Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 15
Down-Regulation of miR-138 Alleviates Inflammatory Response and Promotes Wound Healing in Diabetic Foot Ulcer Rats via Activating PI3K/AKT Pathway and hTERT
Authors Wang J, Zhao X, Tian G, Liu X, Gui C, Xu L
Received 7 February 2022
Accepted for publication 1 April 2022
Published 14 April 2022 Volume 2022:15 Pages 1153—1163
DOI https://doi.org/10.2147/DMSO.S359759
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ming-Hui Zou
Jian Wang,1,2 Xiaodan Zhao,3 Guichang Tian,2 Xiaochao Liu,2 Chengyan Gui,2 Lin Xu1,4
1School of Medicine, Cheeloo College of Medicine, Shandong University, Jinan, 250012, People’s Republic of China; 2Department of Orthopedics, Qufu Hospital of TCM, Qufu, 273100, People’s Republic of China; 3Image Center, Shandong Provincial Third Hospital, Jinan, 250000, People’s Republic of China; 4Department of Orthopedics, Yantai Affiliated Hospital of Binzhou Medical University, Yantai, 264100, People’s Republic of China
Correspondence: Lin Xu, Department of Orthopedics, Yantai Affiliated Hospital of Binzhou Medical University, Yantai, 264100, People’s Republic of China, Tel +86-13805350031, Email [email protected]; [email protected]
Objective: To study the role of miR-138 on the repair of diabetic foot ulcer (DFU) and further to explore its possible mechanism.
Materials and Methods: miR-138 inhibitor, IGF-1, LY294002 were used in DFU rat mode, and the mRNA expression of miR-138 was detected. HE staining was used to observe the histological changes of skin ulcer in rats. The level of inflammation, wound healing, and blood vessel formation-related factors were detected by ELISA and immunohistochemical. The expression of VEGF and PI3K/AKT pathway-related proteins were detected by Western blot. To further determine the underlying mechanism of miR-138 in the repair of DFU, telomerase inhibitor BIBR-1232 was used in HUVECs. Dual-luciferase assay was used to determine the target relationship between miR-138 and hTERT. CCK-8, transwell, and tube formation assays were conducted to observe the biological behavior of HUVECs. Inflammatory cytokines and PI3K/AKT pathway-related proteins were also measured by ELISA and Western blot.
Results: The mRNA expression of miR-138 in DFU rat was increased and ulcer of diabetic foot rats was improved after silencing miR-138. The results of cellular bioactivity in vitro experiment were consistent with that in vivo. Meanwhile, after silencing miR-138, the level of inflammatory cytokines was decreased, while the level of anti-inflammatory and healing factors was increased in vivo and vitro. Moreover, the ratios of p-PI3K/PI3K and p-AKT/AKT were upregulated after treated with miR-138 inhibitor and miR-138 was negatively regulated the expression of hTERT. However, the inhibitory effect on inflammatory response and the promotion effect on wound healing of miR-138 inhibitor were reversed by LY294002 and BIBR-1232.
Conclusion: Down-regulation of miR-138 could alleviate inflammatory response and promote wound healing in DFU rats by activating PI3K/AKT pathway and hTERT.
Keywords: miR-138, diabetic foot ulcer, inflammatory response, wound healing, PI3K/AKT pathway, hTERT
Introduction
Diabetes is a widespread metabolic problem in modern society. Due to long-term hyperglycemia, diabetes can cause damage to multiple systems, such as the eyes, kidneys, nerves, blood vessels, etc.1 As one of the serious complications of diabetes, diabetic foot is a chronic progressive disease caused by vascular occlusive disease, which can lead to complications.2 Once foot ulcers occur, the extremities of diabetic patients are very likely to be infected with invasive infection, leading to an increased risk of amputation.3 With the annual increase in the number of diabetic patients in the world, the incidence of diabetic foot ulcer (DFU) is also increasing, which has a great impact on family, society, and economic development. There is an urgent need to develop more effective detection and treatment methods for DFU.
Persistent infection and chronic inflammation are the most common features and complex mechanisms of DFU based on the profound impact of chronic inflammation on immune response aggravation and treatment disorders.4,5 An appropriate inflammatory response is essential to initiate the healing process, but dysregulated and prolonged inflammation may lead to tissue damage, inhibition of epithelialization, and unresolved infection.6 Meanwhile, the combination of altered protein and lipid metabolism in diabetes impeded the process of wound healing and simultaneously reducing the tensile strength of the wound.7 Therefore, inhibiting inflammation and promoting wound healing are essential to improve DFU.
New analytical techniques, especially those related to molecular markers, have contributed to the development of new and effective therapies. MicroRNAs (miRNAs) are endogenous single-stranded non-coding RNA molecules with a length of about 22nt, which are transcribed by RNA polymerase II.8 A series of previous studies have shown that miRNAs play an important role in diabetic wound healing and inflammation.9–11 MiR-138 was first discovered in undifferentiated embryonic stem cells, and it shows a highly conserved state in different organisms. Study has shown that miR-138 negatively regulates hexokinase 1, a key protease of glucose metabolism in HepG2 cells, and might be a potential and promising target for the treatment of type 2 diabetes.12 Another study found that down-regulation of miR-138 has a protective effect on inflammatory injury of chondrocytes.13 Thus, in this study, we hypothesized that miR-138 is involved in the progression of DFU. The roles of miR-138 in inflammatory response and wound healing were investigated through establishment a DFU rat model and further to explore its possible mechanism.
Materials and Methods
Experimental Animal
Male Sprague Dawley (SD) rats weighing 250 ± 30g and aged 8–10 weeks were purchased from Jinan Pengyue Laboratory Animal Co Ltd., license number SYXK (Lu) 20180030. These rats were kept in a clean environment with 55 ± 5% humidity and 23 ± 2°C, and free to drinking water and food. All animal experiments were carried out in strict accordance with the ethical guidelines by the International Council for Laboratory Animal Science (ICLAS) and have been reviewed and approved by the Animal Care and Use Committee of Shandong University.
Establishment of Diabetic Foot Ulcer Model and Groups
Diabetic rats were induced by single intraperitoneal injection of 60 mg/kg streptozotocin (STZ, Sigma-Aldrich Co., Taufkirchen, Germany) dissolved in citrate buffer solution (0.1 M, pH 4.5). Blood glucose level was measured 72 h after injection, and those with fasting blood glucose higher than 16.7 mM were defined as having diabetes. The rats were anesthetized with isoflurane inhalation, and a 4 × 4 mm wound depth reached fascia was made on the hind foot of the diabetic rats.
Animals were divided into 6 groups (n = 8): i) control group, rats were received the same volume of vehicle, and a 4 × 4 mm wound depth reached fascia was made; ii) DFU model group (DFU), rats were established the DFU model; iii) DFU + miR-138 inhibitor negative control group (NC), after established DFU model, rats were injected with miR-138 inhibitor scramble (40ul) by tail vein; iv) DFU + miR-138 inhibitor group (miR-138 inhibitor), after established DFU model, rats were injected with miR-138 inhibitor (40ul)14 by tail vein; v) IGF-I group (after established DFU model, rats were injected with PI3K/AKT pathway activator IGF-I by tail vein); and vi) inhibitor + LY group (after established DFU model, rats were injected with miR-138 inhibitor and PI3K/AKT pathway inhibitor LY294002 by tail vein). At 4 weeks post injection, the rats were anesthetized with 3% sodium pentobarbital (50 mg/kg) and tissue of foot ulcer was collected.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
After extracted by TRIZOL reagent (OmegaBio-tek, Georgia, USA), total RNA was transcribed into cDNA (Baosheng Scientific, Dalian, China). Mastercycler® nexus X2 (Eppendorf, Hamburg, Germany) was used for PCR and sequence of the primers are as follows: miR-138, forward: 5′-GTATTGACTAGATTAATCACTGT-3′ and reverse: 5′-CTCGCTTCGGCAGCACA-3’. U6, forward: 5′- GACCTCTATGCCAACACAGT −3’ and reverse: 5′-AGTACTTGCGCTCAGGAGGA-3’. The cycling conditions were 95°C for 30s, and then 35 cycles of 60°C for 45s and 72°C for 3 min. U6 is the internal reference, and relative expression level of miR-138 was calculated by the 2−ΔΔCt method.
Histopathological Evaluation
The specimens were immersed in 4% paraformaldehyde for fixation, embedded into paraffin and serially cut into 4µm-thick sections. After stained with hematoxylin and eosin (H&E), the specimens were examined under a light microscope (Olympus BX51, Olympus, Japan).
Enzyme-Linked Immunosorbent Assay (ELISA)
After homogenized and centrifuged, the supernatant of foot ulcer tissue was collected and quantitatively assayed for the levels of interleukin (IL)-1β, tumor necrosis factor (TNF)-a, IL-10, and IL-6 were measured according to the instructions of detection kits (Jiancheng Bioengineering Institute, Nanjing, China).
Immunohistochemistry
The foot ulcer tissue was embedded in paraffin, sliced, and rehydrated through gradient ethanol. After blocked with normal goat serum for 20 min, the foot ulcer tissue was incubated with rabbit anti-mouse collagen I (Biorbyt, Cambridge, UK), collagen III (Biorbyt, Cambridge, UK), α-smooth muscle actin (α-SMA, Biorbyt, Cambridge, UK), matrix metalloproteinase-1 (MMP-1, Biorbyt, Cambridge, UK), and CD34 (Abcam, USA) overnight at 4°C, respectively. Then, the foot ulcer tissue was incubated with biotinylated goat anti-rabbit IgG antibodies (Antibodies-online, Germany). Subsequently, the foot ulcer tissue was incubated with streptavidin-biotin complex and stained with diaminobenzidine. After dehydrated in gradient ethanol solution and transparently treated with xylene, the protein expression of MMP-1, collagen I, collagen III, α-SMA, and CD34 in foot ulcer tissue was viewed under 400x magnification.
Western Blot
The total protein concentration of foot ulcer tissue was extracted and measured by bicinchoninic acid (BCA) protein assay kit (Thermo Fisher Scientific, Waltham, USA). After subjected and transferred, the protein membranes were blocked with 5% not-fat milk and incubated with VEGF, PI3K, p-PI3K, Akt, p-Akt, and β-actin primary antibodies (Biorbyt, Cambridge, UK) overnight at 4°C. After incubated with IgG-HRP (Cell Signaling Technology, USA) for 4 h, the blots were visualized with enhanced chemiluminescence (ECL) system and quantified by Image J (NIH) software.
Cell Culture and Grouping
Human umbilical vein endothelial cells (HUVECs) (ATCC, Manassas, VA, USA) were cultured in 1640 medium containing 10% fetal bovine serum, 1% penicillin, and 1% streptomycin at 37°C, 5% CO2. To mediate transfection, 4 µL LipofectamineTM 2000 was gently mixed with 200 µL Opti-MEM transfection medium and incubated for 5 min at room temperature. Then, 5 µL each of miR-138 inhibitor and scramble were mixed with 200 µL Opti-MEM transfection medium, and then incubated at room temperature for 5 min. HUVECs in logarithmic phase were divided into 7 group: i) control group, HUVECs were cultured with 20 mM mannitol; ii) high glucose group (HG), HUVECs were cultured with 25 mM glucose; iii) miR-138 inhibitor negative control group (NC), high glucose-induced HUVECs were transfected with miR-138 inhibitor scramble using the LipofectamineTM 2000 transfection reagent; iv) miR-138 inhibitor group, high glucose-induced HUVECs were transfected with miR-138 inhibitor; v) IGF-I group, high glucose-induced HUVECs were cultured with IGF-I; vi) inhibitor + LY group, high glucose-induced HUVECs were transfected with miR-138 inhibitor and cultured with LY294002; and vii) inhibitor + BIBR group, high glucose-induced HUVECs were transfected with miR-138 inhibitor and cultured with BIBR-1232. The qPT-PCR, ELISA and Western blot experiments were repeated.
CCK-8 Assay
HUVECs were planted into 96-well plates, and CCK-8 solution (PA137267, Pierce) was added at corresponding time points. After cultured for another 4h, the absorbance value (OD) was measured at 450 nm by a THERMOmax microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Transwell Assay
Matrigel was diluted with prechilled RPMI 1640 medium, 100 μL cell suspension (2 × 105 cells/mL) digested and resuspended in RPMI-1640 medium was added into the upper compartment and 600 ul 10% FBS-RPMI culture media was added to the lower chamber. After incubation at 37°C for 24 h, the underlying transmembrane cells were fixed with 5% glutaraldehyde and stained with 0.5% crystal violet for 30 min. The invasion was photographed under the microscope (Revolve FL, Echo Laboratories, USA). Except that the bottom of the Transwell chamber was not coated with matrigel, the procedure of the migration was the same as that of the invasion.
Tube Formation Assay
HUVECs were inoculated in 96-well plates and cured with 50 μL Matrigel (BD Bioscience, USA). After digested and centrifuged, cells were seeded on Matrigel and incubated at 37 °C for 6 h. The tube formation was observed under an Olympus inverted microscope (CKX40, Olympus, New York, NY).
Luciferase Reporter Assay
TargetScan, miRanda and miRDB databases were used to predict the target genes of miR-138. The wild-type and mutant 3ʹUTRs of hTERT and miR-138 were obtained by pGL3/Luciferase vector and cloned into the downstream of luciferase gene. Twenty-four hours after transfection, the luciferase activity was detected by dual luciferase reporting system (Promega, Madison, WI, USA).
Statistical Analysis
The data were presented as means and standard deviations (mean ± SD) and the results were analyzed by SPSS 21.0 software. Multiple comparisons were tested by one-way ANOVA and subsequent analysis using LSD test. Differences were considered statistically significant at p < 0.05.
Results
Effect of miR-138 on the Histological Changes of Skin Ulcer Model in Rats
Figure 1A shows the expression of miR-138 mRNA in DFU group was markedly increased compared with that in Control group (p < 0.01), while down-regulation of miR-138 could markedly decrease the expression of miR-138 mRNA compared with that in DFU group (p < 0.01). Moreover, the expression of miR-138 mRNA in the DFU group and NC group were no significant difference (p > 0.05). The result indicated that miR-138 mRNA was up-regulated in DFU rats and miR-138 inhibitor administration was successful.
 |
Figure 1 Effect of miR-138 on the histological changes of skin ulcer model in rats. (A) The expression of miR-138 mRNA in different groups. (B) The morphological changes of foot ulceration in different groups (×200). **p < 0.01 against Control group; ##p < 0.01 against DFU group; ^^p < 0.01 against NC group; &&p < 0.01 against miR-138 inhibitor group; $$p < 0.01 against IGF-1 group. |
Compared with rats in DFU group and NC group, rats treated with miR-138 inhibitor or IGF-1 showed significant increase of newly developed or aged granulation tissue containing newly developed blood vessels were observed. However, LY294002 could attenuate the positive effects of miR-138 inhibitor in DFU rats. Those results indicated that down-regulation of miR-138 could improve the ulcer of diabetic foot rats by activating PI3K/AKT pathway (Figure 1B).
Effect of miR-138 on the Content of Inflammation and Wound Healing Related Factors in DFU Rats
The results of ELISA (Figure 2A) and immunohistochemistry (Figure 2B) shown that compared with Control group, the level of IL-1β, TNF-α, and IL-6 was dramatically increased in DFU group, while the level of IL-10, MMP-1, collagen I, collagen III, and α-SMA was dramatically decreased (p < 0.01). Moreover, after silencing miR-138 gene or treatment with IGF-1, the levels of pro-inflammatory cytokines were significantly reduced and the levels of IL-10, MMP-1, collagen type I, collagen type III, α-SMA were significantly increased in DFU rats. However, compared with miR-138 inhibitor group, the anti-inflammatory ability and wound healing ability of inhibitor + LY group were significantly receded (p < 0.05). The results elucidated that down-regulation of miR-138 could alleviate inflammatory response and promote wound healing by activating PI3K/AKT pathway in DFU rats.
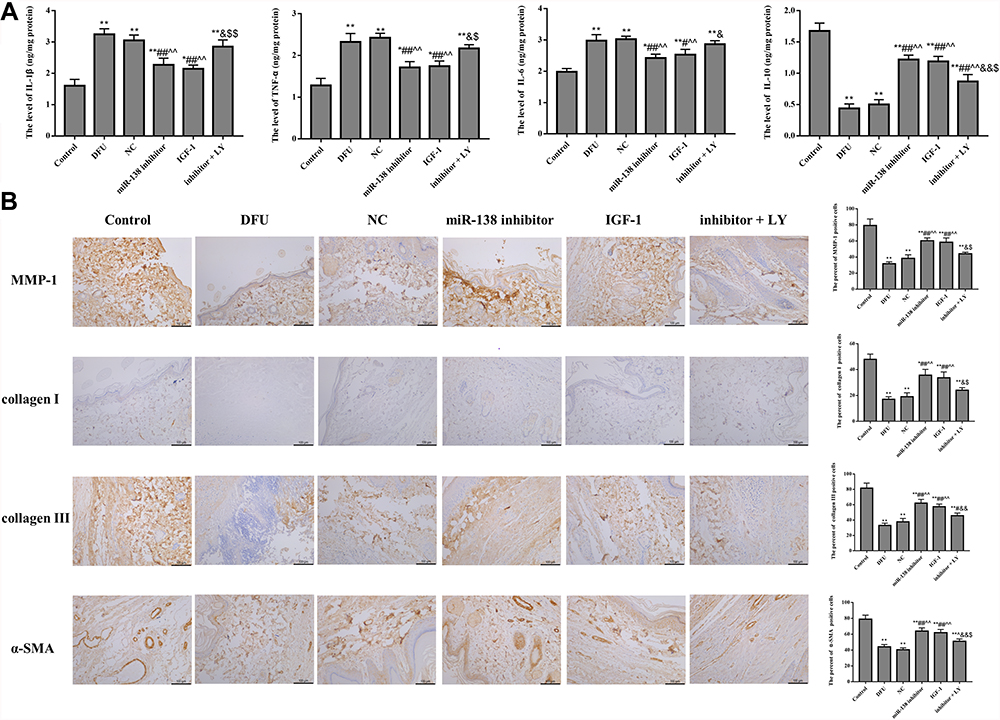 |
Figure 2 Down-regulation of miR-138 alleviates inflammatory response (A) and wound healing ((B), ×200) in DFU rats. *p < 0.05, **p < 0.01 against Control group; #p < 0.05, ##p < 0.01 against DFU group; ^^p < 0.01 against NC group; &p < 0.05, &&p < 0.01 against miR-138 inhibitor group; $p < 0.05, $$p < 0.01 against IGF-1 group. |
Effect of miR-138 on the Expression of PI3K/AKT Pathway and Blood Vessel Formation Related Factors in DFU Rats
The status of neovascularization was evaluated by immunohistochemical staining for CD34 (Figure 3A). The results demonstrated that the arrangement of blood vessels in DFU group was disordered, and the density of blood vessels was significantly reduced. Meanwhile, the number of new blood vessels increased significantly in the miR-138 inhibitor group and IGF-1 group. However, the positive effects of miR-138 inhibitor in DFU rats were destroyed by LY294002.
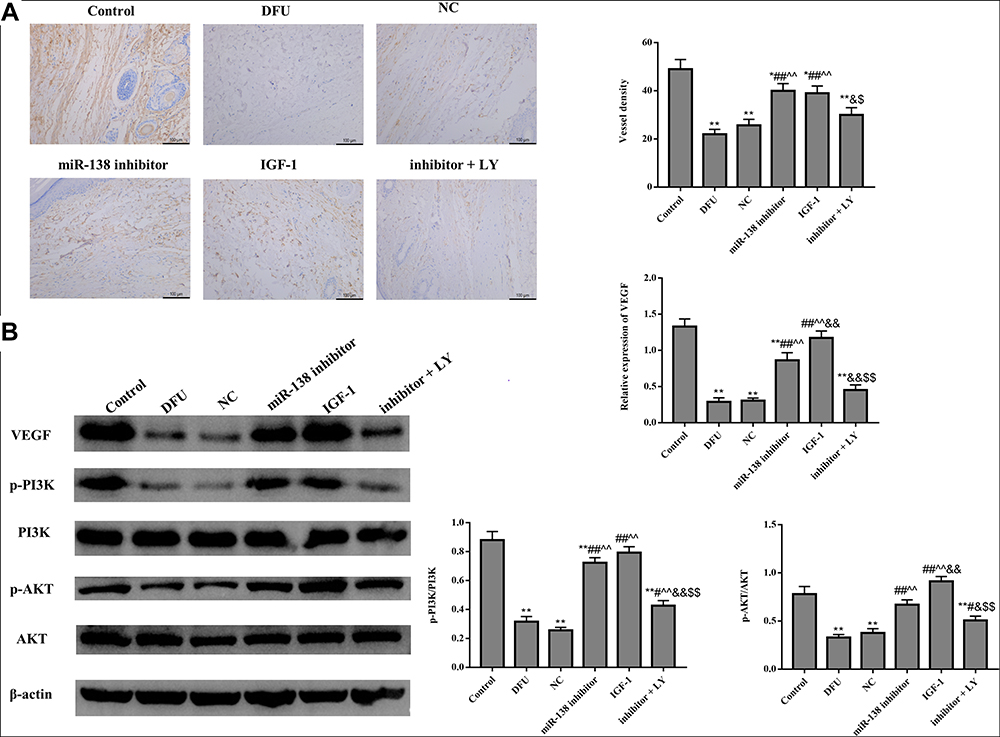 |
Figure 3 Down-regulation of miR-138 improves blood vessel formation in DFU rats. (A) CD34 immunohistochemical staining was used to evaluate vessel density (×200). (B) Western blot was used to detect the expression of blood vessel formation and PI3K/AKT pathway related proteins. *p < 0.05, **p < 0.01 against Control group; #p < 0.05, ##p < 0.01 against DFU group; ^^p < 0.01 against NC group; &p < 0.05, &&p < 0.01 against miR-138 inhibitor group; $p < 0.05, $$p < 0.01 against IGF-1 group. |
Figure 3B shows that compared with Control group, the protein expression of VEGF, p-PI3K, and p-AKT were dramatically down-regulated in DFU group (p < 0.01). Moreover, the expression of those proteins in the miR-138 inhibitor group and IGF-1 group were dramatically increased compared with that in DFU group and NC group (p < 0.05). However, LY294002 treatment could significantly down-regulate the expression of VEGF, p-PI3K, and p-AKT in DFU rats. Taken together, those results indicated that down-regulation of miR-138 could improve blood vessel formation by activating PI3K/AKT pathway in DFU rats.
Effect of miR-138 on hTERT
Software analysis revealed that hTERT was a target of miR-138. In the dual-luciferase report, the luciferase activity of hTERT in the WT 3ʹUTR was reduced, but not in the Mut 3ʹUTR (p < 0.05), suggesting that hTERT is a target gene of miR-138 in cells (Figure 4A).
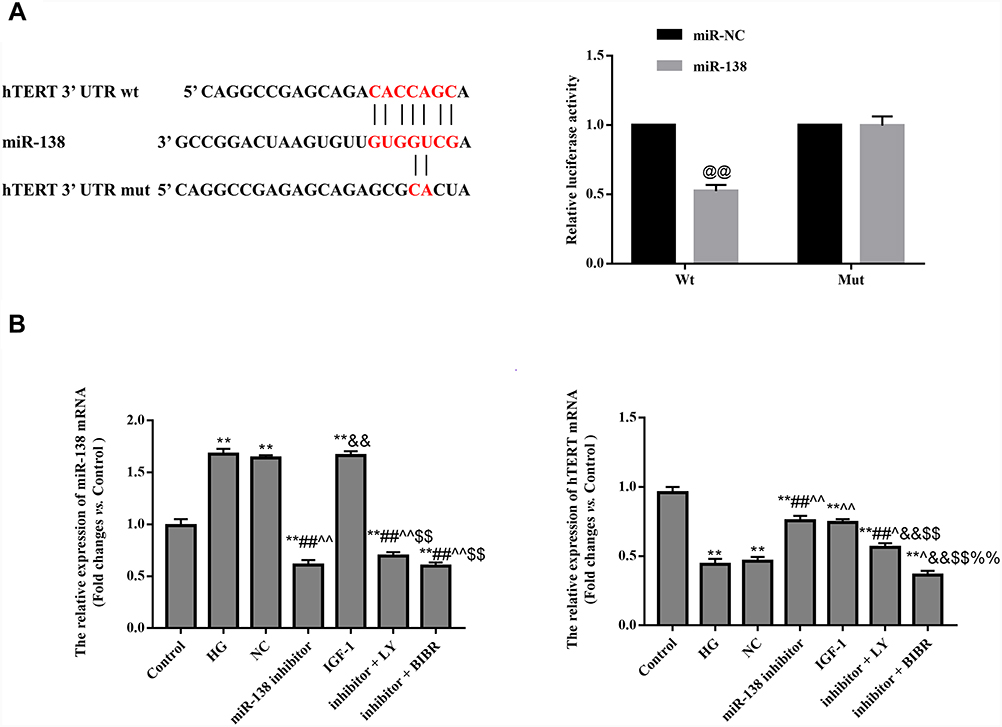 |
Figure 4 Effect of miR-138 on hTERT. (A) hTERT is a target gene of miR-138. (B) qRT-PCR was used to detect the mRNA expression of miR-138 and hTERT. @@p < 0.01 against miR-NC group. **p < 0.01 against Control group; ##p < 0.01 against HG group; ^p < 0.05, ^^p < 0.01 against NC group; &&p < 0.01 against miR-138 inhibitor group; $$p < 0.01 against IGF-1 group; %%p < 0.01 against inhibitor + LY group. |
QRT-PCR found that hTERT expression was significantly increased in the miR-138 inhibitor group compared with that in NC group (p < 0.01), suggesting that miR-138 negatively regulates hTERT expression (Figure 4B).
Effect of miR-138 on the Cellular Bioactivity of HUVECs
CCK-8 assay (Figure 5A), tube formation assay (Figure 5B), and transwell assay (Figure 5C) were conducted to observe the biological behavior of HUVECs. The administration of miR-138 inhibitor significantly increased the proliferation, invasion, migration, and tube formation of HUVECs. However, compared with miR-138 inhibitor group, the cellular bioactivity of HUVECs in inhibitor + LY group and inhibitor + BIBR group were significantly decreased (p < 0.05). The results indicated that down-regulation of miR-138 could promote cellular bioactivity by activating PI3K/AKT pathway and hTERT in HUVECs.
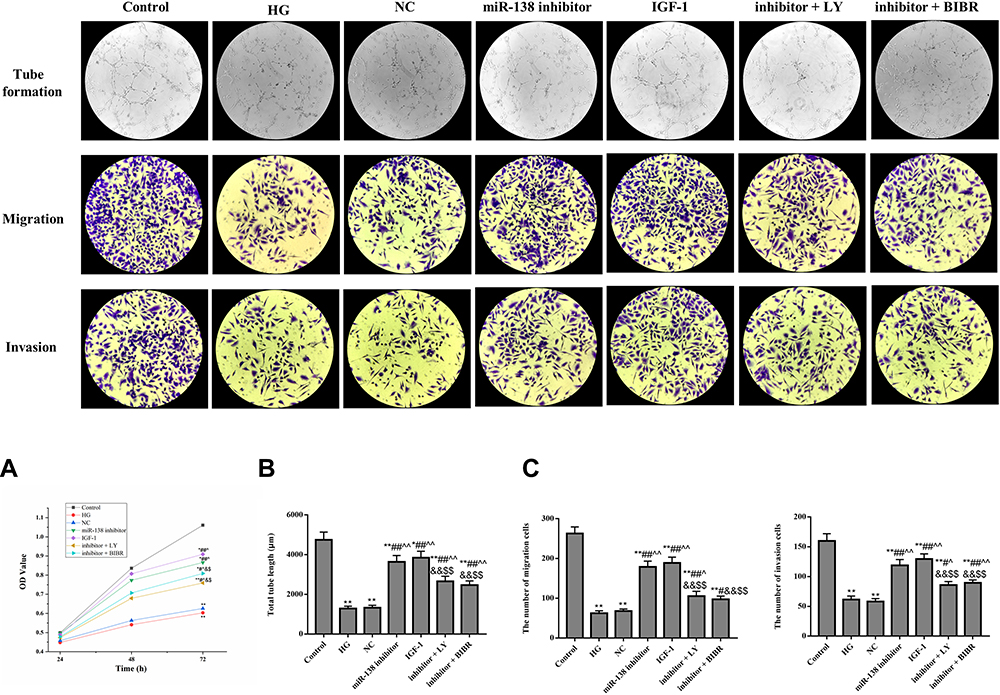 |
Figure 5 Effect of miR-138 on the cellular bioactivity of HUVECs. (A) CCK-8 assay. (B) Tube formation assay (×200). (C) Transwell assay (×200). *p < 0.05, **p < 0.01 against Control group; #p < 0.05, ##p < 0.01 against HG group; ^p < 0.05, ^^p < 0.01 against NC group; &p < 0.05, &&p < 0.01 against miR-138 inhibitor group; $p < 0.05, $$p < 0.01 against IGF-1 group. |
Effect of miR-138 on the Content of Inflammation and PI3K/AKT Pathway Related Factors in HUVECs
Figure 6 shows that compared with HG group and NC group, the level of IL-1β, TNF-α, and IL-6 in miR-138 inhibitor was dramatically decreased, and the level of IL-10, p-PI3K, and p-AKT was dramatically increased (p < 0.01). However, the anti-inflammatory effects of miR-138 inhibitor in HUVECs were reversed by LY294002 or BIBR-1232. This indicated that down-regulation of miR-138 could alleviate inflammatory response by activating PI3K/AKT pathway and hTERT in HUVECs.
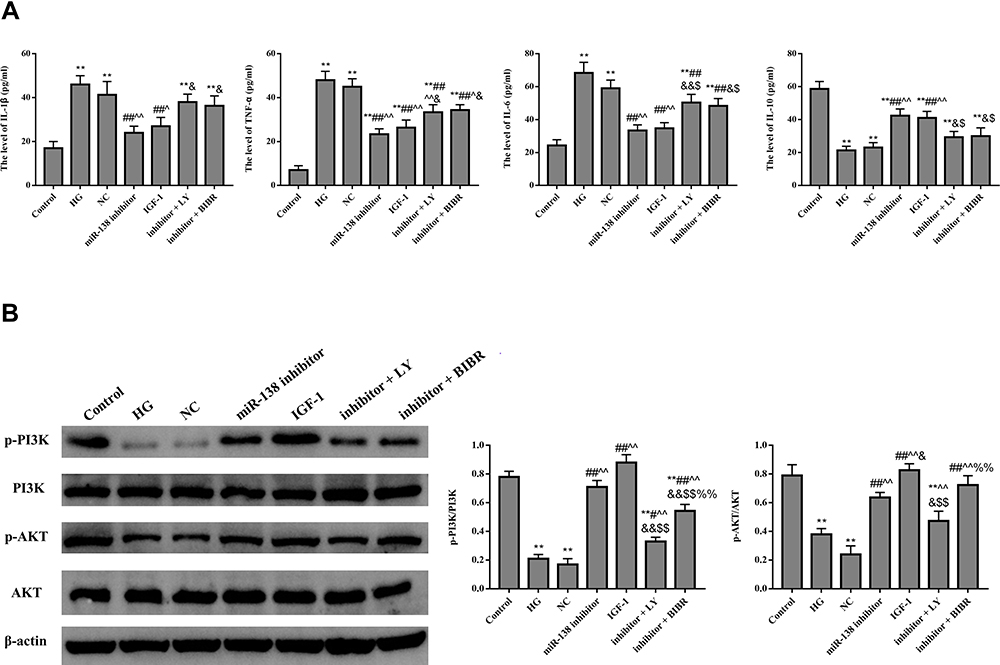 |
Figure 6 Effect of miR-138 on the content of inflammation (A) and PI3K/AKT pathway (B) related factors in HUVECs. **p < 0.01 against Control group; #p < 0.05, ##p < 0.01 against HG group; ^p < 0.05, ^^p < 0.01 against NC group; &p < 0.05, &&p < 0.01 against miR-138 inhibitor group; $p < 0.05, $$p < 0.01 against IGF-1 group, %%p < 0.01 against inhibitor + LY group. |
Discussion
Epidemiological studies have confirmed that DFU is associated with significant morbidity and mortality.15,16 DFU has become a major and growing health issue with the increasing number of diabetic patients. As key regulatory factors of cell physiology, the disorder of miRNAs can lead to many pathological states.17 Increasing evidence suggests that regulation of miRNA activity may influence the course of disease.18 Differentially expressed miRNAs have been found to be associated with type 2 diabetes and its complications.12,19 Relatedly, these miRNAs can be used for therapeutic purposes or as biomarkers for the diagnosis of diabetes and its complications. In order to provide a new feasible scheme for the mechanism research and clinical treatment of DFU, DFU rats and high glucose HUVECs model were established and the roles of miR-138 in DFU was explored in this study.
The pathophysiology of DFU is complex, and the wound healing is a dynamic and complex biological process. It is well known that inflammation is an early response to injury and plays an important role in the healing process. However, excessive inflammation can delay the healing of diabetes, which is an important cause of amputation in diabetic patients.20 Numerous studies have investigated wound healing through HUVECs and found that restoring HUVECs function can accelerate wound healing in DFU.21–23 In this study, the content of IL-1β, TNF-α, and IL-6 were significantly decreased in DFU rats and high glucose-induced HUVECs, and the level of IL-10 was significantly increased after silencing miR-138, indicating that down-regulation of miR-138 could alleviate the inflammatory response of DFU. On the other hand, the formation of granulation tissue that promotes wound healing requires increased collagen synthesis, and wound healing is also associated with scar contraction.24 α-SMA expression is the hallmark of mature myofibroblasts, and MMP-1 has a prominent role in initial cleavage of the extracellular matrix.25,26 Furthermore, angiogenesis is an important factor in the proliferation phase of wound healing. As one of the most effective pro-angiogenic growth factors in skin, VEGF expression level in keloid tissue is higher than that in normal skin.27 The observations of those factors in our study suggested that down-regulation of miR-138 could promote the synthesis of collagen and promote scar contraction in DFU.
The regulation of miRNAs in cellular inflammation, stress response, differentiation and other processes cannot be without signaling pathways.28 As one of the classic signaling pathways in cells, PI3K/AKT regulates a wide range of cellular processes, including proliferation, growth, and angiogenesis.29 A study by Jere et al has shown that PI3K/AKT pathway plays an active role in diabetic wound healing.30 Phosphorylation and dephosphorylation play an important role in most signaling pathways, as these processes directly regulate various aspects of protein function.31 Meanwhile, Akt phosphorylation is a key event that induces telomerase activity in cells.32 Moreover, our research found that human telomerase reverse transcriptase (hTERT) is the target gene of miR-138. hTERT, the catalytic and limiting component of telomerase, is essential for the integrity of linear chromosome ends and regulation of the cell cycle in multicellular eukaryotes.33 To further verify the relationship of miR-138, PI3K/AKT and hTERT in the repair of DFU, PI3K/AKT pathway inhibitor and telomerase inhibitor BIBR-1232 were used. The findings showed that down-regulation of miR-138 could improve DFU by activating PI3K/AKT pathway and hTERT.
This study demonstrated that down-regulation of miR-138 contributes to alleviate inflammatory response and promote wound healing in DFU rats. Further studies on high glucose-induced HUVECs revealed that the positive effect of silencing miR-138 in DFU was related to the activation of PI3K/AKT pathway and hTERT. Taken together, miR-138 may serve as a potential therapeutic target and/or a biomarker for the repair of DFU. In further research, we will explore the efficacy and related mechanisms of miR-138 in the treatment of patients with clinical DFU and provide more theoretical basis for the clinical use of miR-138.
Funding
There is no funding to report.
Disclosure
The authors declared no conflicts of interest in this work.
References
1. Karaa A, Goldstein A. The spectrum of clinical presentation, diagnosis, and management of mitochondrial forms of diabetes. Pediatr Diabetes. 2015;16(1):1–9. doi:10.1111/pedi.12223
2. Bandyk DF. The diabetic foot: pathophysiology, evaluation, and treatment. Semin Vasc Surg. 2018;31(2–4):43–48. doi:10.1053/j.semvascsurg.2019.02.001
3. Yang X, Cao Z, Wu P, Li Z. Effect and mechanism of the Bruton tyrosine kinase (Btk) inhibitor ibrutinib on rat model of diabetic foot ulcers. Med Sci Monit. 2019;25:7951–7957. doi:10.12659/MSM.916950
4. Ramirez HA, Pastar I, Jozic I, et al. Staphylococcus aureus triggers induction of miR-15B-5P to diminish DNA repair and deregulate inflammatory response in diabetic foot ulcers. J Invest Dermatol. 2018;138(5):1187–1196. doi:10.1016/j.jid.2017.11.038
5. Davis FM, Kimball A, Boniakowski A, Gallagher K. Dysfunctional wound healing in diabetic foot ulcers: new crossroads. Curr Diab Rep. 2018;18(1):2. doi:10.1007/s11892-018-0970-z
6. Eming SA, Martin P, Tomic-Canic M. Wound repair and regeneration: mechanisms, signaling, and translation. Sci Transl Med. 2014;6(265):265sr6. doi:10.1126/scitranslmed.3009337
7. Sawaya AP, Stone RC, Brooks SR, et al. Deregulated immune cell recruitment orchestrated by FOXM1 impairs human diabetic wound healing. Nat Commun. 2020;11(1):4678. doi:10.1038/s41467-020-18276-0
8. Saliminejad K, Khorram Khorshid HR, Soleymani Fard S, Ghaffari SH. An overview of microRNAs: biology, functions, therapeutics, and analysis methods. J Cell Physiol. 2019;234(5):5451–5465. doi:10.1002/jcp.27486
9. Goodarzi G, Maniati M, Qujeq D. The role of microRNAs in the healing of diabetic ulcers. Int Wound J. 2019;16(3):621–633. doi:10.1111/iwj.13070
10. Ozdemir D, Feinberg MW. MicroRNAs in diabetic wound healing: pathophysiology and therapeutic opportunities. Trends Cardiovasc Med. 2019;29(3):131–137. doi:10.1016/j.tcm.2018.08.002
11. Alexander M, Hu R, Runtsch MC, et al. Exosome-delivered microRNAs modulate the inflammatory response to endotoxin. Nat Commun. 2015;6:7321. doi:10.1038/ncomms8321
12. Luan B, Sun C. MiR-138-5p affects insulin resistance to regulate type 2 diabetes progression through inducing autophagy in HepG2 cells by regulating SIRT1. Nutr Res. 2018;59:90–98. doi:10.1016/j.nutres.2018.05.001
13. Chunlei H, Chang Z, Sheng L, Yanchun Z, Lulin L, Daozhang C. Down-regulation of MiR-138-5p protects chondrocytes ATDC5 and CHON-001 from IL-1 β-induced inflammation via up-regulating SOX9. Curr Pharm Des. 2020;25(43):4613–4621. doi:10.2174/1381612825666190905163046
14. Liu F, Guo J, Qiao Y, et al. MiR-138 plays an important role in diabetic nephropathy through SIRT1-p38-TTP regulatory axis. J Cell Physiol. 2021;236(9):6607–6618. doi:10.1002/jcp.30238
15. Reardon R, Simring D, Kim B, Mortensen J, Williams D, Leslie A. The diabetic foot ulcer. Aust J Gen Pract. 2020;49(5):250–255. doi:10.31128/AJGP-11-19-5161
16. Peter-Riesch B. The diabetic foot: the never-ending challenge. Endocr Dev. 2016;31:108–134.
17. Chen L, Heikkinen L, Wang C, Yang Y, Sun H, Wong G. Trends in the development of miRNA bioinformatics tools. Brief Bioinform. 2019;20(5):1836–1852. doi:10.1093/bib/bby054
18. Tiwari A, Mukherjee B, Dixit M. MicroRNA key to angiogenesis regulation: miRNA biology and therapy. Curr Cancer Drug Targets. 2018;18(3):266–277. doi:10.2174/1568009617666170630142725
19. Jiménez-Lucena R, Camargo A, Alcalá-Diaz JF, et al. A plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: from the CORDIOPREV study. Exp Mol Med. 2018;50(12):1–12. doi:10.1038/s12276-018-0194-y
20. Feng J, Dong C, Long Y, et al. Kallikrein-binding protein in diabetes impairs wound healing through inducing macrophage M1 polarization. Cell Commun Signal. 2019;17(1):60. doi:10.1186/s12964-019-0376-9
21. Huang X, Liang P, Jiang B, et al. Hyperbaric oxygen potentiates diabetic wound healing by promoting fibroblast cell proliferation and endothelial cell angiogenesis. Life Sci. 2020;259:118246. doi:10.1016/j.lfs.2020.118246
22. Wang TY, Wang W, Li FF, et al. Maggot excretions/secretions promote diabetic wound angiogenesis via miR18a/19a-TSP-1 axis. Diabetes Res Clin Pract. 2020;165:108140. doi:10.1016/j.diabres.2020.108140
23. Xu Y, Yu T, He L, et al. Inhibition of miRNA-152-3p enhances diabetic wound repair via upregulation of PTEN. Aging. 2020;12(14):14978–14989. doi:10.18632/aging.103557
24. Albaugh VL, Mukherjee K, Barbul A. Proline precursors and collagen synthesis: biochemical challenges of nutrient supplementation and wound healing. J Nutr. 2017;147(11):2011–2017. doi:10.3945/jn.117.256404
25. Shinde AV, Humeres C, Frangogiannis NG. The role of α-smooth muscle actin in fibroblast-mediated matrix contraction and remodeling. Biochim Biophys Acta Mol Basis Dis. 2017;1863(1):298–309. doi:10.1016/j.bbadis.2016.11.006
26. Arakaki PA, Marques MR, Santos MC. MMP-1 polymorphism and its relationship to pathological processes. J Biosci. 2009;34(2):43–48. doi:10.1007/s12038-009-0035-1
27. Johnson KE, Wilgus TA. Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair. Adv Wound Care. 2014;3(10):647–661. doi:10.1089/wound.2013.0517
28. Di Leva G, Garofalo M, Croce CM. MicroRNAs in cancer. Annu Rev Pathol. 2014;9:287–314. doi:10.1146/annurev-pathol-012513-104715
29. Ersahin T, Tuncbag N, Cetin-Atalay R. The PI3K/AKT/mTOR interactive pathway. Mol Biosyst. 2015;11(7):1946–1954. doi:10.1039/C5MB00101C
30. Jere SW, Houreld NN, Abrahamse H. Role of the PI3K/AKT (mTOR and GSK3β) signalling pathway and photobiomodulation in diabetic wound healing. Cytokine Growth Factor Rev. 2019;S1359-6101:30013–30019.
31. Taracha A, Kotarba G, Wilanowski T. Metody analizy fosforylacji białek [Methods of analysis of protein phosphorylation]. Postepy Biochem. 2017;63(2):137–142.
32. Peek GW, Tollefsbol TO. Down-regulation of hTERT and Cyclin D1 transcription via PI3K/Akt and TGF-β pathways in MCF-7 Cancer cells with PX-866 and Raloxifene. Exp Cell Res. 2016;344(1):95–102. doi:10.1016/j.yexcr.2016.03.022
33. Daniel M, Peek GW, Tollefsbol TO. Regulation of the human catalytic subunit of telomerase (hTERT). Gene. 2012;498(2):135–146. doi:10.1016/j.gene.2012.01.095
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.