")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 16
Dysregulation of circRNA-0076906 and circRNA-0134944 is Correlated with Susceptibility to Osteoporosis and Osteoporotic Fracture in Postmenopausal Females from the Chinese Han Population
Authors Yang W, Zhang W, Li F, Xu N, Sun P
Received 28 October 2022
Accepted for publication 12 January 2023
Published 10 March 2023 Volume 2023:16 Pages 183—194
DOI https://doi.org/10.2147/PGPM.S394757
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Martin H Bluth
Weijie Yang,* Wei Zhang,* Fengqian Li, Ning Xu, Ping Sun
Department of Orthopedics, Shanghai Eighth People’s Hospital, Shanghai, 200235, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ping Sun, Department of Orthopedics, Shanghai Eighth People’s Hospital, No. 8, Caobao Road, Shanghai, 200235, People’s Republic of China, Email [email protected]
Introduction: Many circRNAs, such as circRNA-0076906 and circRNA-0134944, have been reported to participate in the pathogenesis of osteoporosis via sponging miRNAs in postmenopausal female patients. In this study, we aimed to study potential signaling pathways underlying the role of certain circRNAs, miRNAs and their target genes in the pathogenesis of osteoporotic fracture in postmenopausal females.
Methods: Quantitative real-time PCR was performed to analyze the expression of circRNAs, miRNAs and their targets genes. Luciferase assays were carried out to explore the regulatory relationship between circ_0076906/miR-548i/OGN and circ_0134944/miR-630/TLR4.
Results: Osteoporosis and fracture were positively correlated to the expression of circ_0134944, miR-548i and TLR4, but negatively correlated to the expression of circ_0076906, miR-630 and OGN in the peripheral blood and bone tissue samples of postmenopausal women. Luciferase activities of wild-type circ_0076906 and OGN were inhibited by miR-548i, and the luciferase activities of wild-type circ_0134944 and TLR4 were suppressed by miR-630 in MG-63 and U-2 OS cells. Inhibition of circ_0076906 expression in MG-63 and U-2 OS cells activated the expression of miR-548i and inhibited the expression of OGN. Moreover, the overexpression of circ_0134944 in MG-63 and U-2 OS cells suppressed the expression of miR-630 and enhanced the expression of TLR4.
Conclusion: This study implied that the dysregulation of circRNA-0076906 and circRNA-0134944 modulated their specific signaling and thus contributed to the severity of osteoporosis, increasing the risk of osteoporotic fracture.
Keywords: osteoporotic, circRNA, miRNA, OGN, TLR4
Introduction
Osteoporosis is a frequently diagnosed disease of the bone featured by reduced bone tissue quality as well as a reduced mass of bone tissues. It was determined that > 20 million people suffer from various degrees of osteoporosis at any moment.1,2 Although various therapeutic approaches and agents, such as vitamin D, bisphosphonates, as well as calcium, have been tried to treat osteoporosis and show a great progress in the past few decades, there is still a lack of effective treatments due to the complicated mechanisms implicated in the pathogenesis of osteoporosis.3–5 While current osteoporosis treatments can decrease the danger of bone fracture, fracture can still happen during the treatment although it may not indicate an inadequate treatment response.6–8 In addition, it has been recognized that during the treatments of subjects who have not been treated with similar treatments before, a new fracture substantially elevates the danger of succeeding fractures.9,10 For that reason, it is expected that certain bone fractures may occur in subjects undertaking procedure for the treatment of osteoporosis.11,12
CircRNAs can participate in many biological processes such as cell growth, cell signaling, pathological responses, and physiological responses.13,14 It was shown that circRNAs might act as miRNA sponges via competing with mRNAs for miRNA binding to regulate gene expression.15,16 For example, circ_0076906 has been reported to function as a sponge for miR-1305 to compete with Osteoglycin (OGN), a target gene of miR-1305. Therefore, by regulating the expression of OGN, circ_0076906 could relive osteoporosis and promote osteogenic differentiation of human mesenchymal stem cells (hMSCs) and alleviate the progression of osteoporosis.17 Moreover, circ_0134944 was reported to downregulate miR-127 to participate in the signaling of miR-127-5p/PDX1/SPHK1 pathway, thus inhibiting osteoporosis.18
Osteoglycin (OGN) is a humoral bone anabolic factor derived from muscles.19 OGN level was shown to be linked to the mineralization as well as phenotypes of osteoblasts.20 As a result, OGN may work as a key factor in the regulation of MSC differentiation as well as a therapeutic target of osteoporosis. Subsequently, it was suggested that OGN might be of a key value in the osteogenesis process of senile osteoporosis by regulating the expression of genes specific to osteogenesis, including RUNX2, ALP, OCN, as well as Wnt5b. In a previous study, lentiviral vectors were used to restore OGN expression in osteoporotic SMMSCs to identify the effects of OGN on modulating the osteogenic differentiation of these cells. It was shown that OGN overexpression up-regulated the expression of runt-related transcription factor 2 (RUNX2), alkaline phosphatase (ALP), Osteocalcin (OCN), as well as Wnt Family Member 5B (Wnt5b) but downregulated the expression of adaptor protein 2 (aP2) as well as peroxisome proliferator-activated receptor γ2 (PPARγ2), which defined the phenotypes of adipocytes, indicating that the OGN signaling might play an essential role in managing osteoblast differentiation in the pathogenesis of senile osteoporosis.21
Toll-like receptor 4 (TLR4) is expressed on osteoclast as well as osteoblast surface to serve as important regulators of immune responses during chronic inflammation in the postmenopausal females, eventually leading to the onset of postmenopausal osteoporosis at some point. Kaleta et al found no difference in the genotype frequency of TLR4 in postmenopausal females with and without osteoporosis.22 The administration of methionine was shown to reduce TLR4 expression, which is usually induced in osteoclast precursors by ovariectomy, suggesting that TLR4 is implicated in weight gain, osteoclast genesis, insulin resistance as well as inflammation.23
OGN was reported to play a significant role in osteoporosis,21 and circ_0076906 has been reported to modulate the expression of OGN and alleviate the progression of osteoporosis.17 And circ_0134944 was reported to be upregulated in postmenopausal osteoporosis patients,24 and prevent the process of osteoporosis by overexpressing SPHK1.18 Moreover, in the study which explores the genome-wide circRNAs expression files to identify candidate circRNAs associated with osteoporosis, several circRNAs including circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422 were identified via microarray.24,25 Therefore, in this study, we investigated the expression of these circRNAs to identify the circRNAs associated with osteoporosis fracture and study the potential signaling pathways underlying the role of circRNAs, miRNAs and their target genes in the pathogenesis of osteoporotic fracture in postmenopausal females.
Materials and Methods
Patient Recruitment
In this study, we recruited 306 female subjects during they are being hospitalized due to osteoporosis fracture and divided them into three groups: ie, 1. Osteoporosis (-) Fracture (-) group (N=132, patients with neither osteoporosis nor fracture); 2. Osteoporosis (+) Fracture (-) group (N=112, patients with osteoporosis but no fracture); and 3. Osteoporosis (+) Fracture (+) group (N=62, patients with both osteoporosis and fracture). Osteoporosis is defined based on the BMD evaluation by dual-energy X-ray absorptiometry (DXA). And the fractures were mainly defined via radiological imaging methods as well as clinical manifestation. The information of these patients including their age, height, weight, BMI, bone mineral density (BMD), L1-L4 BMD, femoral neck BMD and total hip BMD was collected and compared among the 3 groups using the Student’s test. In addition, peripheral blood samples were collected from all subjects of the three groups. Furthermore, because some of the subjects in the above three groups underwent surgery, their bone tissue samples were harvested to examine the expression of circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422, miR-548i and miR-630. The subjects subject to bone tissue sample collection were divided into the following 3 groups, ie, 1. Osteoporosis (-) Fracture (-) group (N=12, patients with neither osteoporosis nor fracture); 2. Osteoporosis (+) Fracture (-) group (N=11, patients with osteoporosis but no fracture); and 3. Osteoporosis (+) Fracture (+) group (N=6, patients with both osteoporosis and fracture). The Human Research Ethics Committees of Shanghai Eighth People’s Hospital has approved this research and all methods were performed in accordance with the last vision of the Declaration of Helsinki. Written informed consent was obtained from all participants before the study.
RNA Isolation and Real-Time PCR
Quantitative real-time PCR was performed to analyze the expression of circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422, miR-548i and miR-630 in each collected sample. In brief, total RNA was extracted from the collected samples first by making use of a RNeasy Mini kit (Qiagen GmbH) according to the experimental protocol recommended by the reagent manufacturer. Then, RNA was reverse transcribed into cDNA templates by using a Primer Script one-step reverse transcription assay kit (TaKaRa, Tokyo, Japan) according to the experimental protocol recommended by the assay kit manufacturer. In the next step, real time PCR was carried out by using the synthesized cDNA templates and a SYBR Green Premix Ex Taq II real time PCR assay kit (TaKaRa, Tokyo, Japan) according to the experimental protocol recommended by the assay kit manufacturer. The real time PCR reactions were carried out in an ABI7500 real time PCR machine (Applied Biosystems, Foster City, CA) according to the instrument instructions of the manufacturer. Finally, the relative expression of circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422, miR-548i, TLR4 mRNA, OGN mRNA as well as miR-630 in each sample was determined by using the 2−ΔΔCt approach. The sequences of primers used in this study were shown in Supplementary Table 1.
Cell Culture and Transfection
In order to further explore the regulatory relationship of various target genes, various siRNAs were transfected into MG-63 and U-2 OS cells to evaluate their effects on the expression of miR-548i, circ_0134944, miR-630 and TLR4. In brief, MG-63 and U-2 OS cells were bought from the Shanghai Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and maintained according to the culture conditions recommended by the cell supplier, ie, Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, Grand Island, NY) added with 10% FBS (HyClone, Invitrogen, Carlsbad, CA), 100 U/mL of penicillin as well as 100 μg/mL of streptomycin (Invitrogen, Carlsbad, CA). The cells were incubated in humidified 37 ° C incubator under 5% CO2. The cells were passaged once every 3 days during the experiment. When the cells were 70% confluent, they were divided into two cell models. In cell model 1, the cells were divided into 2 groups, ie 1. NC siRNA group (MG-63 and U-2 OS cells transfected with a negative control siRNA); and 2. circ_0076906 siRNA group (MG-63 and U-2 OS cells transfected with circ_0076906 siRNA). In cell model 2, the cells were also divided into 2 groups, ie 1. a pGL3 group in which the MG-63 and U-2 OS cells were transfected with an empty pGL3 vector (Cat No. E1751, Promega, Madison, WI); and 2. a pGL3-circ_0134944 group in which the MG-63 and U-2 OS cells were transfected with a pGL3 vector carrying circ_0134944. For the transfection experiments, the cells in the various groups were transfected by utilizing Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the experimental protocol recommended by the manufacturer of the transfection reagent. At 48 h after transfection, the transfected cells were harvested to assay the expression of target genes.
Vector Construction, Mutagenesis and Luciferase Assay
The results of binding site prediction indicated that miR-548i could bind to circ_0076906 and OGN, while circ_0134944 and TLR4 were the potential targets of miR-630. In order to confirm the regulatory relationship between miR-548i, circ_0076906 and OGN, luciferase assays were performed. Site-directed mutagenesis was carried out using a Quick Change Site-Directed Mutagenesis assay kit (Stratagene, San Diego, CA) according to the assay protocol to induce mutations in the miR-548i binding sites of circ_0134944 and OGN 3’ UTR, respectively. The wild-type or mutant miR-548i sequences containing circ_0134944 and OGN 3’ UTR binding sites were cloned into pGL3 luciferase reporter vectors (Cat No. E1751, Promega, Madison, WI) to generate vectors carrying miR-548i with wild-type/mutant binding sites of circ_0134944 and OGN 3’ UTR. In the next step, these vectors were co-transfected into MG-63 and U-2 OS cells with miR-548i or negative controls using Lipofectamine 2000 (Invitrogen, USA). At 48 h after the transfection, the luciferase activity of transfected cells was assayed by using a Dual Luciferase reporter assay kit (Promega, Madison, WI) according to the kit protocol.
Luciferase assays were also performed to confirm the regulatory relationship between circ_0134944, TLR4 and miR-630. Similarly, site-directed mutagenesis was carried out by using the Quick Change Site-Directed Mutagenesis assay kit (Stratagene, San Diego, CA) to induce mutations in the miR-630 binding sites of circ_0134944 and TLR4 3’ UTR, respectively. The wild-type/mutant miR-630 sequences were respectively cloned into pGL3 luciferase reporter vectors to generate vectors carrying miR-630 with wild-type/mutant binding sites of circ_0134944 and TLR4 3’ UTR. MG-63 and U-2 OS cells were co-transfected with wild-type/mutant vectors with miR-630 or negative controls by using Lipofectamine 2000 (Invitrogen, USA). At 48 h after the transfection, the luciferase activity of transfected cells was also assayed by using the Dual Luciferase reporter assay kit (Promega, Madison, WI) according to the kit protocol.
Statistical Analysis
The statistical analysis was conducted by utilizing SPSS 19.0 (SPSS, Chicago, IL). The differences among different groups were analyzed by using Student’s t tests. P values were bilateral and were deemed statistically significant at < 0.05.
Results
Patient Characterization
In this study, we recruited 306 female subjects and divided them into three groups: 1. Osteoporosis (-) Fracture (-) (N=132); 2. Osteoporosis (+) Fracture (-) (N=112); and 3. Osteoporosis (+) Fracture (+) (N=62) groups. The information of these patients including their age, height, weight, BMI, BMD, L1-L4 BMD, femoral neck BMD and total hip BMD was summarized in Table 1. No obvious difference was observed between the three groups in terms of patient age, height, weight and BMI, while the L1-L4 BMD, femoral neck BMD and total hip BMD were progressively decreased from group 1 to group 3.
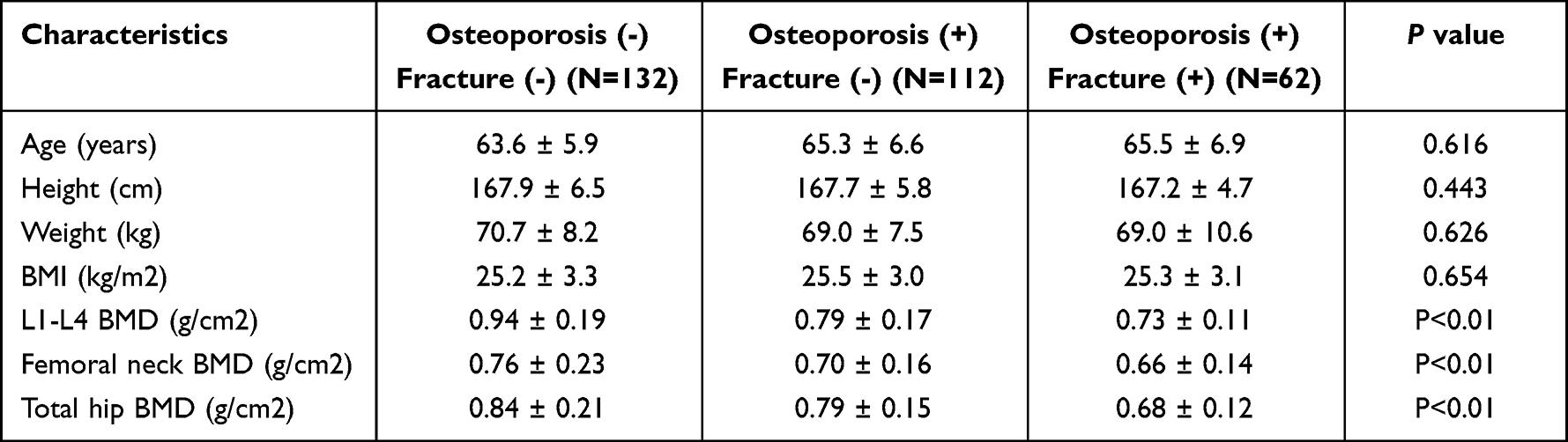 |
Table 1 Basic Patient Information and Characteristics |
Osteoporosis and Fracture Were Closely Correlated with the Expression of circ_0134944, circ_0005692, miR-548i and miR-630 in the Peripheral Blood and Bone Tissue Samples of Postmenopausal Women
Peripheral blood samples were collected from the subjects in the three groups. Quantitative real-time PCR was performed to analyze the expression of circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422, miR-548i and miR-630 in the collected peripheral blood samples. The expression of circ_0134944 was significantly higher in the peripheral blood samples of Osteoporosis (+) Fracture (+) patients and lower in the peripheral blood samples of Osteoporosis (-) Fracture (-) patients when compared with that in the peripheral blood samples of Osteoporosis (+) Fracture (-) patients (Figure 1A). On the contrary, the expression of circ_0076906 was remarkably suppressed in the peripheral blood samples of Osteoporosis (+) Fracture (+) patients and activated in the peripheral blood samples of Osteoporosis (-) Fracture (-) patients (Figure 1B). Besides, no notable difference was observed for the expression of circ_0057340 (Figure 1C), circ_0088422 (Figure 1D), and circ_0005692 (Figure 1E) in the peripheral blood samples of these subjects. Moreover, the expression of miR-548i was significantly higher in the peripheral blood samples of Osteoporosis (+) Fracture (+) patients and lower in the peripheral blood of Osteoporosis (-) Fracture (-) patients (Figure 2A). The expression of miR-630 was remarkably suppressed in the peripheral blood samples of Osteoporosis (+) Fracture (+) patients and activated in the peripheral blood of Osteoporosis (-) Fracture (-) patients (Figure 2B).
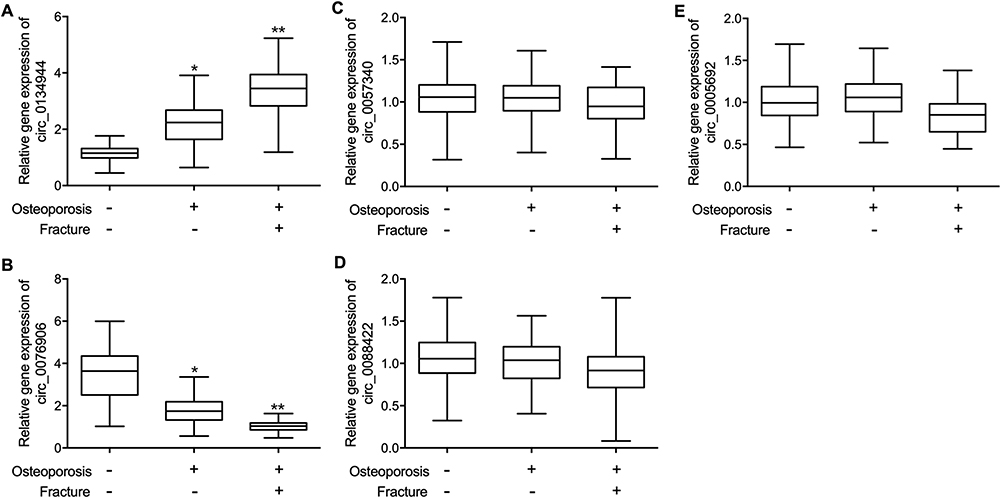 |
Figure 1 The expression of candidate circRNAs in the peripheral blood of postmenopausal women with or without osteoporosis or fracture (*p-value < 0.05 vs Osteoporosis (-) Fracture (-) group; **p-value < 0.0 vs Osteoporosis (+) Fracture (-) group). (A) Circulating expression of circ_0134944 was highest in postmenopausal women from the c. (B) Circulating expression of circ_0076906 was evidently reduced in postmenopausal women from the Osteoporosis (+) Fracture (+) group. (C) Circulating expression of circ_0057340 was similar among all patient groups. (D) Circulating expression of circ_0088422 was similar among all patient groups. (E) Circulating expression of circ_0005692 was similar among all patient groups. |
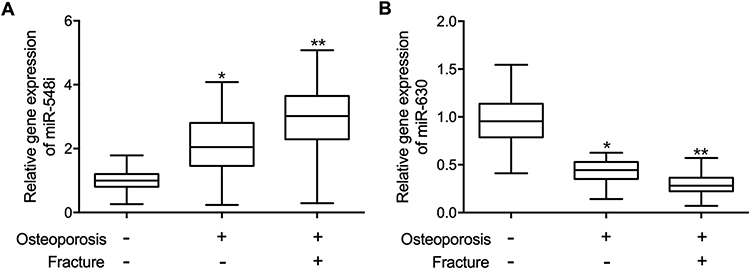 |
Figure 2 The expression of miR-548i and miR-630 in the peripheral blood of postmenopausal women (*p-value < 0.05 vs Osteoporosis (-) Fracture (-) group; **p-value < 0.0 vs Osteoporosis (+) Fracture (-) group). (A) Circulating expression of miR-548i was significantly elevated in the the Osteoporosis (+) Fracture (+) group. (B) Circulating expression of miR-630 was evidently reduced in the Osteoporosis (+) Fracture (+) group. |
Osteoporosis and Fracture Were Closely Correlated with the Expression of circ_0134944 and circ_0005692 in the Bone Tissue Samples of Postmenopausal Women
Furthermore, because some of the subjects in the three groups underwent surgery, their bone tissue samples were harvested to examine the expression of circ_0134944, circ_0057340, circ_0005692, circ_0076906, circ_0088422, miR-548i and miR-630. Similar to the results from the peripheral blood samples, osteoporosis and fracture were positively correlated with the expression of circ_0134944 (Figure 3A), but negatively correlated with the expression of circ_0076906 (Figure 3B) in the bone tissue samples. Furthermore, there was no apparent correlation between osteoporosis and the expression of circ_0057340 (Figure 3C), circ_0088422 (Figure 3D), and circ0005692 (Figure 3E).
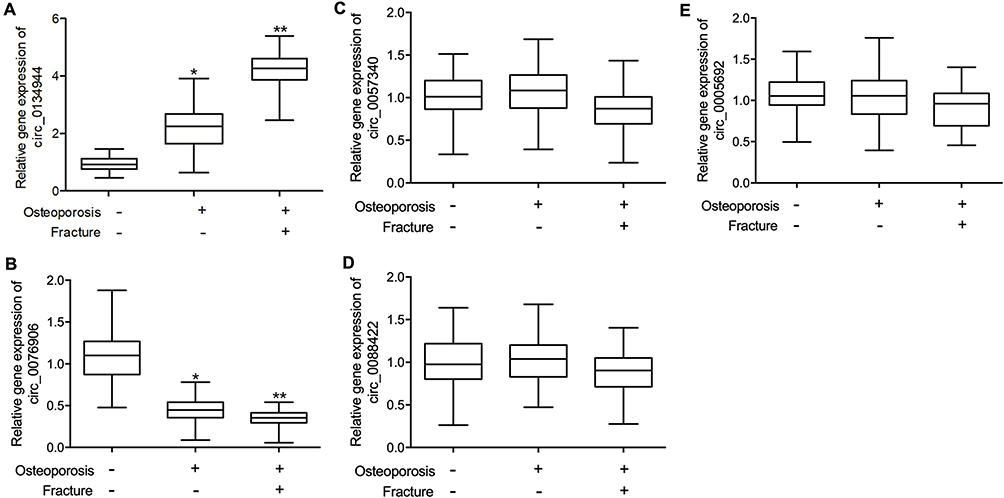 |
Figure 3 Tthe expression of candidate circRNAs in the bone tissue of postmenopausal women (*p-value < 0.05 vs Osteoporosis (-) Fracture (-) group; **p-value < 0.0 vs Osteoporosis (+) Fracture (-) group). (A) Tissue expression of circ_0134944 was significantly increased in the Osteoporosis (+) Fracture (+) group. (B) Tissue expression of circ_0076906 was lowest the Osteoporosis (+) Fracture (+) group. (C) Tissue expression of circ_0057340 was similar among all patient groups. (D) Tissue expression of circ_0088422 was similar among all patient groups. (E) Tissue expression of circ_0005692 was similar among all patient groups. |
Osteoporosis and Fracture Altered the Expression of miR-58i, miR-630, TLR4, and OGN in the Bone Tissue Samples of Postmenopausal Females
The expression of miR-548i was the highest in Osteoporosis (+) Fracture (+) patients (Figure 4A) while the expression of miR-630 was the lowest in Osteoporosis (+) Fracture (+) patients (Figure 4B). Moreover, qPCR was carried out to examine the expression of TLR4 and OGN in the bone tissue. The expression of TLR4 mRNA (Figure 4C) was apparently enhanced in osteoporosis (+) fracture (+) patients along with remarkably suppressed expression of OGN mRNA (Figure 4D) when compared with that in the bone tissue samples of the other two groups.
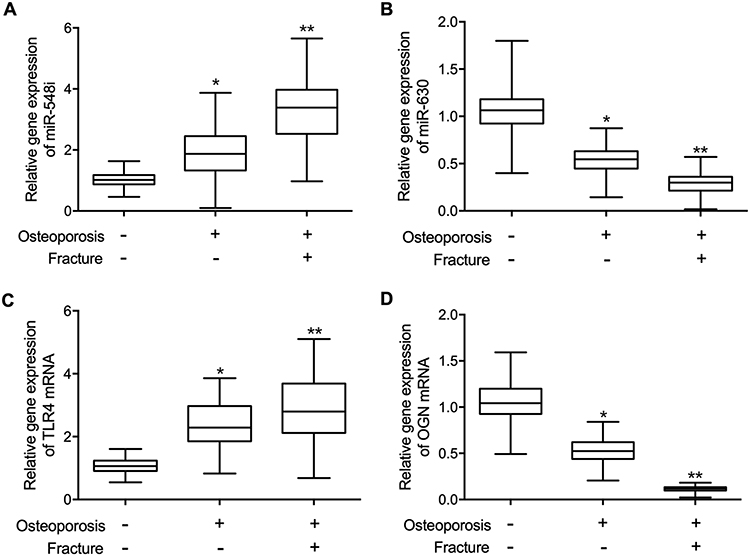 |
Figure 4 The expression of miR-548i, miR-630, TLR4 mRNA and OGN mRNA in the bone tissue of postmenopausal women (*p-value < 0.05 vs Osteoporosis (-) Fracture (-) group; **p-value < 0.0 vs Osteoporosis (+) Fracture (-) group). (A) Tissue expression of miR-548i was highest in the Osteoporosis (+) Fracture (+) group. (B) Tissue expression of miR-630 was lowest in the Osteoporosis (+) Fracture (+) group. (C) Tissue expression of TLR4 mRNA was highest in the Osteoporosis (+) Fracture (+) group. (D) Tissue expression of OGN mRNA was lowest in the Osteoporosis (+) Fracture (+) group. |
The Luciferase Activities of circ_0076906 and OGN Were Suppressed by miR-548i While the Luciferase Activities of circ_0134944 and TLR4 Were Inhibited by miR-630
The results of binding site prediction indicated that miR-548i could bind to circ_0076906 and OGN, while circ_0134944 and TLR4 were the potential targets of miR-630. In order to further explore the regulatory relationship among miRNA, circRNA and their target genes, luciferase vectors were established to test whether miR-548i suppressed the expression of circ_0076906 and OGN, and whether miR-630 repressed the expression of circ_0134944 and TLR4. The luciferase activities of wild-type circ_0076906 (Figure 5A) and OGN (Figure 5B) were remarkably inhibited by miR-548i in MG-63 and U-2 OS cells, while no inhibition was observed for mutant circ_0076906 and OGN. Meanwhile, the luciferase activities of wild-type circ_0134944 (Figure 5C) and TLR4 (Figure 5D) were remarkably inhibited by miR-548i in MG-63 and U-2 OS cells.
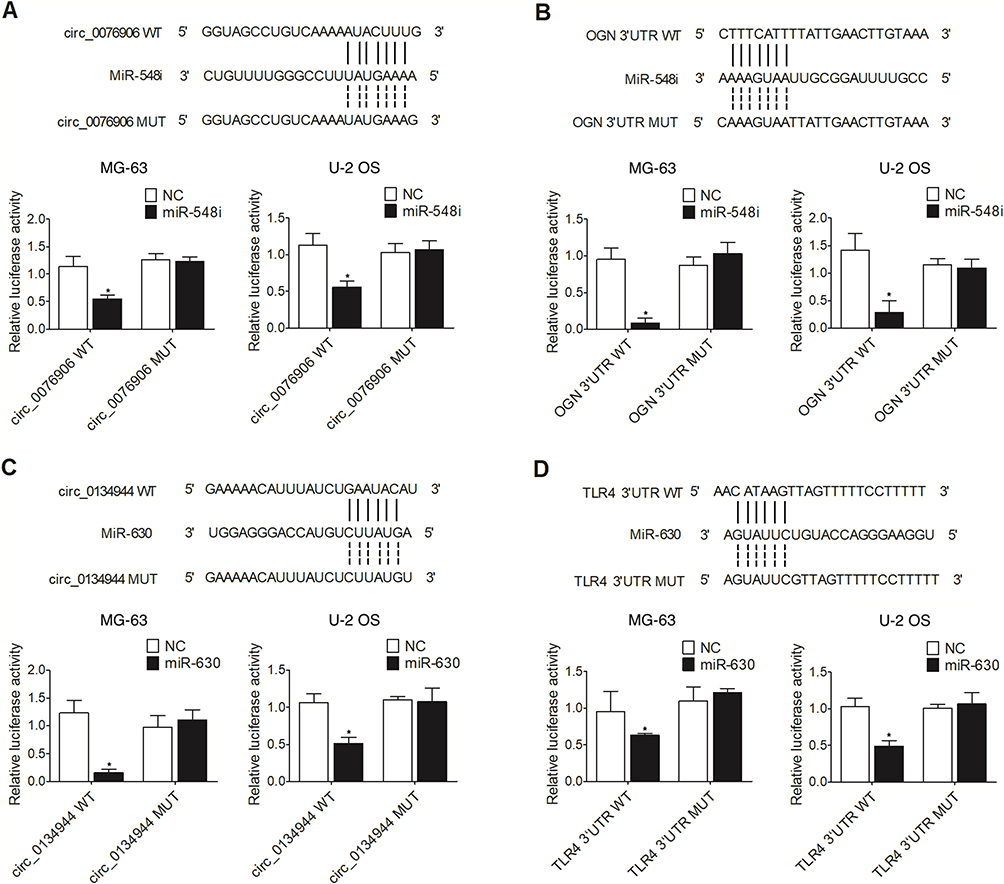 |
Figure 5 The luciferase activities of wild-type circ_0076906 and OGN were inhibited by miR-548i while the luciferase activities of wild-type circ_0134944 and TLR4 were suppressed by miR-630 (*p-value < 0.05 vs NC). (A) Sequence analysis and luciferase assays indicated that miR-548i suppressed the expression of circ_0076906 in MG-63 and U-2 OS cells. (B) Sequence analysis and luciferase assays indicated that miR-548i suppressed the expression of OGN in MG-63 and U-2 OS cells. (C) Sequence analysis and luciferase assays indicated that miR-630 suppressed the expression of circ_0134944 in MG-63 and U-2 OS cells. (D) Sequence analysis and luciferase assays indicated that miR-630 suppressed the expression of TLR4 in MG-63 and U-2 OS cells. |
Different Effects of circ_0076906 siRNA on the Expression of circ_0076906, miR-548i, OGN, circ_0134944, miR-630 and TLR4 in MG-63 and U-2 OS Cells
In order to further explore the regulatory relationship of various target genes, circ_0076906 siRNA was transfected into MG-63 and U-2 OS cells to evaluate its effects on the expression of miR-548i, circ_0134944, miR-630 and TLR4. Significant suppression of circ_0076906 (Figure 6A) as well as OGN mRNA (Figure 6B) was observed in transfected MG-63 and U-2 OS cells, whereas the expression of miR-548i (Figure 6C) was notably enhanced by circ_0076906 siRNA. No remarkable difference was detected for the expression of miR-630 (Figure 6D), circ_0134944 (Figure 6E), and TLR4 mRNA (Figure 6F) in MG-63 and U-2 OS cells.
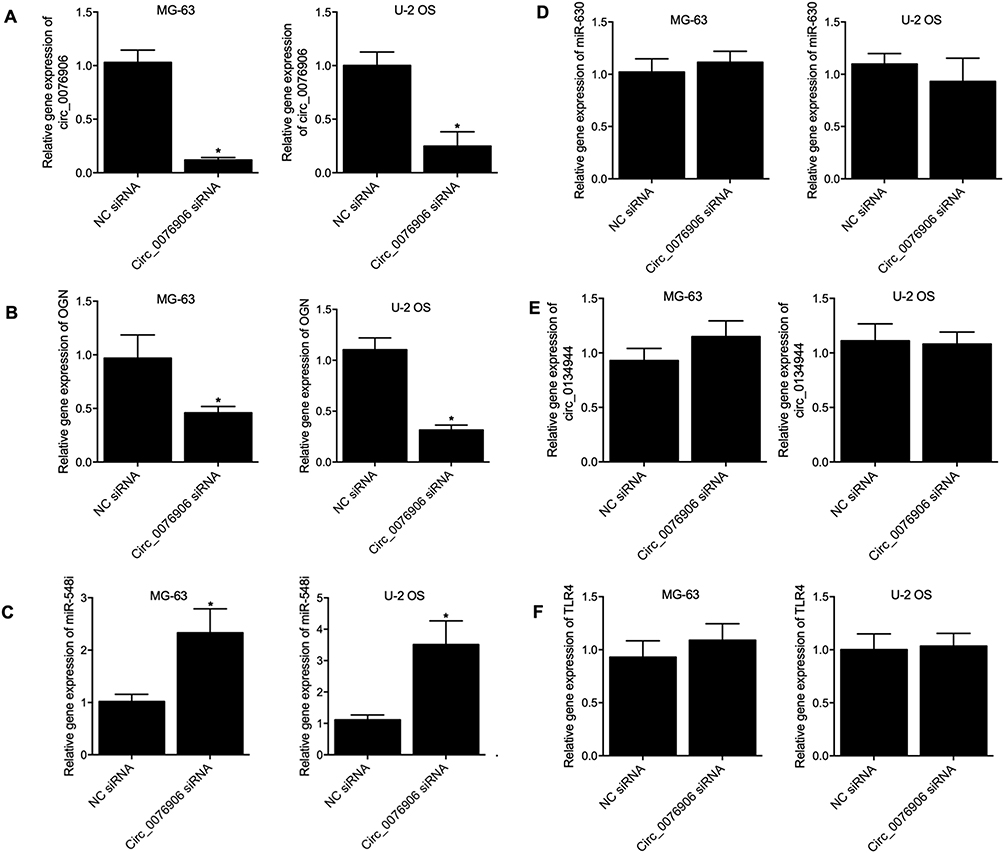 |
Figure 6 Inhibition of circ_0076906 expression in MG-63 and U-2 OS cells activated the expression of miR-548i but inhibited the expression of OGN (*p-value < 0.05 vs NC siRNA). (A) Remarkable suppression of circ_0076906 expression by circ_0076906 siRNA in MG-63 and U-2 OS cells. (B) Notable inhibition of OGN mRNA expression by circ_0076906 siRNA in MG-63 and U-2 OS cells. (C) Significant activation of miR-548i expression by circ_0076906 siRNA in MG-63 and U-2 OS cells. (D) The expression of miR-630 was not affected by circ_0076906 siRNA in MG-63 and U-2 OS cells. (E) The expression of circ_0134944 was not affected by circ_0076906 siRNA in MG-63 and U-2 OS cells. (F) The expression of TLR4 mRNA was not affected by circ_0076906 siRNA in MG-63 and U-2 OS cells. |
Different Effects of pGL3-circ_0134944 on the Expression of circ_0076906, miR-548i, OGN, circ_0134944, miR-630 and TLR4 in MG-63 and U-2 OS Cells
Furthermore, we transfected pGL3-circ_0134944 into MG-63 and U-2 OS cells to examine its effects on the expression of miR-548i, circ_0134944, miR-630 and TLR4. No apparent difference was observed for the expression of circ_0076906 (Figure 7A), miR-548i (Figure 7B), and OGN mRNA (Figure 7C) in MG-63 and U-2 OS cells transfected with pGL3-circ_0134944 when compared with that in the control. PGL3-circ_0134944 remarkably elevated the expression of circ_0134944 (Figure 7D) and TLR4 mRNA (Figure 7F) but suppressed the expression of miR-630 (Figure 7E) in MG-63 and U-2 OS cells.
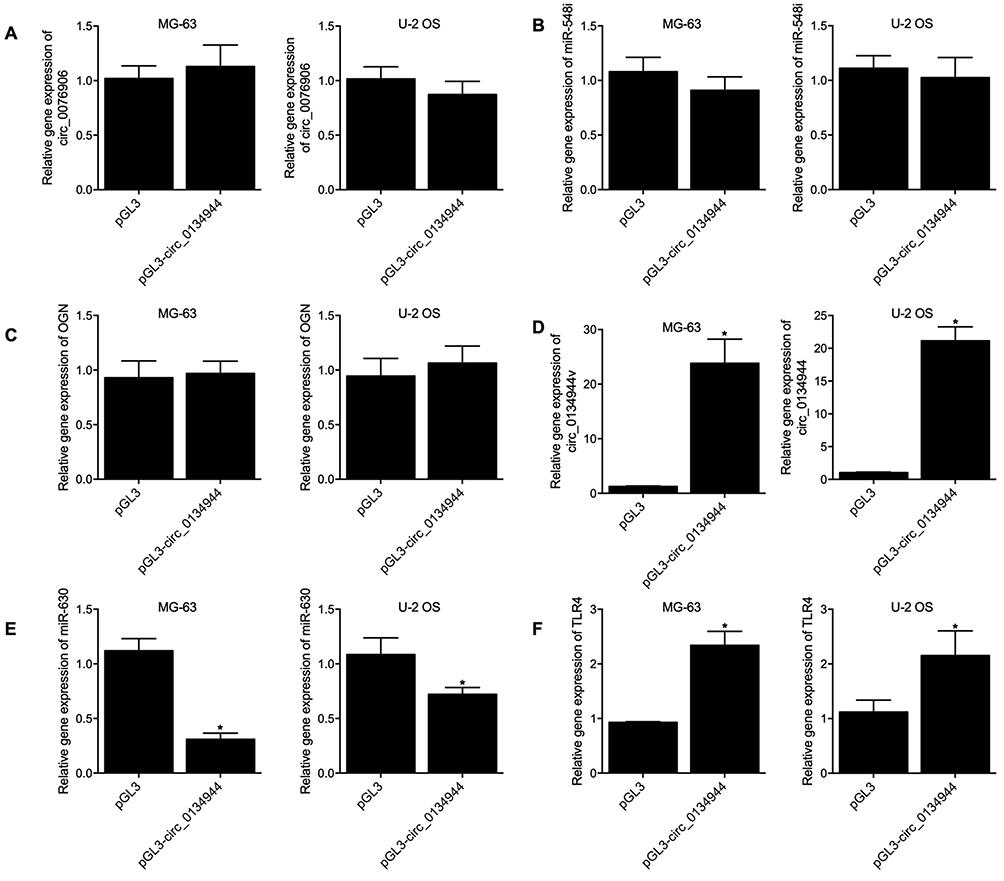 |
Figure 7 Overexpression of circ_0134944 expression in MG-63 and U-2 OS cells suppressed the expression of miR-630 and enhanced the expression of TLR4 (*p-value < 0.05 vs pGL3). (A) The expression of circ_0076906 was not affected by pGL3-circ_0134944 transfection in MG-63 and U-2 OS cells. (B) The expression of miR-548i was not affected by pGL3-circ_0134944 transfection in MG-63 and U-2 OS cells. (C) The expression of OGN mRNA was not affected by pGL3-circ_0134944 transfection in MG-63 and U-2 OS cells. (D) PGL3-circ_0134944 significantly enhanced the expression of circ_0134944 in MG-63 and U-2 OS cells. (E) PGL3-circ_0134944 significantly inhibited the expression of miR-630 in MG-63 and U-2 OS cells. (F) PGL3-circ_0134944 significantly enhanced the expression of TLR4 mRNA in MG-63 and U-2 OS cells. |
Discussion
In this study, we collected peripheral blood and bone tissue samples from postmenopausal females, and performed qPCR to measure the expression of candidate circRNAs and miRNAs. Osteoporosis and fracture were positively correlated with the expression of circ_0134944 and miR-548i, but negatively correlated with the expression of circ_0076906 and miR-630. In addition, we performed qPCR to evaluate the mRNA expression of TLR4 and OGN in bone tissues to explore the effect of osteoporosis and fracture. Osteoporosis and fracture were positively correlated with the expression of TLR4 but negatively correlated with the expression of OGN. Previous sequencing of circRNAs revealed the presence of 387 differentially expressed circRNAs in patients with osteoporosis, especially circ_0134944, circ_0016624, circ_0062466, circ_0057340, circ_0114125, circ_0116994, circ_0014777, circ_0005692, circ_0103518, circ_0115030, circ_0135422, circ_0094768, circ_0068239, circ_0076906, circ_0088422, circ_0133066, circ_0100388, circ_0091574, circ_0108438 as well as circ_0128044. As an example, in several previous studies, numerous circRNAs including circ_0001275 were determined as a biomarker for the diagnosis of postmenopausal osteoporosis.21,24 And to define the actual circiRNAs which most possibly participate in the process of osteoporosis, we observed the expressions of several circRNAs selected based on previous literatures including circ_0076906, circ_0134944, circ_0057340, circ_0088422 and circ_0005692. Accordingly, we found that only circ_0134944 and circ_0076906 were dysregulated in osteoporosis and osteoporosis fracture. Furthermore, we carried out luciferase assays to explore the regulatory network containing circ_0076906/miR-548i/OGN and circ_0134944/miR-630/TLR4. The luciferase activities of wild-type circ_0076906 and OGN were inhibited by miR-548i but the luciferase activities of wild-type circ_0134944 and TLR4 were suppressed by miR-630.
OGN has been proved to be function as a key factor in the regulation of MSC differentiation, by being an effective therapeutic target of osteoporosis.20 Subsequently, OGN was suggested to be associated with the osteogenesis process of senile osteoporosis by regulating the expression of genes specific to osteogenesis, including RUNX2, ALP, and Wnt5b. In previous studies, reduced expression of OGN was noticed in several types of cancers such as stomach cancer, colorectal cancer, breast cancer, as well as laryngeal cancer.26–28 Another research revealed that circ_0076906 competed with OGN to bind to miR-1305 in regulating the osteogenic differentiation of hMSCs. After the expression of miR-1305 or circ_0076906 was separately silenced with siRNAs, it was discovered that the expression of OGN down-regulated by circ_0076906 was recovered. Furthermore, the ALP activity as well as the mineralized matrix in bone tissues was recovered after the transfection of si-miR-1305, suggesting that circ_0076906 may partly control osteogenic differentiation through the miR-1305/OGN pathway. It was shown that OGN exerts an important effect on the pathogenesis of senile osteoporosis through regulating the expression levels of osteogenic as well as adipogenic genes, thus providing a target for treating senile osteoporosis.21 OGN could participate in bone tissue formation via differentiated osteoblasts.29,30 OGN expression might be induced by Cbfa1, a crucial gene in osteoblastogenesis.31 The activity of PPARγ helps to inhibit the maturation of osteoblasts while inducing SMMSC and MMSC differentiation into adipocytes.32 In this study, we found that in osteoporosis (+) fracture (+) patients, the level of OGN mRNA was evidently decreased, and miR-548i serves as a binding target of OGN mRNA. The dysregulation of circ_0076906 was observed along with the dysregulation of OGN mRNA, verifying the regulatory relationship between circ_0076906, miR-548i and OGN.
TLR4 is primarily expressed in myeloid cells such as macrophages, monocytes, granulocytes, chondrocytes, synoviocytes, osteoblasts as well as splenocytes.33,34 TLR has also been implicated in the pathogenesis of musculoskeletal diseases.34,35 TLR4 expression is high in bone marrow, immune cells, adipocytes as well as osteoblasts.23,36,37 Specifically, TLR4 expression was increased in MSCs undergoing osteoblastic differentiation.38 A past research proposed that the polymorphism of C1196T in TLR4 is unrelated to BMD or the incidence of osteoporotic bone fracture in postmenopausal women, although the C1196T polymorphism might be linked to a lower height.22 Surprisingly, it was discovered that methionine downregulated the expression of TLR4 induced by ovariectomy in osteoclast precursors. As a receptor of pathogen recognition that is closely related to insulin resistance, inflammation as well as weight gain, TLR4 also plays a key role in osteoclastogenesis.23,39 In this study, we suppressed the expression of circ_0076906 using siRNA while overexpressing circ_0134944 in MG-63 and U-2 OS cells to examine its effect on the expression of target miRNAs and relevant genes. The suppression of circ_0076906 up-regulated the expression of miR-548i and suppressed the expression of OGN, while the overexpression of circ_0134944 inhibited the expression of miR-630 and activated the expression of TLR4. In this study, we found that in osteoporosis (+) fracture (+) patients, the level of TLR4 mRNA was apparently enhanced, and miR-630 serves as a binding target of TLR4 mRNA. Moreover, the dysregulation of circ_0134944 was observed along with the dysregulation of TLR4 mRNA, verifying the regulatory relationship between circ_0134944, miR-639 and TLR4.
Conclusion
In this study, we established two pathways, circRNA-0076906/miR-548i/OGN and circRNA-0134944/miR-630/TLR4 to reveal the mechanism of participation of circRNA-0076906 and circRNA-0134944 in the pathogenesis of osteoporosis fracture. Accordingly, we found that the dysregulation of circRNA-0076906 and circRNA-0134944 could affect the expression of OGN and TLR4, thus contributing to the progress of osteoporosis. Therefore, it can be concluded that circRNA-0076906 and circRNA-0134944 are associated with risk of osteoporotic fracture in postmenopausal females from the Chinese Han population.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethical Statement
The Human Research Ethics Committees of Shanghai Eighth People’s Hospital has approved this research and all methods were performed in accordance with the last vision of the Declaration of Helsinki. Written informed consent was obtained from all participants before the study.
Funding
There is no funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ensrud KE, Crandall CJ. Osteoporosis. Ann Intern Med. 2017;167:ITC17–ITC32. doi:10.7326/AITC201708010
2. Black DM, Rosen CJ. Postmenopausal Osteoporosis. N Engl J Med. 2016;374:2096–2097. doi:10.1056/NEJMcp1513724
3. Black DM, Rosen CJ, Solomon CG. Clinical practice. Postmenopausal osteoporosis. N Engl J Med. 2016;374(3):254–262.
4. Khosla S, Hofbauer LC. Osteoporosis treatment: recent developments and ongoing challenges. Lancet Diabetes Endocrinol. 2017;5:898–907. doi:10.1016/S2213-8587(17)30188-2
5. Brown C. Osteoporosis: staying strong. Nature. 2017;550:S15–S17. doi:10.1038/550S15a
6. Black DM, Cummings SR, Karpf DB, et al. Randomised trial of effect of alendronate on risk of fracture in women with existing vertebral fractures. Fracture Intervention Trial Research Group. Lancet. 1996;348:1535–1541. doi:10.1016/S0140-6736(96)07088-2
7. Black DM, Reid IR, Boonen S, et al. The effect of 3 versus 6 years of zoledronic acid treatment of osteoporosis: a randomized extension to the HORIZON-Pivotal Fracture Trial (PFT). J Bone Miner Res. 2012;27:243–254. doi:10.1002/jbmr.1494
8. Silverman SL, Christiansen C, Genant HK, et al. Efficacy of bazedoxifene in reducing new vertebral fracture risk in postmenopausal women with osteoporosis: results from a 3-year, randomized, placebo-, and active-controlled clinical trial. J Bone Miner Res. 2008;23:1923–1934. doi:10.1359/jbmr.080710
9. Klotzbuecher CM, Ross PD, Landsman PB, Abbott TA, Berger M. Patients with prior fractures have an increased risk of future fractures: a summary of the literature and statistical synthesis. J Bone Miner Res. 2000;15:721–739. doi:10.1359/jbmr.2000.15.4.721
10. Kanis JA, Johnell O, De Laet C, et al. Tenenhouse A: a meta-analysis of previous fracture and subsequent fracture risk. Bone. 2004;35:375–382. doi:10.1016/j.bone.2004.03.024
11. Turnbull F. Blood pressure lowering treatment trialists C: effects of different blood-pressure-lowering regimens on major cardiovascular events: results of prospectively-designed overviews of randomised trials. Lancet. 2003;362:1527–1535.
12. Duell PB, Santos RD, Kirwan BA, Witztum JL, Tsimikas S, Kastelein JJP. Long-term mipomersen treatment is associated with a reduction in cardiovascular events in patients with familial hypercholesterolemia. J Clin Lipidol. 2016;10:1011–1021. doi:10.1016/j.jacl.2016.04.013
13. Han B, Chao J, Yao H. Circular RNA and its mechanisms in disease: from the bench to the clinic. Pharmacol Ther. 2018;187:31–44. doi:10.1016/j.pharmthera.2018.01.010
14. Salzman J. Circular RNA expression: its potential regulation and function. Trends Genet. 2016;32:309–316. doi:10.1016/j.tig.2016.03.002
15. Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384–388. doi:10.1038/nature11993
16. Zheng Q, Bao C, Guo W, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7:11215. doi:10.1038/ncomms11215
17. Wen J, Guan Z, Yu B, Guo J, Shi Y, Hu L. Circular RNA hsa_circ_0076906 competes with OGN for miR-1305 biding site to alleviate the progression of osteoporosis. Int J Biochem Cell Biol. 2020;122:105719. doi:10.1016/j.biocel.2020.105719
18. Zhang DW, Chen T, Li JX, Wang HG, Huang ZW, Lv H. Circ_0134944 inhibits osteogenesis through miR-127-5p/PDX1/SPHK1 pathway. Regen Ther. 2021;18:391–400. doi:10.1016/j.reth.2021.09.004
19. Kaji H. Interaction between Muscle and Bone. J Bone Metab. 2014;21:29–40. doi:10.11005/jbm.2014.21.1.29
20. Tanaka K, Matsumoto E, Higashimaki Y, et al. Role of osteoglycin in the linkage between muscle and bone. J Biol Chem. 2012;287:11616–11628. doi:10.1074/jbc.M111.292193
21. Chen X, Chen J, Xu D, Zhao S, Song H, Peng Y. Effects of Osteoglycin (OGN) on treating senile osteoporosis by regulating MSCs. BMC Musculoskelet Disord. 2017;18:423. doi:10.1186/s12891-017-1779-7
22. Kaleta B, Walicka M, Sawicka A, et al. Toll-like receptor 4 gene polymorphism C1196T in Polish women with postmenopausal osteoporosis - preliminary investigation. Adv Clin Exp Med. 2015;24:239–243. doi:10.17219/acem/22747
23. Shi H, Kokoeva MV, Inouye K, Tzameli I, Yin H, Flier JS. TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest. 2006;116:3015–3025. doi:10.1172/JCI28898
24. Yu L, Liu Y. circRNA_0016624 could sponge miR-98 to regulate BMP2 expression in postmenopausal osteoporosis. Biochem Biophys Res Commun. 2019;516(2):546–550. doi:10.1016/j.bbrc.2019.06.087
25. Li X, Wu J, Liu S, et al. miR-384-5p targets Gli2 and negatively regulates age-related osteogenic differentiation of rat bone marrow mesenchymal stem cells. Stem Cells Dev. 2019;28:791–798. doi:10.1089/scd.2019.0044
26. Lee JY, Eom EM, Kim DS, Ha-Lee YM, Lee DH. Analysis of gene expression profiles of gastric normal and cancer tissues by SAGE. Genomics. 2003;82:78–85. doi:10.1016/S0888-7543(03)00098-3
27. Wang Y, Ma Y, Lu B, Xu E, Huang Q, Lai M. Differential expression of mimecan and thioredoxin domain-containing protein 5 in colorectal adenoma and cancer: a proteomic study. Exp Biol Med. 2007;232:1152–1159. doi:10.3181/0701-RM-8
28. Rower C, Ziems B, Radtke A, et al. Toponostics of invasive ductal breast carcinoma: combination of spatial protein expression imaging and quantitative proteome signature analysis. Int J Clin Exp Pathol. 2011;4:454–467.
29. Chan CY, Masui O, Krakovska O, et al. Identification of differentially regulated secretome components during skeletal myogenesis. Mol Cell Proteomics. 2011;10:M110 004804. doi:10.1074/mcp.M110.004804
30. Patel MJ, Liu W, Sykes MC, et al. Identification of mechanosensitive genes in osteoblasts by comparative microarray studies using the rotating wall vessel and the random positioning machine. J Cell Biochem. 2007;101:587–599. doi:10.1002/jcb.21218
31. Zambotti A, Makhluf H, Shen J, Ducy P. Characterization of an osteoblast-specific enhancer element in the CBFA1 gene. J Biol Chem. 2002;277:41497–41506. doi:10.1074/jbc.M204271200
32. Kawaguchi H, Akune T, Yamaguchi M, et al. Distinct effects of PPARgamma insufficiency on bone marrow cells, osteoblasts, and osteoclastic cells. J Bone Miner Metab. 2005;23:275–279. doi:10.1007/s00774-005-0599-2
33. Medzhitov R, Preston-Hurlburt P, Janeway CA. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature. 1997;388:394–397. doi:10.1038/41131
34. Midwood K, Sacre S, Piccinini AM, et al. Tenascin-C is an endogenous activator of Toll-like receptor 4 that is essential for maintaining inflammation in arthritic joint disease. Nat Med. 2009;15:774–780. doi:10.1038/nm.1987
35. Wang P, Zhu F, Tong Z, Konstantopoulos K. Response of chondrocytes to shear stress: antagonistic effects of the binding partners Toll-like receptor 4 and caveolin-1. FASEB J. 2011;25:3401–3415. doi:10.1096/fj.11-184861
36. Kikuchi T, Matsuguchi T, Tsuboi N, et al. Gene expression of osteoclast differentiation factor is induced by lipopolysaccharide in mouse osteoblasts via Toll-like receptors. J Immunol. 2001;166:3574–3579. doi:10.4049/jimmunol.166.5.3574
37. Nemoto E, Honda T, Kanaya S, Takada H, Shimauchi H. Expression of functional Toll-like receptors and nucleotide-binding oligomerization domain proteins in murine cementoblasts and their upregulation during cell differentiation. J Periodontal Res. 2008;43:585–593. doi:10.1111/j.1600-0765.2008.01096.x
38. Mo IF, Yip KH, Chan WK, Law HK, Lau YL, Chan GC. Prolonged exposure to bacterial toxins downregulated expression of toll-like receptors in mesenchymal stromal cell-derived osteoprogenitors. BMC Cell Biol. 2008;9:52. doi:10.1186/1471-2121-9-52
39. Vijayan V, Khandelwal M, Manglani K, Gupta S, Surolia A. Methionine down-regulates TLR4/MyD88/NF-kappaB signalling in osteoclast precursors to reduce bone loss during osteoporosis. Br J Pharmacol. 2014;171:107–121. doi:10.1111/bph.12434
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.