Back to Journals » OncoTargets and Therapy » Volume 11
Effect of long non-coding RNA AOC4P on gastrointestinal stromal tumor cells
Authors Hu JC, Wang Q, Jiang LX, Cai L, Zhai HY, Yao ZW, Zhang ML, Feng Y
Received 17 May 2018
Accepted for publication 5 July 2018
Published 26 September 2018 Volume 2018:11 Pages 6259—6269
DOI https://doi.org/10.2147/OTT.S174524
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
This paper has been retracted.
Jin-Chen Hu, 1,* Quan Wang, 2,* Li-Xin Jiang, 1 Li Cai, 3 Hui-Yuan Zhai, 1 Zeng-Wu Yao, 1 Meng-Lai Zhang, 1 Ye Feng 4
1Department of Gastrointestinal Surgery, The Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, China; 2Department of Gastrointestinal and Anal Surgery, The First Hospital of Jilin University, Changchun, China; 3Department of Pathology, The Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, China; 4Department of Gastrointestinal Colorectal and Anal Surgery, China-Japan Union Hospital of Jilin University, Changchun, China
*These authors contributed equally to this work
Objective: In this research, we explored the effect of long non-coding RNA (lncRNA) AOC4P on gastrointestinal stromal tumor (GIST) cells.
Materials and methods: The expression of lncRNA AOC4P in tissues was detected by real-time PCR (RT-PCR). The epithelial–mesenchymal transition (EMT)-related proteins in tissues were analyzed by Western blot. The experiment included negative control group (CN), silence AOC4P group (si AOC4P), and silence negative control group (si CT). RT-PCR, MTT, Scratch, Transwell, and Annexin V-FITC methods were used to detect the expression of lncRNA AOC4P, cell proliferation, cell migration ability, cell invasion ability, and apoptosis, respectively. The EMT-related proteins including TGF-β, ZEB1, Vimentin, Snail, and E-cadherin were analyzed by Western blot.
Results: The expression of lncRNA AOC4P and the expression of EMT-related proteins in high-risk GISTs were higher than that in low- and intermediate-risk GISTs (P< 0.05). It was revealed that cell proliferative migration and invasive ability in si AOC4P group was decreased than that in CN and si CT groups (P< 0.05), and cell apoptosis in si AOC4P group was higher than that in si CT group. The results of Western blot demonstrated that the expression of TGF-β 1, ZEB1, Vimentin, and Snail in si AOC4P group were lower than that in si CT and CN group (P< 0.05), and the expression of E-cadherin in si AOC4P group was higher than that in si CT and CN group (P< 0.05).
Keywords: AOC4P, epithelial–mesenchymal transition, gastrointestinal stromal tumors, long non-coding RNA
Introduction
Gastrointestinal stromal tumor (GIST) was the most common mesenchymal tissue tumors in the alimentary canal.1,2 It was demonstrated that the mortality of GIST was high in human beings, and it mostly occurred in old-aged patients.2 The clinical behaviors of GIST patients include gastrointestinal hemorrhage, enclosed mass in abdomen, and pain of belly, associated with the primary tissues and lump growth.3–5 In addition, unceasing reports have demonstrated that surgical resection for localized primary GIST was the curative mainstay of therapy.6–8 Nevertheless, it still faces great challenges that the risk of recurrence is as high as ~40% and the clinicopathologic features vary greatly.9 Therefore, it is of great significance to find an effective method for the treatment of GIST.
In recent years, long non-coding RNA (lncRNA) has attracted great attention from researchers.10,11 It was demonstrated that the abnormal expression of lncRNA was greatly associated with the occurrence of malignant tumors. The results suggested that lncRNA which participated in epigenetic regulation could regulate cell proliferation, apoptosis, infiltration, and transformation, thereby controlling and regulating tumors. Some lncRNAs including metastasis-associated lung adenocarcinoma transcript 1, highly upregulated in liver cancer, H19, and HOX antisense intergenic RNA, were found by high-throughput screening method, and numerous investigations pointed that lncRNA has the potential to be the therapeutic target of tumors.12
In previous studies, researchers had investigated the differences of lncRNA in gastric carcinoma including GIST and the normal tissues, and five of lncRNAs (TINCR, CCAT2, AOC4P, BANCR, and LINC00857) were significantly elevated in gastric carcinoma.13 TINCR, CCAT2, and BANCR in gastric carcinoma have been documented in previous studies.14–16 These results demonstrated that TINCR upregulates the expression of SP1, followed by mediating the expression of KLF2 mRNA to regulate cell proliferation and apoptosis. It is also found that CCAT2 is associated with lymph gland and distant metastasis and that NF-KB as the unfavorable prognosis symbol affects cell apoptosis through NF-KB pathway.17 It has also been documented that lncRNA containing amine oxidase copper containing four pseudogenes (AOC4P) can inhibit the transformation of hepatoma carcinoma cells through the degradation of the vimentin.18,19 In addition, the investigations have demonstrated that the AOC4P inhibits the degradation of UHRF1 and promotes the occurrence of colorectal carcinoma.19 Therefore, AOC4P as one of lncRNAs exerts important effects on tumors. Epithelial–mesenchymal transition (EMT) is the morphological transformation of epithelial-like cancer cells to an elongated mesenchymal phenotype.20 During EMT, cancer cells stop expressing the adhesion proteins, including epithelial E-cadherin, and increase the expression of mesenchymal phenotype markers, including vimentin and Snail.21 Currently, the roles of lncRNA AOC4P influencing GIST cells to undergo EMT is not widely known or studied.
The targeted therapies of GIST were tyrosine kinase KIT and PDGFRA.22–24 In 2003, Heinrich et al25 further elaborated the role of PDGFRA in the pathogenesis of GISTs that were negative for the expression of c-KIT. These research studies demonstrated that GISTs were predominantly c-KIT or PDGFRA-associated mesenchymal tumors. However, the research of lncRNA AOC4P on GIST is yet to be reported. In this study, we explored the effects of long non-coding RNA AOC4P on GIST to better understand the potential mechanism of GIST therapies.
Materials and methods
Patients and specimens
Fresh GIST samples were obtained from 79 GIST patients between 2012 and 2016. The size of the GISTs ranged from 3 to 17 cm. The medians of the size in high-risk GISTs and low/medium-risk GISTs were 7.25 and 6.10 cm, respectively. All procedures in this study were performed under the regulation of ethical issues in hospital. All participants signed written informed consent form for this study. The study protocol was approved by the Institutional Review Board of China-Japan Union Hospital of Jilin University.
Cells and siRNA transfection
GIST-T1 (heterozygosis mutation type of KIT exon) and GIST-882 (homozygotic mutation type of KIT exon) cells were purchased from Cosmo Bio (Tokyo, Japan) and cultured in DMEM medium (Hyclone, USA) containing 10% fetal bovine serum and 1% penicillin streptomycin (Gibco, USA) at 37°C incubator (Forma, USA) containing 5% of CO2.
GIST-T1 and GIST-882 cells were inoculated into a six-well culture plate in a 37°C incubator overnight. The experiment had three groups to test the effect of AOC4P on GIST including negative control group (CN), silence AOC4P group (si AOC4P), and silence negative control group (si CT). Lipidosome2000 (Amersham Bioscience, Piscataway, NJ, USA) was used to transfect 1 μg si AOC4P and 1 μg si RNA in cells as si AOC4P and si CT group, respectively. The sequences of si-AOC4P are as follows: sense: 5′-GGUAAGGCAUCCUGGCAGAUU-3′ and antisense: 5′-CCAGAGGUAUGGAAACAAAUU-3′.
Quantitative real-time PCR (qRT-PCR) analysis of lnc AOC4P in tissues
The total RNA of GISTs was extracted using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA). About 2 μg of total RNA was reverse transcribed by Superscript™ II according to the manufacturer’s protocol (Thermo Fisher Scientific). Real-time quantitative analysis was performed in the Real-Time Detection System (Thermo Fisher Scientific) by SYBR green I dye detection (Takara, Tokyo, Japan). The primers used in this study are as follows:
lnc AOC4P forward, 5′-AAAGGAGGTGAGAGGGAATGT-3′;
lnc AOC4P reverse, 5′-GCTGGGCACTGGGAGATAC-3′;
U1 forward, 5′-ATACTTACCTGGCAGGGGAG-3′;
U1 reverse, 5′-CAGGGGGAAAGCGCGAACGCA-3′.
The PCR amplifications were performed at 95°C for 3 minutes, followed by 35 cycles of thermal cycling at 95°C for 30 seconds, at 60°C for 15 seconds, and at 72°C for 15 seconds, terminated at 60°C for 15 seconds, and finally stored at 4°C. U1 was used as endogenous control to normalize the differences. Melt curves were performed to complete the whole cycles to ensure that nonspecific products were avoided. Roche LightCycler 480 qRT-PCR (Hoffman-La Roche Ltd., Basel, Switzerland) quantification was performed by normalizing the cycle threshold (Ct) values with U1 Ct and analyzed with the 2−ΔΔCt method.
Western blot analysis of the related EMT proteins in tissues
Total protein of GISTs was extracted by using the Tissue Total Protein Lysis buffer (Sangon Biotech Co., Ltd., Shanghai, China) according to the manufacturer’s protocol. Protein samples (40 μg) were separated on SDS–PAGE gel and transferred to polyvinylidene fluoride nitrocellulose blots. The blots blocked with 5% skim milk for 1 hour were incubated overnight at 4°C with rabbit polyclonal antibody TGF-β1 (1:1,000, orb7087; Biorbyt), rabbit polyclonal antibody ZEB1 (1:1,000, orb254960; Biorbyt, Cambridge, UK), rabbit polyclonal antibody E-cadherin (1:1,000, orb160843; Biorbyt), rabbit polyclonal antibody Vimentin (1:200, orb128909; Biorbyt), and rabbit polyclonal antibody Snail (1:1,000, orb95764; Biorbyt) and washed three times for 5 minutes with TBST (TBS, 1 mL/L Tween 20). Then, these blots were injected with HRP secondary antibodies goat anti rabbit IgG (1:8,000, orb108261; Biorbyt), incubated for 2 hours in dark, and then washed three times for 5 minutes with TBST, and the film was developed in the enhanced chemiluminescence (ECL) in dark room. The expression of protein samples was standardized according to β-actin (GE Healthcare, Piscataway, NJ, USA), then scanned and quantified by the ImageJ software.
Proliferative activity assay
The proliferative activity was measured by MTT method. The transfected cells were prepared for single-cell suspensions and adjusted to 1.0×104/mL. Then, the cell suspensions were inoculated into 96-well plates, in which each of the plates was injected with 100 μL cell suspensions. Then, the 96-well plates were incubated in 37°C incubator containing 5% CO2. The cell suspensions were incubated for 24, 48, 72, and 96 hours, injected with 20 μL MTT (Promega Corporation, Fitchburg, WI, USA) in each well of 96-well plates, and incubated for 3 hours. The results of OD490 were measured and read by Microplate Reader.
Scratch test
Cells in each group were digested with 0.25% trypsin-EDTA (Corning Incorporated, Corning, NY, USA) and made into single cell suspension. The cells were re-inoculated in six-well plates (3×105/well) for 24 hours. Under an aseptic environment, a vertical downward scratch was quickly prepared on the monolayer by the tip of the 10 μL pipette. The medium and cell debris should be removed carefully. The serum-free medium was then added slowly. Under the inverted microscope (10×), images were taken at 0 and 24 hours.
Transwell invasion assay
About 500 μL FBS-free DMEM was added into the chambers (Corning Incorporated) in a 37°C incubator for 2 hours, and the cells were then transfected for 24 hours. FBS-free DMEM was used to suspend the cells, and the cells were adjusted to 25×104/mL. Subsequently, 200 μL of cell suspensions was seeded in the upper chambers of the wells, and the lower chambers were filled with 600 μL DMEM medium containing 10% of FBS to induce cell migration. After incubation for 24 hours, the cells on the filter surface were fixed with 4% formaldehyde, stained with 0.1% crystal violet for 15 minutes, and examined under a microscope. Cells in at least five random microscopic fields (100×) were counted.
Apoptosis assay
Annexin V-FITC/PI double staining was used to detect cell apoptosis. Parietal cells were digested by using 0.25% trypsin. The cell suspension was collected and then centrifuged at 2,000 rpm for 5–10 minutes. Cells were overhung by precooled PBS and centrifuged at 2,000 rpm for 5–10 minutes. Then, 300 μL 1× binding buffer and 5 μL Annexin V-FITC (Beyotime Biotechnology Research Institute) were added and mixed at room temperature at dark incubation for 10 minutes. Add 5 μL PI dye and 200 μL 1× binding buffer, and they were mixed evenly to avoid light at 4°C. Cell apoptosis was detected by flow cytometry (Beckman, Brea, California, USA).
Western blot
Cells were washed by PBS, transfected twice, and added into 200 μL RIPA for lysis (50 mM Tris–HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 1% sodium deoxycholate, and 0.1% SDS). The remaining steps are the same as the tissue detection.
Statistical analysis
All the data were presented as mean ± standard deviation (SD). Data were analyzed with SPSS 19.0 software. Multiple comparisons were evaluated by repeated measures analysis of variance followed by the least significant difference-t test or Kruskal–Wallis test. P<0.05 was considered to be statistically significant.
Results
LncRNA AOC4P was highly expressed in GIST tissues
A total of 79 GIST patients were included with 39 low-risk cases, 14 medium-risk cases, and 26 high-risk cases. At the same time, 79 cases of paracancerous normal tissues were taken. As shown in Figure 1, the expression of AOC4P in GIST tissues was higher than that in normal tissues (P<0.05). The expression of AOC4P in high risk GIST tissues was higher than that in low/medium-risk GIST tissues (P<0.05).
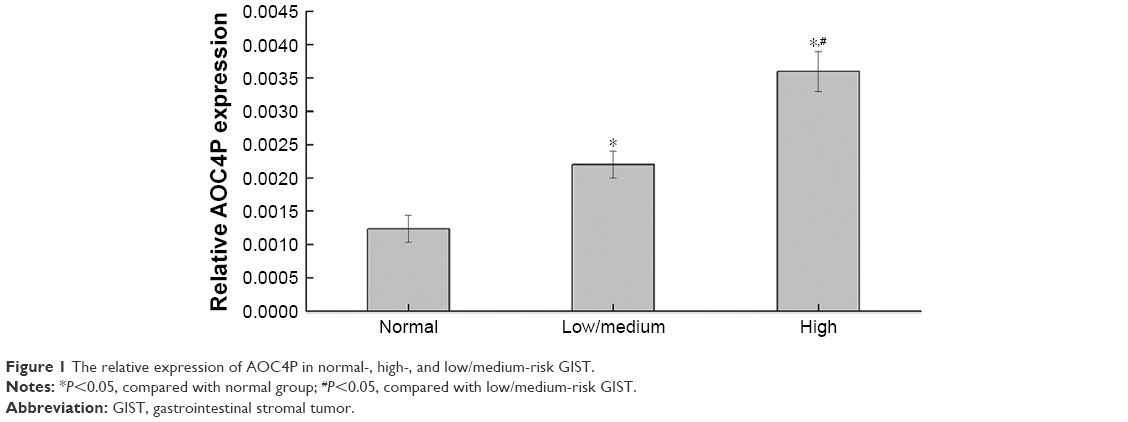 | Figure 1 The relative expression of AOC4P in normal-, high-, and low/medium-risk GIST. |
The expression of EMT-related proteins in GIST patients
As shown in Figure 2, the expression of TGF-β1, ZEB1, Vimentin, and Snail in normal tissues were lower than that in GIST tissues (P<0.05), and the expression of E-cadherin in normal tissues was higher than that in GIST tissues (P<0.05). Compared with high-risk GIST, the expression of TGF-β1, ZEB1, Vimentin, and Snail were significantly decreased in low/medium-risk GIST, while the expression of E-cadherin has significantly increased in low/medium-risk GIST.
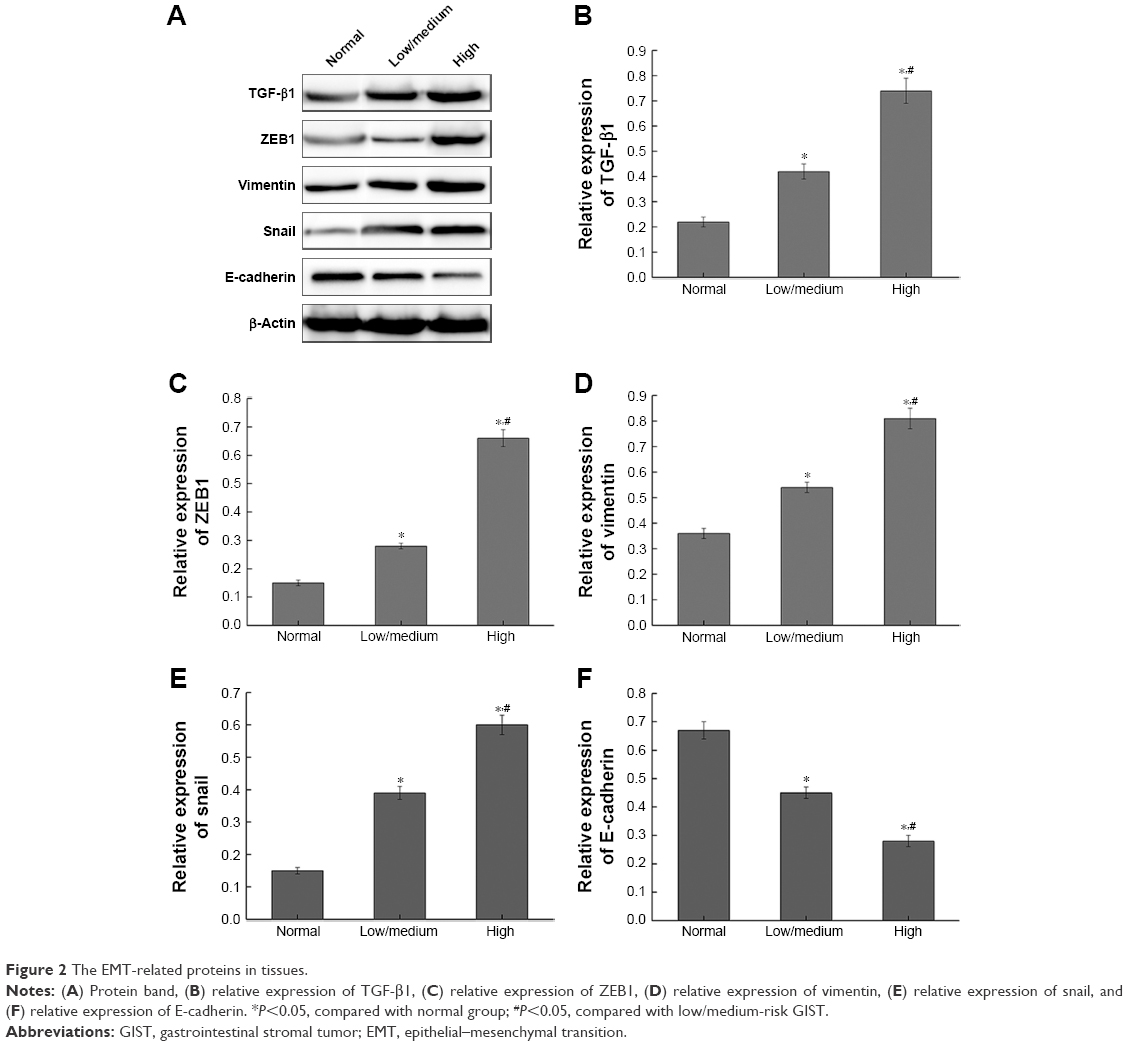 | Figure 2 The EMT-related proteins in tissues. |
Silence of AOC4P inhibited cell proliferation of GIST
As shown in Figure 3A, GIST cells in si AOC4P group were decreased by 60% compared to the si CT group (P<0.05). Simultaneously, cell proliferation in si AOC4P group was significantly attenuated than in the CN group and si CT group, and si-AOC4P group showed a significant difference from si CT group at 72 and 96 hours (P<0.05, Figure 3B). In addition, the expression of si AOC4P in GIST-T1 cells were consistent with GIST882 cells, demonstrating that si AOC4P can inhibit cell proliferation of GIST.
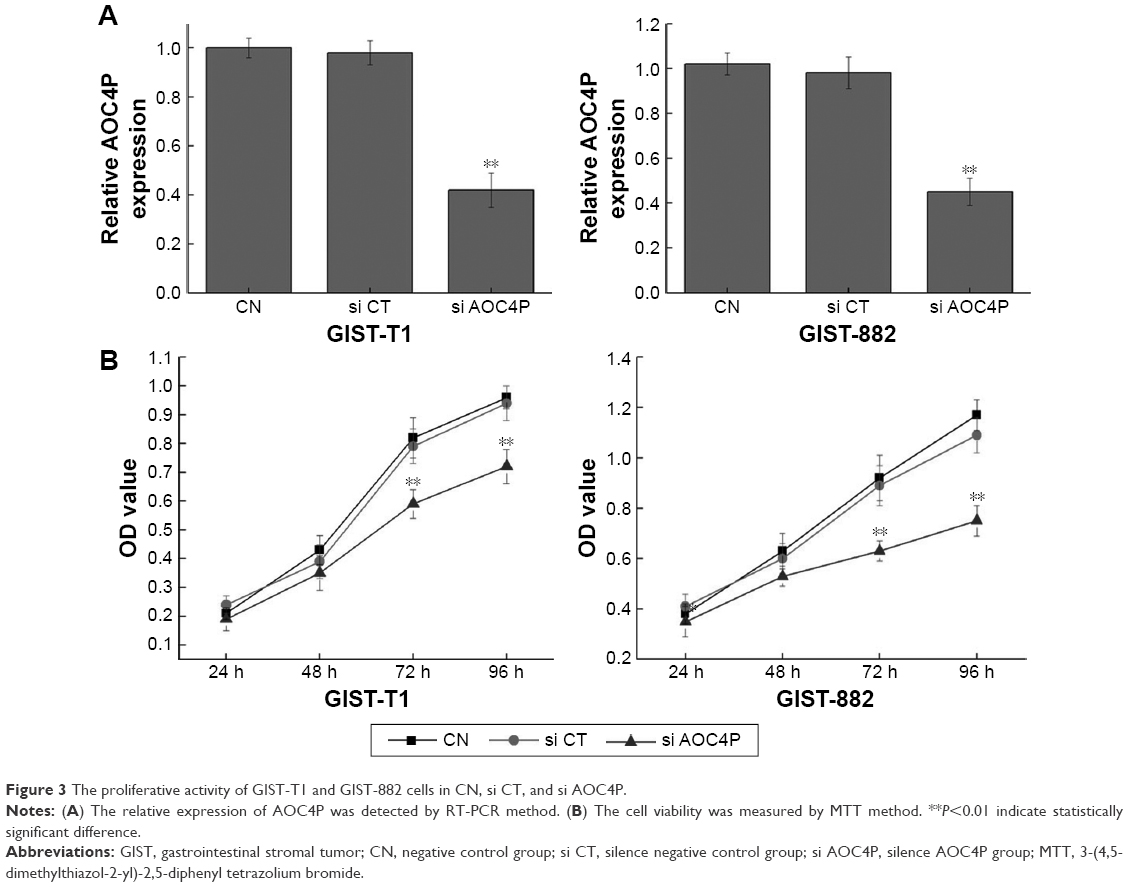 | Figure 3 The proliferative activity of GIST-T1 and GIST-882 cells in CN, si CT, and si AOC4P. |
Silence of AOC4P reduced cell migration ability
As shown in Figure 4, the migration ability in si AOC4P group was significantly decreased than in si CT group (P<0.05). There were no significant differences in si CT group and CN group.
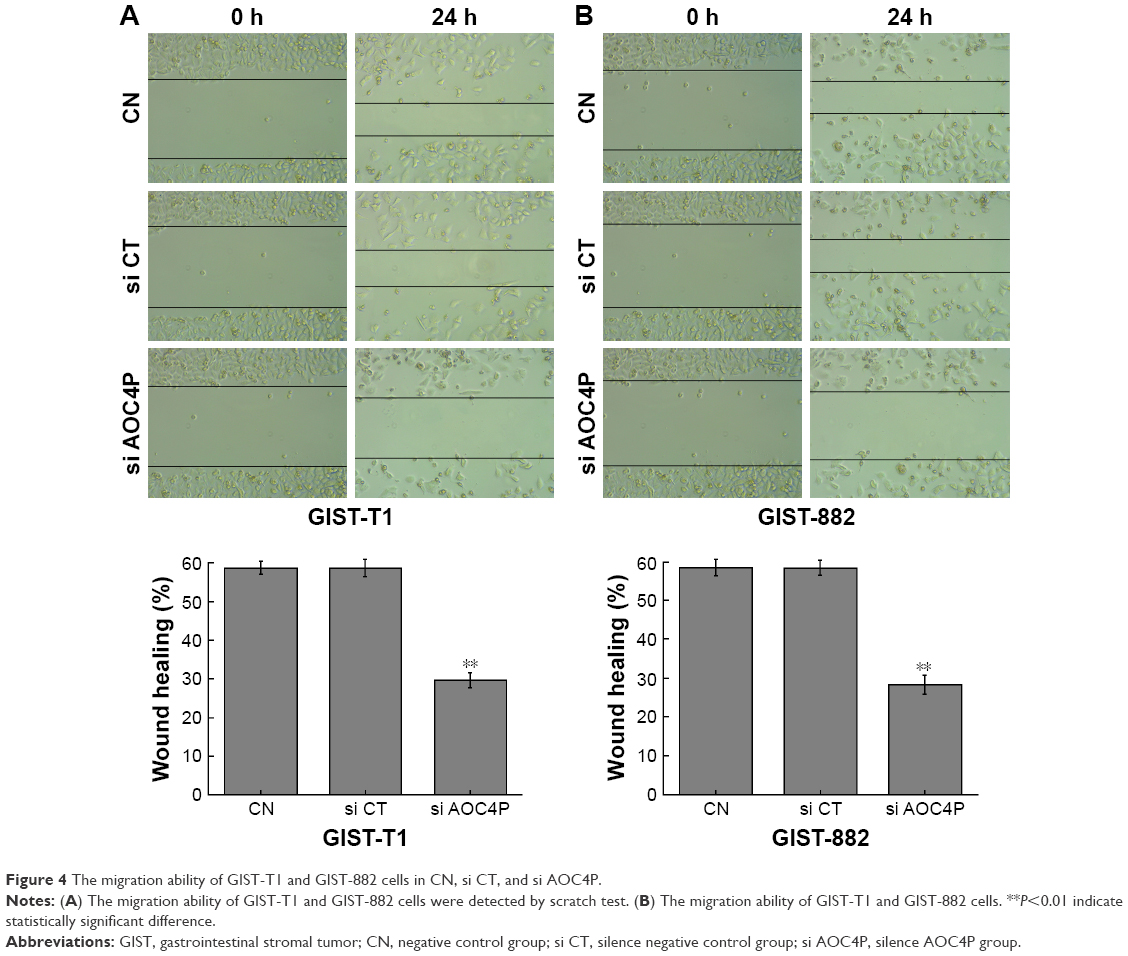 | Figure 4 The migration ability of GIST-T1 and GIST-882 cells in CN, si CT, and si AOC4P. |
Silence of AOC4P reduced cell invasive ability
The results in Figure 5A and B demonstrated that the invasive ability in si AOC4P group was significantly decreased than that in si CT group (P<0.05). Furthermore, the results in GIST-T1 cells were consistent with GIST882 cells, demonstrating that si AOC4P can reduce the invasive ability of GIST. There were no significant differences in cell invasive ability between si CT group and CN group.
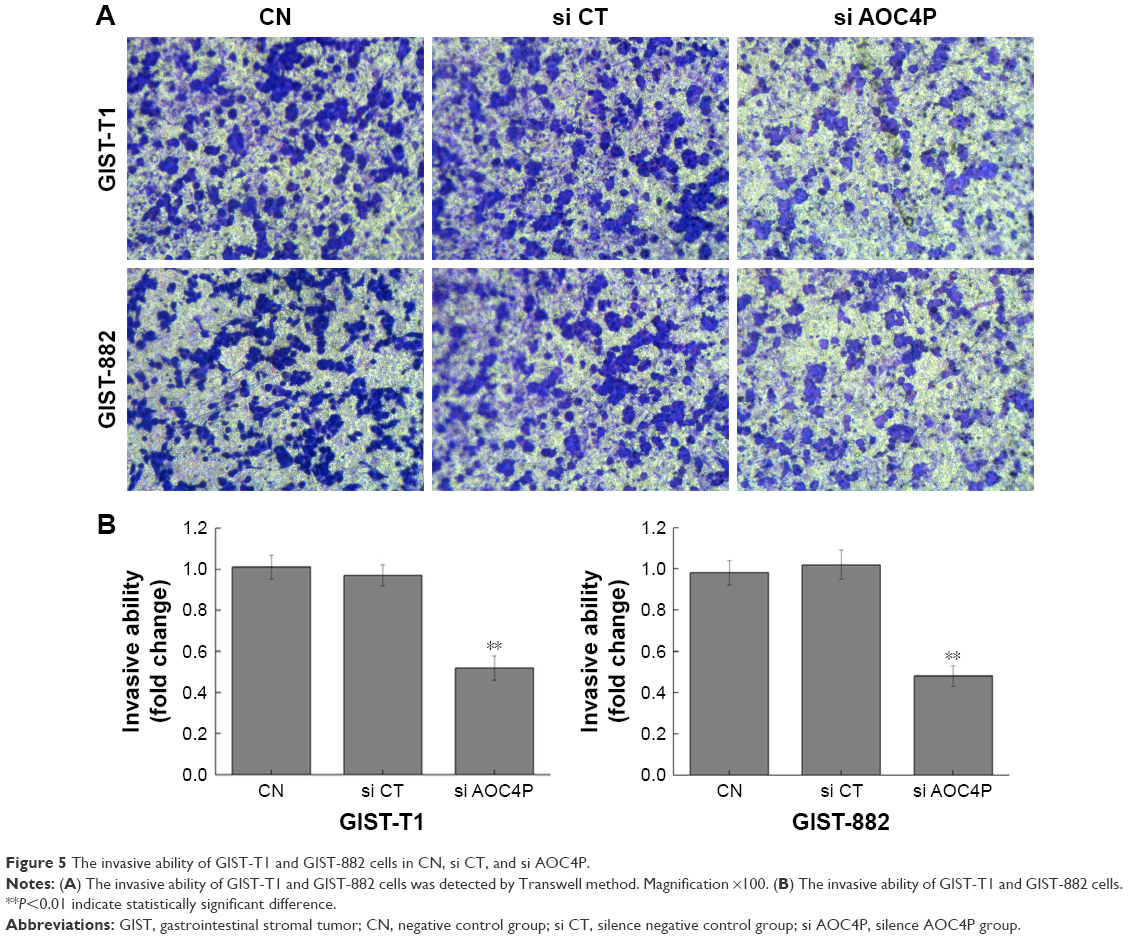 | Figure 5 The invasive ability of GIST-T1 and GIST-882 cells in CN, si CT, and si AOC4P. |
Silence of AOC4P induced cell apoptosis
By Annexin V-FITC staining, it was found that si RNA AOC4P in si AOC4P group induced cell apoptosis compared with the si CT group (Figure 6A and B; P<0.05). Consistently, cell apoptosis rates of GIST-T1 and GIST-882 cells in si AOC4P group were 21.85% and 23.72%, respectively, revealing that cell apoptosis rate in si AOC4P group was higher than that in si CT group (P<0.05).
 | Figure 6 The apoptosis activity of GIST-T1 and GIST-882 cells in CN, si CT, and si AOC4P. |
Effect of AOC4P for GIST on EMT
There was no significant difference in protein expression between si CT and CN groups. Simultaneously, the results of GIST-T1 and GIST-882 cells were consistent. As shown in Figure 7, the results of Western blot demonstrated that the expression of TGF-β1, Vimentin, ZEB1, and Snail in si AOC4P group were lower than that in si CT and CN groups (P<0.05), and E-cadherin in si AOC4P group has increased compared with si CT and CN groups (P<0.05).
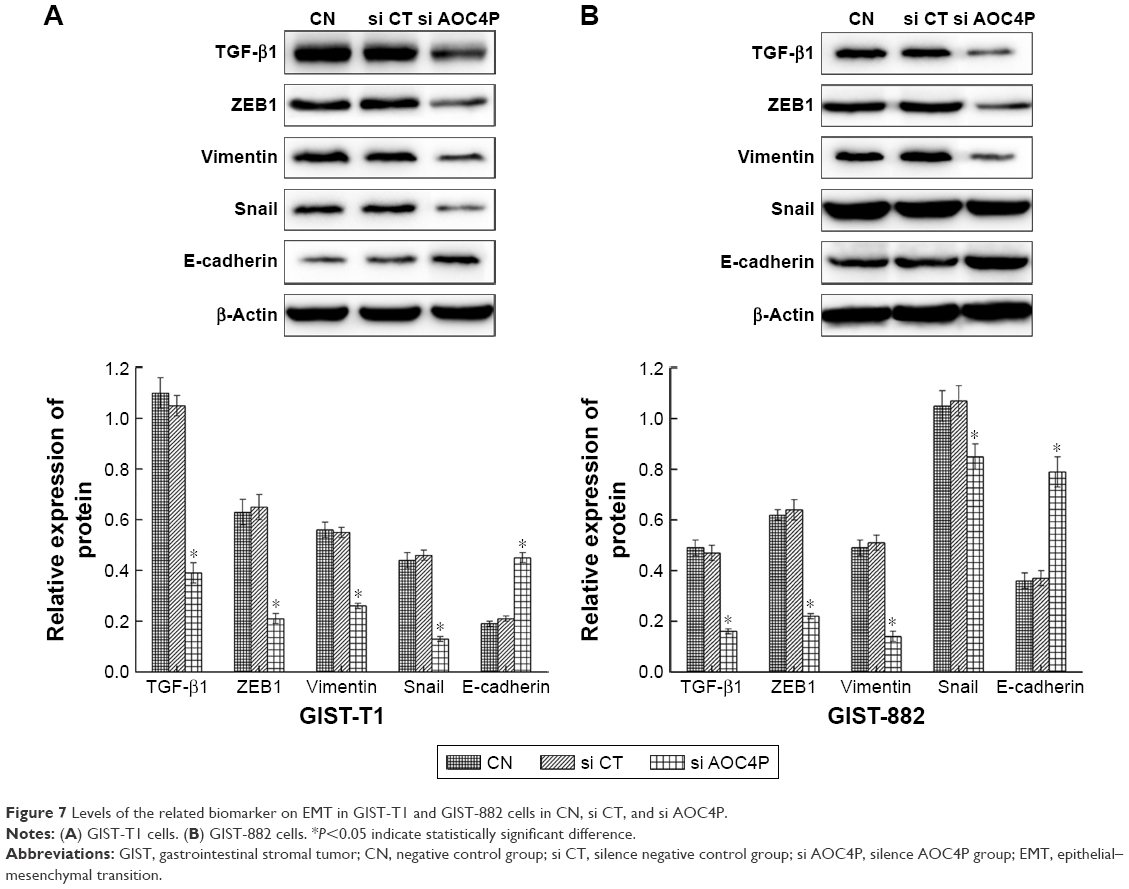 | Figure 7 Levels of the related biomarker on EMT in GIST-T1 and GIST-882 cells in CN, si CT, and si AOC4P. |
Discussion
GIST is defined as a malignantly potential gastroenteric tumor, causing impairments to patients.26,27 Therefore, the therapies to cure GIST have received great attention. Based on investigations, it was illustrated that lncRNA showed potential values in clinical application.28,29 The predecessor pointed that lncRNA AOC4P has inhibited the migration of hepatoma carcinoma cell,19 indicating that AOC4P is an effective diagnosis for tumors. Therefore, in our investigation, we explored the association of AOC4P in GIST, and the results demonstrated that lncRNA AOC4P was highly expressed in GIST patients. Briefly, the expression of lncRNA AOC4P in GIST cells was increased than in normal cells. Our research is consistent with the previous research in 2017,13 indicating that AOC4P is essential for the survival and tumorigenicity of colorectal cancer cells by regulating protein degradation and ubiquitination.
It has been revealed that lncRNA participates in epigenetic regulation, thereby regulating the growth, apoptosis, infiltration, and transformation of tumor cells. To get an insight on the mechanism of AOC4P on GIST, we examined the cell proliferation, migration ability, invasive ability, and apoptosis activity, demonstrating that si AOC4P can inhibit cell proliferation, reduce migration and invasive ability, and promote cell apoptosis. Our data illustrated that silence of lncRNA AOC4P resulted in the decrease of cell proliferation, promoting apoptosis, and reduction of migration and invasive ability. Similar to the function of protein coding gene, lncRNAs can be divided into cancer-promoting lncRNA and cancer-suppressor lncRNA. Based on the investigation, we found that AOC4P was a cancer-promoting lncRNA. This will provide powerful evidence that AOC4P has striking effects on GIST patients.
Although investigations on the association of GIST and lncRNA are sporadic, GIST as an alimentary canal disease is similar to colorectal cancer. Therefore, lncRNA can be used as the marker of diagnosis or prognosis for GIST. Furthermore, lncRNA AOC4P participates in tumor metastasis by inducing EMT. Therefore, our research illustrated the effect of lncRNA AOC4P for GIST on EMT. EMT is characterized as the process of oxophile epithelial cells converting to mesenchymal cells, inducing a series of changes including cell morphology, molecular features, and cell functions.30 EMT enhanced the ability of invasion and metastasis and promoted the growth of tumors.31 The TGF-β signaling pathway plays critical roles in multiple cancer biological processes, including growth, migration, invasion, differentiation, apoptosis, stemness, angiogenesis, and modification of the microenvironment, which can regulate EMT.32,33 Based on the features of EMT, we have explored the effects of si AOC4P on the related makers of EMT, demonstrating that GIST can influence the related factors of EMT.
In this research, we found that silence of AOC4P induced the increase of E-cadherin. It is well known that E-cadherin members develop the connection through immunoglobulin domains, connect with actin, and form the stable intercellular touch.34 Thus, reducing E-cadherin in GIST patients caused decrease of adhesive ability, chromosome elimination, and gene mutation, thereby exacerbating GIST. In addition, the decrease of cell adhesion molecule E-cadherin led to cytokeratin converting into vimentin which was similar to the mesenchymal cell. Vimentin, a major component of the cytoskeleton, is a well-known metastasis marker and a therapeutic target for overexpression in mesenchymal cells and reduces the ability of cells migration.35 In addition, vimentin, one of the markers of EMT, has the functions of cell signal transduction, adhesion, migration, and apoptosis. Our study revealed that the expression of vimentin in si AOC4P group was lower than that in si CT group, demonstrating that lncRNA AOC4P regulates EMT by affecting the production of vimentin. In addition, Snail is one of DNA binding proteins that recognizes and integrates with E-box, thereby inhibiting the expression of E-cadherin.36 In this research, silencing AOC4P could attenuate the expression of Snail in GIST cells, indicating that the expression of E-cadherin was enhanced and the proliferation of tumor was inhibited. Furthermore, Snail can induce loss of cell polarity, enhance cell move ability, and promote metastasis ability, and thus promote tumor invasion. Similar to Snail, ZEB, another important transcription factor, plays a striking effect on the occurrence and development of tumors and promotes tumorigenicity by regulating cell adhesion and microenvironment. It has been documented that Neddle4L regulated by ZEB-1 is a downstream of miR-23a pathway and has activated TGF-β pathway, thus exacerbating the tumors.37 Briefly, silence of AOC4P suppressed the migration of GIST by reducing the production of vimentin and Snail and enhancing the expression of E-cadherin.
Taken together, the administration of lncRNA AOC4P in GIST supports a role in the mechanism of cell proliferative ability, invasion ability, and apoptosis through EMT. At present, tumor size, cell proliferative ability, macroscopic vascular invasion, and cell apoptosis are considered as essential predictors of tumor recurrence. Our research suggested that GIST patients with high expression levels of AOC4P significantly affected EMT factors including vimentin, ZEB1, Snail, and E-cadherin. Our finding supported the concept that the expression level of AOC4P could serve as a prognostic biomarker for GIST.
Conclusion
lncRNA AOC4P silencing can reduce the proliferative ability, decrease the migration and invasion activity, and induce cell apoptosis. These findings added an insight into our understanding of EMT in GIST. Along with further research, we identified that lncRNA AOC4P promoted GIST growth by inducing EMT for better understanding of the molecular mechanisms of lncRNA AOC4P. Most importantly, these studies suggested that lncRNA AOC4P may become a useful and potential prognostic biomarker, as well as an effective target for antimetastasis therapies for GIST. Moreover, the expression level of lncRNA AOC4P in GIST could be applied to predict the risk of disease recurrence and provide an effective idea of therapeutic target. However, no studies address the phenotype changes found in this study and the cells tend to become epithelial-like; therefore, the action of AOC4P lays more toward mesenchymal-to-epithelial transition, rather than EMT. These speculations should be tested in future experiments.
Acknowledgment
This work was supported by Projects of Medical and Health Technology Development Program of Shandong Province (2015WSA06006) and Science and Technology Development Plan Project of Yantai (2015WS016, 2015WS017).
Disclosure
The authors report no conflicts of interest in this work.
References
de Vogelaere K, Aerts M, Haentjens P, de Grève J, Delvaux G. Gastrointestinal stromal tumor of the stomach: progresses in diagnosis and treatment. Acta Gastroenterol Belg. 2013;76(4):403–406. | ||
Comandone A, Boglione A. The importance of mutational status in prognosis and therapy of GIST. Recenti Prog Med. 2015;106(1):17–22. | ||
Fujita K, Kobara H, Mori H, et al. Differences in miRNA expression profiles between GIST and leiomyoma in human samples acquired by submucosal tunneling biopsy. Endosc Int Open. 2015;3(6):E665–E671. | ||
Mehta C, Gumaste VV, Leytin A, Walfish A. An unusual cause of upper gastrointestinal bleeding: duodenal GIST. A case report and literature review. Acta Gastroenterol Belg. 2011;74(2):347–351. | ||
Tiberi L, Faisal A, Rossi M, di Tella L, Franceschi C, Salvioli S. p66(Shc) gene has a pro-apoptotic role in human cell lines and it is activated by a p53-independent pathway. Biochem Biophys Res Commun. 2006;342(2):503–508. | ||
Euanorasetr C. Outcomes and prognostic factors of primary gastric GIST following complete surgical resection: a single surgeon experience. J Med Assoc Thai. 2011;94(1):55–64. | ||
Nie J, Wu Q, Liu W, et al. Ectopic expression of Ligand-of-Numb protein X promoted TGF-beta induced epithelial to mesenchymal transition of proximal tubular epithelial cells. Biochim Biophys Acta. 2009;1792(2):122–131. | ||
Zhang Y, Kwok JS, Choi PW, et al. Pinin interacts with C-terminal binding proteins for RNA alternative splicing and epithelial cell identity of human ovarian cancer cells. Oncotarget. 2016;7(10):11397–11411. | ||
Wozniak A, Rutkowski P, Schöffski P, et al. Tumor genotype is an independent prognostic factor in primary gastrointestinal stromal tumors of gastric origin: a European multicenter analysis based on ConticaGIST. Clin Cancer Res. 2014;20(23):6105–6116. | ||
Nohata N, Abba MC, Gutkind JS. Unraveling the oral cancer lncRNAome: identification of novel lncRNAs associated with malignant progression and HPV infection. Oral Oncol. 2016;59:58–66. | ||
Wang P, Ning S, Zhang Y, et al. Identification of lncRNA-associated competing triplets reveals global patterns and prognostic markers for cancer. Nucleic Acids Res. 2015;43(7):3478–3489. | ||
Zhang H, Guo Y, Song Y, Shang C. Long noncoding RNA GAS5 inhibits malignant proliferation and chemotherapy resistance to doxorubicin in bladder transitional cell carcinoma. Cancer Chemother Pharmacol. 2017;79(1):49–55. | ||
Zhang K, Shi H, Xi H, et al. Genome-wide lncRNA microarray profiling identifies novel circulating lncRNAs for detection of gastric cancer. Theranostics. 2017;7(1):213–227. | ||
Taniue K, Kurimoto A, Sugimasa H, et al. Long noncoding RNA UPAT promotes colon tumorigenesis by inhibiting degradation of UHRF1. Proc Natl Acad Sci U S A. 2016;113(5):1273–1278. | ||
Kogure T, Yan IK, Lin WL, Patel T. Extracellular vesicle-mediated transfer of a novel long noncoding RNA TUC339: a mechanism of intercellular signaling in human hepatocellular cancer. Genes Cancer. 2013;4(7–8):261–272. | ||
Wang L, He Y, Liu W, et al. Non-coding RNA LINC00857 is predictive of poor patient survival and promotes tumor progression via cell cycle regulation in lung cancer. Oncotarget. 2016;7(10):11487–11499. | ||
Xu TP, Liu XX, Xia R, et al. SP1-induced upregulation of the long noncoding RNA TINCR regulates cell proliferation and apoptosis by affecting KLF2 mRNA stability in gastric cancer. Oncogene. 2015;34(45):5648–5661. | ||
Liu Q, Hu X, Zhang X, et al. The TMSB4 pseudogene lncRNA functions as a competing endogenous RNA to promote cartilage degradation in human osteoarthritis. Mol Ther. 2016;24(10):1726–1733. | ||
Wang TH, Lin YS, Chen Y, et al. Long non-coding RNA AOC4P suppresses hepatocellular carcinoma metastasis by enhancing vimentin degradation and inhibiting epithelial-mesenchymal transition. Oncotarget. 2015;6(27):23342–23357. | ||
Kalluri R. EMT: when epithelial cells decide to become mesenchymal-like cells. J Clin Invest. 2009;119(6):1417–1419. | ||
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. | ||
Emile JF, Brahimi S, Coindre JM, et al. Frequencies of KIT and PDGFRA mutations in the MolecGIST prospective population-based study differ from those of advanced GISTs. Med Oncol. 2012;29(3):1765–1772. | ||
Valsangkar N, Sehdev A, Misra S, Zimmers TA, O’Neil BH, Koniaris LG. Current management of gastrointestinal stromal tumors: surgery, current biomarkers, mutations, and therapy. Surgery. 2015;158(5):1149–1164. | ||
Pantaleo MA, Saponara M, Nannini M, Biasco G. Targeted therapies in solid tumours: results and promises. Minerva Med. 2007;98(5):603–623. | ||
Heinrich MC, Corless CL, Duensing A, et al. PDGFRA activating mutations in gastrointestinal stromal tumors. Science. 2003;299(5607):708–710. | ||
Vassos N, Agaimy A, Hohenberger W, Croner RS. Management of liver metastases of gastrointestinal stromal tumors (GIST). Ann Hepatol. 2015;14(4):531–539. | ||
Grecu F, Lapuşneanu A, Ferariu D, et al. Primele rezultate de management chirurgical de gastro-intestinale stromal tumorile (miez)–experienţa de unitate 3 chirurgicale [Early results of the surgical management of gastrointestinal stromal tumors (gist) – the experience of the 3rd surgical unit]. Rev Med Chir Soc Med Nat Iasi. 2012;116(2):506–514. Romanian. | ||
Gao Y, Meng H, Liu S, et al. LncRNA-HOST2 regulates cell biological behaviors in epithelial ovarian cancer through a mechanism involving microRNA let-7b. Hum Mol Genet. 2015;24(3):841–852. | ||
Volders PJ, Verheggen K, Menschaert G, et al. An update on LNCipedia: a database for annotated human lncRNA sequences. Nucleic Acids Res. 2015;43(8):4363–4364. | ||
Sun Y, Li SS, Wang XH, Wang XJ, Yan AH. [Transforming growth factor beta1 regulation of epithelial-mesenchymal transition in esophagus squamous cell carcinoma]. Zhonghua Bing Li Xue Za Zhi. 2008;37(8):542–548. Chinese. | ||
Fujiwara A, Shintani Y, Funaki S, et al. Pirfenidone plays a biphasic role in inhibition of epithelial-mesenchymal transition in non-small cell lung cancer. Lung Cancer. 2017;106:8–16. | ||
Bellomo C, Caja L, Moustakas A. Transforming growth factor beta as regulator of cancer stemness and metastasis. Br J Cancer. 2016;115:761–769. | ||
Yu Y, Xiao CH, Tan LD, Wang QS, Li XQ, Feng YM. Cancer-associated fibroblasts induce epithelial-mesenchymal transition of breast cancer cells through paracrine TGF-β signalling. Br J Cancer. 2014;110(3):724–732. | ||
Makagiansar IT, Avery M, Hu Y, Audus KL, Siahaan TJ. Improving the selectivity of HAV-peptides in modulating E-cadherin-E-cadherin interactions in the intercellular junction of MDCK cell monolayers. Pharm Res. 2001;18(4):446–453. | ||
Inagaki M, Nishi Y, Nishizawa K, Matsuyama M, Sato C. Site-specific phosphorylation induces disassembly of vimentin filaments in vitro. Nature. 1987;328(6131):649–652. | ||
Prokop JW, Liu Y, Milsted A, Peng H, Rauscher FJ 3rd. A method for in silico identification of SNAIL/SLUG DNA binding potentials to the E-box sequence using molecular dynamics and evolutionary conserved amino acids. J Mol Model. 2013;19(9):3463–3469. | ||
Li L, Han LY, Yu M, Zhou Q, Xu JC, Li P. Pituitary tumor-transforming gene 1 enhances metastases of cervical cancer cells through miR-3666-regulated ZEB1. Tumour Biol. 2015;37:15567–15573. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.