Back to Journals » Journal of Pain Research » Volume 9
Dysesthesia symptoms produced by sensorimotor incongruence in healthy volunteers: an electroencephalogram study
Authors Katayama O ![]() , Osumi M, Kodama T, Morioka S
, Osumi M, Kodama T, Morioka S ![]()
Received 17 September 2016
Accepted for publication 10 November 2016
Published 8 December 2016 Volume 2016:9 Pages 1197—1204
DOI https://doi.org/10.2147/JPR.S122564
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Michael Schatman
Osamu Katayama,1,2 Michihiro Osumi,3 Takayuki Kodama,4 Shu Morioka,1,3
1Department of Neurorehabilitation, Graduate School of Health Sciences, Kio University, Nara, 2Department of Rehabilitation, Watanabe Hospital, Aichi, 3Department of Neurorehabilitation Research Center, Kio University, Nara, 4Department of Physical Therapy, Graduate School of Health Sciences, Kyoto Tachibana University, Kyoto, Japan
Objectives: Pathological pain such as phantom limb pain is caused by sensorimotor incongruence. Several studies with healthy participants have clearly indicated that dysesthesia, which is similar to pathological pain, is caused by incongruence between proprioception and/or motor intention and visual feedback. It is not clear to what extent dysesthesia may be caused by incongruence between motor intention and visual feedback or by incongruence between proprioception and visual feedback. The aim of this study was to clarify the neurophysiology of these factors by analyzing electroencephalograms (EEGs).
Methods: In total, 18 healthy participants were recruited for this study. Participants were asked to perform repetitive flexion/extension exercises with their elbows in a congruent/incongruent position while viewing the activity in a mirror. EEGs were performed to determine cortical activation during sensorimotor congruence and incongruence.
Results: In the high-frequency alpha band (10–12 Hz), numeric rating scale scores of a feeling of peculiarity were significantly correlated with event-related desynchronization/synchronization under the incongruence and proprioception conditions associated with motor intention and visual feedback (right inferior parietal region; r=−0.63, P<0.01) and between proprioception and visual feedback (right temporoparietal region; r=−0.49 and r=−0.50, P<0.05). In these brain regions, there was a region in which incongruence between proprioception and visual feedback and between motor intention and visual feedback caused an increase in activity.
Conclusion: The present findings suggest that neural mechanisms of dysesthesia are caused by incongruence between proprioception associated with motor intention and visual feedback and, in particular, are a result of incongruence between proprioception only and visual feedback.
Keywords: sensorimotor incongruence, dysesthesia, electroencephalogram, high-frequency alpha band, event-related desynchronization/synchronization
Introduction
It has been confirmed that sensorimotor incongruence causes pathological pain such as phantom limb pain,1 and several studies with healthy participants have clearly indicated incongruence between proprioception associated with motor intention and visual feedback to be the cause of dysesthesias such as pain, peculiarity, heaviness, the sensation of an extra limb, and nausea.2,3 Moreover, sensorimotor incongruence has been reported to exacerbate pain and sensory perceptions in patients with fibromyalgia,4 those with whiplash-associated disorders,5 and violinists with symptoms of sensory changes6 to a greater extent than in healthy volunteers. To resolve such sensorimotor incongruences and alleviate pathological pain, mirror therapy and virtual visual feedback have been used in clinical practice.7–17 Earlier studies were conducted with the use of active exercise-containing motor intention. In these investigations, participants performed a series of bilateral upper limb movements while viewing a mirror/whiteboard, which created varied degrees of sensorimotor conflict during congruent/incongruent limb movements. Any changes in sensory experience were recorded using qualitative methods. Therefore, it is still not clear whether pain and dysesthesia are caused by incongruences between motor intention and visual feedback or between proprioception and visual feedback. In the current study, conditions for the presence or absence of motor intention were set as follows: incongruence between motor intention and visual feedback (intention condition) and incongruence between proprioception and visual feedback with no motor intention (proprioception condition). We aimed to clarify the neurophysiology of these factors by analyzing electroencephalograms (EEGs). Previous studies have reported attenuation (ie, increased event-related desynchronization [ERD]) of high-frequency alpha power (10–12 Hz) during activation of the cortical sensorimotor system.18–20 Furthermore, high-frequency alpha power has been reported to be attenuated during motor execution or motor imagery.21,22 In this study, we focused on the frequency band of EEGs in the high-frequency alpha rhythm.
Methods
Participants
In total, 18 healthy participants were recruited (3 women and 15 men; age range 23–30 years, mean [standard deviation] age 26.7 years [2.4]). Participants had no current or past physical or mental illnesses (eg, neurological or chronic pain states) and normal or corrected-to-normal vision. Demographic details (including occupation) and a brief medical history (including hand dominance) were acquired from all participants to ensure that the inclusion and exclusion criteria were satisfied. Participants were assessed for their dominant hand using the Edinburgh Handedness Inventory,23 and 17 participants were determined to be right handed. The study was conducted in accordance with the ethical standards of the committee at Kio University and the Declaration of Helsinki of 1964. This study was approved by the ethics committee at Kio University (approval number H26-25) and all participants provided written informed consent, including the use of the images accompanying this publication.
Bimanual coordination test
First, each participant was asked for basic information (ie, name, age) and instructed to remove any jewelry. After resting on a chair for 30 s three times, with a pause between each, the participant was instructed to place his/her left arm on the table so that it was reflected in a mirror and aligned in the sagittal plane. The right arm was outfitted with a portable spring balancer (HNY International Corporation, Tokyo, Japan) and placed behind the mirror (Figure 1). The participant was then asked to perform the following flexion/extension exercises with the elbow.
 | Figure 1 Photographs of the experimental setup. Notes: A mirror was placed between the participant’s right and left arms. The photographs depict the four study conditions: congruence (A), incongruence (B), intention (C), and proprioception (D). |
For the congruence condition, we asked participants to perform flexion/extension exercises with both elbows simultaneously and symmetrically in the same direction. This condition fulfilled motor intention, proprioception, and visual feedback. The motor intention was consistent with the direction of movement of the right elbow. For the incongruence condition, we asked participants to perform flexion/extension exercises with one elbow flexing and the other extending asymmetrically. This condition fulfilled the incongruence between proprioception associated with motor intention and visual feedback. For the intention condition, we asked participants to perform extension exercises with the left elbow only and to have the intention to asymmetrically move the right elbow (while, in fact, the right elbow was not moving). This represented the incongruence between motor intention and visual feedback. For the proprioception condition, we asked participants to perform flexion/extension exercises with the left elbow only, while an examiner moved the right elbow asymmetrically in flexion/extension. This condition fulfilled the incongruence between proprioception and visual feedback. For all conditions, the participant was instructed to watch the reflection of his/her hand in the mirror.
Each participant performed flexion/extension exercises for 10 s with his/her elbows and then performed each condition for 30 s, three times, with a pause between each exercise. EEGs were recorded during each condition. Each participant performed the four conditions in a random order. Between each condition, the participant rested for 5 min. During the break period after each condition, the participant was asked to rate the intensity of 14 different dysesthesia sensations on an 11-point numeric rating scale (NRS; from 0=not at all to 10=very strong) based on a questionnaire designed by Foell et al.3
EEG measurement and data analysis
We recorded EEG activity during every condition. A high-resolution, 64-channel, portable EEG System (ActiveTwo; BioSemi B.V., Amsterdam, the Netherlands) was used for data acquisition (sampling rate, 512 Hz). Electrodes were placed in an elastic head cap (BioSemi B.V.) to continuously record EEG data from 64 scalp locations organized according to the 10–20 system. The positions of electrodes were as follows: Fp1, AF7, AF3, F1, F3, F5, F7, FT7, FC5, FC3, FC1, C1, C3, C5, T7, TP7, CP5, CP3, CP1, P1, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, Pz, CPz, Fpz, Fp2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, T8, TP8, CP6, CP4, CP2, P2, P4, P6, P8, P10, PO8, PO4, and O2. Each electrode was filled with SignaGel® Electrode Gel (Parker Laboratories, Inc., Fairfield, NJ, USA) for signal transduction. We excluded electrodes from further analysis if the impedance exceeded 10 K.
ActiView software (BioSemi B.V.) was used for data collection. The EEG data were analyzed using multimodal EEG analysis software (EMSE Suite 5.4; Source Signal Imaging, La Mesa, CA, USA). EEG data were band-pass filtered in the range of 0.1–50.0 Hz and applied to a common average reference montage. We removed artifacts using previously described methods.24,25 Briefly, artifacts generated by blinking, eye movements, facial muscle activity, or body movements were removed using a specially designed spatial filter in EMSE Suite 5.4 and visual inspection of the frontal EEG trace (Fp1, Fp2). The data were segmented into 2-s segments, with an overlap of 50% accepted. Each participant accumulated 84 valid segments from a total of 135 segments. The segmented data were analyzed using power spectral analysis (Hanning window, 50%).
To quantify event-related changes in EEG power, we used a well-known procedure called ERD/event-related synchronization (ERS).26 One way to quantify event-related oscillatory EEG responses is the so-called ERD method, as originally proposed by Pfurtscheller27 and Pfurtscheller and Aranibar28 in 1977. A relative decrease in the power of a certain frequency band during stimulus processing is called ERD, whereas the opposite (ie, a relative increase in power) is called ERS. ERD/ERS values are calculated between two experimental conditions and expressed as a percentage.27,28 The ERD/ERS method is suitable for studying brain oscillatory correlates of dynamic cognitive processes, evolving in time. In the current study, the ERD/ERS of the individual alpha bands was computed using the equation ERD/ERS% = (E – R)/R × 100, where E indicates the power density at the anticipatory “each condition” and R indicates the power density at “resting on a chair with eyes open for 30 s at baseline”. In addition, two-dimensional scalp topography was determined, consisting of the high-frequency alpha rhythm ERD/ERS% for each condition using EMSE Suite 5.4 with spline interpolation.
Statistical analysis
We confirmed the normality of our data using the Shapiro–Wilk test. We compared the dysesthesia NRS results using the Friedman test, and when the difference from the standard level was <5%, we used the Wilcoxon test with Bonferroni correction to examine the differences among the four congruence/incongruence conditions. The standard level of the Wilcoxon approval was set at 0.8%. One-way analysis of variance was used to test all conditions by comparing each channel’s ERD/ERS%. A post hoc analysis using Tukey’s honest significant difference test was used to compare the congruence condition and the other conditions (ie, incongruence, intention, and proprioception) in every channel. In the congruence condition strongly causing dysesthesia only, we measured the correlation of its NRS results with the ERD/ERS% of all the conditions and channels using Spearman’s rank correlation coefficient. We used the statistical software SPSS (version 10.0J; SPSS Inc., Chicago, IL, USA) for Windows for all calculations.
Results
Comparison of dysesthesia NRS scores in the four conditions
Descriptive NRS results are shown in Figure 2. When comparing the results of dysesthesia NRS scores among the four conditions, the Friedman test showed a significant main effect on peculiarity (χ2 = 11.82, P=0.008) and nausea (χ2= 11.97, P=0.008). The mean (standard deviation) NRS scores for peculiarity were as follows: congruence condition, 0.8 (1.5); incongruence condition, 1.8 (1.7); intention condition, 1.6 (2.0); and proprioception condition, 1.7 (1.9; Figure 2). The post hoc test showed that, compared with the congruence condition, NRS scores for peculiarity were significantly higher in the incongruence (P=0.007), intention (P=0.028), and proprioception (P=0.010) conditions. For nausea, the mean (standard deviation) NRS scores were as follows: congruence condition, 0.2 (0.5); incongruence condition, 0.9 (1.2); intention condition, 1.2 (1.6); and proprioception condition, 1.5 (2.0; Figure 2). Again, compared with the congruence condition, these scores were significantly higher in the proprioception (P=0.007), incongruence (P=0.030), and intention (P=0.017) conditions.
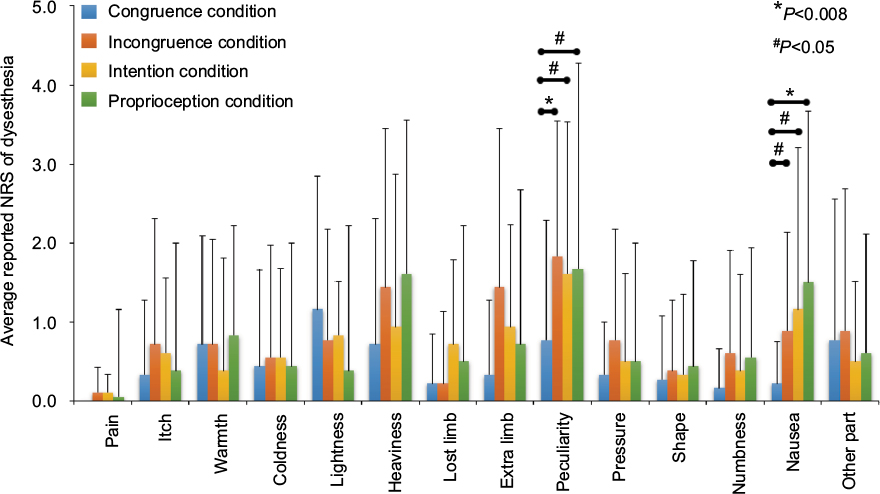 | Figure 2 A bar chart showing the degree of dysesthesia detected under the four conditions. Notes: Bars (mean [standard deviation]) represent the NRS scores for dysesthesia symptoms under the four conditions. The mean NRS scores for peculiarity and nausea were significantly higher under the incongruence, intention, and proprioception conditions than under the congruence condition. Abbreviation: NRS, numeric rating scale. |
Correlation of NRS scores for peculiarity and nausea among each condition
There were significant positive correlations between the NRS scores for peculiarity in the incongruence and intention conditions (r=0.79, P<0.01) and in the incongruence and proprioception conditions (r=0.87, P<0.01). There was also a significant positive correlation between the NRS scores for nausea in the incongruence and intention conditions (r=0.55, P<0.05) and in the incongruence and proprioception conditions (r=0.58, P<0.05; Figure 3).
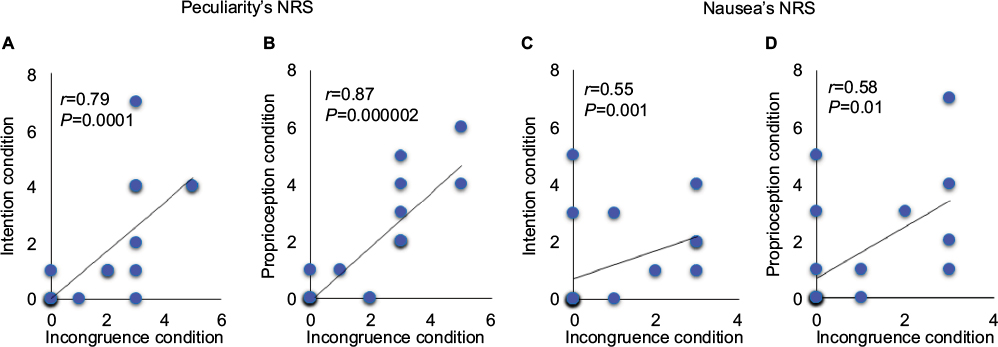 | Figure 3 Correlation analysis of NRS scores for peculiarity (A and B) and nausea (C and D) with each condition. Notes: NRS scores for peculiarity under the incongruence condition were significantly correlated with scores under the intention and proprioception conditions (A and B). In addition, a significant positive correlation was found between NRS scores for nausea under the incongruence and intention conditions (C) and under the incongruence and proprioception conditions (D). Abbreviation: NRS, numeric rating scale. |
Topographic maps of ERD/ERS% of the high-frequency alpha rhythm band
There was no significance seen when comparing each channel’s ERD/ERS% in any condition. However, an inhibited high-frequency alpha rhythm (ie, increased ERD) was observed in several regions. In the congruence condition, ERD of the high-frequency alpha band was observed in the bilateral parietal regions (CP3, CP4). We also found increased ERD in the frontal region (AF3, AFz, AF4) in the incongruence, intention, and proprioception conditions, and a small difference in the parietal region among all the conditions. ERD increased on both sides (CP3, P3, CP4, P4) in the incongruence condition, on the left side (CP3, P3) in the intention condition, and on the right side (CP4, P4) in the proprioception condition (Figure 4).
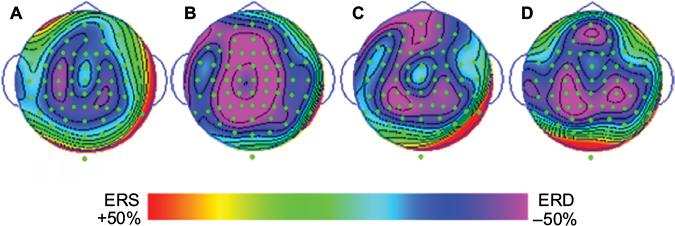 | Figure 4 Topographic maps of ERD/ERS% of the high-frequency alpha rhythm band in the congruence (A), incongruence (B), intention (C), and proprioception (D) conditions. Abbreviations: ERD, event-related desynchronization; ERS, event-related synchronization. |
Correlation of dysesthesia NRS scores and ERD/ERS%
In the high-frequency alpha band, there were significant negative correlations between the NRS scores for peculiarity and ERD/ERS% in the incongruence condition’s P8 channel (right inferior parietal region; r=–0.63, P<0.01), and in the proprioception condition’s TP8 (r=–0.49, P<0.05) and P10 (r=–0.50, P<0.05) channels (right temporoparietal region; Figure 5). There were no significant correlations in similar regions of the incongruence or intention conditions.
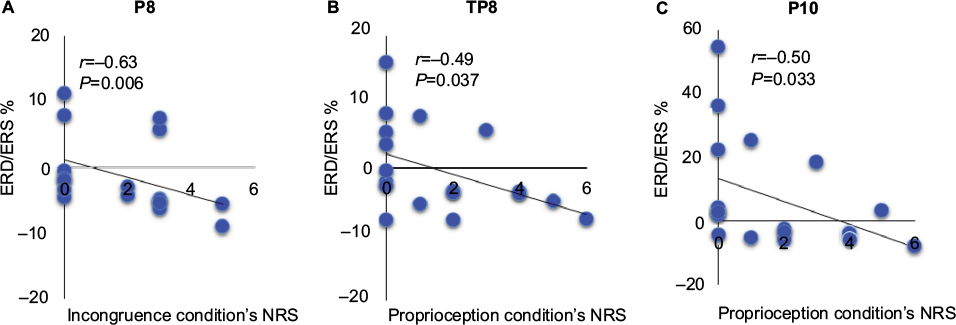 | Figure 5 Correlation analysis of NRS scores for peculiarity and ERD/ERS% in the P8 channel (right inferior parietal region [A]) under the incongruence condition and the TP8 and P10 channels (right temporoparietal region [B and C]) under the proprioception condition. Abbreviations: ERD, event-related desynchronization; ERS, event-related synchronization; NRS, numeric rating scale. |
Discussion
In this study, we investigated the main factors causing dysesthesia related to motor intention, proprioception, and visual feedback. We found that dysesthesia signs of peculiarity and nausea were elicited under conditions of incongruence, intention, and proprioception. However, there were no significant differences between each channel’s ERD/ERS% in any condition. This is because there was incomplete congruence between sensation and movement in all conditions, including the congruence condition.
As in this study, previous studies have reported some dysesthesia in all conditions. We interpret this as meaning that it is impossible to achieve complete congruence under any condition. In the current study, participants in the congruence condition were asked to perform symmetrical flexion/extension exercises with the left arm in front of a mirror and the right arm hidden behind the mirror. However, the real position of the left arm will not be completely congruent with that of the right arm, and we believe that this may cause an extremely small incongruence between proprioception and visual feedback. This may account for the differences in ERD/ERS% channels in every condition.
In addition, it is possible that incongruence between motor intention and visual feedback caused the increased ERD of the high-frequency alpha band in the left parietal region in the incongruence and intention conditions. Furthermore, in the incongruence and the proprioception conditions, incongruence between visual feedback and proprioception resulted in an ERD increase in the right parietal region. Moreover, there was a significant negative correlation between increasing high-frequency alpha bands and subjective perception of dysesthesia.
Dysesthesia NRS scores in the four conditions
Dysesthesia symptoms appeared in all conditions. High NRS scores were observed for peculiarity and nausea compared with other dysesthesia symptoms. We also observed a significant positive correlation in the degree of peculiarity and nausea symptoms between the incongruence condition and the intention and proprioception conditions. Thus, we were able to distinguish the intention and proprioception conditions associated with the presence or absence of motor intention. As a result, we determined that these dysesthesia symptoms were caused only by intention-feedback incongruence or spatial incongruence.
McCabe et al2 and Wall29 have stated that dysesthesia is caused by a warning reaction to sensorimotor incongruence. In line with this, we believe that the dysesthesias observed in the present study were caused by a system alert to sensorimotor incongruence. In addition, McCabe et al2 have stated that an early warning system alerts the individual to abnormalities in information processing and increases negative psychological changes such as threat perception, which ultimately produce pain. Moreover, these monitoring mechanisms can be triggered by conflicts from external (eg, incongruent movements viewed in the mirror) or internal (eg, the aging process leading to inaccurate execution of movements and altered proprioception or disease damage in rheumatoid arthritis, resulting in stiffness) sources. For the incongruence, intention, and proprioception conditions used in the current study, we used external conflict trigger (ie, incongruence between visual feedback from the mirror and the movement) of McCabe et al.2 Therefore, the fact that our participants experienced dysesthesia due to the system of monitoring conflicts corresponds with the findings from previous studies. McCabe et al2 reported that sensorimotor incongruence is the cause of 15% of pain in victims. However, Foell et al3 reported that the perception of pain due to sensorimotor incongruence is less than in other dysesthesias. In addition, Wand et al30 reported that pain does not appear with sensorimotor incongruence and that the pressure pain threshold does not change. Clinical studies have reported that dysesthesia is more severe in patients who are already in pain and who are made to move in conditions of sensorimotor incongruence compared with healthy individuals. In our study, patients reported lower NRS scores for pain compared with other dysesthesias. Therefore, we conclude that it is not sensorimotor incongruence that provokes pain but rather other dysesthesias.
Relationship between brain activity and dysesthesia
In this study, analysis of brain waves revealed significant negative correlations between NRS scores for peculiarity and changes in alpha oscillation in the right inferior parietal region under the incongruence condition and in the right temporoparietal region under the proprioception condition. Shimada et al31 observed an increase in activity in the right inferior parietal lobe with temporal incongruence, causing an image delay between visual feedback and proprioceptive feedback during the use of near-infrared spectroscopy. Balslev et al32 also reported an increase in activity in the right temporoparietal junction in a case of spatial incongruence between movement and visual feedback, as assessed by functional magnetic resonance imaging (fMRI). In addition, in a study on the mechanisms of self-recognition, Jeannerod33 demonstrated that self-recognition of a person’s own body part is possible in a situation of congruence between proprioception and visual feedback, but not without this congruence. From this neuroscientific evidence, the temporoparietal region is believed to be related to the function of detecting incongruence and the distinction between oneself and others. In the current study, in the proprioception condition, we detected congruence and incongruence between the mirror reflection of the left upper limb (visual feedback) and the hidden right upper limb (proprioceptive feedback). Hence, we assume that distinguishing one’s own or another’s body provokes activity in the right temporoparietal region.
In addition, we suggest that incongruence in body representation can be a factor in dysesthesia. Body representation is the body that appears in our brain and that can be perceived with proprioception of our body, even with our eyes closed. However, it can also be perceived with our vision. In other words, body representation is structured by the process of integration between proprioception and vision. Naito et al34 revealed that the right inferior parietal lobule and the inferior frontal gyrus are related to body representation. Berlucchi and Aglioti35 also reported that the posterior parietal cortex is associated with structuring body representation. Our results show that in the incongruence, intention, and proprioception conditions, in contrast to the congruence condition, there is incongruence between the body representation based on the somatosensory system and that based on visual perception and that this can provoke dysesthesia symptoms of peculiarity and nausea and can activate the parietal region. Fink et al36 have previously confirmed an increase in brain activity in the right hemisphere’s dorsolateral prefrontal cortex and posterior parietal cortex using positron emission tomography (PET). Moreover, Nishigami et al37 reported inhibition of the alpha rhythm in the right hemisphere’s posterior parietal cortex, similar to that seen in the incongruence condition in the present study. Therefore, we suggest that in the incongruence condition with motor intention, the proprioception-provoking efference copy and actual movement, coupled with incongruence in visual feedback, preserve the integrity of one’s own destroyed body representation in the right hemisphere’s inferior parietal region. The right temporoparietal region, in which a correlation was observed in NRS scores of peculiarity during the proprioception condition, is a region linked with the theory of mind, empathy, sense of agency, and attention orientation.38 In studies by Farrer and Frith39 and by Farrer et al,40 in cases of incongruence between one’s own movement and sensation feedback, an increase in the activity of the angular gyrus (which is situated in the temporoparietal junction) and the right hemisphere’s inferior parietal lobe was observed during incongruence in visual feedback, as assessed using fMRI and PET.
A limitation of our study is that we could not confirm whether the increased ERD of the high-frequency alpha bands detected in the temporoparietal and right hemisphere’s parietal regions during the incongruence and proprioception conditions was caused by the process of sensory integration or by restructuring of the body representation. Furthermore, we analyzed power spectral density using brain waves for 30 s for each condition; hence, we could not evaluate brain activity at the instant of provoked dysesthesia. In future studies, we will change the configuration of the bimanual coordination test, timing protocols, and EEG analysis methods to more clearly reveal the neural mechanisms of dysesthesia.
Conclusion
We suggest that our results support the previously reported neural mechanisms behind dysesthesia, which is thought to be caused by incongruence between motor intention, proprioception, and visual feedback; in particular, we found that the dysesthesia symptom of “peculiarity” was caused by incongruence between proprioception and visual feedback. Moreover, we observed that the neural mechanisms provoking dysesthesia in the intention condition were different from those related to the incongruence and proprioception conditions. Therefore, with this study’s conditions in mind and assuming that the results would be the same for healthy individuals and patients in chronic pain, we believe that evaluating incongruence between motor intention, proprioception, and visual feedback is important for treating patients in clinical practice.
Acknowledgments
The authors would like to thank the students of the Graduate School of Kio University for their participation in this study. We would also like to thank Ammor Adam Karim for the English translation.
Disclosure
The authors report no conflicts of interest in this work.
References
Harris AJ. Cortical origin of pathological pain. Lancet. 1999;354(9188):1464–1466. | ||
McCabe CS, Haigh RC, Halligan PW, Blake DR. Simulating sensory-motor incongruence in healthy volunteers: implications for a cortical model of pain. Rheumatology. 2005;44(4):509–516. | ||
Foell J, Bekrater-Bodmann R, McCabe CS, Flor H. Sensorimotor incongruence and body perception: an experimental investigation. Front Hum Neurosci. 2013;7:310. | ||
McCabe CS, Cohen H, Blake DR. Somaesthetic disturbances in fibromyalgia are exaggerated by sensory–motor conflict: implications for chronicity of the disease? Rheumatology. 2007;46(10):1587–1592. | ||
Daenen L, Nijs J, Roussel N, Wouters K, Loo MV, Cras P. Sensorimotor incongruence exacerbates symptoms in patients with chronic whiplash associated disorders: an experimental study. Rheumatology. 2012;51(8):1492–1499. | ||
Daenen L, Roussel N, Cras P, Nijs J. Sensorimotor incongruence triggers sensory disturbances in professional violinists: an experimental study. Rheumatology. 2010;49(7):1281–1289. | ||
Ramachandran VS, Rogers-Ramachandran D. Synaesthesia in phantom limbs induced with mirrors. Proc Biol Sci. 1996;263(1369):377–386. | ||
Sumitani M, Miyauchi S, McCabe CS, et al. Mirror visual feedback alleviates deafferentation pain, depending on qualitative aspects of the pain: a preliminary report. Rheumatology. 2008;47(7):1038–1043. | ||
Giraux P, Sirigu A. Illusory movements of the paralyzed limb restore motor cortex activity. Neuroimage. 2003;20(suppl 1):S107–S111. | ||
Mercier C, Sirigu A. Training with virtual visual feedback to alleviate phantom limb pain. Neurorehabil Neural Repair. 2009;23(6):587–594. | ||
Moseley GL. Using visual illusion to reduce at-level neuropathic pain in paraplegia. Pain. 2007;130(3):294–298. | ||
Soler MD, Kumru H, Pelayo R, et al. Effectiveness of transcranial direct current stimulation and visual illusion on neuropathic pain in spinal cord injury. Brain. 2010;133(9):2565–2577. | ||
Katayama O, Iki H, Sawa S, Osumi M, Morioka S. The effect of virtual visual feedback on supernumerary phantom limb pain in a patient with high cervical cord injury: a single-case design study. Neurocase. 2015;21(6):786–792. | ||
McCabe CS, Haigh RC, Ring EFR, Halligan PW, Wall PD, Blake DR. A controlled pilot study of the utility of mirror visual feedback in the treatment of complex regional pain syndrome (Type 1). Rheumatology. 2003;42(1):97–101. | ||
Moseley GL. Graded motor imagery is effective for long-standing complex regional pain syndrome. Pain. 2004;108(1–2):192–198. | ||
Moseley GL. Is successful rehabilitation of complex regional pain syndrome due to sustained attention to the affected limb? A randomised clinical trial. Pain. 2005;114(1–2):54–61. | ||
Moseley GL. Graded motor imagery for pathologic pain: a randomized controlled trial. Neurology. 2006;67(12):2129–2134. | ||
Klimesch W. Memory processes, brain oscillations and EEG synchronization. Int J Psychophysiol. 1996;24(1–2):61–100. | ||
Klimesch W. EEG alpha and theta oscillations reflect cognitive and memory performance. a review and analysis. Brain Res Brain Res Rev. 1999;29(2–3):169–195. | ||
Klimesch W, Doppelmayr M, Russegger H, Pachinger T, Schwaiger J. Induced alpha band power changes in the human EEG and attention. Neurosci Lett. 1998;244(2):73–76. | ||
Pfurtscheller G, Neuper C. Motor imagery activates primary sensorimotor area in humans. Neurosci Lett. 1997;239(2–3):65–68. | ||
Pfurtscheller G, Neuper C, Andrew C, Edlinger G. Foot and hand area mu rhythms. Int J Psychophysiol. 1997;26(1–3):121–135. | ||
Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9(1):97–113. | ||
Ishigaki T, Ueta K, Imai R, Morioka S. EEG frequency analysis of cortical brain activities induced by effect of light touch. Exp Brain Res. 2016;234(6):1429–1440. | ||
Debnath R, Franz EA. Perception of hand movement by mirror reflection evokes brain activation in the motor cortex contralateral to a non-moving hand. Cortex. 2016;81:118–125.26. | ||
Pfurtscheller G, Lopes da Silva FH. Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol. 1999;110(11):1842–1857. | ||
Pfurtscheller G. Graphical display and statistical evaluation of event-related desynchronization (ERD). Electroencephalogr Clin Neurophysiol. 1977;43(5):757–760. | ||
Pfurtscheller G, Aranibar A. Event-related cortical desynchronization detected by power measurements of scalp EEG. Electroencephalogr Clin Neurophysiol. 1977;42(6):817–826. | ||
Wall PD. Introduction. In: Wall PD, Melzack R, editors. Textbook of Pain. 4th ed. Edinburgh: Churchill Livingston; 1999:165–181. | ||
Wand BM, Szpak L, George PJ, Bulsara MK, O’Connell NE, Moseley GL. Moving in an environment of induced sensorimotor incongruence does not influence pain sensitivity in healthy volunteers: a randomised within-subject experiment. PLoS One. 2014;9(4):e93701. | ||
Shimada S, Hiraki K, Oda I. The parietal role in the sense of self-ownership with temporal discrepancy between visual and proprioceptive feedbacks. Neuroimage. 2005;24(4):1225–1232. | ||
Balslev D, Nielsen FA, Paulson OB, Law I. Right temporoparietal cortex activation during visuo-proprioceptive conflict. Cereb Cortex. 2005;15(2):166–169. | ||
Jeannerod M. The mechanism of self-recognition in humans. Behav Brain Res. 2003;142(1–2):1–15. | ||
Naito E, Nakashima Y, Kito T, Aramaki Y, Okada T, Sadato N. Human limb-specific and non-limb-specific brain representations during kinesthetic illusory movements of the upper and lower extremities. Eur J Neurosci. 2007;25(11):3476–3487. | ||
Berlucchi G, Aglioti SM. The body in the brain revisited. Exp Brain Res. 2010;200(1):25–35. | ||
Fink GR, Marshall JC, Halligan PW, et al. The neural consequences of conflict between intention and the senses. Brain. 1999;122(pt 3):497–512. | ||
Nishigami T, Nakano H, Osumi M, Tsujishita M, Mibu A, Ushida T. Central neural mechanisms of interindividual difference in discomfort during sensorimotor incongruence in healthy volunteers: an experimental study. Rheumatology. 2014;53(7):1194–1199. | ||
Decety J, Lamm C. The role of the right temporoparietal junction in social interaction: how low-level computational processes contribute to meta-cognition. Neuroscientist. 2007;13(6):580–593. | ||
Farrer C, Frith CD. Experiencing oneself vs another person as being the cause of an action: the neural correlates of the experience of agency. Neuroimage. 2002;15(3):596–603. | ||
Farrer C, Franck N, Georgieff N, Frith CD, Decety J, Jeannerod M. Modulating the experience of agency: a positron emission tomography study. Neuroimage. 2003;18(2):324–333. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.