Back to Journals » Infection and Drug Resistance » Volume 13
Distribution and Molecular Characterization of Resistance Gene Cassettes Containing Class 1 Integrons in Multi-Drug Resistant (MDR) Clinical Isolates of Pseudomonas aeruginosa
Authors Ahmadian L, Haghshenas MR, Mirzaei B ![]() , Norouzi Bazgir Z, Goli HR
, Norouzi Bazgir Z, Goli HR ![]()
Received 26 May 2020
Accepted for publication 16 July 2020
Published 11 August 2020 Volume 2020:13 Pages 2773—2781
DOI https://doi.org/10.2147/IDR.S263759
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Leila Ahmadian,1,2 Mohammad Reza Haghshenas,1,2 Bahman Mirzaei,3 Zahra Norouzi Bazgir,1,2 Hamid Reza Goli1,2
1Molecular and Cell Biology Research Center, Faculty of Medicine, Mazandaran University of Medical Sciences, Sari, Iran; 2Department of Medical Microbiology and Virology, Faculty of Medicine, Mazandaran University of Medical Sciences, Sari, Iran; 3Department of Medical Microbiology and Virology, School of Medicine, Zanjan University of Medical Sciences, Zanjan, Iran
Correspondence: Hamid Reza Goli
Department of Medical Microbiology and Virology, Faculty of Medicine, Mazandaran University of Medical Sciences, Farah Abad Blv, Khazar Square, Sari, Mazandaran, Iran
Tel +98 1133543081
Fax +98 1133543249
Email [email protected]
Purpose: The integrons, as the mobile exogenous elements, play a prominent role in the spreading of antimicrobial resistance genes from Pseudomonas aeruginosa clinical isolates to other bacteria. This study aimed to investigate the frequency of class 1 integrons andresistance gene cassettes carrying by them in clinical isolates as well as multidrug resistant P. aeruginosa.
Materials and Methods: A total of 100 clinical isolates of P. aeruginosa were collected from 5 hospitals in Mazandaran province, north Iran. The antibiotic susceptibility pattern of the isolates was evaluated using the disk agar diffusion method. Genomic DNAs were extracted and then the presence of class 1 integrons was detected by the PCR test. All PCR products of the positive isolates were sequenced for the detection of resistance gene cassettes by the Sanger method.
Results: Forty-one percent of the clinical isolates were multi-drug resistant. Also, 42% of the isolates were contained class 1 integron, and 61.9% of the integron positive isolates were detected as MDR. We detected 10 different gene cassettes sizing from 0.6 to 3.5 kb in the present study. The sequencing analysis of the internal variable regions of the class 1 integrons showed that the 0.75 kb gene cassette (aadB) was the most frequent resistance gene (54.76%) among all clinical isolates, as well as the MDR isolates. Other resistance genes detected in this study were included: aadA6-orfD (35.71%), aacA4-blaOXA-10 (21.42%), aadB-aacA4-blaOXA-10 (19.04%), blaOXA-10-aacA4-VIM 1 (11.9%), aacA4-catB10 (7.14%), aacA5-aadA1-cmlA5 (7.14%), blaOXA31-aadA2 (4.76%), and aac(3)-Ic-aacA5-cmlA5 (4.76%). To the best of our knowledge, blaOXA-10-aacA4-VIM1 cassette array is detected for the first time in this study.
Conclusion: The treatment of infections caused by P. aeruginosa in this region of Iran is a major problem due to the high prevalence of class 1 integrons. It seems that the high prescription of beta-lactams and aminoglycosides for the treatment of these infections may be replaced by other combination therapy stewardships.
Keywords: Pseudomonas aeruginosa, class 1 integrons, MDR, resistance gene cassettes
Introduction
Pseudomonas aeruginosa is an important opportunistic pathogen causing significant severe infections, especially in immunosuppressed patients.1 As the possession of different antibiotic resistance mechanisms, treatment of the infections caused by this organism is a major concern worldwide.2 Rapid spreading of drug resistance genes among bacteria by horizontal gene transfer mechanism is a challenge in this issue.3 Moreover, integrons have a prominent role in the spreading of resistance genes between different bacteria, particularly Gram-negative ones.4 Integrons, the mobile genetic elements when being part of a transposon or Insertion Sequence (IS), exist in gram-negative bacterial genomes and allow the bacteria to obtain, carry and transfer the variable new genetic elements as well as antibiotic resistance genes.5 The resistance against ß-lactams, carbapenems, chloramphenicol, aminoglycosides, and macrolides can gain by the gene cassettes carried by these genetic elements.6 Integrons are the flexible gene acquisition systems in bacterial genomes that allow the bacteria to gain and spread the new exogenous genes.6
All integrons have at least three essential regions between two conserved ends (5′CS and 3′CS) including intI, a gene encoding integrase enzymes, attI, a site which recognizes by integrases for the integration of the integron cassettes, and a pc promoter which leads to the transcription of encoded genes.7 Also, there is an internal variable region (IVR) between these conserved sections containing antibiotic or disinfectant resistance genes, called gene cassettes.4 Integrons can divide into five classes based on differences in amino acid sequences and relative homology of intI, which class 1 integrons are the most prevalent and clinically significant in P. aeruginosa.4,8 In this system, the new genes are acquired as parts of the gene cassettes.6 Cassettes are one of the simplest genetic mobile elements incapable of movement in the absence of integrons.5 However, the integrons can carry one to eight gene cassettes causing resistance to antimicrobial agents.5
Obtain and expression of the resistance gene cassettes by P. aeruginosa, causing secondary malignant infections in immunocompromised patients,9 can result in infection control problems in the clinical and health settings. Plasmids and transposons carrying antibiotic resistance genes provide the mobility of the integrons.6 These elements probably play a key role in the dissemination of multi-drug resistance (MDR) genes among the strains of P. aeruginosa or other bacteria.7,8 Among various known sub-classes, the class 1 integrons are well known and are the most prevalent among MDR P. aeruginosa clinical isolates.7 Dissemination of antimicrobial resistance genes by the class 1 integrons is a growing health concern, especially among Gram-negative bacteria.4
The present study aimed to investigate the frequency of class 1 integrons and characterization of resistance gene cassettes in clinical isolates as well as multidrug resistant P. aeruginosa collected from patients admitted to educational and therapeutic hospitals of Mazandaran province, north of Iran.
Materials and Methods
Collection of the Samples and Isolation of the Bacteria
A total of 100 non-duplicated (collected from non-repetitive patients) P. aeruginosa strains were isolated from different clinical specimens such as sputum, urine, wound, catheter, blood, and other samples. The specimens obtained from patients admitted to five educational and therapeutic hospitals of Mazandaran province, north of Iran, during 2016–2018. The bacterial isolates identified by the conventional microbiological tests,10 then purified and cultured in TSB medium (Merck, Germany) containing 10% glycerol and were frozen at −20°C until use.
Antimicrobial Susceptibility Testing
Disk agar diffusion method on Mueller-Hinton agar (Merck, Germany) was carried out according to the Clinical and Laboratory Standards Institute (CLSI) guidelines11 to detect the antibiotic resistance pattern of the clinical isolates against 13 antibiotics including ciprofloxacin (5 μg), levofloxacin (5 μg), amikacin (30 μg), tobramycin (10 μg), gentamicin (10 μg), imipenem (10 μg), meropenem (10 μg), doripenem (10 μg), aztreonam (30 μg), cefepime (30 μg), ceftazidime (30 μg), piperacillin (100 μg), and piperacillin/tazobactam (100/10 μg) (Rosco, Denmark). P. aeruginosa ATCC 27853 used as a control strain in antimicrobial susceptibility testing.
Genomic DNA Extraction
The genomic DNAs of the P. aeruginosa clinical isolates were extracted by alkaline lysis method using Sodium Dodecyl Sulfate (SDS) and NaOH.12 First, 3–5 colonies of the overnight grown bacteria were suspended in 60 μL of an alkaline solvent (0.5 gr SDS and 0.4 gr NaOH dissolved in 200 μL distilled water), then warmed at 95°C for 10 minutes. Next, centrifuged at 13000 rpm for 5 min, and 180 μL distilled water added to the mixture. The prepared supernatant was frozen at −20°C until the use as extracted DNA for PCR.
Detection of Class 1 Integrons Using PCR
Detection of class 1 integrons was performed by amplification of the extracted DNAs using specific primers: 5ʹ-CS-F: 5′-GGCATCCAAGCAGCAAGC-3′ and 3ʹ-CS-R: 5′-AAGCAGACTTGACCTGAT-3′)4 in the PCR method. These primer pairs amplify the internal variable regions (IVR) containing resistance gene cassettes between the 5ʹCS and 3ʹCS of the class 1 integrons. Thermal cycler running conditions with 15 μL final volume containing 7.5 μL Master Mix (Ampliqon, Denmark), 300 ng of extracted DNA, 10 pmol of each primer, and distilled water was as follows: one cycle of primary denaturation at 94°C for 5 min, and 35 cycles including denaturation at 94°C for 60 s, primer annealing at 59°C for 60 s, and the extension step at 72°C for 2 min. Also, the final extension step performed for 10 min at 72°C.
Sequencing of the Amplified Gene Cassettes of the Class 1 Integrons
To determine the genes inserted in IVRs of the integrons, first, the PCR products extracted from the electrophoresis gel by AccuPrep Gel Purification Kit (Bioneer Co., Daejeon, Korea) and the next, the extracted products were purified using AccuPrep PCR Purification Kit (Bioneer). The sequencing of the purified products was carried out by Applied Biosystems 3730/3730xl DNA Analyzers Sequencing (ABI, Bioneer). In the next time, we compared the sequenced nucleotides in GenBank databases at the National Center for Biotechnology Information Website (http://www.ncbi.nlm.nih.gov/BLAST/).
Statistical Analysis
The Statistical Package for the Social Sciences (SPSS) software version 22 used for analyzing the data and a comparison of the data performed by the Chi-square test. P-values < 0.05 were reflected as statistical significant.
Ethical Approval Statements
We did not interfere with the subjects and did not have a direct relationship with the patients. We received the clinical samples of the patients without names from the laboratories of the studied hospitals affiliated to the Mazandaran University of Medical Sciences and performed the desired tests on these samples. This study was conducted in accordance with the Declaration of Helsinki, however written informed consent form was provided by the patients or a close relative before hospitalization, and categorizing information of each sample was reserved secret. Also, this study was approved by the Iran National Committee for Ethics in Biomedical Research with the national ethical code (consent ref number) IR.MAZUMS.REC.1398.075 (Supplementary data).
Results
Bacterial Isolates and Antimicrobial Susceptibility Testing
Out of 100 P. aeruginosa clinical isolates, 91% of them obtained from hospitalized patients and the other 9% acquired from out-patients. The resistance pattern of the isolates is shown in Table 1. A comparison of the antibiotic susceptibility pattern among integron positive and negative isolates indicated that the resistance rate among integron positive isolates was higher than integron negative ones. These differences were statistically significant (P-value < 0.05) in terms of all tested antibiotics, except ceftazidime (Table 1). Piperacillin-tazobactam and levofloxacin were the most and least effective antibiotics in the present study, respectively. Moreover, differences in the antibiotic resistance rate between MDR and non-MDR isolates were statistically significant (p < 0.05) about all tested antibiotics (Table 2). The frequency of the MDR phenotype among the isolates was 41% (Table 2), and 42% of all isolates contained the class 1 integrons. Also, 61.9% of the integron positive isolates were detected as MDR, while 63.41% of the MDR isolates were carrying class 1 integrons.
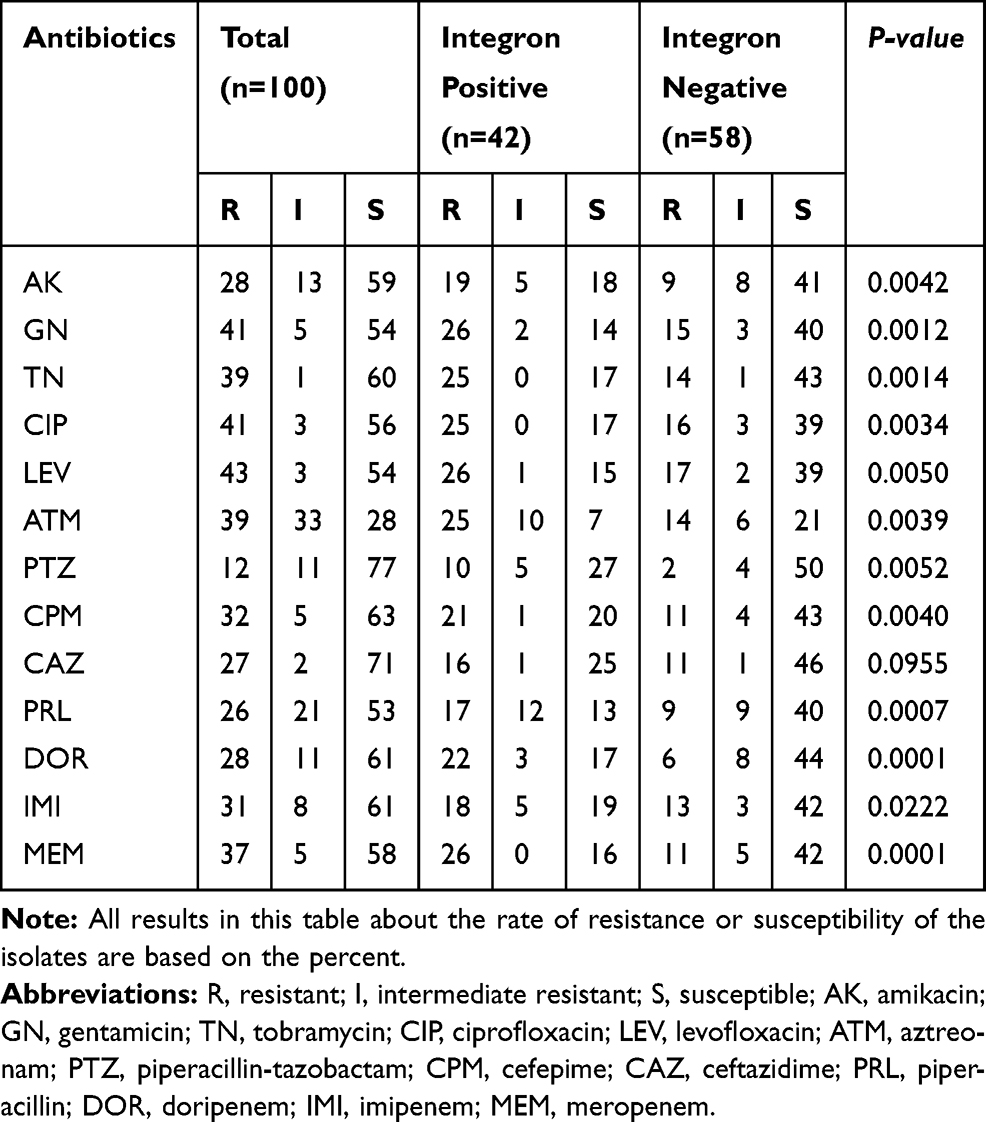 |
Table 1 The Comparison of Antibiotic Resistance Profile Between the Integron Positive and Negative Clinical Isolates of Pseudomonas aeruginosa |
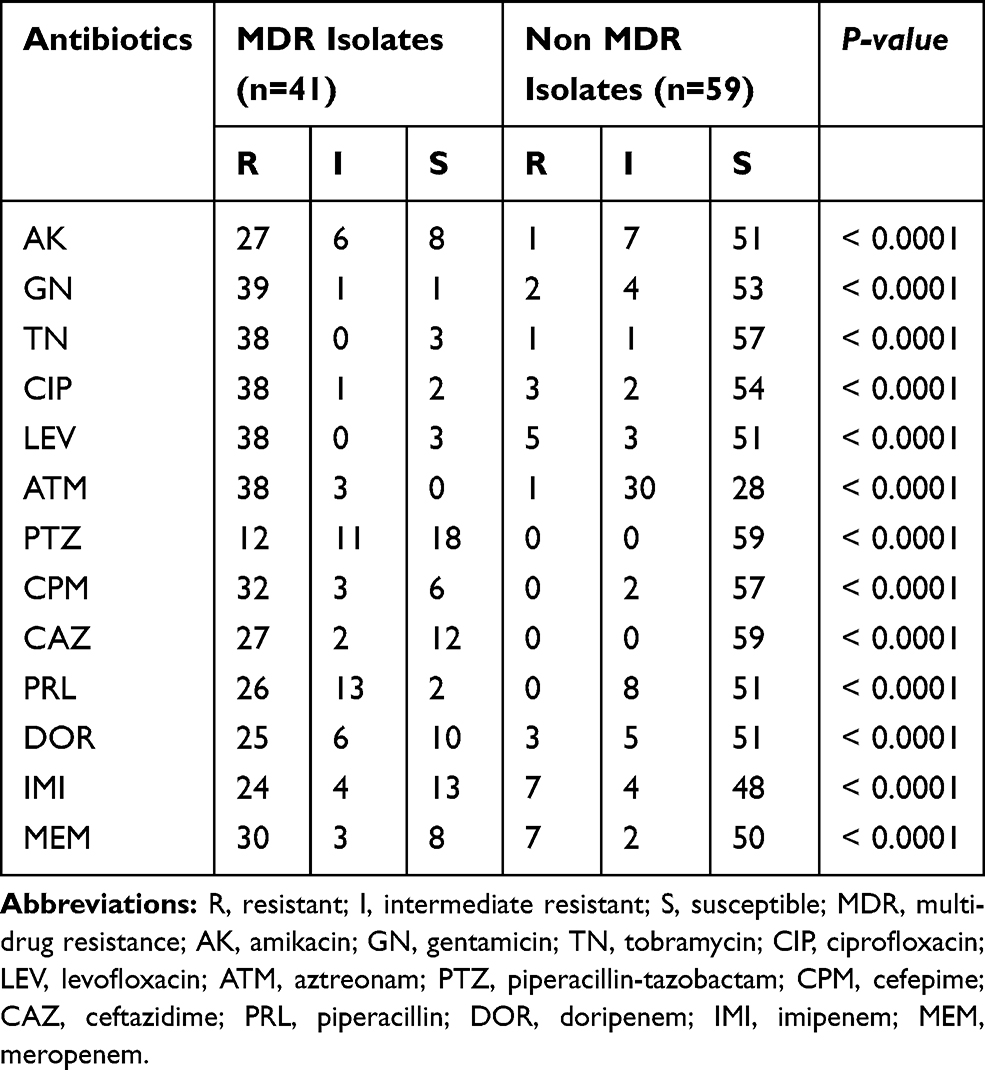 |
Table 2 The Comparison of Resistance Percent Rates Against 13 Tested Antibiotics Among the MDR and Non-MDR Isolates of Pseudomonas aeruginosa |
Class 1 Integrons and Sequencing Analysis
Out of all integron positive isolates, 20 (47.6%) isolates were carrying more than one gene cassette, 13 (30.9%) isolates had two cassettes, and 7 (16.6%) isolates contained three resistance gene cassettes (Table 3). Out of 20 isolates containing more than one gene cassette, 15 (75%) isolates identified as MDR. In the present study, 69 gene cassettes observed in 10 different variable regions detected in the PCR method sizing from 0.6 to 3.5kb (Table 3). The 0.6 kb gene cassette encoding a hypothetical protein found in 2 (4.7%) isolates, while the 0.75 kb resistance cassette was the most frequently observed (54.76%) in the present study, either alone or in combination with other resistance gene cassettes. The frequency of other resistance gene cassettes identified in this study included aadA6-orfD (35.71%), aacA4-blaOXA-10 (21.42%), aadB-aacA4-blaOXA-10 (19.04%), blaOXA-10-aacA4-VIM1 (11.9%), aacA4-catB10 (7.14%), aacA5-aadA1-cmlA5 (7.14), blaOXA-31-aadA2 (4.76), and aac(3)-Ic-aacA5-cmlA5 (4.76). The novelty of this study was the emergence of a new gene cassette array (blaOXA-10-aacA4-VIM1) in this region. All detected gene cassettes, the number of MDR isolates, and the resistance rate of integron positive isolates presented in Table 3. Moreover, the most prevalent MDR strains were isolated from catheters, while the strains collected from the wound and catheter samples detected as more prevalent integron positive isolates (Table 4). Also, we found that 63.6% and 72.7% of our burn isolates were MDR and class 1 integron positive, respectively (Figure 1).
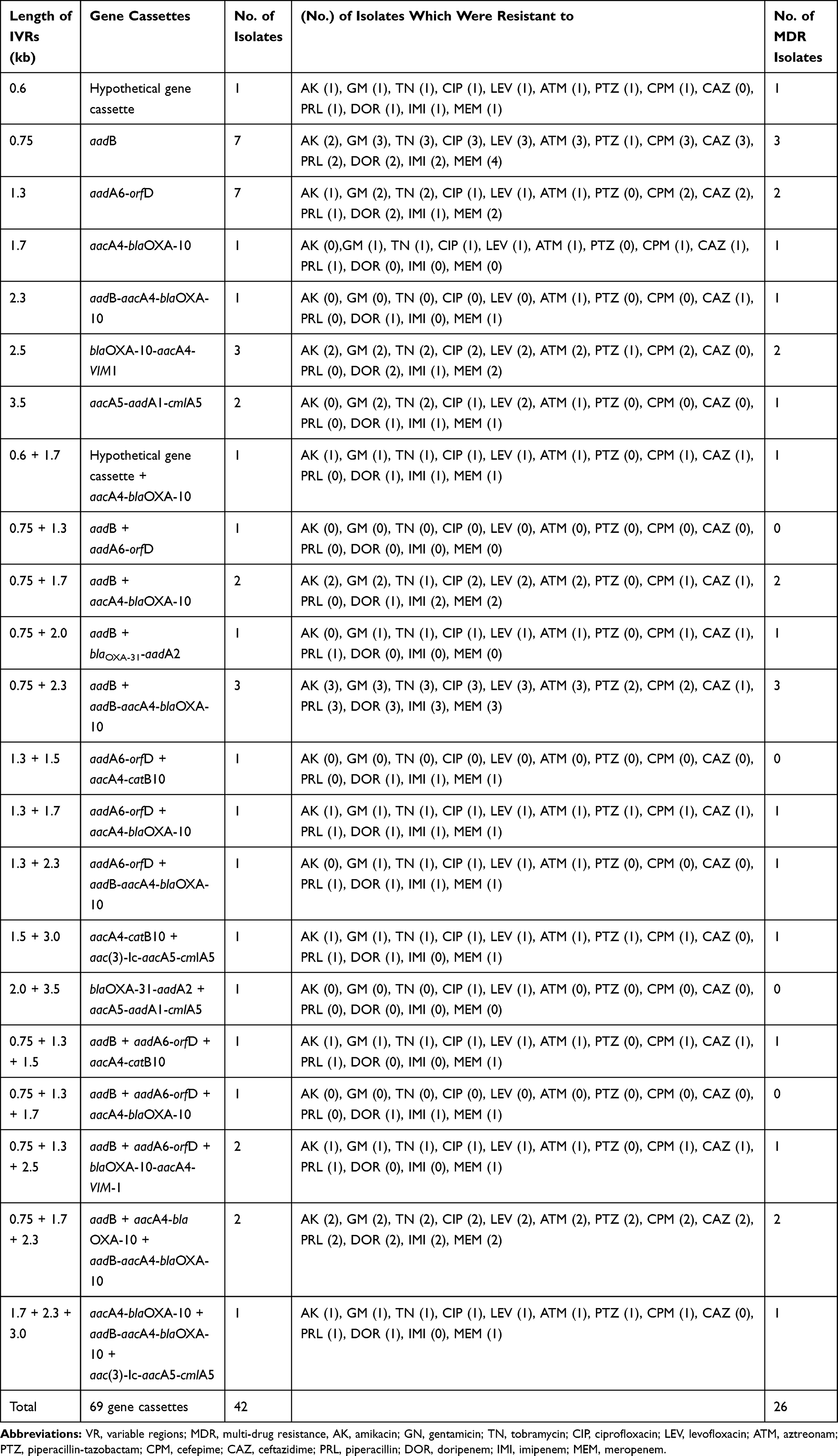 |
Table 3 The Prevalence of Different Resistance Gene Cassette Arrays in the Clinical Isolates of Pseudomonas aeruginosa and Their Correlation with the Resistance Towards Tested Antibiotics |
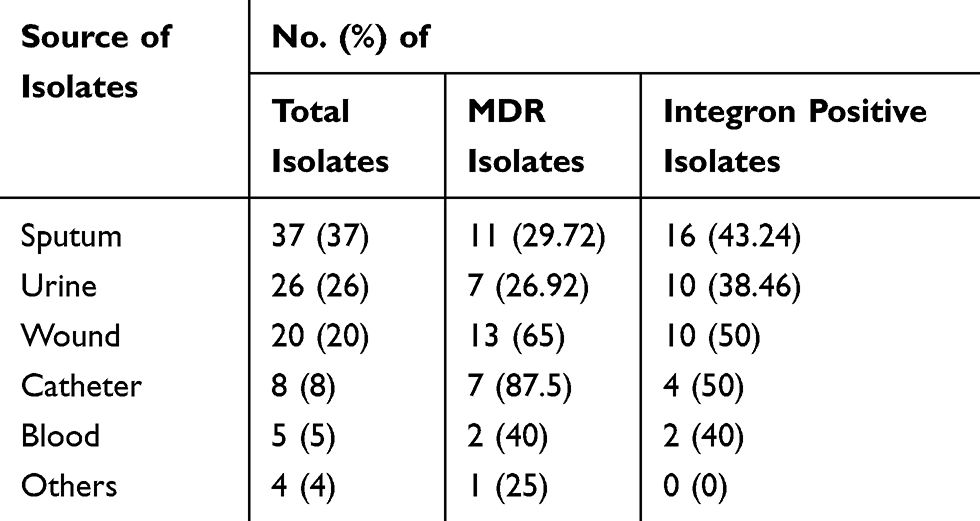 |
Table 4 Prevalence of MDR and Integron Positive Pseudomonas aeruginosa Clinical Isolates Based on the Source of Isolation |
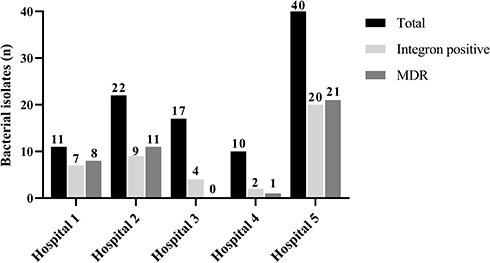 |
Figure 1 The number of integron positive and MDR Pseudomonas aeruginosa clinical isolates according to the hospital sources. |
Discussion
Dissemination of resistance gene cassettes by class 1 integrons among P. aeruginosa clinical isolates causes therapeutic failures as well as the high mortality rate in immunocompromised patients.3 This study exhibited the genotypic characterization of the class 1 integrons detected in the clinical isolates of P. aeruginosa. In the present study, 41% of the isolates were at least resistant to three different classes of antibiotics and detected as multi-drug resistant (MDR). The other study in north Iran showed that 72.2% of their isolates were MDR.13 It seems that the high antibiotic resistance rate of the burn isolates was the major factor of this significant difference in our total MDR isolates and the mentioned study. In contrast, Shahcheraghi et al in Tehran reported 5.4% of their isolates were MDR.14 Other studies in Iran1,2,4,15,16 showed different rates (40% to 97.5%) of MDR clinical isolates of P. aeruginosa based on different sources of samples and hospital wards of their collection. However, we found that P. aeruginosa isolated from the catheters and burn wounds are more capable for the acquisition of antibiotic resistance genes, as exhibited the highest prevalence of MDR phenotypes (Table 4).
On the other hand, our outcomes about the frequency of class 1 integron positive isolates were lower than similar studies carried out in Iran, which reported the rate of 43% to 95.7% class 1 integron positive P. aeruginosa clinical isolates.1,2,4,16-20 It seems that the high incidence of burn isolates in some reviewed studies4,17,18,20 may be the major factor of differences. Generally, the isolates collected from burn injuries show a higher resistance rate may be due to the high antibiotic selection pressure in these patients, as they are high risk for acquiring bacterial infections.21 Moreover, Heidarzadeh et al analyzed the studies in 2000–2018 and reported that the prevalence of class 1 integrons collected from Iranian burn patients varied from 40% to 96.9%.22 This issue confirmed in the present study, as 63.6% and 72.7% of our clinical isolates collected from a burn hospital were MDR and class 1 integron positive, respectively (Figure 1). However, among other hospitals in the current study, we found that these rates in clinical isolates collected from patients hospitalized in infectious centers (Figure 1) were higher than pediatric and heart centers, indicating the lower antibiotic selection pressure in these wards. On the other hand, the inappropriate use of antibiotics may be one of the most important causes, resulting in the high rate of integron dissemination in these regions of Iran.
Furthermore, 26% of the class 1 integron positive isolates in this study were discovered as multi-drug resistant (MDR), while this rate in a previous study was 64%.4 These results confirmed that there is a significant correlation among the presence of class 1 integrons and the emergence of MDR phenotypes of P. aeruginosa. Acquiring the class 1 integrons exposures the bacteria to exogenous genes and inducing antibiotic resistance and development of MDR isolates.14 The low MDR rate among the integron positive isolates in our study indicates that other resistance mechanisms such as efflux pumps, drug inactivation by enzymes, decreased influx, etc. may be the causes of MDR phenotype emerging in the current study, as the co-existence of class 1 integrons and other resistance mechanisms is the cause of the high rate emergence of MDR phenotype in Gram-negative bacteria.23 The prevalence of class 1 integrons in P. aeruginosa isolates is much variable in different geographical locations. For example, a similar study conducted in Turkey (Northwest neighbor of Iran) reported that just 4.8% of their P. aeruginosa isolates were carrying class 1 integrons.24 In contrast, this rate in Tunisia and Nigeria were >50% of their collected isolates.25,26 However, our results about the frequency of class 1 integrons among MDR P. aeruginosa isolates (63.4%) were comparable with a similar study carried out in Egypt (61.1%).27 This difference may be due to the different policies and management of antibiotics for the treatment of infections in the world. Also, this rate was more varied in the southeast Asia from 38% to 95%,3,28-32 while 22% to 47% of the P. aeruginosa clinical isolates in European countries were contained the class 1 integrons between 2007 to 2013.9,33,34 These reports show the high rate dissemination of class 1 integrons in all regions of the world, and if this problem ignored, these mobile elements can spread more rapidly worldwide, concluding the high antibiotic resistance rates, and serious problems in the treatment of the infections. The rate of MDR phenotype in isolates gained from our burned patients were (88%), while 44% of these isolates were integron positive. In another study conducted in northwest Iran,4 100% of their burn isolates detected as MDR and integron positive. These results may be due to the high antibiotic pressure and long hospitalization of burned patients. Also, 43% of ICU isolates in the present study were MDR, and 47% of them were integron positive, which was similar to the above-mentioned study,4 confirming the high antibiotic pressure of the ICU hospitalized patients.
Concordance with other similar studies conducted recently in Iran,4,8,14 we found that the frequency and variety of resistance gene cassettes are increasing in this country, requiring strong management and practical control. The molecular characterization of class 1 integrons showed that the numerous aminoglycoside modifying enzymes encoding genes are spreading by this mobile genetic element, however, in our study, the aadB gene was the most prevalent gene cassette similar to the results of other studies carried out in Iran.4,14 The aadB gene encodes aminoglycoside(2”) adenylyltransferase [ANT(2”)-Ia] enzyme and leads to resistance against kanamycin, gentamicin, and tobramycin,35 however 65% and 60% of the aadB positive isolates of the present study were resistant to gentamicin and tobramycin, respectively. Also, the aadA6-orfD array (1.3 kb) was the second identified cassette (35.7%). This array was the most frequently reported gene cassette in Thailand, and the third reported cassette in the northwest Iran,4,30 while only one isolate of other Iranian research was aadA6-orfD positive.14 The aadA6 encodes the other type of aminoglycoside modifying enzyme and is associated with the resistance against streptomycin and spectinomycin.3,35 The orfD has an unidentified role in the bacteria and may encode a protein that is important for the bacterial community and their ecology.35 The aacA4 gene, encoding the aminoglycoside (6ʹ)-N-acetyltransferase [AAC(6ʹ)-II] enzyme, was detected in 4 different cassettes sizing 1.5, 1.7, 2.3, and 2.5 kb, identified in 22 (52.38%) isolates in combination with other genes in the current study, while this gene has a key role in resistance towards gentamicin, tobramycin, netilmicin, and kanamycin.35 However, among the aacA4 positive isolates of this study, 77% and 72% of the isolates were resistant to gentamicin and tobramycin, respectively. The blaOXA-10 gene confers resistance to some β-lactams such as carboxypenicillins and ureidopenicillins,24 however, the aacA4-blaOXA-10 array (1.7 kb) was identified in 21.4% of our integron positive isolates. On the other hand, the first report of aadB-aacA4-blaOXA-10 (2.3 kb), aac(3)-Ic-aacA5-cmlA5 (3.0 kb) and aacA5-aadA1-cmlA5 (3.5kb) arrays from Iran were in 2017 from Tabriz by Goli et al.4 Surprisingly, these resistance gene cassette arrays detected in 8 (19.04%), 2(4.7%), and 3 (7.1%) integron positive isolates of the present study, too. The aac(3)-Ic gene, encoding aminoglycoside 3-N-acetyltransferase, is associated with the resistance to gentamicin and was the first reported in P. aeruginosa from Italy in 2003.35,36 The aacA5 (aac(3)-Ie) gene encodes the aminoglycoside 3-N-acetyltransferase leading the resistance against gentamicin.36 The cmlA5 gene is associated with the resistance to chloramphenicol by encoding an efflux pump.37
To the best of our knowledge, we identified one new cassette array, including blaOXA-10-aacA4-VIM1 in 5 (11.9%) clinical isolates of P. aeruginosa (Figure 2) for the first time. The VIM-1 encodes a class B extended-spectrum beta-lactamase, which degrades the most β-lactams, including penicillins and carbapenems.38 However, among the blaOXA-10-aacA4-VIM1 positive isolates of the present study, more than 60% of them were resistant to β-lactams.
 |
Figure 2 The size and structure of resistance gene cassettes between two conserved ends (5′CS and 3′CS) of the class 1 integrons detected in the clinical isolates of Pseudomonas aeruginosa. |
Conclusion
Overall, the results of this study and other similar studies demonstrated the high rate of antibiotic resistance, MDR phenotype and the presence of class 1 integrons in the clinical isolates of Pseudomonas aeruginosa in Iran and other countries. Since the existence of class 1 integrons has a strong role in spreading the antibiotic resistance genes, emerging the multidrug resistant strains, among the clinical isolates of P. aeruginosa, it seems that we need correct antibiotic stewardship worldwide to overtop this problem. We think that the variation of antibiotic usage policy in hospitalized patients according to the results of routine antimicrobial susceptibility testing, especially for Gram-negative bacteria, may help to receive this issue. On the other hand, the evaluation of integron presence in the clinical isolates of the bacteria by the PCR method may need to be a routine test in hospitals, helping the best management and effective control and treatment of bacterial infections.
Acknowledgments
We thank the laboratory staff of BuAli Sina, Imam Khomeini, Zare, Fatemeh Zahra, and Razi hospitals for providing patients’ information and the collection of the clinical isolates.
Disclosure
The authors declare no conflict of interest.
References
1. Sharifi H, Pouladfar G, Shakibaie MR, et al. Prevalence of β-lactamase genes, class 1 integrons, major virulence factors and clonal relationships of multidrug-resistant Pseudomonas aeruginosa isolated from hospitalized patients in southeast of Iran. Iran J Basic Med Sci. 2019;22:806–812. doi:10.22038/ijbms.2019.35063.8340
2. Nikokar I, Tishayar A, Flakiyan Z, et al. Antibiotic resistance and frequency of class 1 integrons among Pseudomonas aeruginosa, isolated from burn patients in Guilan, Iran. Iran J Microbiol. 2013;5:36.
3. Poonsuk K, Tribuddharat C, Chuanchuen R. Class 1 integrons in Pseudomonas aeruginosa and Acinetobacter baumannii isolated from clinical isolates. South Asian J Trop Med Public Health. 2012;43:376.
4. Goli HR, Nahaei MR, Rezaee MA, et al. Prevalence and molecular characterization of Class 1 integrons among clinical isolates of Pseudomonas aeruginosa in Northwest of Iran. Mol Genet Microbiol Virol. 2017;32:109–115. doi:10.3103/S0891416817020057
5. Hall RM. Integrons and gene cassettes: hotspots of diversity in bacterial genomes. Ann New York Academy Sci. 2012;1267:71–78. doi:10.1111/j.1749-6632.2012.06588.x
6. Gillings MR. Integrons: past, present, and future. Microbiol Mol Biol Rev. 2014;78:257–277. doi:10.1128/MMBR.00056-13
7. Salimizadeh Z, Hashemi Karouei SM, Hosseini F. Dissemination of Class 1 Integron among different multidrug resistant Pseudomonas aeruginosa Strains. Med Lab J. 2018;12:36–42. doi:10.29252/mlj.12.4.36
8. Mobaraki S, Aghazadeh M, Soroush Barhaghi MH, et al. Prevalence of integrons 1, 2, 3 associated with antibiotic resistance in Pseudomonas aeruginosa isolates from Northwest of Iran. BioMed. 2018;8:2. doi:10.1051/bmdcn/2018080102
9. Ruiz-Martínez L, López-Jiménez L, Fusté E, et al. Class 1 integrons in environmental and clinical isolates of Pseudomonas aeruginosa. Int J Antimicrob Agents. 2011;38:398–402. doi:10.1016/j.ijantimicag.2011.06.016
10. Sahm DF, Weissfeld A, Trevino E. Baily and Scott’s Diagnostic Microbiology. St Louis: Mosby; 2002.
11. Wayne PA. Performance Standards for Antimicrobial Susceptibility Testing. Clinical and Laboratory Standards Institute. 2018.
12. Šipošová NŠ, Liptáková V, Kvasnová S, et al. Genetic diversity of Acinetobacter spp. adapted to heavy metal polluted environments. Nova Biotechnol Chim. 2017;16:42–47. doi:10.1515/nbec-2017-0006
13. Asadpour L. Antimicrobial resistance, biofilm-forming ability and virulence potential of Pseudomonas aeruginosa isolated from burn patients in northern Iran. J Glob Antimicrob Res. 2018;13:214–220. doi:10.1016/j.jgar.2018.01.018
14. Shahcheraghi F, Badmasti F, Feizabadi MM. Molecular characterization of class 1 integrons in MDR Pseudomonas aeruginosa isolated from clinical settings in Iran, Tehran. FEMS Immunol Med Microbiol. 2010;58:421–425. doi:10.1111/j.1574-695X.2009.00636.x
15. Goudarzi SM, Eftekhar F. Multidrug resistance and integron carriage in clinical isolates of Pseudomonas aeruginosa in Tehran, Iran. Turk J Med Sci. 2015;45:789–793. doi:10.3906/sag-1408-120
16. Zarei-Yazdeli M, Eslami G, Zandi H, et al. Prevalence of class 1, 2 and 3 integrons among multidrug-resistant Pseudomonas aeruginosa in Yazd, Iran. Iran J Microbiol. 2018;10:300–306.
17. Azami S, Abdi AA, Asgarani E. Association between metallo-β-lactamases and integrons with multi-drug resistance in Pseudomonas aeruginosa isolates. J Med Microbiol Infect Dis. 2013;1:46–51.
18. Yousefi S, Nahaei M, Farajnia S, et al. Class 1 integron and imipenem resistance in clinical isolates of Pseudomonas aeruginosa: prevalence and antibiotic susceptibility. Iran J Microbiol. 2010;2:115.
19. Faghri J, Nouri S, Jalalifar S, et al. Investigation of antimicrobial susceptibility, class I and II integrons among Pseudomonas aeruginosa isolates from hospitalized patients in Isfahan, Iran. BMC Res Notes. 2018;11:806. doi:10.1186/s13104-018-3901-9
20. Khosravi AD, Motahar M, Abbasi Montazeri E. The frequency of class1 and 2 integrons in Pseudomonas aeruginosa strains isolated from burn patients in a burn center of Ahvaz, Iran. PLoS One. 2017;12:e0183061. doi:10.1371/journal.pone.0183061
21. Laupland K, Kibsey P, Gregson D, et al. Population-based laboratory assessment of the burden of community-onset bloodstream infection in Victoria, Canada. Epidem Infect. 2013;141:174–180. doi:10.1017/S0950268812000428
22. Heidarzadeh S, Enayati Kaliji Y, Pourpaknia R, et al. A meta-analysis of the prevalence of class 1 integron and correlation with antibiotic resistance in Pseudomonas aeruginosa recovered from Iranian burn patients. J Burn Care Res. 2019;40:972–978. doi:10.1093/jbcr/irz135
23. Livermore DM. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: our worst nightmare? Clin Infect Dis. 2002;34:634–640. doi:10.1086/338782
24. Cicek AC, Saral A, Duzgun AO, et al. Screening of Class 1 and Class 2 integrons in clinical isolates of Pseudomonas aeruginosa collected from seven hospitals in Turkey: a multicenter study. Open J Med Microbiol. 2013;3:227. doi:10.4236/ojmm.2013.34034
25. Chairat S, Ben Yahia H, Rojo-Bezares B, et al. High prevalence of imipenem-resistant and metallo-beta-lactamase-producing Pseudomonas aeruginosa in the Burns Hospital in Tunisia: detection of a novel class 1 integron. J Chemother. 2019;31:120–126. doi:10.1080/1120009X.2019.1582168
26. Odumosu BT, Adeniyi BA, Chandra R. Analysis of integrons and associated gene cassettes in clinical isolates of multidrug resistant Pseudomonas aeruginosa from Southwest Nigeria. Ann Clin Microbiol Antimicrob. 2013;12:29. doi:10.1186/1476-0711-12-29
27. Kadry AA, Serry FM, El-Ganiny AM, et al. Integron occurrence is linked to reduced biocide susceptibility in multidrug resistant Pseudomonas aeruginosa. British J Biomed Sci. 2017;74:78–84. doi:10.1080/09674845.2017.1278884
28. Gu B, Tong M, Zhao W, et al. Prevalence and characterization of class I integrons among Pseudomonas aeruginosa and Acinetobacter baumannii isolates from patients in Nanjing, China. J Clin Microbiol. 2007;45:241–243. doi:10.1128/JCM.01318-06
29. Chen J, Su Z, Liu Y, et al. Identification and characterization of class 1 integrons among Pseudomonas aeruginosa isolates from patients in Zhenjiang, China. Int J Infect Dis. 2009;13(6):717–721. doi:10.1016/j.ijid.2008.11.014
30. Kiddee A, Henghiranyawong K, Yimsabai J, et al. Nosocomial spread of class 1 integron-carrying extensively drug-resistant Pseudomonas aeruginosa isolates in a Thai hospital. Int J Antimicrob Agents. 2013;42:301–306. doi:10.1016/j.ijantimicag.2013.05.009
31. Khosravi Y, Vellasamy KM, Tay ST, et al. Molecular detection and characterization of Metallo-β-lactamase (MBL) genes and integrons of imipenem–resistant Pseudomonas aeruginosa. A Méndez-Vilas, Ed. 2013;515–521.
32. Wu Y, Li H, Li J. Detection of Pseudomonas aeruginosa carried a new array of gene cassettes within class 1 integron isolated from a teaching hospital in Nanjing, China. J Microbiol. 2008;46:687–691. doi:10.1007/s12275-008-0021-6
33. Nemec A, Krizova L, Maixnerova M, et al. Multidrug-resistant epidemic clones among bloodstream isolates of Pseudomonas aeruginosa in the Czech Republic. Res Microbiol. 2010;161:234–242. doi:10.1016/j.resmic.2010.02.002
34. Henrichfreise B, Wiegand I, Pfister W, et al. Resistance mechanisms of multiresistant Pseudomonas aeruginosa strains from Germany and correlation with hypermutation. Antimicrob Agents Chemother. 2007;51:4062–4070. doi:10.1128/AAC.00148-07
35. Azucena E, Mobashery S. Aminoglycoside-modifying enzymes: mechanisms of catalytic processes and inhibition. Drug Res Updates. 2001;4:106–117. doi:10.1054/drup.2001.0197
36. Riccio ML, Docquier J-D, Dell’Amico E, et al. Novel 3-N-aminoglycoside acetyltransferase gene, aac (3)-Ic, from a Pseudomonas aeruginosa integron. Antimicrob Agents Chemother. 2003;47:1746–1748. doi:10.1128/AAC.47.5.1746-1748.2003
37. Naas T, Mikami Y, Imai T, et al. Characterization of In53, a class 1 plasmid-and composite transposon-located integron of Escherichia coli which carries an unusual array of gene cassettes. J Bacteriol. 2001;183:235–249. doi:10.1128/JB.183.1.235-249.2001
38. Sacha P, Wieczorek P, Hauschild T, et al. Metallo-beta-lactamases of Pseudomonas aeruginosa-a novel mechanism resistance to beta-lactam antibiotics. Folia Histochem Cyto. 2008;46:137–142.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.