Back to Journals » International Journal of Nanomedicine » Volume 21
Engineering the Future: Strategic Advances in Extracellular Vesicle-Mediated Drug Delivery Systems
Received 20 November 2025
Accepted for publication 25 February 2026
Published 30 March 2026 Volume 2026:21 583242
DOI https://doi.org/10.2147/IJN.S583242
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Kamakhya Misra
Xiran Zhang,1,2 Xiaomin Zhang1,2
1Tianjin Key Laboratory of Ophthalmology and Visual Science, Tianjin Eye Institute, Tianjin Eye Hospital, Nankai University Affiliated Tianjin Eye Hospital; Clinical College of Ophthalmology, Tianjin Medical University, Tianjin, 30020, People’s Republic of China; 2Tianjin Key Laboratory of Retinal Functions and Diseases, Tianjin Branch of National Clinical Research Center for Ocular Disease, Eye Institute and School of Optometry, Tianjin Medical University Eye Hospital, Tianjin, People’s Republic of China
Correspondence: Xiaomin Zhang, Email [email protected]
Abstract: Extracellular vesicles (EVs) are lipid bilayer-enclosed nanoparticles naturally secreted by cells that inherently lack replicative capacity and function as endogenous carriers of biological cargo including proteins, nucleic acids, and metabolites for intercellular communication. Leveraging their intrinsic biocompatibility and biomimetic transport properties, EVs have emerged as versatile drug delivery platforms with distinct therapeutic advantages. Recent advancements have developed two precision-engineered derivatives: structurally and cargo-modified engineered EVs, and EV mimetics integrating synthetic nanomaterials. Both types are designed to enhance targeting specificity and therapeutic efficacy, yet their strong intercorrelations frequently cause confusion. This review systematically examines the evolving landscape of EV-based delivery systems by establishing conceptual distinctions between native EVs, engineered EVs, and EV mimetics, while comparatively analyzing their preparation methodologies, clinical translation progress, and performance characteristics as drug carriers. Through systematic discussion of clinical challenges, including safety, clinical feasibility, and cross-laboratory reproducibility, we propose optimization directions integrating artificial intelligence with drug delivery systems, thereby providing insights and methodologies for next-generation EV-inspired therapeutic delivery platforms.
Keywords: extracellular vesicles, engineered extracellular vesicles, extracellular vesicle mimetics, drug delivery system, synthetic nanoparticles
Introduction
Extracellular vesicles (EVs) are nanoscale lipid-bilayer-enclosed particles released by eukaryotic and prokaryotic cells. They lack functional nuclei and replication capacity,1 and mediate intercellular communication by transporting diverse biomolecules, including nucleic acids, proteins, and lipids.2,3 EVs were initially identified in the 1950s by Chargaff and West as “precipitates during blood coagulation”4 and were long considered by researchers to be mere “garbage bins” of cellular metabolites.5,6 It was not until the early 1980s that these vesicles, which range in diameter from 40 nanometers to 1 micrometer and are released by diverse cell types, were designated as “exosomes”.7 In 1996, Raposo demonstrated the antigen-presenting function of immune cell-derived EVs, a finding that fundamentally challenged the view of EVs as non-functional metabolic byproducts and marked the beginning of a new era in research focused on their role as intercellular communication vehicles.8 With the advent of the 21st century, the discovery that EVs could transport functional RNA ignited a surge of scientific interest. However, inconsistencies in nomenclature and classification, together with the absence of standardized isolation and detection protocols, have resulted in poor reproducibility of experimental outcomes and a fragmented progression of the field. These vesicles exhibit diverse origins, natures, and features, which results in heterogeneous nomenclature within the literature based on size (eg microvesicles, nanoparticles), cellular origin (eg oncosomes), proposed functions (eg argosomes, tolerosomes), or extracellular location (eg ectosomes, exosomes). In fact, it was not until the founding of the International Society for Extracellular Vesicles (ISEV) in 2011, the term “extracellular vesicles” was formally proposed. They recommended the classification of EVs considering size —small EVs (sEVs) < 200 nm vs large EVs (lEVs) > 200 nm, and biogenesis pathways —apoptotic bodies (500–1000 nm) derived from programmed cell death, microvesicles (150–500 nm) generated through plasma membrane budding, and exosomes (40–150 nm) originating from endosomal multivesicular bodies.9 Current isolation methods hinder precise separation of co-enriched EV subtypes, with no universal biomarkers confirming biogenesis-dependent characteristics—a consensus reflected by predominant use of “EVs” in contemporary literature, though early studies employed terms like “exosomes” (denoting sEVs isolated via 220 nm filtration or high-speed ultracentrifugation10) and “microvesicles” (lEVs obtained through differential centrifugation, ultracentrifugation, or filtration11). Notably, no cited evidence confirms intracellular origins, which leads researchers to adopt “sEV” for exosome-related and “lEV” for microvesicle-related references.
EVs mediate intercellular communication and remodel the phenotype of target cells through selective cargo sorting during biogenesis, regulated release, and diverse pathways involving ligand-receptor interactions, endocytic uptake, or membrane fusion with target cells with inherent biocompatibility,12 low immunogenicity13 and efficient tissue penetration,14 are promising for biomolecule delivery and non-invasive disease diagnosis.15 Despite these advantages, limitations in production scalability,16 isolation purity,17 predictability in vivo targeting precision, and storage inefficiencies18 persist. To address these issues, engineered EVs have been developed via surface functionalization and cargo engineering for precision-targeted therapy.19,20 Yet, their complex preparation increases cost and technical difficulty, may damage EV membrane integrity, and lacks standardized protocols leading to uncontrollable batch variation. For large-scale clinical use, EV mimetics—artificial nanovesicles mimicking EV membrane structure—including artificial cell-derived vesicles (ACDVs) fabricated via mechanical cell disruption and synthetic vesicles (SVs) constructed de novo from molecular components or hybridized with natural EVs/liposomes, have emerged as synthetic alternatives. However, they cannot replicate natural EV surface ligands or cell tropism, resulting in inferior biocompatibility, tissue penetration and higher immunogenicity.
This review systematically examines native EVs, engineered EVs, and EV mimetics to delineate their conceptual distinctions and interrelationships while evaluating their biomedical potential through comparative analysis of standardized production methodologies, therapeutic applications, and technological limitations. We categorize established production methodologies by biophysical principles and functional objectives; benchmark protocol variations for analogous particle types; and then synthesize clinical translation outcomes across therapeutic domains including oncology, regenerative medicine, and gene therapy. Through critical analysis and discussion, we identify translational issues such as clinical safety, batch-to-batch consistency, and large-scale production capacity. Finally, we propose mechanistic optimizations and innovative strategies for artificial intelligence-integrated drug delivery systems (DDSs) to advance next-generation vesicular nanomedicines.
Production
EVs
EV Characterization
EV characterization is for confirming presence, assessing quantity, distinguishing them from non-EV components, and elucidating their biological properties, commonly conducted across morphology, particle size, and composition. Direct observation of structure and morphology via transmission electron microscope and scanning electron microscope can differentiate EVs from impurity particles. Nanoparticle tracking analysis and dynamic light scattering enable the measurement and analysis of particle size and heterogeneity.1 Precise detection of biomarkers serves as the core basis for distinguishing EV subtypes and validating their functions. Flow cytometry and Western blotting can detect common EV surface markers such as the annexin family. Specific markers including CD9, CD63, CD81, TSG101, ALIX, and GAPDH are used to confirm EV identity.21,22 Additionally, microvesicle-specific markers such as integrins, selectins, and CD40L, which are associated with parental cell surface molecules, can reflect the cellular origin and targeting properties. Integrating multi-omics analyses further allows for the screening of therapeutic targets and diagnostic markers.23 Owing to the lack of unified molecular markers, the aforementioned methods are frequently used in combination and require the exclusion of serum albumin and lipoproteins to ensure the reliability of experimental results.
Differences and Connections in EV Characterization Methods are presented in Figure 1.
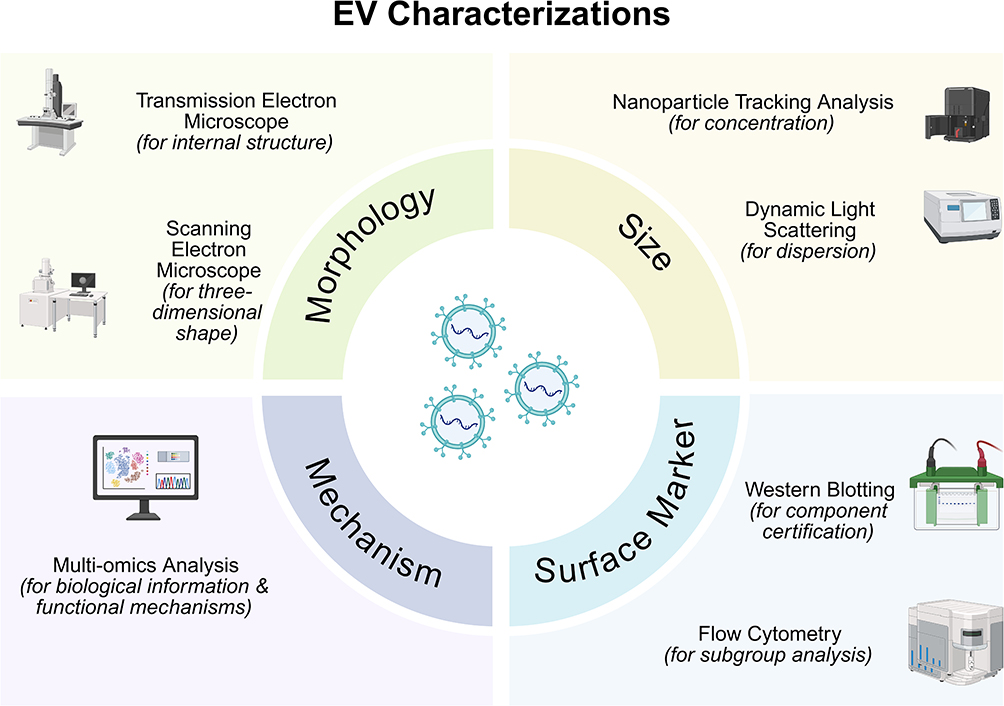 |
Figure 1 Differences and Connections in EV Characterization Methods. Transmission electron microscope (TEM) and scanning electron microscope (SEM) are both employed for the morphological observation of EVs. TEM provides the established morphological evidence for EVs, revealing characteristic features such as the “cup-shaped” or “double-concave disc-shaped” morphology and the lipid bilayer membrane structure. SEM enables the visualization of the three-dimensional architecture of EVs and is utilized to examine inter-particle interactions. Nanoparticle tracking analysis (NTA) and dynamic light scattering (DLS) are techniques for measuring EV size distributions; however, NTA is considered the gold standard for EV quantification, purity assessment, and the analysis of polydisperse samples, while DLS excels as a rapid tool for evaluating sample monodispersity and stability. Western blotting (WB) and flow cytometry (FCM) contribute jointly to establishing the identity and purity of EVs, with WB identifying specific EV protein markers and FCM facilitating subpopulation analysis. Multi-omics analysis, by uncovering biosynthetic pathways, elucidating sources of heterogeneity, discovering disease biomarkers, and revealing molecular mechanisms of action, serves as the core driver propelling future EV research from fundamental characterization toward functional elucidation and clinical application. |
EV Purification
Conventional EV isolation and purification methodologies exhibit inherent trade-offs between yield, purity, and biological integrity, necessitating method selection based on experimental priorities such as reproducibility, target specificity, and operational scalability to address the persistent challenge of balancing yield with purity in EV research.
Precipitation-based EV isolation employs polymers like polyethylene glycol (PEG) or salts to reduce biomolecule solubility and aggregate vesicles from biofluids, providing a cost-effective approach for large-scale preliminary enrichment. However, its limited purity typically necessitates supplementary purification via orthogonal techniques to achieve research-grade vesicle preparations.
Differential centrifugation (DC) employs sequential separation of cellular components through incrementally increased centrifugal forces and durations, initially pelleting high-density particles like nuclei and mitochondria, while retaining lower-density materials in supernatants for subsequent processing. Ultracentrifugation (UC) achieves nanoscale particle resolution through extreme rotational speeds (≥50,000 rpm), thereby generating centrifugal force (~100,000 × g) sufficient to discriminate particles by size and density. Standard protocols for high-purity EV isolation typically employ three steps: (i) remove cells and large debris via low-speed centrifugation (~200–1000 × g, 3–15 min); (ii) filter supernatant through 0.2–0.8 μm membranes to clear residual particles, and subsequently precipitate large cell particles by medium-speed centrifugation (10,000–12,000 × g, 30 min; ≥2 repeats); and (iii) transfer and collect exosome-like small vesicles by ultracentrifugation at (~70,000–100,000 × g, >70 min). The aforementioned steps require strict attention to the k-factor and rotor specifications.10,24–29 Further purification can be achieved through sucrose or iodixanol density gradients to isolate lipid-containing vesicles from contaminants such as proteins and protein-RNA aggregates.8,30–32 Differential ultracentrifugation integrates sequential centrifugation protocols that progressively escalate from low-speed separation of large cellular components to ultracentrifugation forces (100,000 × g) for membrane vesicle purification. This bridges DC and UC through calibrated, stepwise force application to isolate subcellular fractions by size-density parameters.33,34
Density gradient centrifugation (DGC) employs layered media (sucrose,35 cesium chloride,36 iodixanol37) to establish a buoyancy-based separation system where particles migrate under centrifugal force until reaching equilibrium at their intrinsic density zones, thereby enabling high-resolution isolation of EVs from co-isolated proteins or non-vesicular particles through density matching. This method minimizes mechanical shear stress while resolving more precise density variations among biological nanoparticles, outperforming DC and UC in both separation fidelity and particle integrity preservation, particularly for discriminating EV subpopulations from protein aggregates or lipoprotein contaminants based on characteristic buoyant density profiles. Studies have shown that the buoyant density of exosomes in sucrose gradients is 1.07–1.18 g/mL, while that of apoptotic bodies is 1.24–1.28 g/mL.29,38–40
Immunoaffinity separation leverages antigen-antibody specificity to isolate EVs through surface marker targeting, while employing immunoprecipitation for positive EV selection or immunodepletion for contaminant removal.41,42 This technique achieves superior purity relative to DC/UC/DGC by resolving EV subtypes via specific surface epitopes, though yield limitations persist due to the absence of pan-EV biomarkers for comprehensive isolation and inevitable non-specific binding during antibody-based capture.
Size exclusion chromatography (SEC) resolves EVs through hydrodynamic size partitioning using porous stationary phases, where large particles elute first via interstitial flow while smaller molecules experience delayed migration through gel matrix pores. Comparative analyses by Chandrasekera et al demonstrate that precipitation favors high EV yields but introduces copious contaminants, whereas SEC maintains superior purity, though constrained by limited throughput at elevated sample volumes. They suggest that protocols integrating precipitation-based preconcentration with subsequent SEC purification enable high-purity EV recovery from low-abundance biofluids through synergistic yield-purity optimization.43
Ultrafiltration employs semi-permeable membranes under pressure-driven flow to retain EVs while permeating smaller solutes, which achieves EV enrichment through size-based exclusion. However, this method’s high recovery efficiency is countered by copious co-concentrated contaminants that compromise EV integrity and experimental reproducibility through nonspecific protein aggregation and vesicle deformation.44
Current empirical evidence confirms that integrated isolation protocols synergizing complementary separation principles, such as UC, precipitation, and SEC, demonstrate superior efficacy in EV purification by balancing yield-purity tradeoffs inherent to single-method approaches.45 Comparative analyses by Otani et al demonstrated that PEG isolation yields more contaminated products, UC produces lower yields, whereas PEG precipitation coupled with UC outperformed individual PEG or UC methods in both yield and purity for EV isolation from rat plasma.46 Stam et al further reported that 60% of current EV isolation protocols employ combinatorial approaches. They systematically validated SEC initiation followed by low-speed centrifugation as an optimal strategy for achieving maximal purity while maintaining functional EV yields in sequential purification workflows.47
Summaries of the EV separation and purification methods have been showed in Table 1.
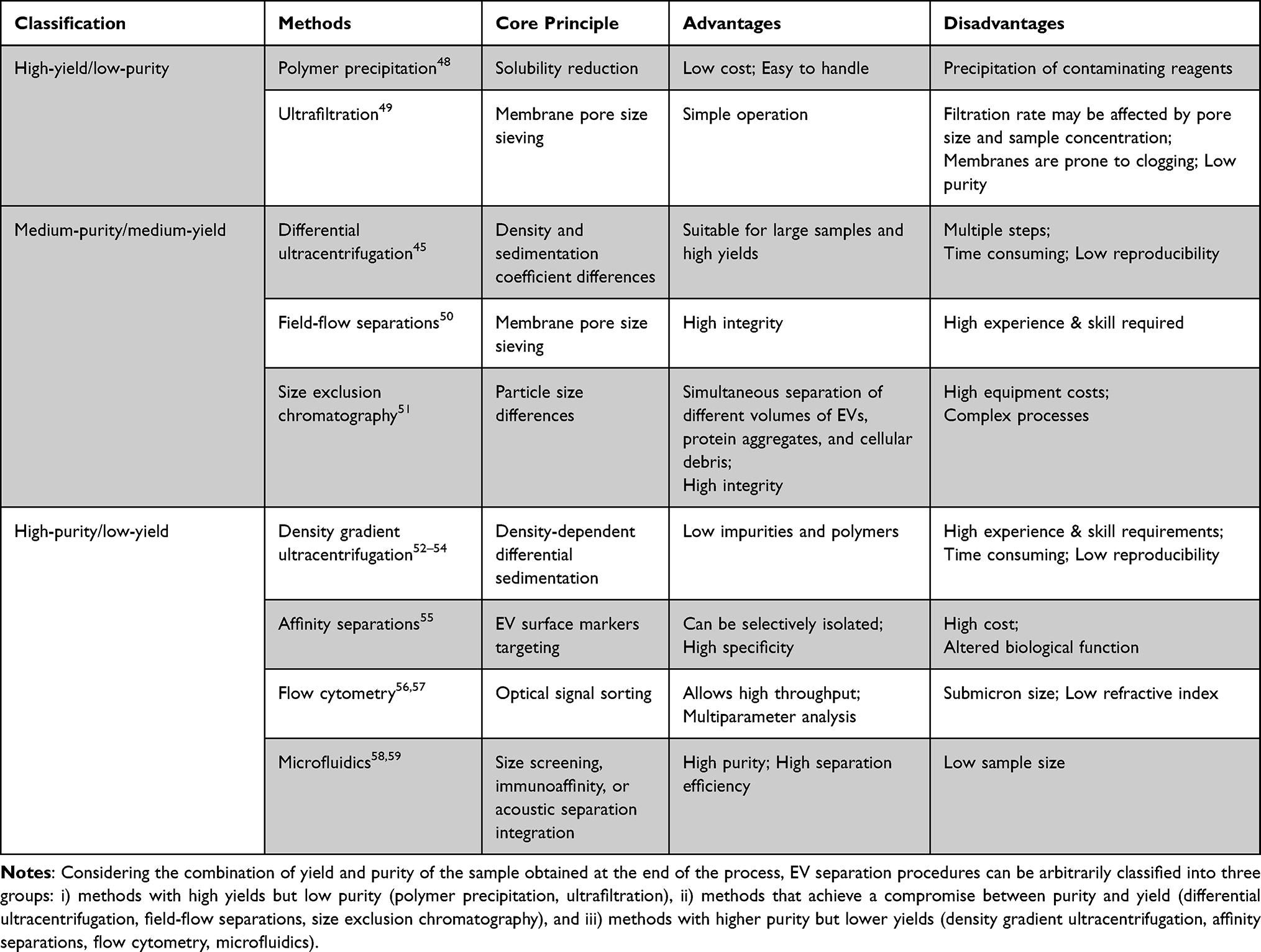 |
Table 1 Summary of EV Separation and Purification Methods |
Engineered EVs
The use of EVs as drug carriers faces challenges including suboptimal cellular targeting specificity, inefficient encapsulation of therapeutic cargoes during donor cell culture, and limited bioavailability of delivered payloads in recipient cells.60 These limitations can be addressed through engineered EV modifications that combine surface functionalization with optimization of cargo composition.
Classification of EV engineering approaches is shown in Figure 2.
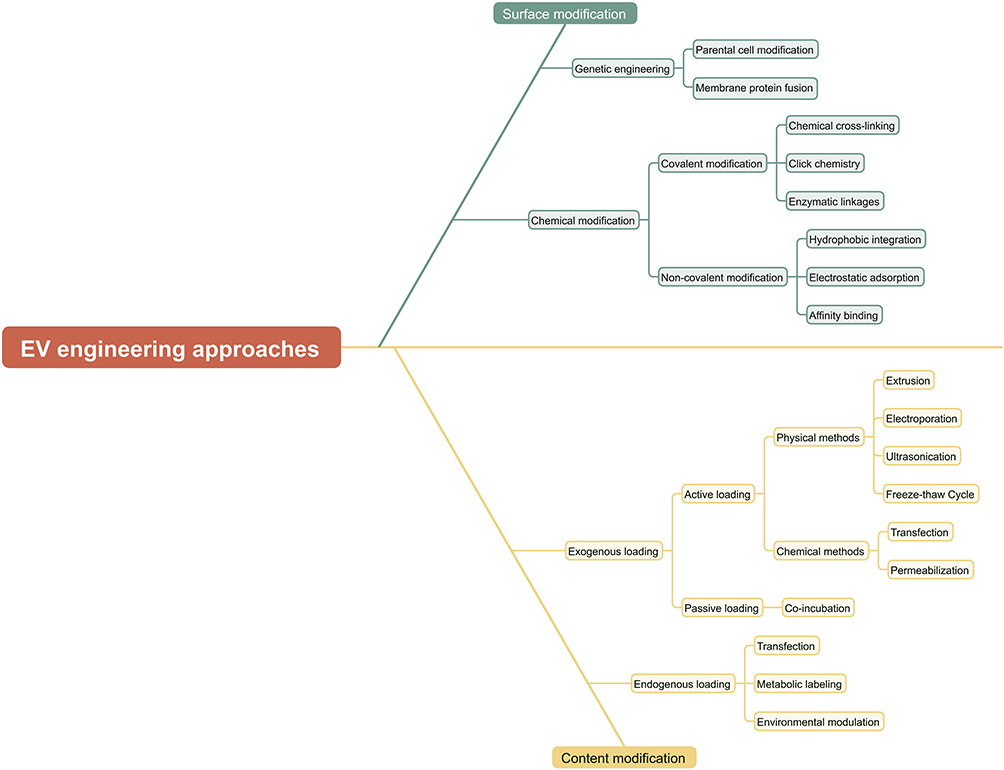 |
Figure 2 Classification of EV engineering approaches. EV modification can be classified into surface modification and cargo modification: surface modification can be further divided into genetic engineering modification and chemical modification based on the modification principle. Among them, chemical modification can be classified into covalent modification and non-covalent modification according to the method of chemical bond formation. Cargo modification can be divided into endogenous loading and exogenous loading based on the modification subject. Exogenous loading consists of active loading and passive loading. Different types of cargos are suitable for different loading methods. |
Surface Modification—Improvement of Targeting
Surface modification involves adding targeting ligands to the surface of EVs through genetic engineering and chemical modification.
Genetic Engineering Methods
Genetic engineering methods for EV surface modification, including parental cell modification61 and membrane protein fusion,62 involve attaching gene sequences that encode guiding proteins or peptides to specific EV surface membrane proteins. Parental cell modification employs genetic engineering to overexpress fusion genes combining transmembrane proteins (eg Lamp2b, PDGFR)63 with targeting peptides or proteins including neuron-targeting RVG peptides, tumor-targeting iRGD peptides,64 and chondrocyte-targeting CAP peptides.65 Membrane protein fusion entails the genetic fusion of natural EV membrane proteins with functional proteins such as luciferase or antibody fragments, which are frequently applied for imaging purposes (eg Gaussia luciferase)66 or immunomodulatory functions (eg PD-1 fusion proteins).67 A key limitation lies in its restriction to genetically encodable targeting motifs present on peptide or protein surfaces and may be degraded by endosomal proteases inside the producing cell. To solve this problem, Hung et al engineered an RVG variant by adding a glycosylation motif. This modification both prevented peptide degradation and enhanced EV delivery to neuroblastoma cells, contributing to the development of novel sEV-based therapeutics.68
Chemical Modification
Covalent Modification
Covalent modification involves the permanent chemical bonding of functional molecules (proteins,69 amines,70 fluorescent dyes71,72) to reactive surface groups on EVs through three primary approaches: (i) Chemical cross-linking (eg1-ethyl-3-(3-(dimethylamino)propyl) carbodiimide (EDC)/N-hydroxysuccinimide (NHS)-mediated amination reaction73 and thiol–maleimide coupling for cysteine residues on the EV surface), (ii) Click chemistry (eg copper-catalyzed azidoalkynyne cycloaddition; strain-promoted azide-alkyne cycloaddition;73 bioorthogonal click chemistry reactions, etc) and (iii) Enzymatic linkages (eg protein ligating enzymes).74
Non-Covalent Modification
Non-covalent modification involves transient molecular interactions (hydrophobic integration, electrostatic adsorption, and affinity binding)75 with EV membranes. Hydrophobic integration utilizes lipid-conjugated molecules like PEG for membrane insertion to enhance stability, prolong circulation half-life, and reduce immunogenicity;76,77 electrostatic adsorption leverages cationic polymer-anionic EV membrane charge interactions;78 and affinity binding includes streptavidin-biotin pairing and antibody-epitope recognition.79 While offering rapid implementation, minimal membrane disruption, and reversible dynamics responsive to environmental stimuli, these approaches exhibit limited stability, making them suitable for only short-term applications or stimuli-responsive drug release. Sherif E. et al demonstrated that sEV PEGylation enhances in vivo circulation duration and plasma concentration. However, subsequent immune monitoring revealed that intravenous administration induced T cell-dependent anti-PEG IgM production can accelerate the clearance of subsequently administered PEGylated sEVs and liposomes, but selectively spare ovalbumin from immune-mediated elimination.80
Modifying EV Content—Improving Treatment Efficiency
Engineering methods for EV content modification can be classified into two categories: endogenous loading and exogenous loading.
Endogenous Loading
Endogenous loading employs genetic engineering of parent cells to facilitate natural encapsulation of therapeutic cargo into EVs during biogenesis. This endogenous approach primarily utilizes three strategic mechanisms: (i) genetic engineering via transfection with gene-editing vectors (plasmids,81 viral vectors82) to drive active packaging of target biomolecules; (ii) metabolic labeling through culture medium supplementation with bio-orthogonal precursor molecules (eg dextran sulfate)83 for subsequent incorporation into EV cargos; and (iii) environmental modulation using hypoxia, inflammatory cytokines, or pharmaceutical agents to stimulate EV production with enhanced cargo-loading capacity.84 Mizrak et al first demonstrated cancer treatment using genetically engineered lEVs that delivered therapeutic mRNA/protein by engineering donor cells to overexpress the suicide gene mRNA and cytosine deaminase-uracil phosphoribosyltransferase fusion protein. Consequently, this achieved significant growth suppression in malignant peripheral nerve sheath tumors.11 This method preserves EVs’ natural structure and biocompatibility but exhibits suboptimal cargo-loading efficiency, limited control over payload quantification, and potential donor cell physiological perturbations that may elevate EV heterogeneity.
Exogenous Loading
Exogenous loading is the process of introducing therapeutic agents into purified EVs. This methodology comprises two distinct strategies: active loading employs physical/chemical methods to induce cargo internalization through energy-dependent mechanisms; passive loading relies on molecules’ intrinsic physicochemical properties for spontaneous transmembrane diffusion without external energy input.
Active Loading
Physical Methods
Extrusion-based engineering involves pressurized passage of drug19,85-EV mixtures through nanoporous membranes,86 which induces transient membrane destabilization followed by vesicle reformation. These synthetic constructs closely mimic natural EVs in key characteristics: size distribution, morphological features, and biomolecular composition,87 while also supporting industrial-grade EV therapeutic production. However, it also causes changes in particle size distribution and membrane potentials.88
Electroporation utilizes brief high-voltage electrical pulses to generate transient nanopores in EV membranes, thereby enabling efficient encapsulation of electrically charged molecules like hereditary materials.89 Wahlgren et al compared chemical transfection versus electroporation for siRNA loading into sEVs Chemical transfection generated contaminating MAPK-1 siRNA-containing micelles inseparable from sEVs, demonstrating its genetic material inapplicability. Thereafter, they used immunoblotting and flow cytometry to examine whether electroporation was the only means of introducing genetic material into sEV, and found that the exogenous siRNA could only be detected in the electroporated exosome samples.90 Electroporation significantly improves loading efficiency up to 30%20,91 compared to passive diffusion and ultrasonication, extrusion, or saponin permeabilization.88,92 However, Sander et al identified key technical limitations: during electroporation, multivalent ions bind to EV membranes while buffer conductivity shifts occur, thereby collectively promoting siRNA aggregation. This phenomenon artificially inflates measured loading efficiency through false-positive signal amplification.93
Ultrasonication disrupts EV membrane integrity through cavitation effects, where collapsing gas bubbles generate shear forces and microjets to create transient nanopores. These pores facilitate efficient transmembrane entry of lipophilic small molecules94,95 and proteins, while achieving incorporation rates of 25%–35%.96 Notably, nucleic acid loading efficiency via ultrasonication remains inferior to electroporation.92,97 Studies have shown that ultrasound treatment with 20% amplitude, six cycles of 30 s on/off, and a 2-minute cooling interval between cycles enables a paclitaxel loading efficiency of 28.29 ± 1.38% in macrophage-derived exosomes. This drug-loaded system exhibits a kinetic profile characterized by an initial burst release within the first 3 hours, followed by sustained release thereafter. After one week, approximately 30% of the drug remains associated with the exosomes, and the system can be stably stored in aqueous solution for up to one month.88
Freeze-thaw cycling facilitates drug encapsulation by mixing therapeutic cargo with EVs at room temperature, followed by 3 repeated cycles of flash freezing (−80°C or liquid nitrogen) and thawing (room temperature98), exploiting ice crystal formation to disrupt plasma membranes.99 This process preserves the bioactivity of protein, RNA,100 and small-molecules,101 but the associated structural alterations risk EV aggregation and polydisperse size distributions, and its loading efficiency remains consistently lower than sonication or extrusion approaches. Regrettably, there are currently no direct quantitative data on the loading efficiency for biomolecules. Efficiency evaluation predominantly relies on comparative analysis with other methods and inference from membrane fusion efficiency.77
Chemical Methods
Calcium chloride (CaCl2)-mediated transfection leverages CaCl2-DNA precipitation complexes to enhance DNA-cell surface interactions by facilitating cellular endocytosis. For sEV-based miRNA delivery, Zhang et al adapted this technique by incorporating heat shock treatment to increase membrane fluidity, thereby enabling sEVs’ uptake of CaCl2-miRNA complexes at ambient temperatures. Comparative analysis revealed this modified approach achieves miRNA loading efficiency comparable to electroporation while eliminating dependence on specialized instrumentation.102
Chemical-mediated EV membrane permeabilization utilizes cholesterol-selective binding reagents to create transient pores for hydrophobic compounds loading. This approach employs two primary reagent categories: organic solvents (eg methanol,103 acetone104) to dissolve membrane components and detergent agents (eg saponins) to complex with membrane cholesterol for inducing controlled porosity. Saponin-based permeabilization is clinically preferred for loading therapeutic proteins into EVs19,105 and significantly enhances cargo-loading capacity compared to incubation techniques.19,106
Passive Loading
Passive EV drug loading via room temperature co-incubation utilizes concentration gradients and molecular hydrophobicity. This approach demonstrates cargo incorporation efficiency proportional to either exogenous substance concentration or lipophilicity. It shows optimized performance for lipophilic chemotherapeutics99,107,108 and anti-inflammatory agents,109 which exhibit enhanced EV loading capacity compared to synthetic liposomes. However, systematic evaluation of five loading strategies (passive incubation, saponin permeabilization, freeze-thaw cycles, ultrasonication, and extrusion) for oxidative enzyme encapsulation revealed that passive loading exhibits the lowest efficiency compared to other active loading methods.
Comparative studies systematically evaluate EV engineering methodologies to balance critical parameters (yield, loading efficiency), with Haney et al demonstrating superior TPP1 protein loading in macrophage-derived sEVs via ultrasonication (70 μg/1011 sEVs) compared to saponin permeabilization (50 μg/1011 sEVs) and transfection (10 μg/1011 sEVs). This establishes that sonication is the optimal strategy for therapeutic cargo encapsulation.105 Ultrasonication and extrusion induce structural reorganization of EVs, thereby enabling peroxidase diffusion through densely packed lipid bilayers. Shao et al observed that ultrasonication and extrusion enhance loading efficacy but risk vesicle aggregation and morphological alterations. In contrast, saponin permeabilization preserves native sEV morphology. Based on these findings, they hypothesized that combining extrusion with saponin permeabilization might synergize these advantages to enhance protein encapsulation efficacy. Their comparative study of SOD-loading strategies (saponin permeabilization, extrusion, saponin-extrusion hybrid) revealed via Western blotting analysis that the combined approach achieved maximal SOD incorporation in sEVs, validating the hypothesis of technique synergy.110 Fuhrmann et al systematically evaluated exogenous drug encapsulation strategies using hydrophobicity-varied porphyrins as model cargoes. They showed that, while EV loading efficiency primarily depends on cargo hydrophobicity, active methods like saponin permeabilization and electroporation substantially enhanced EV incorporation of moderately hydrophobic porphyrins. Particularly, saponin treatment achieved an 11-fold enhancement in hydrophilic porphyrin encapsulation while preserving vesicle delivery competence.11
EV Mimetics
According to MISEV2023 guidelines, EV mimetics encompass two subtypes: ACDVs generated through artificial cellular disruption and SVs created via molecular component synthesis or hybridization. Current preparation strategies fall into three methodological categories, namely top-down, bottom-up, and bio-hybridization approaches.111
Top-down strategies involve disintegrating complex macroscopic units like cells into nanoscale components, and these tiny membrane fragments then self-assemble to produce nanovesicle structures.112 Apart from intact cells, lysed cell membranes113 and EV membranes114 can also be used as raw materials for coating nanoparticles. As a result, top-down-derived vesicles preserve native cellular components during membrane-to-nanovesicle transition, thereby replicating natural EV biocomplexity. Top-down techniques include chemical-induced cell blebbing, sonication nitrogen cavitation, extrusion, filtration, and microfluidic devices. Among them, extrusion is the most widely used strategy, and the yields of EV mimetics can be increased by about 100-fold compared to natural sEVs through extrusion.85 Additionally, nanoparticles can acquire functionalized membrane coatings through metabolic engineering, lipid insertion, membrane hybridization, or genetic modification.115 As their membranes originate directly from cells, these top-down vesicles are classified as ACDVs.
In contrast to top-down methods, the bottom-up approach constructs EV mimetics de novo using cell-derived or synthetic biomolecules, while engineering them into structurally and functionally defined architectures without employing whole cells. These EV-mimetic lipids (eg functionalized liposomes116) are engineered via surface modification techniques such as incubation, bioconjugation, and cell-free protein synthesis to generate diverse structures: peptide/antibody-conjugated liposomes, membrane protein-modified vesicles, protein-embedded nanoparticles, or synthesized nanovesicles.117–119 Though superior for large-scale vesicle production over top-down strategies, the bottom-up approach faces challenges in achieving stable EV mimetics due to significant compositional disparities from natural EVs.
Biohybrid methodologies have emerged to integrate EVs or ACDVs with synthetic nanoparticles via membrane fusion techniques, including freeze-thaw cycling, co-extrusion, incubation, ultrasonication, and film hydration, bridging the technical divide between bottom-up and top-down strategies while enhancing EV mimetic drug-loading capacity and delivery efficiency. This approach strategically merges synthetic nanocarriers’ operational strengths (ease of manufacturing, scalable production, precise process control, and high drug encapsulation rates)120,121 with native membrane components’ biological advantages (low immunogenicity, enhanced biocompatibility and safety, sustained circulatory stability, tissue-targeting specificity, and biological barrier penetration capabilities)122–124 to ultimately generate optimized SVs that combine engineered practicality with innate biological performance.
Current research on EV mimetics faces underdeveloped evaluation standards with inconsistent nomenclature, classification, production, and quality control. In alignment with MISEV2023 guidelines, this study categorically terms all ACDVs and SVs as EV mimetics to ensure terminological precision.
Clinical Applications
With the ongoing exploration and optimization of EV preparation technologies, their inherent advantages—such as biocompatibility, targeted delivery capability, and the capacity to transport bioactive molecules—are being progressively harnessed. This has provided crucial technical support for translating EVs into clinical diagnostic and therapeutic applications and has further propelled in-depth research related to their clinical utilization.
Natural EVs
Natural EVs originate from diverse cellular sources exhibiting unique functional properties. This prioritizes the strategic selection of cell-derived vesicles for targeted therapeutic applications and optimization of scalable production processes with enhanced drug delivery efficiency in EV-based biomedical development.
Clinically and experimentally utilized EVs predominantly originate from mesenchymal stem cells (MSCs) derived from adipose, hepatic, bone marrow, umbilical cord blood, and so on.125–128 These MSC-EVs inherit functional attributes from parental cells through their protein or RNA cargo, while retaining core MSC functionalities: (i) tissue protection and regeneration: MSC-EVs display universal EV markers (CD9, CD63, CD81), MSC-specific identifiers (CD44, CD73, CD90),129 and adhesion molecules (CD29, CD59, CD166)130 enabling targeted homing to injury sites.131 They mitigate tissue damage by suppressing apoptosis (caspase-3/8/9 inhibition)132,133 and pyroptosis (caspase-1 modulation),134 while secreting cytokines, inflammatory mediators, extracellular matrix components, and antimicrobial proteins via paracrine signaling to establish pro-regenerative microenvironments, which are demonstrated in post-trauma repair in cardiac,135 hepatic,136 renal,137 and neural138 cells. Exosomes derived from menstrual blood mesenchymal stem cells effectively suppress endothelial inflammation and advanced atherosclerosis by inhibiting the miR-574-5p-mediated c-Rel/NF-κB pathway, offering new insights into stem cell therapy for inflammatory atherosclerosis.139 Conversely, miRNA-15a-5p secreted by synovial fibroblast-derived extracellular vesicles has been identified as a key signaling molecule mediating chondrocyte apoptosis, playing a critical role in promoting chondrocyte death.140 Gibello et al pioneered the use of autologous serum-derived extracellular vesicles, applying six repeated topical doses over a two-week period to four patients with chronic venous ulcers. After a 30-day treatment period, follow-up evaluations revealed that, compared to standard therapy, EV treatment led to a significant reduction in ulcer area, accompanied by increased fibrosis and microvascular proliferation at the lesion sites.141 (ii) Immunomodulation: through transfer of regulatory miRNAs or proteins, MSC-EVs reprogram inflammatory immune cells (M1 macrophages, dendritic cells, Th1, Th17) into immunosuppressive phenotypes (M2 macrophages, tolerogenic dendritic cells, Tregs) This shift suppresses pro-inflammatory mediators (eg TNF-α, CCL-17, CCL-24) while elevating anti-inflammatory cytokines (eg IL-10) and TGF-β, collectively alleviating inflammation and accelerating tissue regeneration142,143 A Phase I clinical trial conducted by Dai et al combined sEVs derived from ascites fluid with granulocyte-macrophage colony-stimulating factor for colorectal cancer immunotherapy Results demonstrated that this combination therapy is both feasible and safe, providing an alternative immunotherapeutic option for advanced colorectal cancer.144
The next most commonly used human embryonic kidney cell line, HEK293T-derived EVs, demonstrates mass production and cultivation advantages with enhanced genetic engineering potential through plasmid transfection or targeted nucleic acid modifications for precise molecular customization. While immune cell-derived EVs demonstrate functional duality: dendritic cell EVs preserve parental surface signaling molecules to activate antitumor immunity through dendritic cell-dependent and independent mechanisms;145 T-cell EVs exert immunostimulatory effects, and NK-cell EVs deploy cytotoxic molecules (eg FasL, perforin) to induce tumor apoptosis.146 Tumor-derived EVs, bearing tumor-specific antigens that provoke immune activation,147 serve as key components in engineered antitumor EV therapeutics.
Engineered EVs
Surface Modification
Surface-modified EVs demonstrate enhanced targeting precision for delivering therapeutic cargo to hierarchical tissue structures in vivo. Genetic engineering facilitates precise display of tumor-targeting antibody fragments (scFv) on EV surfaces. Illustrating this approach, Chen et al engineered a scaffold protein with dual-displays of αPD-L1 and αCD3 scFv fragments on HEK293T-derived EV membranes. These bispecific αPD-L1 × αCD3 BiTExos mediate T-cell redirection to PD-L1+ tumor cells through dual antigen recognition, simultaneously activating T-cell proliferation and cytotoxic effector functions while blocking PD-1/PD-L1 immunosuppressive signaling to enhance antitumor immunity.148 Genetically engineered EVs demonstrate enhanced therapeutic precision, as exemplified by Gao et al’s PSMAscFv-EVN-GSDMD platform. This system combines a prostate-specific membrane antigen-targeting scFv with the pyroptosis-inducing N-terminal domain of gasdermin D, which synergistically activates antitumor immunity as well as pyroptosis-mediated immunogenic cell death in prostate cancer models.149 Wiklander et al developed Fc-functionalized EVs as modular delivery vehicles. By incubating these Fc-EVs with HER2 monoclonal antibody trastuzumab, they demonstrated enhanced tumor-targeting efficacy. This significantly increased cellular uptake in HER2-positive breast cancer cells while maintaining therapeutic payload integrity.150
Surface-engineered EVs demonstrate organ-specific delivery capabilities through advanced bioconjugation techniques. Luo et al achieved spinal cord targeting by conjugating CAQK homing peptides to neural stem cell-derived sEVs via copper-free click chemistry, subsequently electroporating CCL2-specific siRNAs into these vesicles for precise delivery to spinal cord injury sites, thus, effectively enhancing neural repair outcomes. Simultaneously, Lu et al implemented a bio-orthogonal strategy where MSCs were metabolically labeled with N-azidoacetyl-mannosamine to generate azide-modified sEVs. These azide-modified sEVs were then covalently coupled with dibenzocyclooctyne-functionalized targeting ligands through strain-promoted click reactions, thereby creating chimeric antigen receptor-expressing sEVs for liver-directed therapies.151 Lv et al modified the surface of M2 macrophage-derived, virus macrophage inflammatory protein-II-rich EVs with a cardiac-targeting peptide and platelet membrane. This approach synergistically enhanced the vesicles’ targeting specificity toward myocardial tissue and their endocytic efficiency, thereby reprogramming M1 pro-inflammatory macrophages into an M2 anti-inflammatory phenotype, providing a novel immunomodulatory therapeutic paradigm for addressing excessive inflammatory responses in viral myocarditis.152
Engineered EVs demonstrate remarkable capabilities in crossing the blood-brain barrier (BBB) for targeted therapeutic delivery to specific brain regions. Alvarez et al transfected Lamp2b into the dendritic cells through plasmid transfection and created chimeric variants fused with RVG for neural targeting and muscle-specific peptide (MSP) for myocyte homing. The resultant MSP-Lamp2b and RVG-Lamp2b EVs exhibited cell-type-specific tropism in mice, with RVG-modified EVs achieving neuronal targeting and MSP-EVs showing muscle specificity. This culminated in a significant reduction of β-amyloid 1–42 levels in murine models, thereby validating EV-based strategies for neurodegenerative disease intervention.20
Cargo Loading
EVs exhibit remarkable versatility as delivery platforms capable of co-loading diverse therapeutic cargoes spanning small-molecule drugs, nucleic acids, and proteins. Small-molecule payloads enable cellular proliferation inhibition,153,154 signaling pathway modulation,155,156 and targeted cytotoxicity.157 Nucleic acid cargos include protein-coding plasmids or mRNAs for exogenous gene expression and regulatory RNAs for interference. Protein therapeutics, whether native or engineered, function intracellularly to activate or suppress specific biological pathways.158 However, these biomacromolecules inherently lack cell membrane permeability,159,160 which necessitates EV-mediated delivery to achieve intracellular bioactivity.
Small Molecule Drugs
EVs exhibit intrinsic therapeutic advantages including low immunogenicity, minimal cytotoxicity, enhanced bioavailability, and inherent targeting specificity.122,123,161,162 Small molecule therapeutics, particularly chemotherapeutic agents, often demonstrate suboptimal pharmacokinetics characterized by non-specific biodistribution, rapid systemic clearance, limited aqueous solubility, and toxicity.163 EV encapsulation of these pharmaceuticals enables engineered nanoformulations that enhance the therapeutic index through improved target tissue accumulation while mitigating adverse effects via reduced effective dosage requirements.
Therapeutics of engineered EV loaded with small-molecule have been showed in Table 2.
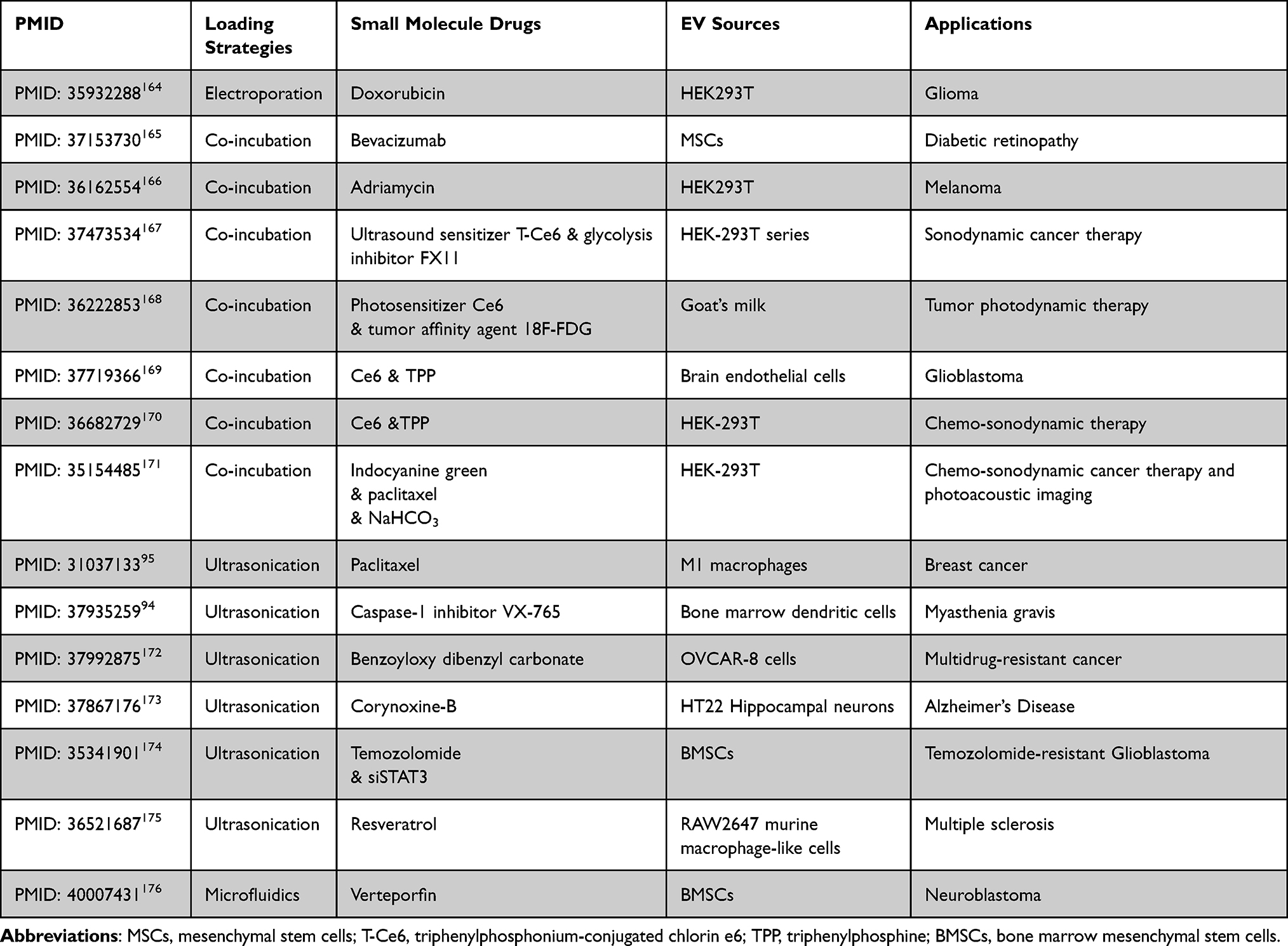 |
Table 2 Engineered EV Loaded with Small-Molecule Therapeutics: Comparative Analysis of Loading Strategies, Physicochemical Properties, EV Sources, and Pharmacological Applications in Drug Delivery |
Protein Cargoes
Protein-based therapeutics, including recombinant proteins,177 monoclonal antibodies,178 and protein kinase,179 are prone to compromised efficacy due to immune recognition, rapid systemic clearance, enzymatic degradation, or off-target molecular sequestration180 Engineered EVs address these limitations by enhancing proteolytic stability through lipid bilayer encapsulation, prolonging circulation half-life via reduced immunogenicity, and enabling receptor-specific targeting through surface modifications.181 This DDS synergistically optimizes pharmacological performance by preserving structural integrity, enhancing cellular uptake efficiency, and minimizing nonspecific interactions, thereby amplifying therapeutic outcomes in protein-based interventions.182
Therapeutics of engineered EV loaded with protein have been showed in Table 3.
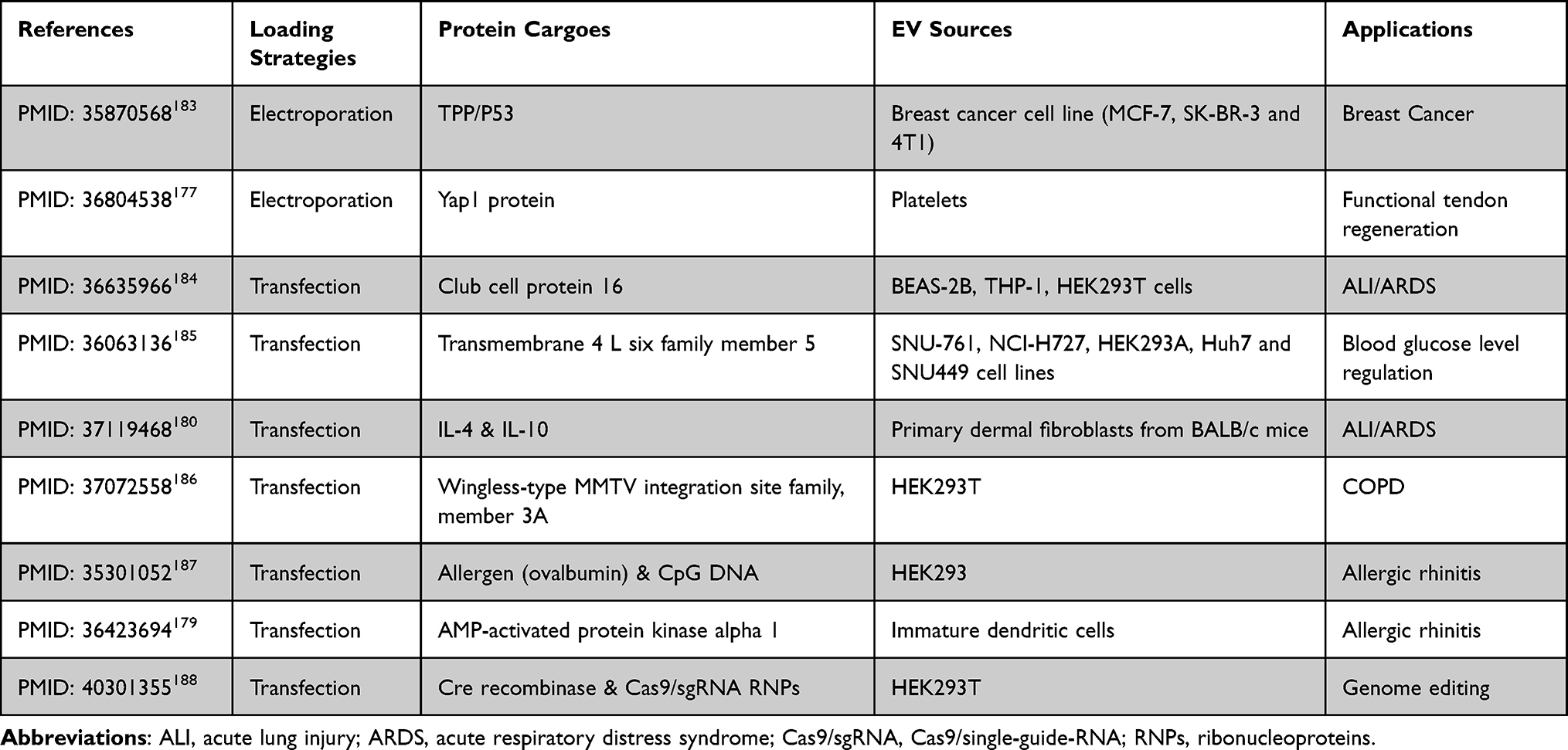 |
Table 3 Engineered EV Loaded with Protein Therapeutics: Comparative Analysis of Loading Strategies, Physicochemical Properties, EV Sources, and Pharmacological Applications in Drug Delivery |
Nucleic Acid-Based Cargoes
siRNA, miRNA, and antisense oligonucleotides (ASOs) demonstrate particular utility in organ repair research. However, these polyanionic biomolecules face intrinsic pharmacological challenges due to their cell membrane-impermeable nature, susceptibility to nuclease degradation, and electrostatic repulsion.189 This necessitates engineered delivery platforms to ensure intracellular bioavailability and therapeutic functionality.
Therapeutics of engineered EV loaded with nucleic acid have been showed in Table 4.
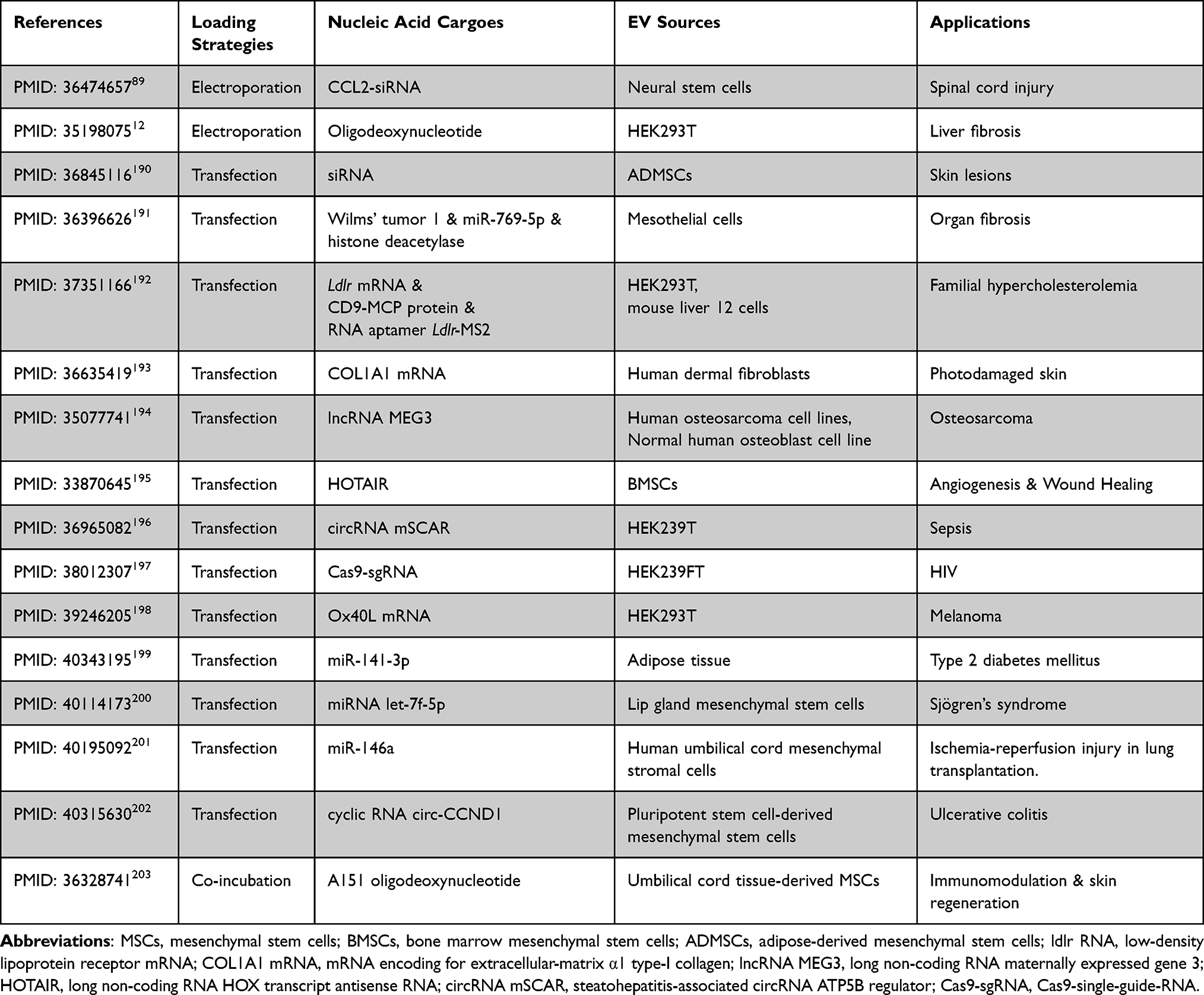 |
Table 4 Engineered EV Loaded with Nucleic Acid Therapeutics: Comparative Analysis of Loading Strategies, Physicochemical Properties, EV Sources, and Pharmacological Applications in Drug Delivery |
EV Mimetics
Although EV mimetics remain under-investigated in clinical trials, they address critical limitations of natural EVs, notably batch-to-batch heterogeneity204 and low yield.205 Through adaptable manufacturing of ACDVs via extrusion, these synthetic systems achieve parental cell-mimetic biological properties with enhanced uniformity in vesicle size, membrane architecture, and cargo composition, which demonstrates significant translational potential for industrial-scale EV-based DDSs. Concurrently, hybrid SVs engineered through the strategic integration of EV components with synthetic nanoparticles are emerging as modular platforms for precision therapeutic targeting as they combine endogenous biocompatibility with engineered functionalization capabilities.
Therapeutics of EV mimetics have been showed in Table 5.
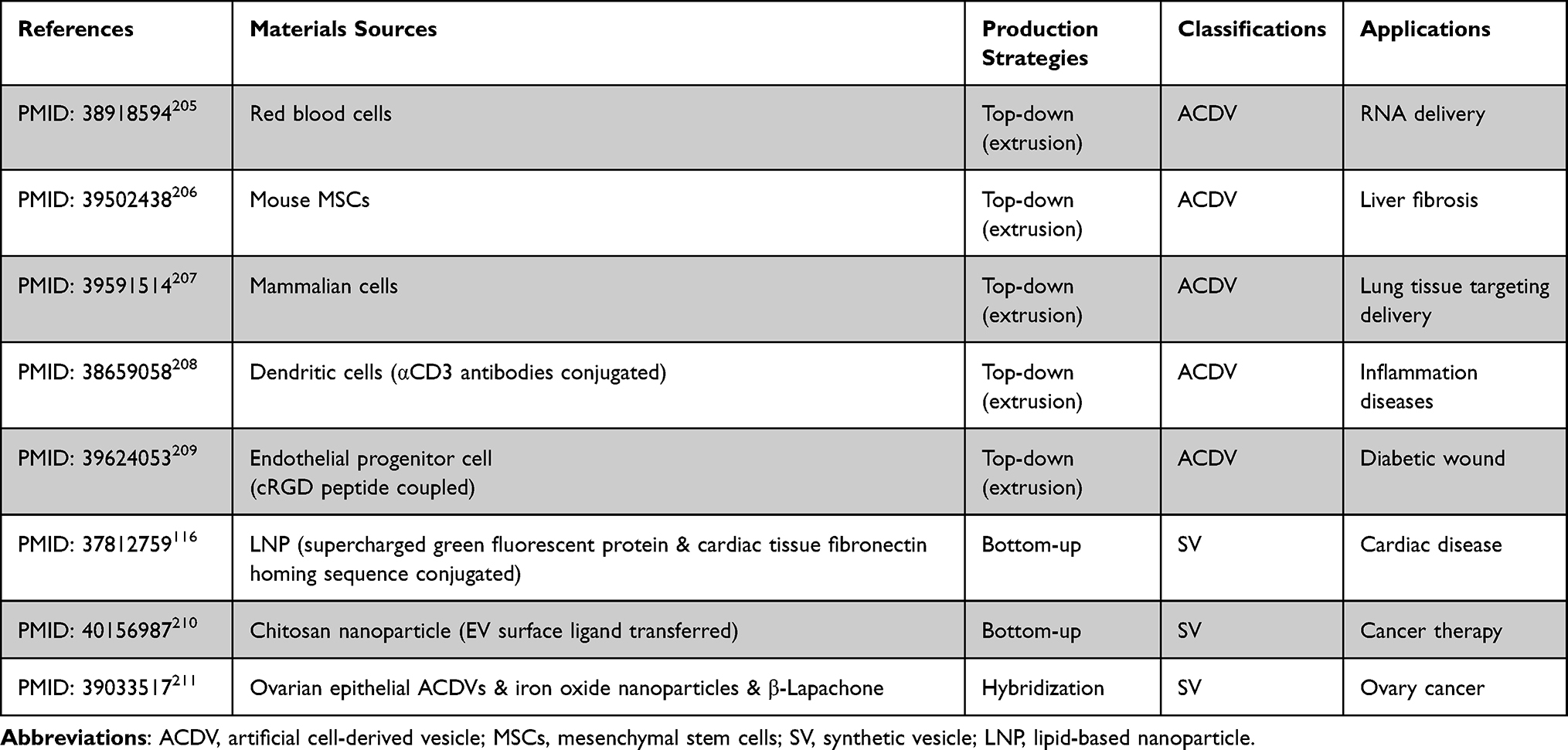 |
Table 5 EV Mimetics Therapeutics: Comparative Analysis of Material Sources, Production Strategies, Classifications, and Pharmacological Applications in Drug Delivery |
Other Synthetic Nanoparticles
Conventional synthetic nanocarriers currently dominate clinical applications as drug delivery vehicles for proteins, small-molecule therapeutics, and other payloads. Thus, they share functional parallels with engineered EVs in target specificity, payload optimization, and mitigation of nonspecific tissue toxicity, which makes direct comparisons contextually redundant.212 This review specifically addresses the transformative therapeutic potential in gene therapy by synthetic nanoparticles, which is yet underexploited in engineered EVs.
Gene therapy employs nucleic acid-based interventions to modulate gene expression or cellular function through four principal strategies: (i) gene replacement, through functional gene delivery to compensate for defective alleles; (ii) gene silencing, via RNA-based regulators213,214 to suppress aberrant gene expression; (iii) gene addition, utilizing DNA/RNA constructs215–217 to introduce ectopic gene expression; and (iv) gene editing, deploying CRISPR-Cas systems218,219 to induce precise genomic modifications for functional gain or loss. The macromolecular size and polyanionic nature of these therapeutic agents necessitate vectors to overcome cellular membrane impermeability and ensure intracellular delivery efficacy.
Classifications of synthetic nanoparticles have been showed in Table 6.
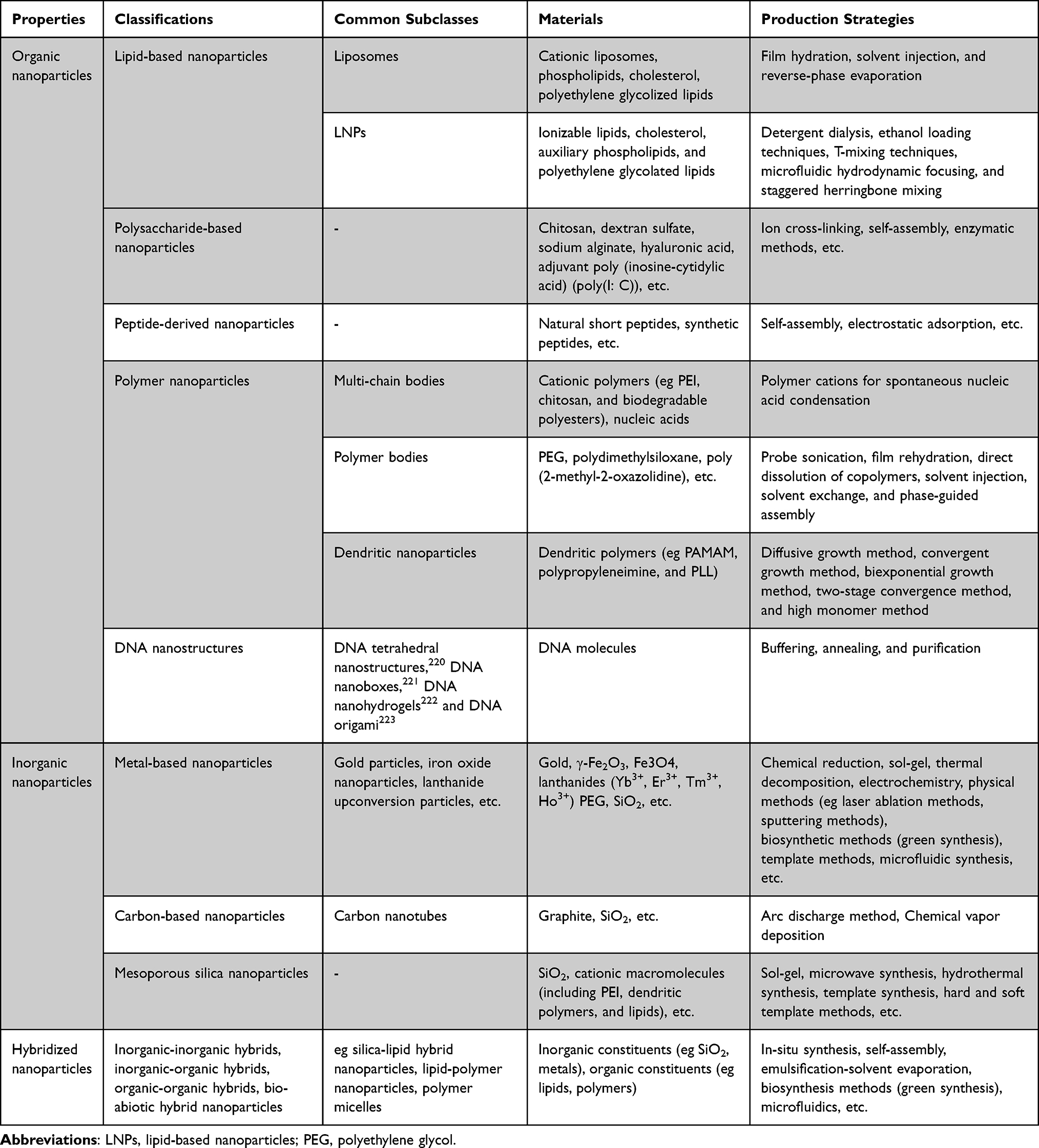 |
Table 6 Classifications of Synthetic Nanoparticles: Sorted by Their Subclasses, Materials, and Production Strategies |
Gene Editing
Gene editing is a biotechnology for modifying genetic information through insertion, deletion, replacement, or modification of specific genomic sequences.224 CRISPR-based strategies now dominate clinical applications owing to their programmable DNA targeting capability, which facilitates permanent genomic edits and donor template integration. These advantages have enabled CRISPR’s utilization in treating genetic disorders,225 developing cancer immunotherapies,226 and advancing antiviral treatments.227 The CRISPR-Cas9 system requires two components: (i) Cas9 endonuclease, for inducing double-strand DNA breaks, and (ii) single-guide RNAs (sgRNAs), for sequence-specific targeting. Synthetic nanoparticles are increasingly leveraged as non-viral delivery platforms for CRISPR-Cas9 payloads, thus enhancing transfection efficiency and tissue specificity while minimizing CRISPR-based off-target genomic perturbations.
Zhao et al engineered an oral CRISPR-Cas9 delivery platform using amphiphilic ion-polysaccharide polymer-coated nanocomplexes to disrupt the tumor necrosis factor receptor-associated protein 1 gene in colorectal cancer. They induced mitochondrial dysfunction in tumor cells, synergistically amplifying chemotherapy-driven necrotic cell death and antitumor immunity.228 It also achieved efficient in vivo gene editing in Huntington’s disease models.229 Liyanage et al engineered a CRISPR-Cas9 delivery platform by covalently conjugating SpCas9 nuclease to hydroxyl-terminated PAMAM dendrimers via high-specificity inverse electron-demand Diels-Alder click chemistry, thus constructing a Cas9-dendrimer nanocomplex (D-Cas9) self-assembling with sgRNA into a neutrally charged D-Cas9-RNP. This system circumvented nuclear delivery challenges of large nucleic acids, including nuclease susceptibility, inefficient packaging, and suboptimal expression, while achieving high-efficiency cellular uptake and specificity-driven genome editing across diverse cell lines and genetic targets.230
Vaccines for Infectious Diseases
A vaccine is primarily composed of immunogenic components, such as inactivated or attenuated pathogens, recombinant antigens, or nucleic acids, along with adjuvants to enhance non-specific immune responses and stabilizers to prevent the degradation of vaccine constituents.231 Synthetic nanoparticles have gained significant attention in vaccine development. Their capability to encapsulate antigens enables surface functionalization for targeted delivery, and the nanoparticles act as immunostimulatory adjuvants that delay antigen degradation while activating APCs and lymphocytes. Their vaccine applications are categorized into three primary roles: antigen delivery vehicles, adjuvant delivery vehicles, and self-contained nucleic acid vaccine carriers.232 RNA vaccines offer a non-integrating, degradable, and non-infectious modality with enhanced manufacturability,233,234 thereby justifying their prioritization in this review.
The SARS-CoV-2 vaccines developed during the COVID-19 pandemic operate by delivering viral spike protein antigens via synthetic nanoparticle carriers to trigger adaptive immune responses. mRNA-1273 (Moderna) and BNT162b2 (Pfizer-BioNTech) employed N1-methylpseudouridine-modified mRNA encapsulated in LNPs formulated with SM-102 or ALC-0315 ionizable lipids, respectively, which encoded SARS-CoV-2 spike glycoprotein variants. Real-world efficacy data from 3,950 U.S. health care personnel, first responders, as well as other essential and frontline workers demonstrated 90% protection against SARS-CoV-2 infection following two doses of either vaccine.235 Accumulating evidence indicates that LNPs possess intrinsic immunostimulatory properties, thereby positioning them as potent adjuvants for recombinant protein vaccines. Dai et al systematically evaluated LNP adjuvant efficacy against conventional aluminum-based adjuvants in norovirus vaccine formulations and demonstrated that LNPs enhance recombinant protein immunogenicity by inducing cellular immunity and promoting balanced IgG2a/IgG1 ratios relative to alum. Furthermore, LNPs outperformed aluminum hydroxide in generating Th1-type cytokine-expressing CD4+ and CD8+ T cell immune responses, thus establishing their capacity to synergistically activate cytotoxic T lymphocytes and B cells, which are critical aspects for antiviral and antitumor immunity.236 Synthetic nanoparticles frequently serve as direct carriers for nucleic acid vaccines: Wang et al engineered HBsAg-encapsulating liposomes dually functionalized with mannose-PEG-cholesterol for C-type lectin receptor targeting and lipid A as an adjuvant via emulsification-freeze-drying, subsequently integrating them into microneedle arrays for oral mucosal immunization. They achieved simultaneous humoral and mucosal immunity against hepatitis B surface antigen.237 Separately, Wang et al developed a protein nanoparticle CBU-CpG-E2 vaccine co-encapsulating Behçet’s disease-related CBU1910 antigen, CpG1826 adjuvant, and PLGA-PEG-PLGA hydrogel and demonstrated that combinatorial nanoparticle-hydrogel vaccination elicited enhanced magnitude and durability of humoral immunity compared to single-component systems.238
Cancer Therapies
Cancer Vaccines
Synthetic nanoparticles enhance cancer vaccine efficacy by delivering immunomodulators such as tumor antigens and adjuvants, thereby amplifying tumor-specific T-cell activation through active immunization.239 Cancer vaccine components comprise tumor antigens (tumor-associated or tumor-specific antigens), formulations (whole-cell or antigen-specific), immune adjuvants (eg granulocyte-macrophage colony-stimulating factor [GM-CSF], CpG oligodeoxynucleotide [CpG-ODN], and Toll-like receptor [TLR] agonists240,241), and nanoparticle delivery systems. Among these, tumor-specific neoantigens—novel epitopes recognized by autologous T cells to initiate anti-tumor immunity independently of central tolerance242—uniquely elicit potent CD4+ and CD8+ T-cell responses via dendritic cells.
Chen et al successfully delivered a model tumor neoantigen using engineered porous hollow Mycobacterium tuberculosis (phMTb) that retained surface pathogen-associated molecular patterns for APC recognition and adhesion molecules facilitating APCs invasion, thereby enhancing cross-presentation of tumor neoantigens, dendritic cell activation, migration, and effector/memory CD8+ T cell function243 Zhang et al developed phospholipid-coated mesoporous silica nanoparticles co-delivering ovarian cancer antigen OVA257-264 and the TLR agonist monophosphoryl lipid A adjuvant, thereby improving lymph node-targeted antigen delivery and achieving synergistic tumor antigen-adjuvant immune activation. Clinical trials demonstrate that neoantigenic tumor vaccines exhibit insufficient antigen uptake, poor tumor recognition, weak CD8+ T-cell responses, and impaired cytotoxic activity attributable to passive APC engagement. To address these limitations, Liang et al engineered a tumor-targeting vaccine by co-loading antigen-loaded ginsenoside-Rg3-based LNPs and GM-CSF into a macroporous hydrogel, where Rg3 serves as a targeting ligand for glucose transporter protein-1-overexpressing tumor cells. The Rg3-LNPs anchor antigens onto tumor cell surfaces and enhance antigen presentation and recognition through adjuvant synergy, thereby mitigating immune escape in antitumor therapy.244 Xu et al engineered a spleen- and tumor-dual-targeting vaccine by incorporating cholesterol-conjugated and thiol-modified CpG-ODN, tumor-specific neoantigen peptides, and the LNPD18 vector, which induced monocytic myeloid-derived suppressor cell differentiation into M1 macrophages, activated splenic dendritic cells, facilitated tumor eradication, reprogrammed the tumor microenvironment, and activated CD8+ T cells.245
Other Therapeutic Effects
Beyond their role in tumor vaccines, synthetic nanoparticles serve multifunctional purposes in oncology therapeutics—primarily as targeted drug delivery vehicles that recognize cancer-specific molecular markers to enable site-specific payload release and enhance bioavailability while sparing healthy tissues. Collagen networks critically regulate tumor proliferation, metastasis, and chemoresistance, prompting the development of collagen-targeting strategies: (i) enzymatic collagenolysis, which enhances intratumoral nanoparticle penetration246,247 and (ii) photothermal collagen denaturation, promoting deeper diffusion of therapeutic nanoparticles.248 Resultant collagen fragments further potentiate conventional therapies by improving chemotherapeutic agent permeation, immunotherapeutic antibody distribution, and immune cell infiltration into tumors. Zhou et al pioneered the use of spermine as a tumor-targeting ligand in polymeric micelles encapsulating the hydrophobic chemotherapeutic agent paclitaxel. By exploiting spermine’s specific binding to the polyamine transport system overexpressed on tumor cells, they achieved active targeting for anticancer drug delivery.249
Synthetic nanoparticles additionally remodel the immunosuppressive tumor microenvironment by inhibiting immunosuppressive signals and activating co-stimulatory pathways. Li et al developed a multifunctional MPLO@HA nanomedicine for the targeted treatment of Fusobacterium nucleatum (Fn)-associated colorectal cancer.250 This platform conjugates metformin, oxaliplatin, and lauric acid to oligomethyleneimine, which is subsequently complexed with hyaluronic acid. The system effectively eradicates intra- and extracellular Fn to overcome Fn-induced chemoresistance, disrupt tumor immunosuppression, improve patient prognosis, and pioneer therapies for bacterial-driven cancers. Song et al engineered an STK11@PPCM biomimetic nanoplatform targeting anti-PD-1-resistant, STK11-mutant non-small cell lung cancer. This system utilizes a cationic PBAE polymer that binds NSCLC cells to enable precise delivery of STK11-encoded DNA. By restoring STK11 expression, the platform reactivates autophagy and induces immunogenic cell death, thereby providing novel clinical strategies for STK11-mutated lung adenocarcinoma.251
Synthetic nanoparticles also provide an efficient alternative to chimeric antigen receptor-T cell generation and transduction by overcoming the limitations of conventional viral-based transduction and offering a safer approach for tumor-targeted T cell therapy. Lin et al combined microfluidic extrusion-mediated membrane disruption with LNP vector delivery, significantly enhancing transfection efficiency252 and cargo delivery while reducing production costs. This maintained cell viability while preserving native immune gene expression.253
Discussion
In recent years, research on EV-based DDSs has grown exponentially, with a clear shift from in vitro experiments to preclinical and clinical studies. Approximately 201 studies have been reported using EVs as interventions or research subjects on www.clinicaltrials.gov, predominantly Phase I exploratory trials. These primarily focus on two major areas: therapeutics (including tumor immune stimulation, inflammatory immunosuppression, and tissue regeneration) and diagnostics (eg early screening via liquid biopsy) EVs have been demonstrated to be safe and well-tolerated, with a low overall incidence of adverse events (4.4%; 95% CI: 0.7–22.2%).254 However, conducting large-scale Phase III trials to demonstrate positive efficacy remains constrained.
Current bottlenecks in EV clinical translation primarily stem from two aspects: limitations in production technology and inherent heterogeneity. Deficiencies in production technology remain a persistent challenge, primarily stemming from the inefficiency of separation and purification techniques. This encompasses: (i) an imbalance between purity, recovery, activity, and cost;255 (ii) functional risks: where forces from centrifugation, chemical reagents, or membrane adsorption can readily compromise EV structural integrity, thereby reducing their biological activity and impacting subsequent diagnostic or therapeutic applications; and (iii) internationally unified quality control standards deficiency: while current advanced methods, magnetic bead separation,256 lipid-based nanoprobe separation,257 and solid-phase medium separation technologies258 can partially reduce inter-batch heterogeneity within a single laboratory, differences in separation parameters between laboratories continue to hinder the cross-laboratory reproducibility of data.259 (iv) Further bottlenecks exist in mass production. Existing methods are predominantly suited for small-scale laboratory samples and struggle to meet the high-throughput, rapid processing demands of large-volume clinical specimens. Constraints also arise from limitations in engineered modifications: genetic modifications suffer from low membrane protein fusion efficiency and narrow donor cell applicability, while chemical modifications often cause membrane damage and induce non-specific binding. Both exogenous and endogenous loading strategies face challenges, including the loss of macromolecular bioactivity, inconsistent drug loading, significant batch variability for small-molecule payloads, and uncontrollable encapsulation rates. In vivo delivery is further hampered by phagocytic system clearance, off-target accumulation, and insufficient tissue penetration, resulting in actual targeting efficiency that falls far short of in vitro predictions.9,84,260
EV heterogeneity represents a major bottleneck in clinical translation, directly impeding the standardized production and predictable efficacy of EV-based therapies. On one hand, this heterogeneity complicates the assurance of “batch consistency”: even when derived from an identical cell line, variations in culture conditions or different cell passages can alter the composition and function of the resulting EVs, leading to inconsistent quality in clinical-grade EV products. On the other hand, heterogeneity hampers the prediction of therapeutic outcomes: the composition of EV subpopulations varies among patients, meaning the same EV therapy can yield vastly different clinical results due to individual differences in the recipient’s endogenous EV landscape.
Furthermore, heterogeneity is a determinant of EV immunogenicity and potential toxicity. In fact, the purported “low immunogenicity” of natural EVs is often overstated. They can, on one hand, activate anti-infective and anti-tumor immunity by carrying pathogen-associated molecular patterns, damage-associated molecular patterns, or antigenic peptides, positioning them as ideal candidates for vaccine development.261 Conversely, they can also mediate immune escape by expressing molecules like PD-L1 or releasing immunosuppressive miRNAs, thereby potentially accelerating tumor progression or exacerbating autoimmune diseases.262 The underlying mechanism is that their immune-inert or immune-active properties are highly dependent on their cellular origin and physiological state.8,263 For instance, EVs derived from microbes or activated immune cells may trigger immune responses due to foreign MHC molecules or stimulatory surface components,264,265 whereas those from tumor cells or certain stem cells often exhibit immunosuppressive tendencies.266,267
From a toxicity standpoint, while EVs themselves typically lack direct cytotoxicity, the pathological effects they mediate are characterized by their “indirect” and “covert” nature. For example, EV-mediated amplification of inflammation in sepsis,268 or EV-induced immunosuppression and promotion of metastasis within the tumor microenvironment,269 both represent forms of “secondary damage” orchestrated by EVs from distinct sources. The toxic thresholds and precise cellular targets of these effects remain difficult to quantify.
Furthermore, for artificially engineered nanoscale DDSs, functional modifications to engineered EVs inevitably increase immunogenic risks. The conjugation of exogenous molecules during surface modification may expose novel immunogenic epitopes, while the process of drug loading can disrupt membrane architecture, ultimately potentiating activation of the complement system. The current absence of standardized modifications renders this immunogenicity largely unpredictable.74 While EV mimetics enable scalable production, their artificially synthesized membrane components inherently lack the “self-recognition” proteins present on natural EVs, rendering them more susceptible to clearance by the mononuclear phagocyte system. Additionally, EV mimetics prepared via methods such as cell lysis may retain residual immunostimulatory components.270
In comparison to EV-based DDS, traditional synthetic nanoparticles present toxicity profiles that extend beyond immunogenicity. This includes inherent material toxicity, the potential for chronic damage from accumulated degradation products, and the compounded effects of immunotoxicity and off-target toxicity. Consequently, their overall biosafety profile is generally regarded as inferior to that of EV-based carriers.270,271 Nevertheless, the capacity for large-scale production and the achievement of high batch consistency remain undeniable advantages of synthetic nanoparticle platforms.272
A comparison of the advantages, disadvantages, challenges, and future optimization directions regarding native EVs, engineered EVs and EV mimetics have been showed in Table 7.
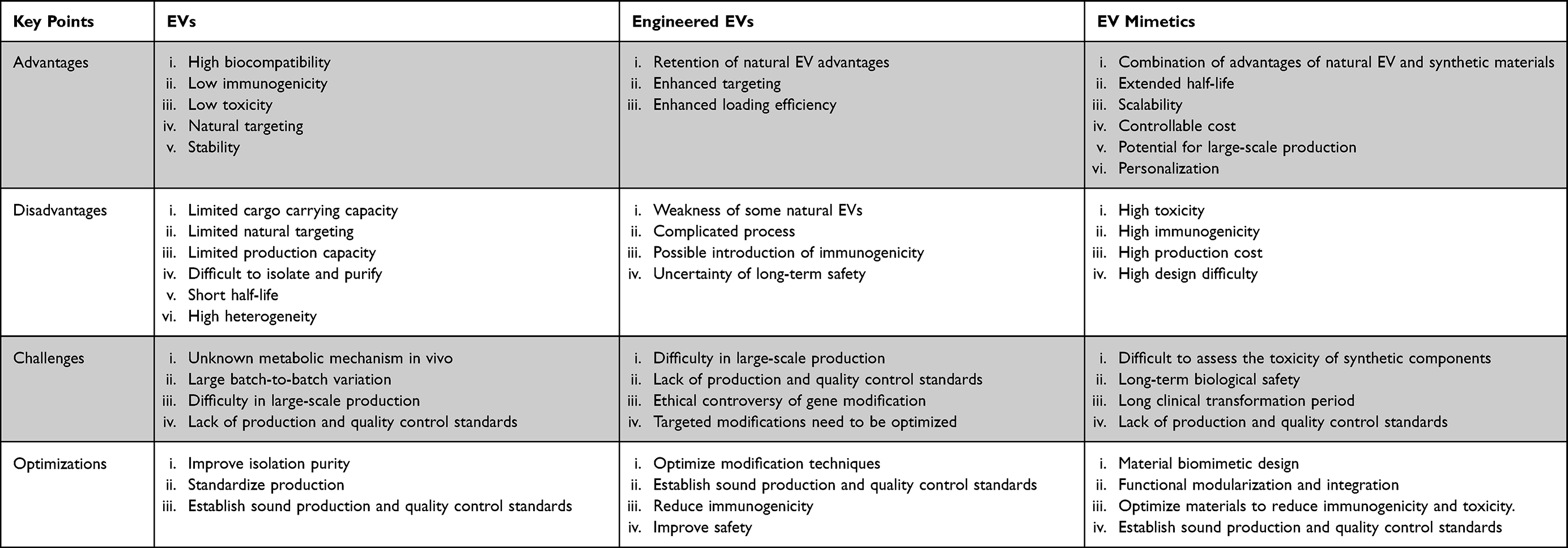 |
Table 7 Comparison of Advantages, Disadvantages, Challenges, and Future Optimization Directions Regarding Native EVs, Engineered EVs, and EV Mimetics |
Conclusion and Outlook
As an emerging DDS, EVs demonstrate significant potential. Their clinical translation and application present a landscape of concurrent challenges and developmental opportunities. Researchers are continuously exploring novel approaches to address the aforementioned limitations.
Regarding production and preparation, studies indicate that integrating tangential flow filtration—a technique amenable to industrial-scale manufacturing—with established EV purification methods facilitates high-purity EV isolation with improved throughput.273–276 For standardized EV production, reproducible, Good Manufacturing Practice (GMP)-compliant process standards are being proposed and progressively refined through clinical trials. For instance, Humbert et al eliminated donor heterogeneity by utilizing identical human induced pluripotent stem cell (hiPSC) donors, enabling the generation of highly reproducible clinical-grade product batches.274 By controlling key process parameters (seeding density, medium composition, cell handling, incubation time, and temperature), they standardized the production workflow from hiPSC-derived cardiovascular progenitor cells (CPCs) vesiculation to product purification, concentration, and sterile filtration, and finally achieved excellent batch consistency between preclinical and clinical-scale batches.274 For characterization, advanced instrumentation such as ImageStream, Amnis, ZetaView, and ONI Nanoimager enables detailed multi-parametric analysis of EV concentration, particle size, zeta potential, and specific exosomal markers, offering a potential integrated alternative to traditional, often standalone methods like flow cytometry, Western blotting, dynamic light scattering, and nanoparticle tracking analysis.277
Furthermore, the integration of artificial intelligence (AI) with nanotechnology is propelling nanoscale diagnostic and DDSs toward more profound precision medicine. AI primarily encompasses the subsets of deep learning (DL) and machine learning (ML). DL is applied to predict interactions between nanocarriers and biological systems278 and to design ligands for precise targeting.279 Researchers have already employed such approaches to engineer iron oxide nanoparticles for targeted, intelligently controlled release in cancer therapy.280 ML algorithms analyze structural features of nanocarriers to predict their physicochemical properties and biological interactions, efficiently processing multi-dimensional data from EV multi-omics, imaging, and biophysics—patterns often unrecognizable by traditional statistical methods.281 Simultaneously, these models can predict individual patient drug responses to customize dosage regimens and release mechanisms, aiming to minimize resistance and adverse effects while enhancing therapeutic efficacy. Through hierarchical modeling and clustering analysis, they can precisely distinguish EV subtypes and their functional differences, thereby helping to overcome heterogeneity bottlenecks. Bayesian parameter optimization and standardized data processing pipelines can further boost research and development and translational efficiency. The use of tools like SHAP and LIME helps decipher model decision-making principles, identifying key functional molecules to provide clear directions for rational EV engineering modifications.282
Although AI-driven EV DDSs represent a promising developmental trajectory, they remain constrained by data sparsity, insufficient model interpretability, and a lack of cross-platform standardization. Furthermore, current models often rely excessively on in vitro data, with insufficient validation of their predictions regarding batch consistency and efficacy correlation in clinical translation. Future efforts should focus on developing few-shot learning and self-supervised algorithms tailored to EV data characteristics, enhancing multimodal data integration and interpretability tools, while advancing federated learning frameworks combined with real-time analysis. By linking AI-driven process optimization with clinical outcome data, this integrated approach will enable the development of more efficient, safer, and personalized EV-based diagnostics and therapies.
Data Sharing Statement
The total number of 282 references can be found in PubMed.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by National Natural Science Foundation of China (82371044), Natural Science Foundation of Tianjin (24JCZDJC00850), and Tianjin Health Commission Key Discipline Special Project (TJWJ2023XK009).
Disclosure
The authors declared that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
2. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30(Volume 30, 2014):255–30. doi:10.1146/annurev-cellbio-101512-122326
3. Zaborowski MP, Balaj L, Breakefield XO, Lai CP. Extracellular vesicles: composition, biological relevance, and methods of study. Bioscience. 2015;65(8):783–797. doi:10.1093/biosci/biv084
4. Couch Y, Buzàs EI, Di Vizio D, et al. A brief history of nearly EV-erything - The rise and rise of extracellular vesicles. J Extracell Vesicles. 2021;10(14):e12144. doi:10.1002/jev2.12144
5. H Rashed M, Bayraktar E, K Helal G, et al. Exosomes: from garbage bins to promising therapeutic targets. Int J Mol Sci. 2017;18(3):538. doi:10.3390/ijms18030538
6. Chargaff E, West R. The biological significance of the thromboplastic protein of blood. J Biol Chem. 1946;166(1):189–197.
7. Trams EG, Lauter CJ, Salem N, Heine U. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochim Biophys Acta. 1981;645(1):63–70. doi:10.1016/0005-2736(81)90512-5
8. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi:10.1084/jem.183.3.1161
9. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
10. Gould SJ, Raposo G. As we wait: coping with an imperfect nomenclature for extracellular vesicles. J Extracell Vesicles. 2013;2. doi:10.3402/jev.v2i0.20389
11. Mizrak A, Bolukbasi MF, Ozdener GB, et al. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol Ther J Am Soc Gene Ther. 2013;21(1):101–108. doi:10.1038/mt.2012.161
12. He F, Li WN, Li XX, et al. Exosome-mediated delivery of RBP-J decoy oligodeoxynucleotides ameliorates hepatic fibrosis in mice. Theranostics. 2022;12(4):1816–1828. doi:10.7150/thno.69885
13. Lu M, Huang Y. Bioinspired exosome-like therapeutics and delivery nanoplatforms. Biomaterials. 2020;242:119925. doi:10.1016/j.biomaterials.2020.119925
14. Morad G, Carman CV, Hagedorn EJ, et al. Tumor-derived extracellular vesicles breach the intact blood-brain barrier via transcytosis. ACS Nano. 2019;13(12):13853–13865. doi:10.1021/acsnano.9b04397
15. Cheng G, Li W, Ha L, et al. Self-assembly of extracellular vesicle-like metal-organic framework nanoparticles for protection and intracellular delivery of biofunctional proteins. J Am Chem Soc. 2018;140(23):7282–7291. doi:10.1021/jacs.8b03584
16. Tian Y, Li S, Song J, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials. 2014;35(7):2383–2390. doi:10.1016/j.biomaterials.2013.11.083
17. Ismail N, Wang Y, Dakhlallah D, et al. Macrophage microvesicles induce macrophage differentiation and miR-223 transfer. Blood. 2013;121(6):984–995. doi:10.1182/blood-2011-08-374793
18. Preservation and Storage Stability of Extracellular Vesicles for Therapeutic Applications - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/29181730/.
19. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release off J Control Release Soc. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
20. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJA. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
21. Kowal J, Arras G, Colombo M, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A. 2016;113(8):E968–977. doi:10.1073/pnas.1521230113
22. Zhang Y, Lan M, Chen Y. Minimal Information for Studies of Extracellular Vesicles (MISEV): ten-Year Evolution (2014-2023). Pharmaceutics. 2024;16(11):1394. doi:10.3390/pharmaceutics16111394
23. Machado FJD, Marta-Enguita J, Herrera M, et al. Transcriptomic profiling of thrombus-derived extracellular vesicles reveals PECAM-1 as a potential inflammatory marker for cardioembolic stroke patients. J Neuroinflammation. 2025;22(1):229. doi:10.1186/s12974-025-03555-8
24. Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–593. doi:10.1038/nri2567
25. Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006;Chapter 3:
26. Jeppesen DK, Hvam ML, Primdahl-Bengtson B, et al. Comparative analysis of discrete exosome fractions obtained by differential centrifugation. J Extracell Vesicles. 2014;3:25011. doi:10.3402/jev.v3.25011
27. The influence of rotor type and centrifugation time on the yield and purity of extracellular vesicles - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/24678386/.
28. Rani S, O’Brien K, Kelleher FC, et al. Isolation of exosomes for subsequent mRNA, MicroRNA, and protein profiling. Methods Mol Biol Clifton NJ. 2011;784:181–195. doi:10.1007/978-1-61779-289-2_13
29. Tauro BJ, Greening DW, Mathias RA, et al. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods San Diego Calif. 2012;56(2):293–304. doi:10.1016/j.ymeth.2012.01.002
30. Shtam TA, Samsonov RA, Volnitskiy AV, et al. Isolation of extracellular micro-vesicles from cell culture medium: comparative evaluation of methods. Biomeditsinskaia Khimiia. 2018;64(1):23–30. doi:10.18097/PBMC20186401023
31. Brennan K, Martin K, FitzGerald SP, et al. A comparison of methods for the isolation and separation of extracellular vesicles from protein and lipid particles in human serum. Sci Rep. 2020;10:1039. doi:10.1038/s41598-020-57497-7
32. Li X, La Salvia S, Liang Y, et al. Extracellular vesicle-encapsulated adeno-associated viruses for therapeutic gene delivery to the heart. Circulation. 2023;148(5):405–425. doi:10.1161/CIRCULATIONAHA.122.063759
33. Obi PO, Souza TFG, Özerkliğ B, et al. Extracellular vesicles released from skeletal muscle post-chronic contractile activity increase mitochondrial biogenesis in recipient myoblasts. J Extracell Vesicles. 2025;14(4):e70045. doi:10.1002/jev2.70045
34. Cao Y, Tan X, Shen J, et al. Morinda Officinalis-derived extracellular vesicle-like particles: anti-osteoporosis effect by regulating MAPK signaling pathway. Phytomedicine Int J Phytother Phytopharm. 2024;129:155628. doi:10.1016/j.phymed.2024.155628
35. Vaidya A, Parande D, Khadse N, et al. Analytical characterization of heterogeneities in mRNA-lipid nanoparticles using sucrose density gradient ultracentrifugation. Anal Chem. 2024;96(14):5570–5579. doi:10.1021/acs.analchem.4c00031
36. Gagnon P, Leskovec M, Prebil SD, et al. Removal of empty capsids from adeno-associated virus preparations by multimodal metal affinity chromatography. J Chromatogr A. 2021;1649:462210. doi:10.1016/j.chroma.2021.462210
37. Ou J, Li K, Yuan H, et al. Staphylococcus aureus vesicles impair cutaneous wound healing through p38 MAPK-MerTK cleavage-mediated inhibition of macrophage efferocytosis. Cell Commun Signal CCS. 2025;23(1):14. doi:10.1186/s12964-024-01994-z
38. Heijnen HF, Schiel AE, Fijnheer R, Geuze HJ, Sixma JJ. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood. 1999;94(11):3791–3799.
39. Keller S, König AK, Marmé F, et al. Systemic presence and tumor-growth promoting effect of ovarian carcinoma released exosomes. Cancer Lett. 2009;278(1):73–81. doi:10.1016/j.canlet.2008.12.028
40. Théry C, Boussac M, Véron P, et al. Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J Immunol Baltim Md 1950. 2001;166(12):7309–7318. doi:10.4049/jimmunol.166.12.7309
41. Wubbolts R, Leckie RS, Veenhuizen PTM, et al. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J Biol Chem. 2003;278(13):10963–10972. doi:10.1074/jbc.M207550200
42. Rabesandratana H, Toutant JP, Reggio H, Vidal M. Decay-accelerating factor (CD55) and membrane inhibitor of reactive lysis (CD59) are released within exosomes during In vitro maturation of reticulocytes. Blood. 1998;91(7):2573–2580.
43. Chandrasekera D, Shah R, van Hout I, et al. Combination of precipitation and size exclusion chromatography as an effective method for exosome like extracellular vesicle isolation from pericardial fluids. Nanotheranostics. 2023;7(4):345–352. doi:10.7150/ntno.82939
44. Duong P, Chung A, Bouchareychas L, Raffai RL. Cushioned-Density Gradient Ultracentrifugation (C-DGUC) improves the isolation efficiency of extracellular vesicles. PLoS One. 2019;14(4):e0215324. doi:10.1371/journal.pone.0215324
45. Small extracellular vesicles isolation and separation: current techniques, pending questions and clinical applications - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/36185597/.
46. Otani K, Fujioka Y, Okada M, Yamawaki H. Optimal isolation method of small extracellular vesicles from rat plasma. Int J Mol Sci. 2019;20(19):4780. doi:10.3390/ijms20194780
47. Stam J, Bartel S, Bischoff R, Wolters JC. Isolation of extracellular vesicles with combined enrichment methods. J Chromatogr B. 2021;1169:122604. doi:10.1016/j.jchromb.2021.122604
48. Fang X, Chen C, Liu B, et al. A magnetic bead-mediated selective adsorption strategy for extracellular vesicle separation and purification. Acta Biomater. 2021;124:336–347. doi:10.1016/j.actbio.2021.02.004
49. Theel EK, Schwaminger SP. Microfluidic approaches for affinity-based exosome separation. Int J Mol Sci. 2022;23(16):9004. doi:10.3390/ijms23169004
50. Petersen KE, Manangon E, Hood JL, et al. A review of exosome separation techniques and characterization of B16-F10 mouse melanoma exosomes with AF4-UV-MALS-DLS-TEM. Anal Bioanal Chem. 2014;406(30):7855–7866. doi:10.1007/s00216-014-8040-0
51. Dong L, Zieren RC, Horie K, et al. Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium. J Extracell Vesicles. 2020;10(2):e12044. doi:10.1002/jev2.12044
52. Palma J, Yaddanapudi SC, Pigati L, et al. MicroRNAs are exported from malignant cells in customized particles. Nucleic Acids Res. 2012;40(18):9125–9138. doi:10.1093/nar/gks656
53. The profiles of microRNAs from urinary extracellular vesicles (EVs) prepared by various isolation methods and their correlation with serum EV microRNAs - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/31987752/.
54. Aalberts M, van Dissel-Emiliani FMF, van Adrichem NPH, et al. Identification of distinct populations of prostasomes that differentially express prostate stem cell antigen, annexin A1, and GLIPR2 in humans. Biol Reprod. 2012;86(3):82. doi:10.1095/biolreprod.111.095760
55. Zhu K, Wang Z, Zong S, et al. Hydrophobic plasmonic nanoacorn array for a label-free and uniform SERS-based biomolecular assay. ACS Appl Mater Interfaces. 2020;12(26):29917–29927. doi:10.1021/acsami.0c03993
56. Welsh JA, Holloway JA, Wilkinson JS, Englyst NA. Extracellular vesicle flow cytometry analysis and standardization. Front Cell Dev Biol. 2017;5:78. doi:10.3389/fcell.2017.00078
57. Monguió-Tortajada M, Gálvez-Montón C, Bayes-Genis A, Roura S, Borràs FE. Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cell Mol Life Sci CMLS. 2019;76(12):2369–2382. doi:10.1007/s00018-019-03071-y
58. He M, Zeng Y. Microfluidic exosome analysis toward liquid biopsy for cancer. J Lab Autom. 2016;21(4):599–608. doi:10.1177/2211068216651035
59. Ingato D, Lee JU, Sim SJ, Kwon YJ. Good things come in small packages: overcoming challenges to harness extracellular vesicles for therapeutic delivery. J Control Release off J Control Release Soc. 2016;241:174–185. doi:10.1016/j.jconrel.2016.09.016
60. Yom-Tov N, Guy R, Offen D. Extracellular vesicles over adeno-associated viruses: advantages and limitations as drug delivery platforms in precision medicine. Adv Drug Deliv Rev. 2022;190:114535. doi:10.1016/j.addr.2022.114535
61. Danilushkina AA, Emene CC, Barlev NA, Gomzikova MO. Strategies for Engineering of Extracellular Vesicles. Int J Mol Sci. 2023;24(17):13247. doi:10.3390/ijms241713247
62. Hood JL. Post isolation modification of exosomes for nanomedicine applications. Nanomed. 2016;11(13):1745–1756. doi:10.2217/nnm-2016-0102
63. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11(7):3183–3195. doi:10.7150/thno.52570
64. Wang C, Li N, Li Y, et al. Engineering a HEK-293T exosome-based delivery platform for efficient tumor-targeting chemotherapy/internal irradiation combination therapy. J Nanobiotechnol. 2022;20(1):247. doi:10.1186/s12951-022-01462-1
65. Zhang H, Yan W, Wang J, et al. Surface functionalization of exosomes for chondrocyte-targeted siRNA delivery and cartilage regeneration. J Control Release off J Control Release Soc. 2024;369:493–505. doi:10.1016/j.jconrel.2024.04.009
66. Lai CP, Mardini O, Ericsson M, et al. Dynamic biodistribution of extracellular vesicles in vivo using a multimodal imaging reporter. ACS Nano. 2014;8(1):483–494. doi:10.1021/nn404945r
67. Zhang H, Gong L, Yu L, et al. Emerging roles of non-coding RNA derived from extracellular vesicles in regulating PD-1/PD-L1 pathway: insights into cancer immunotherapy and clinical applications. Cancer Cell Int. 2025;25(1):188. doi:10.1186/s12935-025-03809-8
68. Hung ME, Leonard JN. Stabilization of exosome-targeting peptides via engineered glycosylation. J Biol Chem. 2015;290(13):8166–8172. doi:10.1074/jbc.M114.621383
69. Whitley JA, Kim S, Lou L, et al. Encapsulating Cas9 into extracellular vesicles by protein myristoylation. J Extracell Vesicles. 2022;11(4):e12196. doi:10.1002/jev2.12196
70. Sul JH, Shin S, Kim HK, et al. Dopamine-conjugated extracellular vesicles induce autophagy in Parkinson’s disease. J Extracell Vesicles. 2024;13(12):e70018. doi:10.1002/jev2.70018
71. Jung KO, Jo H, Yu JH, Gambhir SS, Pratx G. Development and MPI tracking of novel hypoxia-targeted theranostic exosomes. Biomaterials. 2018;177:139–148. doi:10.1016/j.biomaterials.2018.05.048
72. Lee TS, Kim Y, Zhang W, Song IH, Tung CH. Facile metabolic glycan labeling strategy for exosome tracking. Biochim Biophys Acta Gen Subj. 2018;1862(5):1091–1100. doi:10.1016/j.bbagen.2018.02.001
73. Rayamajhi S, Aryal S. Surface functionalization strategies of extracellular vesicles. J Mater Chem B. 2020;8(21):4552–4569. doi:10.1039/d0tb00744g
74. Pham TC, Jayasinghe MK, Pham TT, et al. Covalent conjugation of extracellular vesicles with peptides and nanobodies for targeted therapeutic delivery. J Extracell Vesicles. 2021;10(4):e12057. doi:10.1002/jev2.12057
75. Salunkhe S, null D, Basak M, Chitkara D, Mittal A. Surface functionalization of exosomes for target-specific delivery and in vivo imaging & tracking: strategies and significance. J Control Release off J Control Release Soc. 2020;326:599–614. doi:10.1016/j.jconrel.2020.07.042
76. Lee J, Lee H, Goh U, et al. Cellular engineering with membrane fusogenic liposomes to produce functionalized extracellular vesicles. ACS Appl Mater Interfaces. 2016;8(11):6790–6795. doi:10.1021/acsami.6b01315
77. Sato YT, Umezaki K, Sawada S, et al. Engineering hybrid exosomes by membrane fusion with liposomes. Sci Rep. 2016;6:21933. doi:10.1038/srep21933
78. Tamura R, Uemoto S, Tabata Y. Augmented liver targeting of exosomes by surface modification with cationized pullulan. Acta Biomater. 2017;57:274–284. doi:10.1016/j.actbio.2017.05.013
79. Wang J, Li W, Zhang L, et al. Chemically edited exosomes with dual ligand purified by microfluidic device for active targeted drug delivery to tumor cells. ACS Appl Mater Interfaces. 2017;9(33):27441–27452. doi:10.1021/acsami.7b06464
80. Emam SE, Elsadek NE, Abu Lila AS, et al. Anti-PEG IgM production and accelerated blood clearance phenomenon after the administration of PEGylated exosomes in mice. J Control Release off J Control Release Soc. 2021;334:327–334. doi:10.1016/j.jconrel.2021.05.001
81. Chen SY, Chen YL, Li PC, et al. Engineered extracellular vesicles carrying let-7a-5p for alleviating inflammation in acute lung injury. J Biomed Sci. 2024;31(1):30. doi:10.1186/s12929-024-01019-4
82. Kwak G, Gololobova O, Sharma N, et al. Extracellular vesicles enhance pulmonary transduction of stably associated adeno-associated virus following intratracheal administration. J Extracell Vesicles. 2023;12(6):e12324. doi:10.1002/jev2.12324
83. Wang C, Ju J, Fu C, et al. Metabolically engineered extracellular vesicles released from a composite hydrogel delivery system regulate the microenvironment for periprosthetic osteolysis treatment. J Extracell Vesicles. 2025;14(6):e70098. doi:10.1002/jev2.70098
84. Dave KM, Pinky PP, Manickam DS. Molecular engineering of extracellular vesicles for drug delivery: strategies, challenges, and perspectives. J Control Release off J Control Release Soc. 2025;114068. doi:10.1016/j.jconrel.2025.114068
85. Jang SC, Kim OY, Yoon CM, et al. Bioinspired exosome-mimetic nanovesicles for targeted delivery of chemotherapeutics to malignant tumors. ACS Nano. 2013;7(9):7698–7710. doi:10.1021/nn402232g
86. Yang C, Xue Y, Duan Y, Mao C, Wan M. Extracellular vesicles and their engineering strategies, delivery systems, and biomedical applications. J Control Release off J Control Release Soc. 2024;365:1089–1123. doi:10.1016/j.jconrel.2023.11.057
87. Jo W, Jeong D, Kim J, et al. Microfluidic fabrication of cell-derived nanovesicles as endogenous RNA carriers. Lab Chip. 2014;14(7):1261–1269. doi:10.1039/c3lc50993a
88. Fuhrmann G, Serio A, Mazo M, Nair R, Stevens MM. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J Control Release off J Control Release Soc. 2015;205:35–44. doi:10.1016/j.jconrel.2014.11.029
89. Rong Y, Wang Z, Tang P, et al. Engineered extracellular vesicles for delivery of siRNA promoting targeted repair of traumatic spinal cord injury. Bioact Mater. 2023;23:328–342. doi:10.1016/j.bioactmat.2022.11.011
90. Wahlgren J, De L Karlson T, Brisslert M, et al. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res. 2012;40(17):e130. doi:10.1093/nar/gks463
91. van Rooij T, Skachkov I, Beekers I, et al. Viability of endothelial cells after ultrasound-mediated sonoporation: influence of targeting, oscillation, and displacement of microbubbles. J Control Release. 2016;238:197–211. doi:10.1016/j.jconrel.2016.07.037
92. Lamichhane TN, Jeyaram A, Patel DB, et al. Oncogene knockdown via active loading of small RNAs into extracellular vesicles by sonication. Cell Mol Bioeng. 2016;9(3):315–324. doi:10.1007/s12195-016-0457-4
93. Kooijmans SAA, Stremersch S, Braeckmans K, et al. Electroporation-induced siRNA precipitation obscures the efficiency of siRNA loading into extracellular vesicles. J Control Release off J Control Release Soc. 2013;172(1):229–238. doi:10.1016/j.jconrel.2013.08.014
94. Zhou Y, Du T, Yang CL, et al. Extracellular vesicles encapsulated with caspase-1 inhibitor ameliorate experimental autoimmune myasthenia gravis through targeting macrophages. J Control Release off J Control Release Soc. 2023;364:458–472. doi:10.1016/j.jconrel.2023.11.006
95. Wang P, Wang H, Huang Q, et al. Exosomes from M1-polarized macrophages enhance paclitaxel antitumor activity by activating macrophages-mediated inflammation. Theranostics. 2019;9(6):1714–1727. doi:10.7150/thno.30716
96. Kim MS, Haney MJ, Zhao Y, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations. Nanomedicine. 2018;14(1):195–204. doi:10.1016/j.nano.2017.09.011
97. Lee K, Fraser K, Ghaddar B, et al. Multiplexed profiling of single extracellular vesicles. ACS Nano. 2018;12(1):494–503. doi:10.1021/acsnano.7b07060
98. Raghav A, Jeong GB. A systematic review on the modifications of extracellular vesicles: a revolutionized tool of nano-biotechnology. J Nanobiotechnol. 2021;19:459. doi:10.1186/s12951-021-01219-2
99. Luan X, Sansanaphongpricha K, Myers I, Chen H, Yuan H, Sun D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol Sin. 2017;38(6):754–763. doi:10.1038/aps.2017.12
100. Le Saux S, Aarrass H, Lai-Kee-Him J, et al. Post-production modifications of murine mesenchymal stem cell (mMSC) derived extracellular vesicles (EVs) and impact on their cellular interaction. Biomaterials. 2020;231:119675. doi:10.1016/j.biomaterials.2019.119675
101. Ahmadian S, Jafari N, Tamadon A, Ghaffarzadeh A, Rahbarghazi R, Mahdipour M. Different storage and freezing protocols for extracellular vesicles: a systematic review. Stem Cell Res Ther. 2024;15(1):453. doi:10.1186/s13287-024-04005-7
102. Zhang D, Lee H, Zhu Z, Minhas JK, Jin Y. Enrichment of selective miRNAs in exosomes and delivery of exosomal miRNAs in vitro and in vivo. Am J Physiol Lung Cell Mol Physiol. 2017;312(1):L110–L121. doi:10.1152/ajplung.00423.2016
103. Grzelka K, Matkowski A, Chodaczek G, et al. Pulsed Electric Field (PEF) treatment results in growth promotion, main flavonoids extraction, and phytochemical profile modulation of scutellaria baicalensis georgi roots. Int J Mol Sci. 2024;26(1):100. doi:10.3390/ijms26010100
104. Luengo E, Álvarez I, Raso J. Improving carotenoid extraction from tomato waste by pulsed electric fields. Front Nutr. 2014;1:12. doi:10.3389/fnut.2014.00012
105. Haney MJ, Klyachko NL, Harrison EB, Zhao Y, Kabanov AV, Batrakova EV. TPP1 delivery to lysosomes with extracellular vesicles and their enhanced brain distribution in the animal model of batten disease. Adv Healthc Mater. 2019;8(11):e1801271. doi:10.1002/adhm.201801271
106. Koh HB, Kim HJ, Kang SW, Yoo TH. Exosome-based drug delivery: translation from bench to clinic. Pharmaceutics. 2023;15(8):2042. doi:10.3390/pharmaceutics15082042
107. Falcinelli M, Al-Hity G, Baron S, et al. Propranolol reduces IFN-γ driven PD-L1 immunosuppression and improves anti-tumour immunity in ovarian cancer. Brain Behav Immun. 2023;110:1–12. doi:10.1016/j.bbi.2023.02.011
108. Chandler K, Millar J, Ward G, et al. Imaging of light-enhanced extracellular vesicle-mediated delivery of oxaliplatin to colorectal cancer cells via laser ablation, inductively coupled plasma mass spectrometry. Cells. 2023;13(1):24. doi:10.3390/cells13010024
109. Zhuang X, Xiang X, Grizzle W, et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther J Am Soc Gene Ther. 2011;19(10):1769–1779. doi:10.1038/mt.2011.164
110. Shao X, Zhang M, Chen Y, Sun S, Yang S, Li Q. Exosome-mediated delivery of superoxide dismutase for anti-aging studies in Caenorhabditis elegans. Int J Pharm. 2023;641:123090. doi:10.1016/j.ijpharm.2023.123090
111. Chen Y, Douanne N, Wu T, et al. Leveraging nature’s nanocarriers: translating insights from extracellular vesicles to biomimetic synthetic vesicles for biomedical applications. Sci Adv. 2025;11(9):eads5249. doi:10.1126/sciadv.ads5249
112. Wang X, Zhao X, Zhong Y, Shen J, An W. Biomimetic exosomes: a new generation of drug delivery system. Front Bioeng Biotechnol. 2022;10:865682. doi:10.3389/fbioe.2022.865682
113. Ying M, Zhuang J, Wei X, et al. Remote-loaded platelet vesicles for disease-targeted delivery of therapeutics. Adv Funct Mater. 2018;28(22):1801032. doi:10.1002/adfm.201801032
114. Wang G, Hu W, Chen H, Shou X, Ye T, Xu Y. Cocktail strategy based on NK cell-derived exosomes and their biomimetic nanoparticles for dual tumor therapy. Cancers. 2019;11(10):1560. doi:10.3390/cancers11101560
115. Ai X, Wang S, Duan Y, et al. Emerging approaches to functionalizing cell membrane-coated nanoparticles. Biochemistry. 2021;60(13):941–955. doi:10.1021/acs.biochem.0c00343
116. Cruz-Samperio R, Hicks CL, Scott A, et al. Modular bioorthogonal lipid nanoparticle modification platforms for cardiac homing. J Am Chem Soc. 2023;145(41):22659–22670. doi:10.1021/jacs.3c07811
117. Lu M, Zhao X, Xing H, et al. Liposome-chaperoned cell-free synthesis for the design of proteoliposomes: implications for therapeutic delivery. Acta Biomater. 2018;76:1–20. doi:10.1016/j.actbio.2018.03.043
118. Vázquez-Ríos AJ, Molina-Crespo Á, Bouzo BL, López-López R, Moreno-Bueno G, de la Fuente M. Exosome-mimetic nanoplatforms for targeted cancer drug delivery. J Nanobiotechnol. 2019;17(1):85. doi:10.1186/s12951-019-0517-8
119. Martinez-Lostao L, García-Alvarez F, Basáñez G, et al. Liposome-bound APO2L/TRAIL is an effective treatment in a rabbit model of rheumatoid arthritis. Arthritis Rheum. 2010;62(8):2272–2282. doi:10.1002/art.27501
120. Huang T, Peng L, Han Y, et al. Lipid nanoparticle-based mRNA vaccines in cancers: current advances and future prospects. Front Immunol. 2022;13:922301. doi:10.3389/fimmu.2022.922301
121. Jacob S, Rao R, Gorain B, Boddu SHS, Nair AB. Solid lipid nanoparticles and nanostructured lipid carriers for anticancer phytochemical delivery: advances, challenges, and future prospects. Pharmaceutics. 2025;17(8):1079. doi:10.3390/pharmaceutics17081079
122. Jia G, Han Y, An Y, et al. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials. 2018;178:302–316. doi:10.1016/j.biomaterials.2018.06.029
123. Cui J, Xu Y, Tu H, et al. Gather wisdom to overcome barriers: well-designed nano-drug delivery systems for treating gliomas. Acta Pharm Sin B. 2022;12(3):1100–1125. doi:10.1016/j.apsb.2021.08.013
124. Chai Z, Hu X, Wei X, et al. A facile approach to functionalizing cell membrane-coated nanoparticles with neurotoxin-derived peptide for brain-targeted drug delivery. J Control Release off J Control Release Soc. 2017;264:102–111. doi:10.1016/j.jconrel.2017.08.027
125. Umar S, Debnath K, Leung K, et al. Immunomodulatory properties of naïve and inflammation-informed dental pulp stem cell derived extracellular vesicles. Front Immunol. 2024;15:1447536. doi:10.3389/fimmu.2024.1447536
126. Wu T, Liu Y, Wang S, Shi C. MSC-derived extracellular vesicles: roles and molecular mechanisms for tissue repair. Int J Nanomed. 2025;20:7953–7974. doi:10.2147/IJN.S525394
127. Su J, Wei Q, Ma K, et al. P-MSC-derived extracellular vesicles facilitate diabetic wound healing via miR-145-5p/CDKN1A-mediated functional improvements of high glucose-induced senescent fibroblasts. Burns Trauma. 2023;11:tkad010. doi:10.1093/burnst/tkad010
128. Johnson J, Shojaee M, Mitchell Crow J, Khanabdali R. From mesenchymal stromal cells to engineered extracellular vesicles: a new therapeutic paradigm. Front Cell Dev Biol. 2021;9:705676. doi:10.3389/fcell.2021.705676
129. Kou M, Huang L, Yang J, et al. Mesenchymal stem cell-derived extracellular vesicles for immunomodulation and regeneration: a next generation therapeutic tool? Cell Death Dis. 2022;13(7):580. doi:10.1038/s41419-022-05034-x
130. Harrell CR, Jovicic N, Djonov V, Arsenijevic N, Volarevic V. Mesenchymal stem cell-derived exosomes and other extracellular vesicles as new remedies in the therapy of inflammatory diseases. Cells. 2019;8(12):1605. doi:10.3390/cells8121605
131. Grange C, Tapparo M, Bruno S, et al. Biodistribution of mesenchymal stem cell-derived extracellular vesicles in a model of acute kidney injury monitored by optical imaging. Int J Mol Med. 2014;33(5):1055–1063. doi:10.3892/ijmm.2014.1663
132. wei LJ, Wei L, Han Z, Chen Z. Mesenchymal stromal cells-derived exosomes alleviate ischemia/reperfusion injury in mouse lung by transporting anti-apoptotic miR-21-5p. Eur J Pharmacol. 2019;852:68–76. doi:10.1016/j.ejphar.2019.01.022
133. Chen L, Xiang B, Wang X, Xiang C. Exosomes derived from human menstrual blood-derived stem cells alleviate fulminant hepatic failure. Stem Cell Res Ther. 2017;8(1):9. doi:10.1186/s13287-016-0453-6
134. Wu J, Lin S, Wan B, Velani B, Zhu Y. Pyroptosis in Liver Disease: new Insights into Disease Mechanisms. Aging Dis. 2019;10(5):1094–1108. doi:10.14336/AD.2019.0116
135. Wang K, Jiang Z, Webster KA, et al. Enhanced Cardioprotection by Human Endometrium Mesenchymal Stem Cells Driven by Exosomal MicroRNA-21. Stem Cells Transl Med. 2017;6(1):209–222. doi:10.5966/sctm.2015-0386
136. Nojima H, Freeman CM, Schuster RM, et al. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J Hepatol. 2016;64(1):60–68. doi:10.1016/j.jhep.2015.07.030
137. Gatti S, Bruno S, Deregibus MC, et al. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol Dial Transplant off Publ Eur Dial Transpl Assoc - Eur Ren Assoc. 2011;26(5):1474–1483. doi:10.1093/ndt/gfr015
138. Shiue SJ, Rau RH, Shiue HS, et al. Mesenchymal stem cell exosomes as a cell-free therapy for nerve injury-induced pain in rats. Pain. 2019;160(1):210–223. doi:10.1097/j.pain.0000000000001395
139. Yu J, Liu X, Jin L, et al. Extracellular vesicles derived from menstrual blood-derived mesenchymal stem cells suppress inflammatory atherosclerosis by inhibiting NF-κB signaling. BMC Med. 2025;23(1):565. doi:10.1186/s12916-025-04390-7
140. Zhang Z, Liu L, Ti H, et al. Synovial fibroblast derived small extracellular vesicles miRNA15-29148 promotes articular chondrocyte apoptosis in rheumatoid arthritis. Bone Res. 2025;13(1):61. doi:10.1038/s41413-025-00430-3
141. Gibello L, D’Antico S, Salafia M, et al. First pilot case-control interventional study using autologous extracellular vesicles to treat chronic venous ulcers unresponsive to conventional treatments. Pharmacol Res. 2023;190:106718. doi:10.1016/j.phrs.2023.106718
142. Cao L, Xu H, Wang G, Liu M, Tian D, Yuan Z. Extracellular vesicles derived from bone marrow mesenchymal stem cells attenuate dextran sodium sulfate-induced ulcerative colitis by promoting M2 macrophage polarization. Int Immunopharmacol. 2019;72:264–274. doi:10.1016/j.intimp.2019.04.020
143. Wu XF, Ouyang ZJ, Feng LL, et al. Suppression of NF-κB signaling and NLRP3 inflammasome activation in macrophages is responsible for the amelioration of experimental murine colitis by the natural compound fraxinellone. Toxicol Appl Pharmacol. 2014;281(1):146–156. doi:10.1016/j.taap.2014.10.002
144. Dai S, Wei D, Wu Z, et al. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol Ther J Am Soc Gene Ther. 2008;16(4):782–790. doi:10.1038/mt.2008.1
145. Chen J, Tan Q, Yang Z, et al. Dendritic cell derived-extracellular vesicles engineered to express Interleukin-12 and Anti-CTLA-4 on their surface for combinational cancer immunotherapy. J Extracell Vesicles. 2025;14(4):e70068. doi:10.1002/jev2.70068
146. Zhu L, Kalimuthu S, Gangadaran P, et al. Exosomes derived from natural killer cells exert therapeutic effect in melanoma. Theranostics. 2017;7(10):2732–2745. doi:10.7150/thno.18752
147. Möller A, Lobb RJ. The evolving translational potential of small extracellular vesicles in cancer. Nat Rev Cancer. 2020;20(12):697–709. doi:10.1038/s41568-020-00299-w
148. Chen R, Kang Z, Li W, et al. Extracellular vesicle surface display of αPD-L1 and αCD3 antibodies via engineered late domain-based scaffold to activate T-cell anti-tumor immunity. J Extracell Vesicles. 2024;13(7):e12490. doi:10.1002/jev2.12490
149. Gao K, Xi W, Ni J, et al. Genetically modified extracellular vesicles loaded with activated gasdermin D potentially inhibit prostate-specific membrane antigen-positive prostate carcinoma growth and enhance immunotherapy. Biomaterials. 2025;315:122894. doi:10.1016/j.biomaterials.2024.122894
150. Wiklander OPB, Mamand DR, Mohammad DK, et al. Antibody-displaying extracellular vesicles for targeted cancer therapy. Nat Biomed Eng. 2024;8(11):1453–1468. doi:10.1038/s41551-024-01214-6
151. Lu YT, Chen TY, Lin HH, et al. Small extracellular vesicles engineered using click chemistry to express chimeric antigen receptors show enhanced efficacy in acute liver failure. J Extracell Vesicles. 2025;14(2):e70044. doi:10.1002/jev2.70044
152. Pei W, Zhang Y, Zhu X, et al. Multitargeted immunomodulatory therapy for viral myocarditis by engineered extracellular vesicles. ACS Nano. 2024;18(4):2782–2799. doi:10.1021/acsnano.3c05847
153. Warner KD, Hajdin CE, Weeks KM. Principles for targeting RNA with drug-like small molecules. Nat Rev Drug Discov. 2018;17(8):547–558. doi:10.1038/nrd.2018.93
154. Zhang J, Yang PL, Gray NS. Targeting cancer with small molecule kinase inhibitors. Nat Rev Cancer. 2009;9(1):28–39. doi:10.1038/nrc2559
155. Deng J, Grande F, Neamati N. Small molecule inhibitors of Stat3 signaling pathway. Curr Cancer Drug Targets. 2007;7(1):91–107. doi:10.2174/156800907780006922
156. McCormick F. Small-molecule inhibitors of cell signaling. Curr Opin Biotechnol. 2000;11(6):593–597. doi:10.1016/s0958-1669(00)00149-x
157. Singh AP, Biswas A, Shukla A, Maiti P. Targeted therapy in chronic diseases using nanomaterial-based drug delivery vehicles. Signal Transduct Target Ther. 2019;4:33. doi:10.1038/s41392-019-0068-3
158. Leader B, Baca QJ, Golan DE. Protein therapeutics: a summary and pharmacological classification. Nat Rev Drug Discov. 2008;7(1):21–39. doi:10.1038/nrd2399
159. Scaletti F, Hardie J, Lee YW, Luther DC, Ray M, Rotello VM. Protein delivery into cells using inorganic nanoparticle-protein supramolecular assemblies. Chem Soc Rev. 2018;47(10):3421–3432. doi:10.1039/c8cs00008e
160. Lee YW, Luther DC, Kretzmann JA, et al. Protein delivery into the cell cytosol using non-viral nanocarriers. Theranostics. 2019;9(11):3280–3292. doi:10.7150/thno.34412
161. Bai L, Liu Y, Guo K, et al. Ultrasound facilitates naturally equipped exosomes derived from macrophages and blood serum for orthotopic glioma treatment. ACS Appl Mater Interfaces. 2019;11(16):14576–14587. doi:10.1021/acsami.9b00893
162. Mesenchymal stem cells as natural biofactories for exosomes carrying miR-124a in the treatment of gliomas - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/29016843/.
163. Jiang L, Vader P, Schiffelers RM. Extracellular vesicles for nucleic acid delivery: progress and prospects for safe RNA-based gene therapy. Gene Ther. 2017;24(3):157–166. doi:10.1038/gt.2017.8
164. Zhu Z, Zhai Y, Hao Y, et al. Specific anti-glioma targeted-delivery strategy of engineered small extracellular vesicles dual-functionalised by Angiopep-2 and TAT peptides. J Extracell Vesicles. 2022;11(8):e12255. doi:10.1002/jev2.12255
165. Reddy SK, Ballal AR, Shailaja S, et al. Small extracellular vesicle-loaded bevacizumab reduces the frequency of intravitreal injection required for diabetic retinopathy. Theranostics. 2023;13(7):2241–2255. doi:10.7150/thno.78426
166. Kim YK, Hong Y, Bae YR, et al. Advantage of extracellular vesicles in hindering the CD47 signal for cancer immunotherapy. J Control Release off J Control Release Soc. 2022;351:727–738. doi:10.1016/j.jconrel.2022.09.042
167. Nguyen Cao TG, Truong Hoang Q, Kang JH, et al. Bioreducible exosomes encapsulating glycolysis inhibitors potentiate mitochondria-targeted sonodynamic cancer therapy via cancer-targeted drug release and cellular energy depletion. Biomaterials. 2023;301:122242. doi:10.1016/j.biomaterials.2023.122242
168. Guo R, Jiang D, Gai Y, et al. Chlorine 6-loaded goat milk-derived extracellular vesicles for Cerenkov luminescence-induced photodynamic therapy. Eur J Nucl Med Mol Imaging. 2023;50(2):508–524. doi:10.1007/s00259-022-05978-4
169. Nguyen Cao TG, Kang JH, Kang SJ, et al. Brain endothelial cell-derived extracellular vesicles with a mitochondria-targeting photosensitizer effectively treat glioblastoma by hijacking the blood‒brain barrier. Acta Pharm Sin B. 2023;13(9):3834–3848. doi:10.1016/j.apsb.2023.03.023
170. Nguyen Cao TG, Truong Hoang Q, Hong EJ, et al. Mitochondria-targeting sonosensitizer-loaded extracellular vesicles for chemo-sonodynamic therapy. J Control Release off J Control Release Soc. 2023;354:651–663. doi:10.1016/j.jconrel.2023.01.044
171. Nguyen Cao TG, Kang JH, Kim W, et al. Engineered extracellular vesicle-based sonotheranostics for dual stimuli-sensitive drug release and photoacoustic imaging-guided chemo-sonodynamic cancer therapy. Theranostics. 2022;12(3):1247–1266. doi:10.7150/thno.65516
172. Kang C, Ren X, Lee D, et al. Harnessing small extracellular vesicles for pro-oxidant delivery: novel approach for drug-sensitive and resistant cancer therapy. J Control Release off J Control Release Soc. 2024;365:286–300. doi:10.1016/j.jconrel.2023.11.031
173. Iyaswamy A, Thakur A, Guan XJ, et al. Fe65-engineered neuronal exosomes encapsulating corynoxine-B ameliorate cognition and pathology of Alzheimer’s disease. Signal Transduct Target Ther. 2023;8(1):404. doi:10.1038/s41392-023-01657-4
174. Rehman FU, Liu Y, Yang Q, et al. Heme Oxygenase-1 targeting exosomes for temozolomide resistant glioblastoma synergistic therapy. J Control Release off J Control Release Soc. 2022;345:696–708. doi:10.1016/j.jconrel.2022.03.036
175. Zheng X, Sun K, Liu Y, et al. Resveratrol-loaded macrophage exosomes alleviate multiple sclerosis through targeting microglia. J Control Release off J Control Release Soc. 2023;353:675–684. doi:10.1016/j.jconrel.2022.12.026
176. Piunti C, Micheli S, Giancaterino S, Fusco P, Boi C, Cimetta E. Microfluidic loading of verteporfin into extracellular vesicles for neuroblastoma therapy. Lab Chip. 2025;25(7):1718–1727. doi:10.1039/d4lc01103a
177. Lu J, Yang X, He C, et al. Rejuvenation of tendon stem/progenitor cells for functional tendon regeneration through platelet-derived exosomes loaded with recombinant Yap1. Acta Biomater. 2023;161:80–99. doi:10.1016/j.actbio.2023.02.018
178. Corrado C, Fontana S. Exosomes/extracellular vesicles and targeted tumor immunotherapy. Int J Mol Sci. 2024;25(10):5458. doi:10.3390/ijms25105458
179. Milbank E, Dragano N, Vidal-Gómez X, et al. Small extracellular vesicle targeting of hypothalamic AMPKα1 promotes weight loss in leptin receptor deficient mice. Metabolism. 2023;139:155350. doi:10.1016/j.metabol.2022.155350
180. Salazar-Puerta AI, Rincon-Benavides MA, Cuellar-Gaviria TZ, et al. Engineered extracellular vesicles derived from dermal fibroblasts attenuate inflammation in a murine model of acute lung injury. Adv Mater Deerfield Beach Fla. 2023;35(28):e2210579. doi:10.1002/adma.202210579
181. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
182. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
183. Jiao Y, Tang Y, Li Y, et al. Tumor cell-derived extracellular vesicles for breast cancer specific delivery of therapeutic P53. J Control Release off J Control Release Soc. 2022;349:606–616. doi:10.1016/j.jconrel.2022.07.020
184. Han Y, Zhu Y, Almuntashiri S, et al. Extracellular vesicle-encapsulated CC16 as novel nanotherapeutics for treatment of acute lung injury. Mol Ther. 2023;31(5):1346–1364. doi:10.1016/j.ymthe.2023.01.009
185. Jung JW, Kim JE, Kim E, et al. Liver-originated small extracellular vesicles with TM4SF5 target brown adipose tissue for homeostatic glucose clearance. J Extracell Vesicles. 2022;11(9):e12262. doi:10.1002/jev2.12262
186. Gao L, Sun Y, Zhang X, et al. Wnt3a-loaded extracellular vesicles promote alveolar epithelial regeneration after lung injury. Adv Sci Weinh Baden-Wurtt Ger. 2023;10(18):e2206606. doi:10.1002/advs.202206606
187. Liu W, Ota M, Tabushi M, Takahashi Y, Takakura Y. Development of allergic rhinitis immunotherapy using antigen-loaded small extracellular vesicles. J Control Release off J Control Release Soc. 2022;345:433–442. doi:10.1016/j.jconrel.2022.03.016
188. Liang X, Gupta D, Xie J, et al. Engineering of extracellular vesicles for efficient intracellular delivery of multimodal therapeutics including genome editors. Nat Commun. 2025;16(1):4028. doi:10.1038/s41467-025-59377-y
189. Dowdy SF. Overcoming cellular barriers for RNA therapeutics. Nat Biotechnol. 2017;35(3):222–229. doi:10.1038/nbt.3802
190. Lu W, Zhang J, Wu Y, Sun W, Jiang Z, Luo X. Engineered NF-κB siRNA-encapsulating exosomes as a modality for therapy of skin lesions. Front Immunol. 2023;14:1109381. doi:10.3389/fimmu.2023.1109381
191. Bontempi G, Terri M, Garbo S, et al. Restoration of WT1/miR-769-5p axis by HDAC1 inhibition promotes MMT reversal in mesenchymal-like mesothelial cells. Cell Death Dis. 2022;13(11):965. doi:10.1038/s41419-022-05398-0
192. Yang Z, Ji P, Li Z, et al. Improved extracellular vesicle-based mRNA delivery for familial hypercholesterolemia treatment. Theranostics. 2023;13(10):3467–3479. doi:10.7150/thno.82873
193. You Y, Tian Y, Yang Z, et al. Intradermally delivered mRNA-encapsulating extracellular vesicles for collagen-replacement therapy. Nat Biomed Eng. 2023;7(7):887–900. doi:10.1038/s41551-022-00989-w
194. Huang X, Wu W, Jing D, et al. Engineered exosome as targeted lncRNA MEG3 delivery vehicles for osteosarcoma therapy. J Control Release off J Control Release Soc. 2022;343:107–117. doi:10.1016/j.jconrel.2022.01.026
195. Born LJ, Chang KH, Shoureshi P, et al. HOTAIR-loaded mesenchymal stem/stromal cell extracellular vesicles enhance angiogenesis and wound healing. Adv Healthc Mater. 2022;11(5):e2002070. doi:10.1002/adhm.202002070
196. Fan L, Yao L, Li Z, et al. Exosome-based mitochondrial delivery of circRNA mSCAR alleviates sepsis by orchestrating macrophage activation. Adv Sci Weinh Baden-Wurtt Ger. 2023;10(14):e2205692. doi:10.1002/advs.202205692
197. Stranford DM, Simons LM, Berman KE, et al. Genetically encoding multiple functionalities into extracellular vesicles for the targeted delivery of biologics to T cells. Nat Biomed Eng. 2024;8(4):397–414. doi:10.1038/s41551-023-01142-x
198. Zickler AM, Liang X, Gupta D, et al. Novel endogenous engineering platform for robust loading and delivery of functional mRNA by extracellular vesicles. Adv Sci Weinh Baden-Wurtt Ger. 2024;11(42):e2407619. doi:10.1002/advs.202407619
199. Wang Z, Ma T, Bai G, et al. Adipose tissue-derived extracellular vesicles loaded with miR-141-3p regulate obesity-induced insulin resistance by targeting glycogen synthesis and gluconeogenesis. Int J Nanomed. 2025;20:5709–5726. doi:10.2147/IJN.S511842
200. Xie Y, Chai M, Xing Y, Zhou P, Wei P, Hua H. miRNA let-7f-5p-encapsulated labial gland MSC-derived EVs ameliorate experimental Sjögren’s syndrome by suppressing Th17 cells via targeting RORC/IL-17A signaling axis. J Nanobiotechnol. 2025;23(1):228. doi:10.1186/s12951-025-03308-y
201. Yang X, Hong S, Yan T, et al. MiR-146a engineered extracellular vesicles derived from mesenchymal stromal cells more potently attenuate ischaemia-reperfusion injury in lung transplantation. Clin Transl Med. 2025;15(4):e70298. doi:10.1002/ctm2.70298
202. Li L, Lam I, Wang J, Yu H, Chan C, Cai S. Epigenetic mechanism of iPSC-MSC-EVs in colonic epithelial cell pyroptosis in ulcerative colitis cell models via modulation of ELF3/miR-342-3p/KDM6B axis and histone methylation. Int Immunopharmacol. 2025;157:114704. doi:10.1016/j.intimp.2025.114704
203. Camões SP, Bulut O, Yazar V, et al. 3D-MSCs A151 ODN-loaded exosomes are immunomodulatory and reveal a proteomic cargo that sustains wound resolution. J Adv Res. 2022;41:113–128. doi:10.1016/j.jare.2022.01.013
204. Carney RP, Mizenko RR, Bozkurt BT, et al. Harnessing extracellular vesicle heterogeneity for diagnostic and therapeutic applications. Nat Nanotechnol. 2025;20(1):14–25. doi:10.1038/s41565-024-01774-3
205. Biagiotti S, Canonico B, Tiboni M, et al. Efficient and highly reproducible production of red blood cell-derived extracellular vesicle mimetics for the loading and delivery of RNA molecules. Sci Rep. 2024;14(1):14610. doi:10.1038/s41598-024-65623-y
206. Gopal A, Gangadaran P, Rajendran RL, et al. Extracellular vesicle mimetics engineered from mesenchymal stem cells and curcumin promote fibrosis regression in a mouse model of thioacetamide-induced liver fibrosis. Regen Ther. 2024;26:911–921. doi:10.1016/j.reth.2024.10.005
207. Liu Y, Ling Y, Tai W. Mechanical extrusion of the plasma membrane to generate ectosome-mimetic nanovesicles for lung targeting. Mol Pharm. 2025;22(1):304–315. doi:10.1021/acs.molpharmaceut.4c00927
208. Lin M, Lei S, Chai Y, et al. Immunosuppressive microvesicles-mimetic derived from tolerant dendritic cells to target T-lymphocytes for inflammation diseases therapy. J Nanobiotechnol. 2024;22(1):201. doi:10.1186/s12951-024-02470-z
209. Liu S, Wan G, Jiang T, et al. Engineered biomimetic nanovesicles-laden multifunctional hydrogel enhances targeted therapy of diabetic wound. Mater Today Bio. 2024;29:101330. doi:10.1016/j.mtbio.2024.101330
210. Chen R, Bhavsar C, Lourie R, Li S, Wu SY. Development of an innovative extracellular vesicle mimetic delivery platform for efficient miRNA delivery to tumours. Biomaterials. 2025;321:123282. doi:10.1016/j.biomaterials.2025.123282
211. Wang S, Mao Y, Rong S, et al. Engineering magnetic extracellular vesicles mimetics for enhanced targeting chemodynamic therapy to overcome ovary cancer. ACS Appl Mater Interfaces. 2024;16(30):39021–39034. doi:10.1021/acsami.4c06862
212. Van der Meel R, Mham F, Vader P, Van Solinge WW, Eniola-Adefeso O, Schiffelers RM. Extracellular vesicles as drug delivery systems: lessons from the liposome field. J Control Release. 2014;195:72–85. doi:10.1016/j.jconrel.2014.07.049
213. Kay MA. State-of-the-art gene-based therapies: the road ahead. Nat Rev Genet. 2011;12(5):316–328. doi:10.1038/nrg2971
214. Urits I, Swanson D, Swett MC, et al. A review of patisiran (ONPATTRO®) for the treatment of polyneuropathy in people with hereditary transthyretin amyloidosis. Neurol Ther. 2020;9(2):301–315. doi:10.1007/s40120-020-00208-1
215. Wesselhoeft RA, Kowalski PS, Anderson DG. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat Commun. 2018;9(1):2629. doi:10.1038/s41467-018-05096-6
216. Dangouloff T, Servais L. Clinical evidence supporting early treatment of patients with spinal muscular atrophy: current perspectives. Ther Clin Risk Manag. 2019;15:1153–1161. doi:10.2147/TCRM.S172291
217. McKay PF, Hu K, Blakney AK, et al. Self-amplifying RNA SARS-CoV-2 lipid nanoparticle vaccine candidate induces high neutralizing antibody titers in mice. Nat Commun. 2020;11(1):3523. doi:10.1038/s41467-020-17409-9
218. Gillmore JD, Gane E, Taubel J, et al. CRISPR-Cas9 in vivo gene editing for transthyretin amyloidosis. N Engl J Med. 2021;385(6):493–502. doi:10.1056/NEJMoa2107454
219. Ruan GX, Barry E, Yu D, Lukason M, Cheng SH, Scaria A. CRISPR/Cas9-mediated genome editing as a therapeutic approach for Leber congenital amaurosis 10. Mol Ther J Am Soc Gene Ther. 2017;25(2):331–341. doi:10.1016/j.ymthe.2016.12.006
220. Lee H, Lytton-Jean AKR, Chen Y, et al. Molecularly self-assembled nucleic acid nanoparticles for targeted in vivo siRNA delivery. Nat Nanotechnol. 2012;7(6):389–393. doi:10.1038/nnano.2012.73
221. Bujold KE, Hsu JCC, Sleiman HF. Optimized DNA “Nanosuitcases” for encapsulation and conditional release of siRNA. J Am Chem Soc. 2016;138(42):14030–14038. doi:10.1021/jacs.6b08369
222. Fu X, Chen T, Song Y, et al. mRNA delivery by a pH-responsive DNA nano-hydrogel. Small Weinh Bergstr Ger. 2021;17(29):e2101224. doi:10.1002/smll.202101224
223. Parsons MF, Allan MF, Li S, et al. 3D RNA-scaffolded wireframe origami. Nat Commun. 2023;14(1):382. doi:10.1038/s41467-023-36156-1
224. Eid A, Mahfouz MM. Genome editing: the road of CRISPR/Cas9 from bench to clinic. Exp Mol Med. 2016;48(10):e265. doi:10.1038/emm.2016.111
225. Meisel R. CRISPR-Cas9 gene editing for sickle cell disease and β-Thalassemia. N Engl J Med. 2021;384(23):e91. doi:10.1056/NEJMc2103481
226. Stadtmauer EA, Fraietta JA, Davis MM, et al. CRISPR-engineered T cells in patients with refractory cancer. Science. 2020;367(6481):eaba7365. doi:10.1126/science.aba7365
227. Rajan A, Shrivastava S, null J, Kumar A, Singh AK, Arora PK. CRISPR-Cas system: from diagnostic tool to potential antiviral treatment. Appl Microbiol Biotechnol. 2022;106(18):5863–5877. doi:10.1007/s00253-022-12135-2
228. Zhao K, Yan Y, Jin XK, et al. An orally administered gene editing nanoparticle boosts chemo-immunotherapy in colorectal cancer. Nat Nanotechnol. 2025. doi:10.1038/s41565-025-01904-5
229. Ling S, Zhang X, Dai Y, et al. Customizable virus-like particles deliver CRISPR-Cas9 ribonucleoprotein for effective ocular neovascular and Huntington’s disease gene therapy. Nat Nanotechnol. 2025. doi:10.1038/s41565-024-01851-7
230. Liyanage W, Kannan G, Kannan S, Kannan RM. Efficient intracellular delivery of CRISPR-Cas9 ribonucleoproteins using dendrimer nanoparticles for robust genomic editing. Nano Today. 2025;61:102654. doi:10.1016/j.nantod.2025.102654
231. World Health Organization. WHO Expert Committee on Biological Standardization. World Health Organ Tech Rep Ser. 2014;2014(987):1–266, backcover.
232. Aldosari BN, Alfagih IM, Almurshedi AS. Lipid nanoparticles as delivery systems for RNA-Based vaccines. Pharmaceutics. 2021;13(2):206. doi:10.3390/pharmaceutics13020206
233. Del Poeta M, Wormley FL, Lin X. Host populations, challenges, and commercialization of cryptococcal vaccines. PLoS Pathog. 2023;19(2):e1011115. doi:10.1371/journal.ppat.1011115
234. Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines - a new era in vaccinology. Nat Rev Drug Discov. 2018;17(4):261–279. doi:10.1038/nrd.2017.243
235. Thompson MG, Burgess JL, Naleway AL, et al. Interim estimates of vaccine effectiveness of BNT162b2 and mRNA-1273 COVID-19 vaccines in preventing sars-cov-2 infection among health care personnel, first responders, and other essential and frontline workers - eight U.S. locations, December 2020-March 2021. MMWR Morb Mortal Wkly Rep. 2021;70(13):495–500. doi:10.15585/mmwr.mm7013e3
236. Dai W, Xing M, Sun L, et al. Lipid nanoparticles as adjuvant of norovirus VLP vaccine augment cellular and humoral immune responses in a TLR9- and type I IFN-dependent pathway. J Virol. 2024;98(12):e0169924. doi:10.1128/jvi.01699-24
237. Wang T, Zhen Y, Ma X, Wei B, Li S, Wang N. Mannosylated and lipid A-incorporating cationic liposomes constituting microneedle arrays as an effective oral mucosal HBV vaccine applicable in the controlled temperature chain. Colloids Surf B Biointerfaces. 2015;126:520–530. doi:10.1016/j.colsurfb.2015.01.005
238. Wang L, Ramirez A, Felgner J, et al. Development of a single-dose Q fever vaccine with an injectable nanoparticle-loaded hydrogel: effect of sustained co-delivery of antigen and adjuvant. Drug Deliv. 2025;32(1):2476144. doi:10.1080/10717544.2025.2476144
239. Hu Z, Ott PA, Wu CJ. Towards personalized, tumour-specific, therapeutic vaccines for cancer. Nat Rev Immunol. 2018;18(3):168–182. doi:10.1038/nri.2017.131
240. Zhang J, Huang Q, Yang H, et al. Lipid coating of mesoporous silica nanoparticles leads to efficient antigen delivery to lymph nodes for cancer vaccination. ACS Appl Bio Mater. 2025. doi:10.1021/acsabm.5c00403
241. Xu X, Diao L, Wang J, et al. Maximizing efficacy of cancer nanovaccines and immune cells landscape in responders and non-responders to immunotherapy. Adv Sci Weinh Baden-Wurtt Ger. 2025. doi:10.1002/advs.202416756
242. Lang F, Schrörs B, Löwer M, Türeci Ö, Sahin U. Identification of neoantigens for individualized therapeutic cancer vaccines. Nat Rev Drug Discov. 2022;21(4):261–282. doi:10.1038/s41573-021-00387-y
243. Chen MH, Jiang J, Chen H, et al. Reinforcing cancer immunotherapy with engineered porous hollow mycobacterium tuberculosis loaded with tumor neoantigens. J Immunother Cancer. 2025;13(2):e010150. doi:10.1136/jitc-2024-010150
244. Liang S, Gao S, Fu S, et al. Screening natural cholesterol analogs to assemble self-adjuvant lipid nanoparticles for antigens tagging guided therapeutic tumor vaccine. Adv Mater Deerfield Beach Fla. 2025. doi:10.1002/adma.202419182
245. Xu Y, Wang B, Huang Y, et al. Targeting antigen-presenting cells to enhance the tumor-spleen immunity cycle through liposome-neoantigen vaccine. Adv Sci Weinh Baden-Wurtt Ger. 2025. doi:10.1002/advs.202500021
246. Yin T, Chen H, Ma A, et al. Cleavable collagenase-assistant nanosonosensitizer for tumor penetration and sonodynamic therapy. Biomaterials. 2023;293:121992. doi:10.1016/j.biomaterials.2022.121992
247. Huang HY, Chen LQ, Sun W, et al. Collagenase IV and clusterin-modified polycaprolactone-polyethylene glycol nanoparticles for penetrating dense tumor tissues. Theranostics. 2021;11(2):906–924. doi:10.7150/thno.47446
248. Raeesi V, Chan WCW. Improving nanoparticle diffusion through tumor collagen matrix by photo-thermal gold nanorods. Nanoscale. 2016;8(25):12524–12530. doi:10.1039/c5nr08463f
249. Chen Y, Yang C, Mao J, Li H, Ding J, Zhou W. Spermine modified polymeric micelles with pH-sensitive drug release for targeted and enhanced antitumor therapy. RSC Adv. 2019;9(20):11026–11037. doi:10.1039/c9ra00834a
250. Li X, Wu M, Wu Y, et al. Multifunctional nanodrug for simultaneously combating chemoresistance and immunosuppression in Fusobacterium nucleatum-associated colorectal cancer. Acta Biomater. 2025;195:406–420. doi:10.1016/j.actbio.2025.02.013
251. Song Z, Wang Q, Xiong H, et al. Bionic gene delivery system activates tumor autophagy and immunosuppressive niche to sensitize anti-PD-1 treatment against STK11-mutated lung adenocarcinoma. J Nanobiotechnol. 2025;23(1):312. doi:10.1186/s12951-025-03404-z
252. Giulimondi F, Digiacomo L, Renzi S, et al. Optimizing transfection efficiency in CAR-T cell manufacturing through multiple administrations of lipid-based nanoparticles. ACS Appl Bio Mater. 2024;7(6):3746–3757. doi:10.1021/acsabm.4c00103
253. Lim J, Oh D, Cheng M, et al. Enhancing chimeric antigen receptor T-Cell generation via microfluidic mechanoporation and lipid nanoparticles. Small Weinh Bergstr Ger. 2025;21(17):e2410975. doi:10.1002/smll.202410975
254. Van Delen M, Derdelinckx J, Wouters K, Nelissen I, Cools N. A systematic review and meta-analysis of clinical trials assessing safety and efficacy of human extracellular vesicle-based therapy. J Extracell Vesicles. 2024;13(7):e12458. doi:10.1002/jev2.12458
255. Witwer KW, Buzás EI, Bemis LT, et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J Extracell Vesicles. 2013;2. doi:10.3402/jev.v2i0.20360
256. He J, Li H, Mai J, et al. Profiling extracellular vesicle surface proteins with 10 µL peripheral plasma within 4 h. J Extracell Vesicles. 2023;12(9):e12364. doi:10.1002/jev2.12364
257. Weerakkody JS, Tseng T, Topper M, et al. Photosensitive nanoprobes for rapid isolation and size-specific enrichment of synthetic and extracellular vesicle subpopulations. Adv Funct Mater. 2024;34(34):2400390. doi:10.1002/adfm.202400390
258. Zhang J, Gao Z, Xiao W, et al. A simplified and efficient extracellular vesicle-based proteomics strategy for early diagnosis of colorectal cancer. Chem Sci. 2024;15(44):18419–18430. doi:10.1039/d4sc05518g
259. Torres Crigna A, Fricke F, Nitschke K, et al. Inter-laboratory comparison of extracellular vesicle isolation based on ultracentrifugation. Transfus Med Hemotherapy Off Organ Dtsch Ges Transfusionsmedizin Immunhamatologie. 2021;48(1):48–59. doi:10.1159/000508712
260. Ma Y, Dong S, Grippin AJ, et al. Engineering therapeutical extracellular vesicles for clinical translation. Trends Biotechnol. 2025;43(1):61–82. doi:10.1016/j.tibtech.2024.08.007
261. Burgelman M, Vandendriessche C, Vandenbroucke RE. Extracellular vesicles: a double-edged sword in sepsis. Pharmaceuticals. 2021;14(8):829. doi:10.3390/ph14080829
262. Okoye IS, Coomes SM, Pelly VS, et al. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity. 2014;41(1):89–103. doi:10.1016/j.immuni.2014.05.019
263. Buzas EI. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23(4):236–250. doi:10.1038/s41577-022-00763-8
264. Budden CF, Gearing LJ, Kaiser R, Standke L, Hertzog PJ, Latz E. Inflammasome-induced extracellular vesicles harbour distinct RNA signatures and alter bystander macrophage responses. J Extracell Vesicles. 2021;10(10):e12127. doi:10.1002/jev2.12127
265. Shlomovitz I, Erlich Z, Arad G, et al. Proteomic analysis of necroptotic extracellular vesicles. Cell Death Dis. 2021;12(11):1059. doi:10.1038/s41419-021-04317-z
266. Marar C, Starich B, Wirtz D. Extracellular vesicles in immunomodulation and tumor progression. Nat Immunol. 2021;22(5):560–570. doi:10.1038/s41590-021-00899-0
267. Huber V, Fais S, Iero M, et al. Human colorectal cancer cells induce T-cell death through release of proapoptotic microvesicles: role in immune escape. Gastroenterology. 2005;128(7):1796–1804. doi:10.1053/j.gastro.2005.03.045
268. Menck K, Sönmezer C, Worst TS, et al. Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J Extracell Vesicles. 2017;6(1):1378056. doi:10.1080/20013078.2017.1378056
269. Buzás EI, Tóth EÁ, Sódar BW, Szabó-Taylor KÉ. Molecular interactions at the surface of extracellular vesicles. Semin Immunopathol. 2018;40(5):453–464. doi:10.1007/s00281-018-0682-0
270. Lindenbergh MFS, Stoorvogel W. Antigen presentation by extracellular vesicles from professional antigen-presenting cells. Annu Rev Immunol. 2018;36:435–459. doi:10.1146/annurev-immunol-041015-055700
271. De Jong WH, Borm PJA. Drug delivery and nanoparticles: Applications and hazards. Int J Nanomed. 2008;3(2):133–149. doi:10.2147/ijn.s596
272. Li G, Chen T, Dahlman J, et al. Current challenges and future directions for engineering extracellular vesicles for heart, lung, blood and sleep diseases. J Extracell Vesicles. 2023;12(2):e12305. doi:10.1002/jev2.12305
273. Extracellular vesicle manufacture via FACTORY: fully automated collection technology and optimum machinery for clinical translational applications - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/40750491/.
274. Andriolo G, Provasi E, Brambilla A, et al. GMP-grade methods for cardiac progenitor cells: cell bank production and quality control. Methods Mol Biol Clifton NJ. 2021;2286:131–166. doi:10.1007/7651_2020_286
275. Del Saz-Lara A, López de Las Hazas MC, Tomé-Carneiro J, et al. Milk and cheesemaking whey as a sustainable source of extracellular vesicles: exploring large-scale isolation methods with industrial and potential therapeutic applications. Pharmacol Res. 2025;220:107925. doi:10.1016/j.phrs.2025.107925
276. Kim Y, Jeon S, Lee K, Shin S. Advanced extracellular vesicle isolation: a hybrid electrokinetic-tangential flow filtration approach for improved yield, purity, and scalability. Anal Chem. 2025;97(31):16759–16768. doi:10.1021/acs.analchem.5c01168
277. Mao YA, Shi X, Sun P, et al. Nanomedicines for cardiovascular diseases: lessons learned and pathways forward. Biomaterials. 2025;320:123271. doi:10.1016/j.biomaterials.2025.123271
278. Noury H, Rahdar A, Romanholo Ferreira LF, Jamalpoor Z. AI-driven innovations in smart multifunctional nanocarriers for drug and gene delivery: a mini-review. Crit Rev Oncol Hematol. 2025;210:104701. doi:10.1016/j.critrevonc.2025.104701
279. Sanna V, Sechi M. Therapeutic potential of targeted nanoparticles and perspective on nanotherapies. ACS Med Chem Lett. 2020;11(6):1069–1073. doi:10.1021/acsmedchemlett.0c00075
280. Aram E, Moeni M, Abedizadeh R, et al. Smart and multi-functional magnetic nanoparticles for cancer treatment applications: clinical challenges and future prospects. Nanomaterials. 2022;12(20):3567. doi:10.3390/nano12203567
281. Kumar A, Lunawat AK, Kumar A, et al. Recent trends in nanocarrier-based drug delivery system for prostate cancer. AAPS Pharm Sci Tech. 2024;25(3):55. doi:10.1208/s12249-024-02765-2
282. Tiwari A, Widodo, Krisnawati DI, Tzou K, Kuo T. Machine learning for extracellular vesicles enables diagnostic and therapeutic nanobiotechnology. J Nanobiotechnol. 2026;24(1). doi:10.1186/s12951-025-03952-4
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Mir-218-5p from Extracellular Vesicles of Endometrium in Patients with Recurrent Implantation Failure Impairs Pre-Implantation Embryo Development

Cai L, Lv M, Wei J, Liu C, Li Y, Liao Z, Li T, Zhang H, Xi L, Sui C
International Journal of Nanomedicine 2025, 20:5661-5679
Published Date: 1 May 2025