy 2022, 15:1155-1165
Published Date: 24 August 2022
Relationship Between Asthma Control as Measured by the Asthma Impairment and Risk Questionnaire (AIRQ) and Patient Perception of Disease Status, Health-Related Quality of Life, and Treatment Adherence
Reibman J, Chipps BE, Zeiger RS, Beuther DA, Wise RA, McCann W, Gilbert I, Eudicone JM, Gandhi HN, Harding G, Cutts K, Coyne KS, Murphy KR, George M
Journal of Asthma and Allergy 2023, 16:59-72
Published Date: 5 January 2023
Mometasone/Indacaterol/Glycopyrronium (MF/IND/GLY) and MF/IND at Different MF Strengths versus Fluticasone Propionate/Salmeterol Xinafoate (FLU/SAL) and FLU/SAL+ Tiotropium in Patients with Asthma
van Zyl-Smit RN, Chapman KR, Kerstjens HAM, Gessner C, Sagara H, Tanase AM, Hosoe M, Pethe A, Lawrence D, Tillmann HC, D'Andrea P
Journal of Asthma and Allergy 2023, 16:123-134
Published Date: 20 January 2023
Examining Influenza Vaccination Patterns Among Young Adults with Asthma: Insights into Knowledge, Attitudes, and Practices
Al-Qerem W, Alassi A, Jarab A, Al Bawab AQ, Hammad A, Alasmari F, Alazab B, Abu Husein D, AL Momani N, Eberhardt J
Patient Preference and Adherence 2023, 17:2899-2913
Published Date: 10 November 2023
Real-World Disease Burden and Healthcare Resource Utilization Among Patients with COPD and Asthma Using Triple Therapy (FF/UMEC/VI) in the United States
Igboekwe E, Verma S, Paczkowski R
International Journal of Chronic Obstructive Pulmonary Disease 2024, 19:281-296
Published Date: 25 January 2024