Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Effects of a Postbiotic Saccharomyces and Lactobacillus Ferment Complex on the Scalp Microbiome of Chinese Women with Sensitive Scalp Syndrome
Authors Wang Y, Li J ![]() , Wu J, Gu S, Hu H, Cai R, Wang M, Zou Y
, Wu J, Gu S, Hu H, Cai R, Wang M, Zou Y ![]()
Received 4 May 2023
Accepted for publication 11 August 2023
Published 21 September 2023 Volume 2023:16 Pages 2623—2635
DOI https://doi.org/10.2147/CCID.S415787
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Yue Wang,1,2 Jun Li,3 Jianming Wu,3 Shihong Gu,3 Huishu Hu,3 Rongjuan Cai,2 Man Wang,4 Yue Zou3
1Institute of Shanghai Oriental Beauty Valley, Shanghai Institute of Technology, Shanghai, People’s Republic of China; 2School of Perfume and Aroma Technology, Shanghai Institute of Technology, Shanghai, People’s Republic of China; 3R&D Center, JALA (Group) Co., Ltd, Shanghai, People’s Republic of China; 4Department of Nutrition, Shanghai Jiao Tong University Affiliated Sixth People’s Hospital South Campus, Shanghai, People’s Republic of China
Correspondence: Yue Zou, R&D Center, JALA (Group) Co., Ltd, Building 2, No. 333, Guiping Road, Xuhui District, Shanghai, 200233, People’s Republic of China, Email [email protected]
Introduction: Sensitive scalp is one of the most frequent complaints related to sensitive skin syndrome, characterized by unpleasant sensory reactions in the absence of visible signs of inflammation. In this study, the effects of topical application of postbiotic Himalaya-derived Saccharomyces and Lactobacillus ferment complex (SLFC) on the bacterial and fungal scalp microbiome at the taxonomic level and alleviation of sensitive skin syndrome were investigated.
Methods: Firstly, healthy female participants (aged 30– 45) were classified into a healthy scalp group and a sensitive scalp group based on the questionnaire. Thereafter, topical application of SLFC on sensitive scalp as well as scalp microbiome was evaluated, with the difference in the distribution of microbial taxa between healthy and sensitive scalp communities was assessed using 16S rRNA and ITS1 sequencing analysis. In addition, the effect of SLFC on scalp microbiome at the species level for Cutibacterium acnes, Staphylococcus epidermidis, and Malassezia restricta was evaluated by the qPCR assessment.
Results: After treatment with SLFC for 28 days, the abundance of Staphylococcus, Lawsonella, and Fusarium in the sensitive scalp group was highly significantly increased (p < 0.001), while the abundance of Cutibacterium and Malassezia was highly significantly decreased (p < 0.001). Furthermore, the self-assessment questionnaire indicated a syndrome alleviation effect of 100% after 28 days with a twice-daily application of the SLFC.
Discussion: The obtained results would help to better understand the microbial community of the sensitive scalp and provide useful information on utilization of SLFC for maintaining a healthy scalp and modulating the scalp microbiome.
Keywords: sensitive scalp, scalp microbiome, postbiotics, microbial diversity, questionnaire analysis
Introduction
Sensitive scalp is a skin syndrome caused by the hyper-reactivity to environmental stimuli, which might cause inflammatory symptoms and abnormal sensory reactions of the scalp including pruritus, prickling, tightness, pain, and burning, but without visible signs of inflammation.1 Various circumstances, such as the atmospheric environment, heat, pollution, hair care products, diet, emotions, and skin microbial composition, have been suggested to be relevant factors that could result in erythema of the scalp, redness, dandruff, hair loss, and scalp disease.2 It has been reported that sensitive scalp is a frequent phenomenon all over the world, especially in women, which could be triggered by physical, chemical, and psychological factors.3 For example, the proportion of sensitive scalp among Chinese women was reportedly as high as 57% based on the “3S Questionnaire” investigation, while 44% of French survey respondents reported suffering from sensitive scalp with higher levels in women than that in men.1,4
In people with sensitive skin, the decreased skin tolerance threshold can impair the skin barrier function and result in abnormalities of the cutaneous nervous system.5 However, there is no specific treatment for sensitive scalp due to the complexity of its etiology and pathogenesis.6 Visible signs of inflammation are usually absent in sensitive skin, which is traditionally not regarded as a serious disease, although it has become a common condition that severely impacts the quality of everyday life.7 Concerns regarding skin sensitivity have gradually increased in recent years due to acute environmental stressors and inappropriate utilization of cosmetic products.8 For people with a sensitive scalp, mild and diluted anionic surfactant products are often recommended.2,5 As reported previously, application of targeted cosmetic products and maintenance of daily skin care habits might play a critical role in the management of sensitive scalp syndrome.8
Although the triggering factors of sensitive skin have been extensively investigated, the underlying pathophysiological mechanisms remain poorly understood.9 However, recent studies have demonstrated that sensitive scalp and scalp disorders have a strong correlation with the commensal scalp microbiome.10,11 In fact, the skin microbiota contributes to nearly all aspects of skin health and function.12 For example, abnormal sebum accumulation and a perturbed microbiome might disrupt the scalp barrier function, which was suggested to be the direct cause of sensitive scalp syndrome.13 The human skin sebum has a complex chemical composition including triglycerides, fatty acids, wax esters, cholesterol, cholesterol esters, and squalene, which are important components that play protective roles in the skin barrier maintenance of scalp health.6,13 Commensal microbes, including fungi and bacteria, can degrade sebum components, such as triglycerides and esters, into diglycerides, monoglycerides, and free fatty acids.14 Insufficient or excess sebum might cause various skin diseases, such as scarring, alopecia, and acne vulgaris.15 Therefore, the scalp physiological condition could be modulated by the scalp microbiome through regulation of the microenvironment.
In the past decade, numerous studies have been devoted to determining the taxonomic composition and diversity of bacterial and fungal communities inhabiting the scalp in different populations.11,16 In adults with healthy skin, the skin microbial composition is surprisingly stable and may contribute to the establishment of cutaneous homeostasis as well as modulate the inflammatory response.17 Conversely, dysbiosis of the skin microbiome has been frequently reported to be associated with scalp disorders and diseases.18,19 Hence, modulating the scalp microbiome and preserving the cutaneous homeostasis might provide viable strategies for the restoration and maintenance of skin health. In recent years, applications of prebiotics, probiotics, and postbiotics in skin care products emerged as a trend with beneficial effects on the skin microbiome.20–22
In this study, a postbiotic complex was prepared by mixing the fermentation filtrate of Saccharomyces cerevisiae and Lactobacillus paracasei at a mass ratio of 1:1.5–2. The complex was found to be rich in amino acids, water-soluble vitamins, organic acids, fatty acids, polysaccharides, and large amounts of oligopeptides. The Saccharomyces and Lactobacillus fermentation filtrates are marketed as HiMurchaSin® and LacSherTongba®, respectively. HiMurchaSin® is fermented by a wild-type S. cerevisiae Y017 from the Himalayan region in a polar-like environment, while L. paracasei B003 of LacSherTongba® was screened from traditional finger millet (Eleusine coracana) wine of Sherpas in the Himalayan region.23,24 The aim of this work was to study the functional effect of SLFC on sensitive scalp and evaluate its capacity to modulate the scalp microbiome. The obtained results will aid the development of functional postbiotic products for the treatment of sensitive scalp and other skin dysfunctions in the future.
Materials and Methods
Subject Recruitment and Classification
The research complied with the World Medical Association Declaration of Helsinki. Ethical approval was provided by the Institutional Ethics Committee of Shanghai Jiao Tong University Affiliated Sixth People’s Hospital South Campus (Approval No.: 2022-KY-07-01). Prior to any study-related procedures, all subjects provided written informed consent. In addition, a questionnaire was designed to evaluate the situation of sensitive scalp based on a reported method which had been established and validated previously.4 The participants were healthy females (aged 30–45) and were classified into a healthy scalp group and a sensitive scalp group based on the questionnaire. A total of 60 female volunteers were screened after questionnaire evaluation (Table 1), out of which 30 subjects with a healthy scalp (HS) and 30 subjects with a sensitive scalp (SS) were enrolled. All participants provided information regarding medical history and daily habits. The volunteers were non-smokers, free from any cutaneous diseases, did not consume or apply antibiotics or systemic antifungals for 1 month prior to sampling, did not use hair-loss treatment for at least 3 months prior to sampling, and did not use hair products (such as for dyeing, bleaching, permanent waving, and straightening) on the scalp and hair for at least 4 weeks prior to sampling.
 |
Table 1 Assessment of Symptoms of Scalp Situation |
Self-Assessment Questionnaire
The volunteers were asked to use a neutral shampoo every 2 days for a period of 4-weeks prior to the beginning of the study to standardize the scalp condition.25 Volunteers with a sensitive scalp (SS group) were treated with SLFC two times per day (in 12 h intervals) for a period of 4 weeks to evaluate its effect on sensitive scalp condition and skin microbiome. The efficacy of SLFC was subjectively evaluated by a self-assessment questionnaire during the 28 days of utilization. At day 0 (D0), day 14 (D14), and day 28 (D28), the SS group was presented with statements based on different parameters of the cosmetic attributes, effectiveness, and consumer opinion, and were asked to rate each statement with a score ranging from 0 to 3, where 0 = Completely disagree, 1 = Disagree, 2 = Agree and 3 = Completely agree. The percentage was calculated for each statement, and satisfaction was defined as scores ranging from 2 to 3.26
Sampling of the Scalp Microbiome
The scalp surface was sampled by means of a swab procedure according to the previously reported method,27 with minor modifications as follows. Volunteers were advised not to wash their scalp for 2 days prior to the sampling procedure. Samples from the scalp (vertex or crown of the head) were obtained. Briefly, a sterile cotton swab was first soaked in collection solution (0.15 M NaCl and 0.1% Tween 20), then rubbed onto the scalp surface (approximately 30 times for at least 20 seconds between the hairs) in a zig-zag pattern to cover a total surface of 4×4 cm2 in a non-overlapping manner.11 Thereafter, the head of each swab was cut and stored in a sealed tube containing 5 mL of collection buffer solution. In addition, sterile cotton swabs were treated the same way as negative controls. As described previously,24 the swabs were immediately stored at −80 °C and subjected to DNA isolation within 24 h.
DNA Extraction for High-Throughput Sequencing
Genomic DNA was extracted using the commercial Fast DNA Spin Kit (MP Biomedicals, USA) using the specific protocol provided by the supplier. Each sample (2 mL) was generated from the stored collection tube and centrifugated at 10,000g for 30 min at 4 °C to obtain the microbial cell pellets.11 For DNA extraction, the obtained cells were re-suspended in 500 μL of sterile TE buffer (10 mM Tris-HCl; 1 mM EDTA, pH 7.4). Subsequently, cell suspensions were added with 3 μL of Ready-Lyse lysozyme (250 U/μL) (Epicentre, Wisconsin, USA) and incubated for 12 h at 37 °C, followed by bead beating at 6.5 m/s for 45s, and repeated after 5 minutes using the Fast Prep 24 machine (MP Biomedicals, California, USA). The remaining steps were performed according to the manufacturer’s protocol and the extracted DNA was stored at −80 °C for subsequent sequencing.
PCR Amplification and Sequencing
The extracted microbial DNA (∼1 ng) was used for polymerase chain reaction (PCR) to obtain amplicons using universal primers.28 For bacteria, the V3-V4 variable region in the 16S rRNA gene was amplified by PCR using a conventional thermocycler (GeneAmp 9700, ABI), with primers 380F (5′- ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′- GGACTACHVGGGTWTCTAAT-3′). For fungi, the ITS1 region of the rRNA gene was amplified using the primers ITS1F (5′- CCTACGGGNGGCWGCAG-3′) and ITS2R (5′- GACTACHVGGGTATCTAATCC-3′). PCR reactions for bacteria and fungi were carried out in triplicate using 1 μL of primer (5 μM), 2 μL of 10× CoralLoad Concentrate, 10 μL of HotStart Taq Plus MasterMix (Qiagen, Germany), 2 μL of template DNA, and 4 μL of nuclease-free water. PCR amplicons were purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, USA), libraries were prepared using the NEXTFLEX Rapid DNA-Seq Kit (Bioo Scientific, USA), and sequencing was performed using a MiSeq platform (Illumina, USA) to generate 300-bp paired-end reads according to the standard protocols provided by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China).
Processing of Sequencing Data
The processing of sequencing data was performed according to the reported methods,29,30 with minor modifications as follows. The raw data from high-throughput sequencing were collated, quality-filtered using the q2-demux plugin, and denoised using the DADA2/Debrur sequence noise reduction method to obtain the Amplicon Sequence Variant (ASV) representative sequences and abundance information. Based on the representative sequence and abundance information of ASV, a series of statistical and visual analyses were carried out, including species taxonomy analysis, community diversity analysis, species difference analysis, correlation analysis, phylogenetic analysis, and functional prediction analysis. Chimeric sequences were identified and removed using UCHIME. Taxonomy was assigned using Silva version 138 (99% of the full-length sequences of OTUs) for bacterial 16S sequences using a confidence threshold of 70% and Unite version 8.0 for fungal 28S sequences.28
Quantitative PCR (qPCR)
Amplification primers were designed according to the previously reported method and probes were labeled with the fluorescent reporter dye 6-FAM (Fluorescein) at the 5′ end and BHQ1 as a quencher at the 3′ end.26 Total RNA was extracted using a Trizol RNA isolation kit (Key GEN, Nanjing, China), and a total of 500 ng RNA was reverse-transcribed into cDNA using a Prime Script RT reagent kit (Takara, Shiga, Japan). The qRT-PCR was performed using SYBR® Premix Ex Taq™ II (Takara, Shiga, Japan), with the primers listed in Table 2. The qPCR reactions comprised 10 μL Premix Ex Taq™ II, 2 μL of primer mix (5 μM), 2 μL of the 10 μM probe, and nuclease-free water to a final volume of 20 μL. Subsequently, each DNA sample (2 μL) or dilutions of DNA standards were added to the 96-well assay plate, after which the plate was sealed and placed in an ABI 7500 real-time PCR machine. The RT-PCR temperature program encompassed 40 cycles of 95 °C for 30 s, 60 °C for 30 s, and 70 °C for 1 min. The data were processed, and the copy numbers were calculated using the ABI 7500 software. The qPCR experiments were conducted in triplicate and summarized using the median value.26
 |
Table 2 Sequences of qPCR Primers and Probes for the Quantification of Bacterial and Fungal Taxa |
Statistical Analysis
All quantitative data are presented as means ± standard deviations (X±SD). Differences with p-values <0.05 were considered statistically significant. The taxonomical data were expressed as relative abundance. After obtaining the data on community abundance, we used the Kruskal–Wallis H-test to analyze the differences of microbial communities among the groups. The alpha diversity was calculated from amplicon reads using the metric Shannon index and species were observed after rarefying from 100 sequences at a step size of 6000 for V3 as well as for ITS1 amplicons using QIIME v1.9. Beta diversity was analyzed by calculating the weighted UniFrac distances for the bacterial population. The data were analyzed online on the Majorbio I-Sanger Cloud Platform (www.i-sanger.com).
Results
Taxonomic Profile of the Bacterial Community
After treatment with SLFC for 28 days, the bacterial and fungal microbiomes of the scalps of Chinese women (30 with healthy scalps and 30 with sensitive scalps) were analyzed. Cloning and sequencing of the conserved ribosomal regions (16S for bacteria and 28S-ITS for fungi) was used to identify the major bacterial and fungal scalp colonizers. The alpha diversity of the scalp microbiome in the HS and SS groups was analyzed based on the obtained diversity indices (Figure 1). The Ace, Chao, and Sobs indices of the HS group were relatively lower (p > 0.1) compared to the SS group before SLFC intervention (SS0), while the Simpsoneven index was relatively higher (p > 0.1) than that of SS0. In the SS group treated with SLFC, the alpha diversity (Ace, Chao, Sobs, and Simpsoneven index) changed obviously (p < 0.01) on day 28 (SS28), indicating the modulating effect of SLFC on the scalp microbiome.
 |
Figure 1 Alpha diversity of scalp bacteria microbiome in HS and SS groups intervened by SLFC (*0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01). Abbreviations: HS, healthy scalp; SS, sensitive scalp, SS14; SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. Notes: (A) Ace index, (B) Chao index, (C) Sobs index, (D) Simpsoneven index. |
To address the potential differences in the composition of the microbiome at the genus level, β-diversity analysis was performed through hierarchical clustering and Principal Coordinate Analysis (PCoA) based on Bray–Curtis distances followed by PERMANOVA statistical analysis (Figure 2). As shown in Figure 2, differential microbiota composition was observed after SLFC treatment for 14 and 28 days. Similarly, β-diversity analysis at the species level revealed a significant shift in microbiota composition after treatment for 14 or 28 days (R2=0.0862, p=0.001), which suggested a change of bacterial diversity at the genus level.
 |
Figure 2 Beta diversity analysis of bacterial communities in the SS group stratified by SLFC treatment. Abbreviations: SS, sensitive scalp; SS14, SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. |
Moreover, there was a significant shift in microbiota composition at the genus level on day 28 (SS28) compared to SS0 (Figure 3). In the sensitive scalp group before (SS0) and after treatment (SS14 and SS28) with SLFC, Cutibacterium predominated in the bacterial microbiome, followed by Staphylococcus, and Lawsonella (Figure 3A). These three taxa accounted for >90% of all bacteria at all tested stages. The proportion of Staphylococcus increased significantly (p < 0.01) after 28 days (SS28) of treatment (Figure 3B). At the same time, Staphylococcus was more abundant than Cutibacterium at day 28, indicating a significant decrease (p < 0.001) of Cutibacterium after SLFC treatment (Figure 3B). For SS14 and SS28, the abundance of Cutibacterium decreased, while the proportion of Staphylococcus and Lawsonella increased as compared to SS0, which revealed the potential benefits of SLFC as the proportion of the three taxa was close to HS group (Figure 3A and B). It also suggested more treatment days with SLFC might reconstruct the bacterial community to normalize the flora distribution of the sensitive scalp to a healthy degree. These results indicated that SLFC could significantly modulate the composition of the bacterial community in the sensitive scalp group.
 |
Figure 3 Relative abundance of the predominant bacteria at the genus level (*0.01 < P ≤ 0.05, ***P ≤ 0.001). Abbreviations: HS, healthy scalp; SS, sensitive scalp; SS14, SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. Notes: (A) Principal component analysis of scalp microbiota stratified by SLFC treatment; (B) genus-level differences in relation to SLFC treatment. |
Taxonomic Profiles of Fungal Community
The alpha diversity for the fungal population was significantly different in the scalp of healthy subjects (HS) compared to the sensitive scalp subjects (SS0) at the beginning of the study (Figure 4). The Ace, Chao1, and Sobs indices of the HS group were significantly lower (p < 0.01) compared to the SS group before SLFC intervention (SS0). After treatment with SLFC for 14 and 28 days, the alpha diversity of the fungal microbiome greatly changed compared to SS0 (Figure 4). As shown in Figure 4, the alpha diversity of the fungal community at day 28 (SS28) greatly changed (p < 0.01) compared to SS0, while there was no significant difference compared to the HS group. These results suggested that treatment with SLFC for 28 days could modulate the fungal microbiome of the sensitive scalp group by changing the fungal population and composition.
 |
Figure 4 Alpha diversity of the scalp fungal microbiome in the SS group treated with SLFC (**0.001 < P ≤ 0.01). Notes: (A) Ace index, (B) Chao index, (C) Sobs index. |
In the sensitive scalp group, the β-diversity analysis at the species level revealed a significant change (R=0.045, p=0.002) of the fungal community after 14 and 28 days of intervention (Figure 5), which confirmed the effects of SLFC on the sensitive scalp microbiome.
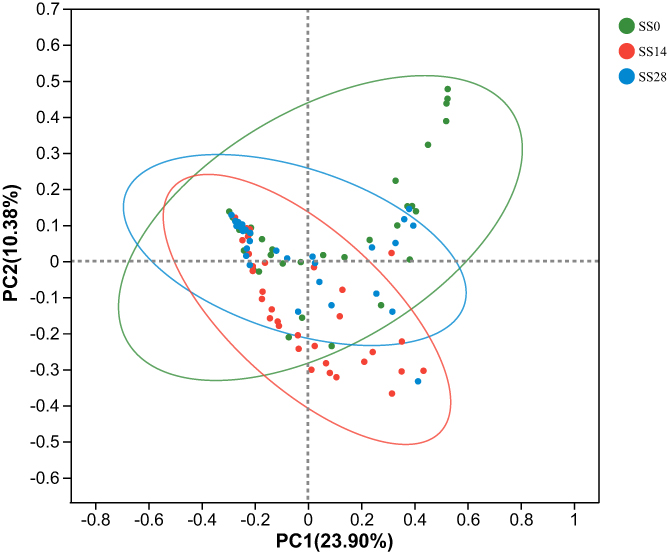 |
Figure 5 Beta diversity analysis of the fungal community. Abbreviations: SS, sensitive scalp; SS14, SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. |
The dominant fungal taxa of all the tested groups were mostly Basidiomycota, with the predominant genus being Malassezia (Figure 6). At the genus level, the samples were mainly composed of Malassezia, Apiotrichum, and Fusarium (Figure 6A). Compared with the HS group, the relative abundance of Malassezia was significantly higher (p < 0.05) in the sensitive scalp group (SS0) before utilization of SLFC. At day 28, however, the proportion of Malassezia in the SS group (SS28) exhibited an obvious decrease (p<0.01) compared to day 0 (Figure 6C) and was similar to that of the HS group (Figure 6B). Conversely, the relative abundance of Fusarium significantly increased after 28 days of SLFC intervention (Figure 6B). These results demonstrated that the postbiotic SLFC could also change the fungal diversity in addition to the bacterial diversity, illustrating its superior capacity to modulate the skin microbiomes of women with sensitive scalp syndrome.
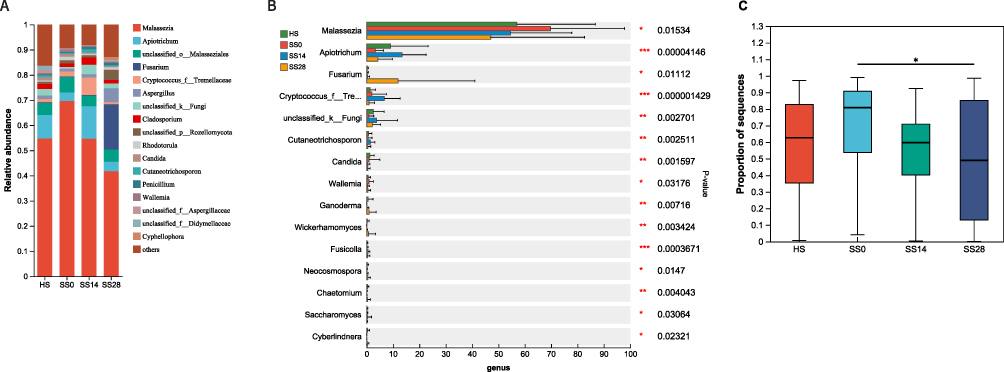 |
Figure 6 Relative abundance of the predominant fungi at the genus level (*0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01, ***P ≤ 0.001). Abbreviations: HS, healthy scalp; SS, sensitive scalp; SS14, SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. Notes: (A) Principal component analysis of microbiota stratified SLFC treatment, (B) changes of scalp microbiota in relation to SLFC treatment, (C) changes of Malassezia in SS group. |
qPCR Assessment
The qPCR evaluation was carried out at the species level for Cutibacterium acnes, Staphylococcus epidermidis, and Malassezia restricta. In the sensitive scalp group, the absolute abundance of C. acnes increased significantly at day 28 compared to day 0 (p<0.001) and day 14 (p<0.01) (Figure 7A). Similarly, the absolute abundance of S. epidermidis increased significantly at day 28 compared to day 0 (p<0.01) and day 14 (p<0.05) (Figure 7B). As shown in Figure 7C, a significant decrease was observed in the absolute abundance of M. restricta in the sensitive scalp group treated with SLFC for 28 days (p<0.001). In the healthy scalp group, the average ratio of C. acnes/S. epidermidis was lower than that of the non-treated sensitive scalp group by 89.52% (Figure 7D). After treatment with SLFC, the ratio decreases by 69.11% and 84.24% on day 14 and day 28 as compared to day 0, respectively (Figure 7D). In the sensitive scalp group with SLFC treatment, the ratio of C. acnes/S. epidermidis on day 28 was not significantly different from the healthy scalp group (Figure 7D).
 |
Figure 7 Quantification of representative taxa at the species level by qPCR (*0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01, ***P ≤ 0.001). Abbreviations: HS, healthy scalp; SS, sensitive scalp; SS14; SLFC treatment for 14 days; SS28, SLFC treatment for 28 days. Notes: (A) the absolute abundance of C. acnes, (B) the absolute abundance of S. epidermidis, (C) the absolute abundance of M. restricta, (D) ratio of the absolute abundance of C. acnes to S. epidermidis. |
Evaluation of the Effect of SLFC Treatment on Sensitive Scalp Syndrome
The efficacy of SLFC treatment for 28 days was subjectively evaluated by a self-assessment questionnaire, answered by each subject on days 14 and 28. Subject satisfaction was defined as scores ranging from 2 (agree) to 3 (completely agree). In the sensitive scalp group, the self-assessment questionnaire revealed an overall syndrome alleviation effect of 100% after 14 and 28 days of treatment with the postbiotic SLFC (Table 3). Additionally, the self-assessment questionnaire demonstrated that 100% of subjects felt alleviation of itching symptoms, 96% felt alleviation of prickling, tightness, pain, and dandruff symptoms, while 92% felt alleviation of burning symptoms. These results suggested that SLFC might serve as a promising product for the treatment of sensitive scalp syndrome among Chinese women. The results of this work suggested that SLFC could significantly alleviate sensitive scalp symptoms.
 |
Table 3 Symptom Alleviation Effect of SLFC on Sensitive Scalp Syndrome |
Discussion
The inclusion of prebiotics and postbiotics in cosmeceutical products provides the opportunity to balance the skin microbiome to achieve overall skin health. In this work, postbiotic SLFC was evaluated for its ability to modulate the scalp microbiome and alleviate sensitive scalp syndrome among Chinese women.
Bacterial 16S rRNA sequence analysis revealed that the distribution of bacteria showed little difference between the healthy and sensitive scalp groups, while the fungal communities exhibited significant differences based on fungal ITS1 sequencing. This was consistent with a previous report, which indicated that bacterial community of individuals with scalp problems was not statistically different from that of participants with a healthy scalp.31 At the same time, the results also indicated that the fungal microbiome might play an important role in sensitive scalp syndrome among Chinese women.
After treatment with SLFC for 28 days, the microbial community of the sensitive scalp group changed significantly. Changes of Ace, Chao, Sobs, and Simpsoneven indices reflected a reduction of species diversity and better evenness of the scalp microbiome. The potential active components of postbiotic SLFC mainly included polysaccharides, antimicrobial peptides, amino acids, vitamin B6, lactic acid, and other organic acids. Lactic acid and other organic acids could reduce the pH of the skin microenvironment and consequently restrict the colonization and reproduction of skin pathogenic bacteria.32 Antimicrobial peptides synthesized by probiotics have an inhibitory effect on Gram-positive bacteria with a narrow antibacterial spectrum. In addition, probiotics could also compete with pathogenic bacteria for food and energy by consuming the nutrients required by pathogenic bacteria.33 These compounds likely play a certain antibacterial role in decreasing the species diversity of the scalp skin to achieve better evenness, which is of great significance to maintaining a steady state of resident bacteria.
The bar plot of the bacterial community at the genus level showed that the relative abundance of Staphylococcus and other skin-resident bacteria increased significantly, while the abundance of Cutibacterium, Lawsonella, Acinetobacter and Pseudomonas decreased significantly. The resident flora of the facial skin is closely related to skin health by forming a biofilm on the skin surface. Firstly, these microorganisms have a space-occupying protective effect and act as a physical barrier to prevent the invasion of exogenous pathogenic bacteria. Secondly, they can also inhibit or kill pathogenic bacteria by producing antibacterial peptides to achieve a more healthful balance.34 The bar plot of the fungal community showed the dominant fungal taxa were Basidiomycota, with the predominant genus being Malassezia, which was consistent with previous findings in Chinese individuals.29 Malassezia spp. are lipophilic yeasts that are resident in the skin of humans and other mammals. At present, 11 out of the 17 species of Malassezia are known to be involved in human skin microecology. Since the growth of most Malassezia spp. is dependent on lipids (except for M. pachydermatis), they are mainly distributed in areas rich in sebum, such as the scalp, where they account for 50–80% of the total number of colonizing fungi in healthy people.35 Most of the symptoms in the population with sensitive scalps were scalp itching and Malassezia spp. colonization of the scalp showed a positive correlation with itching.11,36,37 In the healthy scalp, changes in the abundance of Malassezia can disrupt the scalp barrier and induce keratinocytes to produce pro-inflammatory cytokines such as IL-1, IL-2, IL-4, IL-6, IFN-γ, and TNF-α, causing skin inflammation and desquamation.38 For sensitive scalp, the problem of fungus maladjustment might also disrupt the scalp barrier and result in skin inflammation. It should be noted that inflammation could eventually destroy hair follicles and the skin of the scalp, causing scalp problems such as itching, dandruff, oily hair, and hair loss.39,40 As reported previously, the ratio of M. restricta to M. globosa was observed to be significantly higher (p < 0.01) in the dandruff scalp as compared with healthy scalp.11 After treatment with SLFC, significant changes in fungus proportion (such as decrease of Malassezia) and scalp symptoms were observed (Figure 6 and Table 3), which suggested its important role on regulating the fungus composition and relieving the sensitive scalp. However, the relationship between the microbial composition and sensitive scalp should be further investigated in the future. As the postbiotic SLFC is a fermentation complex product of S. cerevisiae and L. paracasei, we found it could effectively strengthen the skin barrier in a previous study,41 which could effectively prevent the excessive colonization of Malassezia on the scalp and hair follicles, thereby combating the dysbiosis of the sensitive scalp.
Taxonomic profile analysis revealed that SLFC maintained the biodiversity of the microbial community and could regulate the proportion of microbiota at the phylum and genus level. However, little information was available on the association between target taxa with the sensitive scalp syndrome, and the absolute microbial abundance of taxa should be further elucidated to discern the underlying associations.26 Cutibacterium and Staphylococcus were predominant in the bacterial microbiome, while Malassezia was the predominant fungus in the scalp microbiome of the participants in this study. As reported previously, C. acnes, S. epidermidis, and M. restricta were the major microbial species identified on the scalp, and the ratio of C. acnes/S. epidermidis should be evaluated for obtaining the equilibrium information of microbial populations colonizing the scalp.42 Accordingly, a qPCR evaluation was carried out on the tested samples for these three characteristic species. The assessment indicated that the absolute abundance of C. acnes and S. epidermidis both increased significantly, but the ratio of C. acnes/S. epidermidis decreased, which was in accordance with the microbiome changes at the genus level after the application of SLFC for 28 days. In addition, a significant decrease in M. restricta was also observed. In a previous study, treatment with Lindera strychnifolia root extract induced a significant increase in the proportion of C. acnes and S. epidermidis in the scalp of Chinese adults, which was similar to the effect of SLFC treatment.31 M. restricta is well known for its association with a number of skin conditions, including the sensitive scalp.28,43 For example, increased M. restricta colonization in adults was found to potentially induce hair loss, as this species secretes proteases.44 Therefore, the changes in the abundance of M. restricta are of great importance for maintaining the homeostasis of the scalp, and SLFC might modulate the dysbiosis of the skin microbiome to alleviate sensitive scalp syndrome.
In addition, the symptoms of sensitive scalp syndrome were greatly alleviated according to the results of the self-assessment questionnaire. The obtained information demonstrated that postbiotic SLFC could influence the skin microbiome by modulating the composition of the bacterial and fungus (including Cutibacterium, Staphylococcus and Malassezia) community and adjusting microbiota of the sensitive scalp to a healthy balance to provide additional benefits to the participants affected by the sensitive scalp.
Few studies to date focused on the relationship between the sensitive scalp and scalp microbiota.28 The result of this work suggested that SLFC could significantly alleviate sensitive scalp symptoms, which might be attributed to the modulating effect of the postbiotic product on the skin microbiome.
Data Sharing Statement
The sequence dataset has been deposited in the NCBI Sequence Reads Archive (SRA) Database (Accession Number: PRJNA940264).
Acknowledgments
The authors gratefully acknowledge all the study participants who provided skin physiological parameters and specimens for the study.
Funding
This work was funded by JALA (Group) Co., Ltd. and Shanghai Union Studies Program from Shanghai Association for the Promotion of Science and Technology (Grant No. LM201832).
Disclosure
The authors declare no conflicts of interest related to the publication of this work.
References
1. Ma L, Guichard A, Humbert P, et al. Evaluation of the severity and triggering factors of sensitive scalp in Chinese females. J Cosmet Dermatol. 2016;15(3):219–225. doi:10.1111/jocd.12203
2. Park KH, Kim J, Oh B, et al. Evaluation of factors triggering sensitive scalp in Korean adult women. Skin Res Technol. 2019;25(6):862–866. doi:10.1111/srt.12747
3. Brenaut E, Barnetche T, Le Gall‐Ianotto C, et al. Triggering factors in sensitive skin from the worldwide patients’ point of view: a systematic literature review and meta‐analysis. J Eur Acad Dermatol Venereol. 2020;34(2):230–238. doi:10.1111/jdv.15985
4. Misery L, Rahhali N, Ambonati M, et al. Evaluation of sensitive scalp severity and symptomatology by using a new score. J Eur Acad Dermatol Venereol. 2011;25(11):1295–1298. doi:10.1111/j.1468-3083.2010.03968.x
5. Brenaut E, Misery L, Legeas C, et al. Sensitive scalp: a possible association with the use of hair conditioners. Front Med. 2021;7:596544. doi:10.3389/fmed.2020.596544
6. Fan L, Jia Y, Cui L, et al. Analysis of sensitive skin barrier function: basic indicators and sebum composition. Int J Cosmet Sci. 2018;40(2):117–126. doi:10.1111/ics.12442
7. Viodé C, Rouquier A, Mias C, et al. Specific protection of sensitive skin against environmental stress by maintenance and improvement of barrier function. J Eur Acad Dermatol Venereol. 2022;36:13–20. doi:10.1111/jdv.17910
8. Fan L, He C, Jiang L, et al. Brief analysis of causes of sensitive skin and advances in evaluation of anti‐allergic activity of cosmetic products. Int J Cosmet Sci. 2016;38(2):120–127. doi:10.1111/ics.12283
9. Talagas M, Misery L. Role of keratinocytes in sensitive skin. Front Med. 2019;6:108. doi:10.3389/fmed.2019.00108
10. Tao R, Li R, Wang R. Skin microbiome alterations in seborrheic dermatitis and dandruff: a systematic review. Exp Dermatol. 2021;30(10):1546–1553. doi:10.1111/exd.14450
11. Saxena R, Mittal P, Clavaud C, et al. Comparison of healthy and dandruff scalp microbiome reveals the role of commensals in scalp health. Front Cell Infect Microbiol. 2018;8:346. doi:10.3389/fcimb.2018.00346
12. Flowers L, Grice EA. The skin microbiota: balancing risk and reward. Cell Host Microbe. 2020;28(2):190–200. doi:10.1016/j.chom.2020.06.017
13. Ma L, Guichard A, Cheng Y, et al. Sensitive scalp is associated with excessive sebum and perturbed microbiome. J Cosmet Dermatol. 2019;18(3):922–928. doi:10.1111/jocd.12736
14. Ro BI, Dawson TL. The role of sebaceous gland activity and scalp microfloral metabolism in the etiology of seborrheic dermatitis and dandruff. J Investig Dermatol Symp Proc. 2005;10(3):194–197. doi:10.1111/j.1087-0024.2005.10104.x
15. Smith KR, Thiboutot DM. Thematic review series: skin lipids. sebaceous gland lipids: friend or foe? J Lipid Res. 2008;49:271–281. doi:10.1194/jlr.R700015-JLR200
16. Clavaud C, Jourdain R, Bar-Hen A, et al. Dandruff is associated with disequilibrium in the proportion of the major bacterial and fungal populations colonizing the scalp. PLoS One. 2013;8(3):e58203. doi:10.1371/journal.pone.0058203
17. Chen YE, Fischbach MA, Belkaid Y. Skin microbiota–host interactions. Nature. 2018;553(7689):427–436. doi:10.1038/nature25177
18. Xu Z, Wang Z, Yuan C, et al. Dandruff is associated with the conjoined interactions between host and microorganisms. Sci Rep. 2016;6(1):1–9. doi:10.1038/s41598-016-0001-8
19. Soares RC, Camargo-Penna PH, de Moraes V, et al. Dysbiotic bacterial and fungal communities not restricted to clinically affected skin sites in dandruff. Front Cell Infect Microbiol;2016:157. doi:10.3389/fcimb.2016.00157
20. Maguire M, Maguire G. The role of microbiota, and probiotics and prebiotics in skin health. Arch Dermatol Res. 2017;309(6):411–421. doi:10.1007/s00403-017-1750-3
21. Lolou V, Panayiotidis MI. Functional role of probiotics and prebiotics on skin health and disease. Fermentation. 2019;5(2):41. doi:10.3390/fermentation5020041
22. Li M, Fan A, Mao J, et al. The prebiotic effect of triple biotic technology on skin health. J Cosmet Dermatol Sci Appl. 2021;11(4):304–319. doi:10.4236/jcdsa.2021.114025
23. Jun L, Zhang Z, Zhou J, Wang Y, Xie T, Zou Y. Study on skincare properties of Saccharomyces cerevisiae Y017 isolated from Tibetan wine starter.
24. Zhou J, Jun L, Zhang Z, et al. A strain of lactobacillus isolated from Himalayan ragi vinasse and its application in skincare.
25. Saxena R, Mittal P, Clavaud C, et al. Longitudinal study of the scalp microbiome suggests coconut oil to enrich healthy scalp commensals. Sci Rep. 2021;11(1):1–14. doi:10.1038/s41598-021-86454-1
26. Iglesia S, Kononov T, Zahr AS. A multi-functional anti-aging moisturizer maintains a diverse and balanced facial skin microbiome. J Appl Microbiol. 2022;133(3):1791–1799. doi:10.1111/jam.15663
27. Grice EA, Kong HH, Conlan S, et al. Topographical and temporal diversity of the human skin microbiome. Science. 2009;324:1190–1192. doi:10.1126/science.1171700
28. Grimshaw SG, Smith AM, Arnold DS, et al. The diversity and abundance of fungi and bacteria on the healthy and dandruff affected human scalp. PLoS One. 2019;14(12):e0225796. doi:10.1371/journal.pone.0225796
29. Zheng Y, Wang Q, Ma L, et al. Alterations in the skin microbiome are associated with disease severity and treatment in the perioral zone of the skin of infants with atopic dermatitis. Eur J Clin Microbiol Infect Dis. 2019;38(9):1677–1685. doi:10.1007/s10096-019-03598-9
30. Suzuki K, Inoue M, Cho O, et al. Scalp microbiome and sebum composition in Japanese male individuals with and without androgenetic alopecia. Microorganisms. 2021;9(10):2132. doi:10.3390/microorganisms9102132
31. Filaire E, Dreux A, Boutot C, et al. Characteristics of healthy and androgenetic alopecia scalp microbiome: effect of Lindera strychnifolia roots extract as a natural solution for its modulation. Int J Cosmet Sci. 2020;42(6):615–621. doi:10.1111/ics.12657
32. Sobrino-López A, Martín-Belloso O. Use of nisin and other bacteriocins for preservation of dairy products. Int Dairy J. 2008;18(4):329–343. doi:10.1016/j.idairyj.2007.11.009
33. B XLA, Q MIN, YS C, et al. An high-throughput sequencing approach to the preliminary analysis of bacterial communities associated with changes in amino acid nitrogen, organic acid and reducing sugar contents during soy sauce fermentationScienceDirect. Food Chem. 2021;349:129131. doi:10.1016/j.foodchem.2021.129131
34. Shan W, Ying L, Lin M. Progress in skin microecology in the field of atopic dermatitis. China Med Abstr. 2016;33(2):122–127.
35. Meray Y, Gencalp D, Guran M. Putting it all together to understand the role of Malassezia spp. Dandruff Etiol. 2018;183(6):893–903.
36. Sun L, Wang X, Li X, et al. The survey of self-perceived sensitive facial/scalp skin in women in Shanghai. J Clin Dermatol. 2017;46(3):157–160.
37. Turner GA, Hoptroff M, Harding CR. Stratum corneum dysfunction in dandruff. Int J Cosmet Sci. 2012;34(4):298–306. doi:10.1111/j.1468-2494.2012.00723.x
38. Wheeler ML, Limon JJ, Underhill DM. Immunity to commensal fungi: detente and disease. Annu Rev Pathol. 2017;12(1):359–385. doi:10.1146/annurev-pathol-052016-100342
39. Saunders CW, Scheynius A, Heitman J. Malassezia fungi are specialized to live on skin and associated with dandruff, eczema, and other skin diseases. PLoS Pathog. 2012;8(6):e1002701. doi:10.1371/journal.ppat.1002701
40. Gaitanis G, Magiatis P, Hantschke M, Bassukas ID, Velegraki A. The malassezia genus in skin and systemic diseases. Clin Microbiol Rev. 2012;25(1):106–141. doi:10.1128/CMR.00021-11
41. Ren H, Tang X, Chen M. Sensitive scalp and microecologic barrier. China Surfactant Deterg Cosmet. 2020;50(9):638–642.
42. Clavaud C, Jourdain R, Bar-Hen A, et al. Dandruff is associated with disequilibrium in the proportion of the major bacterial and fungal populations colonizing the scalp. PLoS One. 2013;8(3). doi:10.1371/annotation/bcff4a59-10b7-442a-8181-12fa69209e57
43. DeAngelis YM, Gemmer CM, Kacvinsky JR, Keneally DC, Schwartz JR, Dawson TL. Three etiologic facets of dandruff and seborrheic dermatitis: Malassezia fungi, sebaceous lipids, and individual sensitivity. J Investig Dermatol Symp Proc. 2005;10(3):295–297. doi:10.1111/j.1087-0024.2005.10119.x
44. Jo JH, Deming C, Kennedy EA, Conlan S, Polley EC, Ng WI; NISC Comparative Sequencing Program. Diverse human skin fungal communities in children converge in adulthood. J Invest Dermatol. 2016;136(12):2356–2363. doi:10.1016/j.jid.2016.05.130
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.