Back to Journals » Infection and Drug Resistance » Volume 11
Enriched whole genome sequencing identified compensatory mutations in the RNA polymerase gene of rifampicin-resistant Mycobacterium leprae strains
Authors Lavania M, Singh I, Turankar RP ![]() , Gupta AK, Ahuja M, Pathak V, Sengupta U
, Gupta AK, Ahuja M, Pathak V, Sengupta U
Received 19 September 2017
Accepted for publication 13 November 2017
Published 25 January 2018 Volume 2018:11 Pages 169—175
DOI https://doi.org/10.2147/IDR.S152082
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Mallika Lavania,1 Itu Singh,1 Ravindra P Turankar,1 Anuj Kumar Gupta,2 Madhvi Ahuja,1 Vinay Pathak,1 Utpal Sengupta1
1Stanley Browne Laboratory, The Leprosy Mission Trust India, TLM Community Hospital Nand Nagari, 2Agilent Technologies India Pvt Ltd, Jasola District Centre, New Delhi, India
Abstract: Despite more than three decades of multidrug therapy (MDT), leprosy remains a major public health issue in several endemic countries, including India. The emergence of drug resistance in Mycobacterium leprae (M. leprae) is a cause of concern and poses a threat to the leprosy-control program, which might ultimately dampen the achievement of the elimination program of the country. Rifampicin resistance in clinical strains of M. leprae are supposed to arise from harboring bacterial strains with mutations in the 81-bp rifampicin resistance determining region (RRDR) of the rpoB gene. However, complete dynamics of rifampicin resistance are not explained only by this mutation in leprosy strains. To understand the role of other compensatory mutations and transmission dynamics of drug-resistant leprosy, a genome-wide sequencing of 11 M. leprae strains – comprising five rifampicin-resistant strains, five sensitive strains, and one reference strain – was done in this study. We observed the presence of compensatory mutations in two rifampicin-resistant strains in rpoC and mmpL7 genes, along with rpoB, that may additionally be responsible for conferring resistance in those strains. Our findings support the role for compensatory mutation(s) in RNA polymerase gene(s), resulting in rifampicin resistance in relapsed leprosy patients.
Keywords: leprosy, rifampicin resistance, compensatory mutations, next generation sequencing, relapsed, MDT, India
Introduction
Since the introduction of effective multidrug therapy (MDT) in 1985, India is still inhabited by more than 60% of the leprosy patients of the world – a situation that has remained unchanged despite massive decline in the global prevalence of the disease. However, the attainment of the prevalence rate (PR) of 0.66 per 10,000 population indicated the progress of elimination.1 At this crucial stage of elimination, the emergence of drug resistance in Mycobacterium leprae (M. leprae) to rifampicin is being reported from different endemic populations of India.2,3 If this emergence of rifampicin resistance is not checked immediately with the administration of an alternative drug regimen, the elimination program, which relies on rifampicin-based MDT, will delay the achievement of the leprosy control program of the country. Although rifampicin-based MDT, recommended by the World Health Organization (WHO; 1982), has been successful in reducing the number of leprosy cases around the world, the global registered prevalence of leprosy at the end of 2015 was still 176,176, and the new cases reported globally in 2015 was 211,973. The number of registered cases and new cases almost remained steady for many years.4 MDT was originally designed to prevent the emergence and spread of drug-resistant strains and contained rifampicin, dapsone, and clofazimine. Rifampicin, the backbone of MDT, is a bactericidal drug, which binds to the beta subunit of the RNA polymerase encoded by rpoB and inhibits transcription. Recent reports have, however, indicated the occurrence of rifampicin resistance in several endemic areas such as Brazil, India, and People’s Republic of China.2,3,5–7 At this present stage, it is important to monitor the emergence of rifampicin-resistant mutants in endemic populations where transmission of M. leprae infection is continuing. On the other hand, resistance to dapsone has been reported since the late 1960s. Moreover, a strain showing resistance to both dapsone and rifampicin was reported in 1993.8 At present, further reports indicating the emergence of M. leprae strains resistant to multiple drugs are also emerging.2,9,10
As M. leprae cannot be cultured in vitro, molecular drug susceptibility testing, including PCR-sequencing, for determining resistant strains provides a practical alternative. At present, the rapid detection and control of such drug-resistant strains is essential in countries approaching leprosy elimination levels, such as India. As new cases are continuing, which is considered a major indicator for disease transmission, there is still a need for better measures to control leprosy. Furthermore, relapsed cases with drug-resistant strains could be a new source of disease transmission. Therefore, at this juncture of elimination, it will be essential to conduct a thorough investigation of relapsed multibacillary (MB) cases and all new cases to identify the pattern of existence of primary and secondary resistance to anti-leprosy drugs such as rifampicin and dapsone. Whole genome sequencing (WGS) of M. leprae directly from clinical samples is a good tool for identifying mutations that cause drug resistance, especially those that are not known or well characterized. In biopsy samples, M. leprae genome copies can vary in number, but the relative proportion of bacterial DNA is minute in comparison to host DNA. Direct sequencing of mixed human and bacterial DNA does not yield significant proportions of sequence reads that map to the bacterial genome. For this reason, we used a capture probe-based bacterial DNA enrichment method which selectively isolates bacterial DNA from host DNA prior to sequencing. With the help of this approach, the present study focuses on identifying compensatory mutations in genes other than, but related to, rpoB in rifampicin-resistant strains of M. leprae.
Materials and methods
Ethical approval
Ethical clearance for this study was approved by the The Leprosy Mission Trust India Ethical Committee, held on August 29, 2014, under the regulations of the Indian Council of Medical Research. Written informed consent was obtained from all subjects before collection of biological samples.
Source of biopsies for DNA extraction
A total of ten relapsed leprosy patients from hospitals of The Leprosy Mission Trust India (TLMTI) and one reference control strain (Br4923) from BEI Resources, USA, were included in this study. Among these ten strains, five were resistant to rifampicin, characterized by molecular methods and PCR-sequencing. The remaining five strains were drug sensitive. M. leprae DNA was extracted from these ten clinical biopsy specimens using the Blood and Tissue DNA extraction kit (QIAGEN, USA). Single-nucleotide polymorphism (SNP) subtyping data was generated by PCR sequencing11 from DNA before carrying out WGS for the remaining DNA.
Library preparation and sequencing
Target capture sequencing libraries were prepared with Illumina-compatible SureSelect QXT Library Prep Reagent Kit (Agilent Technologies, Santa Clara, CA, USA) at Genotypic Technology Pvt. Ltd., Bangalore, India. Briefly, 50 ng Qubit-quantified genomic DNA was fragmented and adapter-tagged using SureSelect QXT Enzyme (Table S1). The fragmented and adapter-tagged DNA was purified with HighPrep magnetic beads and then amplified by eight cycles of PCR. The PCR-enriched products were purified with HighPrep beads, followed by library quality control check using the Agilent 2100 Bioanalyzer.
Target enrichment was undertaken according to the manufacturer’s instructions using SureSelect RNA capture baits. In-solution hybridization was done as per the SureSelect QXT hybridization method. After hybridization, the captured biotinylated probe-target hybrids were pulled down by using streptavidin-coated magnetic beads (Dynabeads MyOne Streptavidin T1, ThermoFisher Scientific, Waltham, MA, USA). The magnetic beads were washed according to the manufacturer’s instructions and resuspended in 15 μl nuclease-free water. The captured DNA libraries were amplified by 12 cycles of PCR with the inclusion of the appropriate indexing primer for each sample.
The final PCR product (sequencing library) was purified with HighPrep beads, followed by quantification by a Qubit fluorometer (ThermoFisher Scientific, MA, USA), and fragment size distribution was analyzed on an Agilent 2100 Bioanalyzer.
Sequencing was carried out on an Illumina Hi-Seq 2500 instrument and the Illumina paired-end raw reads were quality checked using FastQC. Illumina raw reads were processed for adapters and trimming of low-quality bases. The processed reads were aligned with M. leprae TN reference using a Burrows–Wheeler Aligner (BWA) 0.7.5 algorithm, and the variants were identified with Samtools 1.2 and Bcftools 1.2 filtered for a read depth threshold of greater than 20, and a quality threshold of more than Q30. Filtered variants were further annotated with SnpEff to get the information on the genes, protein change, and the impact of the variation.
Results
Demographics and diagnosis
Among the five rifampicin-resistant patients, one patient was a defaulter who took MDT for 2 months only and remaining four patients were those who relapsed after completion of a full course of MDT. One of the patients (1027) had a dual infection of leprosy and tuberculosis. The demographic and clinical details of these five resistant patients together with the five sensitive ones are mentioned in Table 1A and B.
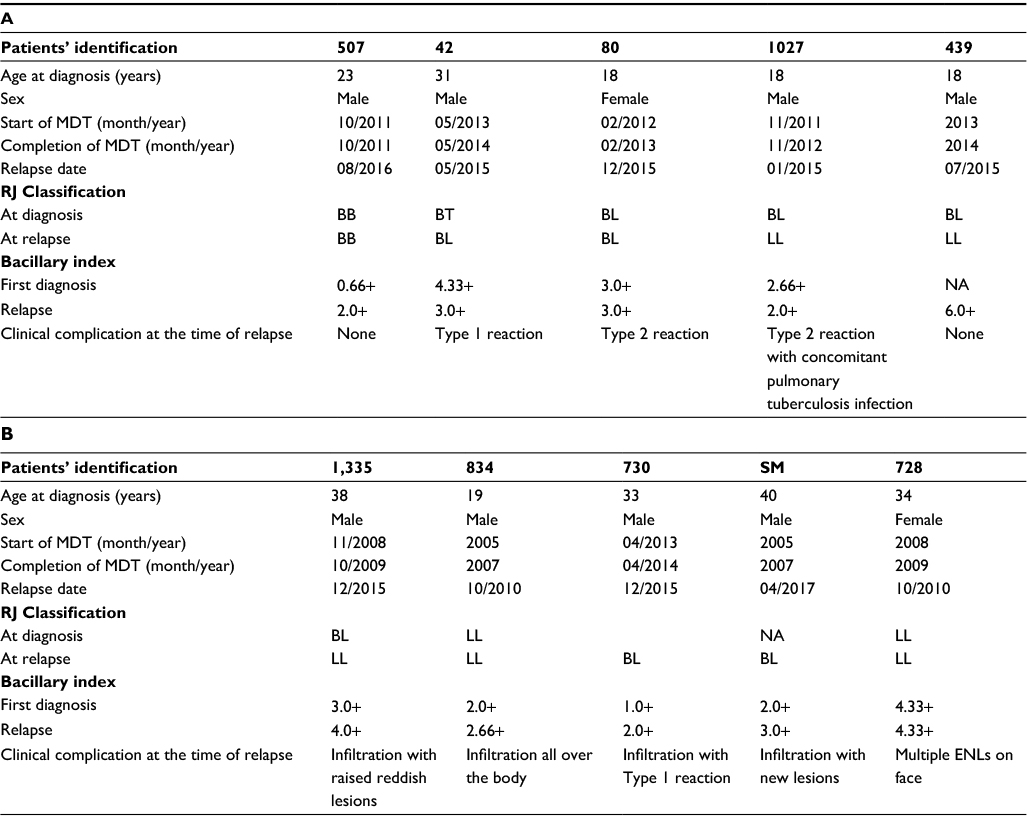 | Table 1 Demographic and clinical details of rifampicin-resistant cases (A). Demographic and clinical details of rifampicin-sensitive cases (B). |
All resistant strains were characterized for drug-susceptibility testing by PCR sequencing. We found the already published rpoB mutations in three patients (507, 42, and 80) at codon positions Leu436Meth, Asp441Tyr, and Ser456Leu, except in two (1027 and 439) who had mutations at codon positions Gln442His and Ser437Pro (Table 2). These two patients did not respond to the treatment. Gln442His mutation in patient 1027 and Ser437Pro in patient 439 of rpoB gene did not confer resistance to rifampicin, which was confirmed by mouse foot pad growth earlier.12 The patient 1027 continuously visited the hospital with recurrent Type 2 reaction and other complications despite being administered thalidomide. He was later diagnosed with a pulmonary tuberculosis infection along with leprosy. He was started on anti-tuberculosis treatment (ATT) as well, but the infection was unalleviated. We tested the same specimen with Mycobacterium tuberculosis-specific rpoB primers, undertook DNA sequencing, and found him susceptible for the TB rpoB gene. This led us to consider the possibility that other mutations at other genes play a role in conferring resistance in such cases.
 | Table 2 Mutations in rpoB and rpoC genes by WGS in molecularly characterized rifampicin-resistant strains Note: *Amino acid. Abbreviations: RIF, rifampicin; NM, no mutation; WGS, whole genome sequencing. |
Genetic polymorphisms
Whole genome read coverage was adequately generated from the 10 M. leprae isolates, and the reference strain for comparative genomic analysis. Among these ten samples, 11,791 variants were identified, including 10,833 (92 %) SNPs and 958 (8.0%) Indels. As there is no previous database to filter out common variants, we focused on rpoB and its associated genes. After analyzing the data of the five rifampicin-resistant strains, we found that two strains showed mutation in the rpoC gene (Patient ID 1027 and 439) along with the rpoB gene (Table 2) as well as a non-synonymous mutation in the resistance nodulation division (RND) family transporter efflux pump gene mmpL 7. These two genes are mainly responsible for causing resistance among the rest of the SNPs present in these strains.
Analysis of rifampicin-resistant strains revealed 1,308 variants, of which there were 44 synonymous, 101 non-synonymous, 112 intergenic, and 12 frame-shift mutations. On genome comparison of all ten strains, we observed seven unique SNPs. While comparing resistant strains with other sensitive strains, it revealed 16 unique SNPs that were present only in these strains (Table 3). An insertion of eight nucleotides (TTTCTTAT at position 508802) was found in ML0411, encoding the Pro-Pro-Glu (PPE) serine-rich antigen.
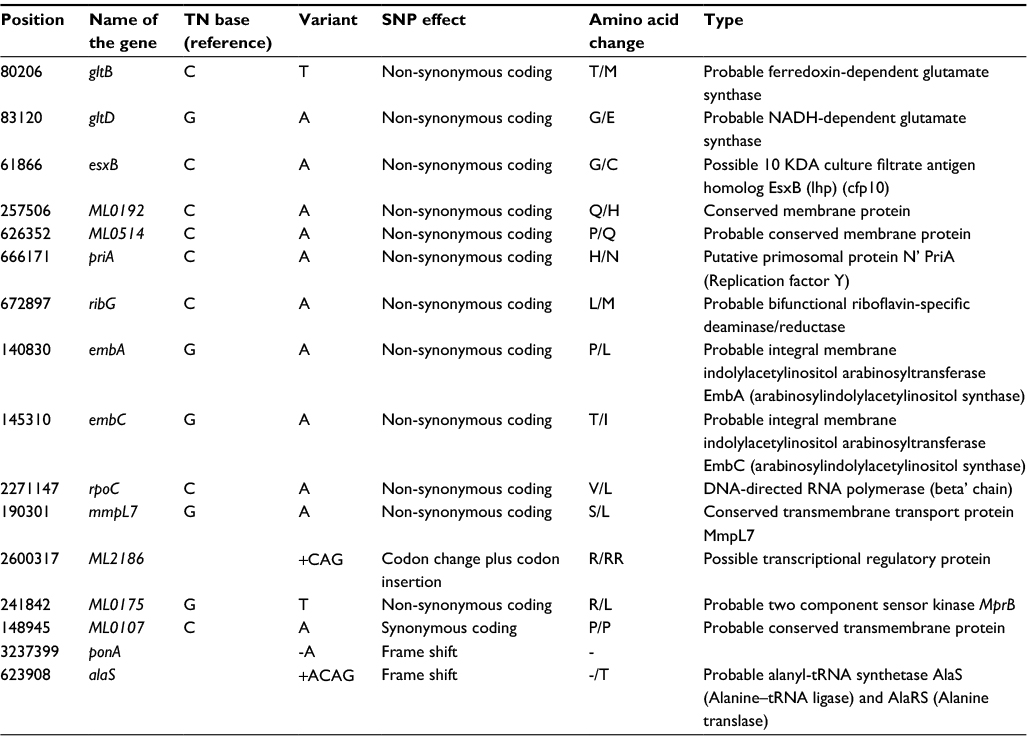 | Table 3 List of unique 16 SNPs present in resistant strains Abbreviation: SNP, single-nucleotide polymorphism. |
Discussion
The emergence of rifampicin resistance in M. leprae after more than 30 years of MDT is not an unexpected outcome. The occurrence of dapsone resistance also took nearly the same duration. However, the emergence of rifampicin-resistant M. leprae during the phase of elimination may cause a major setback to the public health program. Drug-resistant leprosy infection can be caused by transmission of already resistant strains (primary resistance) or by selection of resistance-conferring mutations during inadequate therapy (secondary resistance). Our own work2,3 has shown the presence of rifampicin, dapsone, and ofloxacin resistance cases in highly endemic areas in several states of India. M. leprae isolates resistant to single and multiple drugs have already been encountered.2,13 Although the genome of M. leprae has undergone massive gene decay,14 the rifampicin resistance determining region (RRDR) region of the organism and the associated genes are still functional. In order to monitor the transmission dynamics of drug-resistant leprosy, a comparison of M. leprae strains was carried out by WGS of strains derived from rifampicin-resistant leprosy patients in this study.
Rifampicin inhibits bacterial RNA synthesis by binding strongly to RNA polymerase, preventing attachment of the enzyme to DNA and thus blocking the initiation of transcription. Bacterial resistance to rifampicin is caused by mutations leading to a change in the structure of the beta subunit of RNA polymerase. Resistance to rifampicin is one of the main causes of treatment failure.15 Various studies have shown that resistance to rifampicin in M. tuberculosis as well as in M. leprae is mainly due to a single mutation in an 81-bp region in the rpoB gene.16,17 Furthermore, it has been reported earlier that there are some alternative mechanisms of rifampicin resistance in various bacterial species such as rox gene-mediated mono-oxygenation of rifampicin18 and duplication of the rpoB gene as rpoB2 in Nocardia.19 In our study, all strains belonged to the SNP subtype 1D, which is the predominant genotype prevalent in many countries such as India, Bangladesh, Nepal, Madagascar, Malawi, and the French West Indies.11,20,21
The coexistence of the two mycobacteria has been demonstrated in archaeological samples by Donoghue et al,22 who identified DNA from both tuberculosis and leprosy in the archeological samples from the Roman period to the 13th century from several sites around the world. On review of data from three leprosy referral centers in Hyderabad, India, from 2000 to 2013, Rawson et al23 identified three cases of concomitant disease. Similarly, we observed in one of our samples a patient (Pt1027) who is suffering from both TB and leprosy.
This study provides a better explanation for the presence of the rifampicin-resistance phenotype in leprosy cases in India. It has identified unique SNPs, including non-synonymous SNPs in gltB, gltD, and esxB and mutation in ctpC and mmpL7 transporter genes. The mmpL7 gene in M. tuberculosis was studied and characterized by Pasca et al.24 It encodes a protein which belongs to RND drug transporter family. The MmpL7 protein is of 920 amino acids with a predicted molecular mass of 95.1 kDa, contains 12 transmembrane domains (TMDs) having two large hydrophilic extracytoplasmic domains between TMD-1 and TMD-2 and between TMD-7 and TMD-8. All of these characteristics have been described as typical of RND efflux pumps.25 The MmpL7 protein confers a high level of resistance to isoniazid.24 However, a functional assay is required to determine whether either of these variant genes/transporters confers any degree of rifampicin resistance in M. leprae. In our study, the rpoC gene has a non-synonymous SNPs sequence in two patients – 1027 and 439. Mouse foot pad assay was done for the patient 1027 to check whether the mutation in 442 codon position was conferring resistance. In our previous study, we observed that this 442 codon mutation was not conferring resistance in mice.12 Therefore, it may be suggested that these compensatory mutations in rpoC and/or transporter genes play a role in conferring resistance in this strain. Thus, our study suggested the role of compensatory mutation in rpoC and possibly of drug transporter genes in contributing to acquired resistance in rifampicin-resistant strains in M. leprae.
Acknowledgments
The authors thank the Indian Council of Medical Research (Grant No. ECD/Ad-hoc/leprosy/2014-113/Fy. 14-15/19/Delhi/NGO-ECD-I) and England and Wales Foundation (205T03) for the financial support. The authors are likewise grateful to Atul Roy and Manish Gardia for assisting us in the sample collection, and thank the superintendent and staff of TLM Hospitals for their help and assistance during the work.
Disclosure
The authors report no conflicts of interest in this work.
References
NLEP. NLEP Annual report 2015–2016. Available from: www.nlep.nic.in. Accessed July 20, 2017. | ||
Lavania M, Jadhav RS, Chaitanya VS, et al. Drug resistance patterns in Mycobacterium leprae isolates from relapsed leprosy patients attending The Leprosy Mission (TLM) Hospitals in India. Lepr Rev. 2014;85(3):177–185. | ||
Lavania M, Nigam A, Turankar RP, et al. Emergence of primary drug resistance to rifampicin in Mycobacterium leprae strains from leprosy patients in India. Clin Microbiol Infect. 2015;21(12):e85–e86. | ||
World Health Organization. Leprosy today. Fact sheet [updated February 2017]. Available from: http://www.who.int/mediacentre/factsheets/fs101/en/. Accessed July 20, 2017. | ||
da Silva Rocha A, Cunha Md, Diniz LM, et al. Drug and multidrug resistance among Mycobacterium leprae isolates from Brazilian relapsed leprosy patients. J Clin Microbiol. 2012;50(6):1912–1917. | ||
Contreras Mejía Mdel C, Porto Dos Santos M, Villarouco da Silva GA, et al. Identification of primary drug resistance to rifampin in Mycobacterium leprae strains from leprosy patients in Amazonas State, Brazil. J Clin Microbiol. 2014;52(12):4359–4360. | ||
Liu D, Zhang Q, Sun Y, et al. Drug resistance in Mycobacterium leprae from patients with leprosy in China. Clin Exp Dermatol. 2015;40(8):908–911. | ||
González AB, Maestre JL, Hernández O, et al. Survey for secondary dapsone and rifampicin resistance in Cuba. Lepr Rev. 1993;64(2):128–135. | ||
Cambau E, Perani E, Guillemin I, Jamet P, Ji B. Multidrug resistance to dapsone, rifampicin, and ofloxacin in Mycobacterium leprae. Lancet. 1997;349(9045):103–104. | ||
Matsuoka M, Kashiwabara Y, Namisato M. A Mycobacterium leprae isolate resistant to dapsone, rifampin, ofloxacin and sparfloxacin. Int J Lepr Other Mycobact Dis. 2000;68(4):452–455. | ||
Monot M, Honoré N, Garnier T, et al. Comparative genomic and phylogeographic analysis of Mycobacterium leprae. Nat Genet. 2009;41(12):1282–1289. | ||
Lavania M, Hena A, Reja H, et al. Mutation at codon 442 in the rpoB gene of Mycobacterium leprae does not confer resistance to rifampicin. Lepr Rev. 2016;87(1):93–100. | ||
Maeda S, Matsuoka M, Nakata N, et al. Multidrug resistant Mycobacterium leprae from patients with leprosy. Antimicrob Agents Chemother. 2001;45(12):3635–3639. | ||
Cole ST, Eiglmeier K, Parkhill J, et al. Massive gene decay in the leprosy bacillus. Nature. 2001;409(6823):1007–1011. | ||
Mitchison DA, Nunn AJ. Influence of initial drug resistance on the response to short-course chemotherapy of pulmonary tuberculosis. Am Rev Respir Dis. 1986;133(3):423–430. | ||
Telenti A, Imboden P, Marchesi F, et al. Detection of rifampicin-resistance mutations in Mycobacterium tuberculosis. Lancet. 1993; 341(8846):647–650. | ||
Honore N, Cole ST. Molecular basis of rifampin resistance in Mycobacterium leprae. Antimicrob Agents Chemother. 1993;37(3):414–418. | ||
Hoshino Y, Fujii S, Shinonaga H, et al. Monooxygenation of rifampicin catalyzed by the rox gene product of Nocardia farcinica: structure elucidation, gene identification and role in drug resistance. J Antibiot (Tokyo). 2010;63(1):23–28. | ||
Ishikawa J, Chiba K, Kurita H, Satoh H. Contribution of rpoB2 RNA polymerase beta subunit gene to rifampin resistance in Nocardia species. Antimicrob Agents Chemother. 2006;50(4):1342–1346. | ||
Kuruwa S, Vissa V, Mistry N. Distribution of Mycobacterium leprae strains among cases in a rural and urban population of Maharashtra, India. J Clin Microbiol. 2012;50(4):1406–1411. | ||
Lavania M, Jadhav RS, Turankar RP, Chaitanya VS, Singh M, Sengupta U. Single nucleotide polymorphisms typing of Mycobacterium leprae reveals focal transmission of leprosy in high endemic regions of India. Clin Microbiol Infect. 2013;19(11):1058–1062. | ||
Donoghue HD, Marcsik A, Matheson C, et al. Co-infection of Mycobacterium tuberculosis and Mycobacterium leprae in human archaeological samples: a possible explanation for the historical decline of leprosy. Proc Biol Sci. 2005;272(1561):389–394. | ||
Rawson TM, Anjum V, Hodgson J, et al. Leprosy and tuberculosis concomitant infection: a poorly understood, age-old relationship. Lepr Rev. 2014;85(4):288–295. | ||
Pasca MR, Guglierame P, De Rossi E, Zara F, Riccardi G. mmpL7 gene of Mycobacterium tuberculosis is responsible for isoniazid efflux in Mycobacterium smegmatis. Antimicrob Agents Chemother. 2005;49(11):4775–4777. | ||
Tseng TT, Gratwick KS, Kollman J, et al. The RND permease superfamily: an ancient, ubiquitous and diverse family that includes human disease and development proteins. J Mol Microbiol Biotechnol. 1999;1(1):107–125. |
Supplementary material
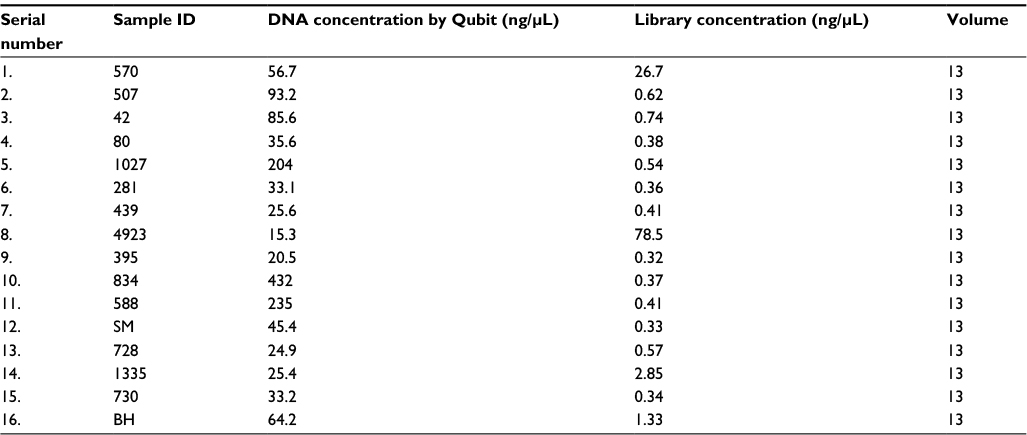 | Table S1 Concentration of DNA from biopsies and after library preparation Abbreviations: BB, borderline leprosy; BL, borderline lepromatous; BT, borderline tuberculoid; LL, lepromatous leprosy; MDT, multidrug therapy; NA, not available; RJ, Ridley–Jopling. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.