Back to Journals » Research and Reports in Tropical Medicine » Volume 17
Distinct Genetic Footprint of Adaptation Observed Within Anopheles gambiae Taxa from The Gambia
Authors Milugo TK, Oriero EC, Hamid-Adiamoh M, Sadio A, Jawara MS, Gibba B, Ndiath MO ![]() , Clarkson CS, Miles A, Rajatileka S, Mohammed NI, Erhart A, D’Alessandro U, Assogba BS, Amambua-Ngwa A
, Clarkson CS, Miles A, Rajatileka S, Mohammed NI, Erhart A, D’Alessandro U, Assogba BS, Amambua-Ngwa A
Received 26 February 2026
Accepted for publication 24 June 2026
Published 16 July 2026 Volume 2026:17 605379
DOI https://doi.org/10.2147/RRTM.S605379
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Mario Rodríguez-Pérez
Trizah Koyi Milugo,1,* Eniyou Cheryll Oriero,1,* Majidah Hamid-Adiamoh,1,* Abdoulaye Sadio,1 Musa S Jawara,1 Balla Gibba,2 Mamadou Ousmane Ndiath,1 Chris S Clarkson,3 Alistair Miles,4 Shavanthi Rajatileka,5 Nuredin I Mohammed,1 Annette Erhart,1 Umberto D’Alessandro,1 Benoit Sessinou Assogba,1 Alfred Amambua-Ngwa1,5
1Medical Research Council Unit The Gambia at LSHTM, Atlantic Boulevard, Fajara, The Gambia; 2National Malaria Control Program, Ministry of Health, Banjul, The Gambia; 3Liverpool School of Tropical Medicine, Liverpool, UK; 4Ellison Institute of Technology, Oxford, UK; 5Wellcome Sanger Institute, Hinxton, UK
*These authors contributed equally to this work
Correspondence: Alfred Amambua-Ngwa, Medical Research Council Unit The Gambia at LSHTM, Atlantic Boulevard, Fajara, The Gambia, Email [email protected] Benoit Sessinou Assogba, Medical Research Council Unit The Gambia at LSHTM, Atlantic Boulevard, Fajara, The Gambia, Email [email protected]
Purpose: Emerging evidence suggests presence of novel or unclassified taxa within the Anopheles gambiae s.l. species complex, the principal vector of malaria in sub-Saharan Africa. The response of these new lineages to standard vector control tools and/or adaptation to selection pressure is currently unknown, underscoring the need for further characterization. We performed a genome-wide selective scan (GWSS) analysis to gain genomic insights into the local adaptation of An. gambiae Bissau molecular form (hereafter "Bissau”), a recently described taxon from The Gambia.
Results: Genomic regions under selection in Bissau primarily code for clusters of gene families associated with metabolic resistance mechanisms against insecticides, specifically Cyp6aa/Cyp6p, Cyp9K, and Gste. Notably, a distinct signal of selection for the Choline acetyltransferase gene (ChAT) was detected in the chromosome arm 2R of Bissau. In the sibling taxa An. coluzzii, two distinctive signals of selection overlapping E-ubiquitin protein and N-acetyltransferase genes were evident. These unique genes warrant further functional validation and monitoring to confirm their potential role as molecular markers for target site resistance. A selective sweep spanning clusters of odorant receptor (OR) genes, usually associated with chemosensory, was also unique to Bissau. Diplotype clustering of OR regions revealed six groups with low heterozygosity and non-synonymous mutations, suggesting local adaptation to fundamental biological traits such as host seeking and habitat selection. Haplotype clustering analysis ruled out adaptive sharing of OR alleles, depicting ongoing independent evolution of chemosensory perception in Bissau population. Contrastingly, the sharing of adaptive alleles was evident between An. gambiae s.s and Bissau in regions coding metabolic resistance genes, potentially facilitating the spread of insecticide resistance across taxa.
Conclusion: Common and taxon-specific signatures of selection are evolving in An. gambiae s.l. needing further studies and monitoring to guide targeted vector control efforts.
Keywords: Bissau molecular form, malaria vectors, insecticide resistance, sweeps selection, Anopheles gambiae
Introduction
The Anopheles gambiae complex (Diptera: Culicidae) is a primary malaria vector in sub-Saharan (SSA).1 The complex comprises nine sibling species, namely Anopheles arabiensis, Anopheles coluzzii, An. gambiae s.s, Anopheles melas, Anopheles merus, An. quadriannulatus, Anopheles bwambae, Anopheles amharicus and Anopheles fontenillei, that are morphologically identical but genetically dissimilar.2–4 The dissimilarity is, in part, a consequence of local adaptation to survive and breed in diverse environments. When adapting to new environments, beneficial alleles may accumulate in the genome and become fixed in the mosquito population over time.5 Genomic regions harbouring these alleles, detected as signatures of selection (ie, selective sweeps), can offer insight into mosquito evolutionary history and genetic diversity.6 Indeed, selective sweeps have been explored to understand resistance mechanisms in malaria mosquitoes, underlying adaptation to vector control measures:7 The most reported are sweeps that host metabolic (ie cytochrome P450 and GSTe-2) and target site (ie Vgsc, Rdl and Ace-1) resistance genes.8–10
In West Africa, the most commonly reported target site mutations in An. gambiae complex includes kdr-w (L1014F) and N1575Y in the Vgsc gene which are associated with resistance to pyrethroids and DDT.11,12 The others include Ace-1 G119S mutation associated with resistance to carbamates and organophosphates, as well as the A302G, A302S and A296S-RDL mutations associated with resistance to dieldrin.11,13–15 For metabolic resistance, commonly reported variants are in cytochrome P450 CYP6P3, CYP6M2, and CYP9K1 and these are associated with resistance to the pyrethroid deltamethrin.16,17 Additionally, it is not unusual to observe the co-occurrence of multiple resistance mechanisms, including L119F-GSTe2 that confers resistance to DDT.18,19 Importantly, additional and new resistance-associated variants continue to emerge across African malaria vector species driving phenotypic adaptation to pesticides, anthropogenic events and/or changing climate.
Geographically, emerging and primary vectors exist in sympatry, allowing both ecological and genomic interactions.20 Genomic interactions between these species may include adaptive introgression of alleles that have important consequences, such as resistance to insecticides, immune response and/or vector competence.21,22 Notably, change in mosquito behavior and ecological interactions can also be driven by chromosomal inversion for example the 2La inversion in An. gambiae and An. coluzzii that has been associated with change in resting preferences, desiccation tolerance, thermal resistance and resistance to insecticides.23,24 These vector adaptations can impact malaria transmission dynamics and erode successes towards malaria elimination, underscoring the need to characterize genetic diversity and evolutionary mechanism in all molecular forms and cryptic taxa of An. gambiae s.l. across local populations to inform the design of effective and targeted vector control measures.
In the Gambia, primary analysis of the genomes of the An. gambiae Bissau molecular form (hereafter Bissau) identified mutations associated with metabolic resistance.25,26 However, an exhaustive genomic scan of signatures of local adaptation in Bissau in relation to sibling species has not been conducted. As the taxa could be identified across different ecoregions of the Gambia, different populations may have taken parallel evolutionary trajectories. Moreover, it is unknown if there is adaptive gene flow between Bissau and other sibling taxa, important for designing and implementing vector control measures in The Gambia.
Our study seeks to unravel the genetic architecture underlying local adaptation in Bissau mosquitoes from The Gambia using whole-genome variation data from the An. gambiae genome project (AG1000G) Phase 3(Ag3). By examining patterns of selection and genetic exchange among taxa, we aim to identify selective sweeps while looking for evidence of gene flow across taxa.
Materials and Methods
Data Processing and Code Availability
The genomic data used in this analysis were obtained from the Anopheles gambiae genome project (AG1000G) phase 3(Ag3). Analysis was restricted to 2658 mosquito samples identified as An. gambiae s.l, collected between 2011 and 2021, across different sites in The Gambia (Table 1). To reduce background noise and ensure statistical robustness27,28 in haplotype frequencies, selection scan analysis was performed for sites with a minimum sample size set at 10 or a subset of 100 maximum at 100 for larger sample size, thereby balancing the census size for comparability.
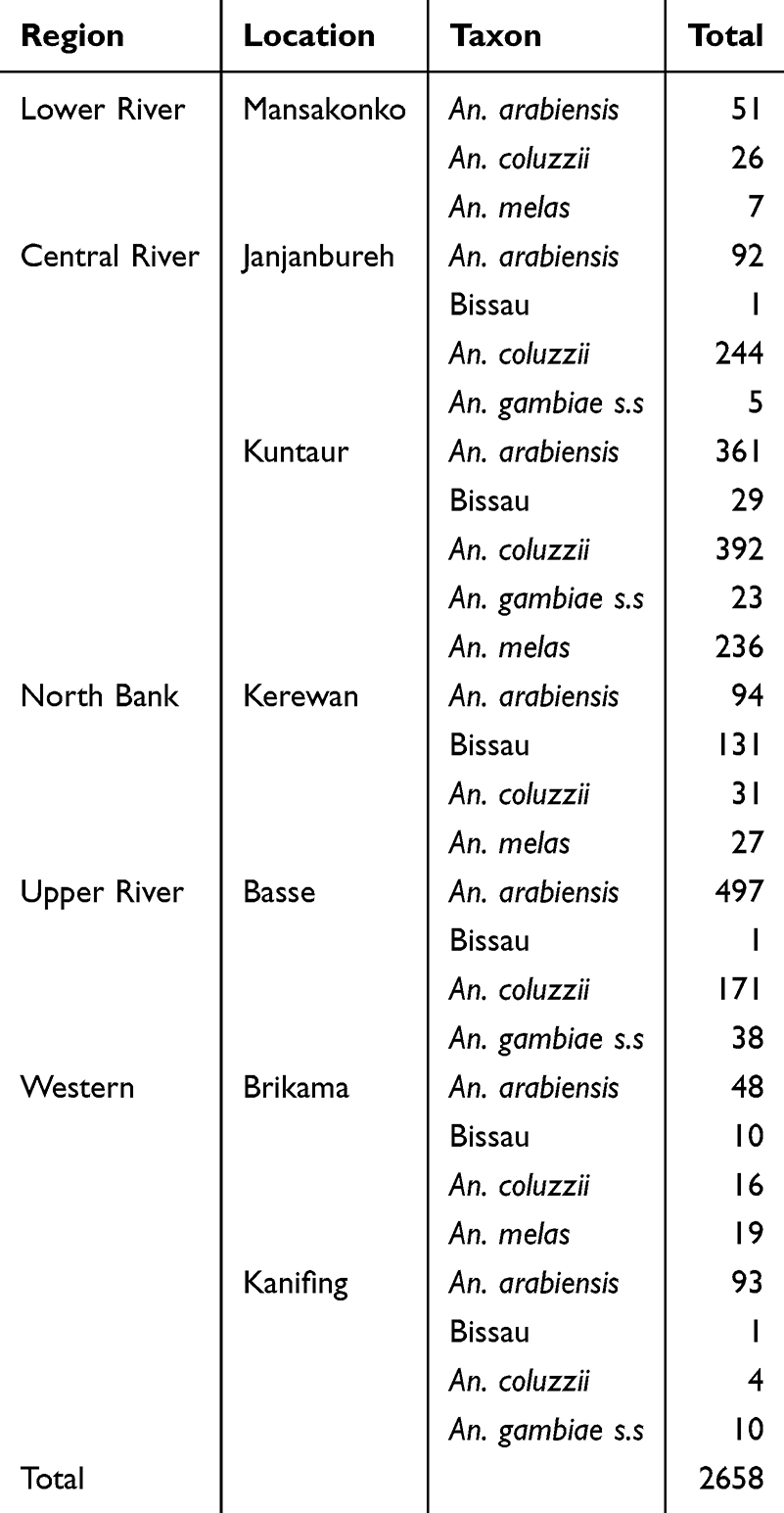 |
Table 1 Sampling Site and Number of An. gambiae Complex Mosquitoes Collected in the Gambia |
Signals of Recent Selection
To detect selective sweeps, three complementary haplotype-based statistical methods, H12,29 integrated haplotype score (iHS)30 and Cross-Population, Extended Haplotype Homozygosity (XP-EHH)31 were used.32 Each chromosome arm, ie, 2 (R&L), 3 (R&L) and X were assessed separately for signal of selection using H12. Non-overlapping windows were computed and calibrated for each taxon and cohort to account for variation in demographic history.22 Thereafter, regions with significant peaks of selective sweep were identified for each taxa and population cohort. Since the sensitivity of H12 to selective signals depends on population demographic history, which can potentially lead to false positives when detecting true sweeps, a window size was calibrated for each cohort rather than set globally. This calibration was done independently for each cohort using calibrate_h12_window_size function implemented in the malariagen_data package.33 The optimal window size was identified by plotting the distribution of H12 value across a range of candidate window sizes and selecting the value at which the 95th percentile of the H12 distribution was at or below 0.1. Following H12 computation, true signals were identified as genomic regions containing a maximum H12 value that was an outlier relative to the genome-wide distribution, accompanied by a characteristic peak architecture with roughly exponential decay on both flanks. Peak boundaries were then defined as the genomic interval over which H12 values remained continuously elevated above background on each side of the maximum. To identify selective sweeps between taxa, XP-EHH (function xpehh_gwss) from the malariagen_data python package33 was used. Additionally, the iHS statistic derived from extended haplotype homozygosity (EHH) was used to identify recent positive selection within population cohorts.31 This statistic detects incomplete selection sweeps where alleles have not reached fixation.30,34 We employed the function ihs_gwss from the malarigen_data python package.33 Candidate SNPs under selection were defined as those exceeding the 99th percentile of genome-wide iHS scores and XP-EHH, corresponding to the top 1% of outlier loci. To identify genome regions with unusual levels of haplotype sharing between population cohorts, the H1X statistic was computed using the function H1X_gwss.35 H1X detects haplotype sharing occurring at high frequencies between population cohorts, unlike traditional haplotype homozygosity statistics (ie, H1 and H12) that analyse patterns within a single population.35
Haplotype Analysis
To explore population structure and haplotype similarity, we computed pairwise haplotype distances within candidate sweep regions. Sub-sampling was done to reduce biases arising from uneven geographic representation. For An. coluzzii, up to 20 individuals were randomly selected per administrative region (admin1_iso), retaining all samples where fewer than 20 were available. All An. gambiae samples were retained. For Bisaau taxon, 50 individuals were randomly sampled from the heavily represented GM-N region, while samples from other regions (excluding GM-U) were retained. Random sampling was performed without replacement using a fixed seed for reproducibility. Haplotype distances were computed across concatenated sweep regions using the haplotype_pairwise_distances function in the malariagen_data Application Programming Interface (API). Hierarchical clustering (Ward’s method) was used to construct a condensed distance matrix and the results visualised as clustered heatmap. Furthermore, diplotype cluster analysis was performed in region 3R:31,000,000–31,080,000 coding for Odorant Receptors (OR) genes. Here, clustering of diplotypes was achieved with complete-linkage hierarchical clustering.36
Fixation Index
Genetic divergence between different cohorts of An. gambiae was assessed using Fst following the approach of Hudson et al37 Fst is a statistic that measures genetic differentiation based on allele frequency differences between populations.38
Genetic Structure at Signature of Selection
We used Principal Coordinate Analysis (PCoA) and Uniform Manifold Approximation and Projection (UMAP) to analyze genetic structure. Analyses were restricted to genomic regions under sweep selection following H12 analysis. Biallelic SNPs with a minor allele frequency (MAF) below 0.05 were retained and used to compute pairwise genetic distances between individuals (allel.pairwise_distance function from the scikit-allel Python library). The resulting city-block distance matrix was used to compute PCoA (function PCoA in scikit-bio package). UMAP (umap-learn Python package), for non-linear dimensionality reduction, was computed using the filtered biallelic sites and configured as follows: n_neighbors=15, representing the number of neighbors, balancing local and global structure, min_dist=0.5, signifying the desired separation between close points in the embedding space while preventing excessive cluster compression n_components=2, indicating the target embedding dimension for visualization and metric=“euclidean” specifying the metric used to compute the distance.39–41 This allowed fine-scale visualisation of population structure based on regions under selection. This is consistent with previously published applications of UMAP in population genomics.42
Neighbor Joining Tree
Evolutionary relationship within the An. gambiae complex was investigated using an unrooted Neighbor-Joining tree based on genomic regions under selection. The tree was built using city block (Manhattan) distance metric and visualised using function plot_njt from malariagen_data Python API.
Pairwise Ancestral Relatedness Within and Between Populations
We used the Refined Identity-by-Descent (IBD) program from Beagle software suite to examine shared ancestry and detect segments of IBD. The Refined IBD program uses probability to detect IBD segments, both short and long, based on genotype data and Hidden Markov Models.43 Briefly, 100,000 biallelic SNPs from each arm of chromosome 3 were selected and pruned for linkage disequilibrium (LD) using PLINK2. Genotype data were phased using Eagle v2.4.1, a fast and accurate phasing algorithm suited for large-scale genomic data. Refined IBD was used to detect IBD segments at a minimum length of 0.125 centiMorgans (cM) and logarithm of odds (LOD) threshold of 3. This filtering ensured accurate detection of recent shared ancestry across individuals in the dataset.
Results
Overall, our finding revealed contrasting patterns of sweeps among taxa, with loci hosting chemosensory genes being under strong selection in Bissau population compared to sibling taxa. Additionally, we observed weak genetic differentiation among taxa with gene flow occurring between Bissau and An. gambiae s.s. compared to other sibling pairs.
Genetic Relatedness Among Gambian Anopheles gambiae s.l. Populations
Identity-by-Descent (IBD) analysis on chromosome 3R revealed strong relatedness between Bissau and An. gambiae s.s. (supplementary Figure S1a and b) and partial affinity with An. coluzzii, while An. arabiensis and An. melas formed distinct genetic isolation (supplementary Figure s1c). In addition, PCoA revealed that An. arabiensis and An. melas formed distinct clusters separating from the remaining sibling species (supplementary Figure S2).
Common and Taxon-Specific Selective Signatures in Gambian Anopheles gambiae s.l. Populations
Genome-wide selection scans indicated insecticides as a major force behind adaptive evolutionary changes in the An. gambiae complex population: Selective sweeps were concentrated in regions harbouring metabolic resistance genes, including CYP450 monooxygenase (ie Cyp6aa/Cyp6p, Cyp9k) and glutathione S-Transferase (Gste) (Figure 1A). Strong signals were detected around the Voltage-gated sodium channel (Vgsc) locus on chromosome arm 2L in An. gambiae s.s (peak at 0.56) and An. coluzzii (peak at 0.28) (2–3 kb, Figure 1A). Downstream of Vgsc was a weak sweep (peak at 0.15) hosting carboxylesterase gene (Coeae2f) exclusive to An. gambiae s.s. In An. coluzzii, two prominent selective signatures (peak at respectively 0.37 and 0.31) hosting fumarylacetoacetate and E3 ubiquitin protein ligase genes were detected (26–30 kb, Figure 1A).
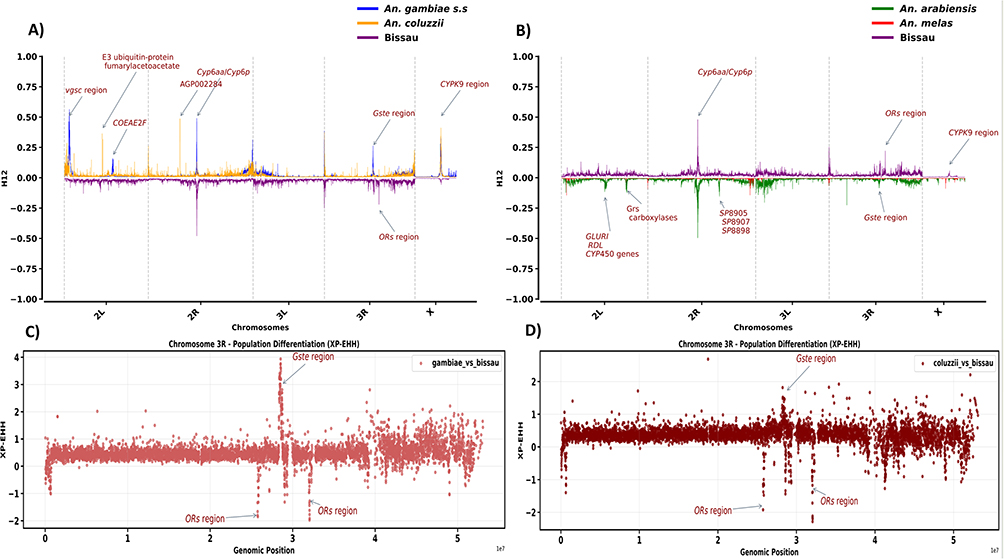 |
Figure 1 Genome-wide selection scan (GWSS) using H12 statistic (A) Bissau vs An. gambiae and An. coluzzii (B) Chromosome arm 3R shows locus hosting ORs genes as absent in sibling species An. arabiensis and An. melas. Genes of interest are highlighted with red, y-axis indicates H12 score per chromosome for different cohorts (species), x-axis indicates the chromosomal arm (C) Manhattan plot showing signature of selection as detected by XP-EHH across chromosome arm 3R which host the OR and Gste genes. The XP-EHH score represents that of Bissau in comparison to An. gambiae s.s. (D) Manhattan plot showing pairwise XP-EHH score comparison between Bissau and An. coluzzii. Y-axis indicates XP-EHH values: X-axis indicates chromosome regions. Candidate genes are detected in the top 1% of XP-EHH. Negative scores signify selection in Bissau while positive scores signify selection in sibling taxa. |
On chromosome arm 2R, selective sweeps were centred on Cyp6aa/Cyp6p gene (shared across all taxa), with peack≈0.49 for An. gambiae s.s and Bissau and weak (< 0.1) for An. coluzzii, and alpha N-acetyltransferase gene restricted to An. coluzzii at 0.23 peak (Figure 1A). On chromosome arm X, the signal for Cyp9k1 (15 kb) overlapped with a cuticular protein gene (CPR125) (0.34, 0.57 respectively for An. gambiae s.s and An. coluzzii), suggesting combined metabolic and cuticular resistance mechanisms (Figure 1A). Whilst the selective sweeps were generally weak on chromosome arm 3L across all taxa (peak < 0.15) (Figure 1A), a clear signal at Gste (28 kb) was evident on chromosome arm 3R in An. gambiae s.s. (peak 0.26) and Bissau (peak 0.14), and the odorant receptor (OR) signal further downstream, restricted to Bissau only (peak 0.22) (Figure 1A). Interestingly, extending H12 analysis of sibling species (An. arabiensis and An. melas) failed to detect ORs sweep signal, though Gste and CYP450 signals were evident (Figure 1B) from further sweep selection analysis. Consequently, these two species were excluded.
Pairwise XP-EHH scans of chromosome arm 3R revealed distinct selective pressure driving genetic divergence amongst taxa. A strong signal of selection was consistently detected in the Gste genes cluster (28.5kb) in both An. gambiae s.s (XP-EHH score >2) and An. coluzzii (XP-EHH score < 2), indicative of recent insecticide-driven selective sweeps (Figure 1C and D). However, in Bissau population, the selection was most evident at loci associated with sensory perception, specifically olfactory receptor regions (ORs, 32 kb; OR29, 25.74 kb), suggesting species-specific local adaptation which could be related to environment or host-seeking behaviour (Figure 1C and D).
iHS Analysis Identified Candidate Genes Under Selection in Insecticide Detoxification, Immune and Sensory Reception
The iHS analysis performed on An. gambiae complex genome identified 159 genes under recent selection (see supplementary Table S1). These genes spanned multiple functional pathways, including sensory reception, immune response and detoxification of xenobiotics (insecticides), consistent with ongoing local adaptation in a heterogeneous environment (Figure 2). SNPs associated with insecticide resistance showed consistently elevated iHS scores (>5), highlighting persistent selective pressure from either vector control interventions or agricultural pesticides (Figure 2).
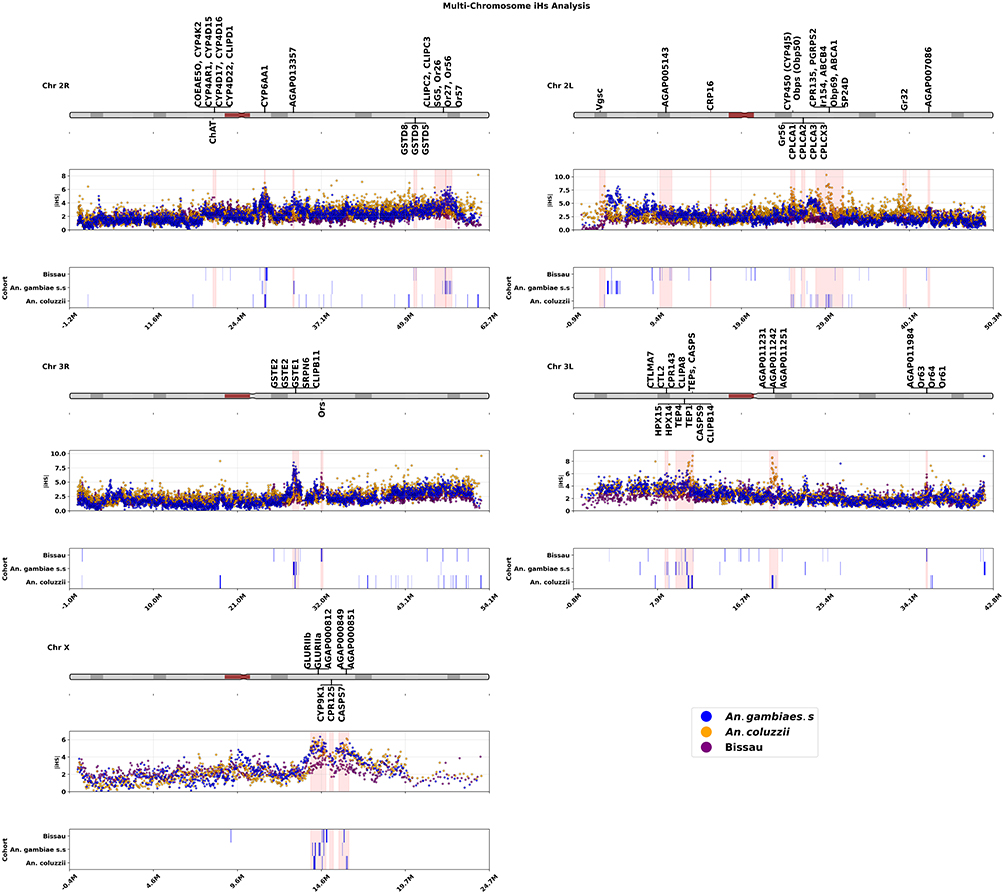 |
Figure 2 Chromosomal distribution maps and integrated haplotype scores (iHS) showing loci under selection in An. gambiae sibling species defined by specific colours. Each panel shows a chromosomal arm (labelled top left of each panel). The rows in each panel represent the karyotype (top row), iHS values for each SNP plotted on y-axis against the SNP position on the x-axis). The bottom row marks the regions with elevated iHS, with the gene labels shown on the kayotype in the top row. |
Selection Signatures of Genes Involved in Insecticide Metabolic and Cuticular Resistance Pathway
Genome scan of An. gambiae species genomes with iHS, much like H12, did not detect selection at the Vgsc gene in chromosome arm 2L within the Bissau population (Figure 2 and supplementary Table S1). Nonetheless, strong selection signals were evident across multiple metabolic resistance genes, particularly the CYP450 family on multiple chromosomal arms. For example, in An. coluzzii, selection targeted Cyp4 genes on chromosome arms 2L (Cyp4J5, Cyp4J9, Cyp4J10) and 2R (Cyp4K2, Cyp4AR1, Cyp4D15, Cyp4D17, Cyp4D16, Cyp4D22) whilst in Bissau, Cyp4C genes (Cyp4C35, Cyp4C36, Cyp4C27) were under selection on chromosome arm 3R. Similarly, Cyp6 family genes (Cyp6AA1-2, Cyp6P1-5, Cyp6P15, Cyp6AD1), known mediators of pyrethroid resistance, were consistently under selection in all sibling taxa on chromosome 2R. The Gste cluster genes (Gste1-8), conferring resistance to DDT and pyrethroids, were under selection on chromosome arm 3R of An. gambiae s.s and An. coluzzii (Figure 2 and supplementary Table S1). Beyond CYP450 and Gste, selection targeted additional unique detoxification loci. In Bissau population, Choline acetyltransferase gene (ChAT, 20 kb) on chromosome arm 2R, a rarely studied resistance candidate compared to well-established Ace-1 gene associated with resistance to carbamates and organophosphates, showed strong selection.
Carboxylesterases were also implicated: Coeae5O on 2R was under selection in both Bissau and An. coluzzii, while Coeae2F, associated with organophosphate resistance, was selected in An. gambiae s.s. (Figure 3 and supplementary Table S1). Strikingly, candidate regions extended beyond classical detoxification pathways. The genes linked to cuticle formation, including CPR16 (2L, 15 kb) and CPLC (CPLCA1–3, CPLCX3, 26–27 kb) in Bissau, CPR135 (2L, 28 kb) and CPR144 (2L, 29 kb) in An. coluzzii, and CPR143 (on 3L) in An. gambiae s.s., were also under selection (Figure 3 and supplementary Table S1). Additionally, ABC (ATP-binding cassette) transporter family genes, associated with xenobiotic detoxification, were enriched in An. coluzzii (2L), including ABCA1, ABCA2, ABCB4, and ABCC7 (Figure 2 and supplementary Table S1).
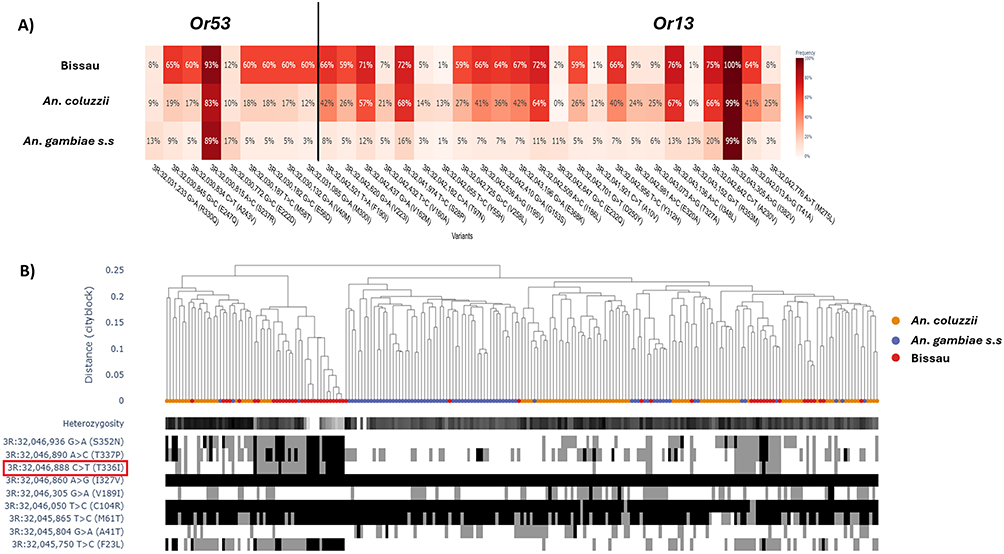 |
Figure 3 Allele frequencies and haplotype diversity across OR genes in Bissau. (A) Heat map of allele frequencies for non-synonymous mutations in OR53 and OR13. (B) Diplotype clustering at OR15 showing a unique allele (C>T: T336I) in Bissau. Hierarchical clustering dendrogram aligned with observed heterozygosity, displayed as a horizontal bar. The leaves of the dendrogram are colored by the species of the individual. Substitutions in each individual are presented below the dendrogram with blocks of the same color indicating shared mutation. |
Selection Signatures of Genes Involved in Immune Response Pathway
Besides insecticide-driven adaptation, iHS analysis revealed strong selection on immune-related genes across An. gambiae complex, highlighting immunity as an additional driver of ongoing evolution. The chromosomal distribution of genes under selection of these genes also varied among taxa. In Bissau, the selection targeted G-protein-coupled receptor neuropeptide (GPRNP1) on chromosome arm 2R, CLIPB14 on chromosome arm 3L, and glutathione peroxidase 2 (GPXH2) on chromosome X. These loci (GPRNP1 and GPXH2) were under selection in other taxa (Figure 2 and supplementary Table S1). In An. coluzzii, the selection signatures included CLIP-domain serine protease on chromosome 2R, serine protease (SP24D) on chromosome 2L, serine protease inhibitor (SRPN4) on chromosome arm 3R, peptidoglycan recognition proteins (PGRPs 2–3), TEPs genes (TEP1, TEP3, TEP6, TEP10, TEP11 and TEP18), short caspase (CASPS9-13), and fibrinogen on chromosome 3L; (Figure 2 and supplementary Table S1). In An. gambiae s.s., the selection of immune-related genes was more widespread, involving the clusters of C-type lectins (CTLMA7 and CTL2), HPX genes (HPX1, HPX14, HPX15), CLIPA8, and TEP4 on chromosome arm 3L (Figure 2). Additional selection peaks on 2R included CLIPC2–3 and salivary gland proteins, while clusters of CLIPB genes (CLIPB11–12, CLIPB18), Mitogen-activated protein kinase, and serine protease inhibitors (SRPN5–6, SRPN16) were detected on chromosome arm 3R (28–29 kb) (Figure 2 and supplementary Table S1).
Selection Signatures of Genes Involved in Sensory Receptor Pathway
iHS analysis revealed selection signatures on genes associated with gustation, olfaction and host seeking, highlighting a behavioural adaptation to environmental chemical cues. In Bissau, the candidate loci selected included gustatory receptor 56 (Gr56) on chromosome arm 2L (26–27 kb), a cluster of olfactory receptor (ORs) genes (OR60-61, OR63-64) on chromosome arm 3L and multiple ORs and odorant-binding proteins (Obp43, Or13, Or15, Or17, Or30, Or46, Or53, Or55) on chromosome 3R (31–32 kb) (Figure 2 and supplementary Table S1). In An. coluzzii, sweep selection signals were detected at gustatory receptor 32 (GR32) and a cluster of odorant binding proteins (Obp71, Obp49-55 region 39 kb) and Obp 69 (29–31 kb) on chromosome arm 2L (Figure 2 and supplementary Table S1). In An. gambiae s.s, the chemoreceptor genes (Or26-27, Or56-57, Obp19) and ionotropic receptors (IR41t.1–2), were under selection (Figure 2 and supplementary Table S1). Across taxa, selection sweeps were also detected in glutamate receptors (GLURIIa, GLURIIb) on the X chromosome, with GLURIIc additionally under selection in An. gambiae s.s. and An. coluzzii (Figure 2 and supplementary Table S1). The divergent but overlapping patterns of selection on chemosensory pathways reflect their role in ecological adaptation and host-seeking behavior in Anopheles gambiae s.l.
Whilst selective sweeps spanned multiple chemosensory gene families (ie IR, OR, GR and OBP), OR genes emerged as the most divergent with Bissau populations carrying distinct, high-frequency variants that were rare or absent in sibling taxa (supplementary Figure S3). For example, amino acid changes were more abundant in ORs 53 and 13 in the Bissau population than in sibling taxa (Figure 3A). These were particularly distinct as most variants had allele frequencies >60 in Bissau, contrary to very low frequencies in sibling taxa, except for the S237R and I382V variant that were nearing fixation in all taxa (Figure 3A). These OR mutations occurring at high frequency have not been validated for association with any phenotype. Diplotype clustering revealed a region with low heterozygozity within the Bissau population, suggesting positive selection (Figure 3B). Besides, distinct patterns of diplotype clustering was observed for other ORs across sibling taxa suggesting divergent adaptation mechanisms (supplementary Figure S3).
Adaptive Gene Flow and Parallel Evolution of Insecticide Resistance Reveal Closer Relatedness Between Bissau and An. gambiae s.s.
Haplotype cluster analysis revealed asymmetric adaptive gene flow across An. gambiae complex with the strongest haplotype sharing occurring between An. gambiae s.s. and Bissau than any other pair of sibling taxa (Supplementary Figure S4). Genome-wide clustering of sweep regions identified three major haplotype groups, one of which was mainly shared between Bissau and An. gambiae s.s. (Supplementary Figure S4a). Consistently, pairwise Fst analysis showed Bissau to be more genetically differentiated from An. coluzzii (Fst 0.168) than An. gambiae s.s. (Fst 0.124) (supplementary Figure s4b). Additionally, UMAP clustering grouped other sibling taxa distinctly with Bissau and An. gambiae s.s apart from the other sibling taxa (Supplementary Figure S4c).
At insecticide metabolic resistance loci, adaptive sharing was also most evident between Bissau and An. gambiae s.s. Haplotype cluster analysis at Cyp9K1 (15kb, chromosome X) revealed a haplotype shared between the two taxa, alongside a haplotype common to all taxa (Supplementary Figure S4d). Similarly, at the CYP450 (chromosome 2R, 28.8 kb) and Gste (chromosome 3R, 28 kb) metabolic resistance clusters, Bissau shared haplotypes with An. gambiae s.s. but not with An. coluzzii (supplementary Figure S4e and Figure S5).
Within the Bissau population, haplotype cluster analysis of the olfactory receptor genes region (chromosome 3R, 32 kb) revealed an isolated haplotype cluster in the North Bank region (Figure 4A). While gene flow was evident amongst Bissau population cohorts (Figure 4A), low-level genetic differentiation was observed in the North Bank population (Fst, 0.001 and 0.012) relative to the other sampling locations (Figure 4B). PCoA and Neighbor-joining analysis confirmed this sub-clustering, identifying the North_Bank population as a relatively separate cluster within the Bissau cohorts (Figure 4C and D). Additionally, haplotype clustering that focused solely on geographical distribution, revealed Bissau to be largely distributed in the North (Figure 4E).
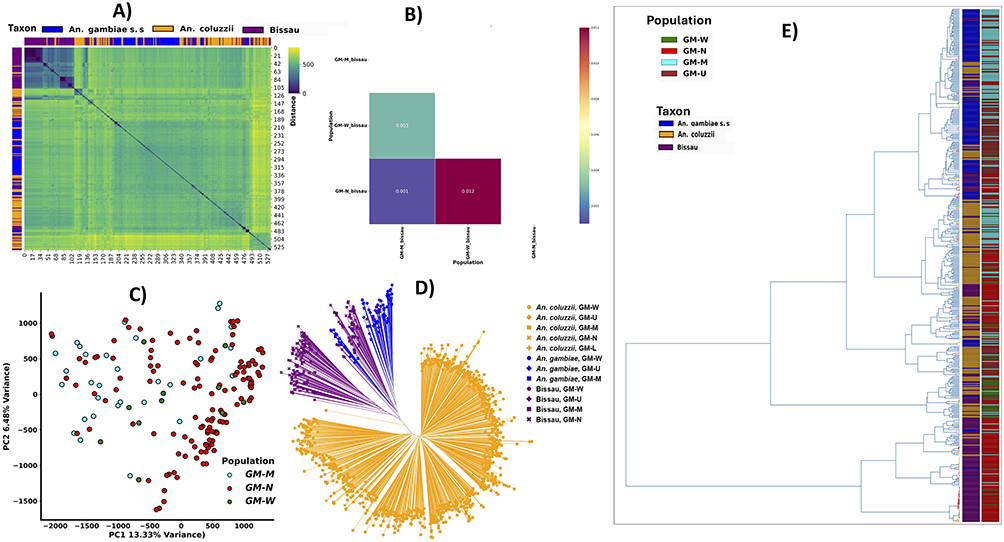 |
Figure 4 (A) Haplotype clustering within a region containing OR genes. Individuals are ordered by sampling regions and dark blue colour indicate haplotype that are shared between populations (B) Fst analysis showing GM-N (North-bank) cohorts of Bissau population displaying some level of genetic differentiation (C) PCoA showing low genetic variation between An. gambiae lineages (D) NJ tree showing relationships across all cohort and taxa (E) Clustering of haplotypes by geographical origin. |
Discussion
We analyzed An. gambiae complex genome from The Gambia for signatures of selective sweep to gain insights into their adaptive responses to selection pressures. There were strong selective sweeps, mainly associated with resistance to insecticide, vector competence and mosquito behavioral adaptation. The selection signatures of resistance encompassed well-known mechanisms including target, cuticular and detoxification enzymes. Interestingly, the patterns of selection varied among species whereas spatial variation was limited. This somehow reflects vector control strategies in The Gambia where insecticides are applied across the country than varying across regions. In contrast, the observed variation in adaptation to insecticides among species could be attributed to behavioral adaptation where species display temporal divergence in breeding peak and therefore a different impact of insecticides.
Detection of selective sweeps associated with metabolic resistance is consistent with other previous studies. For example, we detected several Cyp6 family8,44–46 and Cyp4 family (ie Cyp4AR1, Cyp4D15, Cyp4D17, and Cyp4J5) aligning with recent findings from the Sahel region.47 Another consistent finding was the detection of Cyp9K1 gene, which was recently linked to type I and II pyrethroids resistance in mosquitoes from East and Central Africa.48,49 These further highlight Cyp9K1 as a potential driver of insecticide resistance in An. gambiae s.l across the continent. Interestingly, haplotype clustering analysis supported introgression rather than independent local evolution as a potential factor underlying CYP9K1 haplotype evolution among sibling taxa. Other metabolic resistance genes under selection among An. gambiae s.l and reported in several malaria endemic countries across the African continent are Gste18,49–52 and carboxylesterase (Coeae2F) genes.53 As is for Cyp9k1, haplotype clustering constructed on genomic regions that host Gste revealed adaptive sharing between taxa pairs, illuminating the potential of metabolic resistance genes spreading across sibling taxa through adaptive sharing. We note, however, that haplotypes sharing between sibling taxa within An. gambiae s.l complex at loci associated with insecticide resistance may also reflect incomplete lineage sorting (ILS) or more plausible parallel evolution, given their close evolutionary relationship, calling for a more detailed analyses across species and the continent.
Another notable finding was the detection of selective sweeps associated with individual species, further suggesting species-specific response to selective pressure. For example, sweeps that have not previously been associated with insecticide resistance were evident for An. coluzzii. The unique sweeps were detected at genomic regions around alpha N-acetyltransferase (AGAP002284) and E3 ubiquitin-protein ligase. Both genes have been detected in other mosquito populations, although their role in insecticide resistance remains largely unexplored.54,55 However, alpha N-acetyltransferase has been implicated in detoxification of xenobiotics including agricultural pollutants some of which share a similar chemical class with some insecticides used in vector control.56 Currently, An. coluzzii is among the most geographically widespread malaria vectors in the Gambia and could be in contact with some pesticides (eg neonicotinoids) used in agriculture, which may be driving such selection. Some of these are also commonly incorporated as synergists to increase toxicity against pyrethroid-resistant mosquitoes.57 Possibly, these novel mutations could be allowing An. coluzzii to be more adapted to insecticide pressure, enabling its spatial spread across different ecologies. Indeed, resistance against neonicotinoids insecticides was recently detected in An. gambiae population from Cameroon following phenotypic bioassay.58 Nonetheless, the role of these novel loci under selection in An. coluzzii should be functionally confirmed.
The selective signature in Bissau around the Choline acetyltransferase gene (ChAT) on chromosome arm 2R was unexpected as it is rather the well-known acetylcholinesterase (Ace-1) resistance gene that was previously found under selection, a common occurrence in parts of West Africa.53 Both transferase and esterase enzymes are involved in the metabolism of acetylcholine (Ach) and it is possible that the higher frequency of ChAT associated alleles is being driven by selection pressures from sustained insecticide application and a favorable fitness cost. To preserve the effectiveness of available insecticides, we suggest further monitoring of this novel variant alongside other conventional insecticide resistance markers. Phenotypic analysis to help identify the class of insecticides driving selection at this locus is also important.
The genomic region around the ATP-Binding Cassette (ABC) genes being under selection was another noticeable finding in An. coluzzii. Although the connection between ABC transporters family and insecticides resistance is not well established for mosquitoes, they have been linked to pyrethroid resistance in some agricultural pests including Plutella xylostella.59–61 Recently, ABCs were found to be highly expressed in a deltamenthrin-resistant strain of Anopheles sinensis, a primary malaria vector in China and southeast Asia.62 One of the mechanisms by which ABC transporters are thought to modulate resistance against deltamenthrin is through alteration of cuticle thickness, thus reducing insecticide penetration.63 We detected selective sweeps spanning several cuticular proteins, including CPR125, commonly associated with rigid cuticle, potentially lending credence to cuticle thickness playing a role in enhancing insecticide resistance in An. gambiae s.l. Cuticle thickening mechanism of insecticide resistance and its confluence with immune response was further supported by evidence of selection of CLIP gene sub-family, an immune response gene previously linked to resistance phenotype.64
For An. gambiae s.s., currently decreasing in The Gambia, there were multiple strong sweeps under selection compared to sibling taxa. Concerningly, the sweeps under selection code for genes linked to both target site (Vgsc) and metabolic insecticide resistance (Cyp6aa/Cyp6p, Cyp9k1 and Gste). This could be in response to intensive insecticidal interventions against malaria over the last couple of decades. If these variants are selected directionally to fixation, they may enable resistance against different classes of insecticides. The outcome could be An. gambiae s.s expansion and re-establishment across The Gambia, thereby undermining malaria elimination efforts.65
Besides resistance, our sweep analysis detected immune-related alleles that are known to shape mosquito-plasmodium interaction in An. gambiae s.s. An example is HPX15 gene commonly associated with defence against pathogens and long-term fertility.66 Additionally, under selection in An. gambiae s.s. was the Mitogen-activated protein kinase (AGAP029511) gene that plays a key role in P. falciparum male gametogenesis, also exflagellation, an important step for parasite development in mosquito mid-gut.67 Several genes of the TEPs family, key components of the innate immune systems of mosquitoes, were under selection including TEP-1 that is commonly associated with malaria parasite clearance and works alongside CLIP serine protease.65,67,68
For the recently described Bissau population, distinct and shared selective signatures were also revealed. For example, like An. coluzzii it had ChAT under selection but lacked signals at regions coding for target site genes, corroborating previous findings by Caputo et al25 Comparable observations have been reported for Anopheles funestus where metabolic genes and not target site are frequently implicated in insecticide resistance.69–71 Broadly, target site resistance is expected to succeed metabolic resistance as non-insecticide xenobiotics can exert selection pressure on metabolic genes well before mosquitoes encounter insecticides used for malaria control. An alternative explanation is that mutations conferring metabolic resistance are generally suggested to have lower fitness cost than target site alterations, thus favoring their fixation. However, the resistant-associated haplotypes of Vgsc genes can still spread to Bissau through adaptive introgression as reflected in shared haplotype across the three taxa in chromosome arm 2L.
As seen for the primary malaria vectors (An. gambiae s.s. and An. coluzzii), the CLIP serine protease (CLIP14) was under selection in Bissau. CLIP14 is associated with melanization response against Plasmodium, and this finding could suggest Bissau is involved in malaria transmission. The ecological interaction of Bissau with the human host was further suggested by the selective sweep signals in the region coding for odorant receptors such as OR30, an indole-sensitive odorant receptor in some mosquito species and Drosophila melanogaster.72 Besides, in the common fly Musca domestica, both ORs 30 and 46 are categorized as indole-sensitive receptors.73 Indole serves dual role, in some mosquito’s species being an oviposition cue while in others inhibiting host attraction.74 The OR53 associated with host-seeking in mosquitoes was found to have multiple non-synonymous mutations occurring at high frequency, possibly driven by an ongoing change in the behavior of Bissau.75,76 Similarly, OR13 had appreciable frequencies of nonsynonymous substitutions, suggesting that it is under a strong positive selection in Bissau compared to other sibling species. As the alleles were not shared between Bissau and other sibling species, the selection may be an independent evolutionary event in this taxon.
A possible reason for OR evolution in Bissau could be the need to adapt and establish an ecological niche. For Anopheles mosquitoes, alleles associated with sensory perception are generally less studied. Yet local adaptation involving sensory genes may modify sensory perception and subsequently behavior. In Drosophila melanogaster (Fruit fly) for example, the OR13 is linked to pheromone detection although it is unclear whether mosquitoes retain comparable functions.77 Nonetheless, comprehensive understanding of OR genes in Anopheles mosquitoes is needed as they can serve as targets for vector control strategies, through the disruption of important biological traits such as oviposition or host attraction.
The OR sweep signatures were detected across spatially widespread Bissau cohorts. Whilst some level of geographical segregation was evident among Bissau cohorts, the similar OR sweep patterns across all sub clusters was surprising. A similar sweep pattern was observed for loci involved in metabolic resistance (cyp6a/p and Gste), where no geographic location emerged as a resistance hotspot in The Gambia, although the Gambian river is expected to impose a strong barrier to gene flow between its banks. This may be due to the small size of The Gambia, less than 50 Km at its widest point and around 480 km extending inland, and recent studies show mosquitoes to be capable of long-distance migration.78 With reference to vector control, these findings imply that for small countries like The Gambia resistance management may include using the same insecticides over a wide geographical scale and that insecticide rotation should follow periodic/temporal rather than spatial approach.
Interestingly, we failed to detect selection at loci commonly associated with response to climate conditions such as heat shock proteins, implying that adaptation to insecticide resistance more than climate change could be shaping evolution of emerging species. Overall, these results illuminate further the genetic landscape of An. gambiae s.l in The Gambia and reveal the need to continue genomic surveillance to understand and address the changing landscape of insecticide resistance in the region. Nonetheless, phenotypic analysis to further substantiate the genomic analysis are needed. A possible limitation on the interpretation of our findings, especially on gene flow barriers, could be the sampling scheme that was based on administrative regions rather than ecotypes.
Conclusions
This study constitutes a comprehensive scan of patterns of sweep selection and gene flow encompassing insecticide and non-insecticide resistance genes. The key findings are that An. gambiae sibling species in The Gambia display varying insecticide resistance mechanisms, with some resistance genes being under selection in one taxon while undetectable in others. Besides, the study validates the presence of key insecticide resistance gene previously detected in this region and reveals the likelihood of emergence of novel insecticide resistance markers. Indeed, phenotypic association and experimental functional validation are needed to support the association of genetic variants in these signatures of selection in adaptive process such as insecticide resistance. Selection of genes associated with sensory perception in the novel taxon, Bissau, as well as those implicated in insecticide resistance, immune system and parasite development especially CLIP14 commonly associated with melanization response against Plasmodium, calls for in-depth bionomic investigations of this species and its role in parasite-vector interactions and malaria transmission dynamics in the far west Africa. This work represents a major step towards advancing our understanding of the genomic structure and evolution of emerging malaria vectors, with the potential of informing vector control efforts in The Gambia.
Abbreviations
ABC, ATP-Binding Cassette; ACE-1, acetylcholinesterase; Ach, acetylcholine; ChAT, choline acetyltransferase; CNVs, copy number variants; GR, gustatory receptor; iHS, integrated haplotype scores; MAF, minor allele frequency; OR, odorant receptor; PCoA, principal coordinate analysis; UMAP, Uniform Manifold Approximation and Projection; XP-EHH, cross-Population, Extended Haplotype Homozygosity.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author AAN upon reasonable request.
Acknowledgments
We acknowledge all the members of the Vector Biology team at MRCG at LSHTM and The National Malaria Control Program, The Gambia, for facilitating sampling. We thank the staff of the Welcome Trust Sanger Institute Sample Logistics, Sequencing and Informatics facilities for their contributions. Genomic sequences were freely accessed online and through long-standing collaboration with MalariaGEN.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research reported in this publication was funded by a National Institute of Health and Care Research (NIHR) under award No: NIHR134717; The H3Africa Pan African Malaria Genomic Epidemiology Network (PAMGEN) funded by the Science for Africa Foundation (SFA) grant (H3A/18/002). The content expressed by the authors does not necessarily represent the official views of the NIHR or SFA.
Disclosure
Umberto D’Alessandro reports Grants or contracts from MRC UKRI; Data safety/advisory board participation from Medicine for Malaria Venture, Serum Institute of India, outside the submitted work. Alfred Amambua-Ngwa reports Support for the manuscript from MalariaGEN Vector Observatory, GSM3, NIHR research group, PAMGEN-H3 Africa, during the conduct of the study, outside the submitted work. The authors declare that they have no other financial and non-financial competing interests.
References
1. Takken W, Charlwood D, Lindsay SW. The behaviour of adult Anopheles gambiae, sub-Saharan Africa’s principal malaria vector, and its relevance to malaria control: a review. Malar J. 2024;23(1):161. doi:10.1186/s12936-024-04982-3
2. Loughlin SO. The expanding Anopheles gambiae species complex. Pathog Glob Health. 2020;114(1):1. doi:10.1080/20477724.2020.1722434
3. Coetzee M, Hunt RH, Wilkerson R, et al. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa. 2013;3619(3). doi:10.11646/zootaxa.3619.3.2.
4. Barrón MG, Paupy C, Rahola N, et al. A new species in the major malaria vector complex sheds light on reticulated species evolution. Sci Rep. 2019;9(1):14753. doi:10.1038/s41598-019-49065-5
5. Rehmann CT, Ralph PL, Kern AD. Evaluating evidence for co-geography in the Anopheles–Plasmodium host–parasite system. G3 GenesGenomesGenetics. 2024;14(3):1.
6. Booker TR. The structure of the environment influences the patterns and genetics of local adaptation. Evol Lett. 2024;8(6):787–16. doi:10.1093/evlett/qrae033
7. Morjan CL, Rieseberg LH. How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles. Mol Ecol. 2004;13(6):1341–1356. doi:10.1111/j.1365-294X.2004.02164.x
8. Weedall GD, Riveron JM, Hearn J, et al. An Africa-wide genomic evolution of insecticide resistance in the malaria vector Anopheles funestus involves selective sweeps, copy number variations, gene conversion and transposons. PLOS Genet. 2020;16(6):e1008822. doi:10.1371/journal.pgen.1008822
9. Wagah MG, Korlević P, Clarkson C, et al. Genetic variation at the Cyp6m2 putative insecticide resistance locus in Anopheles gambiae and Anopheles coluzzii. Malar J. 2021;20(1):234. doi:10.1186/s12936-021-03757-4
10. Nagi SC, Lucas ER, Egyir-Yawson A, et al. Parallel evolution in mosquito vectors—a duplicated esterase locus is associated with resistance to pirimiphos-methyl in Anopheles gambiae. Mol Biol Evol. 2024;41(7). doi:10.1093/molbev/msae140.
11. Zouré AA, Badolo A, Francis F. Resistance to insecticides in Anopheles gambiae complex in West Africa: a review of the current situation and the perspectives for malaria control. Int J Trop Insect Sci. 2021;41(1):1–13. doi:10.1007/s42690-020-00173-0
12. Jones CM, Liyanapathirana M, Agossa FR, et al. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc Natl Acad Sci U S A. 2012;109(17):6614–6619. doi:10.1073/pnas.1201475109
13. Wondji CS, Dabire RK, Tukur Z, Irving H, Djouaka R, Morgan JC. Identification and distribution of a GABA receptor mutation conferring dieldrin resistance in the malaria vector Anopheles funestus in Africa. Insect Biochem Mol Biol. 2011;41(7):484–491. doi:10.1016/j.ibmb.2011.03.012
14. Badolo A, Bando H, Traoré A, et al. Detection of G119S ace-1 R mutation in field-collected Anopheles gambiae mosquitoes using allele-specific loop-mediated isothermal amplification (AS-LAMP) method. Malar J. 2015;14. doi:10.1186/s12936-015-0968-9
15. Elanga-Ndille E, Nouage L, Ndo C, et al. The G119S Acetylcholinesterase (Ace-1) target site mutation confers carbamate resistance in the major malaria vector Anopheles gambiae from Cameroon: a challenge for the coming IRS implementation. Genes. 2019;10(10):790. doi:10.3390/genes10100790
16. Djouaka RF, Bakare AA, Coulibaly ON, et al. Expression of the cytochrome P450s, CYP6P3 and CYP6M2 are significantly elevated in multiple pyrethroid resistant populations of Anopheles gambiae s.s. from Southern Benin and Nigeria. BMC Genomics. 2008;9:538. doi:10.1186/1471-2164-9-538
17. Fotso-Toguem Y, Tene-Fossog B, Mugenzi LMJ, et al. Genetic diversity of cytochrome P450s CYP6M2 and CYP6P4 associated with pyrethroid resistance in the major malaria vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon. Genes. 2022;14(1):52. doi:10.3390/genes14010052
18. Riveron JM, Yunta C, Ibrahim SS, et al. A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Genome Biol. 2014;15(2):R27. doi:10.1186/gb-2014-15-2-r27
19. Mitchell SN, Rigden DJ, Dowd AJ, et al. Metabolic and target-site mechanisms combine to confer strong DDT Resistance in Anopheles gambiae. PLoS One. 2014;9(3):e92662. doi:10.1371/journal.pone.0092662
20. Barron MG, Paupy C, Rahola N, et al. A new species in the Anopheles gambiae complex reveals new evolutionary relationships between vector and non-vector species. Evolutionary Biology. Sci Rep. 2019;9(1):14753.
21. Clarkson CS, Weetman D, Essandoh J, et al. Adaptive introgression between Anopheles sibling species eliminates a major genomic island but not reproductive isolation. Nat Commun. 2014;5:4248. doi:10.1038/ncomms5248
22. Norris LC, Main BJ, Lee Y, et al. Adaptive introgression in an African malaria mosquito coincident with the increased usage of insecticide-treated bed nets. Proc Natl Acad Sci U S A. 2015;112(3):815–820. doi:10.1073/pnas.1418892112
23. Adeogun AO, Adetunji O, Babalola AS, et al. Spatial eco-climatic gradients drive adaptive 2La chromosomal inversion polymorphism in Anopheles coluzzii, a major malaria vector in Nigeria. Int J Health Geogr. 2026;25(1):31. doi:10.1186/s12942-026-00465-7
24. Ayala D, Acevedo P, Pombi M, et al. Chromosome inversions and ecological plasticity in the main African malaria mosquitoes. Evol Int J Org Evol. 2017;71(3):686–701. doi:10.1111/evo.13176
25. Caputo B, Marco CD, Pichler V, et al. Speciation within the Anopheles gambiae complex: high-throughput whole genome sequencing reveals evidence of a putative new cryptic taxon in ‘far-west’ Africa. Res Sq. 2024:
26. Abdoulaye S, Milugo TK, Oriero, et al. Genetic divergence and lower frequencies of insecticide resistance markers in the novel Anopheles gambiae Bissau molecular form in The Gambia. Sci Rep. 2026;16:5540. doi:10.1038/s41598-026-35295-x
27. Willing EM, Dreyer C, van OC. Estimates of genetic differentiation measured by FST do not necessarily require large sample sizes when using many SNP markers. PLoS One. 2012;7(8):e42649. doi:10.1371/journal.pone.0042649
28. Ferrer-Admetlla A, Liang M, Korneliussen T, Nielsen R. On detecting incomplete soft or hard selective sweeps using haplotype structure. Mol Biol Evol. 2014;31(5):1275–1291. doi:10.1093/molbev/msu077
29. Garud NR, Rosenberg NA. Enhancing the mathematical properties of new haplotype homozygosity statistics for the detection of selective sweeps. Theor Popul Biol. 2015;102:94–101. doi:10.1016/j.tpb.2015.04.001
30. Voight BF, Kudaravalli S, Wen X, Pritchard JK. A map of recent positive selection in the human genome. PLOS Biol. 2006;4(3):e72. doi:10.1371/journal.pbio.0040072
31. Sabeti PC, Reich DE, Higgins JM, et al. Detecting recent positive selection in the human genome from haplotype structure. Nature. 2002;419(6909):832–837. doi:10.1038/nature01140
32. Moss S, Pretorius E, Ceesay S, et al. Genomic surveillance of Anopheles mosquitoes on the Bijagós Archipelago using custom targeted amplicon sequencing identifies mutations associated with insecticide resistance. Parasit Vectors. 2024;17(1):10. doi:10.1186/s13071-023-06085-5
33. Jon BL, Alistair M, Md AR, et al. (2026). malariagen/malariagen-data-python: v15.8.0.
34. Grossman SR, Andersen KG, Shlyakhter I, et al. Identifying recent adaptations in large-scale genomic data. Cell. 2013;152(4):703–713. doi:10.1016/j.cell.2013.01.035
35. Mwinyi SH, Bennett KL, Nagi SC, et al. Genomic analysis reveals a new cryptic taxon within the Anopheles gambiae complex with a distinct insecticide resistance profile in the coast of East Africa. Mol Ecol. 2025;34(10):e17762. doi:10.1111/mec.17762
36. Nagi SC, Lucas ER, Egyir-Yawson A, et al. Parallel evolution in mosquito vectors – a duplicated esterase locus is associated with resistance to pirimiphos-methyl in An. gambiae. Evolutionary Biology. 2024;2024:1.
37. Hudson RR, Slatkin M, Maddison WP. Estimation of levels of gene flow from DNA sequence data. Genetics. 1992;132(2):583–589. doi:10.1093/genetics/132.2.583
38. Bhatia G, Patterson N, Sankararaman S, Price AL. Estimating and interpreting FST: the impact of rare variants. Genome Res. 2013;23(9):1514–1521. doi:10.1101/gr.154831.113
39. McInnes L, Healy J, Melville J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. (arXiv:1802.03426). arXiv(2020).
40. Diaz-Papkovich A, Anderson-Trocmé L, Gravel S. A review of UMAP in population genetics. J Hum Genet. 2021;66:85–91. doi:10.1038/s10038-020-00851-4
41. Diaz-Papkovich A, Anderson-Trocmé L, Ben-Eghan C, Gravel S. UMAP reveals cryptic population structure and phenotype heterogeneity in large genomic cohorts. PLoS Genet. 2019;15(11):e1008432. doi:10.1371/journal.pgen.1008432
42. Browning BL, Browning SR. Detecting identity by descent and estimating genotype error rates in sequence data. Am J Hum Genet. 2013;93(5):840–851. doi:10.1016/j.ajhg.2013.09.014
43. Ibrahim SS, Amvongo-Adjia N, Wondji MJ, Irving H, Riveron JM, Wondji CS. Pyrethroid resistance in the major malaria vector anopheles funestus is exacerbated by overexpression and overactivity of the P450 CYP6AA1 Across Africa. Genes. 2018;9(3):140. doi:10.3390/genes9030140
44. Kusimo MO, Mackenzie-Impoinvil L, Ibrahim SS, et al. Pyrethroid resistance in the New World malaria vector Anopheles albimanus is mediated by cytochrome P450 CYP6P5. Pestic Biochem Physiol. 2022;183:105061. doi:10.1016/j.pestbp.2022.105061
45. Ibrahim SS, Riveron JM, Stott R, Irving H, Wondji CS. The cytochrome P450 CYP6P4 is responsible for the high pyrethroid resistance in knockdown resistance-free Anopheles arabiensis. Insect Biochem Mol Biol. 2016;68:23–32. doi:10.1016/j.ibmb.2015.10.015
46. Ibrahim SS, Muhammad A, Hearn J, et al. Molecular drivers of insecticide resistance in the Sahelo-Sudanian populations of a major malaria vector Anopheles coluzzii. BMC Biol. 2023;21:125. doi:10.1186/s12915-023-01610-5
47. Hearn J, Djoko Tagne CS, Ibrahim SS, et al. Multi-omics analysis identifies a CYP9K1 haplotype conferring pyrethroid resistance in the malaria vector Anopheles funestus in East Africa. Mol Ecol. 2022;31(13):3642–3657. doi:10.1111/mec.16497
48. Djoko Tagne CS, Kouamo MFM, Tchouakui M, et al. A single mutation G454A in the P450 CYP9K1 drives pyrethroid resistance in the major malaria vector Anopheles funestus reducing bed net efficacy. Genetics. 2025;229(1):iyae181. doi:10.1093/genetics/iyae181
49. Kouamo MFM, Ibrahim SS, Hearn J, et al. Genome-wide transcriptional analysis and functional validation linked a cluster of epsilon glutathione S-transferases with insecticide resistance in the major malaria vector Anopheles funestus across Africa. Genes. 2021;12(4):561. doi:10.3390/genes12040561
50. Tchigossou GM, Atoyebi SM, Akoton R, et al. Investigation of DDT resistance mechanisms in Anopheles funestus populations from northern and southern Benin reveals a key role of the GSTe2 gene. Malar J. 2020;19(1):456. doi:10.1186/s12936-020-03503-2
51. Diallo M, Kolley ES, Dia AK, et al. Evolution of the Ace-1 and Gste2 mutations and their potential impact on the use of carbamate and organophosphates in IRS for Controlling Anopheles gambiae s.l. the major malaria mosquito in Senegal. Pathogens. 2022;11(9):1021. doi:10.3390/pathogens11091021
52. Zhong D, Degefa T, Zhou G, et al. Esterase-mediated pyrethroid resistance in populations of an invasive malaria vector Anopheles stephensi from Ethiopia. Genes. 2024;15(12):1603. doi:10.3390/genes15121603
53. Lucas ER, Nagi SC, Kabula B, et al. Copy number variants underlie major selective sweeps in insecticide resistance genes in Anopheles arabiensis. PLOS Biol. 2024;22(12):e3002898. doi:10.1371/journal.pbio.3002898
54. Fadel AN, Ibrahim SS, Sandeu MM, et al. Exploring the molecular mechanisms of increased intensity of pyrethroid resistance in Central African population of a major malaria vector Anopheles coluzzii. Evol Appl. 2024;17(2):e13641. doi:10.1111/eva.13641
55. Odhiambo CA, Derilus D, Impoinvil LM, et al. Key gene modules and hub genes associated with pyrethroid and organophosphate resistance in Anopheles mosquitoes: a systems biology approach. BMC Genomics. 2024;25(1):665. doi:10.1186/s12864-024-10572-z
56. Karagianni EP, Kontomina E, Davis B, et al. Homologues of xenobiotic metabolizing N-acetyltransferases in plant-associated fungi: novel functions for an old enzyme family. Sci Rep. 2015;5:12900. doi:10.1038/srep12900
57. Ngufor C, Fongnikin A, Rowland M, N’Guessan R. Indoor residual spraying with a mixture of clothianidin (a neonicotinoid insecticide) and deltamethrin provides improved control and long residual activity against pyrethroid resistant Anopheles gambiae sl in Southern Benin. PLoS One. 2017;12(12):e0189575. doi:10.1371/journal.pone.0189575
58. Fouet C, Ashu FA, Ambadiang MM, Tchapga W, Wondji CS, Kamdem C. Clothianidin-resistant Anopheles gambiae adult mosquitoes from Yaoundé, Cameroon, display reduced susceptibility to SumiShield® 50WG, a neonicotinoid formulation for indoor residual spraying. BMC Infect Dis. 2024;24(1):133. doi:10.1186/s12879-024-09030-8
59. Wu C, Chakrabarty S, Jin M, Liu K, Xiao Y. Insect ATP-binding cassette (ABC) transporters: roles in xenobiotic detoxification and bt insecticidal activity. Int J Mol Sci. 2019;20(11):2829. doi:10.3390/ijms20112829
60. Shan J, Sun X, Li R, Zhu B, Liang P, Gao X. Identification of ABCG transporter genes associated with chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag Sci. 2021;77(7):3491–3499. doi:10.1002/ps.6402
61. Pignatelli P, Ingham VA, Balabanidou V, Vontas J, Lycett G, Ranson H. The Anopheles gambiae ATP-binding cassette transporter family: phylogenetic analysis and tissue localization provide clues on function and role in insecticide resistance. Insect Mol Biol. 2018;27(1):110–122. doi:10.1111/imb.12351
62. He Q, Yan Z, Si F, Zhou Y, Fu W, Chen B. ATP-binding cassette (ABC) transporter genes involved in pyrethroid resistance in the malaria vector anopheles sinensis: genome-wide identification, characteristics, phylogenetics, and expression profile. Int J Mol Sci. 2019;20(6):1409. doi:10.3390/ijms20061409
63. Kefi M, Balabanidou V, Sarafoglou C, et al. ABCH2 transporter mediates deltamethrin uptake and toxicity in the malaria vector Anopheles coluzzii. PLoS Pathog. 2023;19(8):e1011226. doi:10.1371/journal.ppat.1011226
64. Saizonou H, Impoinvil LM, Derilus D, et al. Transcriptomic analysis of Anopheles gambiae from Benin reveals overexpression of salivary and cuticular proteins associated with cross-resistance to pyrethroids and organophosphates. BMC Genomics. 2024;25(1):348. doi:10.1186/s12864-024-10261-x
65. Edi CV, Djogbénou L, Jenkins AM, et al. CYP6 P450 enzymes and ACE-1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLOS Genet. 2014;10(3):e1004236. doi:10.1371/journal.pgen.1004236
66. Shaw WR, Teodori E, Mitchell SN, et al. Mating activates the heme peroxidase HPX15 in the sperm storage organ to ensure fertility in Anopheles gambiae. Proc Natl Acad Sci U S A. 2014;111(16):5854–5859. doi:10.1073/pnas.1401715111
67. Saab SA, Cardoso-Jaime V, Kefi M, Dimopoulos G. Advances in the dissection of Anopheles–Plasmodium interactions. PLOS Pathog. 2025;21(3):e1012965. doi:10.1371/journal.ppat.1012965
68. Hamid-Adiamoh M, Jabang AMJ, Opondo KO, Ndiath MO, Assogba BS, Amambua-Ngwa A. Distribution of Anopheles gambiae thioester-containing protein 1 alleles along malaria transmission gradients in The Gambia. Malar J. 2023;22(1):89. doi:10.1186/s12936-023-04518-1
69. Tchouakui M, Mugenzi LMJ, Wondji MJ, Tchoupo M, Njiokou F, Wondji CS. Combined over-expression of two cytochrome P450 genes exacerbates the fitness cost of pyrethroid resistance in the major African malaria vector Anopheles funestus. Pestic Biochem Physiol. 2021;173:104772. doi:10.1016/j.pestbp.2021.104772
70. Tchouakui M, Riveron Miranda J, Mugenzi LMJ, et al. Cytochrome P450 metabolic resistance (CYP6P9a) to pyrethroids imposes a fitness cost in the major African malaria vector Anopheles funestus. Heredity. 2020;124(5):621–632. doi:10.1038/s41437-020-0304-1
71. Mugenzi LMJ, Tekoh TA, Ibrahim SS, et al. The duplicated P450s CYP6P9a/b drive carbamates and pyrethroids cross-resistance in the major African malaria vector Anopheles funestus. PLOS Genet. 2023;19(3):e1010678. doi:10.1371/journal.pgen.1010678
72. Ruel DM, Vainer Y, Yakir E, Bohbot JD. Identification and functional characterization of olfactory indolergic receptors in Drosophila melanogaster. Insect Biochem Mol Biol. 2021;139:103651. doi:10.1016/j.ibmb.2021.103651
73. Pitts RJ, Huff RM, Shih SJ, Bohbot JD. Identification and functional characterization of olfactory indolergic receptors in Musca domestica. Insect Biochem Mol Biol. 2021;139:103653. doi:10.1016/j.ibmb.2021.103653
74. Dekel A, Sar-Shalom E, Vainer Y, Yakir E, Bohbot JD. The ovipositor cue indole inhibits animal host attraction in Aedes aegypti (Diptera: culicidae) mosquitoes. Parasit Vectors. 2022;15(1):422. doi:10.1186/s13071-022-05545-8
75. Huff RM, Pitts RJ. Functional conservation of Anopheline linalool receptors through 100 million years of evolution. Chem Senses. 2022;47:bjac032. doi:10.1093/chemse/bjac032
76. Tsitoura P, Iatrou K. Positive allosteric modulation of insect olfactory receptor function by ORco agonists. Front Cell Neurosci. 2016;10:275. doi:10.3389/fncel.2016.00275
77. Wang B, Liu Y, He K, Wang G. Comparison of research methods for functional characterization of insect olfactory receptors. Sci Rep. 2016;6(1):32806. doi:10.1038/srep32806
78. Huestis DL, Dao A, Diallo M, et al. Windborne long-distance migration of malaria mosquitoes in the Sahel. Nature. 2019;574(7778):404–408. doi:10.1038/s41586-019-1622-4
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.