Back to Journals » International Journal of Nanomedicine » Volume 21
Engineering Extracellular Vesicles for Tumor Targeted Therapy: Source Optimization, Modification, and Clinical Application
Authors Sui J, Qin H, Zhang Z, Lv X, Lin X, Liu Z, Zhao X, Liu X, Zhang H ![]()
Received 4 January 2026
Accepted for publication 17 March 2026
Published 9 April 2026 Volume 2026:21 592579
DOI https://doi.org/10.2147/IJN.S592579
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Misra
Jiaxin Sui,1– 5,* HanBin Qin,2– 6,* Zile Zhang,2– 5,7 Xiaojing Lv,1– 5 Xinhua Lin,2– 6 Zhonglu Liu,2– 5 Xin Zhao,2– 5 Xuexia Liu,8 Hua Zhang2– 5
1Qingdao University Medical College, Qingdao University, Qingdao, Shandong Province, People’s Republic of China; 2Department of Head and Neck Surgery, Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, Shandong Province, People’s Republic of China; 3Shandong Provincial Key Laboratory of Neuroimmune Interaction and Regulation, Yantai, Shandong Province, People’s Republic of China; 4Shandong Provincial Clinical Research Center for Otorhinolaryngologic Diseases, Yantai, Shandong Province, People’s Republic of China; 5Yantai Key Laboratory of Otorhinolaryngologic Diseases, Yantai, Shandong Province, People’s Republic of China; 6School of Clinical Medicine, Shandong Second Medical University, Weifang, Shandong Province, People’s Republic of China; 7Second Clinical Medical College, Binzhou Medical University, Yantai, Shandong Province, People’s Republic of China; 8Shandong Stem Cell Engineering Technology Research Center, Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, Shandong Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xuexia Liu, Email [email protected] Hua Zhang, Email [email protected]
Abstract: Cancer remains a leading cause of global morbidity and mortality, yet conventional therapies, including surgery, radiotherapy, and chemotherapy, are often limited by invasiveness, systemic toxicity, and drug resistance. In this context, extracellular vesicles (EVs) have emerged as a promising cell-free nanotherapeutic platform. As endogenous nanocarriers, EVs enable precise, targeted delivery of diverse bioactive cargoes (eg, nucleic acids, chemotherapeutics, immunomodulators) to tumor tissues, thereby enhancing therapeutic efficacy while minimizing off-target effects, which is the key advantages for their application in tumor targeted therapy. This review systematically summarizes the characteristics of animal-derived and plant-derived EVs and highlights their translational applications in multiple cancers via immune activation, targeted delivery, tumor microenvironment remodeling, and anti-angiogenesis. We further introduce advanced bioengineering strategies for EV modification to optimize cargo loading, targeting specificity, and in vivo stability, particularly frontier innovations such as artificial intelligence-assisted design and microfluidic manufacturing that improve the precision, controllability, and scalability of engineered EVs. Compared to synthetic nanocarriers, EVs exhibit unique advantages, including excellent biocompatibility, low immunogenicity, and superior ability to cross biological barriers. However, the clinical application of EV-based therapies faces notable challenges, including EV heterogeneity, scalability of production, standardization of characterization methods, cargo loading efficiency, and long-term safety concerns. This review emphasizes the transformative potential of engineered EVs in advancing tumor targeted therapy and improving outcomes for patients with refractory or metastatic tumors.
Keywords: extracellular vesicles, EVs, engineered EVs, tumor targeted therapy, tumor microenvironment, clinical translation
Introduction
Cancer incidence and mortality have surged globally in recent years, presenting a major obstacle to extending life expectancy. Notably, there is an increasing trend of cancer affecting younger populations.1,2 Consequently, cancer treatment has emerged as one of the most pressing challenges of our time. Conventional therapeutic strategies, including surgical resection, chemotherapy, radiotherapy, as well as emerging technologies such as photodynamic therapy and acoustic power therapy, are utilized for disease management.3 While these approaches can partially control cancer progression, their effectiveness is often limited and frequently accompanied by severe adverse effects, such as chemotherapy-induced leukopenia, hypothyroidism, and elevated alanine or glutamine transaminases.3 These limitations hinder the realization of the growing demand for precision medicine. Recent research highlights the pivotal role of extracellular vesicles (EVs) in tumor angiogenesis, invasion, metastasis, drug resistance, and remodeling of the tumor microenvironment (TME), positioning them as highly promising therapeutic targets for cancer therapy.4
EVs are a heterogeneous population of lipid bilayer-delimited particles naturally released from cells, typically ranging in diameter from 40 nm to 160 nm and originating from endosomes.5 Characterized by a lipid bilayer structure, they encapsulate diverse cellular components such as DNA, RNA, proteins, and other biomolecules.6 Functioning as signaling vesicles, EVs mediate intercellular communication via autocrine, endocrine, paracrine, or distal secretion pathways,7 thereby participating in numerous physiological and pathological processes. These properties underpin the significant potential of EVs for therapeutic applications. However, a majority limitation is that most natural EVs are sequestered by the liver and spleen and subsequently cleared by the immune system before reaching target sites. Furthermore, the inherent complexity of natural EVs can compromise drug delivery efficacy.
Engineered EVs, as a type of EVs, are functionally enhanced nanocarriers generated by modifying natural secreted EVs via techniques including electroporation, freeze-thaw cycles, and sonication. Compared with unmodified native EVs, they display substantially improved drug loading capacity, targeting specificity, and resistance to rapid in vivo clearance.8 As a promising drug delivery system, engineered EVs hold core advantages over other synthetic nanocarriers such as liposomes and polymeric nanoparticles, primarily due to their natural biological origin as inherent intercellular communication messengers. When derived from autologous or same-species cells,9 they exhibit exceptionally low immunogenicity and high biocompatibility, thereby reducing rapid immune clearance. They also possess intrinsic targeting properties and strong membrane penetration capacity, allowing efficient traversal of complex physiological barriers including the blood-brain barrier. Furthermore, both their surface components and internal cargo can be flexibly modified, and source cells themselves can be strategically engineered, enabling versatile multifunctionalization.10–12 Nevertheless, several critical challenges remain to be addressed for clinical translation, most notably their high inherent heterogeneity, limited scalability of large-scale production, and difficulties in quality control and standardization. These distinctive advantages and unresolved trade-offs together underscore the importance of developing rational and systematic engineering strategies.
This review distinguishes itself from previously published articles in several critical aspects. Unlike most existing reviews that focus predominantly on mammalian EVs or single engineering strategies, this work provides a system-level integration of the entire engineered EVs pipeline, ranging from source selection to clinical translation. We conduct a comparative analysis of animal-derived and plant-derived EVs, emphasizing their unique advantages in scalability and biocompatibility. Moreover, this review uniquely highlights the emerging role of AI-guided design in optimizing cargo loading and targeting specificity, a forward-looking perspective that has not been comprehensively addressed in the current literature. These features collectively enhance the novelty and practical value of this review. (Figure 1)
 |
Figure 1 Schematic roadmap illustrating the overall structure and logical framework of this review on engineered EVs for tumor-targeted therapy. The review systematically summarizes five core sections: EV sources including animal-derived and plant-derived extracellular vesicles; engineering modifications including intracellular cargo loading and extracellular targeting strategies; tumor-specific design principles for different cancer types; anti-tumor therapeutic applications including targeted delivery, immune activation, tumor microenvironment remodeling, and anti-angiogenesis; summary and future perspectives focusing on current challenges and translational prospects involving artificial intelligence-assisted design, microfluidic manufacturing, and clinical translation. |
Source of EVs
EVs of diverse sources exhibit notable differences in cellular composition, modes of intercellular communication, and roles in both physiological and pathological processes. Based on their biological origin, EVs can be broadly categorized into two main types: those derived from animal cells and those derived from plant tissues. In accordance with MISEV2023 guidelines, we use the term “extracellular vesicles” as the generic descriptor. (Table 1)
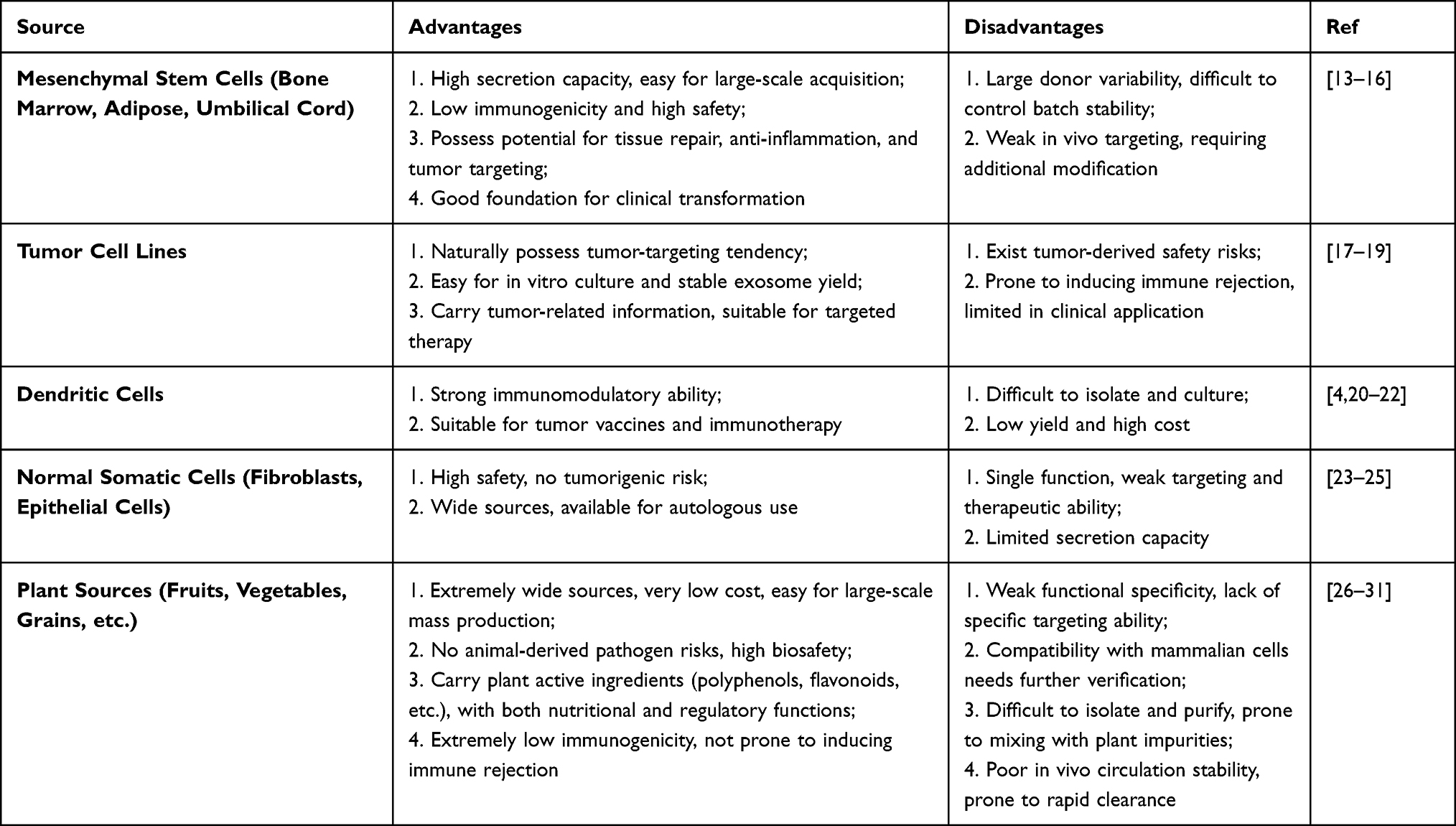 |
Table 1 Advantages and Disadvantages of Different EVs Sources |
EVs of Animal Origin
Animal-derived EVs are predominantly secreted by mesenchymal stem cells (MSCs), tumor cells, macrophages, natural killer cells (NK cells), dendritic cells (DCs) and other cell types. These EVs exhibit different biological functions depending on their cellular origin (Figure 2).
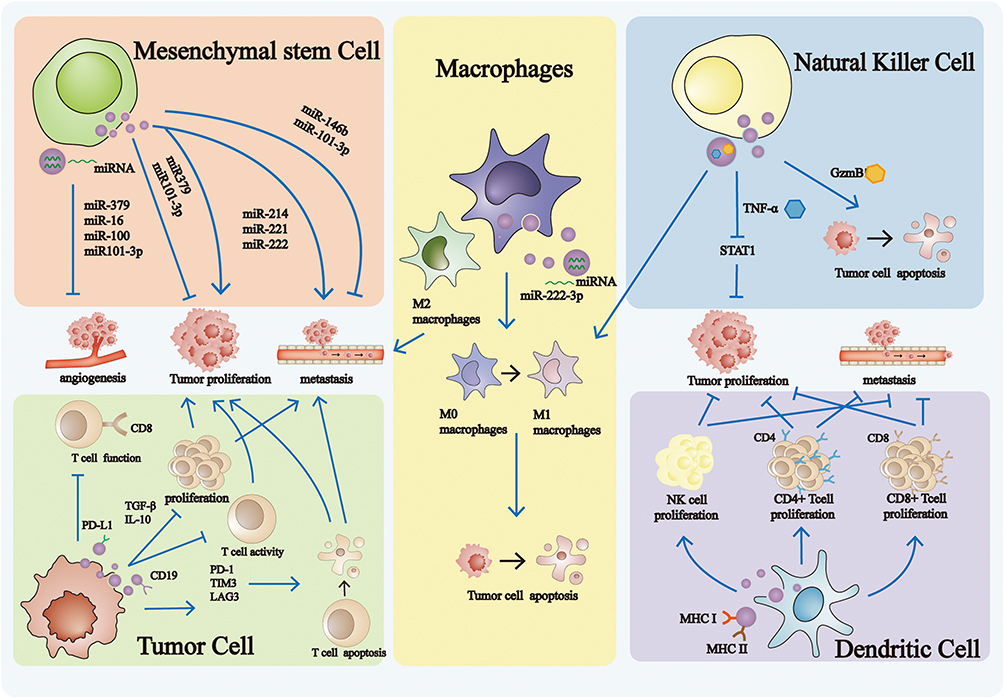 |
Figure 2 Mechanism of Action of Animal-derived extracellular vesicles. MSC-derived EVs regulate tumor progression by modulating of angiogenesis, metastasis, and proliferation. Macrophages-derived EVs influence tumor behavior by promoting metastasis and inducing tumor cell apoptosis. NK cell-derived EVs suppress tumor growth by inducing apoptosis and inhibiting proliferation. Tumor cells-derived EVs contribute to tumor regulation by facilitating metastasis and enhancing proliferation. DC-derived EVs modulate tumor development by involvement in metastasis and regulation of proliferation activity. |
MSC-derived EVs play a significant role in regulating tumor progression by modulating the TME, influencing processes such as tumor cell proliferation, invasion, migration, and apoptosis.13 Currently, research on bone marrow MSC (BM-MSC)-derived EVs has primarily focused on their regenerative capacities, including suppressing inflammation, promoting neovascularization, and protecting hepatocytes and cardiomyocytes from death.32 However, their impact on tumor biology is less explored. Intriguingly, despite this limited focus, existing studies reveal that BM-MSC-derived EVs can paradoxically promote tumor development by enhancing tumor progression and maintaining cancer stemness.32 For instance, Yao et al reported that circ_0030167, highly expressed in BM-MSC-derived EVs, functions as a sponge for miR-338-5p. This interaction elevates Wif1 expression and suppresses the Wnt/β-catenin signaling pathway, ultimately driving pancreatic cancer progression and stemness. In contrast to the pro-tumorigenic effects observed with BM-MSC-derived EVs, EVs derived from human umbilical cord MSCs (hUC-MSCs) exhibit potent anti-tumor effects. These EVs are enriched with tumor-suppressive miRNAs, such as miR-21-5p and miR-128-3p,33,34 which effectively inhibit tumor proliferation, migration, and invasion by downregulating targets like ZNF367 and Galectin-3. Furthermore, Xu et al demonstrated that hUC-MSCs-derived EVs overexpressing miR-451a can reduce ADAM10 levels, thereby inhibiting epithelial-mesenchymal transition (EMT) in tumor cells14 and attenuating tumor progression. Thus, hUC-MSCs-derived EVs represent a source of miRNAs that robustly suppress key oncogenic processes. However, their therapeutic translation faces challenges: collection requires ethical approval and donor consent, sample variability can lead to inconsistencies in EV characteristics, stable culture conditions are necessary, and EV production yield is relatively low, complicating large-scale isolation.14 Given the limitations of hUC-MSCs-derived EVs, adipose-derived stem cells (ADSCs)-derived EVs emerge as a promising alternative source. ADSCs possess robust proliferative capacity, strong immunomodulatory properties, and critically, high EV yield. Their EVs are easily obtainable and retain the beneficial immunomodulatory properties of the parent cells.13 Importantly, ADSCs-derived EVs can effectively deliver therapeutic cargo. For example, miR-145, a well-known tumor suppressor, inhibits prostate cancer progression by activating the caspase-3/7-mediated apoptotic pathway and suppressing the anti-apoptotic activity of Bcl-xL. Takahara et al demonstrated that delivering miR-145 via ADSCs-derived EVs significantly inhibited the proliferation of human prostate cancer cells.15 These findings collectively highlight the potential of ADSCs-derived EVs as valuable adjuvants in cancer treatment.
Tumor cell-derived EVs, including those from lung cancer, ovarian cancer, breast cancer, head and neck squamous cell carcinoma (HNSCC), oral squamous cell carcinoma (OSCC), osteosarcoma (OS), and lymphoma (LYM), are enriched in immunosuppressive molecules. For instance, EVs derived from B-cell lymphoma can transiently activate CD19-specific CAR T cells by carrying CD19 antigens. However, prolonged exposure leads to T cell apoptosis and conversion into regulatory T cells (Tregs) due to the presence of immunosuppressive molecules such as TGF-β, PD-L1, TIM3, and LAG3. Inhibition of the TGF-β/Smad3 signaling pathway can reverse these effects, suggesting that combining CAR T cell therapy with TGF-β inhibitors may present a novel therapeutic strategy for B-cell lymphoma.17,35,36 Additionally, colorectal cancer cell-derived EVs contain high levels of ADAM17, which disrupts vascular endothelial cadherin localization on vascular endothelial cells, increases vascular permeability, and facilitates the formation of a pre-metastatic niche.18 Notably, these tumor cell-derived EVs exhibit prolonged circulation in the bloodstream and retain tumor heterogeneity. Crucially, their intrinsic tumor-targeting capabilities,19 while exploited by the tumor for its own progression, present a unique opportunity. These properties make them potentially ideal natural vehicles for delivering therapeutic agents (eg, drugs, siRNAs, or immune modulators) specifically to tumor cells or the tumor microenvironment.
EVs derived from immune cells-such as macrophages, NK cells, and DCs-play crucial roles in modulating immune responses. Specifically, macrophage-derived EVs are involved in multiple biological processes, including regulating inflammation, promoting angiogenesis, combating infection, modulating tissue fibrosis, and enhancing anti-tumor immunity.4 Notably, EVs from classically activated macrophages (M1) have been shown to promote M1 polarization and enhance the efficacy of anti-programmed cell death receptor ligand 1 (anti-PD-L1) therapy in cancer.20 Similarly, NK cell-derived EVs are of great interest due to their ability to selectively kill tumor cells and activated immune cells. They achieve this by delivering cytotoxic proteins (eg, perforin and granzymes), cytokines (eg, IFN-γ and TNF-α), and activating receptors (eg, NKG2D), while sparing resting cells. These EVs offer several advantages: natural targeting capabilities, systemic distribution via the circulatory system, and enhanced uptake efficiency in the acidic tumor microenvironment. Their potent anti-tumor effects stem from a dual mechanism of action—direct cytotoxicity mediated by the FasL/Fas pathway and perforin, combined with immunomodulation through NKG2D-mediated tumor recognition. These properties make them promising candidates for anti-tumor therapy and the regulation of immune homeostasis.21,37–39 Finally, DCs, as the most potent antigen-presenting cells, secrete EVs that express tumor antigens, major histocompatibility complex (MHC) molecules, and T-cell co-stimulatory molecules.22,40,41 Consequently, these DC-derived EVs can activate immune responses,38,39 stimulate the proliferation of CD4+ and CD8+ T cells as well as NK cells,22,42 and thereby facilitate the elimination of tumor cells, bacteria, or viruses. Collectively, EVs from these key immune effectors represent powerful tools for immune modulation and therapeutic intervention.
In conclusion, animal-derived EVs offer distinct advantages over conventional therapeutic agents. They exhibit superior biocompatibility43 and minimal intrinsic biotoxicity, undergo controlled degradation in vivo, and possess the inherent ability to efficiently cross biological barriers. Of particular importance for therapeutic applications, these EVs demonstrate extended circulation times within the bloodstream and exhibit enhanced tropism towards tumor tissues, leading to their preferential accumulation at pathological sites. This targeted biodistribution significantly increases the local concentration of therapeutic cargo (eg, intrinsic bioactive molecules or engineered payloads), thereby enhancing therapeutic efficacy while minimizing off-target effects. Furthermore, their natural anti-inflammatory properties contribute to tissue repair processes, such as wound healing. The unique properties of specific EV subtypes—such as tumor-targeting capability, immunomodulatory function, or efficient cargo delivery capacity—provide a flexible platform for developing novel individualized treatment strategies. Strategically combining EV-based therapies with established treatments, such as chemotherapy or surgery, holds considerable promise for achieving synergistic effects, improving treatment outcomes, and offering substantial potential for clinical translation. However, realizing this potential requires addressing challenges related to scalable production, standardized characterization, and comprehensive clinical validation.
EVs of Plant Origin
Plant-derived EVs have garnered significant attention as a promising frontier in cancer treatment and targeted drug delivery.26 They are broadly categorized by botanical origin into fruit-, herb- (medicinal plant-), and vegetable-derived subtypes, each of which exhibits distinct biological activities and therapeutic potential (Figure 3).
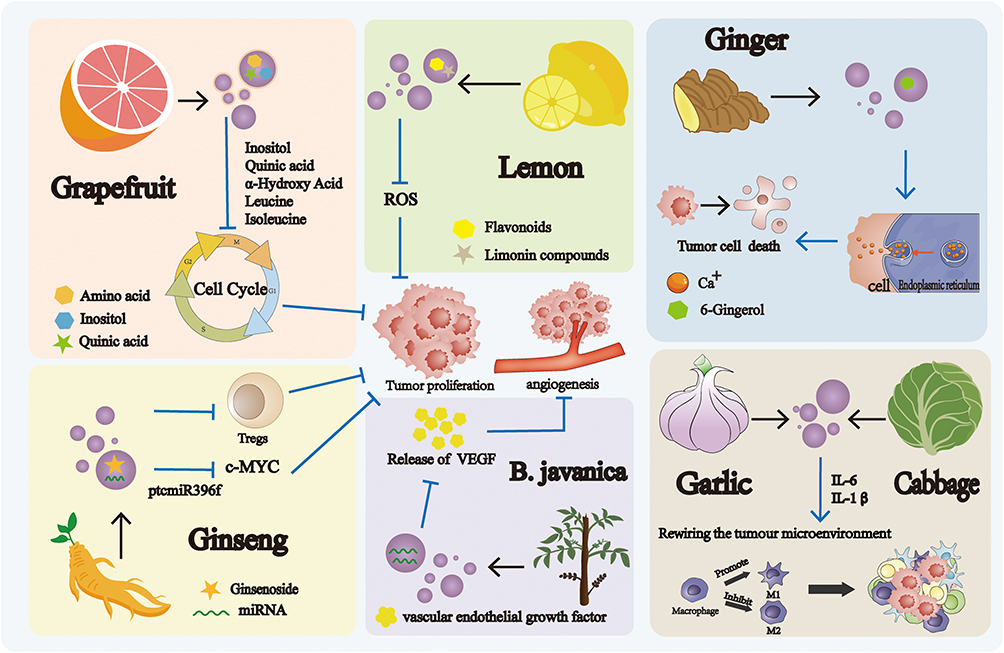 |
Figure 3 Mechanism of Action of plant-derived extracellular vesicles. EVs isolated from grapefruit, lemon, and ginseng exert anti-tumor effects through the modulation of tumor cell proliferation. EVs derived from B. javanica regulate tumors progression by influencing angiogenesis. EVs obtained from ginger induce tumor cell death, thereby contributing to tumor suppression. |
Fruit-derived EVs exert anti-tumor through diverse mechanisms. As a drug delivery platform, grapefruit-derived EVs enhance drug penetration and tissue retention, while reducing renal clearance and off-target drug accumulation in the liver and spleen.44 Beyond carrier function, they exert intrinsic anti-tumor activity by inducing G2/M phase arrest via downregulating cyclin B1/B2 and upregulating p21; inhibiting the PI3K/AKT and MAPK/ERK pathways; suppressing ICAM-1 and histone expression; and triggering tumor cell apoptosis via PARP-1 cleavage. Notably, grapefruit-derived EVs are rich in α-hydroxy acids, which selectively disrupt tumor cell energy metabolism with minimal toxicity to normal cells. Active components such as naringenin and flavonoids further synergize to exert anti-inflammatory and antioxidant effects, forming a multi-component, multi-target anti-tumor network with excellent biosafety. Despite these advantages, critical challenges remain for clinical translation. Specifically, the molecular basis for tumor selectivity and differential cellular responses remains incompletely defined. The long-term in vivo stability of bioactive components (eg, α-hydroxy acids and naringenin) and potential high-dose off-target effects require further investigation.44–46 The synergistic mechanisms between these active components, their optimal ratio for maximal efficacy, and the structure-activity relationships among lipid composition, drug loading capacity, and tumor selectivity also remain unclear. Inconsistent preparation and characterization methods further hinder inter-study comparability and clinical translation. Thus, future research integrating systematic omics analysis and rational engineering modifications will be crucial to fully exploit grapefruit-derived EVs as a next-generation anti-tumor nanoplatform.47,48 Lemon-derived EVs, containing flavonoids, citrullinated compounds, and lipids,26 inhibit gastric cancer by inducing S-phase arrest and apoptosis. This anticancer effect is mediated by reactive oxygen species (ROS) generation, GADD45A upregulation, and ROS-mediated MAPK/p53 activation, leading to caspase-3 cleavage. However, the molecular triggers of ROS production and the crosstalk between flavonoids, citrullinated compounds, and lipids in this pathway remain poorly defined, limiting mechanistic understanding and therapeutic optimization.44 For clinical translation, these EVs display favorable gastrointestinal stability, enabling effective oral delivery and avoiding the invasiveness of injectable formulations. Preclinical studies show significant in vivo tumor inhibition with low toxicity49–51 and anti-inflammatory effects26 that may attenuate pro-tumorigenic microenvironments. Nevertheless, the long-term gastrointestinal stability of bioactive components and potential dose-dependent off-target effects in normal gastrointestinal tissues require further evaluation. Additionally, lemon-derived EVs have been reported to mediate renal interspecies communication, a protective mechanism still under preliminary exploration.47,52,53 Watermelon-derived EVs act as natural miRNA carriers with inherent TME tropism and no known drug interactions. They also contain Vitamin C and lycopene, which alleviate tumor-associated oxidative stress and inflammation, potentially improving treatment tolerance.27 However, the molecular basis of TME tropism (specific mediating molecules/receptors), functional synergy between miRNAs and antioxidants, long-term in vivo stability of cargo/antioxidants, and dose-dependent off-target effects on adjacent normal tissues have not been rigorously characterized.27
Herb-derived EVs exhibit unique immunomodulatory properties. Ginseng-derived EVs, rich in ginsenosides, have prominent biological advantages: they exert anti-inflammatory effects by downregulating inflammatory pathways and scavenging ROS in immune and intestinal epithelial cells.54,55 Notably, their ability to cross the blood-brain barrier, promote angiogenesis, and target tumors28 broadens their application in central nervous system diseases and tumors. Mechanistically, they effectively ameliorate T cell exhaustion via the mTOR-T-bet axis, activate ERK and AKT/mTOR pathways to promote cell proliferation and angiogenesis, and display anti-aging potential by downregulating apoptosis and senescence-related genes.56 For tumor immunomodulation, they induce M2-to-M1 polarization of tumor-associated macrophages via the TLR4-MyD88, reprogram macrophages by stimulating CCL5/CXCL9 secretion to convert “cold” tumors to “hot” tumors, recruit CD8⁺ T cells, and synergize with PD-1 antibody to enhance efficacy without systemic toxicity.29 Despite these merits, critical gaps hinder clinical translation: the specific bioactive components (ginsenosides, miRNAs, lipids) mediating immunomodulation remain unclarified; dose-dependent immune balance and potential off-target activation require systematic evaluation; crosstalk between regulated signaling cascades (eg, ERK/AKT/mTOR) and their driving EV components is unclear; molecular determinants of tumor targeting and blood-brain barrier penetration are poorly defined, and and long-term immune safety of repeated administration needs rigorous validation, which is a key prerequisite for their clinical application. Similarly, Burcea javanica oil (BJO) transforms the immunosuppressive TME to activate anti-tumor immunity and inhibit tumor growth.57 Its derived EVs suppress vascular endothelial growth factor (VEGF) secretion and angiogenesis.30 Aloe-derived EVs, rich in antioxidants, promote tissue repair by activating Nrf2-mediated antioxidant response, reducing pro-inflammatory IL-6 and IL-1β in RAW264.7 macrophages, and promoting angiogenesis in human umbilical vein endothelial cells. They also serve as carriers for indocyanine green in phototherapy. However, significant variability in particle size and composition between EVs from different aloe parts (peel versus gel) poses challenges for standardized preparation.
Vegetable-derived EVs show multi-organ protective effects: Ginger-derived EVs, containing lipids, proteins, and 6-gingerol,58 induce G1/S arrest by modulating cyclin D1, cyclin E, phosphorylated Rb, and p21.59,60 They also facilitate hepatic interspecies communication, thereby modulating detoxification and antioxidant genes to confer hepatoprotection.61 However, the key bioactive cargo (specific lipids, RNAs, or 6-gingerol) mediating their cell cycle regulation and hepatoprotection remains unclear due to cargo complexity. In mitigating chemotherapy-induced toxicity, bitter melon-derived EVs alleviate doxorubicin (DOX)-induced cardiotoxicity by enhancing cardiomyocyte survival, reducing ROS production, and improving cardiac function/structure—without impairing DOX’s antitumor efficacy.62 Celery-derived EVs, enriched with diacylglycerol and phosphatidic acid, exhibit superior cellular uptake across cell types (eg, A549, HUVEC, C42B) compared to EV from lemon, ginger, or grape.63 In terms of anti-inflammatory and skin protection effects, cabbage-derived EVs protect keratinocytes by inhibiting lipopolysaccharide (LPS)-induced IL-6/IL-1β signaling, suggesting potential applications in colorectal cancer and inflammatory bowel disease.64 Garlic-derived EVs, rich in phosphatidic acid and functional miRNAs, have strong immunomodulatory and anti-tumor capacities: their miR-396e modulates PFKFB3 to regulate macrophage metabolic reprogramming and inhibit obesity-related tumor development; they also stimulate beneficial gut bacteria to release outer membrane vesicles, indirectly regulating systemic metabolism/immunity via gut-brain axis or gut-liver axis for novel tumor intervention. Corn-derived EVs have good modifiability: polyethylene glycol (PEG) modification prolongs their blood circulation half-life and enhances tumor tissues distribution. They also possess intrinsic immunostimulatory activity, activating macrophages to release inflammatory factors and indirectly inhibit colon cancer cell proliferation. However, unmodified corn-derived EVs are rapidly cleared in vivo. While PEG modification mitigates this, it increases preparation complexity and raises uncertainties: the potential impact of PEG modification on the intrinsic immunostimulatory activity remains unconfirmed; long-term biosafety (eg, PEG-induced immunogenicity) is unevaluated; and optimal PEG modification conditions (molecular weight, modification degree) for maximizing circulation half-life while maintaining bioactivity remain undefined.
Collectively, plant-derived EVs present significant clinical advantages over animal-derived counterparts: superior stability and prolonged circulation times enhance bioavailability by evading immune clearance;65 compatibility with oral or inhalation administration and absence of zoonotic pathogens improve safety;23 smaller size enables deeper tissue penetration and tumor targeting;66 and scalability enables cost-effective large-scale production.31,59 These intrinsic properties position them as highly viable candidates for commercialization.
However, critical challenges remain. First, there is significant heterogeneity in plant-derived EVs, as their biological characteristics are influenced by multiple factors including plant species, growth environment, harvesting stage, and extraction methods, leading to natural variations in composition and efficacy between batches that complicate standardized production. Second, their targeting capability is fundamentally limited; while they exhibit some inflammatory tropism, they lack specific recognition mechanisms, resulting in substantial accumulation in normal tissues and low targeting efficiency. Third, the drug loading system faces technical bottlenecks, manifesting as low loading efficiency, poor controllability, and potential interference from endogenous active components that may compromise the intended therapeutic effects of loaded drugs. Fourth, long-term safety concerns persist, as plant-derived EVs could potentially trigger immune responses, and the biocompatibility of cross-species applications requires systematic evaluation. These inherent limitations severely constrain their application in precision medicine, currently restricting their use to delivery scenarios with lower targeting requirements.
Engineering Modifications of EVs
Engineered modifications of EVs can be achieved through various approaches, primarily categorized into biological modification, immune modification, physical modification, and chemical modification.24,35,67–71 These methods can be employed either individually or in combination to achieve synergistic effects.3 Based on the site of modification, EV engineering strategies are classified into intracellular and extracellular modifications. Intracellular modifications typically focus on loading therapeutic substances to inhibit tumor initiation and metastasis, whereas extracellular modifications primarily aim to enhance the tumor-targeting capabilities of engineered EVs. (Table 2)
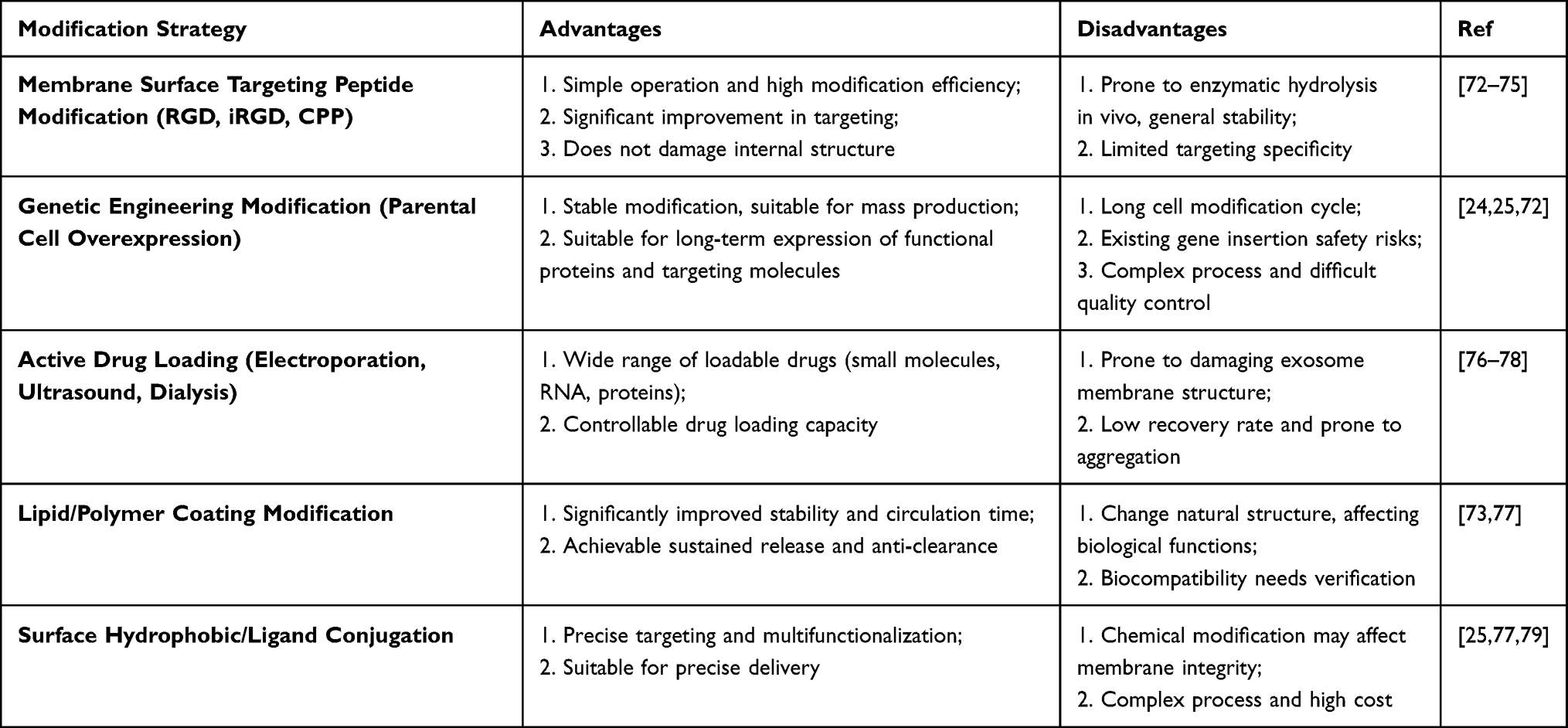 |
Table 2 Advantages and Disadvantages of Common Exosome Modification Strategies |
Intracellular Modifications
EVs comprise structural proteins, specific cargos (eg, proteins, miRNAs, mRNAs, and long non-coding RNAs),6 enabling them to transport a diverse range of therapeutic molecules.7,80,81 Intracellular modifications optimize the composition of EV cargo to ensure stability, protect against enzymatic degradation, and enhance anti-tumor efficacy.79 This is achieved either by modulating endogenous expression within donor cells or by directly introducing exogenous substances into isolated EVs. Endogenous modifications involve genetically engineering donor cells (eg, transfection with specific miRNAs or therapeutic protein genes) or modulating their microenvironment (eg, hypoxia, pharmacological stimulation), prompting the cells to naturally secrete EVs carrying the target molecules. This approach preserves the natural properties of EVs but may be constrained by cellular heterogeneity. Exogenous modifications, conversely, directly load active molecules into pre-isolated exogenous using techniques such as electroporation or chemical transfection. While straightforward, exogenous loading can potentially compromise EV membrane integrity.72 Both strategies offer distinct advantages: endogenous modification is often more suitable for basic research and large-scale production, whereas exogenous modification facilitates precise control over drug loading quantities.
Exogenous Loading
Exogenous loading actively introduces specific therapeutic molecules (eg, nucleic acids, proteins, or small-molecule drugs) into isolated and purified EVs using physical, chemical, or biological methods. Common EV isolation techniques include ultracentrifugation and size exclusion chromatography, while the choice of loading methods depends critically on the physicochemical properties of the target molecule.76,82 Co-incubation is a rapid and straightforward method, but its applicability is largely restricted to hydrophobic small molecules. Hydrophilic macromolecules (eg, siRNA), however, typically require methods like electroporation or sonication to enhance loading efficiency, though these carry a risk of damaging the EV membrane integrity. Additionally, electroporation can induce EV aggregation and fusion. Extrusion is predominantly employed for loading lipophilic compounds or creating hybrid vesicles but often results in heterogeneous vesicle population.77 Chemically-assisted methods (eg, saponin permeabilization) significantly improve the loading of hydrophilic drug but necessitate careful removal of potentially cytotoxic residues.77
Endogenous Loading
Endogenous loading integrates functional biomolecules (eg, viral antigens, mRNAs, or therapeutic proteins) into EVs by leveraging the donor cell’s own biosynthetic and secretory machinery. Target molecules are introduced into EV-secreting cells via techniques such as transfection, co-incubation, or electroporation. Following introduction, the cells are cultured, allowing the target molecules to be naturally incorporated into EVs during their biogenesis. Finally, the engineered EVs are isolated and purified using techniques like ultracentrifugation. Compared to exogenous loading, endogenous loading is generally simpler for biomolecules and better preserves EV integrity. However, its loading efficiency can be variable and is often influenced by cell type and culture conditions.24,25,72
Extracellular Modification
Extracelluar modification primarily targets the EV membrane surface to enhance tumor-targeting specificity, prolong systemic circulation time, and maximize therapeutic efficacy.77,79 Modifying agents encompass a wide array, including liposomes, aptamers, vesicular stomatitis virus G proteins, Nefmut proteins, stimulatory signals, targeting peptides, and chemicals compounds.72–74 These strategies can be broadly categorized into three mainstream approaches: genetic engineering, chemical modification, and physical modification. Each category possesses distinct advantages and disadvantages in key dimensions such as stability, homogeneity, operational complexity, and impact on the native membrane structure.
Genetic Engineering Modification
This approach involves genetic manipulation at the cellular level. By transfecting parent cells to express fusion proteins—such as targeting peptides fused with EV-enriched transmembrane proteins like LAMP2 or CD63—functional molecules are naturally displayed on the EV membrane during biogenesis. The fusion proteins are synthesized and precisely anchored by the cells themselves, ensuring uniform modification, strong binding, and high batch-to-batch consistency while avoiding the uncertainties associated with external manipulations. The entire process requires no in vitro chemical or physical treatment, thereby preserving the native lipid bilayer structure and functional proteins of EVs and offering excellent biocompatibility. Once a stable engineered cell line is established, consistently modified EVs can be continuously produced, laying a foundation for clinical translation and large-scale manufacturing. However, this method involves multiple complex steps, including gene cloning, cell transfection, and the screening of stable clones, making it time-consuming and costly. Additionally, excessively large or structurally complex fusion proteins may interfere with the proper folding and trafficking of transmembrane proteins, reducing EV yield. There is also the risk that foreign fusion proteins may alter the immunological properties of EVs, potentially triggering unforeseen immune responses.
Chemical Modification
This strategy is applied to isolated and purified EVs, where functional molecules are covalently linked to functional groups on the membrane surface via chemical reactions. Covalent bonding ensures that the functional molecules remain stably attached in complex in vivo environments, enabling effective delivery to target sites. A wide variety of molecules—including small-molecule drugs, peptides, antibodies, nucleic acids, and fluorescent probes—can be conjugated, offering great flexibility. Techniques such as click chemistry are particularly notable for their rapid reaction kinetics, high efficiency, specificity, and suitability for standardized and modular assembly. However, reaction conditions—such as the use of crosslinkers or metabolic labeling—may disrupt the native conformation of membrane lipids and proteins, compromising EV integrity and bioactivity. Furthermore, side reactions or residual unreacted groups may introduce cytotoxicity, posing safety concerns. Modification efficiency is also highly dependent on the availability of surface functional groups, which can lead to heterogeneous labeling and batch-to-batch variability.
Physical Modification
This method relies on non-covalent interactions—such as electrostatic or hydrophobic forces—or membrane fusion to adsorb or anchor functional molecules onto the EV surface. It typically involves simple procedures such as co-incubation or extrusion, requiring no complex chemical synthesis or genetic engineering, thus lowering the technical barrier. This approach avoids harsh chemical reactions, thereby preserving the native biological characteristics of EVs. Moreover, by designing different lipid molecules or polymers, multiple functions can be rapidly incorporated. However, the weak non-covalent interactions are prone to dissociation under physiological conditions—due to factors such as shear stress or ionic strength—leading to premature release of functional molecules and loss of targeting or therapeutic efficacy. The modification process is also difficult to control precisely, often resulting in highly heterogeneous EV populations with variable numbers and types of modifications, which affects experimental reproducibility and treatment predictability. Additionally, modifications may alter surface properties—for instance, cationic polymers can significantly change the zeta potential of EVs—potentially influencing their in vivo distribution, cellular uptake pathways, and biosafety profile.72–74,79
In summary, genetic engineering is best suited for basic research and long-term production where uniformity, stability, and membrane integrity are paramount. Chemical modification excels in scenarios requiring robust and versatile molecular conjugation, though careful evaluation of its impact on membrane structure is essential. Physical modification offers a convenient route for rapid preliminary functional validation. In practice, researchers often combine multiple strategies based on the characteristics of the target molecule, therapeutic requirements, and production feasibility to synergistically optimize the performance of engineered EVs.
Design Principles of Engineered EVs for Tumor-Specific Therapy
The design of engineered EVs must be precisely tailored to the core pathological characteristics of tumors to effectively address specific therapeutic challenges. This “tumor-specific” approach is pivotal for translating EV-based nanomedicines into clinical practice.
To achieve this goal, rational modification strategies are required according to the distinct immune and pathological features of different tumor types. For immunosuppressive tumors, such as triple-negative breast cancer, which are characterized by a profoundly suppressive immune microenvironment and insufficient immune cell infiltration, a triple-modification strategy integrating “targeted recognition – microenvironmental response – immune activation” is recommended. This involves displaying tumor-specific antigen antibodies (eg, anti-CD156) on the EV surface for precise targeting, incorporating pH-responsive peptides (eg, GALA) to mediate controlled cargo release in the acidic tumor niche, and co-loading pro-pyroptotic molecules (eg, GSDME-N mRNA) with immunosuppressive gene inhibitors (eg, DDR1 shRNA) to trigger immunogenic cell death and reverse immune suppression.83
For drug-resistant tumors such as non-small cell lung cancer, whose progression is dominated by chemotherapy/targeted therapy resistance and a pro-tumor microenvironment, a dual-path design centered on “tumor microenvironment (TME) reprogramming + drug-resistance mechanism reversal” is appropriate. EVs derived from M1-type macrophages can be employed to repolarize tumor-associated macrophages through their inherent anti-inflammatory properties. Alternatively, engineered EVs can be designed to reverse lncRNA Mir100hg-mediated H3K14 lactylation-driven resistance while co-delivering drug-resistance gene inhibitors (eg, miR-21 inhibitor) to resensitize tumors to treatment.84
For tumors with well-defined specific surface antigens, such as HER2-positive breast cancer and colorectal cancer, an “antigen targeting – cargo synergy” system should be constructed. Utilizing gene fusion technology to display antigen-specific binding moieties (eg, HER2 antibody fragments) on the vesicle surface and co-encapsulating chemotherapeutic agents (eg, 5-fluorouracil) with drug-resistance gene inhibitors can balance precise targeted killing with resistance reversal.83,85
For invasive tumors in special anatomical sites, such as glioblastoma, which are constrained by physiological barriers (eg, the blood-brain barrier) and driven by oncogenic proliferation, functions of “barrier penetration – gene editing” must be enhanced. This can be achieved by modifying EVs with targeting peptides like RVG29 to traverse biological barriers and loading tools such as CRISPR-Cas9 ribonucleoprotein via electroporation to specifically knockdown oncogenes like IDH1.86
Finally, for pan-cancers exhibiting strong target heterogeneity and lacking clear single targets, the core approach lies in decorating the surface with stimulus-responsive components (eg, pH-sensitive polymers, MMP-responsive peptides) to adapt to diverse tumor microenvironments, while carrying universal anti-tumor molecules (eg, tumor-suppressive miRNAs, metabolic disruptors) to achieve broad-spectrum efficacy through mechanisms such as modulating the TME and interfering with tumor metabolism.
Engineered EVs for Targeted Therapy in Tumors Across Different Systems
Engineered EVs, serving as a novel cell-free therapeutic modality, exhibit significant potential for tumors treatment across various systems, including the respiratory, digestive, nervous, and female reproductive systems (Table 3). The following sections will provide a detailed elaboration on their applications across various cancers—ranging from respiratory to reproductive system malignancies—through mechanisms including targeted drug delivery, nucleic acid delivery, immune activation and tumor microenvironment remodeling, as well as anti-angiogenesis therapy.
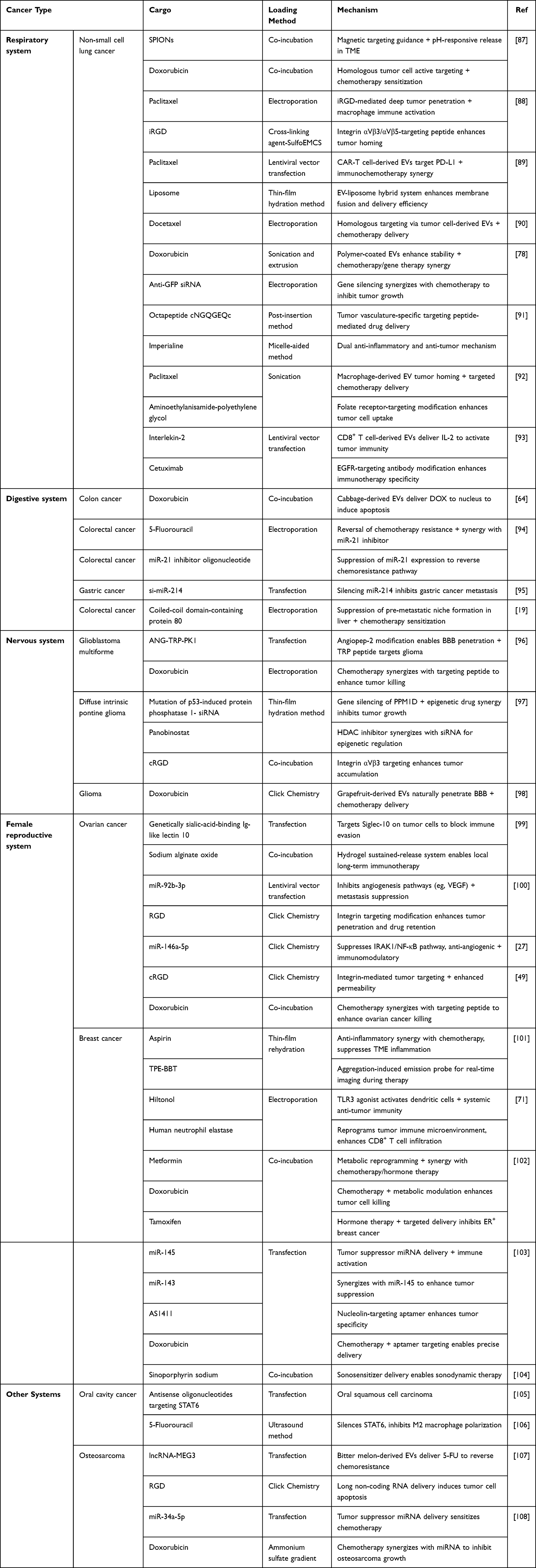 |
Table 3 Engineered EVs for Targeted Therapy in Tumors Across Different Systems |
Immune Activation and Tumor Microenvironment Remodeling
The occurrence and development of malignant tumors depend not only on the malignant proliferation of tumor cells themselves but are also closely related to the TME in which they reside. The TME is a complex ecosystem composed of tumor cells, immune cells, fibroblasts, vascular endothelial cells, as well as various cytokines and signaling molecules.109–112 During tumor evolution, the TME often exhibits highly immunosuppressive characteristics, manifested by exhausted effector T cell function, infiltration of immunosuppressive immune cells such as Tregs, and the abundance of various immune checkpoint molecules and inhibitory cytokines. This immunosuppressive microenvironment acts as a “protective umbrella” for tumor cells,113 enabling them to evade recognition and attack by the immune system, thereby leading to tumor immune escape, progression, and resistance to conventional therapies and immunotherapies. Consequently, effectively reversing the immunosuppressive state of the TME and reactivating and recruiting antitumor immune cells—transforming “immune-cold” tumors into “immune-hot” tumors—has become a core scientific issue and research frontier in the field of cancer immunotherapy.35 To achieve this goal, researchers are actively developing various strategies to synergistically accomplish immune activation and microenvironment remodeling. In recent years, with the rapid advancement of nanobiotechnology, novel delivery systems such as engineered EVs and synthetic nanoparticles have emerged as powerful technological platforms for precisely regulating the TME, owing to their excellent biocompatibility, targeting capabilities, and ease of functionalization. This section will focus on reviewing the latest progress and potential mechanisms in regulating the TME and activating antitumor immune responses through engineering strategies.
Numerous studies have highlighted the significant application potential of engineered EVs in modulating the tumor immune microenvironment. Huang et al encapsulated anti-PD-L1 single-chain variable fragments (scFv) into EVs derived from lung cancer cells.89 This strategy effectively prolonged the survival of mouse models with lung cancer by inhibiting tumor cell PD-L1 expression, preventing T cell exhaustion, and enhancing paclitaxel-induced immunogenic cell death. However, the homologous tumor cell-derived EVs used in this approach may inherently carry tumor-promoting substances, raising concerns about their long-term safety.89 In contrast, Wang et al employed ginseng-derived EVs for engineering, which may offer better biosafety. By fusing them with lung cancer cell membranes, the resulting formulation significantly enhanced dendritic cell (DC) phagocytosis of tumor antigens and promoted their maturation via the Toll-like receptor 4 (TLR4) signaling pathway, thereby activating tumor-specific cytotoxic T lymphocytes. This provides a new strategy for suppressing lung cancer recurrence and metastasis.114 Nevertheless, the interaction mechanisms between plant-derived EVs and the human immune system are not fully elucidated, and the risk of immunogenicity in cross-species applications cannot be overlooked. To further improve immune activation efficiency, Xu et al innovatively developed a hybrid nano-vaccine by fusing tumor-derived EVs with DC membrane vesicles and incorporating the TLR4 agonist MPLA. This engineering strategy endowed the vaccine with three key functions: CCR7-mediated active lymph node targeting, efficient antigen presentation, and potent immune activation.114 In a preclinical model of head and neck squamous cell carcinoma, the vaccine reduced tumor volume by up to 90%, accompanied by an increased proportion of effector T cells and a decrease in regulatory T cells. However, its complex preparation process and difficulties in component characterization pose significant challenges for large-scale production and quality control. Beyond the strategies mentioned above, studies have also shown that integrating human neutrophil elastase and a TLR3 agonist into α-lactalbumin-modified breast cancer cell-derived EVs can effectively activate DCs and CD8+ T cells, thereby enhancing systemic anti-tumor immunity.71 Although this approach robustly activates innate immunity, the in vivo pharmacokinetics of its multiple active components and their potential synergistic toxicity still require systematic evaluation. In an oral squamous cell carcinoma (OSCC) model, EVs encapsulating STAT6-targeting antisense oligonucleotides effectively suppressed STAT6 expression, thereby inhibiting macrophage polarization to the M2 phenotype and curbing tumor progression.105 However, most of these strategies rely on tumor cell-specific targeted delivery, whose efficiency may be significantly compromised in highly heterogeneous solid tumors, and their off-target effects on other immune cell subsets remain unknown. For malignant melanoma, Wang et al developed hybrid EVs by fusing melanoma cells with macrophages, achieving dual precise targeting of both tumor foci and lymph nodes.115 Through efficient tumor antigen delivery and activation of co-stimulatory signals, this approach significantly enhanced T cell activation and remodeled the immunosuppressive tumor microenvironment (TME).115 Additionally, in reversing chemotherapy resistance, Meng et al loaded 5-fluorouracil into bitter melon-derived EVs, which significantly downregulate the expression of the NOD-like receptor thermal protein domain-associated protein 3 (NLRP3). This strategy not only enhanced OSCC sensitivity to 5-fluorouracil by suppressing NLRP3 but also prolonged drug retention and duration of action at the lesion site, demonstrating potent anti-tumor effects.106 It is important to note that natural EVs exhibit substantial batch-to-batch variability, and controlling drug loading efficiency and release kinetics with precision remains challenging, which undermines the reliability and consistency of their clinical application. In summary, engineered EVs represent a versatile platform for reprogramming the immunosuppressive TME and activating systemic anti-tumor immunity. While promising, their clinical translation requires standardized production, improved targeting precision, and thorough evaluation of long-term safety.
Targeted Delivery of Drugs
EVs have emerged as a highly promising platform for the targeted delivery of therapeutic agents, leveraging their innate biocompatibility, low immunogenicity, and natural tropism to specific tissues. In recent years, engineered EVs have been extensively explored to improve the precision and efficacy of cancer treatments, particularly in the delivery of chemotherapeutic drugs and nucleic acid-based therapeutics. These advanced systems not only enhance drug stability and prolong systemic circulation but also facilitate tumor-specific accumulation and penetration, thereby maximizing antitumor effects while minimizing off-target toxicity.116–119 The following sections summarize key advances in the use of EVs for the targeted delivery of chemotherapeutic agents and nucleic acids, highlighting innovative engineering strategies, representative applications, and remaining challenges in translational development.
Targeted Delivery of Chemotherapeutic Drugs
Significant breakthroughs have been achieved in the field of chemotherapeutic drug delivery using EV-based systems. By engineering EVs derived from various sources, including lung cancer cells, celery, and milk, researchers have successfully achieved efficient loading of multiple chemotherapeutic drugs such as doxorubicin and paclitaxel. Notably, studies by demonstrated that celery-derived EVs loaded with DOX not only exhibited excellent biocompatibility but also achieved 3–5 times higher uptake rates in lung cancer cells compared to free DOX. This strategy effectively prolonged the plasma half-life of DOX, increased its concentration in tumor tissues, and significantly promoted cancer cell apoptosis.21 However, the natural tropism of EVs imposes limitations on targeting specificity. To address this challenge, various surface modification strategies have been developed, including the conjugation of targeting ligands such as superparamagnetic iron oxide nanoparticles and interleukin-2. These innovations not only effectively mitigate chemotherapy-associated toxicities but also significantly enhance targeted delivery efficiency. For instance, Wang’s team developed a dual-targeting system composed of NSCLC cell-derived EVs and transferrin-modified SPIONs.87,93 This system combines the natural tumor-homing capability of EVs with magnetic guidance, enabling precise drug release within the acidic tumor microenvironment. Similarly, Chen’s team conjugated the integrin αVβ3/αVβ5-binding peptide iRGD to milk-derived EVs. Loading these modified EVs with paclitaxel achieved deep tumor penetration unattainable with conventional chemotherapy, effectively inhibiting lung cancer cell proliferation and activating macrophage-mediated tumor cell killing.88 While these innovations significantly enhance targeting efficiency, the complexity of their preparation processes and the feasibility of large-scale production require further validation. Of particular concern is that such engineering modifications may alter the innate properties of EVs, affecting their biodistribution and safety profiles, and currently, there is a lack of systematic toxicological assessment data.
In the treatment of digestive system tumors, plant-derived EVs demonstrate unique advantages. You et al confirmed that cabbage-derived EVs could effectively deliver DOX to the nucleus of colon cancer cells to exert cytotoxic effects.64 Zhang’s team developed ginger-derived EVs loaded with folic acid and DOX, enabling active and specific targeting of colon cancer cells.66 Compared to free DOX, these Plant-derived extracellular vesicle delivery systems offer multiple advantages: they significantly improve the stability of chemotherapeutic agents, prolong circulation time, increase intratumoral drug concentration, and reduce systemic toxicity through enhanced targeting, thereby improving treatment safety while maintaining efficacy.66 These groundbreaking advances provide novel strategies for the reformulation of chemotherapeutic drugs and open new avenues for precision cancer therapy. Collectively, these studies underscore the potential of EV-based systems to enhance the precision and safety of chemotherapeutic drug delivery. Future efforts should focus on optimizing targeting strategies, scaling up production, and conducting systematic toxicological assessments to facilitate clinical adoption.
Delivery of Nucleic Acid Drugs
In the field of nucleic acid drug delivery, EVs serve as natural carriers with unique advantages. Lung cancer cell-derived EVs possess inherent homologous targeting ability. Loaded with microRNAs such as miR-30c, they can synergistically inhibit lung cancer progression through multiple mechanisms including suppressing proliferation and metastasis, and modulating the tumor microenvironment. To further enhance targeting, Yang’s team utilized the LXY30 peptide, which targets α3β1 integrins, to modify bone marrow MSC-derived EVs, successfully delivering molecules like miR-30c, effectively inducing apoptosis in NSCLC cells and enhancing tumor-targeting capability.120 In treating diffuse intrinsic pontine glioma, Shan’s team developed cRGD-modified macrophage-derived EVs co-loaded with panobinostat and PPM1D siRNA. This strategy ingeniously combines the precise targeting of cRGD, the blood-brain barrier penetration inherent to macrophage-derived EVs, and the synergistic effects of the therapeutic agents, significantly improving survival rates in diffuse intrinsic pontine glioma (DIPG) models.97 Furthermore, EV platforms show unique value in overcoming chemoresistance. Liang’s team co-loaded 5-fluorouracil and a miR-21 inhibitor into milk-derived EVs, successfully reversing 5-fluorouracil resistance in colorectal cancer and increasing drug efficacy by 2–3 times.98 Similarly, grape-derived EVs loaded with metformin, DOX, and tamoxifen effectively suppressed breast cancer progression by inducing apoptosis and necrosis. However, these innovative strategies still face numerous challenges: EV drug-loading efficiency varies considerably, targeting strategies require further optimization, and the synergistic mechanisms with existing chemotherapy regimens are not fully elucidated. Particularly in scale-up production and quality control, the consistency, stability, and in vivo pharmacokinetics of EV-based drugs remain insufficiently studied.
Engineered EVs can enhance intratumoral drug concentration, effectively mitigate systemic toxicity, reverse drug resistance, improve chemotherapeutic sensitivity, and inhibit tumor growth and metastasis by facilitating the targeted delivery of chemotherapeutic drugs, RNA, and other therapeutic molecules. These properties position engineered EVs as a promising novel strategy and conceptual framework for tumor treatment. In conclusion, EV-mediated nucleic acid delivery offers a promising approach to overcome chemoresistance and enhance targeted therapy. Addressing challenges in loading efficiency, targeting optimization, and scalable manufacturing will be crucial for advancing these strategies toward clinical use.
Anti-Angiogenesis
In ovarian cancer research, Wang’s team developed RGD peptide-modified EVs derived from ovarian cancer cells that overexpress miR-92b-3p, demonstrating a dual anti-tumor mechanism: RGD functionalization confers tumor-specific targeting capability, while overexpressed miR-92b-3p significantly inhibits tumor metastasis by suppressing key angiogenic pathways.100 Similarly, Corvigno’s team utilized watermelon-derived EVs to deliver miR-146a-5p. This strategy not only directly inhibited endothelial cell activity but also indirectly downregulated the expression of pro-angiogenic factors by modulating the IRAK1/NF-κB/SERPINE1 signaling axis, exhibiting a synergistic, multi-targeted anti-angiogenic effect.27 These studies collectively demonstrate that engineered EVs can achieve multi-pathway synergistic therapy through precise molecular regulation, providing new ideas for cancer treatment.
Collectively, these studies highlight the unique value of EVs as natural nanocarriers in anti-angiogenic therapy: their inherent biocompatibility, adaptability to surface modifications, and capacity for multi-mechanistic synergy offer promising directions for the development of next-generation anti-tumor angiogenic therapies. Notably, EVs derived from different plant sources exhibit distinctive functional advantages, providing a diverse array of carrier options for future personalized anti-angiogenic treatment. Taken together, engineered EVs exhibit significant potential in anti-angiogenic therapy by enabling multi-targeted regulation and synergistic effects. Their natural biocompatibility and modifiability make them attractive candidates for developing next-generation anti-angiogenic formulations.
Current Challenges and Future Prospects
EVs are natural intercellular messengers that transfer genetic materials and regulate recipient cell functions. Featuring low toxicity, minimal immunogenicity, biodegradability, and high loading capacity, engineered EVs can be functionalized with therapeutic cargos (RNA, chemotherapeutics, proteins) and targeting moieties to specifically intervene in tumorigenesis, progression, and metastasis. Preclinical evidence has validated their antitumor efficacy in multiple cancer types, underscoring their considerable translational potential in cancer therapy.121–123
Consistent with this translational potential, several clinical trials support the translational potential of engineered EVs, such as plant-derived EVs (NCT01294072), mesenchymal stromal cell EVs delivering KrasG12D siRNA for pancreatic cancer (NCT03608631), and dendritic cell EVs vaccines for non‑small cell lung cancer (NCT01159288). Future trials should focus on unmet clinical needs, local administration to bypass protein corona interference, and rigorous safety evaluation.
To further advance this translation and address existing challenges, the application of artificial intelligence (AI) in the design and clinical application of engineered EVs has become a frontier in biomedicine, offering innovative solutions to overcome existing bottlenecks. AI technology enables efficient processing of EVs spectral and imaging data: for instance, the Chat EV system utilizes deep learning to analyze Raman spectra of hepatocellular carcinoma (HCC)-derived EVs, significantly improving diagnostic accuracy. Additionally, AI algorithms applied in EVs proton detection further enhance the transparency and reliability of EVs-based diagnosis.16 In engineered EVs design, AI not only boosts design efficiency but also optimizes their functional properties—building on its well-established potential in tissue engineering, where it facilitates the design of complex three-dimensional structures and personalized medicine.75,78 By integrating AI with tissue engineering, researchers can precisely design and fabricate biomaterials, accelerating the clinical translation of regenerative medicine products.90 Furthermore, AI’s application provides a novel perspective for cancer management: it optimizes the nanomedicine design process by analyzing large-scale datasets, improving molecular analysis accuracy and early diagnostic capabilities.91 This advancement enhances drug targeting and personalized dosing, while reducing nanotoxicity to improve overall therapeutic efficacy—directly addressing key challenges in engineered EVs translation.
Despite these promising advances, clinical translation is hampered by four critical bottlenecks. First, pronounced heterogeneity and insufficient standardization, leading to poor reproducibility. Second, impaired targeting due to protein corona formation. Third, safety concerns including membrane damage, unpredictable immunogenicity, and potential pro‑tumor risks. Fourth, poor stability and storage stability. To address these limitations, strategies including advanced purification, optimized loading, cryopreservation, lyophilization, EV‑liposome hybrid, and protein corona characterization have been developed.77,124,125
To further advance clinical translation and industrial application of engineered EVs for cancer therapy, the following prioritized key next steps are imperative: First, establish standardized isolation, characterization, and quality control to resolve heterogeneity, enhance reproducibility, and underpin clinical translation. Second, develop high‑efficiency, tumor‑specific targeting strategies via rational engineering, including optimized surface modification and EVs‑liposome hybrid systems to overcome protein corona interference and enhance targeting specificity. Third, realize scalable, controllable, and reproducible production of clinical‑grade EVs. Forth, conduct systematic safety, pharmacokinetic, and pharmacodynamic studies in large animals. Fifth, accelerate clinical trials with optimized design for robust proof-of-concept.
Collectively, engineered EVs research is transitioning from local optimization to systemic reconstruction. Future advances will depend on standardized analytical pipelines, intelligent biomimetic engineering, stimuli‑responsive design, and interdisciplinary integration of AI, single‑cell omics, and in vivo imaging.
Conclusion
EVs are promising biocompatible nanocarriers for precision tumor therapy. This review covers EVs source optimization, engineering modifications, and tumor-specific design, highlighting their roles in targeted delivery, immune activation, tumor microenvironment remodeling, and anti-angiogenesis. Despite challenges in heterogeneity, targeting, safety, and scalable production, advanced strategies are driving translation. Technological advances will enable EVs to become a transformative therapeutic modality for refractory tumors and boost precision oncology.
Abbreviations
TME, tumor microenvironment; MSC, Mesenchymal Stem Cells; NK, Natural Killer; DC, Dendritic Cells; BM-MSCs, bone marrow MSC; hUC-MSCs, human Umbilical Cord Mesenchymal Stem Cells; EMT, Epithelial-Mesenchymal Transition; ADSCs, Adipose-derived Stem Cells; HNSCC, Head and Neck Squamous Carcinoma Cells; OSCC, oral squamous cell carcinoma; OS, Osteosarcoma; LYM, lymphoma; anti-PD-L1, anti-programmed cell death receptor ligand 1; M1, Classically Activated Macrophages; MHC, Major Histocompatibility Complex; LPS, lipopolysaccharide; ROS, Reactive Oxygen Species; BJO, B. javanica oil; VEGF, Vascular Endothelial Growth Factor; DOX, Doxorubicin; NSCLC, Non-Small Cell Lung Cancer; SPIONs, superparamagnetic iron oxide nanoparticles; scFv, Single-chain Variable Fragment; TLR4, Toll-like receptors 4; CCDC80, Coiled-coil Domain Containing 80; GBM, Glioblastomas; BBB, Blood-Brain Barrier; LRP1, Low-density Lipoprotein Receptor-related Protein 1; ANG, Angiopep-2; DIPG, Diffuse Intrinsic Pontine Glioma; PPM1D, P53-induced Protein Phosphatase 1; CNS, Central Nervous System; CAV-1, Caveolin-1; DAMPs, Damage-associated Molecular Patterns; TLR3, Toll-like Receptor 3; NLRP3, NOD-like receptor thermal protein domain associated protein 3; PVP40, Polyvinylpyrrolidone 40; EVs, Extracellular vesicles.
Acknowledgments
The authors would like to express their deepest gratitude to all those who contributed to the completion of this work. We are profoundly grateful to our supervisor and colleagues, for their invaluable guidance, insightful feedback, and unwavering support throughout this review. Their expertise and encouragement were instrumental in shaping this review.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Shandong Provincial Natural Science Foundation (ZR2025MS1322). Shandong Provincial Natural Science Foundation (ZR2024LSW017). Shandong Provincial Natural Science Foundation (ZR2023QH460).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–23. doi:10.3322/caac.21660
2. Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12–49. doi:10.3322/caac.21820
3. Zhang M, Hu S, Liu L, et al. Engineered exosomes from different sources for cancer-targeted therapy. Signal Transduct Target Ther. 2023;8(1):124. doi:10.1038/s41392-023-01382-y
4. Chen H. Advances in macrophage-derived exosomes as immunomodulators in disease progression and therapy. Int Immunopharmacol. 2024;142(Pt B):113248. doi:10.1016/j.intimp.2024.113248
5. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
6. Wang L. Exosomes containing miRNAs targeting HER2 synthesis and engineered to adhere to HER2 on tumor cells surface exhibit enhanced antitumor activity. J Nanobiotechnol. 2020;18(1):153. doi:10.1186/s12951-020-00711-5
7. El Andaloussi S, Mäger I, Breakefield XO, Wood MJA. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12(5):347–357. doi:10.1038/nrd3978
8. Zhang Q, Yu M, Liu L, Tian W. Recent advances of engineered exosomes and challenges on clinical translational research. Chin J Tissue Eng Res. 2023;27(19):3052–3060. doi:10.12307/2023.636
9. Ma C. Application of engineered exosomes in tumor therapy. Am J Transl Res. 2025;17(2):736–747. doi:10.62347/KIXF4662
10. Patil J, Singh A, Bhalerao S, et al. Engineered exosomes as programmable nanocarriers for targeted theranostic and translational applications. ACS Appl Bio Mater. 2025;8(12):10506–10529. doi:10.1021/acsabm.5c01251
11. Iqbal Z, Rehman K, Mahmood A, et al. Exosome for mRNA delivery: strategies and therapeutic applications. J Nanobiotechnol. 2024;22(1):395. doi:10.1186/s12951-024-02634-x
12. Liu JJJ, Liu D, Sky T, Wong AST. Exosomes in cancer nanomedicine: biotechnological advancements and innovations. Mol Cancer. 2025;24(1):166. doi:10.1186/s12943-025-02372-0
13. Hassanzadeh A, Rahman HS, Markov A, et al. Mesenchymal stem/stromal cell-derived exosomes in regenerative medicine and cancer; overview of development, challenges, and opportunities. Stem Cell Res Ther. 2021;12(1):297. doi:10.1186/s13287-021-02378-7
14. Xu Y, Lai Y, Cao L, et al. Human umbilical cord mesenchymal stem cells-derived exosomal microRNA-451a represses epithelial–mesenchymal transition of hepatocellular carcinoma cells by inhibiting ADAM10. RNA Biol. 2021;18(10):1408–1423. doi:10.1080/15476286.2020.1851540
15. Lin R, Wang S, Zhao RC. Exosomes from human adipose-derived mesenchymal stem cells promote migration through Wnt signaling pathway in a breast cancer cell model. Mol Cell Biochem. 2013;383(1–2):13–20. doi:10.1007/s11010-013-1746-z
16. Zeng H, Wang Q, Wang F. Extracellular vesicles derived from citrus plants: innovative applications in therapeutics and drug delivery. J Nanobiotechnol. 2026. doi:10.1186/s12951-026-04210-x
17. Hao Y, Chen P, Guo S, et al. Tumor-derived exosomes induce initial activation by exosomal CD19 antigen but impair the function of CD19-specific CAR T-cells via TGF-β signaling. Front Med. 2024;18(1):128–146. doi:10.1007/s11684-023-1010-1
18. Li K, Xue W, Lu Z, et al. Tumor-derived exosomal ADAM17 promotes pre-metastatic niche formation by enhancing vascular permeability in colorectal cancer. J Exp Clin Cancer Res. 2024;43(1). doi:10.1186/s13046-024-02991-3
19. Huang C, Zhou Y, Feng X, Wang J, Li Y, Yao X. Delivery of engineered primary tumor-derived exosomes effectively suppressed the colorectal cancer chemoresistance and liver metastasis. ACS Nano. 2023;17(11):10313–10326. doi:10.1021/acsnano.3c00668
20. Choo YW, Kang M, Kim HY, et al. M1 macrophage-derived nanovesicles potentiate the anticancer efficacy of immune checkpoint inhibitors. ACS Nano. 2018;12(9):8977–8993. doi:10.1021/acsnano.8b02446
21. Lugini L, Cecchetti S, Huber V, et al. Immune surveillance properties of human NK cell-derived exosomes. J Immunol. 2012;189(6):2833–2842. doi:10.4049/jimmunol.1101988
22. Zitvogel L, Regnault A, Lozier A, et al. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med. 1998;4(5):594–600. doi:10.1038/nm0598-594
23. Zhao B, Lin H, Jiang X, et al. Exosome-like nanoparticles derived from fruits, vegetables, and herbs: innovative strategies of therapeutic and drug delivery. Theranostics. 2024;14(12):4598–4621. doi:10.7150/thno.97096
24. Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. IJN. 2020;15:6917–6934. doi:10.2147/IJN.S264498
25. Lai RC, Yeo RWY, Tan KH, Lim SK. Exosomes for drug delivery — a novel application for the mesenchymal stem cell. Biotechnol Adv. 2013;31(5):543–551. doi:10.1016/j.biotechadv.2012.08.008
26. Yang M, Liu X, Luo Q, Xu L, Chen F. An efficient method to isolate lemon derived extracellular vesicles for gastric cancer therapy. J Nanobiotechnol. 2020;18(1):100. doi:10.1186/s12951-020-00656-9
27. Corvigno S, Liu Y, Bayraktar E, et al. Enhanced plant-derived vesicles for nucleotide delivery for cancer therapy. Npj Precis Onc. 2024;8(1):86. doi:10.1038/s41698-024-00556-3
28. Kim J, Zhu Y, Chen S, et al. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. J Nanobiotechnol. 2023;21(1):253. doi:10.1186/s12951-023-02006-x
29. Han X, Wei Q, Lv Y, et al. Ginseng-derived nanoparticles potentiate immune checkpoint antibody efficacy by reprogramming the cold tumor microenvironment. Mol Ther. 2022;30(1):327–340. doi:10.1016/j.ymthe.2021.08.028
30. Yan G, Xiao Q, Zhao J, et al. Brucea javanica derived exosome-like nanovesicles deliver miRNAs for cancer therapy. J Control Release. 2024;367:425–440. doi:10.1016/j.jconrel.2024.01.060
31. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4:1867. doi:10.1038/ncomms2886
32. Yao X, Mao Y, Wu D, et al. Exosomal circ_0030167 derived from BM-MSCs inhibits the invasion, migration, proliferation and stemness of pancreatic cancer cells by sponging miR-338-5p and targeting the Wif1/Wnt8/β-catenin axis. Cancer Lett. 2021;512:38–50. doi:10.1016/j.canlet.2021.04.030
33. Du L, Tao X, Shen X. Human umbilical cord mesenchymal stem cell-derived exosomes inhibit migration and invasion of breast cancer cells via miR-21-5p/ZNF367 pathway. Breast Cancer. 2021;28(4):829–837. doi:10.1007/s12282-021-01218-z
34. Xie X, Ji J, Chen X, et al. Human umbilical cord mesenchymal stem cell-derived exosomes carrying hsa-miRNA-128-3p suppress pancreatic ductal cell carcinoma by inhibiting Galectin-3. Clin Transl Oncol. 2022;24(3):517–531. doi:10.1007/s12094-021-02705-7
35. Lu Y, Zheng J, Lin P, et al. Tumor microenvironment-derived exosomes: a double-edged sword for advanced T cell-based immunotherapy. ACS Nano. 2024;18(40):27230–27260. doi:10.1021/acsnano.4c09190
36. Tang Q, Yang S, He G, et al. Tumor-derived exosomes in the cancer immune microenvironment and cancer immunotherapy. Cancer Lett. 2022;548:215823. doi:10.1016/j.canlet.2022.215823
37. He S, Su L, Hu H, et al. Immunoregulatory functions and therapeutic potential of natural killer cell-derived extracellular vesicles in chronic diseases. Front Immunol. 2023;14:1328094. doi:10.3389/fimmu.2023.1328094
38. Yin W, Ouyang S, Li Y, Xiao B, Yang H. Immature dendritic cell-derived exosomes: a promise subcellular vaccine for autoimmunity. Inflammation. 2013;36(1):232–240. doi:10.1007/s10753-012-9539-1
39. Zang J, Mei Y, Zhu S, et al. Natural killer-based therapy: a prospective thought for cancer treatment related to diversified drug delivery pathways. Pharmaceutics. 2024;16(7):939. doi:10.3390/pharmaceutics16070939
40. Xia J, Miao Y, Wang X, Huang X, Dai J. Recent progress of dendritic cell-derived exosomes (Dex) as an anti-cancer nanovaccine. Biomed Pharmacother. 2022;152:113250. doi:10.1016/j.biopha.2022.113250
41. Théry C, Duban L, Segura E, Véron P, Lantz O, Amigorena S. Indirect activation of naïve CD4+ T cells by dendritic cell-derived exosomes. Nat Immunol. 2002;3(12):1156–1162. doi:10.1038/ni854
42. Du Z, Huang Z, Chen X, et al. Modified dendritic cell-derived exosomes activate both NK cells and T cells through the NKG2D/NKG2D-L pathway to kill CML cells with or without T315I mutation. Exp Hematol Oncol. 2022;11(1):36. doi:10.1186/s40164-022-00289-8
43. Zhang X, Zhang H, Gu J, et al. Engineered extracellular vesicles for cancer therapy. Adv Mater. 2021;33(14):2005709. doi:10.1002/adma.202005709
44. Zhang Y, Zhang R, Zhang T, et al. Restoration of tendon repair microenvironment by grapefruit exosome-loaded microneedle system for tendinopathy therapy. Front Bioeng Biotechnol. 2025;13:1615650. doi:10.3389/fbioe.2025.1615650
45. Huang R, Jia B, Su D, et al. Plant exosomes fused with engineered mesenchymal stem cell-derived nanovesicles for synergistic therapy of autoimmune skin disorders. J Extracell Vesicles. 2023;12(10):e12361. doi:10.1002/jev2.12361
46. Castelli G, Logozzi M, Mizzoni D, et al. Ex vivo anti-leukemic effect of exosome-like grapefruit-derived nanovesicles from organic farming—the potential role of ascorbic acid. Int J Mol Sci. 2023;24(21):15663. doi:10.3390/ijms242115663
47. Tembo KM, Wang X, Bolideei M, et al. Exploring the bioactivity of MicroRNAs originated from plant-derived exosome-like nanoparticles (PELNs): current perspectives. J Nanobiotechnol. 2025;23(1):563. doi:10.1186/s12951-025-03602-9
48. Feng J, Xiu Q, Huang Y, Troyer Z, Li B, Zheng L. Plant-derived vesicle-like nanoparticles as promising biotherapeutic tools: present and future. Adv Mater. 2023;35(24):2207826. doi:10.1002/adma.202207826
49. Xiao Q, Zhao W, Wu C, et al. Lemon-derived extracellular vesicles nanodrugs enable to efficiently overcome cancer multidrug resistance by endocytosis-triggered energy dissipation and energy production reduction. Adv Sci. 2022;9(20):2105274. doi:10.1002/advs.202105274
50. Urzì O, Cafora M, Ganji NR, et al. Lemon-derived nanovesicles achieve antioxidant and anti-inflammatory effects activating the AhR/Nrf2 signaling pathway. iScience. 2023;26(7):107041. doi:10.1016/j.isci.2023.107041
51. Gasparro R, Gambino G, Duca G, et al. Protective effects of lemon nanovesicles: evidence of the Nrf2/HO-1 pathway contribution from in vitro hepatocytes and in vivo high-fat diet-fed rats. Biomed Pharmacother. 2024;180:117532. doi:10.1016/j.biopha.2024.117532
52. Zhang L, Li S, Cong M, et al. Lemon-derived extracellular vesicle-like nanoparticles block the progression of kidney stones by antagonizing endoplasmic reticulum stress in renal tubular cells. Nano Lett. 2023;23(4):1555–1563. doi:10.1021/acs.nanolett.2c05099
53. Liu D, Gao J, Wu X, Han L. Plant-derived exosome-like nanoparticles as promising biotherapeutic tools: recent advances and challenges. Smart Mater Med. 2025;6(2):285–304. doi:10.1016/j.smaim.2025.07.003
54. Kim J, Zhang S, Zhu Y, Wang R, Wang J. Amelioration of colitis progression by ginseng-derived exosome-like nanoparticles through suppression of inflammatory cytokines. J Ginseng Res. 2023;47(5):627–637. doi:10.1016/j.jgr.2023.01.004
55. Yang S, Li W, Bai X, et al. Ginseng-derived nanoparticles alleviate inflammatory bowel disease via the TLR4/MAPK and p62/Nrf2/Keap1 pathways. J Nanobiotechnol. 2024;22(1):48. doi:10.1186/s12951-024-02313-x
56. Choi W, Cho JH, Park SH, et al. Ginseng root-derived exosome-like nanoparticles protect skin from UV irradiation and oxidative stress by suppressing activator protein-1 signaling and limiting the generation of reactive oxygen species. J Ginseng Res. 2024;48(2):211–219. doi:10.1016/j.jgr.2024.01.001
57. Meng J, Yu Z, Chen H, et al. Brucea javanica oil emulsion significantly improved the effect of anti–programmed cell death protein-1 immunotherapy. Phytomedicine. 2022;107:154446. doi:10.1016/j.phymed.2022.154446
58. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
59. Karna P, Chagani S, Gundala SR, et al. Benefits of whole ginger extract in prostate cancer. Br J Nutr. 2012;107(4):473–484. doi:10.1017/S0007114511003308
60. Chen CY, Chen YN, Shiau JP, Tang JY, Hou MF, Chang HW. Ginger-derived 3HDT exerts antiproliferative effects on breast cancer cells by apoptosis and DNA damage. Int J Mol Sci. 2023;24(6):5741. doi:10.3390/ijms24065741
61. Zhuang X, Deng ZB, Mu J, et al. Ginger-derived nanoparticles protect against alcohol-induced liver damage. J Extracell Vesicles. 2015;4:28713. doi:10.3402/jev.v4.28713
62. Ye C, Yan C, Bian SJ, et al. Momordica charantia L.-derived exosome-like nanovesicles stabilize p62 expression to ameliorate doxorubicin cardiotoxicity. J Nanobiotechnol. 2024;22(1):464. doi:10.1186/s12951-024-02705-z
63. Lu X, Han Q, Chen J, et al. Celery (Apium graveolens L.) exosome-like nanovesicles as a new-generation chemotherapy drug delivery platform against tumor proliferation. J Agric Food Chem. 2023;71(22):8413–8424. doi:10.1021/acs.jafc.2c07760
64. You JY. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact Mater. 2021;6(12):4321–4332. doi:10.1016/j.bioactmat.2021.04.023
65. C M, D N, C X, et al. Plant exosome nanovesicles (PENs): green delivery platforms. Mater Horiz. 2023;10(10):3879–3894. doi:10.1039/d3mh01030a
66. Zhang M, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol Ther. 2016;24(10):1783–1796. doi:10.1038/mt.2016.159
67. Ma X, Yao M, Gao Y, et al. Functional immune cell-derived exosomes engineered for the trilogy of radiotherapy sensitization. Adv Sci. 2022;9(23):2106031. doi:10.1002/advs.202106031
68. Jiang J, Lu Y, Chu J, et al. Anti-EGFR ScFv functionalized exosomes delivering LPCAT1 specific siRNAs for inhibition of lung cancer brain metastases. J Nanobiotechnol. 2024;22(1):159. doi:10.1186/s12951-024-02414-7
69. Yu X, Yang X, Horte S, Kizhakkedathu JN, Brooks DE. A pH and thermosensitive choline phosphate-based delivery platform targeted to the acidic tumor microenvironment. Biomaterials. 2014;35(1):278–286. doi:10.1016/j.biomaterials.2013.09.052
70. Hu S, Ma J, Su C, et al. Engineered exosome-like nanovesicles suppress tumor growth by reprogramming tumor microenvironment and promoting tumor ferroptosis. Acta Biomater. 2021;135:567–581. doi:10.1016/j.actbio.2021.09.003
71. Huang L, Rong Y, Tang X, et al. Engineered exosomes as an in situ DC-primed vaccine to boost antitumor immunity in breast cancer. Mol Cancer. 2022;21(1):45. doi:10.1186/s12943-022-01515-x
72. Wang Z, Mo H, He Z, Chen A, Cheng P. Extracellular vesicles as an emerging drug delivery system for cancer treatment: current strategies and recent advances. Biomed Pharmacother. 2022;153:113480. doi:10.1016/j.biopha.2022.113480
73. Xia T, Liu Z, Du Y, et al. Bifunctional iRGD-Exo-DOX crosses the blood-brain barrier to target central nervous system lymphoma. Biochem Pharmacol. 2024;223:116138. doi:10.1016/j.bcp.2024.116138
74. Barkovich KJ, Zhao Z, Steinmetz NF. iRGD-targeted physalis mottle virus-like nanoparticles for targeted cancer delivery. Small Sci. 2023;3(8):2300067. doi:10.1002/smsc.202300067
75. Yang Z, Tian T, Kong J, Chen H. ChatExosome: an Artificial Intelligence (AI) agent based on deep learning of exosomes spectroscopy for Hepatocellular Carcinoma (HCC) diagnosis. Anal Chem. 2025;97(8):4643–4652. doi:10.1021/acs.analchem.4c06677
76. Fu S, Wang Y, Xia X, Zheng JC. Exosome engineering: current progress in cargo loading and targeted delivery. NanoImpact. 2020;20:100261. doi:10.1016/j.impact.2020.100261
77. Donoso-Quezada J, Ayala-Mar S, Gonzalez-Valdez J. State-of-the-art exosome loading and functionalization techniques for enhanced therapeutics: a review. Crit Rev Biotechnol. 2020;40(6):804–820. doi:10.1080/07388551.2020.1785385
78. Jhan YY, Palou Zuniga G, Singh KA, Gaharwar AK, Alge DL, Bishop CJ. Polymer-coated extracellular vesicles for selective codelivery of chemotherapeutics and sirna to cancer cells. ACS Appl Bio Mater. 2021;4(2):1294–1306. doi:10.1021/acsabm.0c01153
79. Ahmed W, Mushtaq A, Ali S, Khan N, Liang Y, Duan L. Engineering approaches for exosome cargo loading and targeted delivery: biological versus chemical perspectives. ACS Biomater Sci Eng. 2024;10(10):5960–5976. doi:10.1021/acsbiomaterials.4c00856
80. Ashrafizadeh M, Rabiee N, Kumar AP, Sethi G, Zarrabi A, Wang Y. Long noncoding RNAs (lncRNAs) in pancreatic cancer progression. Drug Discov Today. 2022;27(8):2181–2198. doi:10.1016/j.drudis.2022.05.012
81. Meena JK, Wang JH, Neill NJ, et al. MYC induces oncogenic stress through RNA decay and ribonucleotide catabolism in breast cancer. Cancer Discov. 2024;14(9):1699–1716. doi:10.1158/2159-8290.CD-22-0649
82. Liu C, Su C. Design strategies and application progress of therapeutic exosomes. Theranostics. 2019;9(4):1015–1028. doi:10.7150/thno.30853
83. Wang F, Lin S, Yu Z, et al. Recent advances in microfluidic-based electroporation techniques for cell membranes. Lab Chip. 2022;22(14):2624–2646. doi:10.1039/D2LC00122E
84. Shi L, Li B, Tan J, et al. Exosomal lncRNA Mir100hg from lung cancer stem cells activates H3K14 lactylation to enhance metastatic activity in non-stem lung cancer cells. J Nanobiotechnol. 2025;23(1):156. doi:10.1186/s12951-025-03198-0
85. Zhu L, Liu J, Li J, Wang N, Zhao Y, Su H. Research progress on HER2-specific chimeric antigen receptor T cells for immunotherapy of solid tumors. Front Immunol. 2025;16:1514994. doi:10.3389/fimmu.2025.1514994
86. Vasave R, Bhawale R, Khan O, et al. Translational potential of surface engineered extracellular vesicles as theranostic nanocarriers in cancer management: recent developments from bench to bedside. ACS Appl Bio Mater. 2025;8(8):6647–6675. doi:10.1021/acsabm.5c00955
87. Wang J, Zhu X, Jiang H, Ji M, Wu Y, Chen J. Cancer cell-derived exosome based dual-targeted drug delivery system for non-small cell lung cancer therapy. Colloids Surf B Biointerfaces. 2024;244:114141. doi:10.1016/j.colsurfb.2024.114141
88. Chen J, Cao F, Cao Y, Wei S, Zhu X, Xing W. Targeted therapy of lung adenocarcinoma by the nanoplatform based on milk exosomes loaded with paclitaxel. J Biomed Nanotechnol. 2022;18(4):1075–1083. doi:10.1166/jbn.2022.3278
89. Zhu T, Chen Z, Jiang G, Huang X. Sequential targeting hybrid nanovesicles composed of chimeric antigen receptor T-cell-derived exosomes and liposomes for enhanced cancer immunochemotherapy. ACS Nano. 2023;17(17):16770–16786. doi:10.1021/acsnano.3c03456
90. Wang Y, Guo M, Lin D, et al. Docetaxel-loaded exosomes for targeting non-small cell lung cancer: preparation and evaluation in vitro and in vivo. Drug Deliv. 2021;28(1):1510–1523. doi:10.1080/10717544.2021.1951894
91. Lin Q, Qu M, Zhou B, et al. Exosome-like nanoplatform modified with targeting ligand improves anti-cancer and anti-inflammation effects of imperialine. J Control Release. 2019;311–312:104–116. doi:10.1016/j.jconrel.2019.08.037
92. Kim MS, Haney MJ, Zhao Y, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations. Nanomedicine. 2018;14(1):195–204. doi:10.1016/j.nano.2017.09.011
93. Cho H, Jung I, Ju H, Baek MC, Yea K. Engineered CD8+ T cell-derived extracellular vesicles induce enhanced anti-cancer efficacy and targeting to lung cancer cells. Cytokine. 2023;169:156249. doi:10.1016/j.cyto.2023.156249
94. Liang G, Zhu Y, Ali DJ, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnol. 2020;18(1):10. doi:10.1186/s12951-019-0563-2
95. Wu H, Fu M, Liu J, et al. The role and application of small extracellular vesicles in gastric cancer. Mol Cancer. 2021;20(1):71. doi:10.1186/s12943-021-01365-z
96. Do AD, Kurniawati I, Hsieh CL, Wong TT, Lin YL, Sung SY. Application of mesenchymal stem cells in targeted delivery to the brain: potential and challenges of the extracellular vesicle-based approach for brain tumor treatment. Int J Mol Sci. 2021;22(20):11187. doi:10.3390/ijms222011187
97. Shan S, Chen J, Sun Y, et al. Functionalized macrophage exosomes with panobinostat and PPM1D-siRNA for diffuse intrinsic pontine gliomas therapy. Adv Sci. 2022;9(21):e2200353. doi:10.1002/advs.202200353
98. Niu W, Xiao Q, Wang X, et al. A biomimetic drug delivery system by integrating grapefruit extracellular vesicles and doxorubicin-loaded heparin-based nanoparticles for glioma therapy. Nano Lett. 2021;21(3):1484–1492. doi:10.1021/acs.nanolett.0c04753
99. Li Q, Song Q, Zhao Z, et al. Genetically engineered artificial exosome-constructed hydrogel for ovarian cancer therapy. ACS Nano. 2023;17(11):10376–10392. doi:10.1021/acsnano.3c00804
100. Wang J, Wang C, Li Y, et al. Potential of peptide-engineered exosomes with overexpressed miR-92b-3p in anti-angiogenic therapy of ovarian cancer. Clin Transl Med. 2021;11(5):e425. doi:10.1002/ctm2.425
101. Suo M, Shen H, Lyu M, et al. Biomimetic nano-cancer stem cell scavenger for inhibition of breast cancer recurrence and metastasis after FLASH-radiotherapy. Small. 2024;20(29):e2400666. doi:10.1002/smll.202400666
102. Farheen J, Iqbal MZ, Lu Y, Tang Z, Kong X. Vitis vinifera Kyoho-derived exosome-like nanoparticles-based drug delivery and therapeutic modalities for breast cancer therapy. J Drug Deliv Sci Technol. 2024;92:105332. doi:10.1016/j.jddst.2023.105332
103. Bahreyni A, Mohamud Y, Ashraf Nouhegar S, Zhang J, Luo H. Synergistic viro-chemoimmunotherapy in breast cancer enabled by bioengineered immunostimulatory exosomes and dual-targeted coxsackievirus B3. ACS Nano. 2024;18(5):4241–4255. doi:10.1021/acsnano.3c09491
104. Liu Y, Bai L, Guo K, et al. Focused ultrasound-augmented targeting delivery of nanosonosensitizers from homogenous exosomes for enhanced sonodynamic cancer therapy. Theranostics. 2019;9(18):5261–5281. doi:10.7150/thno.33183
105. Jayaseelan VP, Paramasivam A. Engineered exoASO-STAT6: a potent monotherapy for oral squamous cell carcinoma. Future Oncol. 2022;18(25):2767–2769. doi:10.2217/fon-2022-0496
106. Yang M, Luo Q, Chen X, Chen F. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J Nanobiotechnol. 2021;19(1):259. doi:10.1186/s12951-021-00995-1
107. Huang X, Wu W, Jing D, et al. Engineered exosome as targeted lncRNA MEG3 delivery vehicles for osteosarcoma therapy. J Control Release. 2022;343:107–117. doi:10.1016/j.jconrel.2022.01.026
108. Tian W, Niu X, Feng F, et al. The promising roles of exosomal microRNAs in osteosarcoma: a new insight into the clinical therapy. Biomed Pharmacother. 2023;163:114771. doi:10.1016/j.biopha.2023.114771
109. Chang S, Li L, Liu Y, Huang G, Pan ST. Advances in tumor micro-environment and immunotherapy of squamous cell carcinoma of the tongue. Biochimica et Biophysica Acta. 2025;189483. doi:10.1016/j.bbcan.2025.189483
110. Biederstädt A, Rezvani K. Engineered natural killer cells for cancer therapy. Cancer Cell. 2025. doi:10.1016/j.ccell.2025.09.013
111. Chen Z, Li J, Wang J, et al. Bacteria modulate tumor-associated macrophages to combat cancer. Crit Rev Oncol/Hematol. 2025:104983. doi:10.1016/j.critrevonc.2025.104983
112. Fu S, Gao C, Yu Q, et al. Design of nano-delivery systems for pancreatic cancer immunotherapy. Cancer Lett. 2025;635:218098. doi:10.1016/j.canlet.2025.218098
113. Marquez CA, Oh CI, Ahn G, Shin WR, Kim YH, Ahn JY. Synergistic vesicle-vector systems for targeted delivery. J Nanobiotechnol. 2024;22(1):6. doi:10.1186/s12951-023-02275-6
114. Xu J, Liu H, Wang T, et al. CCR7 mediated mimetic dendritic cell vaccine homing in lymph node for head and neck squamous cell carcinoma therapy. Adv Sci. 2023;10(17):e2207017. doi:10.1002/advs.202207017
115. Wang S, Li F, Ye T, et al. Macrophage-tumor chimeric exosomes accumulate in lymph node and tumor to activate the immune response and the tumor microenvironment. Sci Transl Med. 2021;13(615):eabb6981. doi:10.1126/scitranslmed.abb6981
116. Yang Q, Li S, Ou H, et al. Exosome-based delivery strategies for tumor therapy: an update on modification, loading, and clinical application. J Nanobiotechnol. 2024;22(1):41. doi:10.1186/s12951-024-02298-7
117. Lu Q, Kou D, Lou S, et al. Nanoparticles in tumor microenvironment remodeling and cancer immunotherapy. J Hematol Oncol. 2024;17(1):16. doi:10.1186/s13045-024-01535-8
118. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
119. Zhang J. Exosome and exosomal microrna: trafficking, sorting, and function. Genomics Proteomics Bioinf. 2015;13(1):17–24. doi:10.1016/j.gpb.2015.02.001
120. Yang M, Zhou W, Han X, et al. Modified bone marrow mesenchymal stem cells derived exosomes loaded with MiRNA ameliorates non-small cell lung cancer. J Cell Mol Med. 2024;28(18):e70115. doi:10.1111/jcmm.70115
121. Shimizu A, Sawada K, Kimura T. Pathophysiological role and potential therapeutic exploitation of exosomes in ovarian cancer. Cells. 2020;9(4):814. doi:10.3390/cells9040814
122. Abdullaev B, Rasyid SA, Ali E, et al. Effective exosomes in breast cancer: focusing on diagnosis and treatment of cancer progression. Pathol Res Pract. 2024;253:154995. doi:10.1016/j.prp.2023.154995
123. Moghadasi S, Elveny M, Rahman HS, et al. A paradigm shift in cell-free approach: the emerging role of MSCs-derived exosomes in regenerative medicine. J Transl Med. 2021;19(1):302. doi:10.1186/s12967-021-02980-6
124. Dietz L, Oberländer J, Mateos-Maroto A, et al. Uptake of extracellular vesicles into immune cells is enhanced by the protein Corona. J Extracell Vesicles. 2023;12(12):e12399. doi:10.1002/jev2.12399
125. Lennaárd AJ, Mamand DR, Wiklander RJ, El Andaloussi S, Wiklander OPB. Optimised electroporation for loading of extracellular vesicles with doxorubicin. Pharmaceutics. 2021;14(1):38. doi:10.3390/pharmaceutics14010038
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Application of Biomimetic Cell Membrane-Coated Nanocarriers in Cardiovascular Diseases

Zhao X, Chen W, Wu J, Shen Y, Xu B, Chen Z, Sun Y
International Journal of Nanomedicine 2025, 20:8249-8289
Published Date: 26 June 2025
Advances of Drug-Loaded Microsphere Technology for Targeted Immunotherapy Against Prostate Cancer
Feng W
International Journal of Nanomedicine 2025, 20:11479-11489
Published Date: 20 September 2025
Innovative Applications of Nucleic Acid Aptamers in Colorectal Cancer Diagnosis and Therapy: From Selection Optimization to Clinical Translation
Lu JF, Zhang XR, Liu Y, Zhang CL, Wang L, Liu ZQ, Cai J, Yi HW
International Journal of Nanomedicine 2025, 20:13437-13456
Published Date: 6 November 2025
Engineered Exosomes Co-Delivering EGF and FGF Ameliorate Androgenetic Alopecia in a Mouse Model
Lai Y, Wu J, Tan T, Gao B, Han J, Liu Z, Ding X
International Journal of Nanomedicine 2026, 21:572518
Published Date: 16 February 2026
Nanomedicine in 2026: Illustrative Quantitative Analyses of EPR Heterogeneity, Clinical Trial Attrition, and Emerging Horizons for Active Nanotherapeutics
Fayez SM
International Journal of Nanomedicine 2026, 21:618407
Published Date: 11 June 2026