Back to Journals » International Journal of Nanomedicine » Volume 19
Engineered Extracellular Vesicles in Chronic Kidney Diseases: A Comprehensive Review
Received 29 November 2023
Accepted for publication 19 February 2024
Published 7 March 2024 Volume 2024:19 Pages 2377—2393
DOI https://doi.org/10.2147/IJN.S452393
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. RDK Misra
Kaming Xue,1 Bobin Mi2
1Department of Integrated Traditional Chinese and Western Medicine, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China; 2Department of Orthopedics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China
Correspondence: Bobin Mi, Department of Orthopedics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China, Email [email protected]
Abstract: Chronic kidney diseases (CKD) present a formidable global health challenge, characterized by a deficiency of effective treatment options. Extracellular vesicles (EVs), recognized as multifunctional drug delivery systems in biomedicine, have gained accumulative interest. Specifically, engineered EVs have emerged as a promising therapeutic approach for targeted drug delivery, potentially addressing the complexities of CKD management. In this review, we systematically dissect EVs, elucidating their classification, biogenesis, composition, and cargo molecules. Furthermore, we explore techniques for EV engineering and strategies for their precise renal delivery, focusing on cargo loading and transportation, providing a comprehensive perspective. Moreover, this review also discusses and summarizes the diverse therapeutic applications of engineered EVs in CKD, emphasizing their anti-inflammatory, immunomodulatory, renoprotective, and tissue-regenerating effects. It critically evaluates the challenges and limitations in translating EV therapies from laboratory settings to clinical applications, while outlining future prospects and emerging trends.
Keywords: extracellular vesicles, nanomaterials, chronic kidney diseases, drug delivery systems, application
Graphical Abstract:
Introduction
Chronic Kidney Disease (CKD) is a pervasive global health issue, marked by its significant impact on individuals and healthcare systems worldwide.1 CKD is defined by the progressive deterioration of kidney function over time, leading to a range of complications and posing a substantial burden on affected individuals and society as a whole. The prevalence of CKD continues to rise, driven by factors such as aging populations and the increasing prevalence of conditions like diabetes and hypertension, which are major risk factors for CKD.2 As a result, it is paramount to address the challenges associated with CKD management, as current approaches have limitations and are often unable to halt or reverse the progression of the disease.
The current landscape of CKD management is fraught with several challenges. One of the foremost issues is the delayed diagnosis of CKD, as the disease often remains asymptomatic until it reaches an advanced stage.3 This delays the initiation of therapeutic interventions, reducing their effectiveness. Furthermore, the progressive nature of CKD and the insidious development of complications, including cardiovascular disease, anemia, and bone disorders, necessitates multifaceted management approaches that can be cumbersome for both patients and healthcare providers.4 Treatments for CKD primarily focus on symptom management, and patients with advanced CKD often require renal replacement therapy, such as dialysis or kidney transplantation, which come with their own set of challenges, including significant costs and limited availability of donor organs.
In light of these challenges, there is a growing recognition of the potential of EVs as a novel approach to CKD therapy.5 EVs, tiny vesicles secreted by various cell types, are involved in numerous physiological processes, including immune response regulation, tissue repair, and waste elimination, all of which are vital to maintain kidney health. Additionally, they play a role in pathological conditions, such as inflammation, fibrosis, and oxidative stress, which are central to CKD pathogenesis.6 The contents of EVs in urine, including miRNAs, mRNAs, and proteins, show promise as potential biomarkers for this condition, providing a glimmer of hope for improved diagnostics and understanding of disease mechanisms.7 The multifaceted role of EVs in kidney disease underscores their potential as therapeutic targets and diagnostic tools in this and other renal disorders.
With the advances in the field of nanobiotechnology, engineered EVs have emerged as a promising therapeutic alternative for diseases. Engineered EVs exhibit several key advantages over their natural counterparts in CKD therapy. Firstly, these engineered EVs can be functionally modified to target specific kidney cells or affected areas, thus enhancing their precision and therapeutic effectiveness. For instance, engineered red blood cell-derived EVs with designed peptides could target with kidney injury molecule-1 to alleviate kidney injury.8 Secondly, they provide a controlled platform for payload delivery, enabling the loading of therapeutic agents such as drugs or RNA molecules with precision and efficiency. For instance, Yoon et al reported that melatonin-engineered EVs boosted regenerative abilities of CKD patient-derived MSCs, indicating a promising strategy for developing autologous MSC-based therapeutics for patients with CKD.9 Thirdly, the stability of engineered EVs can be improved, extending their circulation time in the bloodstream and ultimately enhancing their therapeutic impact. Fourth, these modifications can also reduce the risk of an immune response, rendering engineered EVs safer for long-term CKD treatment.10 Lastly, engineered EVs can be produced at a larger scale than natural EVs, making them more feasible for clinical applications in CKD therapy.11 These combined advantages position engineered EVs as a promising avenue for the development of targeted and effective CKD treatments, with the potential to enhance patient outcomes and reduce the burden of this chronic condition.
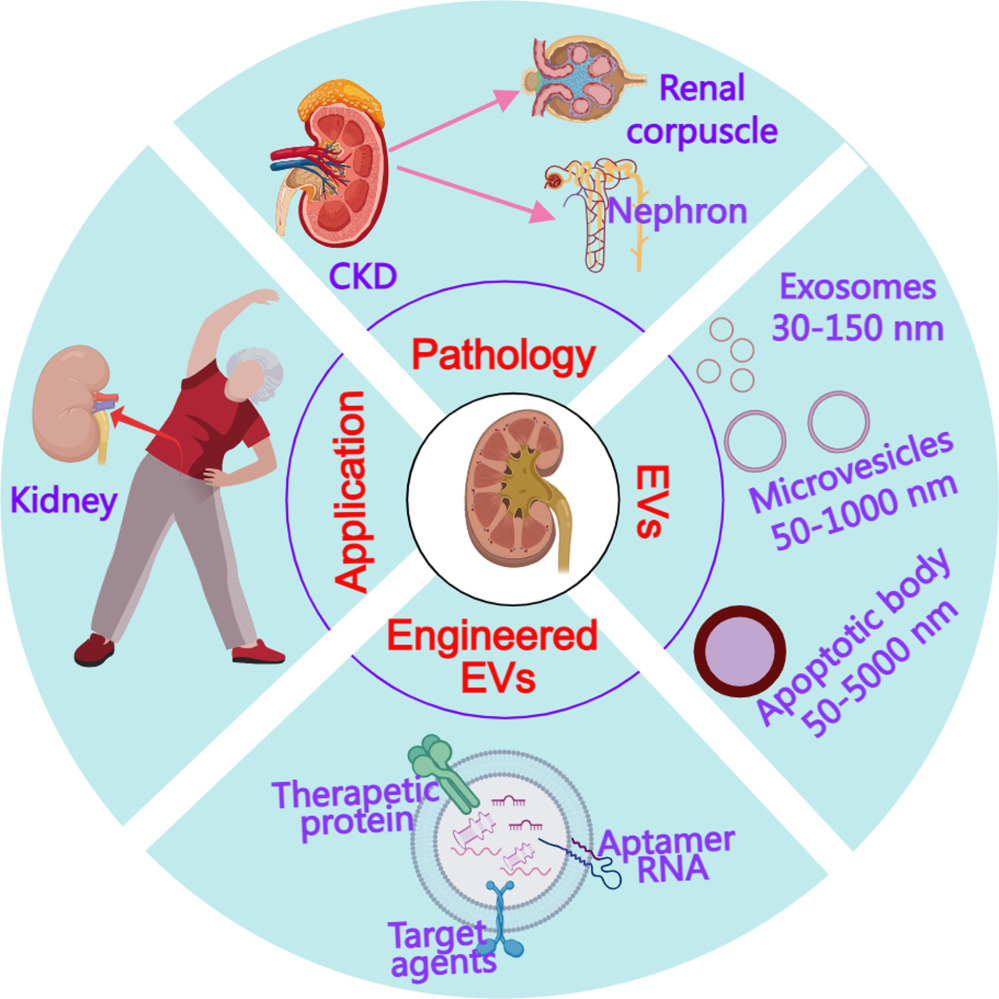
Therefore, in this review, we aim to elucidate the multifaceted involvement of engineered EVs in CKD therapy. We provide an in-depth examination of their potential to address the multiple challenges inherent to CKD management. Through the synthesis of critical research findings and the emphasis on the imperative need for ongoing investigations, this review serves as an invaluable resource for developing pioneering innovative treatments for CKD. Ultimately, the exploration of engineered EV-based therapies offers considerable promise in the complex landscape of CKD management, potentially yielding more efficacious interventions and enhancing the quality of life for individuals grappling with this relentless disease (Figure 1).
 |
Figure 1 The present paper overview of chronic kidney disease (CKD) pathology, explores the involvement of extracellular vesicles (EVs) in CKD progression and therapy, and examines the methods of enhancing EV function through Engineered EVs, along with their potential application in treating CKD. The pink arrows represent the components of CKD. The red arrow represents kidney derived from people. |
Types and Biogenesis of EVs
Definition and Classification of EVs
EVs represent a complex domain of cell biology, providing valuable insights into intercellular communication and their critical roles in numerous physiological and pathological processes. These remarkable cell-derived membranous structures encompass exosomes and microvesicles, each distinctive in its origin and characteristics.12 Exosomes, typically measuring 30–150 nanometers in diameter, emerge from the endosomal system, originating within multivesicular bodies (MVBs), and are subsequently liberated into the extracellular milieu upon fusion of these MVBs with the cell’s plasma membrane.13 In contrast, microvesicles, somewhat larger with a size ranging from 100–1000 nanometers, diverge from the endosomal exosome biogenesis route.14 Instead, they are directly shed from the plasma membrane of cells, carrying a rich cargo of proteins, lipids, and genetic material. Furthermore, within this diverse landscape of EVs, apoptotic bodies occupy their niche, emerging from cells undergoing programmed cell death, known as apoptosis.15
These classifications provide a framework for understanding the heterogeneity of EVs and highlight their significance in mediating intercellular communication (Table 1). Exosomes, with their well-defined biogenesis pathway, are particularly intriguing. They serve as conduits for the transfer of crucial cellular components, including proteins, lipids, and genetic material, to neighboring or distant cells.16 The unique mechanism of their biogenesis within MVBs, followed by their release via fusion with the plasma membrane, underscores their role as specialized messengers in cell-to-cell communication.17 Microvesicles, on the other hand, exhibit a distinct mode of origin. Unlike exosomes, they are directly shed from the plasma membrane, carrying a diverse cargo of cellular materials. This dichotomy in biogenesis mechanisms between exosomes and microvesicles suggests a broad range of roles in intercellular signaling, making them integral players in various physiological and pathological contexts.18 Intriguingly, apoptotic bodies represent a specialized class of EVs that emerge during programmed cell death, or apoptosis. As larger vesicles, they encapsulate the remnants of the dying cell, including fragmented DNA and organelles. This unique characteristic position apoptotic bodies as potential participants in the clearance of cellular debris and the regulation of immune responses.19 Furthermore, their contents, which may include bioactive molecules, can potentially influence the surrounding microenvironment, affecting neighboring cells and influencing tissue homeostasis. For instance, Liu et al reported that transplanted MSCs release apoptotic bodies, which upon uptake by recipient endothelial cells, activate lysosome functions and enhancing autophagy, thus revealing the role of donor cell apoptosis in MSC therapeutic effects.20
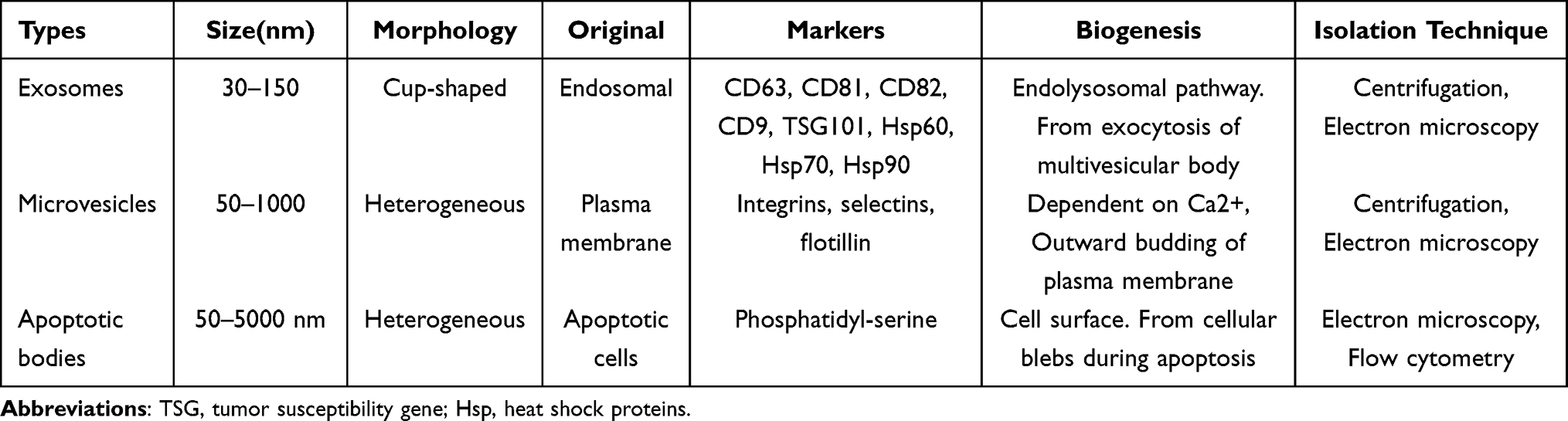 |
Table 1 Characteristic of Different EVs |
Biogenesis and Release of EVs
The release of EVs enables the transfer of their cargo, including proteins, lipids, and various nucleic acids (such as RNA and DNA), to recipient cells. This content exchange drives intercellular communication and significantly influences numerous physiological and pathological processes. Understanding the biogenesis and release of EVs is vital for unraveling their roles in health and disease.21 Exosomes and microvesicles are involved in a multitude of processes, from contributing to normal cell functions to playing critical roles in various diseases. These vesicles are now recognized as an additional mode of cellular communication, allowing cells to exchange vital information in the form of their molecular cargo.22 Despite the growing knowledge in this field, there is still much to learn about the specifics of EV biogenesis and release. In particular, the precise regulation of these processes and the factors that influence them are active areas of research. Additionally, as the field expands, the cargo within EVs, including microRNAs, mRNAs, and proteins, has garnered significant attention due to its potential as a means of intercellular signaling and as a source of biomarkers for various diseases.23
Proteins within EVs encompass a broad spectrum of enzymes, receptors, and signaling molecules that can influence recipient cells upon EV uptake, modulating cellular processes and pathways.24 Simultaneously, the presence of lipids, including phospholipids, cholesterol, and other lipid species, contributes to EV membrane integrity and facilitates potential interactions with target cells.25 Nucleic acids, a significant component of EV cargo, include small and long non-coding RNA molecules, which play a pivotal role in post-transcriptional gene regulation. The transfer of EV-delivered noncoding RNAs to target cells can lead to gene expression changes.26,27 Additionally, EVs may carry messenger RNAs (mRNAs) that can be translated into proteins upon uptake by recipient cells, potentially altering the protein expression profile of the target cells.28 Furthermore, the presence of DNA, including genomic DNA, within some EVs adds another layer of complexity to their cargo composition, and ongoing research seeks to uncover the functional significance of DNA within EVs and its potential impact on recipient cells.29 Understanding the diverse composition and cargo of EVs is instrumental in harnessing their potential in diagnostic and therapeutic applications, as these cargo molecules can profoundly influence cellular processes, signaling pathways, and even epigenetic changes, making EVs versatile mediators of intercellular communication in both normal and pathological contexts.
Role of EVs in CKD
EVs play multifaceted roles in the pathophysiology of CKD, from modulating physiological processes within the kidney to serving as potential biomarkers for disease detection and monitoring. In the following section, we will summary the current finding of EVs in the CKD (Figure 2).
 |
Figure 2 EVs originating from various sources such as stem cells, the kidney, and other organs play a crucial role in regulating both the physiological functions and pathological processes within the kidney. Additionally, EVs derived from plasma hold potential as biomarkers for detecting CKD. The pink arrow represents EVs could be transported to kidney. The brown arrows represent the biogenesis and release of EVs. |
Physiological Functions of EVs in the Kidney
EVs play pivotal roles in kidney physiology, orchestrating a sophisticated network of cellular communication and molecular regulation within this vital organ. These membrane-bound, nano-sized particles serve as messengers for various cell types, transporting a cargo of macromolecules, with miRNA taking a prominent role.30 The miRNAs encapsulated within EVs play a crucial part in orchestrating gene expression within recipient cells through precise base-pairing interactions with mRNAs, resulting in either the degradation or translational inhibition of target mRNAs. This intricate regulatory mechanism enables EVs to influence gene expression profiles and facilitate the exchange of genetic information among kidney cells, contributing to the maintenance of normal physiological processes.31 For instance, Intramuscular injection of exosome-encapsulated miR-29, directed by the RVG peptide, effectively attenuated muscle wasting induced by unilateral ureteral obstruction (UUO) and partially depressed renal fibrosis, evidenced by decreased expression of TRIM63/MuRF1, FBXO32/atrogin-1, TGF-β, alpha-smooth muscle actin, fibronectin, and collagen 1A1.32
Notably, urine serves as a unique reservoir of EVs originating from the kidney, as these vesicles are released into the urine, either from within the kidney or after crossing the glomerular filtration barrier.33 It has been reported that the majority of EVs detectable in the urine of normal people are originated almost all nephron segments, including glomerular podocytes and tubular cells at proximal convoluted tubule, thick ascending limb of the Henle’s loop, distal convoluted tubule and collecting duct.34 EVs released in urine carry specific miRNAs that reflect kidney health or pathology, so urine-based EVs bring some new biomarkers to the diagnosis of kidney disease. In a previous study, Chen et al investigated the potential of urinary exosomal tRNA-derived small noncoding RNAs as novel biomarkers for diagnosing lupus nephritis in systemic lupus erythematosus patients, demonstrating upregulated tRF3-Ile-AAT-1 and tiRNA5-Lys-CTT-1 levels as distinguishing factors, with implications for immune regulation and activity assessment in lupus nephritis.35 These findings offer broad prospects for the diagnosis and monitoring of kidney disease, making it possible for non-invasive techniques to replace invasive techniques, such as kidney biopsy.
In addition to their diagnostic potential, EVs also play a critical role in kidney repair and regeneration. These vesicles have the capacity to support tissue recovery and repair within the kidney. Stem cell-derived EVs, in particular, have shown promise in enhancing renal recovery. For instance, investigated the mechanism by which stem cell-derived EVs alleviate vascular calcification in CKD, finding that their effect on inhibiting cellular apoptosis and calcification partly relies on exosomal microRNA-381-3p (miR-381-3p), which directly targets nuclear factor of activated T cells 5 (NFAT5) expression, as validated in high phosphate-stimulated human aortic smooth muscle cells (HA-VSMCs) and 5/6 subtotal nephrectomy (SNx) rat models, suggesting a potential therapeutic target for CKD.36 This highlights the regenerative potential of EVs and their role in mitigating the consequences of kidney damage.
EVs have emerged as crucial mediators of intercellular communication within the intricate microenvironment of the kidney.37 These membrane-surrounded structures, released by nearly all cell types within the kidney, participate in a diverse array of signaling pathways, influencing both normal physiological processes and the development of kidney-related diseases. In the context of kidney physiology, EVs facilitate intercellular crosstalk, enhancing communication between various cell types within the nephron.38 They contribute to the maintenance of tissue homeostasis and play a pivotal role in the regulation of kidney function. Furthermore, EVs extend their influence beyond the kidney, acting as potent intercellular messengers that can communicate with distant cells in the body, thereby influencing systemic processes.39,40
In summary, EVs in the kidney serve multifaceted roles, functioning as molecular messengers that mediate intercellular communication, regulators of gene expression, potential biomarkers for kidney health and disease, promoters of renal repair and regeneration, and emerging tools for targeted therapeutic intervention. The exploration of EVs in kidney physiology represents a promising avenue for advancing our understanding of kidney function and developing novel strategies for diagnosing and treating kidney diseases, ultimately improving patient outcomes.
Pathological Alterations of EVs in CKD
In the context of CKD, EVs undergo pathological alterations that significantly impact the disease’s progression. CKD is characterized by a progressive loss of renal function and involves various cellular and molecular changes, many of which are mediated or influenced by EVs. One of the key pathological alterations observed in CKD is the increased release of EVs from various cell types within the kidney and the systemic circulation.41 These EVs carry a cargo of pro-inflammatory, pro-fibrotic, and pro-apoptotic molecules, contributing to the perpetuation of kidney damage. The inflammation observed in CKD is, in part, driven by EVs released from immune cells and injured renal cells, which contain pro-inflammatory cytokines, chemokines, and adhesion molecules, creating a sustained inflammatory response within the kidney—a central feature of CKD progression. For instance, Lv et al investigated the role of exosome mRNA released from tubular epithelial cells (TECs) in propagating tubulointerstitial inflammation associated with CKD, finding that increased release of exosomes containing inflammatory cytokine mRNA, particularly CCL2, from TECs occurred in acute and chronic kidney injury models and in response to albumin stimulation, with subsequent internalization by macrophages leading to enhanced inflammatory response and macrophage migration.42
Furthermore, the fibrosis observed in CKD, characterized by the excessive deposition of extracellular matrix proteins, is influenced by EVs. These EVs transport fibrotic markers such as transforming growth factor-beta (TGF-β) and connective tissue growth factor (CTGF), which stimulate fibroblast activation and collagen deposition, exacerbating renal fibrosis.43 In one study, Chen et al identified that renal tubular cell-derived exosomes, stimulated by β-catenin, transport osteopontin (OPN) to fibroblasts, promoting their proliferation and activation via CD44 binding, contributing to renal fibrosis, a mechanism validated in both experimental models and CKD patients, underscoring the significance of exosome-mediated communication in renal fibrosis pathogenesis.44 Alongside inflammation and fibrosis, EVs contribute to endothelial dysfunction in CKD. EVs released in this context promote oxidative stress, inflammation, and the disruption of nitric oxide signaling within the endothelium. The transfer of miRNAs through EVs plays a pivotal role in CKD and coronary artery disease (CAD). The combination of CKD and CAD results in altered miRNA-mediated vascular intercellular communication, which can promote CKD-induced endothelial dysfunction.45 Specific miRNAs carried by EVs are found to be dysregulated in patients with CAD and CKD, affecting endothelial function and cellular crosstalk. This phenomenon highlights the importance of miRNA-containing EVs in the pathogenesis of these diseases.46 Consequently, endothelial dysfunction ensues, leading to complications such as hypertension and an increased risk of cardiovascular events, both of which are common comorbidities in CKD patients.47
Moreover, the renal cells injured in CKD release EVs carrying stress and apoptosis-related molecules, including heat shock proteins, caspases, and mitochondrial DNA fragments.48,49 These EVs perpetuate cell injury within the kidney, thus further exacerbating the loss of renal function. The tubulointerstitial injury, marked by tubular atrophy and interstitial fibrosis, is another key pathological feature of CKD influenced by EVs. These vesicles deliver pro-inflammatory and pro-fibrotic signals to tubular and interstitial cells, contributing to this injurious process. In a previous study, Li et al investigated how hypoxia induces tubulointerstitial inflammation in the kidney and found that increased tubular expression of HIF-1α leads to selective shedding of microRNA-23a (miRNA-23a)-enriched exosomes, which, when taken up by macrophages, triggers their pro-inflammatory activation via suppression of the ubiquitin editor A20, indicating a potential therapeutic target for ameliorating tubulointerstitial inflammation.50
EVs also participate in immune dysregulation within the CKD environment. By modulating immune responses, they may contribute to immune system dysfunction and the chronic inflammatory state often observed in CKD patients. Their influence on the balance between pro-inflammatory and anti-inflammatory immune cell subsets is a complex aspect of the disease pathogenesis.51 For instance, EVs derived from infiltrated macrophage amplify vascular calcification in CKD patients through active SMAD3/RUNX2 signaling.6 Furthermore, EVs are intricately involved in the regulation of mineral metabolism in CKD, which transport factors that impact phosphate and calcium homeostasis, which can contribute to the development of mineral and bone disorders, a common complication in CKD.52
EVs as Biomarkers for CKD Detection
Electrolyte disturbances are common in patients with CKD, making the exploration of novel biomarkers imperative for early detection and management. Emerging evidence suggests that EVs hold promise as potential biomarkers for CKD. These small vesicles, released by various cells including renal epithelial cells, carry molecular cargo reflective of their cellular origin and physiological state.53 In the context of CKD, EVs may contain proteins, lipids, nucleic acids, and metabolites that can provide insights into renal pathophysiology and disease progression. Importantly, EVs are found in various biofluids, including urine and blood, offering non-invasive avenues for sample collection.54 By analyzing the composition and abundance of EVs, researchers aim to identify specific biomarkers indicative of CKD severity, progression, and response to therapy. In a previous study, Haney et al investigated mRNA content in urinary small EVs and gene expression in kidney and bladder tissues of rats with upper and lower urinary tract obstruction, finding unique gene signatures for each obstruction type in urinary sEVs, suggesting their potential as biomarkers for diagnosing and prognosticating upper urinary tract obstruction (UUTO) independently of bladder function and architecture.55 In another study, Tao et al identify novel biomarkers for CKD diagnosis using EV-derived mRNA from kidney tissue microarray data, resulting in the confirmation of AEBP1 levels in plasma EVs as a promising biomarker and demonstrating good diagnostic efficacy for CKD.56 Furthermore, EVs exhibit stability in circulation, potentially enabling longitudinal monitoring of CKD progression and treatment efficacy. For instance, Saejong et al reported that high expression of miR-21 in plasma exosomes, but not in whole plasma, was indicative of high-grade interstitial fibrosis and tubular atrophy, offering a potential for more frequent monitoring and earlier management compared to renal biopsy.57 However, the heterogeneity of EV populations poses challenges in pinpointing specific subsets relevant to CKD pathogenesis. Despite these hurdles, continued efforts in elucidating the intricate roles of EVs in CKD pathophysiology and refining analytical methodologies will be pivotal in harnessing the full potential of EVs as biomarkers for CKD detection.
Engineering EVs for Therapeutic Applications
Methods for EV Modification and Engineering
In regenerative medicine, EVs have been studied extensively, with various engineering approaches employed to enhance their regenerative potential. These methods include preconditioning, drug loading, and surface modification to potentiate the therapeutic outcomes of naturally secreted EVs.58,59 The limitations of natural EVs have also led to the development of artificial EVs, which offer advantages in terms of production yield and isolation methodologies, making them an attractive alternative for therapeutic applications. Material design plays a pivotal role in the modification and engineering of EVs. Researchers have utilized genetic engineering and chemical modifications to improve the stability, release rate, and biodistribution of EVs.60 Additionally, hydrogel and nano-formulation strategies have been explored to increase the persistence of EVs in specific tissues or organs, further enhancing their therapeutic potential.61
One of the key methods is the reversible heterodimerization of Cas9-fusions with EV sorting partners, facilitating the loading of CRISPR/Cas9 into EVs.62 This innovative approach has turned EVs into a powerful gene-editing tool. For instance, Ilahibaks et al investigated EV-mediated CRISPR/Cas9 ribonucleoprotein (RNP) delivery targeting the Pcsk9 gene in mouse hepatocytes ex vivo, successfully inactivating Pcsk9 and reducing plasma LDL-C levels, offering insights for future in vivo studies utilizing this approach for Pcsk9 gene targeting.63 Furthermore, the combination of light-induced dimerization and lipid modifications, such as Myristoylation-Palmitoylation-Palmitoylation, has proven highly efficient for loading Cas9 into EVs, resulting in approximately 25 Cas9 molecules per EV and significant functional gene editing in various cellular assays.64 Additionally, surface modification of EVs has been extensively explored, with researchers developing methods to tether ligands to the EV surface, which modification allows for specific targeting of cells, significantly enhancing the ability of EVs to deliver cargo to the desired cellular targets.65 A general and mild modification method, involving the reduction of disulfide groups to maleimide reactive thiols, has been used for this purpose. For instance, the use of surface albumin-binding peptide (ABP)-displayed EVs as drug delivery systems could enhanced renal retention and therapeutic efficacy of ABP-EVs loaded with small molecule drugs or proteins, suggesting a promising strategy for engineering EVs as versatile therapeutics for kidney diseases.66 Ligand-conjugated EVs, such as transferrin-conjugated milk-derived EVs, have demonstrated enhanced cell targeting and cargo delivery capabilities, with in vivo imaging confirming their potential for tumor enrichment.67
Targeting Strategies for EVs in CKD
Targeted strategies employing EVs in the context of CKD represent a promising direction in the field of renal therapeutics. Mesenchymal stem cells (MSCs), with their well-documented regenerative properties, have been a focal point of interest. Paracrine functions of MSCs involve the secretion of soluble factors and the release of EVs, such as exosomes and microvesicles. Recent revelations suggest that, in numerous animal disease models, the therapeutic effects of MSCs are primarily mediated by the EVs they release.68 This realization opens new avenues for the use of MSC-derived extracellular vesicles as a regenerative medicine therapy, potentially circumventing some of the challenges associated with whole-cell therapies.69 These findings underscore the therapeutic potential of EVs in various disease contexts and their promise as a novel frontier in regenerative medicine. Prior studies highlighted innovative approaches centered around EVs derived from kidney mesenchymal stem cells (KMSCs) that have been genetically engineered to express erythropoietin (EPO), a hormone crucial for stimulating red blood cell production.70 This innovation emerges as a potential solution to address renal anemia, a common and challenging complication associated with CKD. By modifying EV-producing cells genetically to express EPO, researchers have harnessed the power of these small vesicles as targeted delivery vehicles for therapeutic cargo. The fundamental concept underlying this approach is that EPO-expressing EVs, or EPO(+)-EVs, can be effectively administered to CKD mice via intraperitoneal injection.
Loading and Delivery of Therapeutic Cargo into EVs
The loading and delivery of therapeutic cargo into EVs is a pivotal aspect of harnessing EVs as a platform for targeted therapeutic interventions, particularly in the context of CKD. In CKD, the cargo might encompass molecules such as proteins, nucleic acids (eg, mRNA, microRNAs), or small molecules designed to address renal anemia, fibrosis, inflammation, or other CKD-related concerns.71 Once the cargo is chosen, the next phase involves loading it into EVs, which can be accomplished through a variety of techniques. One common method is electroporation, where an electric field is employed to create temporary pores in the EV membrane, allowing cargo molecules to permeate into the vesicles (Figure 3).72 Alternatively, the cargo can be introduced during the natural biogenesis of EVs, ensuring encapsulation within the vesicles as they form.44 Rigorous quality control measures are essential at this stage, involving confirmation of the successful loading of cargo into EVs and an evaluation of the stability of the cargo within the vesicles. The viability and integrity of the cargo-loaded EVs are fundamental to the efficacy of the therapeutic approach.
 |
Figure 3 Strategies for modifying endogenous EVs for therapeutic cargo incorporation. |
Following successful cargo loading, the subsequent step involves the isolation and purification of these cargo-loaded EVs. Multiple techniques are available for this purpose, including ultracentrifugation, size-exclusion chromatography, or commercially available EV isolation kits.73,74 The objective is to effectively separate the cargo-loaded EVs from unmodified EVs and other cellular debris, ensuring the purity of the cargo carriers. Additionally, comprehensive characterization of the cargo-loaded EVs is paramount, encompassing confirmation of cargo presence within the vesicles, assessment of vesicle size, concentration, and surface markers, which is pivotal for comprehending the properties of the engineered EVs and ascertaining their suitability for therapeutic use.75
The ultimate goal of this intricate process is to deliver the cargo-loaded EVs to the target cells or tissues, which, in the context of CKD, would typically be the kidneys. The route of administration is determined by the therapeutic strategy. For CKD, the use of intraperitoneal, intravenous, local, or inhalation routes may be considered, each offering distinct advantages depending on the intended therapeutic effects.76 Once the cargo-loaded EVs successfully reach their target cells, they release their therapeutic cargo, setting in motion a cascade of events that can influence gene expression, modulate cellular functions, or interact with specific cellular pathways to achieve the desired therapeutic effects. Continuous monitoring and assessment of the therapeutic response are critical. This typically involves the measurement of relevant clinical parameters such as hemoglobin levels, renal function indicators, and other disease-specific markers. These assessments are essential for determining the effectiveness of the cargo-loaded EVs in achieving the intended therapeutic outcomes.77
Therapeutic Applications of Engineered EVs in CKD
Anti-Inflammatory and Immunomodulatory Effects of Engineered EVs
Engineered EVs hold immense promise as a therapeutic approach for CKD by virtue of their potent anti-inflammatory and immunomodulatory effects. Engineered EVs have been developed to enhance their stability and targeted delivery of therapeutic molecules, and they play a pivotal role in CKD therapy by addressing multiple aspects of the disease pathology. One of the key contributions of engineered EVs is their ability to reduce inflammation within the kidney. By delivering anti-inflammatory agents, such as interleukin-10 (IL-10), these EVs can effectively dampen the inflammatory response, which is a hallmark of CKD.78 Furthermore, incorporate tumour necrosis factor-α (TNF-α)/interferon-γ (IFN-γ) into MSCs-EVs could significantly improve kidney tissue regeneration and function in mice by suppressing inflammation and enhancing cellular proliferation, suggesting their potent therapeutic potential for CKD (Figure 4).79 Besides, engineered EVs not only exhibit promise in mitigating inflammation and promoting tissue regeneration within the kidney but also demonstrate efficacy in addressing specific pathological mechanisms underlying renal disorders, such as kidney stone formation. For instance, ADSC-derived miR-20b-3p-incorporated EVs demonstrate protective effects against kidney stone formation by mitigating oxalate-induced cell autophagy and inflammatory responses, as evidenced by decreased miR-20b-3p levels in hyperoxaluria-induced models and the inhibition of ATG7 and TLR4 pathways.80
 |
Figure 4 The therapeutic potential of TNF-α/interferon-γ (IFN-γ)-primed MSC-derived extracellular vesicles (TI-EVs) in chronic kidney disease involves inflammation suppression. (A) A schematic representation illustrating the utilization of TI-EVs within a bioactive scaffold for kidney tissue regeneration, focusing on inflammation and fibrosis suppression. (B) Morphological, size, and surface marker characteristics of TI-EVs demonstrated. Scale bars indicate 100 nm. (C) Quantitative analysis comparing CD9, CD63, and CD81 expression levels in TI-EVs and unconditioned MSC-derived EVs (UC-EVs). (D) Confocal immunofluorescence images displaying Arg1 expression in LPS/IFN-γ-stimulated RAW264.7 cells post-treatment with PDRN and/or TI-EVs. (E). Immunofluorescence images depicting CD206 expression two weeks after scaffold implantation. The arrows indicate the different components were incorporated into scaffold. Reprinted with permission from Ko KW, Park SY, Lee EH, Yoo YI, Kim DS. Integrated bioactive scaffold with polydeoxyribonucleotide and stem-cell-derived extracellular vesicles for kidney regeneration. ACS Nano. 2021;15(4):7575–7585. Copyright 2021 American Chemical Society.79 |
In addition to their anti-inflammatory properties, engineered EVs have demonstrated a remarkable ability to modulate immune microenvironment during CDK progression. These engineered EVs, through their unique cargo of bioactive molecules, can effectively regulate various aspects of immune responses within the kidney tissue.81 By modulating specific immune cells, including macrophages, T cells, and dendritic cells, engineered EVs can attenuate inflammatory processes, promote tissue repair, and regulate immune cell function. This modulation of the immune microenvironment holds significant therapeutic potential for mitigating CKD progression and improving renal function in affected individuals. In a previous study, Guo et al explored how epidermal growth factor receptor (EGFR) mimotope immunization regulates the immune microenvironment in CKD, demonstrating that it inhibits macrophage activation through tubular epithelial cell (TEC)-derived exosomes, consequently ameliorating renal fibrosis in a unilateral ureteric obstruction (UUO) model.82 In a related investigation, researchers discovered that the distinctive composition of EVs derived from MSCs in lean as opposed to those in metabolic syndrome (MetS) pigs, suggesting that Lean-EVs enriched with anti-inflammatory TGF-β, effectively induce regulatory T cells and alleviate renal injury in pigs with MetS and renal artery stenosis, highlighting the potential of modulating EV cargo to regulate the immune microenvironment for renal repair.83
In summary, the multifaceted therapeutic potential of engineered EVs, spanning from their anti-inflammatory and immunomodulatory effects to their ability to target specific pathological mechanisms like kidney stone formation, underscores their promising role in the comprehensive management of chronic kidney disease.
Renoprotective Effects and Regeneration of Damaged Tissue of Engineered EVs
Prior studies reported the promising potential of EVs in conferring renoprotective effects and stimulating the regeneration of damaged renal tissue, particularly in the context of CKD.84 Several aspects emerge as key themes in this body of research. First, EVs, especially engineered MSC-EVs, have demonstrated significant renoprotective properties.85 In a previous study, MSCs engineered to overexpress miRNA-let7c (miR-let7c-MSCs) exhibit renoprotective effects by selectively homing to injured kidneys, upregulating miR-let7c expression, attenuating kidney injury, downregulating fibrotic genes, and inhibiting TGF-β1-driven fibrosis in vitro, suggesting their potential as a therapeutic approach for CKD.86 In another study, MSCs-derived EVs engineered with anti-let-7i-5p oligonucleotides effectively reduce renal fibrosis by inhibiting the let-7i-5p level in kidney tubular epithelial cells, suppressing extracellular matrix deposition and epithelial-mesenchymal transition, ultimately improving kidney function through activation of the TSC1/mTOR signaling pathway, suggesting a promising therapeutic approach for chronic kidney disease-associated renal fibrosis.87
CKD often involves chronic and progressive damage to kidney tissue, and these engineered EVs appear to mitigate this damage effectively. They do so by alleviating the inflammatory responses that contribute to tissue injury and by reducing fibrosis, a key hallmark of CKD progression. For instance, Liu et al reported that TGF-β1 stimulated-EVs exert renoprotective effects by preventing renal interstitial fibroblast apoptosis and promoting fibroblast survival and proliferation through the transfer of TNFAIP8, highlighting their crucial role in renal fibrogenesis and potential as therapeutic targets for fibrotic chronic kidney disease.88 Additionally, there is a growing emphasis on the development of targeted therapies using EVs. These therapies involve engineering EVs to specifically home in on injured kidney tissue, which not only enhances their therapeutic efficacy but also has implications for personalized treatment strategies in CKD management.89 For instance, Ji et al introduced a new approach for treating CKD by using superparamagnetic iron oxide nanoparticles (SPION)-decorated MSCs-derived EVs with high expression of carboxyl terminus of Hsc70-interacting protein (CHIP), which selectively target injured renal sites under a magnetic field and deliver CHIP to alleviate fibrosis, offering a promising nanoplatform for CKD therapy (Figure 5).90
 |
Figure 5 MSC-EVs-CHIP alleviate fibrosis through CHIP delivery. (A) TEM images: MSC-EVs and MSC-EVs-CHIP. Scale: 100 nm. Size measured by DLS. (B) Immunofluorescence: α-SMA (Red), Slc5a1 (Green), DAPI (Blue) (Upper). Fibronectin via immunohistochemistry (Middle). HE staining for renal inflammation. CD66 (Red) for inflammatory infiltration. DAPI (Blue) for nuclei. Scale: 100 µm (Below). (C) Renal tissue images: UUO rats (n = 6/group), treatments: sham, PBS, MSC-EVs (10 mg/kg), MSC-EVs-CHIP (10 mg/kg). (D) Sirius red staining: n = 3, higher magnification. Scale: 1 mm (top), 100 µm (bottom). (E) Masson trichrome staining: renal tissue sections, n = 3. Scale: 100 µm. Reprinted with permission from Ji C, Zhang J, Shi L, et al. Engineered extracellular vesicle-encapsulated CHIP as novel nanotherapeutics for treatment of renal fibrosis. NPJ Regen Med. 2024;9(1):3. Under Creative Commons CC BY License.90 |
In summary, engineered EVs have renoprotective effects by reducing renal fibrosis, and offering targeted treatment strategies. These findings suggest a potential paradigm shift in CKD management towards personalized and effective therapeutic interventions utilizing EV-based approaches.
Targeting Fibrosis and Oxidative Stress in CKD
CKD is characterized by a complex interplay of pathological processes, including fibrosis and oxidative stress, which can lead to progressive kidney damage. Prior studies have explored innovative approaches to harness the regenerative and anti-fibrosis properties of EVs to combat these key factors in CKD progression.
Several studies have investigated the potential of EVs derived from various sources, such as MSCs and peritoneal M2 macrophages, as well as EVs encapsulated in specialized matrices like collagen or self-assembling peptides for CKD therapy.91 They can be designed to address specific aspects of the disease, such as hypoxia, which is often associated with kidney damage. By making these EVs hypoxia-sensitive, researchers have created a means of precise imaging and treatment for hypoxic kidney regions, which is a significant step towards targeted intervention in CKD. In a previous study, Cheng et al developed a nano-sized hypoxia-sensitive coassembly (Pc/C5A@EVs) composed of sulfonated aluminum phthalocyanine (Pc), the macrocyclic amphiphile C5A, and MSC-EVs, which specifically targeted hypoxic kidneys, allowing enhanced hypoxia-sensitive imaging and regenerative effects by inducing M1-to-M2 macrophage transition and inhibiting HIF-1α expression and NF-κB signaling pathway, offering a promising strategy for precise kidney hypoxia diagnosis and efficient kidney injury treatment (Figure 6).92
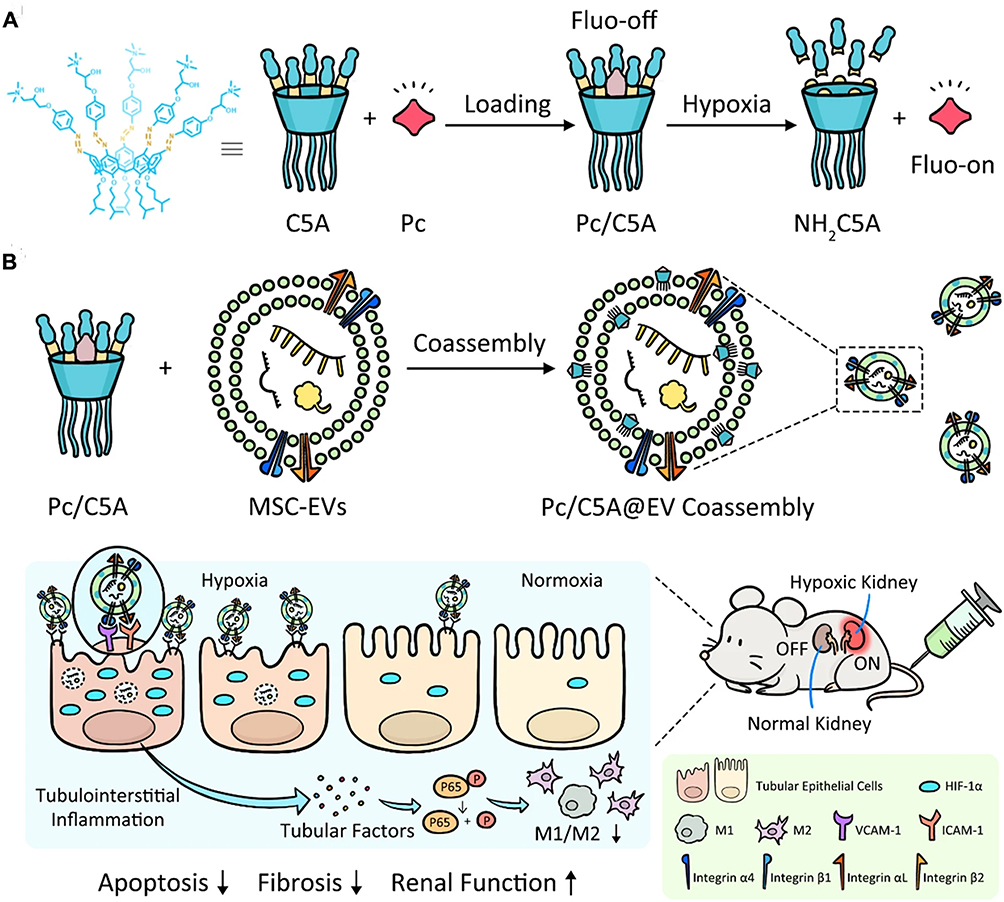 |
Figure 6 Diagram demonstrating the preparation process of Pc/C5A@EV (A) and its application in conducting simultaneous hypoxia-sensitive imaging and therapeutic intervention in injured kidneys (B). The arrows represent the next synthesis process of Pc/C5A@EV. Reprinted with permission from Cheng YQ, Yue YX, Cao HM, et al. Coassembly of hypoxia-sensitive macrocyclic amphiphiles and extracellular vesicles for targeted kidney injury imaging and therapy. J Nanobiotechnol. 2021;19(1):451. Under Creative Commons CC BY License.92 |
Notable, accumulative interests have been attracted to the potential of engineered EVs to mitigate fibrosis and oxidative stress in CKD. One such mechanism is the regulation of cellular viability. Engineered EVs have been shown to enhance cell growth, regeneration and differentiation, which can counteract the tissue damage caused by fibrosis and oxidative stress. In a previous study, researchers devised an anticalcification strategy involving engineered exosome delivery of lncRNA-ANCR to Gli1+ cells (progenitors of smooth muscle cells), effectively inhibiting their differentiation into osteoblast-like cells and thereby preventing calcification and autologous pathological vessels, offering a promising noninvasive treatment approach for vascular calcification of CKD.93 In addition, engineered EVs have been found to inhibit apoptosis, a process of programmed cell death that contributes to kidney damage in CKD. By preventing or reducing apoptosis in renal cells, these EVs can help preserve and repair kidney tissue. For instance, the engineered melatonin-stimulated MSCs-derived EVs shown suppressed apoptosis resident kidney cells in kidney tissues, alleviation of CKD severity, and functional kidney recovery, suggesting it could be served as a promising therapeutic agent for CKD treatment through regulation of apoptosis.94
Engineered EVs also play a role in angiogenesis, the formation of new blood vessels. This process is crucial for ensuring adequate blood supply to the kidneys, and the studies suggest that these EVs are effective in promoting angiogenesis. This, in turn, can help improve renal function by ensuring that the kidney tissue receives the necessary oxygen and nutrients. For instance, Chen et al demonstrated that neurotrophic factor-loaded EVs significantly mitigate peritubular capillary (PTC) loss in tubulointerstitial fibrosis through activation of the Sirtuin 1 (SIRT1)/endothelial nitric oxide synthase (eNOS) signaling pathway, enhancing angiogenesis and protecting endothelial cells against injury in both in vivo and in vitro models, suggesting a potential therapeutic mechanism for renal fibrosis.95
Taken together, the comprehensive approach of engineered EVs in CKD therapy is based on their ability to target multiple aspects of the disease pathology. They reduce inflammation, improve mitochondrial function, promote cellular repair, and address renal anemia. Moreover, their stability and targeted delivery make them a promising platform for the precise administration of therapeutic molecules, further enhancing their therapeutic impact. While more research is needed to refine the application of engineered EVs in CKD therapy and ensure their safety and efficacy, the current findings offer exciting possibilities for innovative and effective treatments for this challenging condition.
Challenges and Perspectives
Translating engineered EV therapies from promising laboratory experiments to effective clinical applications faces a spectrum of challenges and limitations. A fundamental challenge is achieving standardization, as it is imperative to establish uniform procedures for the isolation, characterization, and production of EVs.96 This standardization is essential to guarantee the reproducibility and dependability of outcomes, which are critical factors for regulatory endorsement and clinical safety. However, it is crucial to acknowledge the limitations of translating findings from animal models to human diseases, particularly in the context of CKD. Animal models often differ significantly from human diseases in their pathophysiology, and these differences can pose challenges in extrapolating preclinical results to clinical settings.97 Additionally, differences between study protocols among research groups can lead to difficulties in comparing results and establishing effective treatment strategies. Moreover, the lack of standardization in the production of EVs further complicates the clinical translation process. Variability in isolation methods, cell sources, and culture conditions can result in variations in EVs composition and therapeutic efficacy, emphasizing the need for standardized protocols in EV production.98
Another substantial challenge is navigating the intricate regulatory landscape, representing a significant obstacle to the development of EV therapies.99 EV-based treatments are required to meet stringent safety and efficacy standards akin to other biologics, necessitating comprehensive documentation and rigorous adherence to continuously evolving regulatory frameworks. Additionally, the shift from laboratory-scale to large-scale manufacturing is a complex task.100 Scalable production methods must be developed to meet the demands of clinical trials and potential widespread use. Determining the optimal dosing and delivery methods for EV therapies also remains a challenge, requiring in-depth research to ascertain how to administer EVs effectively to target tissues while avoiding unintended off-target effects.101 Moreover, ensuring the long-term safety of EV therapies demands comprehensive monitoring for potential side effects and immune responses, especially in the context of chronic diseases like CKD.
However, amidst these challenges, there are promising prospects and emerging trends in the field of engineered EV therapies. Advanced characterization techniques, such as single-vesicle analysis and RNA sequencing, are revolutionizing our understanding of EV composition and function, paving the way for personalized medicine.102 The concept of tailoring EV therapies to individual patients based on their unique genetic makeup holds immense potential, potentially transforming treatment approaches and enhancing clinical outcomes. Additionally, the integration of EV therapies with other cutting-edge technologies, including gene editing and CRISPR technology, presents exciting possibilities. These combination therapies could synergistically enhance the therapeutic potential of EVs, offering multifaceted solutions to complex diseases.103 The development of reliable biomarkers specific to EV-based treatments is crucial, enabling accurate monitoring of treatment responses and prediction of outcomes.
In conclusion, while challenges persist in the translation of engineered EV therapies, the field is advancing at a remarkable pace. With ongoing efforts in standardization, advanced characterization, personalized medicine, combination therapies, and biomarker development, the prospects for EV-based treatments are increasingly promising. Collaborative research initiatives and rigorous clinical trials are pivotal in determining the feasibility and effectiveness of EV therapies, holding the key to transforming the landscape of medical treatments for chronic kidney disease and various other diseases. As research continues to unfold and innovative strategies emerge, engineered EV therapies are poised to revolutionize patient care, offering targeted, effective, and personalized solutions to complex health challenges.
Acknowledgments
We deeply appreciate the support from all participants.
Disclosure
The authors declare that they have no competing interests.
References
1. Glassock RJ, Warnock DG, Delanaye P. The global burden of chronic kidney disease: estimates, variability and pitfalls. Nat Rev Nephrol. 2017;13(2):104–114. doi:10.1038/nrneph.2016.163
2. de Boer IH, Khunti K, Sadusky T, et al. Diabetes management in chronic kidney disease: a consensus report by the American Diabetes Association (ADA) and Kidney Disease: improving Global Outcomes (KDIGO). Kidney Int. 2022;102(5):974–989. doi:10.1016/j.kint.2022.08.012
3. Martin P, Awan AA, Berenguer MC, et al. Executive Summary of the KDIGO 2022 clinical practice guideline for the prevention, diagnosis, evaluation, and treatment of hepatitis c in chronic kidney disease. Kidney Int. 2022;102(6):1228–1237. doi:10.1016/j.kint.2022.07.012
4. Noels H, Jankowski J. Increased risk of cardiovascular complications in chronic kidney disease: introduction to a compendium. Circ Res. 2023;132(8):899–901. doi:10.1161/CIRCRESAHA.123.322806
5. Zhou S, Cheing GLY, Cheung AKK. Role of exosomes and exosomal microRNA in muscle-Kidney crosstalk in chronic kidney disease. Front Cell Dev Biol. 2022;10:10 951837. doi:10.3389/fcell.2022.951837
6. Li Q, Zhang C, Shi J, et al. High-Phosphate-stimulated macrophage-derived exosomes promote vascular calcification via let-7b-5p/TGFBR1 axis in chronic kidney disease. Cells. 2022;12(1):161. doi:10.3390/cells12010161
7. Gudehithlu KP, Hart P, Joshi A, et al. Urine exosomal ceruloplasmin: a potential early biomarker of underlying kidney disease. Clin Exp Nephrol. 2019;23(8):1013–1021. doi:10.1007/s10157-019-01734-5
8. Tang TT, Wang B, Li ZL, et al. Kim-1 targeted extracellular vesicles: a new therapeutic platform for RNAi to Treat AKI. J Am Soc Nephrol. 2021;32(10):2467–2483. doi:10.1681/ASN.2020111561
9. Yoon YM, Lee JH, Song KH, Noh H, Lee SH. Melatonin-stimulated exosomes enhance the regenerative potential of chronic kidney disease-derived mesenchymal stem/stromal cells via cellular prion proteins. J Pineal Res. 2020;68(3):e12632. doi:10.1111/jpi.12632
10. Ceccotti E, Saccu G, Herrera Sanchez MB, Bruno S. Naive or engineered extracellular vesicles from different cell sources: therapeutic tools for kidney diseases. Pharmaceutics. 2023;15(6):1715. doi:10.3390/pharmaceutics15061715
11. Grange C, Skovronova R, Marabese F, Bussolati B. Stem Cell-derived extracellular vesicles and kidney regeneration. Cells. 2019;8(10):1240. doi:10.3390/cells8101240
12. Yang B, Lin Y, Huang Y, Zhu N, Shen YQ. Extracellular vesicles modulate key signalling pathways in refractory wound healing. Burns Trauma. 2023;11:11 tkad039. doi:10.1093/burnst/tkad039
13. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
14. Madkhali AM, Mobarki AA, Ghzwani AH, et al. Elevated levels of procoagulant microvesicles and tissue-factor bearing microvesicles in malaria patients. Int J Gen Med. 2023;16:1205–1215. doi:10.2147/IJGM.S402212
15. Yang J, Zhang X, Wang G, et al. ApoSEVs-mediated modulation of versatile target cells promotes diabetic wound healing: unveiling a promising strategy. Int J Nanomed. 2023;18:6955–6977. doi:10.2147/IJN.S436350
16. Hu Z, Chen G, Zhao Y, et al. Exosome-derived circCCAR1 promotes CD8 + T-cell dysfunction and anti-PD1 resistance in hepatocellular carcinoma. Mol Cancer. 2023;22(1):55. doi:10.1186/s12943-023-01759-1
17. Hu Y, Tao R, Chen L, et al. Exosomes derived from pioglitazone-pretreated MSCs accelerate diabetic wound healing through enhancing angiogenesis. J Nanobiotechnol. 2021;19(1):150. doi:10.1186/s12951-021-00894-5
18. Zhao M, Liu S, Wang C, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate mitochondrial damage and inflammation by stabilizing mitochondrial DNA. ACS Nano. 2021;15(1):1519–1538. doi:10.1021/acsnano.0c08947
19. Zhou M, Li YJ, Tang YC, et al. Apoptotic bodies for advanced drug delivery and therapy. J Control Release. 2022;351:394–406.
20. Liu H, Liu S, Qiu X, et al. Donor MSCs release apoptotic bodies to improve myocardial infarction via autophagy regulation in recipient cells. Autophagy. 2020;16(12):2140–2155. doi:10.1080/15548627.2020.1717128
21. Liao Y, Zhang Z, Ouyang L, Mi B, Liu G. Engineered extracellular vesicles in wound healing: design, paradigms, and clinical application. Small. 2023;20(7):e2307058. doi:10.1002/smll.202307058
22. Zhang Y, Tang Y, Du S, Li P, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;15:6917–6934. doi:10.2147/IJN.S264498
23. Buzas EI. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23(4):236–250. doi:10.1038/s41577-022-00763-8
24. Mi B, Chen L, Xiong Y, et al. Saliva exosomes-derived UBE2O mRNA promotes angiogenesis in cutaneous wounds by targeting SMAD6. J Nanobiotechnol. 2020;18(1):68. doi:10.1186/s12951-020-00624-3
25. Ferreira JV, da Rosa Soares A, Ramalho J, et al. LAMP2A regulates the loading of proteins into exosomes. Sci Adv. 2022;8(12):eabm1140. doi:10.1126/sciadv.abm1140
26. Chen C, Zheng H, Luo Y, et al. SUMOylation promotes extracellular vesicle-mediated transmission of lncRNA ELNAT1 and lymph node metastasis in bladder cancer. J Clin Invest. 2021;131(8). doi:10.1172/JCI146431
27. Lin Z, Xiong Y, Zeng R, et al. Circulating MiRNA-21-enriched extracellular vesicles promote bone remodeling in traumatic brain injury patients. Exp Mol Med. 2023;55(3):587–596. doi:10.1038/s12276-023-00956-8
28. Segel M, Lash B, Song J, et al. Mammalian retrovirus-like protein PEG10 packages its own mRNA and can be pseudotyped for mRNA delivery. Science. 2021;373(6557):882–889. doi:10.1126/science.abg6155
29. Bitto NJ, Cheng L, Johnston EL, et al. Staphylococcus aureus membrane vesicles contain immunostimulatory DNA, RNA and peptidoglycan that activate innate immune receptors and induce autophagy. J Extracell Vesicles. 2021;10(6):e12080. doi:10.1002/jev2.12080
30. Yu Y, Chen M, Guo Q, et al. Human umbilical cord mesenchymal stem cell exosome-derived miR-874-3p targeting RIPK1/PGAM5 attenuates kidney tubular epithelial cell damage. Cell Mol Biol Lett. 2023;28(1):12. doi:10.1186/s11658-023-00425-0
31. Ding H, Li LX, Harris PC, Yang J, Li X. Extracellular vesicles and exosomes generated from cystic renal epithelial cells promote cyst growth in autosomal dominant polycystic kidney disease. Nat Commun. 2021;12(1):4548. doi:10.1038/s41467-021-24799-x
32. Wang H, Wang B, Zhang A, et al. Exosome-mediated miR-29 transfer reduces muscle atrophy and kidney fibrosis in mice. Mol Ther. 2019;27(3):571–583. doi:10.1016/j.ymthe.2019.01.008
33. El Fekih R, Hurley J, Tadigotla V, et al. Discovery and validation of a urinary exosome mRNA signature for the diagnosis of human kidney transplant rejection. J Am Soc Nephrol. 2021;32(4):994–1004. doi:10.1681/ASN.2020060850
34. Erdbrugger U, Blijdorp CJ, Bijnsdorp IV, et al. Urinary extracellular vesicles: a position paper by the urine task force of the international society for extracellular vesicles. J Extracell Vesicles. 2021;10(7):e12093. doi:10.1002/jev2.12093
35. Chen S, Zhang X, Meng K, et al. Urinary exosome tsRNAs as novel markers for diagnosis and prediction of lupus nephritis. Front Immunol. 2023;14:1077645. doi:10.3389/fimmu.2023.1077645
36. Liu Y, Guo Y, Bao S, Huang H, Liu W, Guo W. Bone marrow mesenchymal stem cell-derived exosomal microRNA-381-3p alleviates vascular calcification in chronic kidney disease by targeting NFAT5. Cell Death Dis. 2022;13(3):278. doi:10.1038/s41419-022-04703-1
37. Behrens F, Holle J, Kuebler WM, Simmons S. Extracellular vesicles as regulators of kidney function and disease. Intensive Care Med Exp. 2020;8(Suppl 1):22. doi:10.1186/s40635-020-00306-2
38. Zhou X, Zhao S, Li W, et al. Tubular cell-derived exosomal miR-150-5p contributes to renal fibrosis following unilateral ischemia-reperfusion injury by activating fibroblast in vitro and in vivo. Int J Biol Sci. 2021;17(14):4021–4033. doi:10.7150/ijbs.62478
39. Zhang W, Zheng X, Yu Y, et al. Renal cell carcinoma-derived exosomes deliver lncARSR to induce macrophage polarization and promote tumor progression via STAT3 pathway. Int J Biol Sci. 2022;18(8):3209–3222. doi:10.7150/ijbs.70289
40. Wang B, Zhang A, Wang H, et al. miR-26a limits muscle wasting and cardiac fibrosis through exosome-mediated microRNA transfer in chronic kidney disease. Theranostics. 2019;9(7):1864–1877. doi:10.7150/thno.29579
41. Zhou X, Zhang W, Yao Q, et al. Exosome production and its regulation of EGFR during wound healing in renal tubular cells. Am J Physiol Renal Physiol. 2017;312(6):F963–F70. doi:10.1152/ajprenal.00078.2017
42. Lv LL, Feng Y, Wu WJ, et al. Exosomal CCL2 from tubular epithelial cells is critical for albumin-induced tubulointerstitial inflammation. J Am Soc Nephrol. 2018;29(3):919–935. doi:10.1681/ASN.2017050523
43. Jalal D, Sanford B, Ten Eyck P. Detection of pro angiogenic and inflammatory biomarkers in patients with CKD. Sci Rep. 2021;11(1):8786. doi:10.1038/s41598-021-87710-0
44. Chen S, Zhang M, Li J, et al. beta-catenin-controlled tubular cell-derived exosomes play a key role in fibroblast activation via the OPN-CD44 axis. J Extracell Vesicles. 2022;11(3):e12203. doi:10.1002/jev2.12203
45. Lin X, Shan SK, Xu F, et al. The crosstalk between endothelial cells and vascular smooth muscle cells aggravates high phosphorus-induced arterial calcification. Cell Death Dis. 2022;13(7):650. doi:10.1038/s41419-022-05064-5
46. Zietzer A, Steffen E, Niepmann S, et al. MicroRNA-mediated vascular intercellular communication is altered in chronic kidney disease. Cardiovasc Res. 2022;118(1):316–333. doi:10.1093/cvr/cvaa322
47. Jankowski J, Floege J, Fliser D, Bohm M, Marx N. Cardiovascular disease in chronic kidney disease: pathophysiological insights and therapeutic options. Circulation. 2021;143(11):1157–1172. doi:10.1161/CIRCULATIONAHA.120.050686
48. Cao H, Cheng Y, Gao H, et al. In vivo tracking of mesenchymal stem cell-derived extracellular vesicles improving mitochondrial function in renal ischemia-reperfusion injury. ACS Nano. 2020;14(4):4014–4026. doi:10.1021/acsnano.9b08207
49. Wen J, Ma Z, Livingston MJ, et al. Decreased secretion and profibrotic activity of tubular exosomes in diabetic kidney disease. Am J Physiol Renal Physiol. 2020;319(4):F664–F73. doi:10.1152/ajprenal.00292.2020
50. Li ZL, Lv LL, Tang TT, et al. HIF-1alpha inducing exosomal microRNA-23a expression mediates the cross-talk between tubular epithelial cells and macrophages in tubulointerstitial inflammation. Kidney Int. 2019;95(2):388–404. doi:10.1016/j.kint.2018.09.013
51. Lv LL, Feng Y, Wu M, et al. Exosomal miRNA-19b-3p of tubular epithelial cells promotes M1 macrophage activation in kidney injury. Cell Death Differ. 2020;27(1):210–226. doi:10.1038/s41418-019-0349-y
52. Zhang L, Li S, Cong M, et al. Lemon-derived extracellular vesicle-like nanoparticles block the progression of kidney stones by antagonizing endoplasmic reticulum stress in renal tubular cells. Nano Lett. 2023;23(4):1555–1563. doi:10.1021/acs.nanolett.2c05099
53. Han L, Wang S, Li J, Zhao L, Zhou H. Urinary exosomes from patients with diabetic kidney disease induced podocyte apoptosis via microRNA-145-5p/Srgap2 and the RhoA/ROCK pathway. Exp Mol Pathol. 2023;134 104877. doi:10.1016/j.yexmp.2023.104877
54. Burrello J, Monticone S, Burrello A, et al. Identification of a serum and urine extracellular vesicle signature predicting renal outcome after kidney transplant. Nephrol Dial Transplant. 2023;38(3):764–777. doi:10.1093/ndt/gfac259
55. Haney NM, Kim CJ, Kuczler MD, et al. Novel urinary tract obstruction marker discovery by multi-marker profiling of urinary extracellular vesicles derived from a rat UTO model. Am J Clin Exp Urol. 2023;11(2):136–145.
56. Tao Y, Wei X, Yue Y, et al. Extracellular vesicle-derived AEBP1 mRNA as a novel candidate biomarker for diabetic kidney disease. J Transl Med. 2021;19(1):326. doi:10.1186/s12967-021-03000-3
57. Saejong S, Townamchai N, Somparn P, et al. MicroRNA-21 in plasma exosome, but not from whole plasma, as a biomarker for the severe interstitial fibrosis and tubular atrophy (IF/TA) in post-renal transplantation. Asian Pac J Allergy Immunol. 2022;40(1):94–102. doi:10.12932/AP-101019-0656
58. Kim HY, Kwon S, Um W, et al. Functional extracellular vesicles for regenerative medicine. Small. 2022;18(36):e2106569. doi:10.1002/smll.202106569
59. Yuan X, Li L, Liu H, et al. Strategies for improving adipose-derived stem cells for tissue regeneration. Burns Trauma. 2022;10:10 tkac028. doi:10.1093/burnst/tkac028
60. Lin Y, Yan M, Bai Z, et al. Huc-MSC-derived exosomes modified with the targeting peptide of aHSCs for liver fibrosis therapy. J Nanobiotechnol. 2022;20(1):432. doi:10.1186/s12951-022-01636-x
61. Xiong Y, Chen L, Liu P, et al. All-in-one: multifunctional hydrogel accelerates oxidative diabetic wound healing through timed-release of exosome and fibroblast growth factor. Small. 2022;18(1):e2104229. doi:10.1002/smll.202104229
62. Aslan C, Zolbanin NM, Faraji F, Jafari R. Exosomes for CRISPR-Cas9 delivery: the cutting edge in genome editing. Mol Biotechnol. 2023;1:2.
63. Ilahibaks NF, Kluiver TA, de Jong OG, et al. Extracellular vesicle-mediated delivery of CRISPR/Cas9 ribonucleoprotein complex targeting proprotein convertase subtilisin-kexin type 9 (Pcsk9) in primary mouse hepatocytes. J Extracell Vesicles. 2024;13(1):e12389. doi:10.1002/jev2.12389
64. Osteikoetxea X, Silva A, Lazaro-Ibanez E, et al. Engineered Cas9 extracellular vesicles as a novel gene editing tool. J Extracell Vesicles. 2022;11(5):e12225. doi:10.1002/jev2.12225
65. Salunkhe S, Chitkara D, Mittal A, Chitkara D, Mittal A. Surface functionalization of exosomes for target-specific delivery and in vivo imaging & tracking: strategies and significance. J Control Release. 2020;326:326 599–614. doi:10.1016/j.jconrel.2020.07.042
66. Liu S, Lv K, Wang Y, et al. Improving the circulation time and renal therapeutic potency of extracellular vesicles using an endogenous ligand binding strategy. J Control Release. 2022;352:1009–1023. doi:10.1016/j.jconrel.2022.11.017
67. Chen C, Sun M, Liu X, et al. General and mild modification of food-derived extracellular vesicles for enhanced cell targeting. Nanoscale. 2021;13(5):3061–3069. doi:10.1039/D0NR06309F
68. Deng D, Li X, Zhang JJ, et al. Biotin-avidin system-based delivery enhances the therapeutic performance of MSC-derived exosomes. ACS Nano. 2023;17(9):8530–8550. doi:10.1021/acsnano.3c00839
69. Gao L, Qiu F, Cao H, et al. Therapeutic delivery of microRNA-125a-5p oligonucleotides improves recovery from myocardial ischemia/reperfusion injury in mice and swine. Theranostics. 2023;13(2):685–703. doi:10.7150/thno.73568
70. Choi HY, Kim TY, Lee M, et al. Kidney mesenchymal stem cell-derived extracellular vesicles engineered to express erythropoietin improve renal anemia in mice with chronic kidney disease. Stem Cell Rev Rep. 2022;18(3):980–992. doi:10.1007/s12015-021-10141-x
71. Kim S, Lee SA, Yoon H, et al. Exosome-based delivery of super-repressor IkappaBalpha ameliorates kidney ischemia-reperfusion injury. Kidney Int. 2021;100(3):570–584. doi:10.1016/j.kint.2021.04.039
72. Rong Y, Wang Z, Tang P, et al. Engineered extracellular vesicles for delivery of siRNA promoting targeted repair of traumatic spinal cord injury. Bioact Mater. 2023;23:328–342. doi:10.1016/j.bioactmat.2022.11.011
73. Yuan J, Yang H, Liu C, et al. Microneedle patch loaded with exosomes containing MicroRNA-29b prevents cardiac fibrosis after myocardial infarction. Adv Healthc Mater. 2023;12(13):e2202959. doi:10.1002/adhm.202202959
74. Lai JJ, Chau ZL, Chen SY, et al. Exosome processing and characterization approaches for research and technology development. Adv Sci. 2022;9(15):e2103222. doi:10.1002/advs.202103222
75. Xiong Y, Lin Z, Bu P, et al. A whole-course-repair system based on neurogenesis-angiogenesis crosstalk and macrophage reprogramming promotes diabetic wound healing. Adv Mater. 2023;35(19):e2212300. doi:10.1002/adma.202212300
76. Zheng H, Ji J, Zhao T, Wang E, Zhang A. Exosome‑encapsulated miR‑26a attenuates aldosterone‑induced tubulointerstitial fibrosis by inhibiting the CTGF/SMAD3 signaling pathway. Int J Mol Med. 2023;51:2.
77. Liang M, Zhang D, He W, Jin J. Exosomes from miR-374a-5p-modified mesenchymal stem cells inhibit the progression of renal fibrosis by regulating MAPK6/MK5/YAP axis. Bioengineered. 2022;13(2):4517–4527. doi:10.1080/21655979.2022.2033465
78. Tang TT, Wang B, Wu M, et al. Extracellular vesicle-encapsulated IL-10 as novel nanotherapeutics against ischemic AKI. Sci Adv. 2020;6(33):eaaz0748. doi:10.1126/sciadv.aaz0748
79. Ko KW, Park SY, Lee EH, Yoo YI, Kim DS. Integrated bioactive scaffold with polydeoxyribonucleotide and stem-cell-derived extracellular vesicles for kidney regeneration. ACS Nano. 2021;15(4):7575–7585. doi:10.1021/acsnano.1c01098
80. Shi J, Duan J, Gong H, Pang Y, Wang L, Yan Y. Exosomes from miR-20b-3p-overexpressing stromal cells ameliorate calcium oxalate deposition in rat kidney. J Cell Mol Med. 2019;23(11):7268–7278. doi:10.1111/jcmm.14555
81. Jia Y, Zheng Z, Xue M, et al. Extracellular vesicles from albumin-induced tubular epithelial cells promote the M1 macrophage phenotype by targeting klotho. Mol Ther. 2019;27(8):1452–1466. doi:10.1016/j.ymthe.2019.05.019
82. Guo J, Liu X, Song H, Gu Y, Niu J, Yang L. MHC-1B carried exosomes derived from tubular epithelial cell induced by the EGFR mimotope inhibit macrophage activation in renal fibrosis. Extracel Vesicle. 2023;2:100024. doi:10.1016/j.vesic.2023.100024
83. Song T, Eirin A, Zhu X, et al. Mesenchymal stem cell-derived extracellular vesicles induce regulatory T cells to ameliorate chronic kidney injury. Hypertension. 2020;75(5):1223–1232. doi:10.1161/HYPERTENSIONAHA.119.14546
84. Li Y, Chakraborty A, Broughton BRS, et al. Comparing the renoprotective effects of BM-MSCs versus BM-MSC-exosomes, when combined with an anti-fibrotic drug, in hypertensive mice. Biomed Pharmacother;2021. 144 112256. doi:10.1016/j.biopha.2021.112256
85. Tapparo M, Bruno S, Collino F, et al. Renal regenerative potential of extracellular vesicles derived from miRNA-Engineered mesenchymal stromal cells. Int J Mol Sci. 2019;20:10.
86. Wang B, Yao K, Huuskes BM, et al. Mesenchymal stem cells deliver exogenous MicroRNA-let7c via exosomes to attenuate renal fibrosis. Mol Ther. 2016;24(7):1290–1301. doi:10.1038/mt.2016.90
87. Jin J, Qian F, Zheng D, He W, Gong J, He Q. Mesenchymal stem cells attenuate renal fibrosis via exosomes-mediated delivery of microRNA Let-7i-5p antagomir. Int J Nanomed. 2021;16:3565–3578. doi:10.2147/IJN.S299969
88. Liu X, Liu Z, Wang C, et al. Kidney tubular epithelial cells control interstitial fibroblast fate by releasing TNFAIP8-encapsulated exosomes. Cell Death Dis. 2023;14(10):672. doi:10.1038/s41419-023-06209-w
89. Chen XJ, Jiang K, Ferguson CM, et al. Augmented efficacy of exogenous extracellular vesicles targeted to injured kidneys. Signal Transduct Target Ther. 2020;5(1):199. doi:10.1038/s41392-020-00304-6
90. Ji C, Zhang J, Shi L, et al. Engineered extracellular vesicle-encapsulated CHIP as novel nanotherapeutics for treatment of renal fibrosis. NPJ Regen Med. 2024;9(1):3. doi:10.1038/s41536-024-00348-0
91. Jin C, Wu P, Li L, Xu W, Qian H. Exosomes: emerging therapy delivery tools and biomarkers for kidney diseases. Stem Cells Int. 2021;2021:7844455. doi:10.1155/2021/7844455
92. Cheng YQ, Yue YX, Cao HM, et al. Coassembly of hypoxia-sensitive macrocyclic amphiphiles and extracellular vesicles for targeted kidney injury imaging and therapy. J Nanobiotechnol. 2021;19(1):451. doi:10.1186/s12951-021-01192-w
93. Yan J, Xiao H, Zhou X, et al. Engineered exosomes reprogram Gli1(+) cells in vivo to prevent calcification of vascular grafts and autologous pathological vessels. Sci Adv. 2023;9(29):eadf7858. doi:10.1126/sciadv.adf7858
94. Yea JH, Yoon YM, Lee JH, Yun CW, Lee SH. Exosomes isolated from melatonin-stimulated mesenchymal stem cells improve kidney function by regulating inflammation and fibrosis in a chronic kidney disease mouse model. J Tissue Eng. 2021;12:12 20417314211059624. doi:10.1177/20417314211059624
95. Chen L, Wang Y, Li S, et al. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics. 2020;10(20):9425–9442. doi:10.7150/thno.43315
96. Huang W, Zhu XY, Lerman A, Lerman LO. Extracellular vesicles as theranostic tools in kidney disease. Clin J Am Soc Nephrol. 2022;17(9):1418–1429. doi:10.2215/CJN.16751221
97. Kishi S, Matsumoto T, Brooks CR, Brooks CR. Human reconstructed kidney models. Vitro Cell Dev Biol Anim. 2021;57(2):133–147. doi:10.1007/s11626-021-00548-8
98. Tian Y, Gong M, Hu Y, et al. Quality and efficiency assessment of six extracellular vesicle isolation methods by nano-flow cytometry. J Extracell Vesicles. 2020;9(1):1697028. doi:10.1080/20013078.2019.1697028
99. Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023;33(8):667–681. doi:10.1016/j.tcb.2023.01.002
100. Verweij FJ, Balaj L, Boulanger CM, et al. The power of imaging to understand extracellular vesicle biology in vivo. Nat Methods. 2021;18(9):1013–1026. doi:10.1038/s41592-021-01206-3
101. Chen J, Tan Q, Yang Z, Jin Y. Engineered extracellular vesicles: potentials in cancer combination therapy. J Nanobiotechnol. 2022;20(1):132. doi:10.1186/s12951-022-01330-y
102. Zhang J, Wu J, Wang G, et al. Extracellular vesicles: techniques and biomedical applications related to single vesicle analysis. ACS Nano. 2023;17(18):17668–17698. doi:10.1021/acsnano.3c03172
103. Whitley JA, Cai H. Engineering extracellular vesicles to deliver CRISPR ribonucleoprotein for gene editing. J Extracell Vesicles. 2023;12(9):e12343. doi:10.1002/jev2.12343
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.