Back to Journals » Cancer Management and Research » Volume 12
Effects of Annona muricata Extract on Triple-Negative Breast Cancer Cells Mediated Through EGFR Signaling
Authors Alshaeri HK ![]() , Alasmari MM
, Alasmari MM ![]() , Natto ZS
, Natto ZS ![]() , Pino-Figueroa A
, Pino-Figueroa A
Received 30 August 2020
Accepted for publication 10 November 2020
Published 4 December 2020 Volume 2020:12 Pages 12519—12526
DOI https://doi.org/10.2147/CMAR.S278647
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ahmet Emre Eşkazan
Heba K Alshaeri,1,2 Moudi M Alasmari,3,4 Zuhair S Natto,5 Alejandro Pino-Figueroa2
1Department of Pharmaceutical Sciences, Fakeeh College for Medical Sciences, Jeddah, Saudi Arabia; 2Department of Pharmaceutical Sciences, MCPHS University, Boston, MA, USA; 3College of Medicine, King Saud Bin Abdulaziz University for Health Sciences, Jeddah, Saudi Arabia; 4King Abdullah International Research Center, Jeddah, Saudi Arabia; 5Department of Dental Public Health, Faculty of Dentistry, King Abdulaziz University, Jeddah, Saudi Arabia
Correspondence: Heba K Alshaeri
Department of Pharmaceutical Sciences, Fakeeh College for Medical Sciences, Jeddah 21499, Saudi Arabia
Tel +966 504691039
Fax +966 12 6943899
Email [email protected]
Purpose: To evaluate the antiproliferative activity and the mechanisms of action of Annona muricata ethyl acetate (AMEA) extract and one of its active fractions on BT-20 TNBC cells.
Methods: The triple-negative human breast cancer BT-20 cells were used. After the preparation and extraction of Annona muricata ethyl acetate (AMEA), the ethyl acetate extract was exposed to a preparative thin layer chromatography (TLC) plate. From this preparative TLC plate, eight individual bands were collected. Each band was scraped and removed from the plate and soaked in ethyl acetate. After filtration, all eight fractions were then tested on the BT-20 TNBC cells using the MTS cell viability assay. The expressions of EGFR, p-EGFR, AKT, p-AKT, MAPK, p-MAPK, cyclin D1, and NF-κB p65 were measured using Western blot analysis.
Results: The AMEA showed a significant decrease in NF-κB p65 protein expression and BT-20 cell viability, as determined via the MTS assay. Furthermore, the AMEA was subjected to preparative thin layer chromatography (TLC), and eight fractions were obtained. From the eight fractions, only fraction 4 (F4) showed a significant reduction in cell viability in the MTS assay. Immunoblotting analysis revealed that AMEA and F4 formed an antiproliferative effect. These effects were complemented by a downregulation of cyclin D1 assembly, causing cell-cycle arrest at the G1/S phase. Furthermore, NF-κB was measured because of its involvement in the progression of cancers.
Conclusion: The antiproliferative influence is produced through EGFR-mediated signaling pathways, which include AKT, MAPK, NF-κB, and cyclin D1 inhibition. Further studies will be required to demonstrate the possible applications of this natural product in breast cancer therapy.
Keywords: triple negative, breast cancer, Annona muricata, acetogenins
Introduction
Triple-negative breast cancer (TNBC) has an unfavorable prognosis compared with positive breast cancer (BC).1 This type of cancer is responsible for 15–20% of newly diagnosed BC cases.2,3 It is more common among younger females, mainly of African American or Hispanic origin, and among females of low socioeconomic status.4,5
At present, there is increasing interest and research on natural products to assess their anticancer activity. There are many drugs available in the market that originally come from plants.6 The commonly used anticancer agent paclitaxel is a diterpene from Taxus brevifolia (Taxaceae) that was discovered in 1971.6,7 This drug is mainly used in advanced ovarian cancer treatment.6 Other examples include podophyllotoxin from Podophyllum peltatum, camptothecin from Camptotheca acuminata (Nyssaceae), adriamycin from Streptomyces peucetius, and vincristine and vinblastine from Vinca rosea.6 These drugs have motivated scientists to search for new therapies from plant sources. Thus, exploring the traditional use of these natural products and the potential mechanism of active components may provide new therapeutic resources.
Annona muricata L. (A. muricata or AM) is a plant associated with the Annonaceae family, generally known as graviola or soursop.8 Annona species, such as Annona squamosa and Annona reticulata, are used as a natural product in traditional medicine.9 The main active constituents in these plants are acetogenins followed by alkaloids, phenols, and other compounds. A. muricata can be a sedative, pesticide, astringent, insecticide, vermifuge, antiparasitic hypotensive, and antispasmodic.10 Moreover, it can treat fevers, asthma, pain, and coughs and provide wound and skin therapies.10,11 In addition, it has been used for anticancer therapy in South America, Southeast Asia, and tropical Africa, including Nigeria.6,10,12 The most studied parts of AM are the leaves and seeds, probably because they are the most used traditionally, and they contain a high amount of acetogenins.8 Recently, AM showed great activity as an anticancer agent because it can inhibit cancer cell growth in several types of cancer.8,13 An examination of the application of the ethanolic extract of A. muricata leaves to the myelogenous leukemic K562 cell line indicated that it has an effect on apoptosis, and the possible signaling pathway has not been investigated.14,15
Therefore, the objective of this study was to measure the effect of AMEA and its isolated fractions on cell viability. Mechanistic studies were also performed by measuring the phosphorylation of EGFR and its downstream signaling pathways, including the MAPK and PI3K/AKT pathways. Furthermore, NF-κB was measured because of its involvement in the progression of cancer that overexpresses EGFR, as reported in several studies. This research showed that this extract and its active fraction possibly act as antiproliferative agents through inhibition of the EGFR signaling pathway.
Materials and Methods
Plant Materials
The Annona muricata L. was obtained from KatieKountry (Cleburne, TX) in 2015. A voucher specimen of this plant has been placed at the MCPHS University pharmaceutical laboratory, Boston, MA, United States.
Preparation of the Extracts
The dried leaves of AM were ground into a powder with an herb blender to yield 340 g of powder. The powder was then soaked in 3 L of 90% methanol for 5 days at room temperature. This crude extract was filtered with filter paper four times using 1.5 L of 90% methanol. The filtered extract was then pooled and evaporated at 40°C using a Buchi Rotavapor to yield 301.8 g of a concentrated, dark green extract. This methanol extract was then subjected to a partition between water and ethyl acetate at a ratio of 1:1 in a separatory funnel with shaking for 30 min, and the mixture was then allowed to rest for 24 h until complete separation. Then, the ethyl acetate fraction was dried using a rotary evaporator at 60°C. The ethyl acetate fraction was a thick green, viscous residue, and the final obtained weight was 105 g. This sample was the final AMEA extract that was used for further experiments, as shown in Figure S1.
Preparative Thin Layer Chromatography
The ethyl acetate extract was exposed to a preparative TLC plate eluted with 90% ethyl acetate and 10% hexane. From this preparative TLC plate, eight individual bands were collected. The silica gel of each band was scraped and removed from the plate and extracted with ethyl acetate for 1 day. Each fraction was filtered with filter paper into evaporating flasks to be evaporated by a rotary evaporator at 40°C. The following amounts of fraction were obtained: F1, 0.01 g; F2, 0.03 g; F3, 0.06 g; F4, 0.08 g; F5, 0.03 g; F6, 0.02 g; F7, 0.02 g; and F8, 0.06 g, as shown in Figure S2. All eight fractions were then tested on BT-20 triple-negative breast cancer cells using the MTS cell viability assay.
Cell Culture
The triple-negative human breast cancer BT-20 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). BT-20 cells were cultured in Eagle’s minimum essential medium (EMEM) and supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin at 37°C in a 5% CO2 atmosphere until they were confluent.
Cell Viability Assay
This assay was used to assess the cell viability. Briefly, 1.0 × 104 cells/well were seeded in 96-well plates, and after 24 h of incubation, they were treated with different concentrations (1 μg/mL, 3 μg/mL, 10 μg/mL, 30 μg/mL, and 100 μg/mL) of AMEA or F1-F8. The control group consisted of 0.5% DMSO solution-treated cells. Following this exposure, 10 μL of MTS 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium salt reagent was added to each well, and the cells were then incubated for 3 h. Using a UV plate reader (Synergy HT Multi-Mode Microplate Reader, Biotek, Winooski, VT), the cell viability was determined by measuring the absorbance at 490 nm. The cell viability was normalized with respect to the control wells (100% cell viability).
Nuclear Extraction Preparation
The BT-20 cells were seeded into a 25 cm2 cell culture flask at 3×106 cells per flask for 24 h with complete media. At 24 h after the cell seeding, the cells were received in the vehicle, AMEA (10 μg/mL), or F4 (10 μg/mL) for 24 h. The DMSO concentration was maintained at 0.5%. After 24 h of treatment, the nuclear extract was set as defined in the kit assay protocol (Abcam, Cambridge, MA). The protein concentrations for the nuclear and the cytoplasmic extracts were quantified and measured with the colorimetric bicinchoninic acid assay (BCA; Thermo Fisher Scientific, Cambridge, MA).
Western Blot Analysis
Briefly, the total protein levels of EGFR, p-EGFR, AKT, p-AKT, MAPK, p-MAPK, cyclin D1, and NF-κB p65 were assessed using a BCA kit (Thermo Scientific, Waltham, MA). Then, 20 μg of the whole cell lysate was used from each sample to define all the proteins of interest. For NF-κB p65, levels of 20 μg were used from nuclear and cytoplasmic samples. All the samples were loaded in 4–15% polyacrylamide gels (Bio-Rad, Hercules, CA) for separation and then transferred to nitrocellulose membranes (0.45 μm) using a trans-blot turbo system (Bio-Rad) and 1-step transfer buffers (Thermo Fisher Scientific, Cambridge, MA).
The membranes were incubated overnight at 4°C with the primary antibodies—anti-EGFR (1:1000), anti-EGFR (phospho) (1:500), anti-cyclin D1 (1:1000), AKT (pan) (1:1000), phospho-AKT (1:1000), p42/44 MAP kinase (ERK1/2) (1:1000), phospho-p42/44 MAP kinase (ERK1/2) (1:1000), and NF-κB p65 (1:1000)—diluted in blocking buffers. Then, membranes were incubated with HRP-conjugated goat anti-rabbit IgG (1:2000) secondary antibody (ab97051, Abcam) for 1 h at room temperature. The bands were visualized with a C-DiGit blot scanner (Licor Technologies, Lincoln, NE).
Statistical Analysis
Data were expressed as means ± standard error (SE). Any significant differences were assessed by a one-way analysis of variance (ANOVA) with Bonferroni corrections to compare the control and treated groups. Data were considered significantly different when p < 0.05. The samples were tested in triplicate and each assay was completed in three independent experiments. SPSS (IBM, Armonk, NY) software was used to analyze data and plot graphs.
Results
Effects of AMEA on BT-20 Cell Viability
The cell viability assay was conducted to determine the number of viable cells in the test wells. The BT-20 cells were incubated for 24, 48, and 72 h with various concentrations of AMEA (1 μg/mL, 3 μg/mL, 10 μg/mL, 30 μg/mL, and 100 μg/mL). The DMSO was used as a vehicle control and did not exceed 0.5%.
As shown in Figure 1, AMEA decreased the cell viability in a concentration and time-dependent manner compared to the control. The treatment with AMEA showed significant effects at all concentrations in 24 h (p < 0.01), while 1 μg/mL did not show any inhibition in 48 or 72 h. Among all the time-dependent concentrations, 10 μg/mL had the greatest inhibition on cellular viability (23.7%, 10.9%, and 11.2% for 24, 48, and 72 h, respectively), as shown in Figure 1. However, the percentage of cell viability then slightly increased at concentrations of 30 and 100 μg/mL at 24, 48, and 72 h.
 |
Figure 1 Effect of AMEA on cell viability in different time points 24, 48 and 72 hrs. |
Effects of Fractions (F1-F8) on BT-20 Cell Viability
Eight fractions were separated from AMEA using a preparative thin layer chromatography technique. Each fraction was then evaluated via MTS to measure its effect on the cell viability of BT-20 breast cancer cells. As shown in Figure 2, the cell viability did not change after 24 h of incubation at 1 and 3 μg/mL in all eight fractions. However, only fraction 4 (F4) showed a significant reduction at a concentration of 10 μg/mL with approximately 50% inhibition of cell viability.
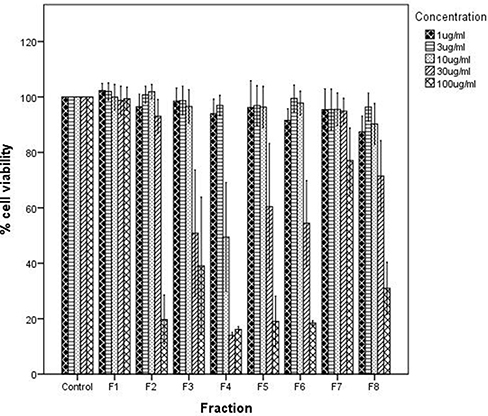 |
Figure 2 Effect of fractions (F1-F8) on cell viability using different concentrations (0, 1, 3, 10, 30 and 100 μg/mL) after 24 h. |
Figure 2 also shows the effect of a concentration of 30 μg/mL on cell viability. The cell viability significantly decreased, compared to the control, at a 30 μg/mL concentration for fraction 3 (F3), fraction 5 (F5), and fraction 6 (F6), but F4 showed a particularly significant reduction at this concentration with less than 20% of the cells viable, which indicated that F4 was the most active fraction in reducing cell viability. Additionally, a significant decrease in cell viability was observed for fraction 2 (F2) and fraction 8 (F8) at a concentration of 100 μg/mL. The concentration of 100 μg/mL also showed a significant reduction of cell viability in F3, F4, F5, and F6 compared to the control (p < 0.01). However, F4 remained the greatest inhibitor of cell viability among all fractions.
Mechanism(s) Involved in AMEA and F4 Antiproliferative Effects
Effects of AMEA and F4 on the EGFR Signaling Pathway
To investigate the inhibitory effect of AMEA and F4 on breast cancer cells, the EGFR, AKT, and MAPK signaling pathways were tested. Briefly, BT-20 cells were treated with either AMEA (10 μg/mL) or F4 (10 μg/mL) for 24 h and subjected to Western blotting. As shown in Figure 3, the phosphorylation levels of EGFR were significantly decreased by AMEA and F4 treatment. Quantitatively, a densitometry analysis of the blots revealed that AMEA and F4 significantly downregulated the EGFR phosphorylation by 90% and 88%, respectively, while the total EGFR levels remained unchanged with both treatments.
 |
Figure 3 The Western blot image showed the effect of AMEA (10 μg/mL) and F4 (10 μg/mL) on the expression of (A) EGFR signaling. (B) the MAPK. (C) AKT. (D) Cyclin D1 on BT-20 cells. |
Effects of AMEA and F4 on MAPK Phosphorylation
The protein expression MAPK is one of the downstream kinases of the EGFR pathway, which regulates several proteins that play essential roles in inducing cell growth and cell survival. In the present study, we measured the phosphorylated and total MAPK with Western blotting. The BT-20 cells were exposed for 24 h to AMEA (10 μg/mL), F4 (10 μg/mL), and DMSO as a vehicle control. The treatment of BT-20 cells with AMEA and F4 significantly downregulated the MAPK protein expression compared to the control. As shown in Figure 3, AMEA reduced the phosphorylated MAPK expression by 61%; however, F4 significantly decreased the phosphorylated protein by 85% compared to the control. The total MAPK expression remained unchanged in all treated groups.
Inhibitory Activity of AMEA and F4 on AKT/PKB Phosphorylation in BT-20 Cells
The AKT is another main protein in EGFR downstream signaling and plays a significant role in the regulation of cell proliferation, differentiation, migration, survival, and invasion. The immunoblotting results, showed in Figure 3, show a reduction in the phosphorylation of AKT in BT-20 cells treated with either AMEA (10 μg/mL) or F4 (10 μg/mL). The densitometric analysis of the bands indicated that AMEA and F4, at a concentration of 10 μg/mL, downregulated phosphorylated AKT by 65% and 80%, respectively, compared to the control. However, the total AKT levels remained unchanged with AMEA and F4 treatment.
Effects of AMEA and F4 on Cyclin D1 Expression
EGFR-induced activation of MAPK and PI3K/AKT pathways plays an essential role in the induction of cyclin D1 and the formation of a complex with CDK4/6 to initiate cell-cycle progression. Thus, the effect of AMEA and F4 on cyclin D1 was investigated using Western blot analysis. The BT-20 cells were exposed to AMEA (10 μg/mL), F4 (10 μg/mL), or DMSO as the control. As shown in Figure 3, the expression level of cyclin D1 was significantly inhibited by 10 ug/mL AMEA (97%) and 10 ug/mL F4 (98%).
Effects of AMEA and F4 on NF-κB p65 Protein Expression
The NF-κB is a transcription factor that plays an important role in cancer formation. As shown in Figure 4A, BT-20 cells that were treated with either AMEA (10 ug/mL) or F4 (10 ug/mL) significantly decreased the NF-κB p65 protein expression in nuclear extracts compared to the control. On the other hand, the treatment of BT-20 cells with either AMEA (10 ug/mL) or F4 (10 ug/mL) had no significant effect in the cytoplasm compared to the control (Figure 4B), indicating that AMEA and F4 could restore the NF-κB p65 protein at the cytoplasmic levels. This compound prevents the translocation of NF-κB to the nuclear level, where it mediates the transcription of several genes involved in cell proliferation.
 |
Figure 4 Effect of AMEA (10 μg/mL) and F4 (10 μg/mL) on NF-κB p65 nuclear protein translocation in BT-20 Cells. (A) Western blot image of cytoplasm NF-κB p65 (upper blot) and β-actin (lower blot). (B) Western blot image of nuclear NF-κB p65 (upper blot) and β-actin (lower blot). |
Discussion
Annona muricata L. (A. muricata or AM) leaves showed great potential as anticancer agents because of their ability to prevent cancer cell growth, as previously described in cancer cell lines such as HCT-116, HT-29, MCF-7, MDA-MB-231, and A549.15,16
A. muricata or graviola contains annonaceous acetogenins that are byproducts of the long-chain C35–C37 fatty acid, which are present in several parts of the plant (leaves, stem, pulp, seeds, etc).17 It was reported that many of these acetogenins apply anticancer effects by inhibiting mitochondrial complex I and ATP syntheses.18 As a consequence, it was suggested that acetogenins might specifically inhibit the growth of cancer cells, which usually use a higher ATP level compared to those of normal cells.12,19 Torres et al revealed that graviola extract reduced cell viability and caused cell death by targeting several pathways that regulate metabolism, the cell cycle, and cell survival and metastasis in pancreatic cancer cells.20 Furthermore, they found that graviola treatment downregulates the expression of hypoxic and glycolysis molecules, such as HIF-1α, NF-κB, GLUT1, GLUT4, hexokinase II, and lactate dehydrogenase-A, in pancreatic cancer. This treatment results in a lower glucose uptake and ATP production.20
To further explore the antiproliferative mechanisms of A. muricata on TNBC, BT-20 cells were investigated. The potential key factor involved in the pathogenesis of TNBC is EGFR. The protein EGFR is commonly overexpressed or constitutively activated in several tumors of epithelial origin, such as breast, lung, and colon cancer. These cancer cells are characterized by uncontrolled proliferation and survival. Thus, novel therapies targeting EGFR and its downstream pathways, such as JAK/STAT, PI3K/AKT, and MAPKs, have been established in cancers with overexpressed EGFR signaling, such as TNBC. In addition, natural compounds have been demonstrated to cause cancer cell apoptosis by preventing the EGFR signaling pathway, including MAPK and PI3K/AKT.20,21 In the current study, we examined the effects of AMEA and F4 on EGFR and its downstream signaling pathways, such as MAPK and PI3K/AKT. The immunoblotting analysis showed that AMEA and F4 significantly suppressed the activation of the EGFR, MAPK, and PI3K/AKT signaling pathways in BT-20 cells. These results indicate that AMEA and F4 may potentially reduce cell proliferation in TNBC through the inhibition of EGFR signaling pathways.
Moreover, it has also been stated that EGFR facilitates NF-κB activation in most cancer cells. The PI3K/AKT signaling pathway is one of the most important mediators that has been identified to activate NF-κB via EGFR stimulation.22 This signaling pathway has been discovered in tumors. Demonstrating this pathway showed that it is accountable for constitutive NF-κB activation in cancer cells.
In fact, when cells are at rest, the NF-κB is associated with the IκB and in an inactive form in the cytoplasm. Once cells are stimulated, IκB is phosphorylated and degraded, which leads to the release of the active NF-κB and activates the target genes transcription that is associated with abnormal proliferation, survival, and the invasion of cancer cells.22
Our study demonstrated that AMEA and F4 remarkably decreased the NF-κB p65 subunit in the nucleus and restored NF-κB p65 protein levels in the cytoplasm. Moreover, these results showed that AMEA and F4 mediated the downregulation of NF-κB activity, which might be an important step for the antiproliferative effect of AMEA and F4 on BT-20 cells. The results also suggested that AMEA and F4 prevented NF-κB activation by suppressing the EGFR signaling pathway. A previous study on lung cancer cells showed that A. muricata extract can effectively minimize the activation of NF-κB and suggested that the presence of NF-κB inhibition is a mechanism in apoptosis.13 However, our results showed that AMEA and F4 treatments decreased cell proliferation in BT-20 cells by suppressing the EGFR and NF-κB signaling pathways. To our knowledge, our findings provided a new mechanism of action for A. muricata, which exhibits antiproliferative effects, as shown in Figure 5, by inhibiting the EGFR-mediated signaling pathways (MAPK, PI3K/AKT, and NF-κB) and causing cell-cycle arrest by suppressing cyclin D1 expression. Thus, this plant does indeed possess anticancer activity, and this effect might be beneficial for treating TNBC.
 |
Figure 5 A diagram shows the antiproliferative mechanism of action of A. muricata extract on TNBC. |
The multiple sequence of events of DNA replication and cell division are closely regulated in the cell cycle. It has been stated that this cycle is mainly controlled in two ways: (1) the posttranslational modifications of proteins that activate the cell from one stage to another and (2) the checkpoint proteins that help to delay or complete cell progression to the next stage.23
Earlier reports have noted that disorders of cell-cycle regulators are general characteristics of human cancers. This result means that targeting cell-cycle progression is considered a possible therapy for human malignancies. Cyclin A, D1, and E along with CDK2, 4, and 6 play important roles in the cell cycle. This result occurs when the cyclins and CDKs work together by phosphorylating the retinoblastoma protein, which allows the G1/S transition progression. Moreover, WAF1-p21 and KIP1-p27, parts of the CIP/KIP group, prevent cell-cycle progression by blocking the activity of the cyclin-CDK complexes.23–25
The effect of multiple antiproliferative agents on cyclin D1 activity has been investigated. It has been proved that the response of EGFR signaling is facilitated by the activation of transcription factors. In fact, one of the most important EGFR signaling pathways is the initiation of cell-cycle progression, the upregulation of cyclin D1, and the downregulation of cyclin-dependent kinase inhibitor proteins (CDKs).22 This pathway provided the rationale for our interest in examining cyclin D1 activity as a consequence of EGFR signaling pathway inhibition.
Thus, the effects of AMEA and F4 on cyclin D1 expression levels as an indicator for the G1 phase were evaluated. An immunoblotting assay targeting a specific antibody against cyclin D1 was conducted. The management of BT-20 cells with either AMEA or F4 showed a significant decrease in the expression level of cyclin D1 compared to the control. These results suggested that AMEA and F4 downregulate cyclin D1 and further cause cell-cycle arrest in the G0 and G1 phases. Our results, in agreement with the previous study by Torres et al, also showed that a decrease in cyclin D1 expression in pancreatic cancer cells, when treated with graviola extract, leads to the induction of G0/G1 cell-cycle arrest.
Overall, the results showed that the effects of this plant were mediated by EGFR signaling pathways that involved MAPK and PI3K/AKT, and NF-κB led to antiproliferative effects and cell-cycle arrest. It has been shown that AMEA and F4 can prevent the activation of an important signaling pathway with multiple downstream biomarkers that are responsible for TNBC tumorigenesis.
The experiments described in this paper were performed in a single cell line, and in their present form, the data are preliminary in nature. It will be more beneficial to investigate the antiproliferative effect of ethanolic extract on other human cell lines. Moreover, the tumor suppressor protein PTEN level depends on AKT signaling. It may help to check the expression of PTEN to complete the signaling axis. In addition, conducting a FACS-based study will determine which cell-cycle stage is blocked by AMEA and fraction 4. Moreover, we did not conduct any morphological examinations, additional dose response studies to support the viability results, or use any cytotoxicity/proliferation assays except the MTS assay. However, we are planning to check these steps in the upcoming research projects.
Conclusion
Our results confirmed that AMEA and F4 significantly inhibited the growth of BT-20 cells. These effects were facilitated by preventing the activation of the EGFR signaling pathways. As a consequence of EGFR inhibition, we found that its major downstream pathways, MAPK and PI3K/AKT, were also suppressed by the treatment. In addition, one result of this study showed that AMEA and F4 reduced the protein expression of NF-κB p65 in the nucleus. Furthermore, AMEA and F4 could decrease the expression of cyclin D1, leading to cell-cycle arrest in the G1 phase. Taken together, these results clearly confirmed the antiproliferative effect of AMEA and F4 in BT-20 cells.
Acknowledgments
We would like to thank Dr. Iman Zaghloul, Dr. Joseph DeMasi and Dr. Swati Betharia for their valuable comments and help in this research project. Part of this manuscript was presented in the Joint Event on 8th World Conference on Women’s Health and Breast Cancer & World Congress on Polycystic Ovarian Syndrome and Fertility, 2018.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Brautigam K, Mitzlaff K, Uebel L, et al. Subtypes of triple-negative breast cancer cell lines react differently to eribulin mesylate. Anticancer Res. 2016;36:2759–2766.
2. Hurvitz S, Mead M. Triple-negative breast cancer: advancements in characterization and treatment approach. Curr Opin Obstet Gynecol. 2016;28:59–69.
3. Khosravi-Shahi P, Cabezon-Gutierrez L, Custodio-Cabello S. Metastatic triple negative breast cancer: optimizing treatment options, new and emerging targeted therapies. Asia Pac J Clin Oncol. 2018;14(1):32–39. doi:10.1111/ajco.12748
4. Carey LA, Perou CM, Livasy CA, et al. Race, breast cancer subtypes, and survival in the Carolina breast cancer study. JAMA. 2006;295(21):2492–2502. doi:10.1001/jama.295.21.2492
5. Schmadeka R, Harmon BE, Singh M. Triple-negative breast carcinoma: current and emerging concepts. Am J Clin Pathol. 2014;141(4):462–477. doi:10.1309/AJCPQN8GZ8SILKGN
6. Liaw CC, Wu TY, Chang FR, et al. Historic perspectives on Annonaceous acetogenins from the chemical bench to preclinical trials. Planta Med. 2010;76:1390–1404.
7. Wani MC, Taylor HL, Wall ME, et al. Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J Am Chem Soc. 1971;93(9):2325–2327. doi:10.1021/ja00738a045
8. Gavamukulya Y, Abou-Elella F, Wamunyokoli F, et al. Phytochemical screening, anti-oxidant activity and in vitro anticancer potential of ethanolic and water leaves extracts of Annona muricata (Graviola). Asian Pac J Trop Med. 2014;7S1:S355–S363. doi:10.1016/S1995-7645(14)60258-3
9. Baskar R, Rajeswari V, Kumar TS. In vitro antioxidant studies in leaves of Annona species. Indian J Exp Biol. 2007;45:480–485.
10. Adewole SO, Ojewole JA. Protective effects of Annona muricata Linn. (Annonaceae) leaf aqueous extract on serum lipid profiles and oxidative stress in hepatocytes of streptozotocin-treated diabetic rats. Afr J Tradit Complement Altern Med. 2008;6:30–41.
11. Iyanda-Joel WO, Ajetunmobi OB, Chinedu SN, et al. Phytochemical, antioxidant and mitochondrial permeability transition studies on fruit-skin ethanol extract of Annona muricata. J Toxicol. 2019;2019:7607031. doi:10.1155/2019/7607031
12. McLaughlin JL. Paw paw and cancer: annonaceous acetogenins from discovery to commercial products. J Nat Prod. 2008;71(7):1311–1321. doi:10.1021/np800191t
13. Zorofchian Moghadamtousi S, Karimian H, Rouhollahi E, et al. Annona muricata leaves induce G(1) cell cycle arrest and apoptosis through mitochondria-mediated pathway in human HCT-116 and HT-29 colon cancer cells. J Ethnopharmacol. 2014;156:277–289. doi:10.1016/j.jep.2014.08.011
14. Yap CV, Subramaniam KS, Khor SW, et al. Annonacin exerts antitumor activity through induction of apoptosis and extracellular signal-regulated kinase inhibition. Pharmacognosy Res. 2017;9(4):378–383. doi:10.4103/pr.pr_19_17
15. Zorofchian Moghadamtousi S, Rouhollahi E, Karimian H, et al. The chemopotential effect of Annona muricata leaves against azoxymethane-induced colonic aberrant crypt foci in rats and the apoptotic effect of Acetogenin Annomuricin E in HT-29 cells: a bioassay-guided approach. PLoS One. 2015;10(4):e0122288. doi:10.1371/journal.pone.0122288
16. Syed Najmuddin SU, Romli MF, Hamid M, et al. Anti-cancer effect of Annona muricata linn leaves crude extract (AMCE) on breast cancer cell line. BMC Complement Altern Med. 2016;16(1):311. doi:10.1186/s12906-016-1290-y
17. Deep G, Kumar R, Jain AK, et al. Graviola inhibits hypoxia-induced NADPH oxidase activity in prostate cancer cells reducing their proliferation and clonogenicity. Sci Rep. 2016;6(1):23135. doi:10.1038/srep23135
18. Degli Esposti M, Ghelli A, Ratta M, et al. Natural substances (acetogenins) from the family Annonaceae are powerful inhibitors of mitochondrial NADH dehydrogenase (complex I). Biochem J. 1994;301(1):161–167. doi:10.1042/bj3010161
19. Oberlies NH, Croy VL, Harrison ML, et al. The Annonaceous acetogenin bullatacin is cytotoxic against multidrug-resistant human mammary adenocarcinoma cells. Cancer Lett. 1997;115(1):73–79. doi:10.1016/S0304-3835(97)04716-2
20. Torres MP, Rachagani S, Purohit V, et al. Graviola: a novel promising natural-derived drug that inhibits tumorigenicity and metastasis of pancreatic cancer cells in vitro and in vivo through altering cell metabolism. Cancer Lett. 2012;323(1):29–40. doi:10.1016/j.canlet.2012.03.031
21. Dai Y, Hogan S, Schmelz EM, et al. Selective growth inhibition of human breast cancer cells by graviola fruit extract in vitro and in vivo involving downregulation of EGFR expression. Nutr Cancer. 2011;63(5):795–801. doi:10.1080/01635581.2011.563027
22. Wee P, Wang Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers (Basel). 2017;9.
23. Duronio RJ, Xiong Y. Signaling pathways that control cell proliferation. Cold Spring Harb Perspect Biol. 2013;5(3):a008904. doi:10.1101/cshperspect.a008904
24. Natto ZS, Parashis A, Steffensen B, et al. Efficacy of collagen matrix seal and collagen sponge on ridge preservation in combination with bone allograft: a randomized controlled clinical trial. J Clin Periodontol. 2017;44(6):649–659. doi:10.1111/jcpe.12722
25. ALHarthi SSY, Natto ZS, Midle JB, et al. Association between time since quitting smoking and periodontitis in former smokers in the national health and nutrition examination surveys (NHANES) 2009 to 2012. J Periodontol. 2019;90(1):16–25. doi:10.1002/JPER.18-0183
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.