Back to Journals » Neuropsychiatric Disease and Treatment » Volume 15
Effects of prenatal exposure to temephos on behavior and social interaction
Authors Martins Laurentino AO, Durante de Medeiros F ![]() , de Oliveira J, da Rosa N
, de Oliveira J, da Rosa N ![]() , Mateus Gomes T, de Medeiros Peretti E
, Mateus Gomes T, de Medeiros Peretti E ![]() , Somariva Prophiro J, Fortunato JJ
, Somariva Prophiro J, Fortunato JJ
Received 5 December 2018
Accepted for publication 29 January 2019
Published 11 March 2019 Volume 2019:15 Pages 669—673
DOI https://doi.org/10.2147/NDT.S193896
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Ana Olívia Martins Laurentino,1 Fabiana Durante de Medeiros,1 Juliana de Oliveira,1,2 Naiana da Rosa,1,2 Tamires Mateus Gomes,1 Eduardo de Medeiros Peretti,1 Josiane Somariva Prophiro,1,2 Jucélia J Fortunato1,2
1Neurobiology Laboratory of Inflammatory and Metabolic Processes, University of Southern Santa Catarina, Tubarão, Santa Catarina, Brazil; 2Postgraduate Program in Health Sciences, University of Southern Santa Catarina, Tubarão, Santa Catarina, Brazil
Abstract: The neurodevelopment period is susceptible to alterations by genetic and environmental factors, such as the exposure to organophosphates (OP). The OP is neurotoxic and has been associated with neurological diseases pathophysiology. The OP temephos is widely used against Aedes aegypti in Brazil’s public health programs.
Purpose: To evaluate behavioral effects of prenatal exposition to temephos in Wistar rats.
Methods: First, we divided pregnant females into groups: those who received temephos diluted in distilled water by gavage between gestational days 6–13 and those who received only distilled water in the same period and volume. Then, we divided pups according to sex and exposure, and we made the behavioral tests on postnatal day 30.
Results: Prenatal exposure to temephos caused hyperactivity, stereotyped behavior, and social impairment in animals.
Conclusion: These results are similar to the altered behavior presented in some neurobiological diseases models, like Attention Deficit Hyperactivity Disorder and Autism Spectrum Disorders, and this study may bring a red alert to the large use of temephos in Brazil, due to the damage caused by its exposure.
Keywords: neurodevelopment, organophosphate, toxicity, temephos
Introduction
Neurodevelopment begins during the prenatal period and there are maternal and fetal mechanisms that provide a normal evolution. However, genetic and environmental factors can promote alterations in its crucial phases.1 Among the environmental factors, exposition to pesticides has been highlighted in the last few years because their neurotoxic action can deregulate the nervous system homeostasis necessary to normal development.2
Organophosphates (OP) are the most used pesticides in the world and emerged in the market to decrease organochlorines (OC) use due to their high bioaccumulation rate.2 However, OP compounds are neurotoxic, acting by the inhibition of acetylcholinesterase (AChE). This inhibition causes an acetylcholine (ACh) accumulation in the central nervous system (CNS), peripheral nervous system (PNS), and in the neuromuscular junctions, causing irreversible damage to normal neurologic function.2,3
Preclinical3 and clinical4 studies suggest that the poisoning by OP can promote behavioral and neurophysiological alterations and can contribute, because of their influence in the cholinergic system, like a potent environmental factor in the genesis of neurobiological disorders that include Autism Spectrum Disorders (ASD), Alzheimer disease (AD), schizophrenia, and Attention Deficit Hyperactivity Disorder (ADHD).5,6
Temephos is one of the most used OP in health programs in Brazil to control vectors like Aedes aegypti, and vectors of diseases like dengue, zika, and chikungunya.6 Due to the lack of studies about the influence of temephos on behavior, the increase in incidence of neurological and psychiatric diseases in recent years in Brazil, and the neurotoxic potential of OP on neurological diseases genesis,5,7,8 we aim to evaluate behavioral effects of prenatal exposure to temephos in Wistar rats.
Materials and methods
The Ethics Committee of the University of Southern Santa Catarina approved this study, and all the procedures followed the Brazilian Guide for the Production, Maintenance, or Use of animals for teaching or scientific research activities established by the National Council for the Control of Animal Experimentation (CONCEA). Throughout the experimental period, the animals were kept in a room with a controlled temperature (22°C±1°C) and lighting system that guaranteed 12 hours of light environment and 12 hours of dark, receiving standardized commercial rations for laboratory rats and water ad libitum. Besides that, all the animals were from the University of Itajaí Valley (UNIVALI).
The mating procedures were followed in accordance with Fortunato et al.9 The matrices were divided into two groups: those who received temephos (100% pure, Sigma-Aldrich Co., St Louis, MO, USA) diluted in distilled water (50 mg/kg) by gavage during gestational days (GD) 6–13, and those who received only distilled water during same period and volume. Gavage was the chosen route because it allowed rapid and complete absorption, while the choice of a 50 mg/kg dose of temephos was based on the study of Mélo et al.7
The litters remained with their biological mothers until postnatal day (PND) 21, which corresponds to the breastfeeding period. Then, pups were randomly distributed in groups according to sex and prenatal exposure, resulting in four groups with members that were not siblings: a female control group (CG), a male CG, a female temephos group (TG), and a male TG (n=10/group). The open-field10 and reciprocal social interaction tests (RSIs)11,12 were conducted on PND 30 to evaluate behavior. It is important to emphasize that a blind experimenter performed all the tests. The animals were tested at this age because clinical manifestations of ASD and ADHD usually occur during childhood and youth.
Open-field test
We introduced an animal alone on high left quadrant into an arena made of acrylic with 12 quadrants. First, we made a habituation during 5 minutes and, 24 hours later, the test was conducted. The exploratory parameters corresponded to crossing, characterized by the number of crossed lines in arena and rearing, characterized by the movement of being erect on the hind limbs. Stereotypical parameters corresponded to grooming frequency, ie, self-cleaning movements.10
RSI
This test occurred in the same arena as the Open-field test within 5 minutes. We measured social behavior through interaction between the experimental animal and an intruder animal of the same age, weight, and sex as the animal observed. The intruder did not receive any treatment. The parameters included frequency of smelling, mount, and following.11,12
The analysis was performed using the software GraphPad Prism v.5.0. The one-way ANOVA test was used for parametric data followed by Tukey post hoc and Kruskal–Wallis test. The results were presented as mean±standard error. Wilcoxon test was used for comparisons within the group. Statistical significance was set at P<0.05.
Results
Open-field test
The results of this study demonstrated that prenatal exposition to temephos was able to increase the exploratory parameter, represented by crossings, when compared to both CGs (F=5.840; P=0.0030). The analysis of rearing was not significant. This result suggests that prenatal exposure to temephos can cause an increase in exploratory behavior. Besides that, the analysis of grooming parameters, corresponding to stereotyped movements, showed TG females expressed a significant increase when compared to both CGs (F=7.370; P=0.0008) (Figure 1).
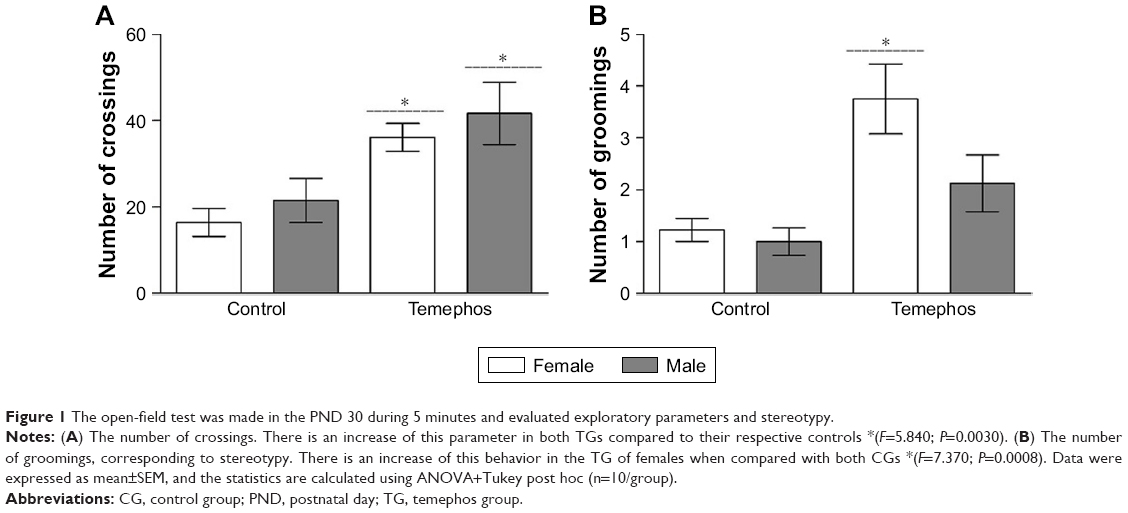 | Figure 1 The open-field test was made in the PND 30 during 5 minutes and evaluated exploratory parameters and stereotypy. |
RSI
The results of the smelling parameter revealed that both TG and CG females presented a significant reduction of this parameter when compared to the GC males (F=6.459; P=0.0022). In addition, there is a tendency for a non-significant reduction of the mount parameter in the TG animals when compared to the CG (Figure 2).
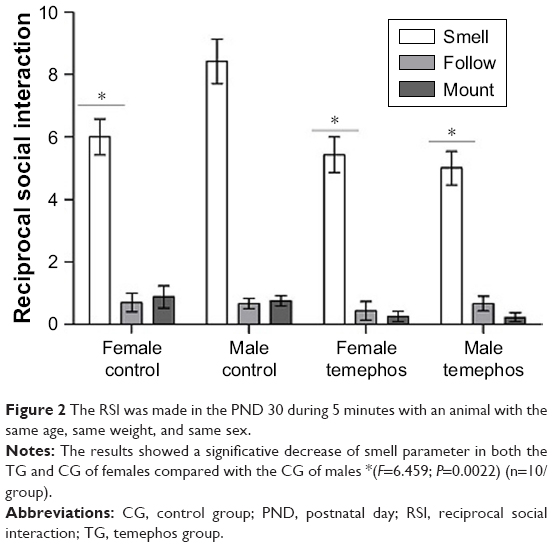 | Figure 2 The RSI was made in the PND 30 during 5 minutes with an animal with the same age, same weight, and same sex. |
Discussion
The neurotoxic effect of OP compounds can cause several alterations in CNS and is associated, as an environmental factor, with physiopathology of neurological diseases such as ASD, ADHD, and Alzheimer’s disease. These diseases have, as clinical manifestations, atypical social behaviors, agitation, and forgetfulness.13
Among the OP, temephos stands out for being widely used in home health programs in Brazil for eradication of the Aedes aegypti, vector of dengue, zika, and chikungunya.7 Considering the knowledge about its use in Brazil and about the effects that it can cause in neurodevelopment,14–16 we aimed to evaluate the behavioral parameters of exploration, stereotypy, and social interaction of rats prenatally exposed to temephos, since these parameters are consistent with behaviors presented by animal models of the aforementioned pathologies.
The results of the exploratory parameters revealed a significant increase of crossings in both TG compared to CG results, with no significant differences between sexes. These results demonstrated prenatal exposure to temephos can cause alterations in exploratory behavior. Similarly, a study conducted by Ricceri et al17 using a protocol of prenatal exposure to OP chlorpyrifos, revealed a significant increase in crossings of the exposed animals compared to the CGs.
The increase in crossings is an important parameter used in animal studies of ADHD. Zhou et al18 evaluated crossings in open-field test and showed that animals with ADHD presented an increase in this parameter, and these were reversed after treatments. In this way, our data demonstrated that prenatal exposure to temephos can promote hyperactive behavior in the animals, which may be associated with clinical manifestations of neurodevelopmental changes, such as ADHD.
Also, clinical studies that aimed to evaluate the concentrations of OP compounds in the urine of American children aged 6–15 years old are related to ADHD diagnoses.19 They suggested that children with higher urinary concentrations of OP were more likely to be diagnosed with this ADHD,19 which is defined as a neurobiological disorder characterized by hyperactive and impulsive behaviors and attention deficits.20 London et al19 affirm that one of the possible etiological agents of ADHD are the OP compounds that would act by deregulating homeostasis of the cholinergic system (CS), which, therefore, would trigger the clinical manifestations.
Nicotinic and muscarinic receptors and their degrading enzymes, such as AChE, which are responsible for the hydrolysis of ACh, compose the CS.19 ACh is an important CNS neurotransmitter and is closely related to cognitive and emotional processes.
Nicotinic receptors are associated with mediation of rapid excitatory transmissions, regulating the release of glutamate, dopamine, and serotonin, and with learning, neurodevelopmental, and memory processes. The muscarinic receptors act in brain regions corresponding to the limbic system (hippocampus and amygdala) and have neurological functions associated with emotional, cognitive, and stress modulation responses.2,19
Therefore, behavioral disorders related to the exploratory parameters and the evidences found in several studies about the relationship between ADHD and OP17,20 suggest that prenatal exposure to temephos is capable of causing alterations in exploratory behavior compatible with the characteristics that validate experimental models of ADHD. However, studies that are more specific are necessary using other behavioral and biochemical parameters that could ensure the effects of OP in the etiology of ADHD.
Pre- and perinatal exposure to OP can also affect social interaction and cognition, and it has been associated with developmental delays and diseases that cause severe behavioral and social impairment in early childhood, such as ASD.21–24 Moreover, this study also aimed to evaluate the parameter of reciprocal social interaction through the RSI test and stereotypy in the open-field test, since the animal’s frequency of grooming corresponds to the stereotyped behavior of autistic patients.
Some studies have shown that prenatal exposure to an OP can promote sex-dependent alterations in behavior but they are contradictory. For example, the study conducted by Gómez-Gimenez et al25 shows that exposure to chlorpyrifos in the developmental phase can affect spatial learning more in males than in females. Our results only demonstrate a sex difference in the stereotypy test, which is discussed below.
The results of grooming showed a significant increase in the female TG when compared to both CG. In addition, the male TG results showed a non-significant increase when compared to the CG. Similar results were found in the study by Dam et al.22 They exposed rats to the OP chlorpyrifos and suggested that behavioral changes found in animals can be associated with the clinical manifestations of neurological diseases, such as ASD.
In addition, data from RSI presented a significant decrease in the smelling parameter of both TG and CG of females compared to the CG of males. Besides that, there was a non-significant tendency to decrease the mount parameter of both TG when compared to CG, suggesting social impairment in TG animals. Laugeray et al26 reached similar conclusions after evaluating rats prenatally exposed to an OP. According to them, this exposure was able to cause damage in communication between animals and in social behavior that is similar to the clinical manifestations of ASD and to the impairments found in animal models of autism.
ASD is a neurobiological disorder that mainly affects the areas of behavior and interest, communication, and social interaction.26 Its prevalence is higher in males than in females, but when it occurs in females the clinical manifestations tend to be more severe due to involvement of hormonal factors.24 Its symptomatology is quite heterogeneous and its etiology is not well defined yet. However, a lot of studies demonstrate that interferences caused by environmental factors, such as exposure to OP, during neurodevelopment may be related to the ASD pathophysiology.26,27
The results of the stereotypy allow us to infer that prenatal exposure to temephos was able to alter the animals’ behavior, causing a higher frequency of stereotypical movements, especially in females, which are consistent with the behavior of animal models of autism. This difference in the behavior of males and females of the TGs occurred probably because clinical manifestations of ASD are more expressive in females when the disease affects them.
Besides that, the results of RSI suggested that exposure to temephos can cause impairments in animals’ social behavior, and this fact may be involved with an alteration in the early development and performance of cholinergic receptors, especially in muscarinic receptors. This alteration may also have caused neurodevelopmental impairment in brain areas where these receptors act, like the hippocampus and amygdala, areas responsible for social behavior and cognitive processes.19
Conclusion
The results showed that exposure to temephos in neurodevelopment is able to cause injury to normal behavior of individuals characterized by hyperactivity, stereotypical behavior, and social impairment. These results are similar to the altered behavior presented in some neurobiological diseases models, like ADHD and ASD, and this study may bring a red alert to the large use of temephos in Brazil due to the damage caused by its exposure. Moreover, this is one of the first studies about the effects of temephos on rat behavior. However, further studies using different protocols of exposure are necessary to explain clearly the relation with pathophysiology of these neurobiological diseases.
Acknowledgment
We are thankful to the University of Southern of Santa Catarina and to the members of the Research Group on Neurobiology of Inflammatory and Metabolic Processes for all their support.
Disclosure
The authors report no conflicts of interest in this work.
References
Burnier M, Fricker AF, Hayoz D, Nussberger J, Brunner HR. Pharmacokinetic and pharmacodynamic effects of YM087, a combined V1/V2 vasopressin receptor antagonist in normal subjects. Eur J Clin Pharmacol. 1999;55(9):633–637. | ||
Wong CT, Wais J, Crawford DA. Prenatal exposure to common environmental factors affects brain lipids and increases risk of developing autism spectrum disorders. Eur J Neurosci. 2015;42(10):2742–2760. | ||
Slotkin TA. Cholinergic systems in brain development and disruption by neurotoxicants: Nicotine, environmental tobacco smoke, organophosphates. Toxicol Appl Pharmacol. 2004;198(2):132–151. | ||
Campbell CG, Seidler FJ, Slotkin TA. Chlorpyrifos interferes with cell development in rat brain regions. Brain Res Bull. 1997;43(2):179–189. | ||
Bouchard MF, Chevrier J, Harley KG, et al. Prenatal exposure to organophosphate pesticides and IQ in 7-year-old children. Environ Health Perspect. 2011;119(8):1189–1195. | ||
Greget R, Dadak S, Barbier L, et al. Modeling and simulation of organophosphate-induced neurotoxicity: prediction and validation by experimental studies. Neurotoxicology. 2016;54:140–152. | ||
Mélo MEB, Merlo KC, Fernandes RRC, et al. Ação mutagênica do inseticida organofosforado temefós em células de medula óssea de camundongos. Rev Inst Adolfo Ltz. 2008;67(3):196–201. | ||
Chediak M, Pimenta Jr PFG, Coelho GE, et al. Spatial and temporal country-wide survey of temephos resistance in Brazilian populations of Aedes aegypti. Mem Inst Oswaldo Cruz. 2016;111(5):311–321. | ||
Fortunato JJ, da Rosa N, Martins Laurentino AO, et al. Effects of ω-3 fatty acids on stereotypical behavior and social interactions in Wistar rats prenatally exposed to lipopolysaccarides. Nutrition. 2017;35:119–127. | ||
Levin ED, Addy N, Nakajima A, Christopher NC, Seidler FJ, Slotkin TA. Persistent behavioral consequences of neonatal chlorpyrifos exposure in rats. Brain Res Dev Brain Res. 2001;130(1):83–89. | ||
Hall CS. Emotional behavior in the rat. I. defecation and urination as measures of individual differences in emotionality. J Comp Psychol. 1934;18(3):385–403. | ||
Bernardi MM, Teixeira LP, Ligeiro-de-Oliveira AP, et al. Neonatal lipopolysaccharide exposure induces sexually dimorphic sickness behavior in adult rats. Psychol Neurosci. 2014;7(2):113–123. | ||
Eaton DL, Daroff RB, Autrup H, et al. Review of The toxicology of chlorpyrifos with an emphasis on human exposure and neurodevelopment. Crit Rev Toxicol. 2008;38(Suppl 2):1–125. | ||
Tannahill D, Harris LW, Keynes R. Role of morphogens in brain growth. J Neurobiol. 2005;64(4):367–375. | ||
Scudeler CGS, Arruda EJ, Andrade CFS, et al. Larval susceptibility of two Culex quinquefasciatus populations (Diptera: Culicidae) Temephos® in the city of Naviraí, MS, Brazil. Orbital Electron J Chem. 2015;7(3):270–274. | ||
Salama M, Lotfy A, Fathy K, et al. Developmental neurotoxic effects of malathion on 3D neurosphere system. Appl Transl Genom. 2015;7:13–18. | ||
Ricceri L, Venerosi A, Capone F, et al. Developmental neurotoxicity of organophosphorous pesticides: fetal and neonatal exposure to chlorpyrifos alters sex-specific behaviors at adulthood in mice. Toxicol Sci. 2006;93(1):105–113. | ||
Zhou RY, Han XM, Wang JJ, et al. [Effect of baicalin on behavioral characteristics of rats with attention deficit hyperactivity disorder]. Zhongguo Dang Dai Er Ke Za Zhi. 2017;19(8):930–937. Chinese. | ||
London L, Beseler C, Bouchard MF, et al. Neurobehavioral and neurodevelopmental effects of pesticide exposures. Neurotoxicology. 2012;33(4):887–896. | ||
Ruckart PZ, Kakolewski K, Bove FJ, Kaye WE. Long-term neurobehavioral health effects of methyl parathion exposure in children in Mississippi and Ohio. Environ Health Perspect. 2004;112(1):46–51. | ||
Ventura ALM, Abreu PA, Freitas RCC, Sathler PC, Loureiro N, Castro HC. Colinergic system: revisiting receptors, regulation and the relationship with Alzheimer disease, schizophrenia, epilepsy and smoking. Rev Psiq Clín. 2010;37(2):66–72. | ||
Dam K, Seidler FJ, Slotkin TA. Chlorpyrifos exposure during a critical neonatal period elicits gender-selective deficits in the development of coordination skills and locomotor activity. Brain Res Dev Brain Res. 2000;121(2):179–187. | ||
Baltazar MT, Dinis-Oliveira RJ, de Lourdes Bastos M, et al. Pesticides exposure as etiological factors of Parkinson’s disease and other neurodegenerative diseases-a mechanistic approach. Toxicol Lett. 2014;230(2):85–103. | ||
Lyall K, Croen LA, Sjödin A, et al. Polychlorinated biphenyl and organochlorine pesticide concentrations in maternal mid-pregnancy serum samples: association with autism spectrum disorder and intellectual disability. Environ Health Perspect. 2017;125(3):474–480. | ||
Gómez-Giménez B, Llansola M, Hernández-Rabaza V, et al. Sex-dependent effects of developmental exposure to different pesticides on spatial learning. The role of induced neuroinflammation in the hippocampus. Food Chem Toxicol. 2017;99:135–148. | ||
Laugeray A, Herzine A, Perche O, et al. Pre- and postnatal exposure to low dose glufosinate ammonium induces autism-like phenotypes in mice. Front Behav Neurosci. 2014;8:390. | ||
Sánchez-Santed F, Colomina MT, Herrero Hernández E. Organophosphate pesticide exposure and neurodegeneration. Cortex. 2016;74:417–426. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.