Back to Journals » Infection and Drug Resistance » Volume 13
Emergence of vanA-Type Vancomycin-Resistant Enterococcus faecium ST 78 Strain with a rep2-Type Plasmid Carrying a Tn1546-Like Element Isolated from a Urinary Tract Infection in China
Authors Sun L, Xu J, Wang W, He F ![]()
Received 29 January 2020
Accepted for publication 25 March 2020
Published 1 April 2020 Volume 2020:13 Pages 949—955
DOI https://doi.org/10.2147/IDR.S247569
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Long Sun,1,* Juan Xu,2,* Weizhong Wang,3 Fang He3
1Department of Clinical Laboratory, Hangzhou Women’s Hospital, Hangzhou Maternity and Child Health Care Hospital, Hangzhou, Zhejiang 310008, People’s Republic of China; 2Institute of Hygiene, Zhejiang Academy of Medical Sciences, Hangzhou Medical College, Hangzhou, Zhejiang 310013, People’s Republic of China; 3Department of Clinical Laboratory, Zhejiang Provincial People’s Hospital, People’s Hospital of Hangzhou Medical College, Hangzhou, Zhejiang 310014, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fang He Tel/Fax +86-571-8821-5596
Email [email protected]
Purpose: The emergence of vancomycin-resistant enterococci (VRE) dramatically narrows therapeutic options. Although the prevalence of VRE in China has maintained a low level, VRE outbreaks have been reported in some tertiary hospitals in the developed areas of China. The clonal background of vanA-positive Enterococcus faecium strains has not been well characterized in China. Here, we report the whole-genome sequence of a vanA-type vancomycin-resistant E. faecium belonging to sequence type (ST) 78 isolated from a urinary tract infection in China.
Patients and Methods: A vancomycin-resistant E. faecium was isolated from a 66-year-old male patient diagnosed with brainstem hemorrhage. Antibiotic susceptibility assays were performed according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI). Complete genome sequencing was performed using both the HiSeqTM 4000 platform and the MinION platform. Plasmid, genomic and phylogenetic relationship analysis were further performed.
Results: E. faecium VRE1 was resistant to all antimicrobials tested except for tetracyclines and oxazolidinones. The whole genome of E. faecium VRE1 was composed of one chromosomal DNA and four plasmids. Two virulence genes and five antimicrobial resistance genes were identified. In silico multilocus sequence typing (MLST) showed that it belonged to ST78 (clonal complex CC17), a well-known epidemic clone that is widespread in Europe and the United States. Three antimicrobial resistance genes, including aminoglycoside resistance genes ant(6)-Ia and aph(3ʹ)-III; and glycopeptide resistance gene vanA, were located on a rep2-type plasmid carrying a Tn 1546-like element that has not been reported. The most closely related strain harboring a similar plasmid backbone was recovered from fodder sample in China that differed by 178 cgMLST loci.
Conclusion: Our study characterizes the genomic feature of a vancomycin-resistant E. faecium ST78 strain harboring a vanA-carrying plasmid in China. The ST78 clonal group possessed the potential to emerge as a successful vanA-carrying epidemic lineage in China.
Keywords: Enterococcus faecium, vancomycin-resistant enterococci, vanA, whole-genome sequencing, Tn 1546-like element
Introduction
Enterococcus faecium has emerged as a leading cause of multidrug-resistant infections, such as bacteremia, intra-abdominal infections and urinary tract infections. Vancomycin is a glycopeptide antibiotic that plays an antibacterial role by combining with the peptidoglycan precursor and blocking synthesis of the cell wall. It is the first-line drug for the treatment of Gram-positive bacteria, including methicillin-resistant staphylococcus aureus (MRSA) and multidrug-resistant enterococci. Since the first report of vancomycin-resistant enterococci (VRE) in 1988,1 it has been increasingly reported all over the world, representing a global public health concern.2 Resistance to vancomycin dramatically narrows the therapeutic options in E. faecium infections.
The prevalence of VRE shows significant regional differences. In Europe, VRE are prevalent in the community, and the use of glycopeptide antibiotics (avoparcin) in agriculture is thought to be one of the major factors contributing to the further dissemination of VRE.3 In the United States, because of the increasing clinical use of vancomycin, VRE isolated from hospitals has gradually increased and has become one of the most prevalent nosocomial pathogens.4,5 Compared with Europe and the USA, China has maintained a low prevalence of VRE, with an isolation rate of 1.4% for E. faecium and 0.1% for Enterococcus faecalis in the CHINET surveillance in 2017.6
A total of eight acquired glycopeptide resistance determinants have been reported in enterococci (VanA, VanB, VanD, VanE, VanG, VanL, VanM and VanN), of which VanA is the most frequently encountered worldwide.7 The vanA gene is traditionally associated with a plastic antimicrobial resistance transposon, Tn1546, which also contains the additional genes responsible for vancomycin resistance (vanR, vanS, vanH, vanX, vanY and vanZ). Tn1546 is able to easily transpose into diverse conjugative plasmids, and the horizontal transfer of Tn1546-like elements also plays an important role in the dissemination of vanA-type VRE.8–10
Attributable to the narrow antibiotic spectrum of vancomycin and its highest limit level in the classification management system of antibiotics, VRE are still rare in China. However, the prevalence of VRE in some tertiary hospitals in Shanghai and Beijing has increased, and VRE outbreaks have been reported sporadically in China.11 In this study, a vanA-carrying E. faecium strain was isolated from a male patient hospitalized in a tertiary hospital in Hangzhou, China. The whole genome of the strain was sequenced, and a vanA-harboring plasmid was also analyzed to elucidate its genomic epidemiological characteristics.
Patients and Methods
Patient and Bacterial Isolate
A 66-year-old male patient diagnosed with brainstem hemorrhage was long-term hospitalized in the Department of Rehabilitation Medicine in a tertiary hospital in Hangzhou, Zhejiang Province, China. During his hospitalization, the patient received multiple antimicrobial treatments, including teicoplanin, ceftazidime, sulperazone and meropenem. A vancomycin-resistant E. faecium was isolated from his urine sample on December 3, 2018. The strain was preliminarily identified using the VITEK MS system (bioMérieux, France) and was further confirmed by 16S rRNA gene sequencing.
Antimicrobial Susceptibility Testing
Antibiotic susceptibility assays were performed according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI). In total, ten categories including sixteen antibiotics were tested. They were penicillins (penicillin and ampicillin), aminoglycosides (gentamicin and streptomycin), quinolones (ciprofloxacin, levofloxacin and moxifloxacin), macrolides (erythromycin), lincosamides (clindamycin), tetracyclines (tetracycline and tigecycline), glycopeptides (vancomycin and teicoplanin), nitrofurantoins (nitrofurantoin), lipopeptides (daptomycin) and oxazolidinones (linezolid). The minimal inhibitory concentrations (MICs) of penicillin, vancomycin, teicoplanin, tetracycline and tigecycline were determined using the Etest method. The minimum inhibitory concentrations (MICs) of gentamicin, streptomycin and daptomycin were determined using standard broth microdilution tests. The MICs of other antimicrobial agents were determined using a VITEK 2 system (bioMérieux, France) with Gram-positive antimicrobial susceptibility testing cards (AST-GP67). Antimicrobial susceptibility was determined using the breakpoints approved by the CLSI.12
Genomic DNA Extraction and Whole-Genome Sequencing
Genomic DNA of E. faecium VRE1 was extracted using a QIAamp DNA MiniKit (Qiagen, USA) and then subjected to whole-genome sequencing. Whole-genome sequencing was performed using both the HiSeqTM 4000 platform (Illumina, San Diego, CA, USA), with a 150 bp paired-end protocol, and the MinION (Nanopore, Oxford, UK) platform. The short reads generated by the HiSeqTM 4000 were de novo assembled into contigs using SPAdes. Long Nanopore reads were generated by a MinION Sequencer. Hybrid assembly of both short Illumina reads and long MinION reads was performed using Unicycler v 0.4.7 under conservative mode.13 Complete circular contigs generated were then corrected using Pilon v 1.23 with Illumina reads for several rounds. Then, the whole-genome sequence of E. faecium VRE1 was generated. We obtained five circular contigs, which were represented by a complete chromosome and four plasmids. The whole-genome sequence was annotated by the NCBI Prokaryotic Genomes Annotation Pipeline (PGAP) server.
Identification of Antimicrobial Resistance Genes and Virulence Genes
Acquired antimicrobial resistance genes were identified using the ResFinder 2.1 server by uploading the entire genome sequence to the database. Virulence genes were analyzed using VirulenceFinder 2.0 with a 98% threshold for gene identification and an 80% minimum length.14
Plasmid Analysis
The plasmid sequences were annotated by the NCBI Prokaryotic Genomes Annotation Pipeline (PGAP) server. A graphical map of the vanA-carrying plasmid was converted by CGView Server, complete with labels and footnotes.15 Circular comparisons between pVRE1-VanA and similar plasmids were conducted by BLAST Ring Image Generator (BRIG) as concentric rings.16
Genomic Analysis and Phylogenetic Relationship Analysis
In silico multilocus sequence typing (MLST) analysis was performed using the MLST 2.0 server with the entire genome sequence.14 Plasmid replicon types were analyzed using PlasmidFinder 1.3.14 Identification of insertion elements (ISs) was predicted by the application of ISfinder.17 The phylogenetic relationship between E. faecium VRE1 and other E. faecium strains using a core genome multilocus sequence typing (cgMLST) strategy was performed by BacWGSTdb server. The database currently contains 1758 E. faecium strains, including 61 ST78 strains.18,19
Nucleotide Sequence Accession Numbers
The whole-genome sequence of the strain E. faecium VRE1 and the plasmids was submitted to GenBank under accession number CP040740-CP040744.
Ethical Approval
This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Zhejiang Provincial People’s Hospital. Written informed consent from the patient was exempted by the Ethics Committee of Zhejiang Provincial People’s Hospital because the present study only focused on bacteria. The clinical isolate E. faecium VRE1 was part of the routine hospital laboratory procedure.
Results
MICs of Antimicrobial Susceptibility Testing
The MICs of the antibiotics tested are presented in Table 1. E. faecium VRE1 was resistant to multiple antimicrobials including penicillin, ampicillin, gentamicin, streptomycin, ciprofloxacin, levofloxacin, moxifloxacin, erythromycin, clindamycin, nitrofurantoin, vancomycin, teicoplanin and daptomycin. This strain was susceptible to only three antibiotics, ie, tetracycline, tigecycline and linezolid. E. faecium VRE1 was highly resistant to vancomycin, with an MIC exceeding 256 mg/L.
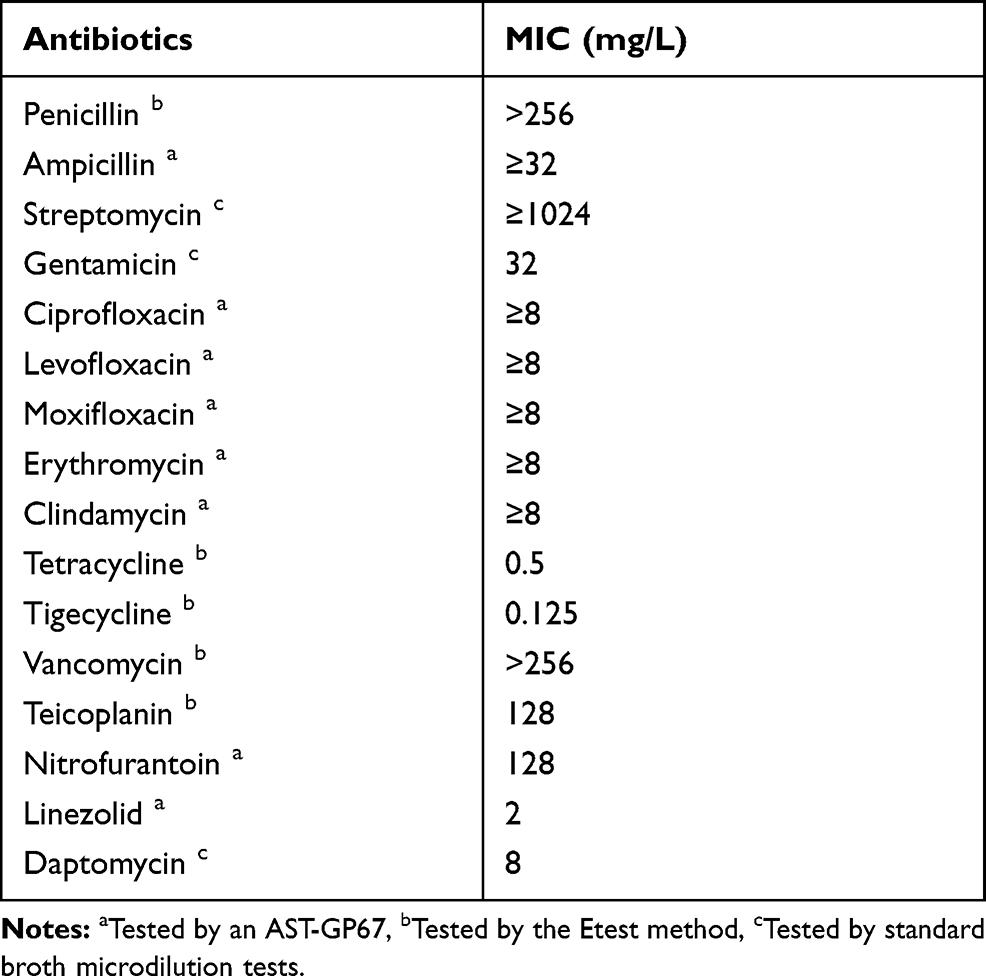 |
Table 1 Minimal Inhibitory Concentrations (MICs) of the Antibiotics for Enterococcus faecium VRE1 |
Genome Characteristics of E. faecium VRE1
The whole-genome sequence of E. faecium VRE1 was composed of one chromosomal DNA comprising 2,718,395 bp and four plasmids with sizes of 238,664 bp, 132,733 bp, 6175 bp and 2056 bp. In the chromosomal DNA, a total of 69 tRNA genes, 3 rRNA operons and 2950 protein-coding sequences were identified by the NCBI Prokaryotic Genomes Annotation Pipeline (PGAP) server. Several IS elements were found in the genome, with the majority belonging to the IS3 and IS6 families. In silico MLST analysis revealed that E. faecium VRE1 belongs to ST78 (clonal complex CC17).
The distributions of the antimicrobial resistance genes and virulence genes in the genome of E. faecium VRE1 are presented in Table 2. Two putative virulence genes, acm and espfm, were detected in the chromosomal DNA, encoding a collagen adhesin precursor and an enterococcal surface protein, respectively. Five acquired antimicrobial resistance genes were identified, ie, macrolide resistance gene msr(C); aminoglycoside resistance genes aac(6ʹ)-Ii, ant(6)-Ia and aph(3ʹ)-III; and glycopeptide resistance gene vanA. Except for aac(6ʹ)-Ii and msr(C) located on the chromosomal DNA, other antimicrobial resistance genes, including vanA, were all located on a 132,733 bp-sized plasmid, pVRE1-VanA. Six additional genes involved in vancomycin resistance, namely, vanR-A, vanS-A, vanH-A, vanX-A, vanY-A and vanZ-A, could also be identified on this plasmid.
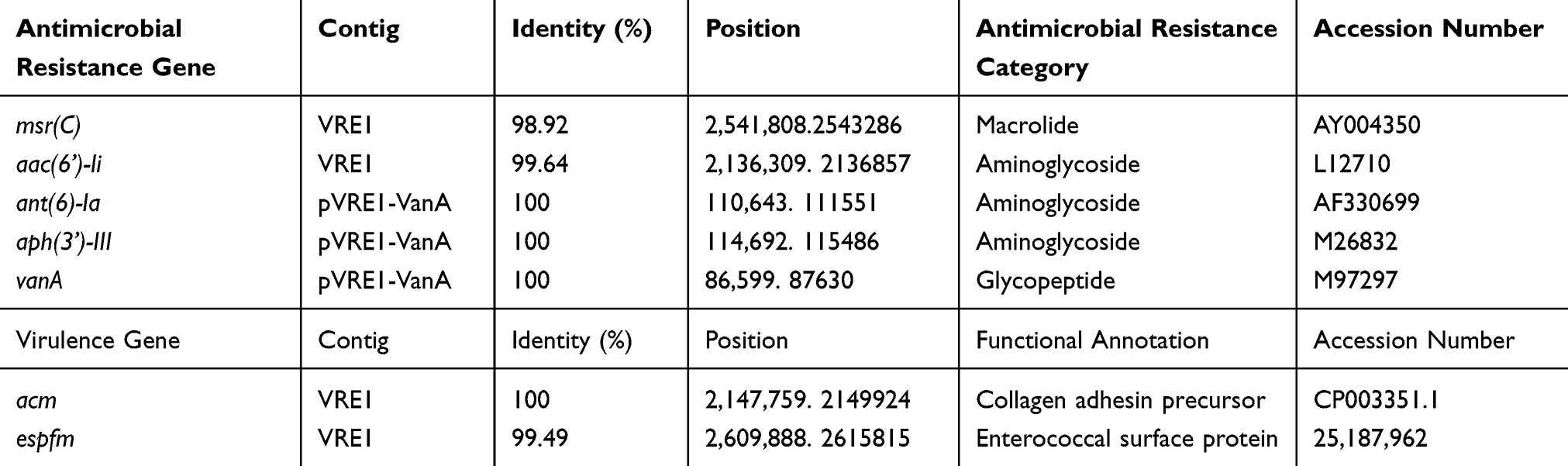 |
Table 2 Antimicrobial Resistance and Virulence Encoded Genes of Enterococcus faecium VRE1 |
Genetic Organization of vanA-Carrying Plasmid
The genetic organization of plasmid pVRE1-VanA is presented in Figure 1. It is a rep2-type plasmid. Several insertion sequences could be identified in this plasmid, including IS1216, IS1542, IS6, IS3, IS30, ISEfm1, ISEfa11, ISEfa4 and IS200/IS605. Among them, IS1216 appeared more frequently than the others. Genome alignment of the complete sequence of pVRE1-VanA with the NCBI GenBank database indicated that pVRE1-VanA appeared to be a novel plasmid.
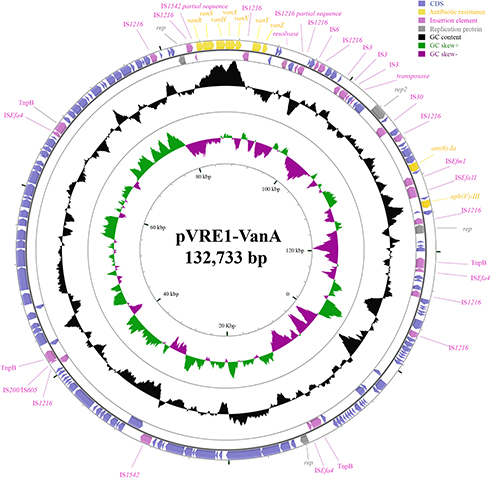 |
Figure 1 Circular representation of the vanA-encoding plasmid pVRE1-VanA. |
Genetic Environment of vanA
The genetic environment of vanA was analyzed, which spanned from 82,560 bp to 95,279 bp of the plasmid pVRE1-VanA. We identified a Tn1546-like element in pVRE1-VanA (Figure 2). Three complete IS1216 sequences and one incomplete IS1216 (IS1216 partial sequence) were found in the element. Among the three complete IS1216 elements, two were distributed on both ends of the element and one was inserted into the vanX-vanY intergenic region. There were IS1542 residues (IS1542 partial sequence) located at the downstream of the first IS1216 and IS1216 residues (IS1216 partial sequence) located at the upstream of the tail IS1216.
 |
Figure 2 Comparison of the homologous regions shared by different types of vanA-carrying plasmids. (A) Typical Tn1546 (E. faecium strain BM4147 plasmid pIP816, accession no. AM932524). (B) Tn1546-like element in this study (E. faecium strain VRE1 plasmid pVRE1-VanA, accession no. CP040742). (C) Similar Tn1546-like element with IS1216 inserted in the vanX-vanY intergenic region (E. faecium strain AUSMDU00004167 plasmid unnamed3, accession no. CP027500). (D) Tn1546-like element most closely related to that in the present study (E. faecium strain 2014-VREF-63 plasmid p63–1, accession no. CP019989). |
Phylogenetic Analysis of E. faecium VRE1
The phylogenetic relationship between E. faecium VRE1 and other E. faecium strains deposited in the NCBI GenBank database was analyzed using the BacWGSTdb server. One phylogenetically related strain was identified in the database: E. faecium strain SC4, with a difference of 178 alleles. E. faecium SC4 was isolated from fodder in 2016 in Beijing, China. E. faecium SC4 also harbors the macrolide resistance gene msr(C) and the virulence genes acm and espfm in its chromosomal DNA. Two additional plasmids could be identified in E. faecium SC4; among them the plasmid p2 (a rep2-type plasmid comprising 142,988 bp, accession no. CP025427) carrying the resistance genes vanA, ant(6)-Ia, aph(3ʹ)-III and erm(B). Genome alignment showed that the plasmid p2 had 80% coverage and 98.92% identity to pVRE1-VanA (Figure 3).
 |
Figure 3 Plasmid sequence alignment of vanA-carrying plasmids that revealed partial sequence identity to pVRE1-VanA. 3: E. faecium E7098 plasmid 3, accession no. LR135256. p2: E. faecium SC4 plasmid p2, accession no. CP025427. Unnamed2: E. faecium VB3025 plasmid unnamed2, accession no. CP040238. p63–1: E. faecium 2014-VREF-63 plasmid p63–1, accession no. CP019989. pEMSRR6: E. faecium SRR6 plasmid pEMSRR6, accession no. MG640601. Unnamed3: E. faecium AUSMDU00004167 plasmid unnamed3, accession no. CP027500. |
Discussion
Since the first clinical VRE isolate was detected in Hangzhou, China, in April 2006, VRE have aroused our serious attention.20 Several studies on VRE from China have been reported, and ST78 was the predominant sequence type in China.21–26 However, the number of studies examining the clonal background of vanA-positive E. faecium strains is still low in China.27
We report the complete genome sequence of a clinically isolated E. faecium ST78 strain harboring vanA from a patient in Hangzhou, China. Genome alignment of the whole sequence of pVRE1-VanA with the NCBI GenBank database indicated that pVRE1-VanA was a novel plasmid. The most closely related plasmid (E. faecium strain E7098 plasmid 3, accession no. LR135256) found in the database had 86% coverage and 99.13% identity to pVRE1-VanA (Figure 3).28 E. faecium strain E7098 was isolated in the Netherlands. Two plasmid replicons (rep2 and rep22) and six resistance genes (ant(6)-Ia, aph(3ʹ)-III, erm(B), lnu(B), tet(L) and vanA) could be identified in plasmid 3.
We also identified a Tn1546-like element in pVRE1-VanA (Figure 2). Compared with the typical Tn1546 element (E. faecium strain BM4147 plasmid pIP816, accession no. KX976485),9 there was an absence of the transposase ORF1 and resolvase ORF2 in pVRE1-VanA. The present Tn1546-like element was similar to the Tn1546-B2-type element, which has the characteristic of the presence of IS1216 upstream of vanR.3,10,29 A similar Tn1546-like element with IS1216 inserted in the vanX-vanY intergenic region was found in the NCBI GenBank database (E. faecium strain AUSMDU00004167 plasmid unnamed3, accession no. CP027500).7 The structure of the present Tn1546-like element was similar to that strain but seems to be more complex (Figure 2). We also found a Tn1546-like element carrying IS1216 and IS1542 elements in similar position to that previously reported in E. gallinarum.30 The most closely related Tn1546-like element to that reported in the present study was the one located on a 287,502 bp repUS15-type plasmid (p63–1) identified in E. faecium strain 2014-VREF-63 in Korea (E. faecium strain 2014-VREF-63 plasmid p63–1, accession no. CP019989).
According to the phylogenetic analysis, the most closely related vancomycin-resistant E. faecium strain, SC4, was found in fodder from Beijing, China with a transferable vanA-carrying plasmid.31 In Hangzhou, the first report of vancomycin-resistant E. faecium was published in 2007 by Qu et al.20 Twenty-one vanA-carrying E. faecium isolates were obtained from inpatients among five hospitals in Hangzhou, from April 2006 to April 2007. The predominant ST was ST78, and a Tn1546-like element with IS1485 inserted between vanXY was found in those isolates. Another research on VRE conducted by Qu et al was published in 2012.23 They collected 45 vancomycin-resistant E. faecium strains from different cities (including Hangzhou) of Zhejiang Province in 2009. In agreement with our results, ST78 resulted to be the predominant ST and all strains carried Tn1546-like elements, although of variable type and different from that described here. These findings highlight the spread also in China of the epidemic ST78. Besides clonal dissemination, horizontal transfer of Tn1546-like elements can contribute to the spread of vancomycin resistance to different enterococcal lineages. Moreover, the presence of additional resistance genes on the Tn1546-carrying pVRE1-VanA plasmid could contribute to the emergence of multidrug-resistant strains in mainland China. Therefore, more studies are required to illuminate the epidemic clones of VanA-positive E. faecium in China.
Conclusion
In summary, our study reports the emergence of a vancomycin-resistant E. faecium ST78 strain with a rep2-type vanA-carrying plasmid in Hangzhou, China. The ST78 lineage possesses the potential to emerge as a successful vanA-carrying epidemic clone. Further studies involving more VanA-producing isolates are warranted to identify reservoirs and monitor the transmission dynamics of vanA genes in China.
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (81702042), Natural Science Foundation of Zhejiang Province (LQ19H200003) and the Zhejiang Provincial Medical and Health Science and Technology plan (2018KY344 and 2019KY311).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Uttley AH, Collins CH, Naidoo J, George RC. Vancomycin-resistant enterococci. Lancet. 1988;331(8575–8576):57–58. doi:10.1016/S0140-6736(88)91037-9
2. Faron ML, Ledeboer NA, Buchan BW. Resistance mechanisms, epidemiology, and approaches to screening for vancomycin-resistant Enterococcus in the health care setting. J Clin Microbiol. 2016;54(10):2436–2447. doi:10.1128/JCM.00211-16
3. van Hal SJ, Espedido BA, Coombs GW, et al. Polyclonal emergence of vanA vancomycin-resistant Enterococcus faecium in Australia. J Antimicrob Chemother. 2017;72(4):998–1001. doi:10.1093/jac/dkw539
4. Weiner LM, Webb AK, Limbago B, et al. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect Control Hosp Epidemiol. 2016;37(11):1288–1301. doi:10.1017/ice.2016.174
5. Sievert DM, Ricks P, Edwards JR, et al. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect Control Hosp Epidemiol. 2013;34(1):1–14. doi:10.1086/668770
6. Hu F, Zhu D, Wang F, Wang M. Current status and trends of antibacterial resistance in China. Clin Infect Dis. 2018;67(suppl_2):S128–s134. doi:10.1093/cid/ciy657
7. Lee RS, Goncalves da Silva A, Baines SL, et al. The changing landscape of vancomycin-resistant Enterococcus faecium in Australia: a population-level genomic study. J Antimicrob Chemother. 2018;73(12):3268–3278. doi:10.1093/jac/dky331
8. Ranotkar S, Kumar P, Zutshi S, et al. Vancomycin-resistant enterococci: troublemaker of the 21st century. J Glob Antimicrob Resist. 2014;2(4):205–212. doi:10.1016/j.jgar.2014.04.002
9. Arthur M, Molinas C, Depardieu F, Courvalin P. Characterization of Tn1546, a Tn3-related transposon conferring glycopeptide resistance by synthesis of depsipeptide peptidoglycan precursors in Enterococcus faecium BM4147. J Bacteriol. 1993;175(1):117–127. doi:10.1128/JB.175.1.117-127.1993
10. Bernasconi OJ, Principe L, Viaggi V, Luzzaro F, Endimiani A. Novel vanA-carrying plasmid in a clinical isolate of Enterococcus avium. Int J Antimicrob Agents. 2019;53(6):876–877. doi:10.1016/j.ijantimicag.2019.04.006
11. Zhu X, Zheng B, Wang S, et al. Molecular characterisation of outbreak-related strains of vancomycin-resistant Enterococcus faecium from an intensive care unit in Beijing, China. J Hosp Infect. 2009;72(2):147–154. doi:10.1016/j.jhin.2009.02.014
12. DiPersio JR, Dowzicky MJ. Regional variations in multidrug resistance among Enterobacteriaceae in the USA and comparative activity of tigecycline, a new glycylcycline antimicrobial. Int J Antimicrob Agents. 2007;29(5):518–527. doi:10.1016/j.ijantimicag.2006.10.019
13. Ruan Z, Wu J, Chen H, Draz MS, Xu J, He F. Hybrid genome assembly and annotation of a pandrug-resistant klebsiella pneumoniae strain using nanopore and illumina sequencing. Infect Drug Resist. 2020;13:199–206. doi:10.2147/IDR.S240404
14. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
15. Grant JR, Stothard P. The CGView server: a comparative genomics tool for circular genomes. Nucleic Acids Res. 2008;36(Suppl_2):W181–184. doi:10.1093/nar/gkn179
16. Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12:402. doi:10.1186/1471-2164-12-402
17. Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(suppl_1):D32–D36. doi:10.1093/nar/gkj014
18. Ruan Z, Feng Y. BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acids Res. 2016;44(D1):D682–687. doi:10.1093/nar/gkv1004
19. Ruan Z, Yu Y, Feng Y. The global dissemination of bacterial infections necessitates the study of reverse genomic epidemiology. Brief Bioinform. 2019;1–10. doi:10.1093/bib/bbz010
20. Qu TT, Chen YG, Yu YS, et al. Molecular characterization of vancomycin-resistant enterococci in Hangzhou, China. J Antimicrob Chemother. 2007;60(6):1403–1405. doi:10.1093/jac/dkm399
21. Cao B, Liu Y, Song S, Li R, Wang H, Wang C. First report of clinical and epidemiological characterisation of vancomycin-resistant enterococci from mainland China. Int J Antimicrob Agents. 2008;32(3):279–281. doi:10.1016/j.ijantimicag.2008.04.005
22. Zheng B, Tomita H, Xiao YH, Wang S, Li Y, Ike Y. Molecular characterization of vancomycin-resistant Enterococcus faecium isolates from mainland China. J Clin Microbiol. 2007;45(9):2813–2818. doi:10.1128/JCM.00457-07
23. Qu TT, Yang Q, Shen P, Wei ZQ, Yu YS. Novel vancomycin-resistance transposon, plasmid replicon types, and virulence factors of vancomycin-resistant Enterococci in Zhejiang, China. Microb Drug Resist. 2012;18(2):183–188. doi:10.1089/mdr.2011.0140
24. Sun HL, Liu C, Zhang JJ, Zhou YM, Xu YC. Molecular characterization of vancomycin-resistant enterococci isolated from a hospital in Beijing, China. J Microbiol Immunol Infect. 2019;52(3):433–442. doi:10.1016/j.jmii.2018.12.008
25. Hua X, Sun L, Shi K, Yu Y. Draft genome sequence of ST203 vanA Enterococcus faecium strain XH877 from China. J Glob Antimicrob Resist. 2016;7:24–25. doi:10.1016/j.jgar.2016.07.004
26. Huang L, Zhang R, Hu Y, et al. Epidemiology and risk factors of methicillin-resistant Staphylococcus aureus and vancomycin-resistant enterococci infections in Zhejiang China from 2015 to 2017. Antimicrob Resist Infect Control. 2019;8:90. doi:10.1186/s13756-019-0539-x
27. Yang JX, Li T, Ning YZ, et al. Molecular characterization of resistance, virulence and clonality in vancomycin-resistant Enterococcus faecium and Enterococcus faecalis: a hospital-based study in Beijing, China. Infect Genet Evol. 2015;33:253–260. doi:10.1016/j.meegid.2015.05.012
28. Arredondo-Alonso S, Top J, Schürch AC, et al. Genomes of a Major Nosocomial Pathogen Enterococcus Faecium are Shaped by Adaptive Evolution of the Chromosome and Plasmidome. bioRxiv; 2019:530725.
29. Novais C, Freitas AR, Sousa JC, Baquero F, Coque TM, Peixe LV. Diversity of Tn1546 and its role in the dissemination of vancomycin-resistant enterococci in Portugal. Antimicrob Agents Chemother. 2008;52(3):1001–1008. doi:10.1128/AAC.00999-07
30. Foglia G, Del Grosso M, Vignaroli C, et al. Molecular analysis of Tn1546-like elements mediating high-level vancomycin resistance in Enterococcus gallinarum. J Antimicrob Chemother. 2003;52(5):772–775. doi:10.1093/jac/dkg445
31. Li N, Yu H, Liu H, et al. Horizontal transfer of vanA between probiotic Enterococcus faecium and Enterococcus faecalis in fermented soybean meal and in digestive tract of growing pigs. J Anim Sci Biotechnol. 2019;10:36. doi:10.1186/s40104-019-0341-x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.