Back to Journals » International Journal of Nanomedicine » Volume 21
Environmental Micro-Nanoplastics and Musculoskeletal Disorders: Current Research and Insights
Authors Ju C, Dong H, Liu R, Ma Y, Luo R, Hu H
Received 4 March 2026
Accepted for publication 16 May 2026
Published 30 May 2026 Volume 2026:21 606793
DOI https://doi.org/10.2147/IJN.S606793
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Cheng Ju,1,2,* Hui Dong,1,2,* Renfeng Liu,1,2,* Yanming Ma,1,2 Rongjin Luo,1,2 Huimin Hu1,2
1Department of Spine Surgery, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China; 2Shaanxi Key Laboratory of Spine Bionic Treatment, Xi’an, Shaanxi, People’s Republic of China
*These authors have contributed equally to this work
Correspondence: Huimin Hu; Rongjin Luo
Department of Spine Surgery, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China
, Email [email protected]; [email protected]
Abstract: In recent years, micro-nanoplastics, as globally prevalent emerging environmental pollutants, can enter the human body through oral ingestion, respiratory inhalation, and dermal contact, and widely distribute in diverse tissues including blood, placenta, lung, bone marrow, and bone, thereby arousing widespread concern regarding their health risks. As an essential functional system of the human body, the skeletal system is involved in mechanical support, metabolic homeostasis, and immune regulation. Disruption of skeletal homeostasis may induce a series of musculoskeletal diseases, such as osteoporosis, rheumatoid arthritis, intervertebral disc degeneration, and osteoarthritis. Accumulating evidence has verified that micro-nanoplastics can accumulate in human bone, cartilage, intervertebral disc, and bone marrow, and destroy skeletal homeostasis via multiple mechanisms, including inflammatory response, oxidative stress, pyroptosis, ferroptosis, cellular senescence, and epigenetic regulation, thus participating in the occurrence and development of musculoskeletal diseases. This paper systematically reviews the sources, classification, environmental distribution, human exposure pathways, and systemic toxic effects of micro-nanoplastics. It emphatically expounds the impacts and potential molecular mechanisms of micro-nanoplastics on osteoporosis, intervertebral disc degeneration, rheumatoid arthritis, osteoarthritis, lupus-related musculoskeletal lesions, and osteosarcoma progression. Furthermore, we summarize the detection methods and research limitations of micro-nanoplastics in skeletal tissues, and prospect future directions in environmental prevention and control, clinical translation, and high-risk population monitoring. This review is expected to provide novel ideas for mechanistic investigation, risk assessment, and clinical translation of micro-nanoplastic-associated skeletal diseases.
Plain Language Summary: 1. This review systematically summarizes the association between environmental micro-nanoplastics and skeletal homeostasis, as well as the latest research evidence in musculoskeletal disorders.
2. This review elucidates the core mechanisms of skeletal system damage induced by micro-nanoplastics, including inflammatory responses, oxidative stress, pyroptosis, ferroptosis, cellular senescence, and epigenetic regulation.
3. This review proposes future research directions and potential strategies for micro-nanoplastics in skeletal disease studies. The diagram illustrates the environmental sources of micro and nanoplastics, exposure routes, disruption of skeletal homeostasis and potential musculoskeletal diseases. On the left, sources include plastic packaging, drinking water, occupational exposure, food or seafood, airborne particles and synthetic textiles. Exposure routes are ingestion, inhalation and dermal contact, shown affecting a human figure. The center shows disruption of skeletal homeostasis, highlighting bone, cartilage, intervertebral disc, bone marrow and synovium, with a balance scale indicating bone formation and resorption. On the right, potential musculoskeletal diseases listed are osteoporosis, osteoarthritis, rheumatoid arthritis, intervertebral disc degeneration, lupus-related synovitis and osteosarcoma-related conditions.Diagram of MNPs sources, exposure routes, skeletal disruption and potential musculoskeletal diseases.
Keywords: microplastics, nanoplastics, skeletal homeostasis, musculoskeletal diseases, ecological environment pollution
Introduction
Microplastics, defined as plastic particles, films, and fibers with diameters less than 5 mm, whereas nanoplastics generally refer to plastic particles within the nanoscale range; however, their size thresholds have not been fully standardized across studies. Together, they have become ubiquitous environmental contaminants.1–3 Over the past few decades, the rapid increase in plastic production, coupled with the strong environmental persistence of plastic waste, has led to the widespread occurrence of micro- and nanoplastic pollution.4–6 Research has documented the presence of Micro- and nanoplastics in soil, freshwater, sediments, plants, animals, the atmosphere, aquatic ecosystems, and even human bodies.7–9 Humans are exposed to micro- and nanoplastics mainly through oral ingestion, inhalation, and potential dermal contact. A growing body of evidence shows that micro- and nanoplastics are present in human blood, placenta, lung tissue, breast milk, liver, spleen, colon, urine, testes, and semen, suggesting that they may cross biological barriers and undergo systemic distribution.9–11 The health risks associated with micro- and nanoplastics have therefore attracted considerable global concern, and these pollutants are regarded as important contributors to ecological degradation and potential threats to human health.12,13 The skeletal system comprises bones, joints, tendons, ligaments, and other components and is essential for mechanical support, mineral storage, hematopoiesis, endocrine regulation, and immune modulation.14–16 Skeletal homeostasis refers to the dynamic equilibrium that maintains bone mass, bone microarchitecture, cartilage integrity, intervertebral disc function, mineral metabolism, and motor function. This process depends on coordinated regulation among osteoblasts, osteoclasts, osteocytes, bone marrow stromal cells, chondrocytes, nucleus pulposus cells, synovial fibroblasts, immune cells, and endocrine factors. Under physiological conditions, osteoblast-mediated bone formation and osteoclast-mediated bone resorption remain dynamically coupled during bone remodeling, whereas cartilage and intervertebral disc tissues preserve joint and spinal homeostasis by maintaining extracellular matrix metabolism and biomechanical function. Disruption of skeletal homeostasis—triggered by inflammation, foreign body exposure, degenerative changes, or metabolic abnormalities—can lead to osteoporosis, fractures, intervertebral disc degeneration, arthritis with motor dysfunction, and immune/inflammatory cascades. These pathological alterations significantly impair human health and quality of life, imposing substantial economic burdens on both society and individuals.17–20 Therefore, investigating the adverse effects of microplastics on the skeletal system is crucial for public health, and exploring potential strategies to mitigate microplastic pollution warrants urgent attention.
The skeletal system may be a potential target of long-term micro- and nanoplastic exposure. After entering the human body, micro- and nanoplastics can access the bloodstream and may be transported to highly vascularized or immunologically active tissues, such as bone marrow and synovium. Skeletal homeostasis is closely coupled with immune and inflammatory regulation, and bone remodeling is jointly influenced by cytokines, oxidative stress, macrophage activation, and osteoimmune signaling. Tissues such as cartilage and intervertebral discs have limited regenerative capacity; therefore, sustained inflammatory stimulation and oxidative injury may increase the risk of structural degeneration and functional impairment. Recent studies have detected microplastics in human bone, cartilage, and intervertebral discs, providing direct evidence of skeletal system exposure.21,22 Another study of human bone marrow samples also showed that microplastics can be present in the bone marrow microenvironment, suggesting that their potential effects may not be confined to bone tissue itself.23 On the basis of these findings, the potential effects of micro- and nanoplastics on skeletal homeostasis and the progression of musculoskeletal diseases warrant further investigation.
Although the body of evidence is increasing, the relationship between micro- and nanoplastic exposure and skeletal system diseases remains insufficiently characterized. Most current studies are based on in vitro experiments, animal models, or cross-sectional observations in humans, which are not sufficient to establish a direct causal relationship in the human body. In addition, analytical methods for detecting micro- and nanoplastics in skeletal system samples have not yet been fully standardized. Previous studies have mainly focused on the environmental distribution, systemic toxicity, or general health risks of micro- and nanoplastics, with limited systematic synthesis of their effects on skeletal homeostasis and musculoskeletal diseases. Existing reviews have begun to address the potential effects of microplastics on bone and bone marrow and have suggested possible links with skeletal disorders such as osteoporosis; however, the field still lacks a comprehensive synthesis centered on skeletal homeostasis, disease-related evidence, mechanisms of action, and detection methods.22,24 Therefore, this review summarizes the current evidence on the relationship between micro- and nanoplastics and skeletal and musculoskeletal diseases, with a focus on osteoporosis, rheumatoid arthritis, intervertebral disc degeneration, osteoarthritis, lupus-related musculoskeletal involvement, and osteosarcoma progression. It further discusses potential mechanisms, including inflammatory responses, oxidative stress, pyroptosis, ferroptosis, cellular senescence, and epigenetic regulation; compares methods for detecting micro- and nanoplastics in biological samples and skeletal system tissues; and outlines future directions for clinical translation, exposure monitoring, and public health prevention and control.
Formation, Classification, and Environmental Distribution of Microplastics and Nanoplastics
Classification, Sources, and Formation of Micro- and Nanoplastics
Microplastics can be classified into five major categories: fragments, fibers, foams, particles, and films. Based on their chemical composition, microplastics are divided into six types: polyethylene (PE), polystyrene (PS), polypropylene (PP), polyurethane (PUR), polyvinyl chloride (PVC), and polyethylene terephthalate (PET).25,26 In terms of origin, microplastics are categorized as primary or secondary. Primary microplastics consist of small synthetic polymers intentionally manufactured for use in exfoliating agents, chemical formulations, abrasive products, plastic product maintenance, and synthetic textile production.27,28 Secondary microplastics originate from the fragmentation of larger plastic items or mesoplastics, primarily driven by environmental processes such as biodegradation, photodegradation, thermo-oxidative degradation, thermal degradation, and hydrolytic decomposition. Ultraviolet irradiation, temperature fluctuations, wave action, and abrasion by sand and gravel can all promote the fragmentation of plastic waste and represent important sources of secondary microplastics in the environment.29–31 We illustrate the formation and classification of microplastics in Figure 1 (All figures were drawn by Figdraw). Nanoplastics are generally defined as plastic particles within the nanoscale range. In this review, nanoplastics are operationally defined as plastic particles measuring 1–1000 nm, although some studies and regulatory frameworks adopt a narrower upper size limit of 100 nm.32,33 Compared with microplastics, nanoplastics have smaller particle sizes, larger specific surface areas, and greater reactivity, properties that may enhance their interactions with biological membranes and facilitate translocation across the blood–brain barrier and other biological barriers, thereby conferring higher bioavailability and potential toxicity.34–37 Over the past 70 years, global plastic production has surged from 1.5 million metric tons to approximately 359 million metric tons. Large quantities of plastic waste are incinerated, landfilled, or discharged directly into the environment, further exacerbating micro- and nanoplastic pollution. Owing to their persistence and resistance to degradation, microplastics and nanoplastics have become a major concern in global environmental health.38–40
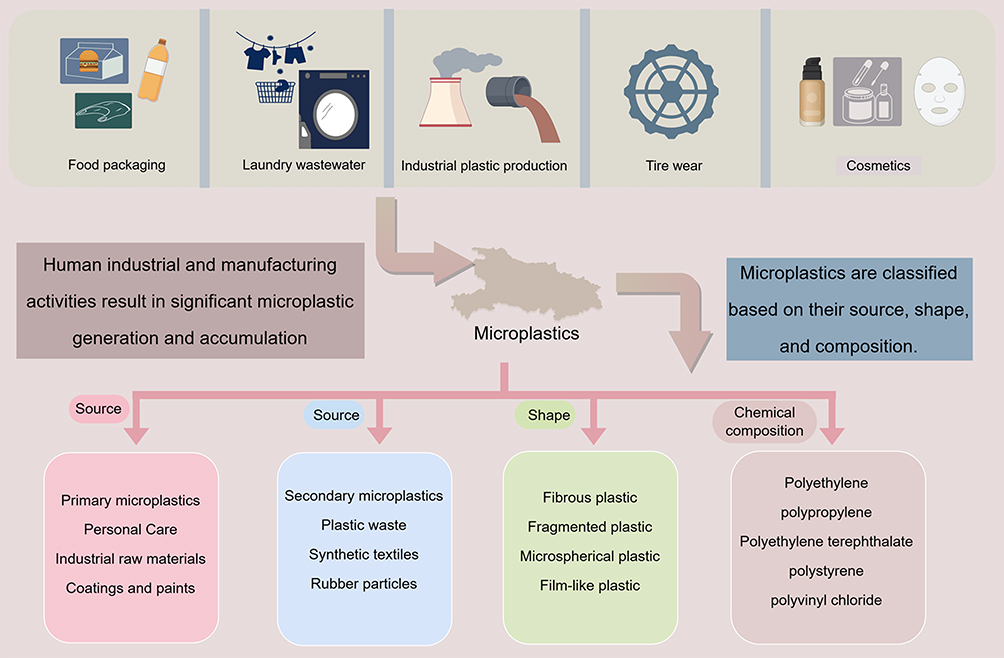 |
Figure 1 The production of micro-nanoplastics is closely linked to human activities, and micro-nanoplastics can be classified based on their source, shape, and composition. |
Environmental Distribution and Ecological Basis of Exposure to Micro- and Nanoplastics
Micro- and nanoplastics are widely distributed across environmental media, including marine waters, freshwater, soil, the atmosphere, sediments, and glaciers. In recent years, the production of microplastics has significantly increased, with concentrations reaching thousands of particles per cubic meter detected in coastal areas of certain marine regions. These minute particles exert substantial environmental impacts, particularly on aquatic organisms, as they accumulate and leach toxic organic and inorganic contaminants.41,42 Furthermore, microplastics are renowned for their stability and resistance to degradation, implying a persistence in the environment for decades.43 Owing to their stability and slow degradation, microplastics can persist in the environment for extended periods and migrate and accumulate across trophic levels through the food chain.44,45 After ingestion by aquatic organisms, microplastics may induce adverse effects such as disrupted feeding behavior, gastrointestinal injury, immune dysfunction, oxidative stress, and growth inhibition.46–49 Their ecotoxicity is closely related to the route of exposure, particle size, morphology, polymer type, and adsorbed contaminants. Direct exposure is typically characterized by acute or localized injury resulting from particle–organism contact, whereas indirect exposure mainly causes chronic toxicity through food-chain transfer and co-exposure to associated contaminants.50–52 These patterns of environmental distribution and ecological accumulation provide an important basis for continuous human exposure to micro- and nanoplastics. Microplastics have been found to exert multiple adverse effects on the environment and organisms (Figure 2).
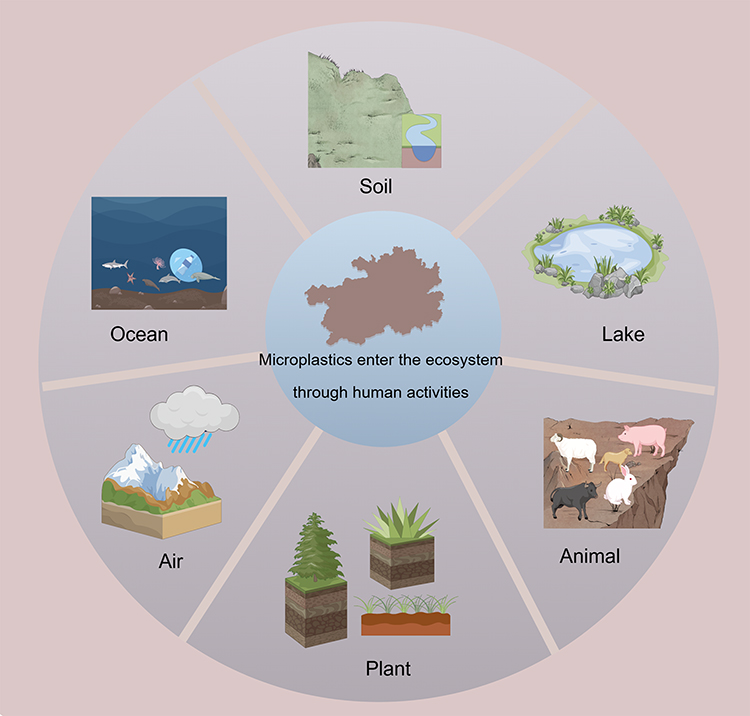 |
Figure 2 The release of micro-nanoplastics severely impacts ecosystems. |
Human Exposure Routes and in vivo Distribution of Micro- and Nanoplastics
Microplastics and nanoplastics have been detected across diverse human samples, with particle sizes ranging from ≥800 nm to 5 mm, exhibiting varied colors and dimensions. These contaminants have been identified in critical biological matrices including lungs, breast milk, liver, spleen, placenta, blood, sputum, colon, saliva, facial tissues, urine, testes, and semen.46,48,53 Humans are exposed to micro- and nanoplastics mainly through oral ingestion, inhalation, and potential dermal contact. Because air, water, soil, and food can all serve as carriers of micro- and nanoplastics, humans may be subject to long-term, low-dose, and multi-route exposure risks.
The Hazards to Human Health from Drinking Water and Food Sources
Aquatic ecosystems are particularly vulnerable to plastic pollution and high-density plastic particle contamination, with approximately 10–20% of microplastics discharged into aquatic environments originating from marine sources.47,54 Reports reveal that over 5.25 trillion particles, totaling 268,940 metric tons, float in the world’s oceans. These alarming figures have raised scholarly concerns regarding the survival and mutagenic potential of aquatic organisms.55–57 Within urban systems, microplastics are released into aquatic environments through urban/industrial wastewater (eg, water treatment facilities, washing machines) runoff or the breakdown of larger debris via physical, biological, and chemical degradation processes. They have been detected in numerous rivers, oceans, and even in polar snow and Antarctic ice layers worldwide.58–60
Drinking water is also an important source of exposure. Previous studies have detected microplastics in tap water, bottled water, and mineral water, with possible sources including contamination of water sources, packaging processes, and pollution along the water supply chain.61–63 Food packaging materials can likewise release microplastics, particularly under high-temperature conditions involving items such as tea bags, plastic kettles, and infant feeding bottles, where particle release may be more pronounced.64–66 During contact between food and plastic packaging, packaging constituents and micro- and nanoplastics may migrate into food, and the gastrointestinal tract is considered an important entry route for nanoplastics into the human body.67–69 After entering the digestive tract, micro- and nanoplastics may affect the intestinal barrier, gut microbiota, and local immune responses, thereby inducing systemic biological effects.
Airborne Sources
Airborne microplastics have recently been detected globally, indicating their ubiquitous presence in the atmosphere and associated inhalation risks to humans.70,71 Studies have reported residual microplastic particles in lung tissue samples of non-smoking adults.72 Jenner et al identified microplastics across all pulmonary regions, predominantly as fibers and fragments.73 Microplastics isolated from bronchoalveolar lavage fluid (BALF) pose a severe concern, as inhaled microplastics have been confirmed to translocate to placental and fetal tissues.74,75 The respiratory epithelium serves as the primary target for inhaled nanoplastics (NPs), with biological reactivity dependent on their physicochemical properties. Atmospheric sources of NPs and microplastics include textiles, dried sludge, agricultural residues, and tire rubber dispersed through mechanical abrasion during vehicle operation.76,77 The pulmonary alveolar surface, spanning approximately 150 m2, features tissue barriers thinner than 1 μm. This barrier enables NP penetration into capillaries, facilitating systemic distribution.78 Autopsy analyses of human lung tissues have confirmed microplastics presence, with polymer particles and fibers detected in 13 out of 20 samples.72 In occupational settings such as plastic production and processing, monitoring of related monomers and metabolites, in addition to particles themselves, may provide useful exposure information. In styrene-related biomonitoring, urinary styrene (StyU) represents styrene itself, whereas mandelic acid (MA) and phenylglyoxylic acid (PGA) are the major urinary metabolites of styrene and can be used to assess human styrene exposure levels.79
Direct Contact
Dermal contact is generally considered a relatively minor route of exposure to microplastics; however, under specific conditions, nanoscale particles may still enter local tissues through sweat glands, hair follicles, or sites of skin barrier disruption.80–82 The stratum corneum serves as an important barrier that prevents particulate matter and chemicals from entering the body, and particle size, surface properties, and skin integrity are key factors influencing transdermal penetration.83,84 Nanoplastics derived from cosmetics and textiles may increase the risk of dermal exposure. Gopinath et al found that nanoplastics extracted from cosmetics exerted cytotoxic effects on keratinocytes, inducing oxidative stress, inhibiting cell proliferation, and promoting cellular senescence.85
Following absorption, microplastics can enter the circulatory system and undergo systemic distribution, potentially harming various organs or tissues. A study by Han et al demonstrated that intratracheally instilled polyethylene (PE)- microplastics accumulated predominantly in mouse lungs, followed by the heart, liver, and gastrointestinal tract.86 Other migration pathways involve the lymphatic system and neural fibers, which contribute to microplastics distribution. In zebrafish, research indicated microplastics accumulation in brain regions of embryos, leading to oxidative DNA damage.87 The transport and distribution of microplastics depend not only on blood flow, tissue affinity, and diffusivity but also on exposure routes and particle characteristics. Yong et al observed the highest bioaccumulation rate of orally administered microplastics in the gut, with an average residence time of ~17 days.88 Additionally, individual health status influences microplastics counts in identical tissues; compared to healthy individuals, higher microplastics levels were detected in the livers of patients with cirrhosis. Thus, the ubiquitous exposure to microplastics and their subsequent systemic adverse effects warrants rigorous attention.89
Systemic Toxic Effects of Micro- and Nanoplastics and Their Relevance to Skeletal Homeostasis
After entering the human body, the health effects of micro- and nanoplastics are not confined to the initial sites of exposure. Current evidence indicates that micro- and nanoplastics may affect the nervous, respiratory, gastrointestinal, reproductive, and immune-related systems through mechanisms involving oxidative stress, inflammatory responses, mitochondrial dysfunction, immune dysregulation, endocrine disruption, and metabolic abnormalities.90–94 For example, particulate exposure can activate neuroinflammatory responses through direct translocation or the systemic release of pro-inflammatory mediators, thereby inducing microglial activation, oxidative stress, and neuronal injury.95,96 Population-based studies suggest that microplastic exposure may be associated with mild cognitive impairment, the risk of Alzheimer’s disease progression, and the incidence of dementia.97,98 Animal studies have also shown that microplastics can affect neurotransmitter metabolism, oxidative stress levels, and behavioral outcomes.99,100
The respiratory system is one of the major entry sites for micro- and nanoplastic exposure. Inhaled microplastics can deposit in the respiratory tract and lung tissue and may disrupt pulmonary homeostasis through local inflammatory responses, epithelial barrier injury, and activation of inflammation-related genes.101,102 The gastrointestinal tract is the primary target site after oral ingestion of micro- and nanoplastics. Previous studies have reported higher fecal microplastic concentrations in patients with inflammatory bowel disease than in healthy individuals, with concentrations correlated with disease severity.103,104 Animal experiments further suggest that exposure to microplastics such as PVC can cause intestinal barrier injury, gut microbiota dysbiosis, metabolic disturbances, and systemic inflammatory responses.105,106 These alterations may further affect the homeostasis of distant organs through disruption of the intestinal barrier, changes in microbial metabolites, and dissemination of immune-inflammatory signals.
Studies on reproductive and developmental toxicity have shown that nanoplastics can impair reproductive capacity across generations, induce abnormal gonadal development, and increase the risk of translocation across the placental barrier.107,108 After pregnant mice were exposed to 20 nm polystyrene nanoplastics, particles were detected in multiple maternal organs as well as in the liver, lungs, heart, kidneys, and brain of fetuses, accompanied by reduced fetal body weight and placental weight.74 These findings suggest that smaller nanoplastics may have greater barrier-penetrating capacity and pose higher developmental toxicity risks.
These systemic effects are linked to skeletal homeostasis at multiple levels. Chronic inflammation and oxidative stress can enhance osteoclast activity, impair osteogenic differentiation, and promote extracellular matrix degradation in cartilage and intervertebral disc cells. Intestinal barrier dysfunction and gut microbiota dysbiosis may affect bone metabolism through the gut–bone axis, while abnormalities in endocrine and energy metabolism may also alter the local and systemic regulatory environment required for bone remodeling. These mechanisms provide a biological basis for understanding the potential skeletal toxicity of micro- and nanoplastics. Given these links, it is necessary to further synthesize the available evidence on the potential effects of micro- and nanoplastics on osteoporosis, rheumatoid arthritis, intervertebral disc degeneration, osteoarthritis, lupus-related musculoskeletal involvement, and osteosarcoma progression. The major systemic toxic effects of micro- and nanoplastics and their potential links with skeletal homeostasis are summarized in Figure 3.
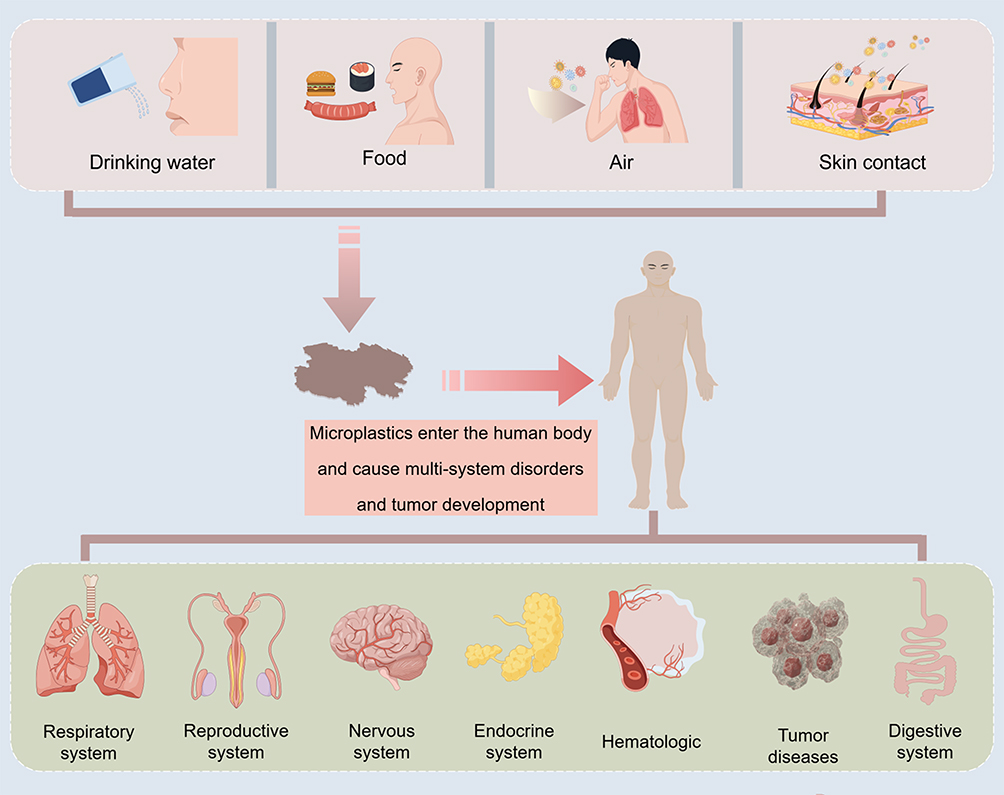 |
Figure 3 Plastics enter the human body through drinking water, food, air, and skin contact, and pose adverse effects on multiple body systems. |
In conclusion, the multi-system issues caused by microplastics in the human body cannot be ignored.
Micro- and Nanoplastics in Skeletal and Musculoskeletal Diseases
The skeletal system is not merely a structural support system; rather, it is a dynamic organ system closely linked to immune, metabolic, endocrine, and inflammatory regulation. In recent years, evidence of microplastics in human bone, cartilage, intervertebral discs, and bone marrow has drawn increasing attention to the potential effects of micro- and nanoplastics on skeletal homeostasis and musculoskeletal diseases. At the disease level, current evidence is mainly concentrated in osteoporosis, rheumatoid arthritis, intervertebral disc degeneration, osteoarthritis, lupus-related musculoskeletal involvement, and osteosarcoma progression. It should be noted that most existing studies are based on cross-sectional human observations, animal experiments, or in vitro mechanistic investigations. Further long-term cohort studies and clinical validation are therefore needed to clarify the potential relationship between micro- and nanoplastic exposure and skeletal diseases. To provide an overview of the current evidence, representative studies linking micro- and nanoplastics to skeletal and musculoskeletal disorders are summarized in Table 1.
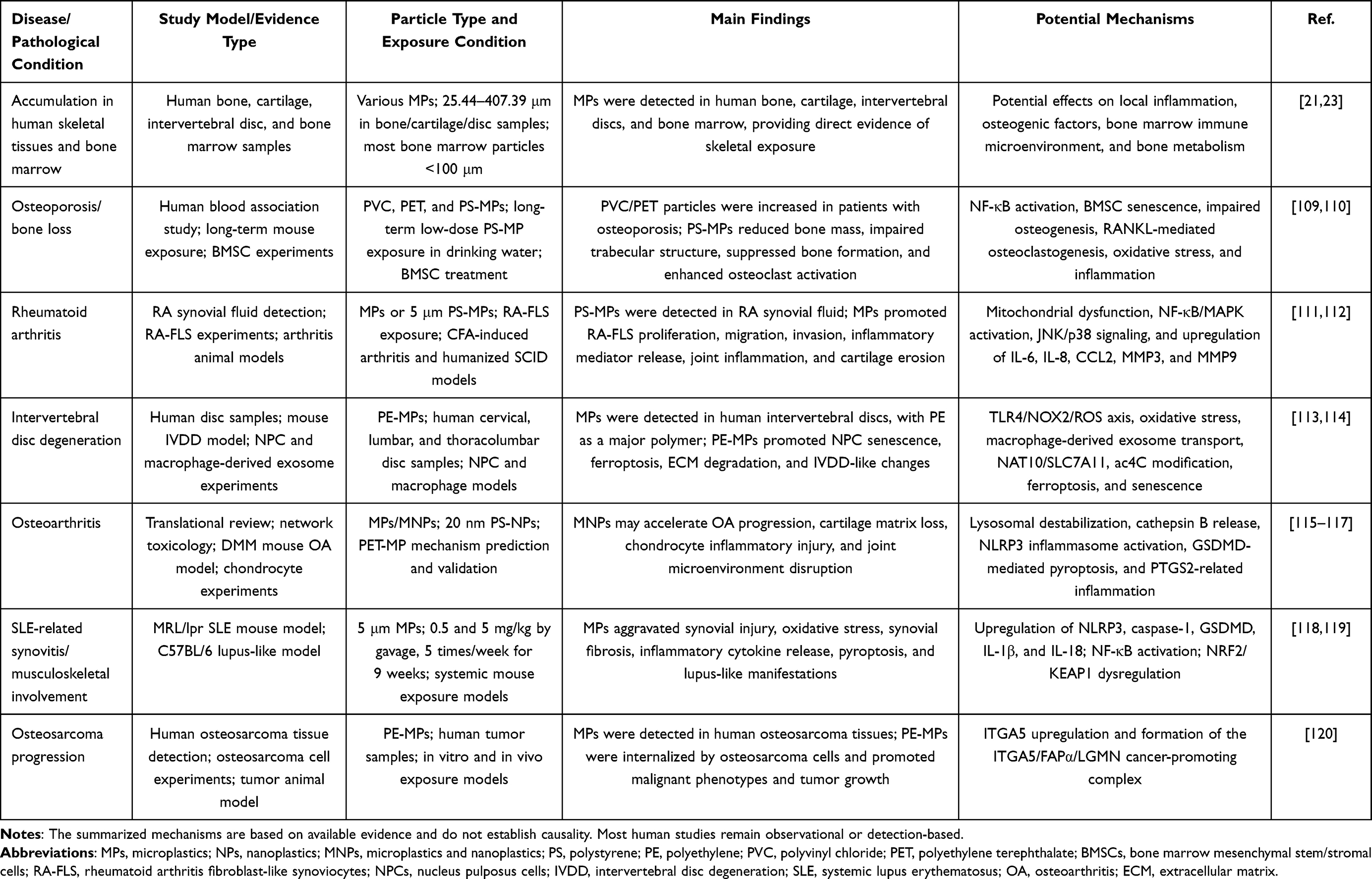 |
Table 1 Summary of Studies Linking Microplastics and Nanoplastics to Skeletal and Musculoskeletal Disorders |
Micro- and Nanoplastics and Osteoporosis
Osteoporosis (OP) is a systemic metabolic bone disease characterized by microarchitectural changes in bone tissue and reduced bone density, which subsequently increase fracture risk, leading to pain, decreased mobility, and deterioration of quality of life, with severe cases posing life-threatening risks.121,122 As the most common skeletal disorder, over 200 million individuals worldwide are affected by OP. Indeed, due to population aging and increasing life expectancy, its incidence continues to rise annually, positioning OP as an increasingly severe global public health concern.123
Bone remodeling is a fundamental process for maintaining bone mass, bone strength, and mineral homeostasis. Under physiological conditions, osteoblast-mediated bone formation and osteoclast-mediated bone resorption remain dynamically coupled. When this balance is disrupted by factors such as aging, menopause, inflammatory responses, endocrine disorders, nutritional deficiency, or metabolic abnormalities, relatively enhanced bone resorption or insufficient bone formation can promote the onset and progression of osteoporosis. Therefore, any exogenous factor capable of affecting osteoblast–osteoclast coupling, the bone marrow microenvironment, or osteoimmune regulation may represent a potential environmental risk factor in osteoporosis research.124–126 Blood samples from 37 OP patients were analyzed for microplastic composition, revealing significantly elevated levels of PVC and PET particles compared to non-osteoporotic individuals. In cellular experiments, the phagocytic capacity of MC3T3 cells incubated with PET or PVC increased over time. Additionally, PET- or PVC-treated cells exhibited higher apoptosis rates, upregulated pro-inflammatory cytokines (IL-1β, TNF-α, and IL-6), and enhanced reactive oxygen species (ROS) responses.109 This human study mainly suggests a potential association between microplastic burden and osteoporosis, but does not establish that PVC or PET microplastics directly cause osteoporosis.
In vivo studies demonstrated that long-term low-dose PS-MP exposure suppressed bone formation, promoted osteoclast activation, caused significant damage to trabecular bone structure, and elevated senescence markers in mice, potentially contributing to OP development. Bone marrow stromal cells (BMSCs) cultured with varying PS-MP concentrations displayed reduced expression of osteogenic markers (Osterix and RUNX2) and increased adipogenic differentiation markers. Mechanistically, the NF-κB pathway played a pivotal role in PS-MP-induced BMSC senescence and osteoclastogenesis, as targeting this pathway reversed MP-mediated BMSC aging.110 In summary, prolonged microplastic accumulation disrupts bone homeostasis by altering the balance between osteogenesis and osteoclasticity, triggering cellular and microenvironmental inflammation, oxidative stress, and impaired osteoblast migration while enhancing osteoclastogenesis.127 Future prospective cohort studies are still needed to further validate the relationships between microplastic exposure levels, reductions in bone mineral density, changes in bone turnover markers, and fracture risk.
Micro- and Nanoplastics and Rheumatoid Arthritis
Rheumatoid arthritis (RA) is a systemic autoimmune inflammatory disease primarily affecting joints. RA can occur in individuals of any age, with higher incidence observed in populations over 40 years old. Women are affected at a frequency 2–3 times greater than men.128 Currently, the pathogenesis of RA is complex, involving abnormal immune cell function, cytokine release, and microbiome-joint axis interactions as key mechanisms. It is characterized by inflammatory changes in synovial tissues, cartilage, and bone, along with less frequent extra-articular inflammation.129
Genetic susceptibility plays a pivotal role, with first-degree relatives of RA patients exhibiting a 2- to 5-fold increased risk compared to the general population.130 Multiple alleles have been associated with elevated RA risk, most notably those related to rheumatoid factor (RF), antibodies targeting post-translationally modified proteins, and T/B lymphocyte migration to synovium. Additionally, environmental factors such as cigarette smoke and dust exposure significantly contribute to aberrant immune system activation, highlighting critical crosstalk between adaptive and innate immune components. Clinically, RA manifests as synovial joint pain, swelling, stiffness, and cartilage/bone destruction.131,132 Fibroblast-like synoviocytes (FLS) in the synovial lining layer have been demonstrated to play a central role in RA pathogenesis. When incubated with varying concentrations of microplastics (0–1.0 mg/mL) for 60 minutes, fluorescently labeled microplastics were internalized into FLSs in a dose- and time-dependent manner. Microplastic stimulation enhanced proliferation markers and migratory capacity of RA-FLSs. Furthermore, microplastic exposure promoted FLS migration in RA and stimulated release of inflammatory cytokines IL-6, IL-1β, and TNFα. In vivo, intra-articular injection of microplastics in mouse knees and paws significantly exacerbated cartilage damage and accelerated arthritis progression.111 It should be noted that the concentrations of microplastics used in some in vitro experiments were higher than realistic environmental exposure levels; therefore, these findings are more appropriate for elucidating potential mechanisms than for directly extrapolating exposure risks to the general population.
Recent studies have further strengthened disease-level evidence linking microplastics to RA. PS-MPs have been detected in the synovial fluid of patients with RA, and 5 μm PS-MPs were shown to be taken up by RA-FLS, activate the NF-κB/MAPK signaling pathways, promote the expression of inflammatory and matrix-degradation-related molecules such as IL-6, IL-8, CCL2, MMP3, and MMP9, and exacerbate joint inflammation and cartilage erosion in animal models.112 Notably, the microplastic concentration range used in these FLS in vitro experiments was 0–1.0 mg/mL, which is higher than most realistic environmental or general population exposure levels. Accordingly, such experimental findings are more suitable for revealing the inflammatory and migratory mechanisms that may be triggered after microplastics are internalized by synovial cells, rather than for directly inferring the risk of RA onset under environmental exposure conditions in the general population.
Overall, current evidence suggests that microplastics may act as environmental particulate stimuli that exacerbate synovial inflammation and joint structural damage in RA. However, direct evidence for their role in RA initiation remains lacking, and these conclusions require further validation using larger patient samples, realistic exposure assessment, and long-term follow-up studies.
Micro- and Nanoplastics and Intervertebral Disc Degeneration
Intervertebral disc degeneration (IDD) is a prevalent cause of joint-related chronic disability in the elderly worldwide. It severely impacts quality of life and imposes substantial socioeconomic burdens. With the aging population and accelerating life pace alongside persistent stress from daily work routines, IDD has become a common underlying pathology for age-related joint dysfunction. As the primary source of low back pain, it serves as the shared pathological basis for multiple spinal disorders including lumbar disc herniation and spinal stenosis.133,134 The intervertebral disc (IVD) comprises the nucleus pulposus (NP), annulus fibrosus (AF), and cartilaginous endplate (CEP), playing critical roles in spinal motion and load bearing. The NP, located centrally within the IVD, contains abundant water and extracellular matrix (ECM) essential for maintaining disc functionality. Key characteristics of IDD include reduced disc height, diminished hydration capacity, and impaired pressure absorption capability. During IVD degeneration, multiple phenotypic changes occur, including decreased NP cell (NPC) quantity, ECM degradation, disorganization of NP and AF tissues, CEP calcification, and microfractures.135,136 Extensive studies demonstrate close associations between inflammation and various pathological processes of IDD, including sustained ECM loss, cell apoptosis, and senescence. Genetic predisposition, aging, obesity, smoking, trauma, and abnormal mechanical loading are generally recognized as contributing factors.20,137
With the widespread use of microplastics, emerging research reveals intimate correlations between microplastic exposure and IDD progression. Raman spectroscopy analyses have detected diverse polymeric microplastic in human cervical, thoracic, and lumbar discs. Animal experiments demonstrate that polyethylene microplastic (PE-MP) exposure induces significant disc degeneration in mice, manifesting as reduced NP thickness, irregular morphology, and decreased cellular density. Mechanistically, microplastic promote disc aging and degeneration by activating the TLR4/NOX2 signaling axis to induce oxidative stress, thereby promoting NP cell senescence and the degenerative disc phenotype.113 Other studies suggest that microplastics may further affect the intervertebral disc microenvironment through intercellular transport and regulated cell death. PE-MPs can be transported to NP cells via macrophage-derived exosomes and induce lipid peroxidation, ferroptosis, cellular senescence, and ECM degradation through NAT10/SLC7A11- and ac4C modification-related pathways, thereby promoting IDD progression.114 In addition, network toxicology and molecular docking analyses further identify AKT1, SRC, CASP3, KDR, PTGS2, PIK3CA, and MAPK14 as key genes mediating PET-MP-driven IVDD progression.138 Overall, IDD is currently one of the areas in skeletal system research on microplastics with relatively direct tissue-level evidence; however, further population-based studies are needed to clarify the relationship between microplastic burden in intervertebral discs and the clinical severity of disc degeneration.
Micro- and Nanoplastics and Osteoarthritis
Osteoarthritis (OA) is a chronic degenerative joint disease characterized by articular cartilage degeneration, subchondral bone remodeling, synovial inflammation, and pain. Aging, obesity, abnormal mechanical loading, joint injury, and low-grade chronic inflammation can all contribute to the onset and progression of OA.139,140 Although clinical evidence regarding the relationship between micro- and nanoplastics and OA remains limited, this area is receiving increasing attention.
Existing translational reviews and mechanistic studies suggest that micro- and nanoplastics may contribute to OA progression through inflammatory responses, oxidative stress, chondrocyte injury, and disruption of the joint microenvironment.115,116 Network toxicology and molecular docking studies also indicate that PET-MPs may affect shared targets and inflammatory pathways related to OA and RA, with molecules such as PTGS2 potentially playing a role in microplastic-associated joint toxicity.117
A recent experimental study further suggested that exposure to 20 nm PS-NPs can accelerate OA progression in mice and promote cartilage matrix loss through lysosomal destabilization, Cathepsin B release, NLRP3 inflammasome activation, and GSDMD-mediated chondrocyte pyroptosis.115 These findings suggest that nanoplastics may act as potential pathological amplifiers within the OA joint microenvironment. However, evidence from human samples and long-term exposure studies remains lacking in the context of OA, and the role of nanoplastics in OA onset and progression requires further validation.
Micro- and Nanoplastics and Lupus-Related Musculoskeletal Involvement
Systemic lupus erythematosus (SLE) is a multisystem autoimmune disease characterized by the presence of autoantibodies targeting nuclear antigens and immune complex deposition. The organs most severely affected by SLE are the kidneys, nervous system, joints, and skin, with persistent inflammation potentially leading to irreversible organ damage. Although SLE can occur at any age, it predominantly affects women of reproductive age. Most patients may develop arthralgia, myalgia, synovitis, or other musculoskeletal manifestations during the disease course. Although SLE is a systemic autoimmune disease, musculoskeletal manifestations such as arthralgia, myalgia, and synovitis are common; therefore, lupus-related osteoarticular involvement may be discussed as extended evidence for the effects of micro- and nanoplastics on the musculoskeletal system.141,142
The pathogenesis of SLE is complex, involving interactions between genetic, environmental, hormonal, and immunological factors, including both innate and adaptive immune dysregulation. A hallmark feature of SLE is the production of autoantibodies forming immune complexes that precipitate in vasculature, driving organ injury. Biological mechanisms involve self-antigens triggering T cell dysfunction, subsequently causing B cell hyperactivation and excessive antibody production.143,144 Given that SLE-associated synovitis and joint involvement are closely related to immune inflammation, pyroptosis, and oxidative stress, the potential effects of micro- and nanoplastics on SLE-related musculoskeletal injury warrant attention.
Chronic microplastic exposure has been shown to significantly exacerbate synovial inflammation in SLE mouse models, evidenced by increased inflammatory cell infiltration, enhanced angiogenesis, and elevated TUNEL-positive cells. These pathological changes correlate with microplastic dosage, with apoptotic rates increasing by 5% after low-dose exposure and 10% after high-dose exposure.118 Mechanistically, microplastics can upregulate the expression and phosphorylation of NF-κB signaling pathway-related proteins in a dose-dependent manner and promote the expression of pyroptosis-related molecules, including NLRP3, CASPASE-1, GSDMD, IL-1β, and IL-18, thereby amplifying synovial inflammation and pyroptotic responses. Dysregulation of the NRF2/KEAP1 antioxidant pathway may also be involved in this process.119
Additionally, microplastic exposure upregulates NF-κB signaling pathway-related protein expression and phosphorylation in a dose-dependent manner. Microplastic may amplify synovial inflammation and pyroptosis by modulating NF-κB pathways, thereby worsening structural and functional synovial damage. Prolonged high-dose microplastic exposure further aggravates renal structural injury in mice, induces lupus-like symptoms, and potentiates pre-existing lupus manifestations. These include abnormal elevations in splenic double-negative T cells, plasma cells, serum autoantibodies, and pro-inflammatory cytokines.119
However, these findings are mainly derived from animal models and do not directly demonstrate that microplastics are an inducing factor for human SLE. Current evidence more strongly supports the possibility that microplastics may exacerbate lupus-related inflammatory responses and musculoskeletal involvement, whereas their role in the onset of human SLE requires further validation.
Micro- and Nanoplastics and Osteosarcoma Progression: Emerging Evidence
Beyond degenerative and inflammatory osteoarticular diseases, the relationship between microplastics and bone tumor progression has also begun to receive attention. Osteosarcoma is one of the most common primary malignant bone tumors and is characterized by high aggressiveness, a strong tendency for metastasis, and a high risk of recurrence. Whether environmental pollutants contribute to remodeling of the bone tumor microenvironment and malignant progression has emerged as an important area of investigation.
A recent study detected microplastics in human osteosarcoma tissues and further showed that PE-MPs can be internalized by osteosarcoma cells, promoting malignant cellular phenotypes and tumor growth. Mechanistic analyses suggested that PE-MPs can upregulate ITGA5 and promote the formation of the pro-tumorigenic ITGA5/FAPα/LGMN complex; blockade of this complex partially reversed the microplastic-mediated pro-tumor effects.120 This study provides emerging evidence for a potential link between microplastics and bone tumor progression. However, research in osteosarcoma remains at an early stage, and the available evidence is mainly derived from a single human tissue-detection study and experimental models. Whether microplastics participate in osteosarcoma initiation, progression, or remodeling of the tumor microenvironment requires further validation in independent cohorts, different plastic particle models, and long-term exposure studies.
Current Mechanisms of Micro- and Nanoplastics Harm to the Skeletal System
Research on the effects of micro- and nanoplastics on the skeletal system is still at an early stage. However, existing evidence suggests that their injurious effects are not mediated by a single pathway, but instead involve multiple processes, including particle uptake, activation of inflammatory signaling, oxidative stress, mitochondrial dysfunction, pyroptosis, cellular senescence, ferroptosis, and disruption of osteoblast–osteoclast coupling. Bone, cartilage, synovium, intervertebral discs, and the bone marrow microenvironment differ in cellular composition and metabolic characteristics; therefore, the modes of action of micro- and nanoplastics may also vary across tissues. Overall, inflammatory responses and oxidative stress are currently the two shared mechanisms with the strongest body of evidence.
Particle Uptake, Tissue Accumulation, and Disruption of the Skeletal Microenvironment
The ability of micro- and nanoplastics to reach skeletal tissues and interact with local cells is a prerequisite for their effects on the skeletal system. Recent studies of human samples have detected microplastics in bone, cartilage, intervertebral discs, and bone marrow, providing direct evidence of skeletal system exposure.21,23 At the cellular level, micro- and nanoplastics can be taken up by osteoblast-like cells, bone marrow stromal cells, synovial fibroblasts, macrophages, and nucleus pulposus cells, potentially affecting lysosomal function, mitochondrial homeostasis, and cellular stress responses. Particle uptake and the resulting disturbance of the local microenvironment represent a common starting point for subsequent mechanisms such as inflammatory responses, oxidative stress, pyroptosis, cellular senescence, and matrix degradation.
Activation of TLR4/NF-κB- and MAPK-Mediated Inflammatory Signaling
Activation of inflammatory signaling is one of the key mechanisms by which micro- and nanoplastics induce skeletal system injury. After entering local tissues, micro- and nanoplastics may activate pattern-recognition receptors through particle stimulation, surface-adsorbed contaminants, cellular stress responses, or the release of damage-associated molecules, among which the TLR4/NF-κB pathway has received considerable attention.145–148 In the canonical inflammatory response, lipopolysaccharide (LPS) is a typical ligand of TLR4 and can activate IRAK, TRAF6, and the IKK complex through the TLR4/MyD88 axis, promoting IκBα degradation and nuclear translocation of NF-κB p65, ultimately inducing the expression of IL-1β, IL-6, TNF-α, COX-2, and various chemokines. Unlike LPS, whether micro- and nanoplastics directly act as TLR4 ligands remains unclear. They are more likely to indirectly amplify TLR4/NF-κB-mediated inflammatory cascades through particle surface properties, adsorbed endotoxins or organic pollutants, oxidative injury, and the release of cellular danger signals.
In skeletal system-related studies, the TLR4/NF-κB pathway is closely associated with impaired osteogenesis, enhanced osteoclastogenesis, synovial inflammation, and extracellular matrix degradation. In osteoporosis models, PS-MPs can activate NF-κB signaling, promote BMSC senescence, and enhance RANKL-related osteoclastogenesis, thereby disrupting the dynamic balance between bone formation and bone resorption.110 In studies of rheumatoid arthritis, PS-MPs have been detected in the synovial fluid of patients with RA, and 5 μm PS-MPs can be taken up by RA-FLS, activate the NF-κB/MAPK signaling pathways, promote the expression of inflammatory and matrix-degradation-related molecules such as IL-6, IL-8, CCL2, MMP3, and MMP9, and aggravate joint inflammation and cartilage erosion in animal models.112 These findings suggest that the NF-κB/MAPK pathways may serve as important signaling nodes through which microplastics promote synovial inflammation, cartilage destruction, and osteoarticular structural damage.
Similar inflammatory pathways may also participate in lupus-related synovitis and other forms of musculoskeletal injury. Studies using SLE models have shown that microplastic exposure can increase the expression and phosphorylation of NF-κB-related proteins in synovial tissue, accompanied by inflammatory cell infiltration, synovial injury, and upregulation of pyroptosis-related molecules.118,119 Collectively, these findings suggest that micro- and nanoplastic-induced inflammatory responses are not confined to a single skeletal disease, but may represent a shared pathological basis across osteoporosis, inflammatory arthropathies, and degenerative diseases. However, most current studies remain limited to animal models and in vitro cellular systems. Whether micro- and nanoplastics drive inflammatory responses in the human skeletal system through a specific TLR4-recognition mechanism requires further validation.
Oxidative Stress, NOX2/ROS, and NRF2/KEAP1 Antioxidant Imbalance
Oxidative stress is one of the more consistent mechanistic signals implicated in micro- and nanoplastic-induced skeletal system injury. After entering cells, micro- and nanoplastics can promote the generation of reactive oxygen species (ROS) through mitochondrial dysfunction, activation of NADPH oxidases, recruitment of inflammatory cells, and contaminants adsorbed onto particle surfaces. Excessive ROS can cause oxidative damage to lipids, proteins, and DNA, while further amplifying inflammatory responses, cellular senescence, and cell death. In the skeletal system, oxidative stress can inhibit osteogenic differentiation of osteoblasts and BMSCs, promote osteoclast activation, and accelerate degeneration of chondrocytes and nucleus pulposus cells.
In osteoporosis-related studies, PET or PVC microplastics were shown to increase ROS levels in MC3T3 cells, accompanied by elevated expression of inflammatory cytokines such as IL-1β, TNF-α, and IL-6.109 In studies of intervertebral disc degeneration, PE-MPs enhanced ROS production through the TLR4/NOX2 signaling axis and induced nucleus pulposus cell senescence and degenerative phenotypes.113 These findings suggest that NOX2-mediated oxidative stress may be an important mechanism by which microplastics impair nucleus pulposus cell function and disrupt intervertebral disc homeostasis.
The NRF2/KEAP1 pathway is an important protective mechanism by which cells respond to oxidative stress. Under physiological conditions, KEAP1 binds to NRF2 and promotes its degradation. When oxidative stress increases, NRF2 is released from the KEAP1 complex and translocates into the nucleus, where it induces the expression of antioxidant genes such as HO-1, NQO1, and GCLC. If micro- and nanoplastic exposure persistently increases the ROS burden while the NRF2/KEAP1-mediated antioxidant response remains insufficient, oxidative damage may accumulate. In models of SLE-related synovitis, microplastic exposure was associated with NRF2/KEAP1 pathway abnormalities, synovial oxidative stress, and aggravated inflammatory injury.119 Future studies could further incorporate indicators such as 8-hydroxy-2′-deoxyguanosine (8-OHdG), lipid peroxidation products, mitochondrial membrane potential, and antioxidant enzyme activity to determine whether micro- and nanoplastics contribute to skeletal system pathology through oxidative DNA damage and mitochondrial dysfunction.
NLRP3 Inflammasome Activation and Pyroptosis
Pyroptosis is a form of inflammatory programmed cell death that is typically associated with NLRP3 inflammasome activation, caspase-1 activation, GSDMD cleavage, and the release of IL-1β and IL-18. After entering cells, micro- and nanoplastics may trigger the NLRP3 inflammasome through lysosomal damage, increased mitochondrial ROS production, and disruption of ionic homeostasis. Compared with apoptosis alone, pyroptosis is accompanied by inflammatory cytokine release and cell membrane rupture, making it more likely to amplify local inflammatory responses.
In synovial and cartilage tissues, pyroptosis may promote synovial inflammation, cartilage matrix degradation, and joint structural damage. Studies of SLE-related synovitis have shown that microplastic exposure can upregulate pyroptosis-related molecules, including NLRP3, CASPASE-1, GSDMD, IL-1β, and IL-18, and aggravate synovial inflammation and tissue injury.119 In the context of osteoarthritis, a recent study suggested that 20 nm PS-NPs can accelerate OA progression and cartilage matrix loss through lysosomal destabilization, Cathepsin B release, NLRP3 inflammasome activation, and GSDMD-mediated chondrocyte pyroptosis.115
These studies indicate that the NLRP3 inflammasome and pyroptosis may represent important mechanisms by which micro- and nanoplastics exacerbate joint inflammation and cartilage injury. However, current evidence is mainly derived from animal models and cellular experiments, and their actual contribution to human osteoarticular diseases requires further validation using patient tissue samples and long-term exposure studies.
Cellular Senescence, SASP, and Disruption of Osteoblast–Osteoclast Coupling
Cellular senescence is an important pathological process in skeletal degeneration and age-related bone diseases. Senescent cells are typically characterized by increased expression of markers such as p16, p21, and SA-β-gal, and can secrete a range of inflammatory cytokines, chemokines, and matrix-degrading enzymes, collectively forming the senescence-associated secretory phenotype (SASP). SASP can persistently alter the local microenvironment and promote chronic inflammation, osteoclast activation, cartilage matrix degradation, and tissue degeneration.
In osteoporosis models, long-term low-dose exposure to PS-MPs can induce BMSC senescence, reduce the expression of osteogenic markers such as Osterix and RUNX2, and enhance RANKL-related osteoclastogenesis.110 These findings suggest that microplastics may alter the balance of bone remodeling by affecting BMSC fate, shifting the bone marrow microenvironment from an osteogenesis-supportive state toward a pro-inflammatory, pro-osteoclastogenic, and low-osteogenic state. In osteoporosis, such changes may lead to reduced bone formation and increased bone resorption. In joint and intervertebral disc tissues, senescence-associated inflammatory mediators and matrix-degrading enzymes may further promote cartilage degeneration and ECM destruction.
Micro- and nanoplastic-induced oxidative stress and inflammatory responses may also feedback to promote cellular senescence, forming a cycle of “increased ROS–amplified inflammation–cellular senescence–disrupted bone homeostasis”. In studies of intervertebral disc degeneration, PE-MPs induced nucleus pulposus cell senescence through the TLR4/NOX2/ROS axis, providing further support for this mechanism.113 Future studies should clarify differences in senescence responses to micro- and nanoplastic exposure among different skeletal cell types, particularly the specific roles of osteoblasts, osteocytes, BMSCs, chondrocytes, synovial cells, and nucleus pulposus cells in SASP formation and disease progression.
Ferroptosis, Exosome-Mediated Transport, and Extracellular Matrix Degradation
In addition to inflammation, oxidative stress, and pyroptosis, ferroptosis may also contribute to microplastic-mediated skeletal system injury. Ferroptosis is an iron-dependent form of cell death associated with lipid peroxidation and is commonly linked to abnormal glutathione metabolism, reduced GPX4 activity, impaired SLC7A11 function, and lipid ROS accumulation. In tissues that are rich in extracellular matrix and have limited regenerative capacity, such as cartilage and intervertebral discs, ferroptosis and lipid peroxidation may further compromise cell survival and promote disruption of ECM homeostasis.
Studies of intervertebral disc degeneration have shown that PE-MPs can be transported to nucleus pulposus cells via macrophage-derived exosomes and induce lipid peroxidation, ferroptosis, cellular senescence, and ECM degradation through NAT10/SLC7A11- and ac4C modification-related mechanisms.114 This finding suggests that microplastics do not necessarily exert toxicity only through direct contact with target cells, but may also indirectly affect skeletal system cells through macrophages, exosomes, and the local inflammatory microenvironment.
ECM degradation is a shared pathological endpoint in diseases such as RA, OA, and IVDD. In RA studies, PS-MPs have been shown to promote the expression of matrix-degradation-related molecules, including MMP3 and MMP9.112 Studies of OA and IVDD also suggest that micro- and nanoplastic exposure may be accompanied by cartilage or nucleus pulposus ECM destruction. Taken together, ferroptosis, exosome-mediated transport, and ECM degradation may jointly contribute to the progression of micro- and nanoplastic-associated degenerative musculoskeletal diseases, although these mechanisms still require validation in further independent studies.
Epigenetic Regulation and Omics-Based Research Directions
At present, the epigenetic mechanisms by which micro- and nanoplastics affect the skeletal system remain insufficiently investigated, but this area has considerable exploratory value. Chronic inflammation, oxidative stress, and metabolic abnormalities may alter DNA methylation, histone modifications, non-coding RNA expression, and chromatin accessibility, thereby affecting osteogenic differentiation, osteoclastogenesis, chondrocyte homeostasis, and nucleus pulposus cell degeneration. Bone homeostasis itself is also influenced by epigenetic regulation, with mechanisms such as DNA methylation, histone modifications, and microRNAs participating in the regulation of bone remodeling, osteogenic differentiation, and osteoclast function.149 Inflammation-related miRNAs, such as miR-146a, miR-155, and miR-21, may be involved in feedback regulation of NF-κB signaling, macrophage polarization, and osteoimmune responses. However, direct evidence demonstrating a decisive role of these epigenetic factors in micro- and nanoplastic-associated skeletal toxicity is still lacking.
Multi-omics technologies may provide new tools for this field. Single-cell RNA sequencing can help characterize the transcriptional responses of different skeletal cell populations after micro- and nanoplastic exposure and reveal changes in cellular heterogeneity, cell-fate transitions, and intercellular communication; this technology has already been applied in studies of orthopedic diseases such as osteoarthritis, rheumatoid arthritis, intervertebral disc degeneration, and osteosarcoma.150 Spatial transcriptomics can further preserve tissue spatial information and may be used to examine changes in inflammatory, metabolic, and extracellular matrix microenvironments surrounding regions of particle accumulation; in recent years, its application in musculoskeletal research has also received increasing attention.151 Metabolomics and lipidomics can be used to identify pathways related to oxidative stress, ferroptosis, and energy metabolic abnormalities. In the future, integrating particle detection techniques, animal disease models, patient tissue samples, and multi-omics analyses may help clarify the cell-type-specific mechanisms by which micro- and nanoplastics contribute to osteoporosis, arthritis, intervertebral disc degeneration, and bone tumors. All mechanistic schematics are presented in Figure 4.
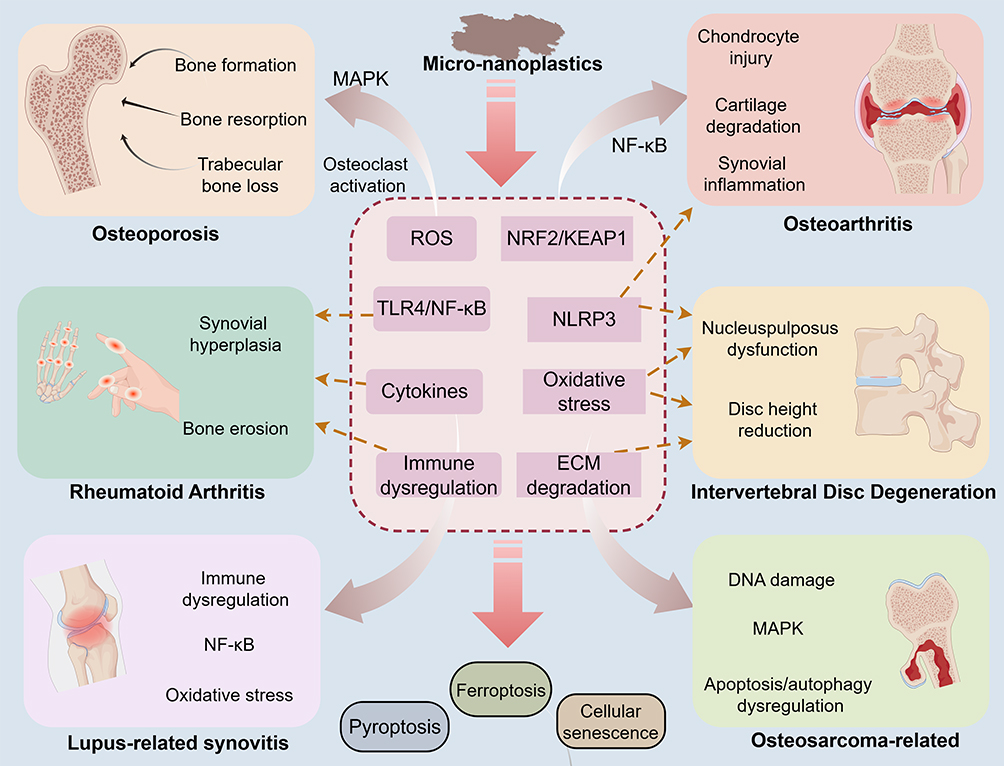 |
Figure 4 Micro-nanoplastics affect skeletal system health through multiple pathways. |
Future Perspectives and Clinical Translation
Current evidence suggests that micro- and nanoplastics may affect skeletal homeostasis through inflammatory responses, oxidative stress, immune dysregulation, cellular senescence, and disruption of osteoblast–osteoclast coupling. However, their long-term effects, dose–response relationships, and causal associations in human populations remain to be clarified. Future research should move beyond descriptive toxicological observations and gradually shift toward exposure assessment, disease risk prediction, clinical monitoring, and public health intervention. For the skeletal system, it will be particularly important to integrate biomonitoring of micro- and nanoplastics with clinical outcomes such as bone mineral density, bone turnover markers, indicators of joint inflammation, imaging changes, and fracture risk, in order to determine whether these pollutants have genuine value for disease prediction and intervention.21,152
Environmental Source Control and Public Health Prevention
Reducing the generation and release of micro- and nanoplastics into the environment remains fundamental to lowering the risk of long-term human exposure. Over recent decades, the use of plastic products has continued to increase, whereas the recycling rate of plastic waste remains limited. Large quantities of plastics can enter the natural environment and gradually form microplastics under the effects of light exposure, thermo-oxidation, hydraulic erosion, and mechanical abrasion.153 Therefore, future efforts should strengthen whole-chain management of plastic production, use, recycling, and waste disposal, including limiting unnecessary single-use plastics, improving waste sorting and recycling systems, reducing the direct discharge of plastic waste, and promoting the development and application of biodegradable or environmentally friendly alternative materials.
From a public health perspective, assessment of skeletal health risks associated with micro- and nanoplastics should be incorporated into a more systematic exposure-monitoring framework. Future studies could integrate regional levels of environmental contamination, micro- and nanoplastic concentrations in drinking water and food, population exposure levels, and skeletal health indicators to evaluate the potential associations of long-term exposure with reduced bone mineral density, abnormal bone turnover, chronic joint inflammation, and fracture risk. As evidence accumulates, monitoring standards for micro- and nanoplastics in food, drinking water, and key occupational settings may be gradually established to inform public health policy and risk management.
Interventions Targeting Food, Drinking Water, and Daily Exposure
Food and drinking water are important sources of human exposure to micro- and nanoplastics. Plastic packaging materials, bottled water, takeaway containers, tea bags, plastic kettles, and infant feeding bottles may release plastic particles during production, storage, or high-temperature use.154,155 Therefore, reducing high-temperature contact between food and plastic materials has practical relevance. In daily life, glass, stainless steel, or ceramic containers may be preferred for food storage and heating, while plastic containers should be avoided for hot foods or microwave heating. For potentially high-risk groups, such as infants, pregnant women, patients with osteoporosis, and patients with rheumatoid arthritis, reducing micro- and nanoplastic exposure from food packaging and drinking water may have preventive value.
Drinking-water filtration technology is another important direction for reducing exposure risk. For micro- and nanoplastic particles that may be present in tap water, bottled water, and household drinking water, future research should further develop efficient, low-cost filtration and adsorption systems suitable for daily use.156 These technologies should focus on removal efficiency across different particle-size ranges, polymer types, and aged plastic particles, and should establish detection and evaluation standards that reflect real drinking-water scenarios. It should be noted that, at this stage, no single filtration method can be assumed to fully eliminate the risk of micro- and nanoplastic exposure. Source pollution control, optimization of water-treatment processes, and end-point monitoring should therefore be advanced together.
Clinical Translation and Monitoring of High-Risk Populations
As micro- and nanoplastics have been detected in human blood, bone marrow, bone, cartilage, intervertebral discs, and other samples, clinical translational research will become an important direction in this field. Existing reviews have noted that human biomonitoring data for microplastics remain limited, with substantial heterogeneity across studies in sample types, detection methods, and analyses linking exposure to health outcomes; a more standardized exposure-assessment framework is therefore urgently needed.157 Future research may prioritize prospective cohort studies that combine biomonitoring of micro- and nanoplastics with skeletal health assessment. For example, micro- and nanoplastic burdens in blood, urine, feces, synovial fluid, or bone marrow samples could be measured across populations differing in age, occupation, and disease risk, and then evaluated together with bone mineral density, bone turnover markers, inflammatory cytokines, joint imaging findings, severity of intervertebral disc degeneration, fracture incidence, and joint function scores to assess the relationship between long-term exposure and the onset or progression of skeletal diseases.
For workers in plastic production, textile manufacturing, packaging, waste treatment, and tire- and rubber-related industries, occupational exposure to micro- and nanoplastics and its potential musculoskeletal health effects should be considered. A systematic review of airborne micro- and nanoplastics showed that both indoor and outdoor air can serve as sources of inhalation exposure, with exposure doses varying across age groups and settings, indicating the practical importance of occupational exposure monitoring and protective strategies.158 Future efforts could establish occupational exposure-monitoring systems that regularly assess airborne particulate levels in workplaces, use of personal protective equipment, blood or urinary exposure biomarkers, and changes in bone mineral density and inflammatory indicators. For patients with established osteoporosis, rheumatoid arthritis, osteoarthritis, or intervertebral disc degeneration, there is currently no evidence to support micro- and nanoplastic testing as a routine clinical screening item. Nevertheless, reducing contact with high-temperature plastic food packaging, drinking effectively treated water, strengthening occupational protection, and lowering exposure to environmental particulates remain feasible recommendations for risk management.
Clinical translation also requires stable and reliable detection workflows. Substantial differences remain across studies in sample collection, tissue digestion, contamination control, particle identification, and quantification methods, limiting the comparability of results. Recent reviews on the detection of micro- and nanoplastics in biological samples have also emphasized that standardization of detection workflows and cross-platform validation are essential prerequisites for future human studies.159 Future work should prioritize standardized detection procedures for blood, synovial fluid, bone marrow, bone, cartilage, intervertebral disc, and other samples, including the use of non-plastic consumables, procedural blanks, negative controls, spiked recovery experiments, spectral-library matching thresholds, and multi-platform cross-validation. Only when detection methods are sufficiently robust can biomonitoring of micro- and nanoplastics be further applied to disease risk assessment and clinical research.
Emerging Intervention Technologies and Potential Therapeutic Directions
Technologies for clearing micro- and nanoplastics from the body remain at an early exploratory stage. Environmental water-treatment studies have attempted to use magnetic nanomaterials to adsorb and separate microplastics from water. For example, nano-Fe3O4 can magnetize various microplastics through surface adsorption and enable their separation under a magnetic field, suggesting that magnetically responsive materials have potential applications in environmental microplastic removal.160 However, such strategies are mainly used for water purification and cannot be directly extrapolated to in vivo clearance in humans.
Research on micro- and nanorobots for microplastic capture and degradation has also mainly focused on environmental water purification. For example, magnetic/enzyme microrobots can capture and degrade target microplastics under an external magnetic field, demonstrating experimental feasibility.161 However, these studies remain at the in vitro or environmental-application stage and are still far from achieving safe, controllable, and effective clearance of micro- and nanoplastics in the human body. For these strategies to be translated into biomedical applications, key issues such as tissue targeting, material biosafety, immune responses, metabolic clearance pathways, long-term toxicity, and secondary damage to normal tissues must be addressed.
By contrast, the more practical intervention directions at present remain reducing external exposure, improving environmental governance, strengthening monitoring of drinking-water and food safety, and conducting long-term follow-up of high-risk populations. For skeletal system diseases, future studies may also explore whether antioxidant, anti-inflammatory, gut microbiota-modulating, bone metabolism-improving, and abnormal osteoclast activation-inhibiting strategies can mitigate micro- and nanoplastic-associated skeletal injury. However, these interventions currently lack direct clinical evidence in the context of human micro- and nanoplastic exposure, and their efficacy and safety will need to be validated gradually through animal experiments, organoid models, and population-based studies.
Overall, prevention and control of micro- and nanoplastic-related skeletal health risks require coordinated efforts across environmental governance, exposure reduction, standardized detection, occupational monitoring, and clinical research. As evidence from human skeletal system samples and mechanistic studies continues to increase, this field is expected to move beyond descriptive toxicological observations toward risk assessment, disease prevention, and clinical translational research. To support future studies in this field, commonly used analytical methods for detecting MNPs and their applicability to skeletal system samples are summarized in Table 2.
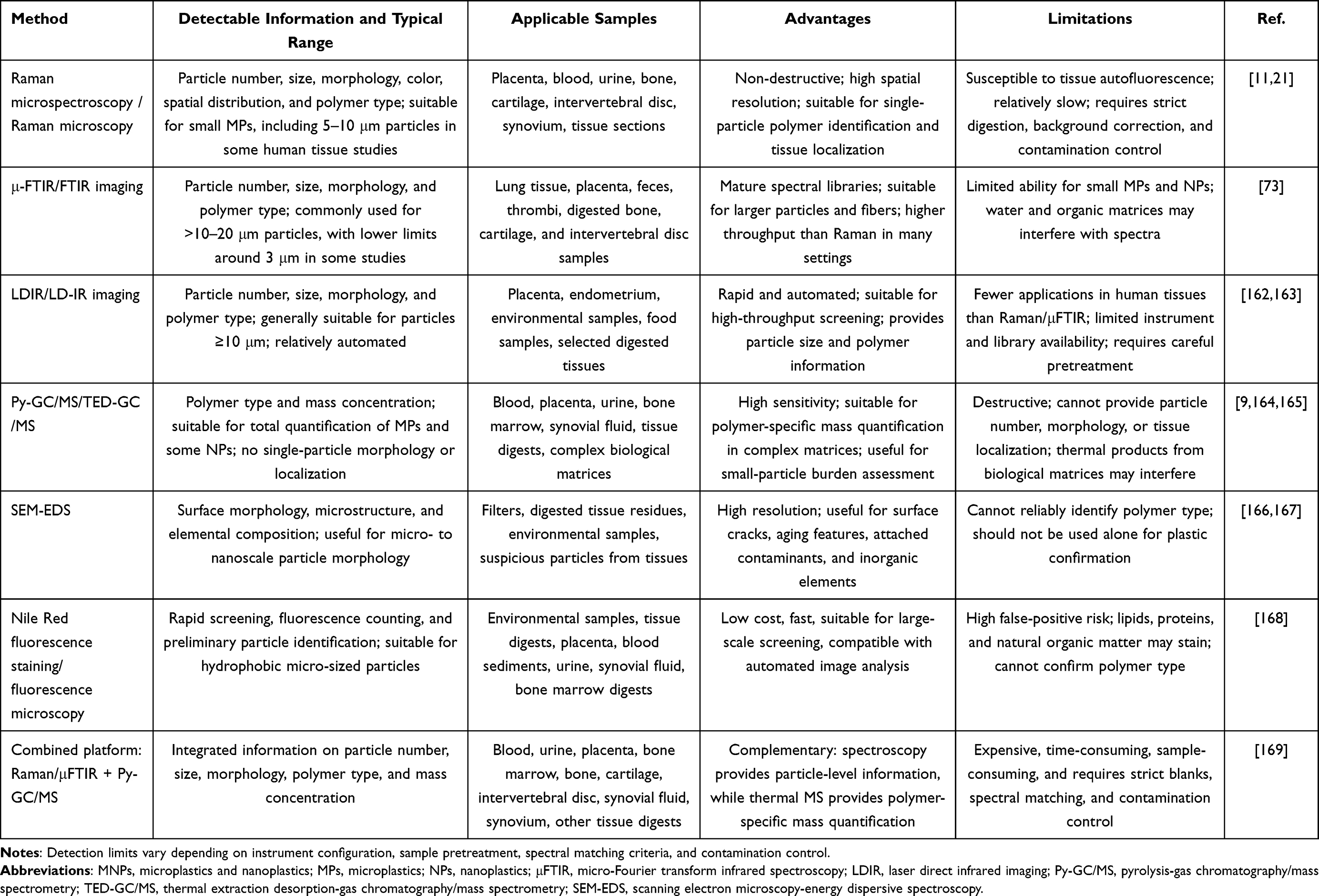 |
Table 2 Analytical Methods for Detecting Microplastics and Nanoplastics in Biological and Skeletal Tissues |
Research Limitations
Although existing studies suggest that micro- and nanoplastics may affect skeletal homeostasis and may be potentially associated with the progression of various musculoskeletal diseases, this field remains at an early stage. Current evidence still has several limitations regarding human causal inference, exposure models, experimental particle characteristics, detection methods, and clinical extrapolation.
In terms of human evidence, available studies remain limited. Microplastics have been detected in human bone, cartilage, intervertebral discs, and bone marrow, providing direct evidence of skeletal system exposure.21,23 Human sample-based or experimental model evidence has also emerged in areas such as osteoporosis, rheumatoid arthritis, and intervertebral disc degeneration. However, most human studies remain cross-sectional observations, tissue-detection studies, or small-sample analyses, making it difficult to determine exposure timing, dose–response relationships, and the temporal sequence between exposure and disease onset. Therefore, current findings are more appropriately interpreted as indicating potential associations and are not sufficient to establish a direct causal relationship between micro- and nanoplastic exposure and human skeletal diseases.
In terms of experimental models, animal and in vitro studies provide important mechanistic clues, but caution is needed when extrapolating these findings to humans. Current studies largely rely on mouse, zebrafish, Caenorhabditis elegans, or cellular models. These models are useful for elucidating mechanisms such as inflammatory responses, oxidative stress, cellular senescence, pyroptosis, and disrupted osteoblast–osteoclast coupling, but their exposure routes, metabolic characteristics, and disease courses differ from those in humans. Some in vitro studies have used micro- and nanoplastic concentrations higher than realistic environmental exposure levels. For example, in RA-FLS-related experiments, the concentration range of microplastics reached 0–1.0 mg/mL. Such conditions are more suitable for observing mechanistic changes such as cellular uptake, inflammatory activation, migration, and invasion, rather than for directly inferring actual exposure risks in the general population.111
From the perspective of real-world exposure, experimental particles also differ from environmental particles. Most studies use commercially available plastic microspheres with uniform particle sizes, regular morphology, and single polymer composition, such as PS-MPs or PS-NPs. By contrast, micro- and nanoplastics in the real environment often undergo aging, oxidation, ultraviolet irradiation, and mechanical abrasion, resulting in more complex morphology and surface properties. They may also adsorb exogenous substances such as heavy metals, persistent organic pollutants, pathogens, or endotoxins. Plastic additives, including phthalates, bisphenol A, flame retardants, and plasticizers, may also act together with plastic particles, resulting in mixed exposure and confounding effects.
Methodologically, detection workflows for micro- and nanoplastics in skeletal system samples have not yet been fully standardized. Samples such as bone, cartilage, intervertebral discs, synovial fluid, and bone marrow have complex matrices, and exogenous contamination or matrix interference may occur during sample collection, digestion, filtration, and detection. Raman spectroscopy, μFTIR, LDIR, Py-GC/MS, SEM-EDS, and fluorescence staining each have advantages and limitations, and a single method usually cannot simultaneously provide information on particle number, particle size, morphology, polymer type, spatial distribution, and mass concentration.169,170
Future studies should establish more unified quality-control procedures, including the use of non-plastic consumables, procedural blanks and negative controls, spiked recovery experiments, clearly defined spectral matching thresholds, blinded re-evaluation, and interlaboratory validation. Long-term prospective studies are still needed in potentially high-risk populations, such as older adults, postmenopausal women, patients with osteoporosis, patients with rheumatoid arthritis, and individuals with long-term occupational exposure. These studies should integrate measurements of human micro- and nanoplastic burden with bone mineral density, bone turnover markers, inflammatory indicators, imaging changes, and clinical outcomes to clarify their true significance in the onset and progression of skeletal diseases.
Overall, current evidence suggests that micro- and nanoplastics may contribute to disrupted skeletal homeostasis and the progression of musculoskeletal diseases. However, insufficient causal evidence in humans, limited extrapolation from experimental models, the complexity of real-world exposure scenarios, and the lack of standardized detection methods continue to constrain the strength of existing conclusions. Future research should gradually move from single toxicological experiments toward an integrated research framework that combines standardized detection, realistic exposure modeling, multi-omics mechanistic analysis, and long-term population follow-up. The potential future strategies for micro- and nanoplastic research and risk management are summarized in Figure 5.
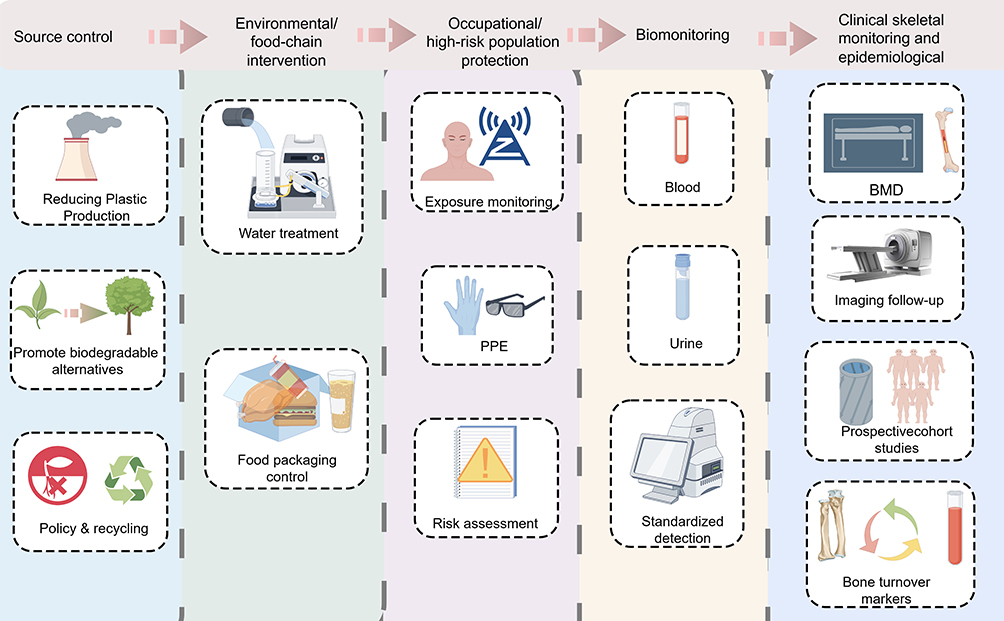 |
Figure 5 Future applications and research perspectives of micro-nanoplastics. |
Conclusions
Micro- and nanoplastics are widespread emerging environmental pollutants that can enter the human body through oral ingestion, inhalation, and potential dermal contact, and they have been detected in multiple tissues. With emerging evidence of microplastics in human bone, cartilage, intervertebral discs, and bone marrow, the skeletal system has become a new area of interest in research on the health effects of micro- and nanoplastics. Current studies suggest that micro- and nanoplastics may disrupt skeletal homeostasis through mechanisms involving inflammatory responses, oxidative stress, mitochondrial dysfunction, cellular senescence, pyroptosis, ferroptosis, and disruption of osteoblast–osteoclast coupling, and may be potentially associated with diseases or pathological conditions such as osteoporosis, rheumatoid arthritis, intervertebral disc degeneration, osteoarthritis, lupus-related musculoskeletal involvement, and osteosarcoma progression.
In terms of strength of evidence, osteoporosis, rheumatoid arthritis, and intervertebral disc degeneration are currently supported by relatively more evidence from human samples, animal models, and cellular mechanistic studies. By contrast, osteoarthritis, lupus-related synovitis, and osteosarcoma progression remain emerging research directions, and their related conclusions require further validation in independent studies. It should be emphasized that most current evidence still derives from animal experiments, in vitro studies, or cross-sectional human observations and is not yet sufficient to establish a direct causal relationship between micro- and nanoplastic exposure and human skeletal diseases.
In the future, this field should build on standardized detection methods and integrate realistic exposure modeling, long-term prospective cohorts, multi-omics analyses, monitoring of bone mineral density and bone turnover markers, and imaging assessment of joints and the spine to further clarify the role of micro- and nanoplastics in the onset and progression of skeletal system diseases. At the same time, reducing environmental plastic pollution, lowering exposure from food and drinking water, strengthening occupational protection, and monitoring high-risk populations remain more practical directions for prevention and control at this stage. As direct exposure evidence and mechanistic studies continue to accumulate, assessment of micro- and nanoplastic-related skeletal health risks is expected to gradually expand from basic toxicological research to clinical translation and public health practice.
Abbreviations
MPs, Microplastics; NPs, Nanoplastics; MNPs, Microplastics and nanoplastics; PS, Polystyrene; PE, Polyethylene; PVC, Polyvinyl chloride; PET, Polyethylene terephthalate; OP, Osteoporosis; RA, Rheumatoid arthritis; IDD, Intervertebral disc degeneration; SLE, Systemic lupus erythematosus; BMSCs, Bone marrow mesenchymal stem/stromal cells; FLS, Fibroblast-like synoviocytes; NPCs, Nucleus pulposus cells; ROS, Reactive oxygen species; NF-κB, Nuclear factor-kappa B; TLR4, Toll-like receptor 4; NOX2, NADPH oxidase 2; NRF2, Nuclear factor erythroid 2-related factor 2; KEAP1, Kelch-like ECH-associated protein 1; SASP, Senescence-associated secretory phenotype; 8-OHdG, 8-hydroxy-2′-deoxyguanosine.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Xi’an Municipal Health Commission Science and Technology Project (2025ms11). The Fundamental Research Funds of Xi’an Jiaotong University (xzy012025150).
Disclosure
The authors declare no competing interests in this work.
References
1. Thompson RC, Olsen Y, Mitchell RP, et al. Lost at sea: where is all the plastic? Science. 2004;304(5672):838. doi:10.1126/science.1094559
2. Mitrano, D.M., P. Wick, and B. Nowack. Placing nanoplastics in the context of global plastic pollution. Nat Nanotechnol, 2021. 16(5): p. 491–28. doi:10.1038/s41565-021-00888-2
3. Osman AI, Hosny M, Eltaweil AS, et al. Microplastic sources, formation, toxicity and remediation: a review. Environ Chem Lett. 2023;1–41. doi:10.1007/s10311-023-01593-3
4. Geyer R, Jambeck JR, Law KL. Production, use, and fate of all plastics ever made. Sci Adv. 2017;3(7):e1700782. doi:10.1126/sciadv.1700782
5. Huang D, Tao J, Cheng M, et al. Microplastics and nanoplastics in the environment: macroscopic transport and effects on creatures. J Hazard Mater. 2021;407:124399. doi:10.1016/j.jhazmat.2020.124399
6. Bui XT, Vo T-D-H, Nguyen P-T, et al. Microplastics pollution in wastewater: characteristics, occurrence and removal technologies. Environ Technol Innovation. 2020;19:101013. doi:10.1016/j.eti.2020.101013
7. Alimi OS, Farner Budarz J, Hernandez LM, et al. Microplastics and nanoplastics in aquatic environments: aggregation, deposition, and enhanced contaminant transport. Environ Sci Technol. 2018;52(4):1704–1724. doi:10.1021/acs.est.7b05559
8. Moeller JN, Loeder MGJ, Laforsch C. Finding microplastics in soils: a review of analytical methods. Environ Sci Technol. 2020;54(4):2078–2090. doi:10.1021/acs.est.9b04618
9. Leslie HA, van Velzen MJM, Brandsma SH, et al. Discovery and quantification of plastic particle pollution in human blood. Environ Int. 2022;163:107199. doi:10.1016/j.envint.2022.107199
10. Lamoree MH, van Boxel J, Nardella F, et al. Health impacts of microplastic and nanoplastic exposure. Nat Med. 2025;31(9):2873–2887. doi:10.1038/s41591-025-03902-5
11. Ragusa A, Svelato A, Santacroce C, et al. Plasticenta: first evidence of microplastics in human placenta. Environ Int. 2021;146:106274. doi:10.1016/j.envint.2020.106274
12. Luo Q, Tan H, Ye M, et al. Microplastics as an emerging threat to human health: an overview of potential health impacts. J Environ Manage. 2025;387:125915. doi:10.1016/j.jenvman.2025.125915
13. Junaid M, Liu S, Chen G, et al. Transgenerational impacts of micro(nano)plastics in the aquatic and terrestrial environment. J Hazard Mater. 2023;443(Pt B):130274. doi:10.1016/j.jhazmat.2022.130274
14. Murphy AC, Muldoon SF, Baker D, et al. Structure, function, and control of the human musculoskeletal network. PLoS Biol. 2018;16(1):e2002811. doi:10.1371/journal.pbio.2002811
15. Nowak A, Ogurkowska M. Bone health and physical activity - the complex mechanism. Aging Dis. 2024;16(6):3400–3420. doi:10.14336/AD.2024.1316
16. Chen S, He T, Zhong Y, et al. Roles of focal adhesion proteins in skeleton and diseases. Acta Pharm Sin B. 2023;13(3):998–1013. doi:10.1016/j.apsb.2022.09.020
17. Li HZ, Zhang J-L, Yuan D-L, et al. Role of signaling pathways in age-related orthopedic diseases: focus on the fibroblast growth factor family. Mil Med Res. 2024;11(1):40. doi:10.1186/s40779-024-00544-5
18. Nandy A, Helderman RCM, Thapa S, et al. Enhanced fatty acid oxidation in osteoprogenitor cells provides protection from high-fat diet induced bone dysfunction. J Bone Miner Res. 2025;40(2):283–298. doi:10.1093/jbmr/zjae195
19. Xiao H, Li W, Qin Y, et al. Crosstalk between lipid metabolism and bone homeostasis: exploring intricate signaling relationships. Research. 2024;7:0447. doi:10.34133/research.0447
20. Chen X, Zhang A, Zhao K, et al. The role of oxidative stress in intervertebral disc degeneration: mechanisms and therapeutic implications. Ageing Res Rev. 2024;98:102323. doi:10.1016/j.arr.2024.102323
21. Yang Q, Peng Y, Wu X, et al. Microplastics in human skeletal tissues: presence, distribution and health implications. Environ Int. 2025;196:109316. doi:10.1016/j.envint.2025.109316
22. Zhao H, Mu S, Wang W, et al. Potential threats of environmental microplastics to the skeletal system: current insights and future directions. Front Endocrinol. 2025;16:1658056. doi:10.3389/fendo.2025.1658056
23. Guo X, Wang L, Wang X, et al. Discovery and analysis of microplastics in human bone marrow. J Hazard Mater. 2024;477:135266. doi:10.1016/j.jhazmat.2024.135266
24. Pelepenko LE, de Oliveira MC, Masaro DA, et al. Effects of microplastics on the bones: a comprehensive review. Osteoporos Int. 2025;36(8):1327–1345. doi:10.1007/s00198-025-07580-4
25. Anderson PJ, Warrack S, Langen V, et al. Microplastic contamination in Lake Winnipeg. Canada Environ Pollut. 2017;225:223–231. doi:10.1016/j.envpol.2017.02.072
26. He S, Jia M, Xiang Y, et al. Biofilm on microplastics in aqueous environment: physicochemical properties and environmental implications. J Hazard Mater. 2022;424(Pt B):127286. doi:10.1016/j.jhazmat.2021.127286
27. Ziani K, Ioniță-Mîndrican C-B, Mititelu M, et al. Microplastics: a real global threat for environment and food safety: a state of the art review. Nutrients. 2023;15(3):617. doi:10.3390/nu15030617
28. Winiarska E, Jutel M, Zemelka-Wiacek M. The potential impact of nano- and microplastics on human health: understanding human health risks. Environ Res. 2024;251(Pt 2):118535. doi:10.1016/j.envres.2024.118535
29. da Costa JP, Santos PSM, Duarte AC, et al. (Nano)plastics in the environment - Sources, fates and effects. Sci Total Environ. 2016;566-567:15–26. doi:10.1016/j.scitotenv.2016.05.041
30. Sharma S, Chatterjee S. Microplastic pollution, a threat to marine ecosystem and human health: a short review. Environ Sci Pollut Res Int. 2017;24(27):21530–21547. doi:10.1007/s11356-017-9910-8
31. Jambeck JR, Geyer R, Wilcox C, et al. Marine pollution. Plastic waste inputs from land into the ocean. Science. 2015;347(6223):768–771. doi:10.1126/science.1260352
32. Gigault J, Halle AT, Baudrimont M, et al. Current opinion: what is a nanoplastic? Environ Pollut. 2018;235:1030–1034. doi:10.1016/j.envpol.2018.01.024
33. Hartmann NB, Hüffer T, Thompson RC, et al. Are we speaking the same language? Recommendations for a definition and categorization framework for plastic debris. Environ Sci Technol. 2019;53(3):1039–1047. doi:10.1021/acs.est.8b05297
34. Ali N, Katsouli J, Marczylo EL, et al. The potential impacts of micro-and-nano plastics on various organ systems in humans. EBioMedicine. 2024;99:104901. doi:10.1016/j.ebiom.2023.104901
35. Liang B, Huang Y, Zhong Y, et al. Brain single-nucleus transcriptomics highlights that polystyrene nanoplastics potentially induce Parkinson’s disease-like neurodegeneration by causing energy metabolism disorders in mice. J Hazard Mater. 2022;430:128459. doi:10.1016/j.jhazmat.2022.128459
36. Prüst M, Meijer J, Westerink RHS. The plastic brain: neurotoxicity of micro- and nanoplastics. Part Fibre Toxicol. 2020;17(1):24. doi:10.1186/s12989-020-00358-y
37. Xie J, ji J, Sun Y, et al. Blood-brain barrier damage accelerates the accumulation of micro- and nanoplastics in the human central nervous system. J Hazard Mater. 2024;480:136028. doi:10.1016/j.jhazmat.2024.136028
38. Wang Q, Bai J, Ning B, et al. Effects of bisphenol A and nanoscale and microscale polystyrene plastic exposure on particle uptake and toxicity in human Caco-2 cells. Chemosphere. 2020;254:126788. doi:10.1016/j.chemosphere.2020.126788
39. Way C, Hudson MD, Williams ID, et al. Evidence of underestimation in microplastic research: a meta-analysis of recovery rate studies. Sci Total Environ. 2022;805:150227. doi:10.1016/j.scitotenv.2021.150227
40. Zhang K, Hamidian AH, Tubić A, et al. Understanding plastic degradation and microplastic formation in the environment: a review. Environ Pollut. 2021;274:116554. doi:10.1016/j.envpol.2021.116554
41. van Emmerik T, Kieu-Le TC, Loozen M, et al. A methodology to characterize riverine macroplastic emission into the ocean. Front Mar Sci. 2018;5:372. doi:10.3389/fmars.2018.00372
42. Isobe A, Iwasaki S, Uchida K, et al. Abundance of non-conservative microplastics in the upper ocean from 1957 to 2066. Nat Commun. 2019;10(1):417. doi:10.1038/s41467-019-08316-9
43. Xiang Y, Jiang L, Zhou Y, et al. Microplastics and environmental pollutants: key interaction and toxicology in aquatic and soil environments. J Hazard Mater. 2022;422:126843. doi:10.1016/j.jhazmat.2021.126843
44. Wang Y, Nan X, Sun H, et al. From insects to mammals! Tissue accumulation and transgenerational transfer of micro/nano-plastics through the food chain. J Hazard Mater. 2024;480:136424. doi:10.1016/j.jhazmat.2024.136424
45. Wang W, Do ATN, Kwon JH. Ecotoxicological effects of micro- and nanoplastics on terrestrial food web from plants to human beings. Sci Total Environ. 2022;834:155333. doi:10.1016/j.scitotenv.2022.155333
46. Barceló D, Picó Y, Alfarhan AH. Microplastics: detection in human samples, cell line studies, and health impacts. Environ Toxicol Pharmacol. 2023;101:104204. doi:10.1016/j.etap.2023.104204
47. Li WC. The occurrence, fate, and effects of microplastics in the marine environment. In: Zeng EY, editor. Microplastic Contamination in Aquatic Environments: An Emerging Matter of Environmental Urgency. 2018:133–173. doi:10.1016/b978-0-12-813747-5.00005-9
48. Liu S, Guo J, Liu X, et al. Detection of various microplastics in placentas, meconium, infant feces, breastmilk and infant formula: a pilot prospective study. Sci Total Environ. 2023;854:158699. doi:10.1016/j.scitotenv.2022.158699
49. Vasse GF, Melgert BN. Microplastic and plastic pollution: impact on respiratory disease and health. Eur Respir Rev. 2024;33(172):230226. doi:10.1183/16000617.0226-2023
50. Bargagli R, Rota E. Microplastic interactions and possible combined biological effects in antarctic marine ecosystems. Animals. 2022;13(1):162. doi:10.3390/ani13010162
51. Castro-Castellon AT, Horton AA, Hughes JMR, et al. Ecotoxicity of microplastics to freshwater biota: considering exposure and hazard across trophic levels. Sci Total Environ. 2022;816:151638. doi:10.1016/j.scitotenv.2021.151638
52. Santonicola S, Volgare M, Cocca M, et al. Impact of fibrous microplastic pollution on commercial seafood and consumer health: a review. Animals. 2023;13(11):1736. doi:10.3390/ani13111736
53. Chen Y, Chen J, Guo J, et al. Release of microplastics during dental procedures and denture wear: impact on dental personnel and patients. J Hazard Mater. 2025;494:138463. doi:10.1016/j.jhazmat.2025.138463
54. Karbalaei S, Golieskardi A, Hamzah HB, et al. Abundance and characteristics of microplastics in commercial marine fish from Malaysia. Mar Pollut Bull. 2019;148:5–15. doi:10.1016/j.marpolbul.2019.07.072
55. Long X, Fu T-M, Yang X, et al. Efficient atmospheric transport of microplastics over Asia and adjacent oceans. Environ Sci Technol. 2022;56(10):6243–6252. doi:10.1021/acs.est.1c07825
56. Bothma F, Uren RC, Iordachescu L, et al. Microplastics in the Indian and South Atlantic oceans translocate to gills, digestive glands, and muscle of the chokka squid loligo reynaudii. Mar Pollut Bull. 2024;202:116371. doi:10.1016/j.marpolbul.2024.116371
57. Huang W, Chen M, Song B, et al. Microplastics in the coral reefs and their potential impacts on corals: a mini-review. Sci Total Environ. 2021;762:143112. doi:10.1016/j.scitotenv.2020.143112
58. Vivekanand AC, Mohapatra S, Tyagi VK. Microplastics in aquatic environment: challenges and perspectives. Chemosphere. 2021;282:131151. doi:10.1016/j.chemosphere.2021.131151
59. Cai T, Gu J, Liang Y, et al. Occurrence of microplastics and distinct plastisphere in aquatic environments of metal mining areas in South China. J Hazard Mater. 2025;491:137910. doi:10.1016/j.jhazmat.2025.137910
60. Zhang S, Wu H, Hou J. Progress on the Effects of Microplastics on Aquatic Crustaceans: a Review. Int J Mol Sci. 2023;24(6).doi:10.3390/ijms24065523
61. Koelmans AA, Mohamed Nor NH, Hermsen E, et al. Microplastics in freshwaters and drinking water: critical review and assessment of data quality. Water Res. 2019;155:410–422. doi:10.1016/j.watres.2019.02.054
62. Oßmann BE, Sarau G, Holtmannspötter H, et al. Small-sized microplastics and pigmented particles in bottled mineral water. Water Res. 2018;141:307–316. doi:10.1016/j.watres.2018.05.027
63. Schymanski D, Goldbeck C, Humpf H-U, et al. Analysis of microplastics in water by micro-Raman spectroscopy: release of plastic particles from different packaging into mineral water. Water Res. 2018;129:154–162. doi:10.1016/j.watres.2017.11.011
64. Shi Y, Li D, Xiao L, et al. The influence of drinking water constituents on the level of microplastic release from plastic kettles. J Hazard Mater. 2022;425:127997. doi:10.1016/j.jhazmat.2021.127997
65. Ali T, Habib A, Muskan F, et al. Health risks posed by microplastics in tea bags: microplastic pollution - a truly global problem. Int J Surg. 2023;109(3):515–516. doi:10.1097/JS9.0000000000000055
66. Zhang Q, Liu L, Jiang Y, et al. Microplastics in infant milk powder. Environ Pollut. 2023;323:121225. doi:10.1016/j.envpol.2023.121225
67. Kharaghani D, DeLoid GM, He P, et al. Toxicity and absorption of polystyrene micro-nanoplastics in healthy and Crohn’s disease human duodenum-chip models. J Hazard Mater. 2025;490:137714. doi:10.1016/j.jhazmat.2025.137714
68. Jin T, Liu Y, Lyu H, et al. Plastic takeaway food containers may cause human intestinal damage in routine life usage: microplastics formation and cytotoxic effect. J Hazard Mater. 2024;475:134866. doi:10.1016/j.jhazmat.2024.134866
69. Kaseke T, Lujic T, Velickovic TC. Nano- and microplastics migration from plastic food packaging into dairy products: impact on nutrient digestion, absorption, and metabolism. Foods. 2023;12(16):3043. doi:10.3390/foods12163043
70. Prata JC. Airborne microplastics: consequences to human health? Environ Pollut. 2018;234:115–126. doi:10.1016/j.envpol.2017.11.043
71. Torres-Agullo A, Karanasiou A, Moreno T, et al. Airborne microplastic particle concentrations and characterization in indoor urban microenvironments. Environ Pollut. 2022;308:119707. doi:10.1016/j.envpol.2022.119707
72. Amato-Lourenço LF, Carvalho-Oliveira R, Júnior GR, et al. Presence of airborne microplastics in human lung tissue. J Hazard Mater. 2021;416:126124. doi:10.1016/j.jhazmat.2021.126124
73. Jenner LC, Rotchell JM, Bennett RT, et al. Detection of microplastics in human lung tissue using μFTIR spectroscopy. Sci Total Environ. 2022;831:154907. doi:10.1016/j.scitotenv.2022.154907
74. Fournier SB, D’Errico JN, Adler DS, et al. Nanopolystyrene translocation and fetal deposition after acute lung exposure during late-stage pregnancy. Part Fibre Toxicol. 2020;17(1):55. doi:10.1186/s12989-020-00385-9
75. Baeza-Martínez C, Olmos S, González-Pleiter M, et al. First evidence of microplastics isolated in European citizens’ lower airway. J Hazard Mater. 2022;438:129439. doi:10.1016/j.jhazmat.2022.129439
76. Traversa A, Mari E, Pontecorvi P, et al. Polyethylene micro/nanoplastics exposure induces epithelial-mesenchymal transition in human bronchial and alveolar epithelial cells. Int J Mol Sci. 2024;25(18):10168. doi:10.3390/ijms251810168
77. Michelini S, Mawas S, Kurešepi E, et al. Pulmonary hazards of nanoplastic particles: a study using polystyrene in in vitro models of the alveolar and bronchial epithelium. J Nanobiotechnology. 2025;23(1):388. doi:10.1186/s12951-025-03419-6
78. Lehner R, Weder C, Petri-Fink A, et al. Emergence of nanoplastic in the environment and possible impact on human health. Environ Sci Technol. 2019;53(4):1748–1765. doi:10.1021/acs.est.8b05512
79. Laffon B, Lema M, Méndez J. Simultaneous high-performance liquid chromatographic determination of urinary mandelic and phenylglyoxylic acids as indirect evaluation of styrene exposure. J Chromatogr B Biomed Sci Appl. 2001;753(2):385–393. doi:10.1016/S0378-4347(00)00583-1
80. Martin L, Simpson K, Brzezinski M, et al. Cellular response of keratinocytes to the entry and accumulation of nanoplastic particles. Part Fibre Toxicol. 2024;21(1):22. doi:10.1186/s12989-024-00583-9
81. Menichetti A, Mordini D, Montalti M. Penetration of microplastics and nanoparticles through skin: effects of size, shape, and surface chemistry. J Xenobiot. 2024;15(1):6. doi:10.3390/jox15010006
82. Song GB, Nam J, Ji S, et al. Deciphering the links: fragmented polystyrene as a driver of skin inflammation. J Hazard Mater. 2024;480:135815. doi:10.1016/j.jhazmat.2024.135815
83. Kantha P, Liu S-T, Horng J-L, et al. Acute exposure to polystyrene nanoplastics impairs skin cells and ion regulation in zebrafish embryos. Aquat Toxicol. 2022;248:106203. doi:10.1016/j.aquatox.2022.106203
84. Cheng S, Hu J, Guo C, et al. The effects of size and surface functionalization of polystyrene nanoplastics on stratum corneum model membranes: an experimental and computational study. J Colloid Interface Sci. 2023;638:778–787. doi:10.1016/j.jcis.2023.02.008
85. Gopinath PM, Saranya V, Vijayakumar S, et al. Assessment on interactive prospectives of nanoplastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Sci Rep. 2019;9(1):8860. doi:10.1038/s41598-019-45139-6
86. Han Y, Song Y, Kim GW, et al. No prominent toxicity of polyethylene microplastics observed in neonatal mice following intratracheal instillation to dams during gestational and neonatal period. Toxicol Res. 2021;37(4):443–450. doi:10.1007/s43188-020-00086-7
87. Sökmen T, Sulukan E, Türkoğlu M, et al. Polystyrene nanoplastics (20 nm) are able to bioaccumulate and cause oxidative DNA damages in the brain tissue of zebrafish embryo (Danio rerio). Neurotoxicology. 2020;77:51–59. doi:10.1016/j.neuro.2019.12.010
88. Yong CQY, Valiyaveettil S, Tang BL. Toxicity of microplastics and nanoplastics in mammalian systems. Int J Environ Res Public Health. 2020;17(5):1509. doi:10.3390/ijerph17051509
89. Horvatits T, Tamminga M, Liu B, et al. Microplastics detected in cirrhotic liver tissue. EBioMedicine. 2022;82:104147. doi:10.1016/j.ebiom.2022.104147
90. Donisi I, Colloca A, Anastasio C, et al. Micro(nano)plastics: an emerging burden for human health. Int J Biol Sci. 2024;20(14):5779–5792. doi:10.7150/ijbs.99556
91. Christensen MS, Vestergaard JM, d’Amore F, et al. Styrene exposure and risk of lymphohematopoietic malignancies in 73,036 reinforced plastics workers. Epidemiology. 2018;29(3):342–351. doi:10.1097/EDE.0000000000000819
92. Goodman KE, Hare JT, Khamis ZI, et al. Exposure of human lung cells to polystyrene microplastics significantly retards cell proliferation and triggers morphological changes. Chem Res Toxicol. 2021;34(4):1069–1081. doi:10.1021/acs.chemrestox.0c00486
93. Lin W, Luo H, Wu J, et al. Polystyrene microplastics enhance the microcystin-LR-induced gonadal damage and reproductive endocrine disruption in zebrafish. Sci Total Environ. 2023;876:162664. doi:10.1016/j.scitotenv.2023.162664
94. Yan Z, Liu Y, Zhang T, et al. Analysis of microplastics in human feces reveals a correlation between fecal microplastics and inflammatory bowel disease status. Environ Sci Technol. 2022;56(1):414–421. doi:10.1021/acs.est.1c03924
95. Chen Y, Nan Y, Xu L, et al. Polystyrene nanoplastics exposure induces cognitive impairment in mice via induction of oxidative stress and ERK/MAPK-mediated neuronal cuproptosis. Part Fibre Toxicol. 2025;22(1):13. doi:10.1186/s12989-025-00633-w
96. MohanKumar SM, Campbell A, Block M, et al. Particulate matter, oxidative stress and neurotoxicity. Neurotoxicology. 2008;29(3):479–488. doi:10.1016/j.neuro.2007.12.004
97. Chen H, Kwong JC, Copes R, et al. Living near major roads and the incidence of dementia, Parkinson’s disease, and multiple sclerosis: a population-based cohort study. Lancet. 2017;389(10070):718–726. doi:10.1016/S0140-6736(16)32399-6
98. Ranft U, Schikowski T, Sugiri D, et al. Long-term exposure to traffic-related particulate matter impairs cognitive function in the elderly. Environ Res. 2009;109(8):1004–1011. doi:10.1016/j.envres.2009.08.003
99. Barboza LGA, Vieira LR, Branco V, et al. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat Toxicol. 2018;195:49–57. doi:10.1016/j.aquatox.2017.12.008
100. Sun J, Peng S, Yang Q, et al. Microplastics/nanoplastics and neurological health: an overview of neurological defects and mechanisms. Toxicology. 2025;511:154030. doi:10.1016/j.tox.2024.154030
101. Gou Z, Wu H, Li S, et al. Airborne micro- and nanoplastics: emerging causes of respiratory diseases. Part Fibre Toxicol. 2024;21(1):50. doi:10.1186/s12989-024-00613-6
102. Chen L, Liu Y, Li H, et al. Size-dependent pulmonary toxicity and whole-body distribution of inhaled micro/nanoplastic particles in male mice from chronic exposure. Environ Sci Technol. 2025;59(14):6993–7003. doi:10.1021/acs.est.4c14232
103. Dutta P, Rabeeah S, Vargas A, et al. A comprehensive narrative review of potential gastrointestinal adverse effects from micro(nano) plastic exposure. Clin Gastroenterol Hepatol. 2026;24(4):881–892. doi:10.1016/j.cgh.2025.11.002
104. Chen X, Zhuang J, Chen Q, et al. Polyvinyl chloride microplastics induced gut barrier dysfunction, microbiota dysbiosis and metabolism disorder in adult mice. Ecotoxicol Environ Saf. 2022;241:113809. doi:10.1016/j.ecoenv.2022.113809
105. Molina E, Benedé S. Is there evidence of health risks from exposure to micro- and nanoplastics in foods? Front Nutr. 2022;9:910094. doi:10.3389/fnut.2022.910094
106. Ullah S, Ahmad S, Guo X, et al. A review of the endocrine disrupting effects of micro and nano plastic and their associated chemicals in mammals. Front Endocrinol. 2022;13:1084236. doi:10.3389/fendo.2022.1084236
107. Medley EA, Spratlen MJ, Yan B, et al. A systematic review of the placental translocation of micro- and nanoplastics. Curr Environ Health Rep. 2023;10(2):99–111. doi:10.1007/s40572-023-00391-x
108. Yu CW, Luk TC, Liao VH. Long-term nanoplastics exposure results in multi and trans-generational reproduction decline associated with germline toxicity and epigenetic regulation in Caenorhabditis elegans. J Hazard Mater. 2021;412:125173. doi:10.1016/j.jhazmat.2021.125173
109. Li Y, Piao G, Hu F, et al. The silent invasion of microplastics polyvinyl chloride and polyethylene terephthalate: potential impact on osteoporosis. J Hazard Mater. 2025;492:138074. doi:10.1016/j.jhazmat.2025.138074
110. Pan C, Hong R, Wang K, et al. Chronic exposure to polystyrene microplastics triggers osteoporosis by breaking the balance of osteoblast and osteoclast differentiation. Toxicology. 2025;510:154017. doi:10.1016/j.tox.2024.154017
111. Lihua C, Zhiyin T. Microplastics aggravates rheumatoid arthritis by affecting the proliferation/migration/inflammation of fibroblast-like synovial cells by regulating mitochondrial homeostasis. Int Immunopharmacol. 2023;120:110268. doi:10.1016/j.intimp.2023.110268
112. Lee SH, Jeong GH, Nam M-K, et al. Polystyrene microplastics activate NF-κB/MAPK signaling in synovial fibroblasts, promoting inflammation and joint destruction in rheumatoid arthritis. J Hazard Mater. 2025;499:140194. doi:10.1016/j.jhazmat.2025.140194
113. Zhang W, Li Z, Wang Z, et al. Polyethylene microplastics promote nucleus pulposus cell senescence by inducing oxidative stress via TLR4/NOX2 axis. Ecotoxicol Environ Saf. 2025;292:117950. doi:10.1016/j.ecoenv.2025.117950
114. Zhang W, Liu K, Li Z, et al. Polyethylene microplastics transported to nucleus pulposus cells via macrophages-derived exosomes, and promote intervertebral disc degeneration through NAT10/SLC7A11 and ac4C modification. Ecotoxicol Environ Saf. 2025;306:119324. doi:10.1016/j.ecoenv.2025.119324
115. Yu T, Zhang H, Sun W, et al. Nanoplastics exposure accelerates the progression of osteoarthritis via lysosomal destabilization-mediated pyroptosis. J Adv Res. 2026. doi:10.1016/j.jare.2026.04.045
116. Malik MA, Wu S, Zhang W, et al. Microplastics as an emerging driver of osteoarthritis: a translational synthesis of environmental exposure, patho-mechanisms, and public health implications. J Transl Med. 2025;23(1):1061. doi:10.1186/s12967-025-07081-2
117. Di J, Wang S, Liu L, et al. Unveiling the pathogenic mechanisms of polyethylene terephthalate-microplastic-driven osteoarthritis and rheumatoid arthritis: PTGS2 signaling hub-oriented toxicity profiling. Ecotoxicol Environ Saf. 2025;303:118816. doi:10.1016/j.ecoenv.2025.118816
118. Chen H, Wan L, Qiu Y, et al. Microplastics exposure induced and exacerbated the development of systemic lupus erythematosus in mice. Sci Total Environ. 2024;909:168586. doi:10.1016/j.scitotenv.2023.168586
119. Zeng W, He S, Zhao Y, et al. Microplastics exposure aggravates synovitis and pyroptosis in SLE by activating NF-κB and NRF2/KEAP1 signaling. Toxics. 2024;12(12):840. doi:10.3390/toxics12120840
120. Zhang W, Huang S, Liu K, et al. Polyethylene microplastics trigger osteosarcoma progression via ITGA5/FAPα/LGMN cancer promoting complex: a novel environmental cancer promoting mechanism. Ecotoxicol Environ Saf. 2025;305:119207. doi:10.1016/j.ecoenv.2025.119207
121. De Mattia G, Maffi M, Mazzantini M. Anabolic treatment for osteoporosis and fragility fracture risk: one year in review 2024. Clin Exp Rheumatol. 2024;42(7):1311–1316. doi:10.55563/clinexprheumatol/7f52vl
122. Genant HK, Cooper C, Poor G, et al. Interim report and recommendations of the world health organization task-force for osteoporosis. Osteoporos Int. 1999;10(4):259–264. doi:10.1007/s001980050224
123. Wright NC, Looker AC, Saag KG, et al. The recent prevalence of osteoporosis and low bone mass in the United States based on bone mineral density at the femoral neck or lumbar spine. J Bone Miner Res. 2014;29(11):2520–2526. doi:10.1002/jbmr.2269
124. Lane NE. Epidemiology, etiology, and diagnosis of osteoporosis. Am J Obstet Gynecol. 2006;194(2 Suppl):S3–11. doi:10.1016/j.ajog.2005.08.047
125. Weivoda MM, Bradley EW. Macrophages and Bone Remodeling. J Bone Miner Res. 2023;38(3):359–369. doi:10.1002/jbmr.4773
126. Zheng HW, Bui AAT, Ensrud KE, et al. Identifying younger postmenopausal women with osteoporosis using USPSTF-recommended osteoporosis risk assessment tools. JAMA Network Open. 2025;8(3):e250626. doi:10.1001/jamanetworkopen.2025.0626
127. Giannandrea D, Parolini M, Citro V, et al. Nanoplastic impact on bone microenvironment: a snapshot from murine bone cells. J Hazard Mater. 2024;462:132717. doi:10.1016/j.jhazmat.2023.132717
128. Scott DL, Wolfe F, Huizinga TW. Rheumatoid arthritis. Lancet. 2010;376(9746):1094–1108. doi:10.1016/S0140-6736(10)60826-4
129. McInnes IB, Schett G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet. 2017;389(10086):2328–2337. doi:10.1016/S0140-6736(17)31472-1
130. Padyukov L. Genetics of rheumatoid arthritis. Semin Immunopathol. 2022;44(1):47–62. doi:10.1007/s00281-022-00912-0
131. Scherer HU, Häupl T, Burmester GR. The etiology of rheumatoid arthritis. J Autoimmun. 2020;110:102400. doi:10.1016/j.jaut.2019.102400
132. Deane KD, Demoruelle MK, Kelmenson LB, et al. Genetic and environmental risk factors for rheumatoid arthritis. Best Pract Res Clin Rheumatol. 2017;31(1):3–18. doi:10.1016/j.berh.2017.08.003
133. Dowdell J, Erwin M, Choma T, et al. Intervertebral Disk Degeneration and Repair. Neurosurgery. 2017;80(3s):S46–s54. doi:10.1093/neuros/nyw078
134. Taylor W, Erwin WM. Intervertebral Disc Degeneration and Regeneration: new Molecular Mechanisms and Therapeutics: obstacles and Potential Breakthrough Technologies. Cells. 2024;13(24):2103. doi:10.3390/cells13242103
135. Kang L, Zhang H, Jia C, et al. Epigenetic modifications of inflammation in intervertebral disc degeneration. Ageing Res Rev. 2023;87:101902. doi:10.1016/j.arr.2023.101902
136. Xia Q, Zhao Y, Dong H, et al. Progress in the study of molecular mechanisms of intervertebral disc degeneration. Biomed Pharmacother. 2024;174:116593. doi:10.1016/j.biopha.2024.116593
137. Bermudez-Lekerika P, Crump KB, Tseranidou S, et al. Immuno-modulatory effects of intervertebral disc cells. Front Cell Dev Biol. 2022;10:924692. doi:10.3389/fcell.2022.924692
138. Zhang Y, Yang Y, Sun Y, et al. Assessing the toxicological impact of PET-MPs exposure on IVDD: insights from network toxicology and molecular docking. J Environ Manage. 2025;373:123830. doi:10.1016/j.jenvman.2024.123830
139. Barnett R. Osteoarthritis. Lancet. 2018;391(10134):1985. doi:10.1016/S0140-6736(18)31064-X
140. Bijlsma JW, Berenbaum F, Lafeber FP. Osteoarthritis: an update with relevance for clinical practice. Lancet. 2011;377(9783):2115–2126. doi:10.1016/S0140-6736(11)60243-2
141. Kiriakidou M, Ching CL. Systemic lupus erythematosus. Ann Intern Med. 2020;172(11):Itc81–itc96. doi:10.7326/AITC202006020
142. Durcan L, O’Dwyer T, Petri M. Management strategies and future directions for systemic lupus erythematosus in adults. Lancet. 2019;393(10188):2332–2343. doi:10.1016/S0140-6736(19)30237-5
143. Li H, Boulougoura A, Endo Y, et al. Abnormalities of T cells in systemic lupus erythematosus: new insights in pathogenesis and therapeutic strategies. J Autoimmun. 2022;132:102870. doi:10.1016/j.jaut.2022.102870
144. Dai X, Fan Y, Zhao X. Systemic lupus erythematosus: updated insights on the pathogenesis, diagnosis, prevention and therapeutics. Signal Transduct Target Ther. 2025;10(1):102. doi:10.1038/s41392-025-02168-0
145. Bu N, Du Q, Xiao T, et al. Mechanism of S -palmitoylation in polystyrene nanoplastics-induced macrophage cuproptosis contributing to emphysema through alveolar epithelial cell pyroptosis. ACS Nano. 2025;19(19):18708–18728. doi:10.1021/acsnano.5c02892
146. Yang JZ, Li J-H, Liu J-L, et al. Multiomics analysis revealed the effects of polystyrene nanoplastics at different environmentally relevant concentrations on intestinal homeostasis. Environ Pollut. 2025;372:126050. doi:10.1016/j.envpol.2025.126050
147. Feng X, Zhang M, Zhao T, et al. Polystyrene microplastics trigger colonic inflammation in rats via the TLR4/NF-κB/COX-2 pathway and modulation of intestinal microbiota. Toxicology. 2025;513:154090. doi:10.1016/j.tox.2025.154090
148. Chen J, Chen X, Xuan Y, et al. Surface functionalization-dependent inflammatory potential of polystyrene nanoplastics through the activation of MAPK/ NF-κB signaling pathways in macrophage Raw 264.7. Ecotoxicol Environ Saf. 2023;251:114520. doi:10.1016/j.ecoenv.2023.114520
149. Husain A, Jeffries MA. Epigenetics and Bone Remodeling. Curr Osteoporos Rep. 2017;15(5):450–458. doi:10.1007/s11914-017-0391-y
150. Wang T, Wang L, Zhang L, et al. Single-cell RNA sequencing in orthopedic research. Bone Res. 2023;11(1):10. doi:10.1038/s41413-023-00245-0
151. Wang H, Cheng P, Wang J, et al. Advances in spatial transcriptomics and its application in the musculoskeletal system. Bone Res. 2025;13(1):54. doi:10.1038/s41413-025-00429-w
152. Vethaak AD, Legler J. Microplastics and human health. Science. 2021;371(6530):672–674. doi:10.1126/science.abe5041
153. Chen J, Wu J, Sherrell PC, et al. How to build a microplastics-free environment: strategies for microplastics degradation and plastics recycling. Adv Sci. 2022;9(6):e2103764. doi:10.1002/advs.202103764
154. Kumar R, Manna C, Padha S, et al. Micro(nano)plastics pollution and human health: how plastics can induce carcinogenesis to humans? Chemosphere. 2022;298:134267. doi:10.1016/j.chemosphere.2022.134267
155. Dhaka V, Singh S, Anil AG, et al. Occurrence, toxicity and remediation of polyethylene terephthalate plastics. A review. Environ Chem Lett. 2022;20(3):1777–1800. doi:10.1007/s10311-021-01384-8
156. Eerkes-Medrano D, Thompson RC, Aldridge DC. Microplastics in freshwater systems: a review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015;75:63–82. doi:10.1016/j.watres.2015.02.012
157. Zuri G, Karanasiou A, Lacorte S. Human biomonitoring of microplastics and health implications: a review. Environ Res. 2023;237(Pt 1):116966. doi:10.1016/j.envres.2023.116966
158. Eberhard T, Casillas G, Zarus GM, et al. Systematic review of microplastics and nanoplastics in indoor and outdoor air: identifying a framework and data needs for quantifying human inhalation exposures. J Expo Sci Environ Epidemiol. 2024;34(2):185–196. doi:10.1038/s41370-023-00634-x
159. Amulya K, Verma P, Sheikh ZUD, et al. Comprehensive strategies for mitigating microplastic pollution: from detection to remediation. Global Chang Biol. 2025;31(10):e70526. doi:10.1111/gcb.70526
160. Shi X, Zhang X, Gao W, et al. Removal of microplastics from water by magnetic nano-Fe(3)O(4). Sci Total Environ. 2022;802:149838. doi:10.1016/j.scitotenv.2021.149838
161. Zhou H, Mayorga-Martinez CC, Pumera M. Microplastic removal and degradation by mussel-inspired adhesive magnetic/enzymatic microrobots. Small Methods. 2021;5(9):e2100230. doi:10.1002/smtd.202100230
162. Zhu L, Zhu J, Zuo R, et al. Identification of microplastics in human placenta using laser direct infrared spectroscopy. Sci Total Environ. 2023;856(Pt 1):159060. doi:10.1016/j.scitotenv.2022.159060
163. Sun J, Sui M, Wang T, et al. Detection and quantification of various microplastics in human endometrium based on laser direct infrared spectroscopy. Sci Total Environ. 2024;906:167760. doi:10.1016/j.scitotenv.2023.167760
164. Garcia MA, Liu R, Nihart A, et al. Quantitation and identification of microplastics accumulation in human placental specimens using pyrolysis gas chromatography mass spectrometry. Toxicol Sci. 2024;199(1):81–88. doi:10.1093/toxsci/kfae021
165. Sorolla-Rosario D, Llorca-Porcel J, Pérez-Martínez M, et al. Microplastics’ analysis in water: easy handling of samples by a new thermal extraction desorption-gas chromatography-mass spectrometry (TED-GC/MS) methodology. Talanta. 2023;253:123829. doi:10.1016/j.talanta.2022.123829
166. Wang ZM, Wagner J, Ghosal S, et al. SEM/EDS and optical microscopy analyses of microplastics in ocean trawl and fish guts. Sci Total Environ. 2017;603-604:616–626. doi:10.1016/j.scitotenv.2017.06.047
167. Pan W, Hao J, Zhang M, et al. Identification and analysis of microplastics in peritumoral and tumor tissues of colorectal cancer. Sci Rep. 2025;15(1):16130. doi:10.1038/s41598-025-98268-6
168. Shim WJ, Song YK, Hong SH, et al. Identification and quantification of microplastics using Nile Red staining. Mar Pollut Bull. 2016;113(1–2):469–476. doi:10.1016/j.marpolbul.2016.10.049
169. Zhang L, Tian J, Zhu X, et al. Cross-platform detection of microplastics in human biological tissues: comparing spectroscopic and chromatographic approaches. J Hazard Mater. 2025;492:138133. doi:10.1016/j.jhazmat.2025.138133
170. Choi S, Lee S, Kim M-K, et al. Challenges and recent analytical advances in micro/nanoplastic detection. Anal Chem. 2024;96(22):8846–8854. doi:10.1021/acs.analchem.3c05948
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.