Back to Journals » OncoTargets and Therapy » Volume 12
DPY30 regulates cervical squamous cell carcinoma by mediating epithelial–mesenchymal transition (EMT)
Authors He F, Zhang L ![]() , Jin P, Liu D, Li A
, Jin P, Liu D, Li A
Received 19 March 2019
Accepted for publication 1 July 2019
Published 2 September 2019 Volume 2019:12 Pages 7139—7147
DOI https://doi.org/10.2147/OTT.S209315
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Feng-xi He,1,* Li-li Zhang,1,* Peng-fei Jin,1 Dan-dan Liu,2 Ai-hua Li1
1Department of Obstetrics and Gynecology, Liaocheng People’s Hospital Affiliated to Shandong First Medical University, Liaocheng 252000, People’s Republic of China; 2Shandong First Medical University, Taian 271016, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ai-hua Li
Department of Obstetrics and Gynecology, Liaocheng People’s Hospital Affiliated to Shandong First Medical University, No. 67 Dongchang West Road, Liaocheng City, Shandong Province 252000, People’s Republic of China
Tel +86 1 356 205 0087
Email [email protected]
Introduction: Set1/MLL complexes are the main histone H3K4 methyltransferases and are crucial regulators of tumor pathogenesis. DPY30 is a fairly uncharacterized protein in the Set1/MLL complex, but it has been reported to regulate tumor growth. However, the exact mechanism by which DPY30 mediates the progression of cervical squamous cell carcinoma (CSCC) remains unknown. In the present study, we investigated the role of DPY30 in CSCC at a molecular level.
Methods: We obtained normal cervical and cervical cancer tissue samples from patients. We used immunohistochemistry and real-time polymerase chain reaction (PCR) to detect DPY30 expression in CSCC tissues. In addition, we used the human cervical cancer cell line to evaluate expression levels of DPY30 and epithelial–mesenchymal transition (EMT) markers in vitro.
Results: Immunohistochemical and real-time PCR analyses showed that DPY30 expression was upregulated in tissue samples from patients with CSCC and that DPY30 levels were associated with EMT markers such as E-cadherin. Furthermore, knock-down of DPY30 by siRNA resulted in a decrease in the proliferation, migration, and invasion of CSCC cells. We also found that DPY30-induced EMT is mediated by the Wnt/β-catenin signaling pathway.
Conclusion: Our results suggest that elevated DPY30 levels may contribute to EMT by activating Wnt/β-catenin signaling in the progression of CSCC.
Keywords: DPY30, cervical squamous cell carcinoma, epithelial–mesenchymal transition, Wnt/β-catenin pathway
Introduction
Cervical cancer is the second most common form of cancer in developing countries and the third most common cause of cancer-related deaths among women.1 The persistent high incidence of cervical cancer and recent trend of decrease in the age of onset poses a significant threat to women’s health.1 The rate of occurrence of cervical cancer has been reported to be high in Uganda and in some Eastern European countries.2 Therefore, there is still a need to develop effective therapeutic strategies for cervical cancer.
DPY30 belongs to the human SET1/MLL complex and is an essential protein required for the activity of SET1/MLL methyltransferase.3,4 DPY30 and its homologs are mainly localized in the nuclei of cells; they are conserved across various species such as yeast, Caenorhabditis elegans, and humans.5 The protein encoded by DPY30 directly controls cell cycle regulators and plays a crucial role in cell proliferation and differentiation. Previous studies have shown that this protein is involved in the regulation of H3K4 methylation, whereby it plays a critical role in the proliferation, invasion, and migration of cells in gastric carcinoma.6 Similarly, DPY30 is known to mediate the differentiation of embryonic stem cells3 and hematopoietic progenitor cells, as well as the proliferation of the latter.4 Moreover, DPY30 has been shown to influence the expression of inhibitors of DNA-binding/differentiation proteins, whereby DPY30 plays a role in regulating cell senescence.7
EMT is a process where in epithelial cells transform into mesenchymal cells by a process involving many reversible and rapid gene changes. An important feature of EMT is the decreased expression of epithelial cells markers, such as E-cadherin, and increased expression of mesenchymal cell markers, such as N-cadherin and vimentin.8 Recent studies have shown that EMT is crucial for the growth and spread of cervical carcinoma.9 Thus far, the exact mechanism underlying the process of EMT remains unclear. The Wnt/β-catenin signaling pathway is well-known for its role in the de-differntiation and proliferation of cancer cells.10 Abnormal activation of this pathway is believed to contribute to the growth of some tumors, particularly epithelial malignancies.11
In this study, we determined whether DPY30 contributes to the progression of CSCC by inducing EMT, as well as investigated the underlying mechanisms in vitro.
Materials and methods
Patients and tissue specimens
We obtained paraffin-embedded tissue samples from patients with CSCC (n=65; mean age 47.3 y; range 27–69 y) and subjects with normal cervical architecture (n=10). All samples were collected between January 2010 and January 2013 at the Pathology Department of Liaocheng People’s Hospital. The tissue samples were matched with the demographic and clinical data of the enrolled patients, and the subjects were classified into different groups based on various parameters, including age, histological type, Federation International of Gynecology and Obstetrics (FIGO) stage, pathological grade, and lymph node metastasis. In addition, 20 pairs of fresh tissue samples of cervical cancer and adjacent normal tissues were obtained from patients who underwent surgery at the Department of Gynecology of Liaocheng People’s Hospital between September 2016 and September 2017. The fresh specimens were stored at −80 °C within 30 min of collection.
Cell culture and siRNA transfection
The human cervical cancer cell line, SiHa, was acquired from the Central Laboratory of Liaocheng People’s Hospital,which was approved by the the ethics committee of Liaocheng People’s Hospital. The cells were then transferred to RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) with 10% fetal bovine serum and cultured at 37 °C in a humidified incubator maintained at atmospheric conditions of 5% CO2. Cells were grown in a 6-well plate, and upon reaching 80% confluence, cells were transfected with either synthesized siRNA (DPY30-siRNA group) or control siRNA sequences (DPY30-NC group) using Lipofectamine® 2000 according to the manufacturer’s instructions. The siRNA sequence of DPY30 was as follows: 5′- GCAGAAGGUAGAUCUCCAGTT-3′.
Wound-healing assay
Cells were transferred to a 6-well plate and cultured until reaching 80–90% confluence. A wound was scratched with a 200 μL pipette tip. The exfoliated cells were washed with phosphate-buffered saline. Under the same culture conditions described above, cells were grown in RPMI-1640 medium to allow for wound healing. Images were acquired at 24-h intervals (0, 24, and 48 h) to assess cell migration.
Transwell invasion
To evaluate cell invasion and migration, cells were cultured in the presence or absence of basement membrane matrix (BD Biosciences, Franklin Lakes, NJ, USA) according to the manufacturer’s instructions. The transfected cells were cultured in the upper chamber (3×104 cells/100 μL) with serum-free medium, while cells in the lower chambers were cultured in 20% fetal bovine serum (8-mm pore; Corning, Life Science, Lowell, MA, USA). The cells were incubated for 24 h at 37 °C with 5% CO2 and then stained with 1% crystal violet (20 min; room temperature); cell numbers in five random regions were counted. The extent of cell invasion and cell migration was determined as mean ± standard deviation of the number of cells per field.
CCK-8 assay
The CCK8 assay was used to assess proliferation of the SiHa cells. First, we adjusted the density of the transfected cells to 3×104/mL. The cells were then transferred to a 96-well plate in 100 μL per well. All experiments were performed in triplicate for the DPY30-siRNA group and the DPY30-NC group. Cells were incubated for 0, 24, 48, and 72 h. Mixtures of CCK-8 and serum-free medium (1:10) were prepared, and 110 μL of the mixture was placed in each well. The optical density was measured at a 450-nm wavelength using a microplate reader (Epoch; BioTek Instruments, Inc., Winooski, VT, USA).
Real-time polymerase chain reaction
Total RNA from the cells and tissue samples was extracted using Trizol (Invitrogen). Reverse transcription was performed according to the guidelines provided by the PrimeScript™ RT reagent Kit with gDNA Eraser (RR047A, Takara). Quantitative real-time PCR was performed to determine the levels of DPY30 mRNA. The primer sequences of DPY30 were as follows: 5′-AAC GCA GGT TGC AGA AAA TCC T −3′ and 5′-TCT GAT CCA GGT AGG CAC GAG −3′. The analysis was performed using the Biosystems 7500 Real-Time PCR System and TB Green™ Premix Ex Taq™ II (Tli RNaseH Plus) (RR820A, Takara), according to the manufacturer’s instructions. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the endogenous control.
Immunohistochemistry
We sectioned the paraffin-embedded tissues to a thickness of 4 μM. DPY30 and E-cadherin expression levels were determined using immunohistochemical staining. The antibodies used for immunohistochemistry were rabbit anti-DPY30 antibody (1:200, Abcam) and rabbit anti-E-cadherin monoclonal antibody (ZSGB-BIO). The procedure was performed according to the instructions of the rabbit two-step kit (PV6001, ZSGB-BIO). Two experienced pathologists, blinded to the subject’s clinical conditions, analyzed the immunohistochemistry results. The semi-quantitative scoring method was used to evaluate the staining intensity of tissues and the percentage of stained cells in hundreds. DPY30 mainly showed nuclear staining, while E-cadherin mainly showed membrane staining. The degree of staining was scored according to the color: 0 for colorless, 1 for light yellow or yellow, 2 for brown or brown, and 3 for dark brown. According to the percentage of stained cells, the scores were as follows: 0 (0–5%), 1 (6–25%), 2 (26–50%), 3 (51–75%) 4 (76–100%). The total score was taken as the product of the two scores. If the score was less than 3, the expression was considered negative, while it was considered positive if the score was greater than or equal to 3.
Western blot assay
Transfected cells were cultured in 6-well plates (density, 3×105 cells/mL) and lysed with lysis buffer (1 μL PMSF, added to 100 μL RIPA) on ice for 30 min, followed by centrifugation at 4 °C for 5 min. The supernatant was collected and protein-loading buffer was added, followed by boiling for 6 min at 100 °C. After protein extraction, SDS-PAGE was used to separate the proteins and transfer them to a poly-vinylidene fluoride membrane (EMD Millipore, Billerica, MA, USA). We blocked the membranes in 5% skim milk at room temperature for 1 h, followed by overnight incubation at 4 °C with specific antibodies. The bound antibodies were detected using secondary horseradish peroxidase-conjugated antibodies, and the protein bands were visualized using an enhanced chemiluminescence system (ProteinSimple, San Jose, CA, USA). The protein bands were then analyzed using Quantity One software Version 4.0.1 (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Experiments were performed in triplicate.
Statistical analysis
Statistical analyses were performed using the SPSS version 22.0 software. Data from the cell experiments are expressed as mean ± SD and analyzed using one-way ANOVA. Intergroup differences were assessed using Student’s t-test. Immunohistochemistry findings were analyzed using the Chi-square test and Pearson's rank correlation test. A P-value of less than 0.05 was considered statistically significant.
Results
DPY30 protein expression in CSCC is elevated and associated with E-cadherin expression
Our immunohistochemistry results showed that DPY30 expression in CSCC tissues was significantly greater than that in normal cervical tissue (***P<0.001; Table S1 and Figure 1). Among the 65 CSCC samples, 43 had varying degrees of positive staining for DPY30. However, DPY30 staining was negative in the 10 normal cervical tissue samples (Table S1 and Figure 1). A comparison of the clinicopathological and immunohistochemistry data of the 65 CSCC samples revealed that DPY30 expression significantly correlated with the depth of interstitial invasion (P=0.006) and the grade of cell differentiation (P=0.000), irrespective of the patient’s age, FIGO stage, and presence or absence of lymph node metastasis (Table 1). These findings indicate that DPY30 expression increases with CSCC progression and that expression of this protein may be related to the invasiveness of the carcinoma.
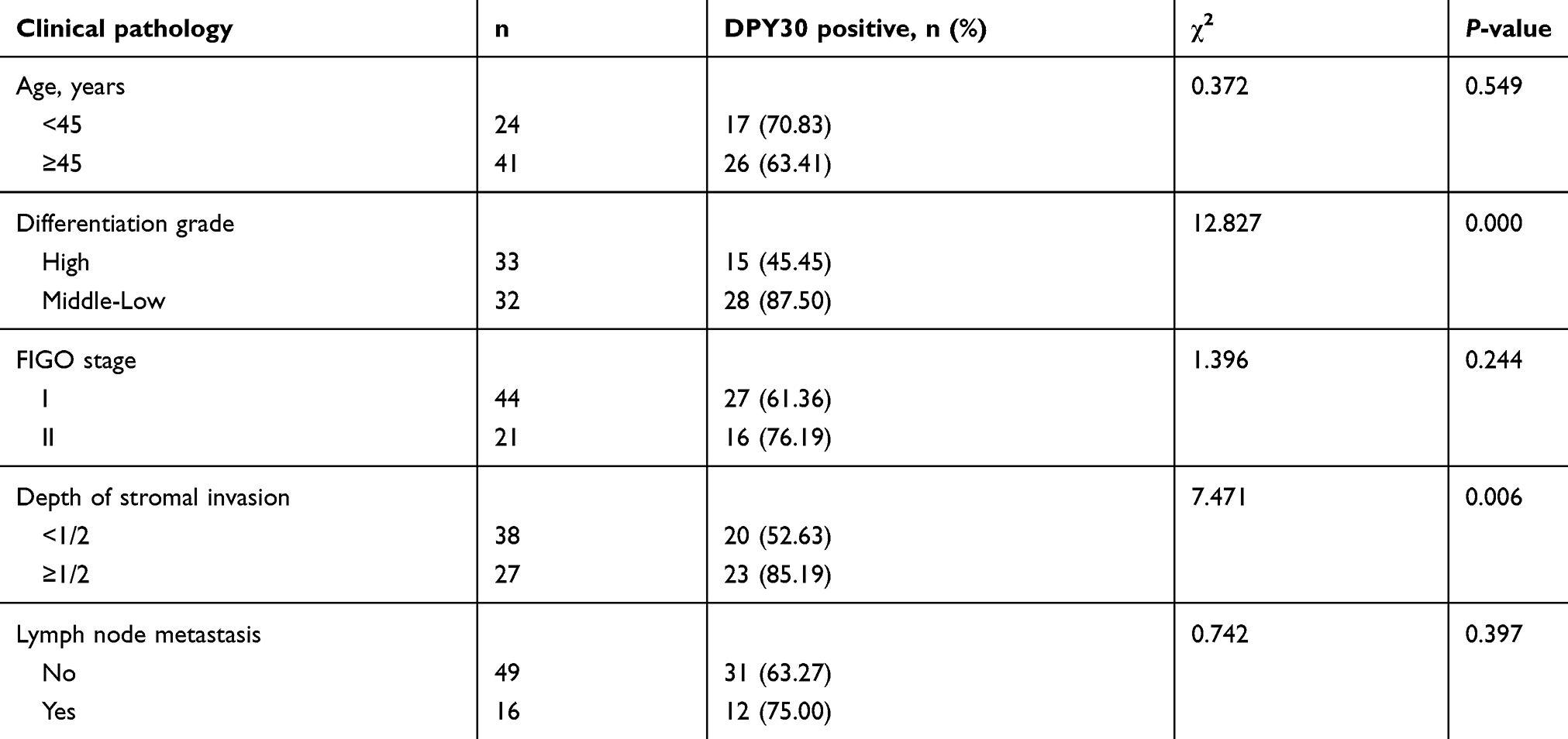 |
Table 1 Relationship between DPY30 expression and clinicopathological characteristics of CSCC |
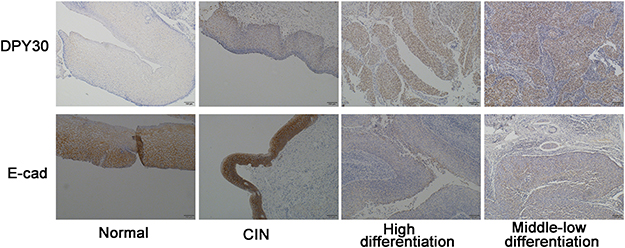 |
Figure 1 DPY30 protein expression increased and correlated with E-cadherin expression in CSCC. Representative immunohistochemistry staining results for DPY30 and E-cadherin in human normal cervical and CSCC tissues. Scale bar=100 μM. |
Additionally, we found a decrease in the EMT marker E-cadherin in the 65 CSCC samples (Table S2). Thus, we analyzed the association between DPY30 and E-cadherin expression (Table 2). Representative samples of immunohistochemical staining of CSCC tissue samples for E-cadherin are shown in Figure 1. The results show that E-cadherin expression decreased with an increase in DPY30 expression (χ2=5.240, c =0.273, r = – 0.284, P=0.022; Table 2), indicating that DPY30 influences EMT in CSCC.
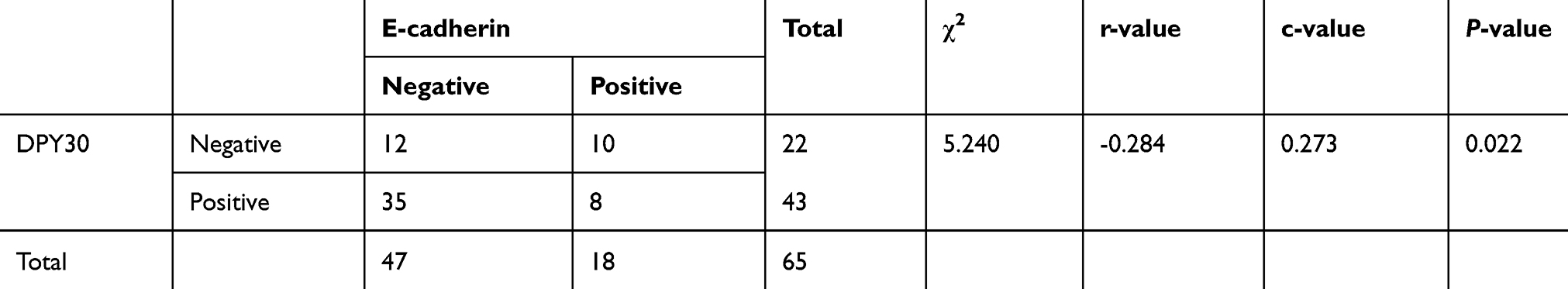 |
Table 2 Statistical analysis of DPY30 and E-cadherin correlation based on immunohistochemistry staining of CSCC tissues |
DPY30 mRNA is upregulated in CSCC tissues
The results of real-time PCR suggested that DPY30 expression was significantly greater in CSCC samples than in the control samples (Figure 2). This suggests that increased DPY30 expression may be an important factor in CSCC development.
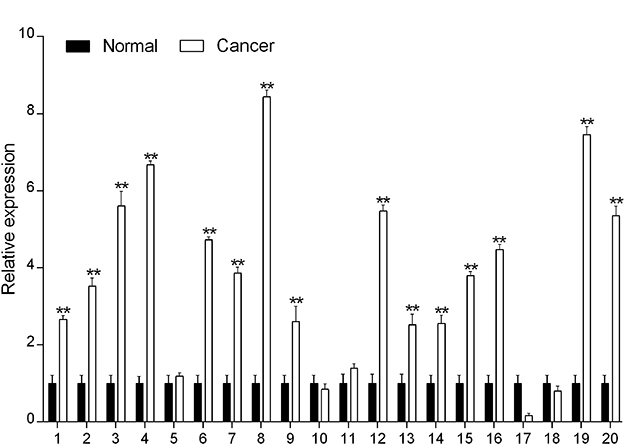 |
Figure 2 Over-expression of DPY30 in CSCC. DPY30 mRNA expression in 20 paired CSCC tissues was measured using real-time PCR. **P<0.01 (Student’s t-test, versus normal). |
DPY30 promotes CSCC cell proliferation, migration, and invasion in vitro
To determine the mechanism by which DPY30 influences the pathogenesis and progression of CSCC, we transfected DPY30 siRNA into SiHa cells to knock-down DPY30 expression. Real-time PCR results showed that the use of DPY30-siRNA reduced the DPY30 mRNA to levels significantly lower than those observed with control siRNA (Figure 3A).
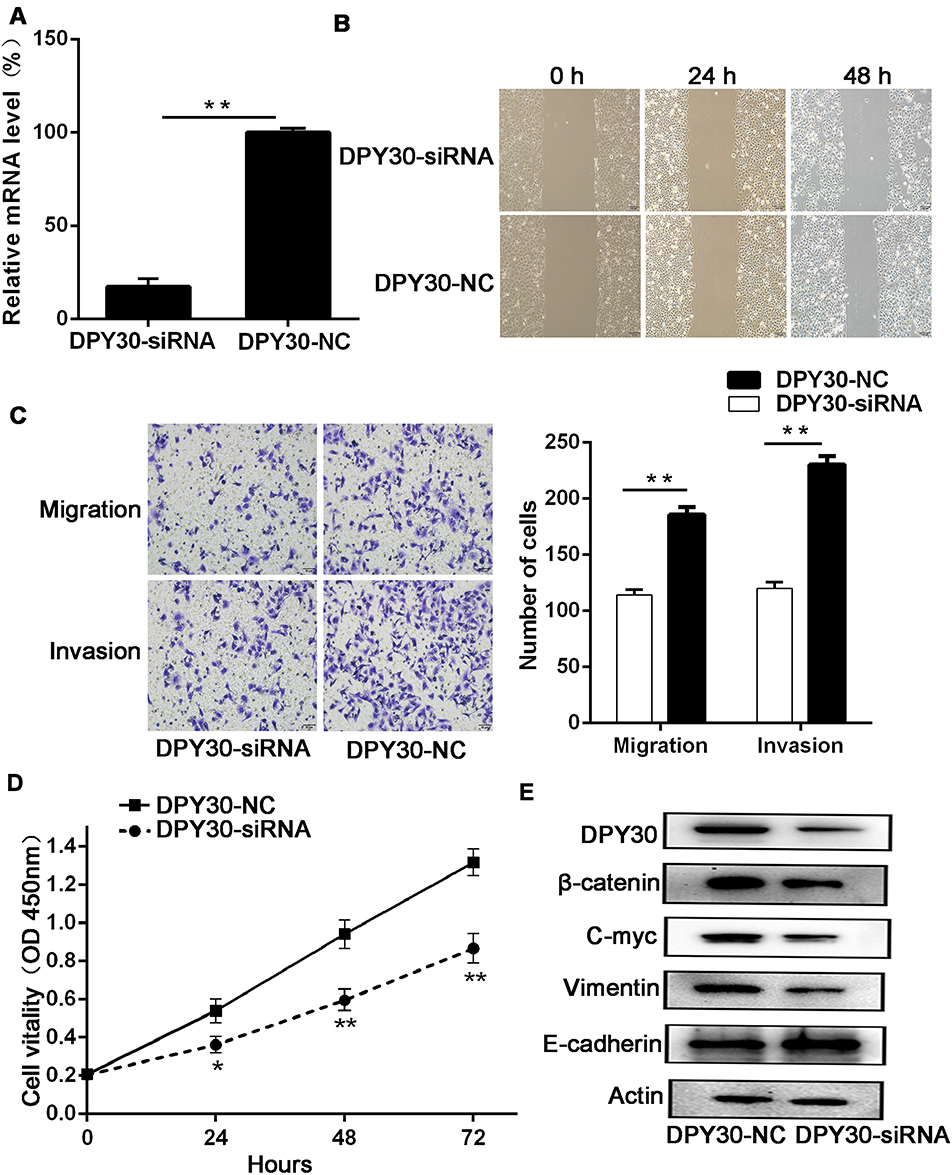 |
Figure 3 DPY30 promotes CSCC cell migration, invasion, and proliferation in vitro and the relationship between DPY30 protein and EMT-related proteins. (A) We knocked down DPY30 expression using DPY30-siRNA and monitored knock-down efficiency using real-time PCR. **P<0.01, vs DPY30-NC. (B) DPY30 expression enhanced Siha cell migration in the wound healing assay. The wound healing rates were calculated at the indicated time points of 0, 24, and 48 h. Scale bar =100 μM. (C) DPY30 promoted migration and invasion of CSCC cells in the Transwell assay. Scale bar =50 μM. Data are all presented as mean ± SD from three independent experiments. *P<0.05; **P<0.01. (D) Effects of DPY30 on CSCC cell proliferation following transfection with siRNA in Siha cells. The CCK-8 assay was used to detect changes in cell viability. Data are presented as mean ± SD from three independent experiments, *P<0.05, **P<0.01; OD, optical density. (E) Western blot analysis showing the relationships between DPY30 expression and expression of EMT-markers (E-cadherin and Vimentin), β-catenin, and c-myc. |
We then assayed the effects of DPY30 on cell migration using the wound-healing and Transwell-migration assays. The results of the wound-healing assay revealed that decreased DPY30 expression significantly accelerated the migration of SiHa cells as compared to normal cells (Figure 3B). In agreement with this finding, knock-down of DPY30 decreased the serum-induced migration of SiHa cells (Figure 3C). In the invasion assay, DPY30-siRNA inhibited the serum-induced SiHa cell invasion to an extent greater than that in the DPY30-NC group (Figure 3C). These findings suggest that DPY30 facilitates cell migration and tumor invasion of CSCC cells.
Using the CCK-8 assay, we found that knock-down of DPY30 expression markedly reduced proliferation (Figure 3D), which indicates that DPY30 promotes CSCC cell proliferation.
DPY30 induces EMT in CSCC cells
EMT is characterized by enhanced motility and cytoskeletal reorganization.12,13 Given our data showing that DPY30 enhances migration and invasion of SiHa cells, we further investigated the molecular mechanisms by which DPY30 influences these processes by assessing the levels of EMT markers. Western blot analysis showed that levels of the epithelial factor E-cadherin were significantly higher, while levels of the mesenchymal factor vimentin were markedly lower in the DPY30-siRNA group than the DPY30-NC group (Figure 3E). These findings indicate that DPY30 promotes EMT of CSCC cells.
DPY30-induced EMT may be mediated by the Wnt/β-catenin pathway
Wnt/β-catenin signaling is a classical Wnt pathway, with β-catenin as the core protein. Nuclear β-catenin binds to the nuclear transcription factor Tcf/Lef, thereby abolishing Tcf/Lef inhibition and subsequently activating a series of target genes, such as c-myc and cyclinD1 that are involved in cell proliferation, as well as genes associated with tumorigenesis, leading to excessive cell proliferation.14 To further elucidate the mechanism by which DPY30 contributes to the regulation of CSCC cell growth and metastasis, we analyzed the effect of DPY30 knock-down on activation of the Wnt/β-catenin pathway. Western blot analysis indicated that DPY30 knock-down significantly decreased β-catenin expression in SiHa cells, with a similar decrease in c-myc expression (Figure 3E). This suggests that DPY30 promotes tumorigenesis in CSCC by activating the Wnt/β-catenin signaling pathway.
Discussion
Cervical cancer is an important health concern, as it is associated with significant mortality and morbidity among women.15 EMT is involved in the development of various tumors, including cervical cancer.9,16,17 Our findings presented here indicate that DPY30 plays a role in the progression of CSCC. Our results also suggest that DPY30 induces EMT in CSCC by activating Wnt/β-catenin signaling.
DPY30 is a common but fairly uncharacterized member of the Set1/MLL complex. Members of the SET1/MLL family of proteins are important transcriptional regulators and are crucial for both normal development and disease pathogenesis.18 The role of the SET1/MLL family in cancer biology is complex. Members of the Set1 complex include the integral cores of Wdr5, Rbbp5, Ash2l, or Dpy30; the core subunits are necessary for full methylation activity19,20 and include oncoproteins as well as tumor suppressors. Wdr5 acts as an oncoprotein in prostate cancer, contributing to androgen-induced proliferation of tumor cells.21 Another protein, Ash2l, interacts with the oncoprotein MYC,22 resulting in MYC/HaRAS-dependent transformation of fibroblasts in the rat embryo. Ash21 overexpression has been implicated in the cell proliferation of several tumors.23 Based on the evidence presented here, we propose that DPY30 plays a similar oncogenic role in CSCC.
DPY30 has been recently reported to be involved in the regulation of cell proliferation and cell-cycle progression.4,6,24 In this study, we provide evidence for the first time that DPY30 expression markedly increases in cervical cancer tissues. In addition, our in vitro experiments confirmed that DPY30 promotes the proliferation, migration, and invasion of CSCC cells. Furthermore, the increased expression of DPY30 correlated with many clinicopathological characteristics, such as depth of stromal invasion and differentiation grade. Therefore, DPY30 may be involved in the development of cervical cancer, which makes it a promising new target for treating cervical cancer.
EMT is responsible for cell reprogramming at a molecular level, whereby cancer cells acquire many new properties; these properties in turn modify their interaction with the tumor microenvironment.25 In this study, we found that expression levels of DPY30 were associated with those of some EMT markers, such as E-cadherin, suggesting that DPY30 participates in EMT. Over-activation of Wnt signaling has been implicated in EMT and the subsequent occurrence of cervical cancer.14 Western blot analysis showed that DPY30 down-regulation significantly reduced β-catenin expression, as well as c-myc expression, a downstream target of β-catenin. This suggests that DPY30 contributes to CSCC pathogenesis by activating the Wnt/β-catenin signaling pathway.
Although we present new insights regarding DPY30 regulation of CSCC, our study does have some limitations. This study was based on a retrospective study to predict the value of DPY30, and thus lacks statistical power. Moreover, further analyses, such as the luciferase reporter gene assay, are necessary to investigate the specific nature of the interaction between DPY30 and the Wnt/β-catenin pathway in CSCC cells.
Conclusion
To summarize, DPY30 expression was significantly elevated in CSCC tissues, and enhance proliferation, migration, and invasion of CSCC cells. Furthermore, DPY30 may promote EMT in CSCC and modulate by the Wnt/β-catenin signaling pathway. Therefore, DPY30 may be a useful biomarker to determine CSCC progression and has potential application as a therapeutic target in cervical cancer.
Ethical approval and informed consent
This study was approved by the ethics committee of Liaocheng People’s Hospital Affiliated to Taishan Medical University. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Written informed consent was obtained from individual participants.
Consent for publication
All data published here are under the consent for publication. Written informed consent was obtained from all individual participants included in the study.
Data availability
The datasets generated and analyzed during the present study are available from the corresponding author on reasonable request.
Abbreviations
CSCC, Cervical squamous cell carcinoma; EMT, Epithelial–mesenchymal transition; PCR, Polymerase chain reaction; FIGO, Federation international of gynecology and obstetrics; GAPDH, Glyceraldehyde-3-phosphate dehydrogenase; PMSF, Phenylmethanesulfonyl fluoride; RIPA, Radio-immunoprecipitation assay; SDS-PAGE, Sodium salt-polyacrylamide gel electrophoresis.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21294
2. Forman D, de Martel C, Lacey CJ, et al. Global burden of human papillomavirus and related diseases. Vaccine. 2012;30(Suppl 5):F12–23. doi:10.1016/j.vaccine.2012.07.055
3. Jiang H, Shukla A, Wang X, Chen WY, Bernstein BE, Roeder RG. Role for Dpy-30 in ES cell-fate specification by regulation of H3K4 methylation within bivalent domains. Cell. 2011;144(4):513–525. doi:10.1016/j.cell.2011.01.020
4. Yang Z, Augustin J, Chang C, et al. The DPY30 subunit in SET1/MLL complexes regulates the proliferation and differentiation of hematopoietic progenitor cells. Blood. 2014;124(13):2025–2033. doi:10.1182/blood-2014-01-549220
5. Wang X, Lou Z, Dong X, et al. Crystal structure of the C-terminal domain of human DPY-30-like protein: a component of the histone methyltransferase complex. J Mol Biol. 2009;390(3):530–537. doi:10.1016/j.jmb.2009.05.061
6. Lee YJ, Han ME, Baek SJ, Kim SY, Oh SO. Roles of DPY30 in the proliferation and motility of gastric cancer cells. PLoS One. 2015;10(7):e0131863. doi:10.1371/journal.pone.0131863
7. Simboeck E, Gutierrez A, Cozzuto L, et al. DPY30 regulates pathways in cellular senescence through ID protein expression. Embo J. 2013;32(16):2217–2230. doi:10.1038/emboj.2013.159
8. Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. doi:10.1038/nrm3758
9. Qureshi R, Arora H, Rizvi MA. EMT in cervical cancer: its role in tumour progression and response to therapy. Cancer Lett. 2015;356(2 Pt B):321–331. doi:10.1016/j.canlet.2014.09.021
10. Wang Z, Dao R, Bao L, et al. Epigenetic reprogramming of human lung cancer cells with the extract of bovine parthenogenetic oocytes. J Cell Mol Med. 2014;18(9):1807–1815. doi:10.1111/jcmm.12165
11. Yang LH, Xu HT, Han Y, et al. Axin downregulates TCF-4 transcription via beta-catenin, but not p53, and inhibits the proliferation and invasion of lung cancer cells. Mol Cancer. 2010;9:25. doi:10.1186/1476-4598-9-254
12. Ye X, Weinberg RA. Epithelial-mesenchymal plasticity: a central regulator of cancer progression. Trends Cell Biol. 2015;25(11):675–686. doi:10.1016/j.tcb.2015.07.012
13. Li W, Kang Y. Probing the fifty shades of EMT in metastasis. Trends Cancer. 2016;2(2):65–67. doi:10.1016/j.trecan.2016.01.001
14. Bello JO, Nieva LO, Paredes AC, Gonzalez AM, Zavaleta LR, Lizano M. Regulation of the Wnt/beta-catenin signaling pathway by human papillomavirus E6 and E7 oncoproteins. Viruses. 2015;7(8):4734–4755. doi:10.3390/v7082842
15. Mabelele MM, Materu J, Ng’ida FD, Mahande MJ. Knowledge towards cervical cancer prevention and screening practices among women who attended reproductive and child health clinic at Magu district hospital, Lake Zone Tanzania: a cross-sectional study. BMC Cancer. 2018;18(1):565. doi:10.1186/s12885-018-4242-8
16. Huang D, Xu W, Xu X, Zhang X, Zhou R, Chen P. EMT influences the expression of CK19 in pleural effusion-derived lung cancer cells and their invasion and metastasis. Oncol Lett. 2016;12(6):5052–5058. doi:10.3892/ol.2016.5315
17. Xu J, Liu D, Niu H, et al. Resveratrol reverses doxorubicin resistance by inhibiting epithelial-mesenchymal transition (EMT) through modulating PTEN/Akt signaling pathway in gastric cancer. J Exp Clin Cancer Res. 2017;36(1):19. doi:10.1186/s13046-016-0487-8
18. Vedadi M, Blazer L, Eram MS, Barsyte-Lovejoy D, Arrowsmith CH, Hajian T. Targeting human SET1/MLL family of proteins. Protein Sci. 2017;26(4):662–676. doi:10.1002/pro.3129
19. Dou Y, Milne TA, Ruthenburg AJ, et al. Regulation of MLL1 H3K4 methyltransferase activity by its core components. Nat Struct Mol Biol. 2006;13(8):713–719. doi:10.1038/nsmb1128
20. Shilatifard A. The COMPASS family of histone H3K4 methylases: mechanisms of regulation in development and disease pathogenesis. Annu Rev Biochem. 2012;81:65–95. doi:10.1146/annurev-biochem-051710-134100
21. Kim JY, Banerjee T, Vinckevicius A, et al. A role for WDR5 in integrating threonine 11 phosphorylation to lysine 4 methylation on histone H3 during androgen signaling and in prostate cancer. Mol Cell. 2014;54(4):613–625. doi:10.1016/j.molcel.2014.03.043
22. Yu M, Schreek S, Cerni C, et al. PARP-10, a novel Myc-interacting protein with poly(ADP-ribose) polymerase activity, inhibits transformation. Oncogene. 2005;24(12):1982–1993. doi:10.1038/sj.onc.1208410
23. Luscher-Firzlaff J, Gawlista I, Vervoorts J, et al. The human trithorax protein hASH2 functions as an oncoprotein. Cancer Res. 2008;68(3):749–758. doi:10.1158/0008-5472.CAN-07-3158
24. Yang Z, Shah K, Khodadadi-Jamayran A, Jiang H. Dpy30 is critical for maintaining the identity and function of adult hematopoietic stem cells. J Exp Med. 2016;213(11):2349–2364.
25. Chockley PJ, Keshamouni VG. Immunological consequences of epithelial-mesenchymal transition in tumor progression. J Immunol. 2016;197(3):691–698. doi:10.4049/jimmunol.1600458
Supplementary materials
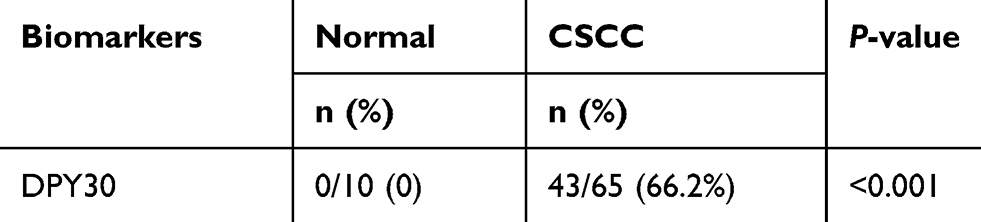 |
Table S1 Statistical analysis of immunohistochemistry staining of DPY30 in normal and CSCC tissues |
 |
Table S2 Statistical analysis of immunohistochemistry staining of E-cadherin in normal and CSCC tissues |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.