Back to Journals » Cancer Management and Research » Volume 18

Dihydrocapsaicin Secreted by RYK Silenced Bone Marrow-Derived Mesenchymal Stem Cells Triggers Apoptosis of Gastric Cancer Cells
Authors Fu Y, Cai Z, Wang Y, Huang M, Huang J ![]()
Received 6 November 2025
Accepted for publication 15 March 2026
Published 21 March 2026 Volume 2026:18 579598
DOI https://doi.org/10.2147/CMAR.S579598
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Yongan Fu, Zongda Cai, Yangqiang Wang, Mingjin Huang, Jinghua Huang
Department of Gastrointestinal Surgery, Quanzhou First Affiliated Hospital to Fujian Medical University, Quanzhou, People’s Republic of China
Correspondence: Jinghua Huang, Department of Gastrointestinal Surgery, Quanzhou First Affiliated Hospital to Fujian Medical University, 250 East Street, Quanzhou city, People’s Republic of China, Tel +8618960466116, Email [email protected]
Introduction: Bone marrow-derived mesenchymal stem cells (BMSCs) are recruited into the gastric cancer (GC) microenvironment and promote progression, though the underlying mechanisms remain unclear. This study investigated how RYK-silenced BMSCs induce GC cell apoptosis, with a focus on the novel role of dihydrocapsaicin (DHC).
Methods: BMSCs were transfected with RYK siRNA or negative controls and co-cultured with NCI-N87 cells to investigate their interactions. Cancer cell cycle and apoptosis were assessed by flow cytometry, while protein levels of Caspase3, Bax, and Bcl-2 in NCI-N87 cells were determined via Western blot. Metabolomics analyzed differential metabolites in BMSCs. Additionally, NCI-N87 cells were treated with DHC, and their proliferation, apoptosis, and apoptosis-related protein expression were evaluated after different DHC treatments.
Results: Compared to NCI-N87 cells cultured alone, co-culture with si-NC-modified BMSCs reduced apoptosis in NCI-N87 cells. However, co-culture with si-RYK-BMSCs significantly increased apoptosis. Additionally, DHC, a metabolic product secreted by BMSCs after RYK interference, suppresses NCI-N87 cell growth, promotes cell death, and increases the expression of apoptosis-related proteins.
Conclusion: RYK-silenced BMSCs induce NCI-N87 apoptosis, likely through increased DHC secretion, highlighting DHC as a novel mediator in GC progression.
Keywords: dihydrocapsaicin, RYK, bone marrow-derived mesenchymal stem cells, gastric cancer, apoptosis
Introduction
Gastric cancer (GC) ranks as one of the most prevalent malignancies globally, being the fifth leading cause of cancer-related mortality worldwide.1 It is also the most common cancer in East Asia, with approximately 990,000 new cases diagnosed annually.2 GC is a multifactorial disease influenced by both environmental and genetic factors. Key risk factors include age, sex, smoking, and Helicobacter pylori infection.3–5 Despite advancements in surgical resection and adjuvant chemotherapy that have improved patient survival, the overall prognosis for gastric cancer remains suboptimal.6 Therefore, it is urgent to elucidate the development mechanism of gastric cancer and find effective treatment strategies.
Mesenchymal stem cells (MSCs) are intricately associated with the initiation and progression of tumors.7,8 Research has demonstrated that circulating MSCs can migrate and localize to tumor sites under the chemotactic influence of growth factors and chemokines secreted by tumor cells and other inflammatory cells, thereby contributing to the formation of the tumor microenvironment.9 The biological properties of MSCs are modulated by the tumor microenvironment, which may enhance their angiogenic capacity, immunosuppressive functions, and anti-apoptotic activities.10 Moreover, MSCs can facilitate the epithelial-mesenchymal transition in tumor cells and protect the survival of cancer stem cells, thereby contributing significantly to tumor initiation and progression. Earlier research has indicated that bone marrow-derived mesenchymal stem cells (BMSCs) can accelerate the development of gastric cancer.11 Acting as a Wnt5a receptor in the non-canonical Wnt signaling pathway, RYK participates in the regulation of the onset and progression of various cancers.12–14 Our prior investigation also demonstrated that knocking down RYK in gastric cancer cells could suppress the malignant behavior of gastric cancer.15 Nevertheless, no studies have explored the impact of RYK depletion in BMSCs on the apoptosis of gastric cancer cells. Metabolomics was employed to identify key metabolites secreted by RYK-silenced BMSCs, as these small molecules often serve as critical mediators of cell-cell communication and can reveal novel pathways linking stromal modulation to cancer cell fate.
Dihydrocapsaicin (DHC), a major active capsaicinoid found in chili peppers, has garnered increasing attention for its pharmacological properties, particularly its anti-tumor potential.16 Emerging evidence suggests that DHC and related capsaicinoids can inhibit cell proliferation and induce apoptosis in various malignancies, including cervical cancer and prostate cancer.17,18 However, the specific mechanism by which DHC affects gastric cancer cells, especially in the context of the tumor microenvironment modulated by mesenchymal stem cells, remains to be fully elucidated. Consequently, this study was designed to examine how RYK-depleted BMSCs influence the apoptosis of gastric cancer cells and to elucidate the associated molecular mechanisms.
Materials and Methods
Materials
Human BMSCs and the gastric cancer cell line NCI-N87 were obtained from Wuhan Pricella Biotechnology Co., Ltd. (Shanghai, China). The Cell Counting Kit-8 and the Annexin V-FITC Apoptosis Detection Kit were acquired from Solaibao Technology Co., Ltd. (Beijing, China). Antibodies targeting Caspase-3, Bax, Bcl-2, RYK, β-Actin, and Caspase-9 were sourced from Abcam (Cambridge, United Kingdom). Anthraquinone, DHC, and Cerulenin were purchased from MCE (New Jersey, USA). The siRNAs, including Si-RYK-1, Si-RYK-2, Si-RYK-3, and the negative control siRNA (si-NC), were provided by GenePharma (Shanghai, China).
Cell Culture and Transfection
Human BMSCs and NCI-N87 cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin at 37 °C in an atmosphere containing 5% CO2.
The transfection procedure was initiated when the confluence of BMSCs reached 70%. Initially, the culture medium was replaced with 1 mL of serum-free medium. Two sterilized EP tubes were prepared, and each tube received 125 μL of Opti-MEM diluent. To one EP tube, 5 μL of Lipofectamine 3000 reagent was added, while to the other, 12.5 μL of siRNA was introduced (the siRNA powder should be pre-dissolved in DEPC water at a concentration of 1 OD/125 μL). After thorough mixing, both EP tubes were incubated at room temperature for 5 minutes. The contents of the two EP tubes were then combined and mixed evenly, followed by an additional incubation at room temperature for 15 minutes. Drops of this combined solution were subsequently added to the appropriate wells of a six-well plate, after which the cells were returned to the incubator for further growth. Four to six hours post-transfection, 1 mL of complete medium containing 20% serum was added to the six-well plate. After 48 hours, the conditioned medium required for the experiment was harvested. This experiment included three control groups: the first group, labeled as si-NC-BMSCs+N87, involved treating NCI-N87 cells with the conditioned medium from the si-NC group of BMSCs for 24 hours; the second group, designated as si-RYK-BMSCS + N87, entailed treating NCI-N87 cells with the conditioned medium from the si-RYK group of BMSCs for 24 hours; the third group, simply referred to as the N87 group, consisted of untreated normal NCI-N87 cells cultured for 24 hours as a baseline comparison.
Quantitative Real-Time PCR (qRT-PCR)
Samples were collected and the total RNA was isolated using a Total RNA Miniprep Kit. Subsequently, cDNA synthesis was carried out following the instructions provided by the manufacturer. The qRT-PCR analysis was performed under these conditions: an initial denaturation step at 95 °C for 5 minutes, then denaturation at 95 °C for 10 seconds, annealing at 60 °C for 30 seconds, and a final extension phase from 65 to 95 °C, increasing the temperature by 0.5 °C every 5 seconds. After completion of the reaction, average cycle threshold (Ct) values were determined for each gene as well as for the reference gene. The relative expression levels of genes were evaluated using the widely accepted method known as 2− ΔΔCT. The sequences of the primers were showed in Table 1.
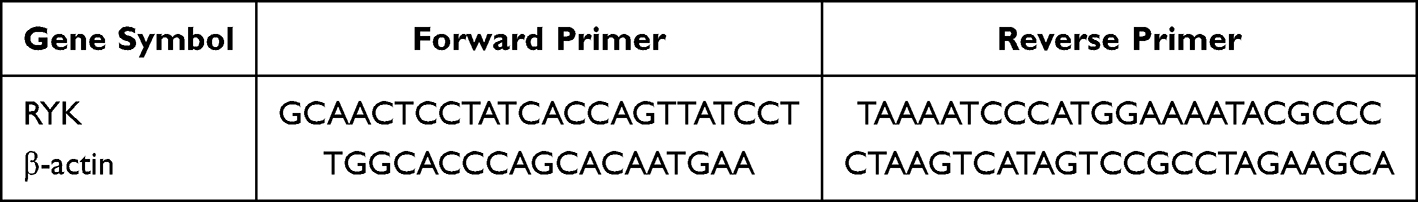 |
Table 1 Primer Sequence |
Cell Apoptosis Assay
The harvested cells (1×106) were first centrifuged at 1500 rpm for 3 minutes using PBS, followed by two rinsing steps. Next, the cell pellet was re-suspended in 300 μL of pre-chilled 1× Annexin V-FITC binding buffer. Afterwards, 5 μL of Annexin V-FITC and 10 μL of propidium iodide (PI) were added to each sample well. The mixture was gently vortexed and then incubated in the dark at room temperature for 10 minutes. Finally, the samples were subjected to analysis via NovoCyte flow cytometry.
Western Blotting
According to the method proposed by Wu et al,19 the detection of apoptosis-related proteins was conducted across different cell groups.
Non-Targeted Metabolomics Assay
Differential metabolites in various cell types were detected using liquid chromatography-mass spectrometry (LC-MS) as previously described.20
Targeted Differential Metabolites Were Detected by LC-MS
Take 100 μL of the cell supernatant sample and add 300 μL of methanol. Vortex for 60 seconds, then centrifuge at 17,000 × g for 15 minutes. Collect the supernatant for subsequent analysis. Standard solutions of varying concentrations were prepared using 75% methanol, and mixed with the internal standard in a 1:1 ratio to generate the calibration standards. The sample loading was performed under the following conditions: the column temperature was maintained at 40°C, with an injection volume of 3 μL and utilizing the positive ion mode. The mobile phase was composed of two components: A (water containing 0.1% formic acid) and B (acetonitrile). Gradient elution was carried out as follows: 4% B (from 0 to 3 min), increasing from 4% to 70% B (from 3 to 10 min), further increasing from 70% to 100% B (from 10 to 36 min), decreasing from 100% to 4% B (from 36 to 37 min), and maintaining 4% B (from 37 to 40 min). The flow rate was kept constant at 400 μL/min. For mass spectrometry, data were collected using an AB 6600 mass spectrometer in positive ion mode. The ESI ion source settings were adjusted as follows: gas temperature set to 500°C, curtain gas pressure at 25 Psi, collision gas pressure at 10 Psi, ion spray voltage at 5500 V, and atomization temperature at 500°C.
CCK-8 Assay
Cell viability was measured using a cell counting kit-8. Briefly, cells (2×104/well) were seeded in 96-well plate and cultured for 48 h. Then, each well was supplement with 10 μL CCK8 solution and incubated for 1 h. Afterwards, the absorbance was measured using a microplate reader at the wavelength of 450 nm.
Statistical Analyses
All experiments were performed independently at least three times. The statistical analysis was conducted using SPSS 26.0, with experimental data presented as the mean ± standard deviation (x ± s). Inter group comparisons were analyzed through One-Way ANOVA, while two-group comparisons were performed using a t-test. The sample size was determined based on preliminary experiments and standard practices in the field to ensure sufficient statistical power to detect significant differences. A p-value of less than 0.05 was regarded as indicating a statistically significant difference.
Results
Construction of RYK-Silenced BMSCs
Si-NC and three different si-RYK was transfected into BMSCs, and the silencing effect of RYK was verified by qPCR (Figure 1A) and WB (Figure 1B and C). The unprocessed original full blot/gel images of Figure 1B were shown in Figure S1. Compared with the si-NC group, the mRNA and protein expression of RYK in si-RYK-1, si-RYK-2, and si-RYK-3 group were significantly decreased in BMSCs (P<0.05). According to the results of silencing RKY, we used si-RYK-3 for follow-up experiments.
 |
Figure 1 Construction of RYK-silenced BMSCs. (A) Transfection efficiency of si-RYKs in BMSCs was verified by qRT-PCR, (B) Transfection efficiency of si-RYKs in BMSCs was verified by WB, (C) Quantitative analysis of RYK protein expression. Data are presented as the mean±SD (n=3). *P <0.05 vs. si-NC. |
Effect of RYK-Silenced BMSCs on Apoptosis of NCI-N87 Cells
As illustrated in Figure 2A, flow cytometry was employed to assess the apoptosis of various NCI-N87 cells. In comparison with the N87 group, a significant reduction in apoptosis was observed in the si-NC-BMSCs+N87 group (P<0.05). These findings indicate that BMSCs possess the ability to suppress apoptosis in NCI-N87 cells. Furthermore, when compared to the si-NC-BMSCs+N87 group, there was a marked increase in apoptosis in the si-RYK-BMSCs+NCI-N87 group (P<0.05) (Figure 2B). Additionally, we further analyzed the expression levels of proteins associated with apoptosis (Caspase3, Bax, and Bcl-2) by Western blotting (Figure 2C). The unprocessed original full blot/gel images of Figure 2C were shown in Figure S2. The quantitative protein analysis results are displayed in Figure 2D. Relative to the N87 group, the expression levels of Caspase3 and Bax proteins in NCI-N87 cells were notably reduced in the si-NC-BMSCs+N87 group (P<0.05), whereas Bcl-2 protein expression was significantly elevated (P<0.05). Conversely, in comparison with the si-NC-BMSCs+N87 group, the si-RYK-BMSCs+N87 group exhibited a significant increase in Caspase3 and Bax protein expression (P<0.05), while Bcl-2 protein expression was substantially decreased (P<0.05).
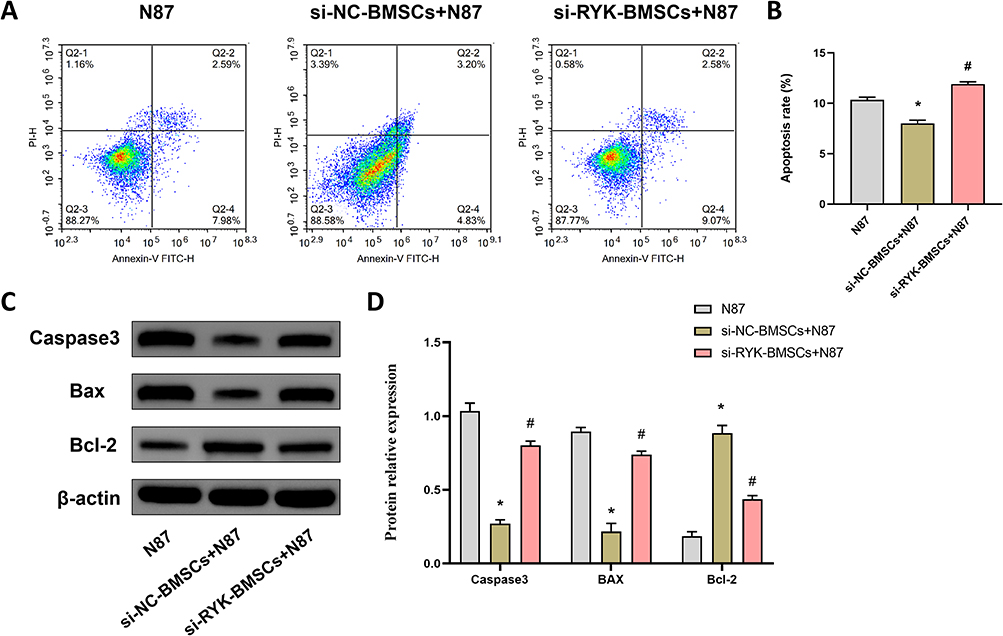 |
Figure 2 Effect of RYK-silenced BMSCs on apoptosis of NCI-N87 cells. (A and B) The apoptosis of NCI-N87 cells was detected by flow cytometry, (C and D) The apoptosis-related protein expressions of Caspase3, Bax, and Bcl-2 in NCI-N87 cells were detected by WB. Data are presented as the mean±SD (n=3). *P <0.05 vs. N87; #P <0.05 vs. si-NC-BMSCs+N87. |
Identification of Metabolites in RYK-Silenced BMSCs
To further analyze the metabolic components of si-RYK-BMSCs in inhibiting the apoptosis of NCI-N87 cells, we used metabolomics to analyze the differential metabolites between si-RYK-BMSCs and si-NC-BMSCs. The volcano plot in Figure 3A showed that there were 104 up-regulated metabolites and 306 down-regulated metabolites in the RYK group (si-RYK-BMSCs) compared with the NC group (si-NC-BMSCs). As shown in Figure 3B, We employed principal component analysis (PCA) to visualize the clustering between the NC and RYK groups and observed a significant separation in the PCA score plots between the two groups. Subsequently, we normalized the metabolites exhibiting the most significant differences, identified the top 10 metabolites with the most substantial up-regulation and the top 12 metabolites with the most pronounced down-regulation, and generated a cluster heatmap to visualize the results (Figure 3C).
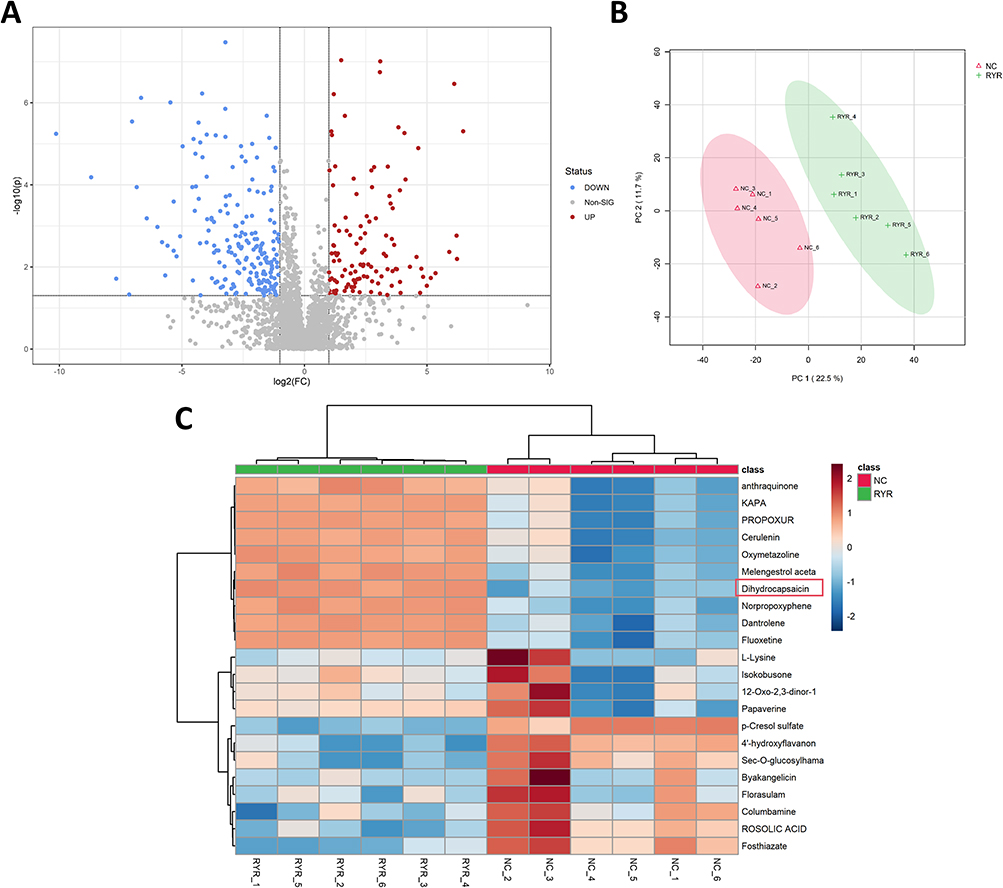 |
Figure 3 Identification of Metabolites in RYK-Silenced BMSCs. (A) Volcano plot showing the differentially expressed metabolites between the BMSCs-si-NC and BMSCs-si-RYK groups (n=6), (B) Score plots of principal component analysis from the discovery set, (C) Heatmap analysis of cell metabolites in the BMSCs-si-NC and BMSCs-si-RYK groups (n=6). The red box in the picture encloses DHC. |
Effect of DHC on Proliferation of NCI-N87 Cells
To further validate the elevated levels of differential metabolites in the BMSCs-si-RYK group, we employed liquid chromatography-mass spectrometry (LC-MS) to quantify the concentrations of three specific metabolites (Anthraquinone, DHC, and Cerulenin) in different cells (Figure 4A–C). The results showed that compared with the BMSCs-si-NC group, the level of DHC in the BMSCs-si-RYK group was significantly increased (P<0.05), whereas the levels of the other two metabolites showed no significant differences (P>0.05). Additionally, we used a CCK-8 assay to investigate the effect of DHC on the proliferation of NCI-N87 cells at 12, 24, 48h (Figure 4D). Compared with the N87 group, the proliferation of NCI-N87 cells were significantly decreased in the 60 μM DHC and 80 μM DHC group at 12, 24, 48h (P<0.05). The results showed that DHC can inhibit the proliferation of NCI-N87 cells.
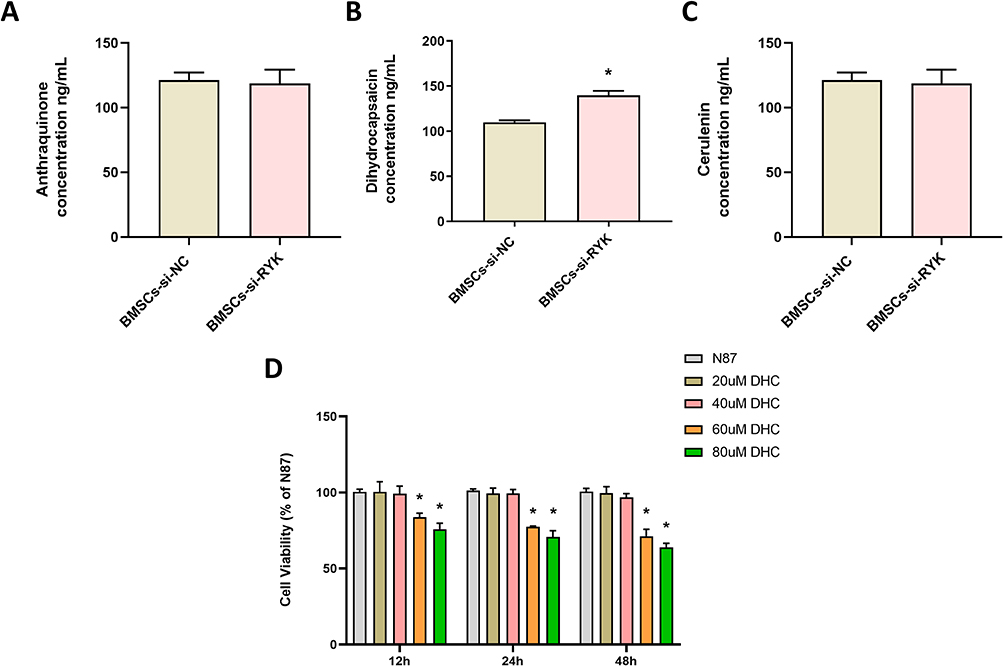 |
Figure 4 Effect of DHC on proliferation of NCI-N87 cells. (A–C) LC-MS was employed to quantify the differences in targeted metabolites across distinct cellular groups, (A) Anthraquinone, (B) Dihydrocapsaicin, (C) Cerulenin, (D) The proliferation of NCI-N87 cells was detected by CCK-8 at 12, 24, and 48h. Data are presented as the mean±SD (n=3). *P <0.05 vs. BMSCs-si-NC or N87. |
Effect of DHC on Apoptosis of NCI-N87 Cells
Subsequently, flow cytometry (Figure 5A) and WB were used to detect the apoptosis of different treated cells. The analysis results of flow cytometry were shown in Figure 5B. Compared with the N87 group, the apoptosis of NCI-N87 cells were significantly increased in the 60 μM DHC and 80 μM DHC group (P<0.05). The apoptosis level of NCI-N87 cells also increased with the increase in DHC dose. Moreover, as shown in Figure 5C and D, the Bax, Caspase3, and Caspase9 protein expression in the 60 μM DHC and 80 μM DHC group were significantly increased (P<0.05) compared with the N87 group. The unprocessed original full blot/gel images of Figure 5C were shown in Figure S3.
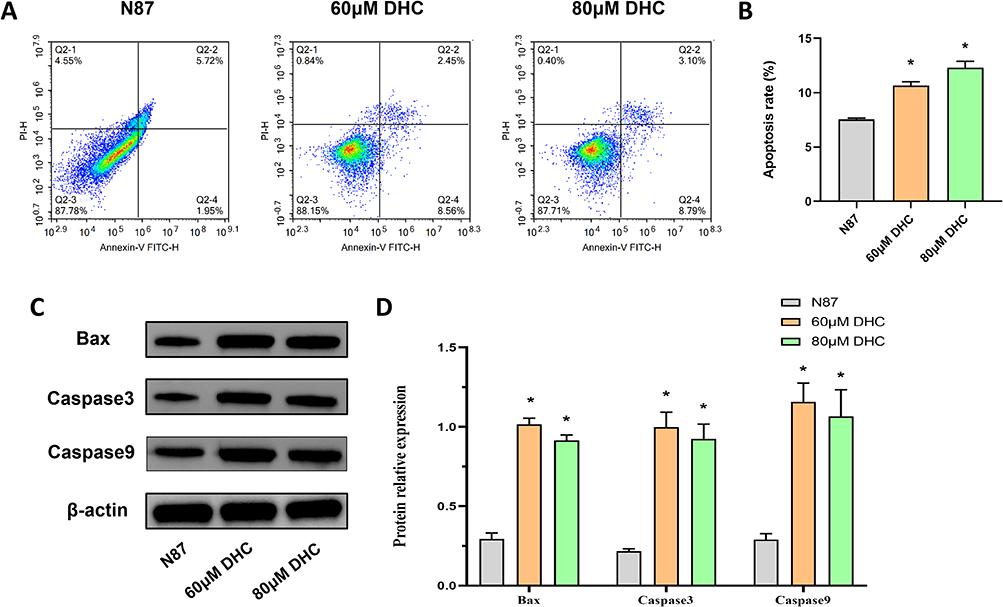 |
Figure 5 Effect of DHC on apoptosis of NCI-N87 cells. (A and B) The apoptosis of NCI-N87 cells was detected by flow cytometry, (C and D) The apoptosis-related protein expressions of Bax, Caspase3, and Caspase9 in NCI-N87 cells were detected by WB. Data are presented as the mean±SD (n=3). *P <0.05 vs. N87. |
Taken together, we concluded that si-RYK intervention in BMSCs promotes the apoptosis in gastric cancer cells by enhancing the release of DHC from BMSCs (Figure 6).
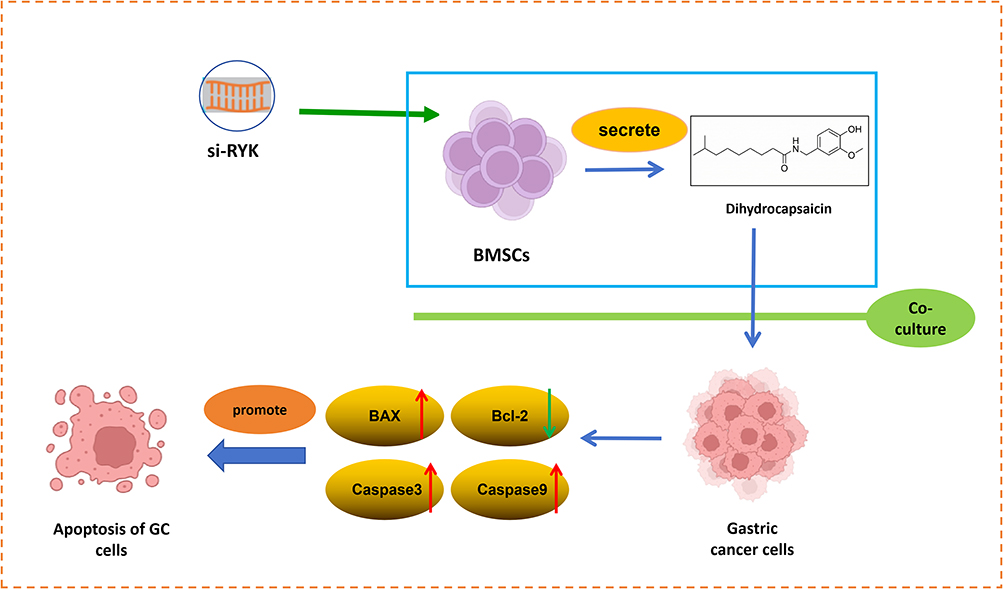 |
Figure 6 The proposed model suggests that DHC secreted by RYK-silenced BMSCs promote the apoptosis of gastric cancer cells. The upward-pointing red arrow indicates an increase in protein expression, while the downward-pointing green arrow indicates a decrease. |
Discussion
When a tumor develops within the body, non-cancerous tissue-derived mesenchymal stem cells (MSCs), such as BMSCs, are recruited to the cancerous tissue and become an integral component of the tumor microenvironment.21 Subsequently, under the influence of inflammatory cytokines and other stromal cells present at the local cancer site, BMSCs undergo differentiation, acquiring novel characteristics, and transform into resident MSCs within the tumor tissue.22 Mu et al found that co-culture of BMSCs with SGC7901 gastric cancer cells could significantly promote the apoptosis of gastric cancer cells.23 In this paper, we found that treating NCI-N87 cells with the conditioned medium from BMSCs could inhibit their apoptosis, which is consistent with the results of Mu et al However, BMSCs modified by gene editing exhibit the ability to suppress tumor progression. Liu et al found that treatment of breast cancer cells with Rictor knockout BMSCs inhibited bone metastasis of breast cancer.24 Chang et al demonstrated that treating gastric cancer cells with BMSC-derived exosomes overexpressing miR-1228 significantly suppressed the apoptosis of gastric cancer cells.25 Thus, inhibiting gastric cancer progression by utilizing genetically modified BMSCs holds significant potential for the treatment of gastric cancer.
RYK belongs to the family of receptor tyrosine kinases, a group of genes that encode proteins typically located on cell membranes. These proteins are capable of detecting external signals and converting them into intracellular biochemical reactions.26–28 A series of studies have demonstrated that the abnormal expression of RYK is significantly associated with various types of cancer, including breast, colorectal, and gastric cancers.29,30 Our earlier research also demonstrated that reducing RYK expression in gastric cancer cells can inhibit liver metastasis of gastric cancer.15 However, there are few studies that have reported the effect of si-RYK modified BMSCs on apoptosis of gastric cancer cells. In this paper, we found that treating NCI-N87 cells with the conditioned medium from si-RYK modified BMSCs could promote the apoptosis compared with the si-NC-BMSCs+N87 group.
Dihydrocapsaicin (DHC), as a spicy component of capsicum, has been found to have analgesic, anti-inflammatory, antioxidant, anti-obesity and anti-cancer effects.31–33 Shi et al found that treating melanoma cell lines with DHC could significantly inhibit the proliferation, migration and metastasis of melanoma cells.34 Xie et al demonstrated that treatment of glioma cell lines with DHC significantly enhanced the apoptosis of glioma cells.35 However, no studies have reported the effect of DHC on the development of gastric cancer. In this study, metabolomics analysis was employed to investigate the metabolite profiles of Si-RyK-modified BMSCs, revealing a significant increase in DHC levels within these modified cells. Subsequently, we treated gastric cancer cell line NCI-N87 with different concentrations of DHC, and found that DHC could significantly promote the apoptosis of NCI-N87 cells and increase the expression of pro-apoptotic protein (Bax, Caspase3, and Caspase9). Taken together, our results suggest that RYK silencing-modified BMSCs can induce the apoptosis of NCI-N87 cells, potentially through increased secretion of DHC. However, this study also has some limitations. This study did not investigate the potential signaling pathways involved in the effect of DHC on apoptosis of gastric cancer cells. We will conduct an in-depth analysis of the signaling pathway by which DHC promotes apoptosis in gastric cancer cells and validate it through both in vivo and in vitro experiments in our subsequent study. In addition, GC is a highly heterogeneous malignancy with diverse molecular subtypes and histological features. In the current study, we primarily utilized the NCI-N87 cell line to elucidate these mechanism. While NCI-N87 is a well-established model, results derived from a single cell line may not fully recapitulate the complex biological behavior of all GC subtypes. Future studies involving a broader panel of GC cell lines, patient-derived xenografts (PDX), or clinical samples are warranted to validate the generalizability of these findings. Although our metabolomics analysis identified DHC as the key differentially expressed metabolite, cell-level studies demonstrated that dihydrocapsaicin could promote apoptosis in NCI-N87 cells. However, future research is necessary to involve more GC cell lines and animal experiments to verify this effect.
Conclusions
In conclusion, our findings indicate that RYK-silenced BMSCs can trigger apoptosis in NCI-N87 cells, potentially through the enhanced secretion of DHC. It should be noted that these results are based on in vitro experiments; therefore, future validation in in vivo models and deeper mechanistic exploration are necessary to confirm the therapeutic potential.
Data Sharing Statement
The data that support the findings of this study are available on request from the corresponding author.
Acknowledgments
This paper has been uploaded to ResearchSquare as a preprint: https://www.researchsquare.com/article/rs-7167954/v1. And the abstract of this paper was presented at the Society (an independent, non-profit ecosystem that aggregates preprints with public peer reviews): https://labs.sciety.org/articles/by?article_doi=10.21203/rs.3.rs-7167954/v1.
Funding
This research was supported by grants from the Fujian Provincial Natural Science Foundation of China [grant numbers: 2021J011405; 2024J011522].
Disclosure
The authors declare no competing interest in this work.
References
1. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–10. doi:10.3322/caac.21834
2. Wu M, Yuan S, Liu K, et al. Gastric cancer signaling pathways and therapeutic applications. Technol Cancer Res Treat. 2024;23:15330338241271935. doi:10.1177/15330338241271935
3. Mamun TI, Younus S, Rahman MH. Gastric cancer-epidemiology, modifiable and non-modifiable risk factors, challenges and opportunities: an updated review. Cancer Treat Res Commun. 2024;41:100845. doi:10.1016/j.ctarc.2024.100845
4. Machlowska J, Baj J, Sitarz M, et al. Gastric cancer: epidemiology, risk factors, classification, genomic characteristics and treatment strategies. Int J Mol Sci. 2020;21(11):4012. doi:10.3390/ijms21114012
5. Yang WJ, Zhao HP, Yu Y, et al. Updates on global epidemiology, risk and prognostic factors of gastric cancer. World J Gastroenterol. 2023;29(16):2452–2468. doi:10.3748/wjg.v29.i16.2452
6. Thrift AP, Wenker TN, El-Serag HB. Global burden of gastric cancer: epidemiological trends, risk factors, screening and prevention. Nat Rev Clin Oncol. 2023;20(5):338–349. doi:10.1038/s41571-023-00747-0
7. Karami Fath M, Moayedi Banan Z, Barati R, et al. Recent advancements to engineer mesenchymal stem cells and their extracellular vesicles for targeting and destroying tumors. Prog Biophys Mol Biol. 2023;178:1–16. doi:10.1016/j.pbiomolbio.2023.02.001
8. Ridge SM, Sullivan FJ, Glynn SA. Mesenchymal stem cells: key players in cancer progression. Mol Cancer. 2017;16(1):31. doi:10.1186/s12943-017-0597-8
9. Lin W, Huang L, Li Y, et al. Mesenchymal stem cells and cancer: clinical challenges and opportunities. Biomed Res Int. 2019;2019:2820853. doi:10.1155/2019/2820853
10. Fu Y, Zhang M, Sui B, et al. Mesenchymal stem cell-derived apoptotic vesicles ameliorate impaired ovarian folliculogenesis in polycystic ovary syndrome and ovarian aging by targeting WNT signaling. Theranostics. 2024;14(8):3385–3403. doi:10.7150/thno.94943
11. Qi C, Shi H, Fan M, et al. Microvesicles from bone marrow-derived mesenchymal stem cells promote Helicobacter pylori-associated gastric cancer progression by transferring thrombospondin-2. Cell Commun Signal. 2023;21(1):274. doi:10.1186/s12964-023-01127-y
12. Keeble TR, Cooper HM. Ryk: a novel Wnt receptor regulating axon pathfinding. Int J Biochem Cell Biol. 2006;38(12):2011–2017. doi:10.1016/j.biocel.2006.07.005
13. Liang Z, Liu L, Gao R, et al. Downregulation of exosomal miR-7-5p promotes breast cancer migration and invasion by targeting RYK and participating in the atypical WNT signalling pathway. Cell Mol Biol Lett. 2022;27(1):88. doi:10.1186/s11658-022-00393-x
14. FAdamo A, Fiore D, De Martino F, et al. RYK promotes the stemness of glioblastoma cells via the WNT/β-catenin pathway. Oncotarget. 2017;8(8):13476–13487. doi:10.18632/oncotarget.14564
15. Fu Y, Chen Y, Huang J, et al. RYK, a receptor of noncanonical Wnt ligand Wnt5a, is positively correlated with gastric cancer tumorigenesis and potential of liver metastasis. Am J Physiol Gastrointest Liver Physiol. 2020;318(2):G352–G360. doi:10.1152/ajpgi.00228.2019
16. Chen Y, Yu J, He H, et al. Dihydrocapsaicin attenuates oxidative stress and apoptosis in acute myocardial infarction via promoting Raf-1/ASK1 complex formation. Phytomedicine. 2025;146:157126. doi:10.1016/j.phymed.2025.157126
17. Boonyarat C, Sakurai H, Hayakawa Y, et al. Dihydrocapsaicin enhances tumor necrosis factor-α-induced apoptosis and G1 cell cycle arrest in human cervical cancer cells through TAK1-mediated NF-κB and EGFR pathways. Int J Mol Sci. 2025;26(11):5011. doi:10.3390/ijms26115011
18. Zhao J, Liu X, Pang Y, et al. Androgen receptor inhibitory activity of dihydrocapsaicin: insights from in vitro, in vivo and in silico studies. J Steroid Biochem Mol Biol. 2026;255:106872. doi:10.1016/j.jsbmb.2025.106872
19. Wu YJ, Wong BS, Yea SH, et al. Sinularin induces apoptosis through mitochondria dysfunction and inactivation of the pI3K/Akt/mTOR pathway in gastric carcinoma cells. Mar Drugs. 2016;14(8):142. doi:10.3390/md14080142
20. Yang L, Hou Y, Du YE, et al. Mirtronic miR-4646-5p promotes gastric cancer metastasis by regulating ABHD16A and metabolite lysophosphatidylserines. Cell Death Differ. 2021;28(9):2708–2727. doi:10.1038/s41418-021-00779-y
21. Zhang H, Cao X, Gui R, et al. Mesenchymal stem/stromal cells in solid tumor microenvironment: orchestrating NK cell remodeling and therapeutic insights. Int Immunopharmacol. 2024;142(Pt B):113181. doi:10.1016/j.intimp.2024.113181
22. Taheri M, Tehrani HA, Dehghani S, et al. Nanotechnology and bioengineering approaches to improve the potency of mesenchymal stem cell as an off-the-shelf versatile tumor delivery vehicle. Med Res Rev. 2024;44(4):1596–1661. doi:10.1002/med.22023
23. Mu L, Sui W, Lin Y, et al. Androgen attenuates antitumor effects of gastric cancer cells by bone marrow mesenchymal stem cells via restricting the JNK signaling activation. Transl Cancer Res. 2019;8(3):917–927. doi:10.21037/tcr.2019.06.04
24. Liu Z, Wang H, He J, et al. Rictor ablation in BMSCs inhibits bone metastasis of TM40D cells by attenuating osteolytic destruction and CAF formation. Int J Biol Sci. 2019;15(11):2448–2460. doi:10.7150/ijbs.37241
25. Chang L, Gao H, Wang L, et al. Exosomes derived from miR-1228 overexpressing bone marrow-mesenchymal stem cells promote growth of gastric cancer cells. Aging. 2021;13(8):11808–11821. doi:10.18632/aging.202878
26. Zhai M, Peng B, Zhu H, et al. Wnt5a/Ryk signaling contributes to bone cancer pain by sensitizing the peripheral nociceptors through JNK-mediated TRPV1 pathway in rats. Pain. 2025;166(3):680–692. doi:10.1097/j.pain.0000000000003426
27. Gonzalez RD, Small GW, Green AJ, et al. RYK Gene expression associated with drug response variation of temozolomide and clinical outcomes in glioma patients. Pharmaceuticals. 2023;16(5):726. doi:10.3390/ph16050726
28. Zhou G, Zhang S, Jin M, et al. Comprehensive analysis reveals COPB2 and RYK associated with tumor stages of larynx squamous cell carcinoma. BMC Cancer. 2022;22(1):667. doi:10.1186/s12885-022-09766-z
29. Habu M, Koyama H, Kishida M, et al. Ryk is essential for Wnt-5a-dependent invasiveness in human glioma. J Biochem. 2014;156(1):29–38. doi:10.1093/jb/mvu015
30. Gorroño-Etxebarria I, Aguirre U, Sanchez S, et al. Wnt-11 as a potential prognostic biomarker and therapeutic target in colorectal cancer. Cancers. 2019;11(7):908. doi:10.3390/cancers11070908
31. Lai Y, Yang N, Chen X, et al. Dihydrocapsaicin suppresses the STING-mediated accumulation of ROS and NLRP3 inflammasome and alleviates apoptosis after ischemia-reperfusion injury of perforator skin flap. Phytother Res. 2024;38(5):2539–2559. doi:10.1002/ptr.8167
32. Laorob T, Ngoenkam J, Nuiyen A, et al. Comparative effectiveness of nitro dihydrocapsaicin, new synthetic derivative capsaicinoid, and capsaicin in alleviating oxidative stress and inflammation on lipopolysaccharide-stimulated corneal epithelial cells. Exp Eye Res. 2024;244:109950. doi:10.1016/j.exer.2024.109950
33. Hudáková T, Šemeláková M, Očenáš P, et al. Chili pepper extracts, capsaicin, and dihydrocapsaicin as potential anticancer agents targeting topoisomerases. BMC Complement Med Ther. 2024;24(1):96. doi:10.1186/s12906-024-04394-5
34. Shi S, Li C, Zhang Y, et al. Dihydrocapsaicin inhibits cell proliferation and metastasis in melanoma via down-regulating β-catenin pathway. Front Oncol. 2022;12:1052365. doi:10.3389/fonc.2021.648052
35. Xie L, Xiang GH, Tang T, et al. Capsaicin and dihydrocapsaicin induce apoptosis in human glioma cells via ROS and Ca2+‑mediated mitochondrial pathway. Mol Med Rep. 2016;14(5):4198–4208. doi:10.3892/mmr.2016.5784
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A 5`-tRNA Derived Fragment NamedtiRNA-Val-CAC-001 Works as a Suppressor in Gastric Cancer
Zheng J, Li C, Zhu Z, Yang F, Wang X, Jiang P, Yan F
Cancer Management and Research 2022, 14:2323-2337
Published Date: 4 August 2022