Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Effect of Insulin-Regulated FOXC2 Expression in Adipocyte Differentiation and Insulin Resistance
Authors Zhang X, Wang Y ![]() , Zuo F, Su Y, Liu H, Wu B, Nian X
, Zuo F, Su Y, Liu H, Wu B, Nian X ![]()
Received 15 May 2020
Accepted for publication 24 July 2020
Published 6 August 2020 Volume 2020:13 Pages 2801—2809
DOI https://doi.org/10.2147/DMSO.S262950
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Xuxiang Zhang,1 Yuming Wang,2 Fan Zuo,1 Yandan Su,3 Hua Liu,3 Bin Wu,1 Xin Nian1
1Department of Endocrinology, First Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China; 2Department of Clinical Laboratory, Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China; 3Department of Clinical Laboratory, First Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China
Correspondence: Xin Nian
Department of Endocrinology, First Affiliated Hospital of Kunming Medical University Email [email protected]
Objective: 1) To investigate the effect of FOXC2 on the differentiation of adipose-derived mesenchymal stem cells. 2) To analyze the mechanism between FOXC2 expression regulation in adipose differentiation and insulin resistance (IR).
Methods: We first amplified the FOXC2 promoter region-512 and cloned it into the luciferase expression vector. The reporter gene system was transfected into the adipose tissue-derived mesenchymal stem cell to study insulin-mediated FOXC2 expression. We also manipulated FOXC2 protein expression by either siRNA or overexpression and studied the differentiation capability of adipose tissue-derived mesenchymal stem cell into adipocytes, as well as the influence on several IR-related genes: GLUT4, PPARγ, UCP1 and PAI-1.
Results: 1) Insulin effectively induced the expression of FOXC2 protein in adipose tissue-derived mesenchymal stem cells under differentiation (P< 0.01). Insulin also induced FOXC2-pro-512T promoter activity significantly (P< 0.01). 2) The stem cell adipose differentiation decreased in the FOXC2 overexpression group. 3) When FOXC2 was overexpressed, the expression of GLUT4, PAI-1 and UCP1 was higher than control groups (p< 0.001). When FOXC2 was down-regulated by siRNA, both GLUT4 and PAI-1’s protein expression were decreased (p< 0.001), and the protein expression of PPARγ was increased (p< 0.001). In the presence of insulin induction, overexpression of FOXC2 led to significantly higher UCP-1 expression (p< 0.001) and lower PAI-1 expression (p< 0.001). The protein expression of GLUT4, PAI-1 (p< 0.001) and UCP-1 (p< 0.05) was decreased in cells transfected with FOXC2 siRNA.
Conclusion: Insulin effectively induced the expression of FOXC2 protein in adipose tissue-derived mesenchymal stem cells under differentiation, possibly through the regulation of the FOXC2-pro-512T promoter activity. The different protein expression of FOXC2 has regulatory effects on several genes related to insulin resistance. FOXC2 is an important regulatory factor in adipocyte differentiation and insulin resistance.
Keywords: FOXC2, expression regulation, adipocyte differentiation, insulin resistance
Introduction
The regulation of fat metabolism involves many factors, and FOXC2 (forkhead box C2) is one of them. FOXC2 is a transcription factor of the winged helix/forkhead transcription factor family,1 and plays an important regulatory role in the metabolism of blood lipids and blood glucose.2 Thus, FOXC2 is considered to have an activity to control obesity and insulin resistance. In addition, Datta et al3 have shown that the overexpression of FOXC2 in human podocyte can cause podocyte function to be impaired and is associated with proteinuria. Some studies suggested that FOXC2 expression is significantly associated with several proteins that are related to insulin resistance (IR), such as PPARγ, UCP-1, PAI-1 and GLUT4.4–6 However, it is not clear how insulin and FOXC2 regulate the expression of these IR-related factors.
Our previous studies suggested that elevated expression of FOXC2 in visceral fat tissue led to decreased blood glucose level and increased HDL, and the TT genotype at the C-512T site of FOXC2 gene had significantly lower rates of HOMA2-IR than the CC and CT genotype populations. In this study, we studied the expression of FOXC2 in preadipocytes, by reporter gene system and by interfering expression with siRNA. Furthermore, we examined the regulatory effects of FOXC2 on insulin-resistance-related genes PPARγ, GLUT4, PAI-1 and UCP-1.
Materials and Methods
The Culture of Adipose Tissue-Derived Mesenchymal Stem Cells
Mesenchymal Stem Cells Complete Culture Medium Kit, Human adipose-derived mesenchymal stem cells were purchased from AbMole Bioscience. IBMX were purchased from Sigma. 0.25% Trypsin-EDTA (1X), Phenol Red, DMEM, and Low Glucose were purchased from Gibco. Australian superior fetal bovine serum FBS were obtained from Cellmax. To obtain differentiated adipocyte, cells were cultured in the L-DMEM (low sugar) medium containing 5% FBS, 1mM dexamethasone, 10ug/mL insulin, and 0.5mM IBMX.
After centrifugation of cell suspension, the cell precipitates were resuspended with whole culture medium and cultured at 37°C in a culture chamber with 5% CO2 and saturated humidity. The complete culture medium was replaced with fresh medium every 1–2 days. Cells were digested and passage when cells a confluence of 80%~90% was reached. Adipose tissue-derived mesenchymal stem cells were inoculated into 24-well plate at a density of 2.5–4.0×104 cells/cm2, 10 days after culture with adipocyte differentiation medium. These cells were stained with oil red O and photographed by electron microscopy.
Insulin Induction
Cells were cultured in a complete or differentiated medium for 48 hours. After rinsing with serum-free DMEM, cells were cultured in serum-free L-DMEM overnight. In the next day, insulin was added to the medium and cells were cultured for another 6 hours before they were collected to study the expression levels of the FOXC2 protein.
Construction of a Luciferase Reporter Vector and Detection of Luciferase Activity
Synthesized cDNA of 512-C and 512-T, in pUC57 plasmids, were purchased from Shuoqin Biotech. KpnI and HindIII were purchased from Thermo. PGL3-basic-vector was a product of Promega. T4 DNA ligase, DH5α receptive bacteria, gel extraction kit, and plasmid mini-extraction kit were from Tiangen.
512-C and 512-T cDNA fragments were cleaved from pUC57 plasmid by Kpn1 and Hind III, and subsequently cloned into PGL3-basic-vector by the same pair of restriction enzymes.
Plasmid Extraction
According to plasmid extraction manufacturer’s instructions.
Identification of Recombinant Plasmid by Enzyme Digestion
The post product of enzymatic digestion was shown by gel electrophoresis. The two recombinant plasmids are named PGL3-basic-FOXC2-pro-512-T and PGL3-basic-FOXC2-pro-512-C, respectively, as following. See Figure 1.
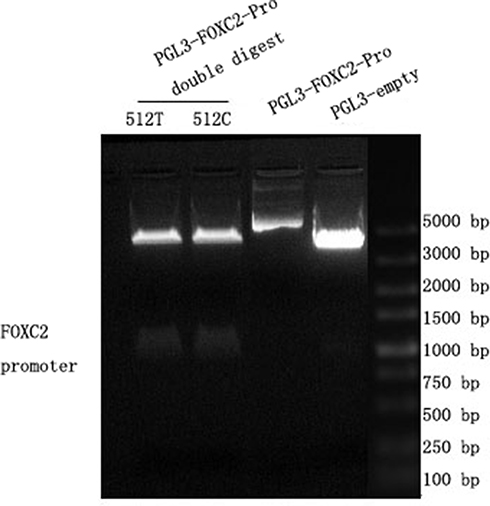 |
Figure 1 Results of plasmid enzymatic digestion. PGL3 basic plasmid was successfully connected into the 512T and 512C pro. In the figure, the right-most band 5 is Marker, band 1 is the double enzyme digestion product of pgl3-basic-FOXC2-pro-512-T, band 2 is the double enzyme digestion product of pgl3-basic-FOXC2-pro-512-C, band 3 is the PGL3-FOXC2-pro, and band 4 is the PGL3- empty. |
Detection of FOXC2 Promoter Activity by Dual Luciferase Reporter Gene Assay
To study the regulation of FOXC2 promoter activity, Dual Luciferase Reporter Gene Assay Kit (Bi Yun Tian) was used. Sea kidney fluorescein reporter plasmid pRL-TK vector was used as internal reference control. The activity of luciferase in cells was detected following the manufacturer’s protocol.
Cell Transfection
The experimental subgroup: pRL-TK vector is the internal reference control, each copy is 25ng; PGL3-vector (empty plasmid or plasmid inserted into promoter mutation), each group 475ng; PGL3-basic-FOXC2-pro-512-C and PGL3-basic-FOXC2-pro-512-T groups, each group 475ng; PGL3-vector group, PGL3-basic-FOXC2-pro-512-C group and PGL3-basic-FOXC2-pro-512-T group were added to insulin 4nM.
siRNA Transfection
Cells were ready for transfection when the confluence reached ~70%, 15min before transfection, the cell culture medium was replaced with serum-free culture medium, 1.5mL per well, a 5ul Lipofectamine 2000 was dissolved in 245ul of serum-free DMEM medium and mixed at room temperature for 5min. 10ul of siRNA at 100 pM was dissolved in 240ul of serum-free DMEM and mixed at room temperature for 5min. Mix the above two tubes and let them rest at room temperature for 20 minutes. The mixture was cultured at 37°C. Then the mixture was added to cells for 6 hours before changing the medium to the differentiation medium.
Plasmid and siRNA Transfection
When cell confluence reached about 70%, the cell-culture medium was changed into a serum-free medium at 15 minutes before transfection. 5ul Lipofectamine 2000 (Invitrogen Lipofectamine) was dissolved in 245ul DMEM serum-free medium and mixed at room temperature for 5min. 4ug plasmid (10ul 100pm siRNA) was dissolved in 240ul DMEM serum-free medium and mixed at room temperature for 5min. Lipofectamine and siRNA were mixed and incubated with cells at room temperature for 20 min, and then incubated for 6h at 37°C before cells were changed into differentiation medium.
Protein Expression Measured by Western Blot
Cells were washed by precooled PBS and kept on ice. Add 10 microliters PMSF (100mM) to 1mL lysate and shake it on ice. 100 microliters of lysis buffer (1 mM PMSF) were added to cells for a 30min incubation on-ice. Cells were scraped and transferred into a 1.5mL centrifuge tube. After 12,000 RPM centrifugation at 4°C for 5 min, the supernatant was transferred to 0.5 mL centrifuge tube. For Western blot, detailed procedure was described previously (See Clinical Lab.2016; 62:781-791). The following commercial primary antibodies were used: FOXC2 Antibody (Elabscience); PPARγ Polyclonal Antibody (rabbit), Glut4 Polyclonal Antibody (rabbit) (SAB); UCP1 Antibody 4E5 (sc-293418, Santa Cruz); GAPDH antibody (Elabscience). The following Secondary antibodies were used: Pro-Light HRP (Beijing Tiangen Biotech); Goat anti-mouse IgG-HRP, Goat anti-rabbit IgG-HRP. Primary antibodies were diluted at a ratio of 1:1000. Secondary antibodies were diluted at a ratio of 1:5000. All samples were prepared in triplicate.
Data Processing
All data were recorded into a computer and statistically analyzed with GraphPad Prism6. Data were statistically analyzed by One-way ANOVA, with a significant level set at p<0.05. The measured data were shown as mean with SD of the group of samples (n=3). The two-group differences were examined by t-test, with a significant level set at p<0.05.
Results
Expression of FOXC2 Protein in Adipose Tissue-Derived Mesenchymal Stem Cells Can Be Effectively Induced by Insulin Under the Differentiation Condition
The expression of FOXC2 protein in adipose tissue-derived mesenchymal stem cells cultured in complete medium or differentiation culture medium was detected. In the complete medium, cells were induced with 3nM insulin. In the differentiation medium, cells were induced with 1nM, 2nM, or 4nM insulin. FOXC2 expression levels under each condition were normalized with GAPDH levels. As shown in Figure 2, although the expression of FOXC2 protein in adipose cells increased after insulin induction, only under in the differentiation medium and with 4nM insulin, the FOXC2 expression level in adipose tissue-derived mesenchymal stem cells was significantly different from the control (P<0.001). In normal medium, the increase was not significant (P=0.191).
 |
Figure 2 FOXC2 protein expression in adipose mesenchymal stem cells. In both normal medium (complete medium) and differentiation medium, FOXC2 protein expression in adipocytes increased after insulin treatment. In differentiation medium, FOXC2 expression was significantly increased in the presence of 4nM insulin. ***P < 0.001. |
Effect of Insulin on FOXC2 Promoter
To study the regulation of FOXC2 promoter activity, we established dual luciferase reporter gene plasmids with two versions of FOXC2 promoter: 512C and −512T. The two plasmids, FOXC2-pro-512C and FOXC2-pro-512T, respectively, as well as control plasmid (PGL3) were transfected separately into adipocytes. To compare firefly luciferase activity induced by the FOXC2 promoter, the renilla luciferase was used as the internal reference. As shown in Figure 3, 4nM insulin was able to significantly induce the activity of the FOXC2-pro-512T promoter. In contrast, the FOXC2-pro-512C promoter was insensitive to insulin of the same concentration.
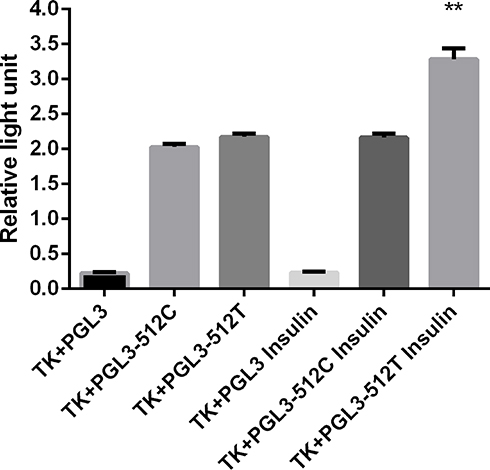 |
Figure 3 Effect of insulin on FOXC2 promoters. Insulin significantly increase the FOXC2-pro-512T promoter activity. **P<0.01. |
Effect of FOXC2 Expression on Fat Formation and Differentiation of Adipocytes
We studied the effect of FOXC2 expression on adipocytes via two approaches: overexpression and knock-down. To overexpress FOXC2, FOXC2 plasmid or empty plasmid was transfected into adipocytes. To knock-down FOXC2, NC siRNA (control) or FOXC2 siRNA was transfected into adipocytes. Expression of FOXC2 in transfected cells was evaluated by Western blot. As shown in Figure 4, FOXC2 expression was significantly increased in cells transfect with FOXC2 plasmid, but reduced in cells transfected with FOXC2 siRNA. Using GAPDH as the internal reference, we also calculated the relative expression levels of FOXC2 (Figure 4).
 |
Figure 4 Expression of FOXC2 protein in adipocytes transfected with FOXC2 overexpression plasmid or siRNA. ***P < 0.001. |
A major function of adipocytes is to produce fat. Thus, the lipogenic differentiation of adipocytes can be studied by Oil Red O staining. To understand the effect of FOXC2 on the adipose tissue-derived mesenchymal stem cells, FOXC2 overexpression or knockdown adipocytes were stained with Oil Red O after days of cell differentiation. The results showed that Oil red staining intensified in cells transfected with FOXC2 siRNA, suggesting that limiting FOXC2 expression led to significant increase of fat content in cells. Consistently, in cells with FOXC2 overexpression, Oil Red O staining decreased, suggesting less fat was produced in these cells. These data suggested that FOXC2 could inhibit the differentiation of adipose tissue-derived mesenchymal stem cells into adipocytes (Figure 5).
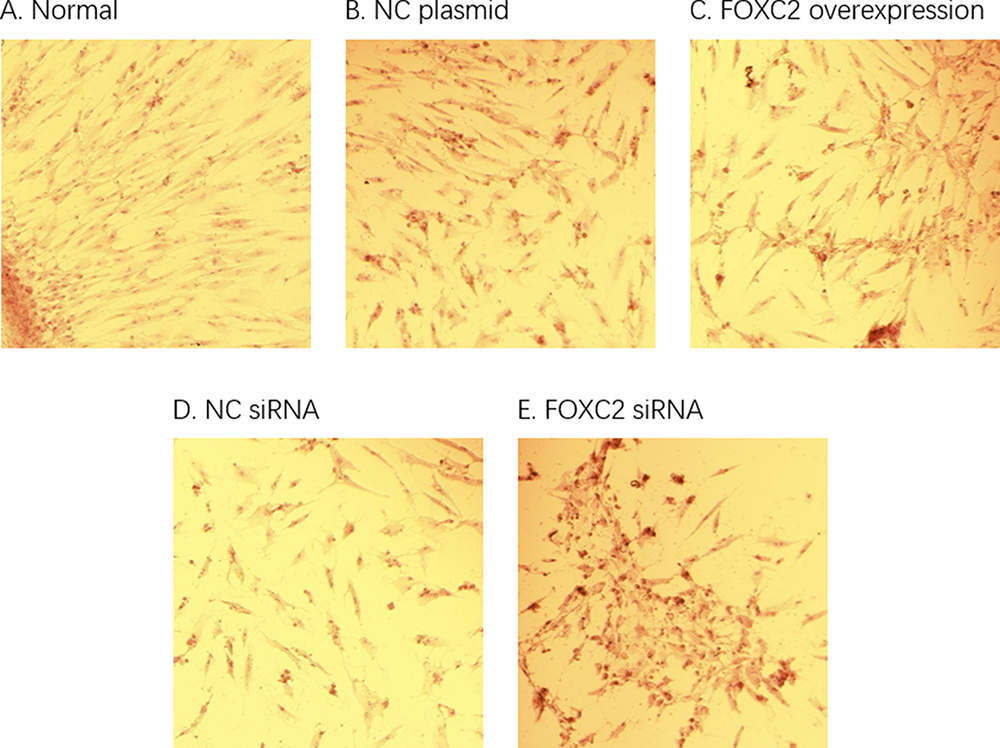 |
Figure 5 Adipocyte differentiation and Oil Red O staining. The (A) group was cultured with normal differentiation. Compared to NC siRNA group (D), the oil red staining was intensified in cells transfected with FOXC2 siRNA (E), indicating that the adipose differentiation of the cells was significantly increased. Compared to NC plasmid (B), oil Red O staining was reduced in FOXC2 overexpression group (C), suggesting a decreased fat formation. |
Effects of FOXC2 Expression on IR-Related Genes
We also examined the regulation of expression of IR-related genes by FOXC2. In the absence of insulin induction, knocking-down of FOXC2 by siRNA led to reduced expression of GLUT4 and PAI-1 (P<0.001), while overexpression of FOXC2 led to increased expression of GLUT4 and PAI-1 (P<0.001). PPARγ protein expression was significantly higher in the FOXC2 siRNA group vs the control group (P<0.001), while UCP1 protein expression was significantly higher in the FOXC2 overexpression cells as compared to control cells (P<0.001). According to the results, FOXC2 enhances the expression of GLUT4, PAI-1, and UCP1, and inhibits the expression of PPARγ.
In the presence of insulin induction, the effects of FOXC2 on GLUT4 and PPARγ were less obvious, suggesting an inhibitory effect of insulin on FOXC2 functions. For UCP1, insulin further enhanced the promoting activity of FOXC2 on UCP1 expression, as siRNA transfected cells showed much lower levels, while FOXC2 overexpression cells showed much higher levels. Insulin had a mixed effect on FOXC2-mediated PAI-1 expression (Figures 6 and 7).
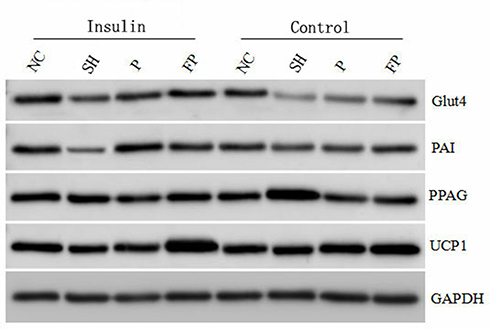 |
Figure 6 The electrophoretogram of the expression of the genes. Control: no insulin. NC: control siRNA; SH: FOXC2 siRNA; P: control Plasmid; FP: FOXC2 Plasmid. |
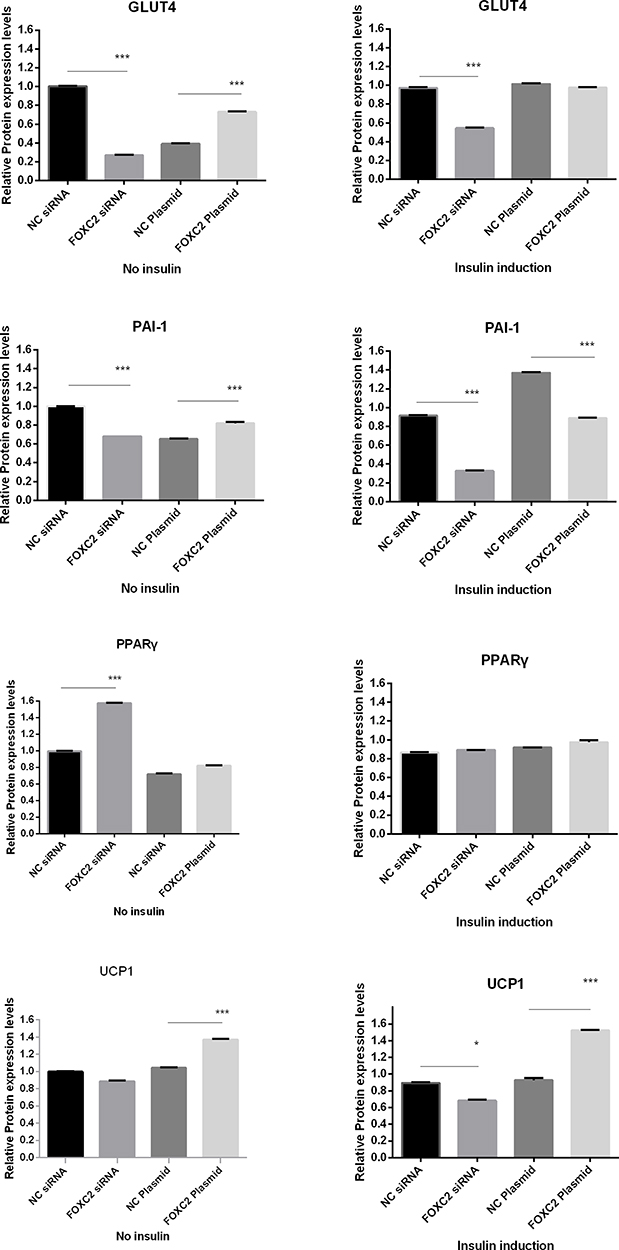 |
Figure 7 Expression of IR-related genes under different FOXC2 expression condition, with or without insulin induction. ***P<0.001. |
Discussion
FOXC2 is closely related to adipocyte differentiation.7 Studies have shown that the increased expression of FOXC2 in adipocytes has pleiotropic effects on genes related to the differentiation and metabolism of adipocytes,8 and it can affect the β-adrenergic sensitivity and intercellular signal transduction9,10 by changing the composition of PKA total enzyme in adipocytes. Therefore, FOXC2 is closely related to sugar metabolism and lipid metabolism. Another study reported that the mRNA level of FOXC2 gene was reduced in subcutaneous adipose tissue of insulin-resistance patients, and the mRNA level of FOXC2 gene in visceral adipose tissue and skeletal muscle was positively correlated with insulin sensitivity.11 FOXC2 and promoter polymorphism play an important role in the regulation of glucose and lipid metabolism. Yildirim-Toruner12 reported that FOXC2 C-512T polymorphism was associated with diabetes mellitus. Additional studies suggested that the FOXC2 C-512T was possibly associated with dysmetabolic syndrome,13 obesity and dyslipidemia.14 Kovacs et al15 reported that the C-512T polymorphism of FOXC2 promoter region was associated with BMI and body fat percentage in Pima population (both male and female). It is also related with basal glucose turnover and fasting plasma triglyceride levels in female.
In this study, insulin can effectively induce the expression of FOXC2 protein in adipose tissue-derived mesenchymal stem cells in the differentiation medium. Higher FOXC2 protein expression levels were observed at higher insulin concentrations. The expression of FOXC2 induced by 4nM insulin was significantly higher than that induced by 0, 1nM and 2nM insulin. These results suggest that the insulin dose-dependently promote the expression of FOXC2 in adipocytes. The present study also found that insulin significantly induced the activity of FOXC2-pro-512T promoter, while the FOXC2-pro-512C promoter was less sensitive to insulin. It suggested that TT genotype of the FOXC2 gene was related to insulin resistance.
Oil red O staining is used to show the amount of lipid in adipose tissue, as the dye is easily dissolved in lipid. After siRNA interferes with FOXC2 expression, red oil staining deepens after stem cell differentiation. Our data suggested that the expression of FOXC2 gene was reversely correlated with the fat content in adipocytes. Thus, FOXC2 gene was closely related to adipocyte differentiation. Davis et al16 confirmed that FOXC2 inhibited the differentiation of 3T3-L1 preadipocytes, and FOXC2 overexpression inhibited fat formation. This study further showed that FOXC2 could also inhibit adipose differentiation in adipose tissue-derived mesenchymal stem cells, suggesting that FOXC2 is closely related to obesity.
The GLUT4 gene plays an important role in the production of IR. GLUT4 is the main gene involved in adipose differentiation and glucose metabolism. It was found that the abnormality of the GLUT4 gene could lead to the IR, as GLUT4 was down-regulated when insulin resistance occurred.17,18 We investigated the effect of different expressions of FOXC2 on the expression of GLUT4. In the process of adipocyte differentiation, when FOXC2 was overexpressed, the expression of GLUT4 was significantly increased. When FOXC2 was silenced, the expression of GLUT4 was significantly decreased. It is indicated that FOXC2 could promote the expression of GLUT4, and FOXC2 could play an antagonistic role in insulin resistance by upregulating the level of GLUT4. During the differentiation of adipose tissue-derived stromal cells, 4nM insulin minimized the effect of FOXC2 on GLUT4 expression. This is in consistent with Figure 2, as insulin induced the expression of endogenous FOXC2. This renders knocking-down FOXC2 siRNA less effective, and overexpression of FOXC2 less obvious.
PAI-1 is a cytokine synthesized and secreted by adipocyte. Studies have found that adipose tissue, especially visceral adipose tissue, produces PAI-1, and the level of PAI-1 in obese people is significantly increased.19 FOXC2 plays an important role in regulating the insulin-induced expression of PAI-1. FOXC2 binds to the PAI-1 promoter at the insulin response component and the adnexal binding site, then upregulating the expression of PAI-1.20 In consistent, our studies showed that expression of PAI-1 was decreased after FOXC2 was silenced, while the expression of PAI-1 was significantly increased when FOXC2 was overexpressed. In the presence of insulin induction, PAI-1 expression was significantly increased, as shown in the control plasmid transfected cells (NC, Figure 6). It is not clear why cells with control siRNA failed to express more PAI-1 after insulin induction. However, FOXC2 siRNA clearly reduced PAI-1 expression with or without insulin, while overexpression failed to further enhance PAI-1 expression in the presence of insulin induction. It is possible that FOXC2 may have a complicated role in the regulation of PAI-1 promoter, and when there is an excessive amount of insulin, FOXC2 turns into an inhibitory factor for the transcription of PAI-1. This could be an antagonistic effect of FOXC2 on insulin resistance, but needs further studies to verify.
Brown adipose tissue (BAT) can selectively express UCP-1. Expression of UCP-1 in brown fat can reduce insulin resistance. Katagiri found that UCP-1 expression in liver increased energy consumption in mice, and reduced body weight and blood glucose level.21 Our study investigated the relationship between FOXC2 expression and UCP-1 expression. Apparently, FOXC2 promoted UCP-1 expression, especially in the presence of insulin induction. FOXC2 showed a synergistic effect with insulin on the expression of UCP-1 during adipocyte differentiation.
The activation of PPARγ in adipocyte has a regulating effect on the insulin signaling pathway and changes the insulin-mediated glucose homeostasis and fat metabolism processes. The activation of PPAR can promote the expression of GLUT1, GLUT2 and GLUT4, and lead to the changes in the expression of some key genes in glucose and lipid metabolism and insulin signal transduction pathway. PPARγ can selectively induce the expression of lipoprotein lipase and acetylcoenzyme A in adipose tissue, and promote the elimination of FFA in adipose tissue, resulting in the retention of fatty acids in adipose tissue and the reduction of available fatty acids in the whole body. Therefore, PPARγ can accelerate the consumption of fatty acids through the metabolic process of glucose and fatty acids, thus improving insulin sensitivity.22 Joakim performed fat biopsies and genetic tests in patients with type 2 diabetes before and after rosiglitazone treatment. The result showed that rosiglitazone treatment increased the number of mitochondria in patients, as well as FOXC2 and mtTFA/Tfam expression. It was suggested that FOXC2 was likely associated with PPARγ, a target of Thiazolidinediones (TZD), and played a role in the mitochondrial response activated by TZD.23 However, in our study, although knocking down FOXC2 led to an increase of PPARγ expression, neither overexpression nor knock-down of FOXC2 had any effect on PPARγ expression when adipocytes were cultured in the insulin medium.
In summary, our data suggest that insulin can effectively induce the expression of FOXC2 protein in adipose tissue-derived mesenchymal stem cells under the differentiation condition, and can significantly induce the FOXC2-pro-512T promoter activity. FOXC2 may inhibit the differentiation of adipose tissue-derived mesenchymal stem cells and reduce the number of adipocytes. Insulin can affect the expression of the insulin resistance-related genes, such as PPARγ, GLUT4, PAI-1 and UCP1, while FOXC2 can affect the expression levels of GLUT4, PAI-1 and UCP1, but has no significant effect on the expression level of PPARγ. Thus, FOXC2 participates in the regulation of insulin resistance but plays different regulatory roles on the insulin-resistance-related genes.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Rutscher K, Wilting J. Transcription factor FOXC2 demarcates the jugular lymphangiogenic region in avian embryos. Lymphology. 2008;41(1):11–17.
2. Jag UR, Zavadil J, Stanley FM. Insulin acts through FOXO3a to activate transcription of plasminogen activator inhibitor type 1. Mol Endocrinol. 2009;23(10):1587–1602. doi:10.1210/me.2008-0421
3. Datta N, Lindfors S, Miura N, et al. Overexpression of transcription factor FOXC2 in cultured human podocytes upregulates injury markers and increases motility. Exp Cell Res. 2016;340:32–42. doi:10.1016/j.yexcr.2015.10.035
4. Yang X, Enerback S, Smith U. Reduced expression of FOXC2 and brown adipogenic genes in human subjects with insulin resistance. Obes Res. 2003;11:1182–1191. doi:10.1038/oby.2003.163
5. Kim JK, Kim HJ, Park SY, et al. Transgenic mice overexpressing FOXC2 are z from high diet-induced insulin resistance in skeletal muscle and liver. Diabetes. 2003;52(Suppl 1):A314.
6. Cederberg A, Gronning LM, Ahren B, Tasken K, Carlsson P, Enerback S. FOXC2 is a winged helix gene that counteracts obesity, hypertriglyceridemia, and diet-induced insulin resistance. Cell. 2001;106:563–573. doi:10.1016/S0092-8674(01)00474-3
7. Gjedde S, Gormsen LC, Riis AL, et al. Reduced expression of uncoupling protein 2 in adipose tissue in patients with hypothyroidism. J Clin Endocrinol Metab. 2010;95(7):3537–3541. doi:10.1210/jc.2009-0907
8. Dahle MK, Gronning LM, Cederberg A, et al. Mechanisms of FOXC2- and FOXD1-mediated regulation of the RI alpha subunit of cAMP-dependent protein kinase include release of transcriptional repression and activation by protein kinase B alpha and cAMP. J Biol Chem. 2002;277(25):22902–22908. doi:10.1074/jbc.M200131200
9. Kim JK, Kim HJ, Park SY, et al. Adipocyte-specific over expression of FOXC2 prevents diet-induced increases in intramuscular fatty acyl CoA and insulin resistance. Diabetes. 2005;54(6):1657–1673. doi:10.2337/diabetes.54.6.1657
10. Ferrell RE, Kimmak MA, Lawrence EC, et al. Candidate gene analysis in primary lymphedema. Lymphat Res Biol. 2008;6(2):69–76. doi:10.1089/lrb.2007.1022
11. Ridderstråle M, Carlsson E, Klannemark M, et al. FOXC2 mRNA expression and a 5ʹuntranslated region polymorphism of the gene are associated with insulin resistance. Diabetes. 2002;51(12):3554–3560. doi:10.2337/diabetes.51.12.3554
12. Yildirim-Toruner C, Subramanian K, El Manjra L, Chen E, Goldstein S, Vitale E. A novel frameshift mutation of FOXC2 gene in a family with hereditary lymphedema-distichiasis syndrome associated with renal disease and diabetes mellitus. Am J Med Genet A. 2004;131(3):281–286. doi:10.1002/ajmg.a.30390
13. Carlsson E, Groop L, Ridderstråle M. Role of the FOXC2-512C>T polymorphism in Type 2 diabetes: possible association with the dysmetabolic syndrome. Int J Obes (Lond). 2005;29(3):268–274. doi:10.1038/sj.ijo.0802876
14. Carlsson E, Almgren P, Hoffstedt J, Groop L, Martin R. The FOXC2 C-512T polymorphism is associated with obesity and dyslipidemia. Obes Res. 2004;12(11):1738–1743. doi:10.1038/oby.2004.215
15. Kovacs P, Lehn-Stefan A, Stumvoll M, et al. Genetic variation in the human winged helix/forkhead transcription factor gene FOXC2 in Pima Indians. Diabetes. 2003;52(5):1292–1295. doi:10.2337/diabetes.52.5.1292
16. Davis KE, Moldes M, Farmer SR. The forkhead transcription factor FoxC2 inhibits white adipocyte differentiation. J Biol Chem. 2004;279(41):42453–42461. doi:10.1074/jbc.M402197200
17. Gaster M, Staehr P, Beck Nielsen H, Schroder HD, Handberg A. GLUT4 is reduced in slow muscle fibers of type 2 diabetic patients: is insulin resistance in type 2 diabetes a slow, type 1 fiber disease. Diabetes. 2001;50(6):1324–1329. doi:10.2337/diabetes.50.6.1324
18. Abel ED, Peron IO, Kimi JK, et al. Adipose selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature. 2001;409(6821):729–733. doi:10.1038/35055575
19. Delporte ML, Funahashi T. Pre and post translational negative effect of adrenoceptor agonists on adiponectin secretion: in vitro and in vivo. Biochem J. 2002;367:677. doi:10.1042/bj20020610
20. Fujita H, Kang M, Eren M, Gleaves LA, Vaughan DE, Kume T. Foxc2 is a common mediator and insulin transforming growth factor beta signaling to regulate plasm inogen activator inhibitor type I gene expression. Circ Res. 2006;98(5):626–634. doi:10.1161/01.RES.0000207407.51752.3c
21. Katagiri S, Park K, Maeda Y. Overexpressing IRS1 in endothelial cells enhances angioblast differentiation and wound healing in diabetes and insulin resistance. Diabetes. 2016;65(9):2760–2771. doi:10.2337/db15-1721
22. Yalverde AM, Benito M, Lorenzo M. The brown adipose cell: a model for understanding the molecular mechanisms of insulin resistance. Acta Physiol Scand. 2005;183:59–73. doi:10.1111/j.1365-201X.2004.01384.x
23. Håkansson J, Eliasson B, Smith U, Enerbäck. S. Adipocyte mitochondrial genes and the forkhead factor FOXC2 are decreased in type 2 diabetes patients and normalized in response to rosiglitazone. Diabetol Metab Syndr. 2011;3:32–40. doi:10.1186/1758-5996-3-32
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.