Back to Journals » Clinical Ophthalmology » Volume 14
Differential miRNA Expression: Signature for Glaucoma in Pseudoexfoliation
Authors Rao A ![]() , Chakraborty M, Roy A, Sahay P, Pradhan A, Raj N
, Chakraborty M, Roy A, Sahay P, Pradhan A, Raj N
Received 29 March 2020
Accepted for publication 7 August 2020
Published 5 October 2020 Volume 2020:14 Pages 3025—3038
DOI https://doi.org/10.2147/OPTH.S254504
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Aparna Rao,* Munmun Chakraborty,* AkashSingha Roy,* Prity Sahay, Amiya Pradhan, Niranjan Raj
Glaucoma Service, LV Prasad Eye Institute, Bhubaneswar, Odisha 751024, India
*These authors contributed equally to this work
Correspondence: Aparna Rao
LV Prasad Eye Institute, Patia, Bhubaneswar, India
Tel +91 674 398 7999/7201
Fax +91 674 398 7130
Email [email protected]
Purpose: To investigate the microRNA (miRNA) profile in patients with different stages of pseudoexfoliation (PXF).
Methods: Peripheral blood of patients with PXF (naïve to medical therapy and with no systemic disease/drugs) with ocular hypertension (OHT) and pseudoexfoliation glaucoma (PXG) was evaluated in triplicate for miRNA profiling using polymerase chain reaction (PCR) arrays. Those identified in the discovery stage were validated with evaluation of serum transforming growth factor-β 1 (TGF-β 1) levels by ELISA. The downstream targets of TGF-β 1 and unfolded protein response (UPR) were analyzed using reverse transcription-quantitative polymerase chain reaction (RT-qPCR). Predicted targets of the identified miRNA and KEGG pathway analysis were done using miRbase and DIANA tools mirPathv3.1.
Results: We found hsa-miR-122-5p, hsa-miR-124-3p and hsa-miR-424-5p to be upregulated in PXG targeting 3 specific pathways namely TGF-β 1, fibrosis/ECM and proteoglycan metabolism with common effectors like SMAD/3/2. The unfolded protein response (UPR) genes were significantly downregulated in all stages of PXF suggesting this as the key mechanism for protein aggregates in PXF syndrome. Serum TGF-β 1 was significantly upregulated as disease progressed to later stages in PXG. This elevation in advanced stages was associated with significantly differential expression of downstream pathways and fibrotic genes in OHT compared to PXG predominantly through the SMAD3, a canonical pathway marker.
Conclusion: Circulatory miRNA differentially regulating TGF-β 1 and downstream targets including UPR genes may be the key mechanisms for glaucoma onset in PXF.
Keywords: pseudoexfoliation, pseudoexfoliation glaucoma, ocular hypertension, miRNA, transforming growth factor
Introduction
Pseudoexfoliation (PXF) is a systemic microfibrillopathy characterized by accumulation of exfoliative material (XFM) in different parts of the eye causing glaucomatous neurodegeneration and irreversible blindness.1 Aberrant extracellular matrix (ECM) remodeling coupled with accumulation of complex protein aggregates in the ocular tissues gradually causes progressive fibrosis leading to trabecular meshwork (TM) dysfunction, obstruction to aqueous humor (AH) outflow, and raised intraocular pressure (IOP).1,2 Clinically, an eye with PXF disease may originally lack obvious PXF deposits (unmanifest disease). This progresses to manifest PXF with normal IOP, the stage with raised IOP (OHT), and lastly to pseudoexfoliative glaucoma with irreversible optic nerve/visual field damage (PXG). These complex protein aggregates are a result of misfolded proteins triggered by endoplasmic reticulum (ER) stress, oxidative stress or impaired chaperone effect.2,3 In response, regulatory processes like enhanced unfolded protein response (UPR) are recruited to increase the degradation of complex protein aggregates.3–5 The mechanism by which these regulatory mechanisms get overwhelmed in advanced disease would give insight into potential stages of functional reversibility by medical/other intervention. Unlike other forms of glaucoma, PXF and PXG are unique in having systemic associations with changes in peripheral blood, therefore making it possible to identify putative markers/changes in the peripheral blood marking transition to the next stage or identifying “eyes at risk”.2–4 The pathogenesis of this unique disease is derived from abnormal elastotic process mediated by stress or inflammatory mediators, breakdown of blood-aqueous barrier, precipitation of ECM components onto tissue surfaces, decrease of chaperone (clusterin) activity and genetic predisposition including LOXL1 polymorphisms.2 Yet, the role of all these processes and how these are altered between PXF stages is unknown.
Our earlier study found decreased matrix metalloproteinase-9 (MMP-9) activity in advanced stages of pseudoexfoliation suggesting exhaustion of MMP mediated ECM degradation in advanced PXF disease.6 Transforming growth factor-β1 (TGF-β1) is a molecule that tightly controls various downstream genes regulating fibrosis, MMP activity, ECM remodeling and unfolded protein responses in cells therapy making this an important molecule in PXF pathogenesis.2,4,5,7 While TGF is known to be upregulated in all stages of PXF, the mechanism by which the cytoprotective effects mediated by TGF are altered to cytotoxic effects in later stages (TGF paradox) is unknown. One of the reasons could be double negative feedback loop connection of the TGF pathway with the microRNA (miRNA) pathway. MicroRNAs are small molecules which act as regulatory switches turning on or off several key post-transcriptional gene modifications mediating transition of cell phenotypes and determining reversibility of epithelial mesenchymal transition, EMT.8,9 These circulatory miRNA may trigger local tissue effects mediating clearing of debris/aggregates, EMT/fibrosis or collagen assembly in the TM and neighboring ECM and may be responsible for the paradoxical effects in advanced disease stages.5 The role of circulatory miRNA and their role in regulating downstream TGF effectors in different PXF stages may give insight into specific miRNAs playing a role in causing a switch from reversibility to irreversibility in the TM and signaling the onset of glaucoma. This study explored the miRNAs differentially expressed in different disease stages of pseudoexfoliation and tested their specific role in glaucoma pathogenesis in this disease.
Patients and Methods
Patients diagnosed with pseudoexfoliation at the glaucoma services from 2011 to 2016 were screened. This study was conducted in accordance with the Declaration of Helsinki after obtaining written informed consent from all patients. This study was approved by the institutional approval of the LV Prasad eye Institute, MTC campus, India.
All patients underwent detailed ocular examination including slit lamp examination, Goldman applanation IOP, gonioscopy, and fundus biomicroscopy. Visual fields were performed for all patients (24–2 SITA standard program) and 2 reliable fields were done before a reliable baseline was established.
PXF was diagnosed in the presence of classical dandruff (classical PXF phenotype) or radial pigmentary (pigmentary PXF) deposits on various ocular structures of the eye like lens capsule, iris or pupil.6,10 While this phenotypic pattern reflects clinical variants of early disease, stages of the disease were categorized as follows: patients with PXF, raised IOP and normal optic nerve/visual field were classified as PXF with OHT while those with optic nerve and corresponding visual field damage were labeled as PXG. Patients with no PXF and normal disc/field attending our services for cataract surgery were taken as normal (henceforth labeled as controls). To avoid bias due to long-term medications, patients naïve to ocular or systemic medical therapy were chosen for analysis. Since this was based on circulatory miRNA, only bilateral cases were considered which excluded patients with eyes in different PXF stages or clinically unilateral disease. Patients with other neurological disorders, systemic disease (diabetes, hypertension), non-consenting absolute glaucoma with blind painful eyes and secondary glaucoma were excluded from the study.
All experiments detailed below were done in triplicates with robust selection of each case and control.
Peripheral Blood Mononuclear Cell Isolation and Total RNA Extraction
Blood samples from all patients recruited (n=17 PXF classical+pigmentary phenotype, n=11 OHT, n=11 PXG) and 11 controls (see supplemental data sheet) were collected in sodium-EDTA vials (around 3 mL) and mixed with equal volume of 1X phosphate-buffered saline (pH 7.2) after which it was overlaid with Ficoll (HiSepTm LSM 1077, HiMedia) (ratio 1:3) ensuring that they did not mix with each other. This was followed by centrifugation at 400 g for 30 minutes at 25°C for separation of serum and buffy coat layer. After careful separation of the supernatant layer, the mononuclear cell pellet was retrieved for further analysis.
Total RNA extraction from isolated PBMC was done with QIAzol® Lysis reagent (QIAGEN) followed by RNA integrity assessment on denaturing agarose gel. The homogenized mixture of 750 μL QIAzol and cells was mixed with 0.2 mL of chloroform followed by vigorous mixing for 15 seconds and centrifugation at 12,000 x g for 15 min at 4°C. The top aqueous phase was separated followed by precipitation with isopropanol and 5 µL of RNase free glycogen (5 µg) and centrifugation at 12,000 x g for 20 minutes at 4°C. This was followed by three washes with 70% ethanol and short centrifugation at 7500 x g for 5 min at 4°C. The RNA was briefly air dried for 5–10 minutes and eluted in 20 µL DEPC water. The purity and concentration of total RNA were determined by measurement of the OD260/280 ratio. Genomic DNA was eliminated from RNA which was then reverse transcribed using the First Strand Synthesis Kit (Qiagen) and subsequently stored at –20°C until further use for mRNA or miRNA expression.
miRNA Extraction and Differential Expression
400 ng of total RNA (containing miRNA) was reverse transcribed using the miScript II RT Kit (Qiagen). The kit contains 5x miScript HiSpec which facilitates the selective conversion of mature miRNAs into cDNA for subsequent mature miRNA profiling using miScript miRNA PCR Arrays (Qiagen, mifinder-MIHS-001Z miscript miRNA PCR array Human finder, miFibrosis- MIHS- 117Z miscript miRNA PCR array Human Fibrosis). The reverse-transcription master mix was prepared by mixing the following components: 4 µL 5x miScript HiSpec Buffer, 2 µL 10x miScript Nucleics Mix, 2 µL miScript Reverse Transcriptase Mix, required volumes of 400 ng total RNA. The volume was adjusted to 20 µL using RNase free water. The mixture was incubated at 60°C for 30 min followed by inactivation step of 5 min at 95°C in a PCR thermal cycler.
The prepared cDNA was diluted with 200 µL of RNase free water ready for real time PCR. The following components were mixed to prepare a reaction mixture for miScript miRNA PCR Arrays: 1375 µL 2x QuantiTect SYBR Green PCR Master Mix, 275 µL 10x miScript Universal Primer,1000 µL RNase-free water and 100 µL Template cDNA. 25 µL of this reaction mixture was added to each well of a 96 well array plate. The thermal cycling conditions used were as follows: Stage 1 (1 cycle): 95°C for 15 min, Stage 2 (40 cycles): 940C for 15 seconds, 55°C for 30 seconds and 70°C for 30 seconds.
For SYBR green based qPCR for differential expression, the thermal cycling conditions used were as follows: Stage 1 (1 cycle): 95°C for 15 min, Stage 2 (40 cycles): 94°C for 15 seconds, 55°C for 30 seconds and 70°C for 30 seconds. Web-based miScript miRNA PCR Array data analysis tool was used for the analysis of real-time PCR data. Once raw threshold cycle (CT) data are uploaded, the tool automatically performs fold-change calculations using the ΔΔCT method of relative quantification and presents the results in simple formats. Two wells of the PCR array plate contain replicate C. elegans miR-39 miScript primer assays that are used as an alternative normalizer for array data (Ce). Other wells contain an assay for a different snoRNA/snRNA that can be used as a normalization control for the array data (SN1=SNORD61 assay, SN2=SNORD68 assay, SN3=SNORD72 assay, SN4=SNORD95 assay, SN5=SNORD96A assay, SN6=RNU6B/RNU6-2 assay). There are also wells that contain replicate miRTC miScript primer assays (miRTC and replicate positive PCR controls (PPC). The miRbase (http://www.mirbase.org/) was used to verify known functions of the miRNA identified in this study. The predicted targets of the identified miRNA differentially expressed and KEGG pathway analysis was done using DIANA tools mirPathv3.1 (http://snf-515788.vm.okeanos.grnet.gr/) with targeted functions and molecular targets being predicted through online miRDG database (http://mirdb.org/).
Data obtained from each assay were deposited onto a public repository (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE135661).
Serum Enzyme Linked Immunosorbent Assay for TGF-β1
Since we wanted to evaluate the role of TGF in glaucoma pathogenesis we chose to study TGF levels in serum in eyes with (PXG) and without glaucoma (PXF). Serum was collected from the peripheral blood (n=5 in each group of PXF (PXF and PXG) compared to 5 controls) after being allowed to clot at room temperature for 10–15 minutes followed by centrifugation at 1000 g x 15 minutes. Transforming growth factor, TGF-β1, concentration was analyzed by colorimetric immunoassays using the Quantikine TGF-β1 Elisa kit (DB100B, R & D Systems). For this, each sample was incubated overnight with monoclonal anti-TGF-β1 antibody with gentle shaking at 4°C This was followed by incubation with 1X biotinylated secondary antibody for an hour at room temperature (RT) followed by incubation with 100 µL of streptavidin horseradish peroxidase for 45 min at RT. Color detection was enabled by adding 100 µL of 3,3ʹ,5,5ʹ-tetramethylbenzidine into each well for 30 min at RT followed by 50 µL of 0.2 M sulfuric acid to stop the reaction. Absorbance was measured on an ELISA microplate spectrophotometer (Epoch, Biotek, USA) set at 450 nm.
mRNA Expression of Transforming Growth Factor Related Predicted Targets
A fixed concentration 2.1 µg of total RNA from each PBMC sample was reverse transcribed using cDNA kit (RT2 first strand kit 330404, Qiagen) as per manufacturer’s protocol. Reverse transcription mix (10 µL) was mixed with 10 µL genomic DNA elimination mix and incubated at 42°C for 15 min, 95°C for 5 min in a thermal cycler. 91 µL of RNase free water was added to the reaction mixture (cDNA mix). The PCR components mix was prepared with 1350 µL 2x RT2 SYBRR Green Mastermix, 102 µL cDNA mix, 1248 µL RNase-free water. 25 µL of this reaction mixture was loaded onto each well of fibrosis array (PHAS-120ZRT2 Profiler array), (PHAS-035YCRT2 Profiler array) and unfolded protein response (PHAS-089YCZRT2 Profiler array). The thermal cycling conditions used for this reaction were 42° for 25 min followed by 95° for 5 minutes. The data obtained were subjected to robust analysis using the online Qiagen software to generate Ct values. Threshold and linear baseline correction were automatically calculated for each individual reaction which was used to calculate fold change (FC) as described previously.
Statistical Analysis
Statistical analysis was calculated using Stata Corp (version 10, USA). Student’s t-test was used to compare differences between two groups while Chi square was used to compare proportions between the groups with alpha error set at p<0.05. For clinical variables among different stages and phenotypes with controls, one way ANOVA or Kruskal–Wallis test was used for comparisons. Since the correlation between two eyes of a PXF patient is not proven, no adjustment was made for inter-eye similarities and each eye was considered as a separate unit.
Results
Of 94 patients screened in the study period, 50 patients with no systemic associations and fulfilling all inclusion criteria, were selected who were naïve to any form of medical therapy with a mean age of 60±9.9 years, Table 1. Three patients of each stage were taken into the discovery stage and the rest entered the validation phase wherein selected markers were validated using different assays as detailed previously (see supplemental data for detailed patient recruitment and flow of analysis). The patients with PXG were significantly older than earlier forms of PXF or controls, Table 1. PXG eyes also had higher baseline IOP and lower baseline visual field indices than earlier forms of PXF which were comparable with controls.
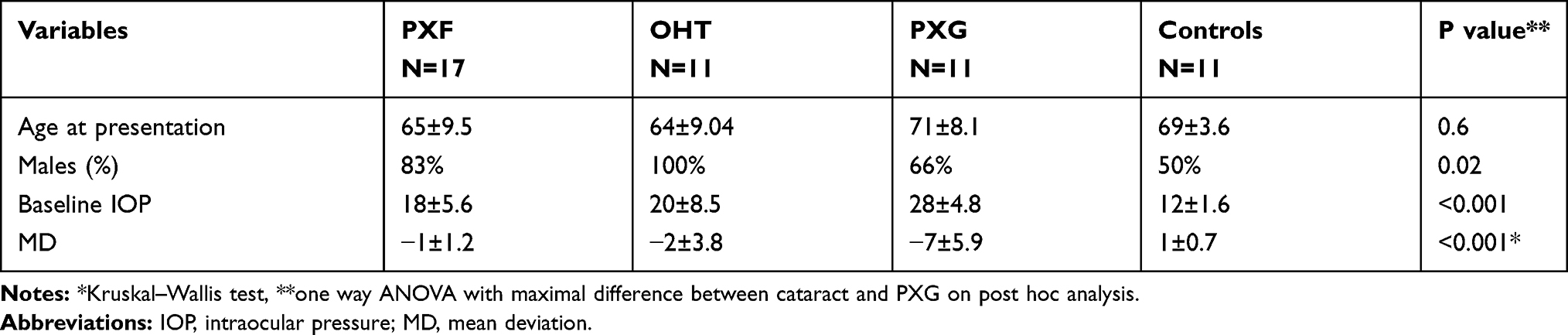 |
Table 1 Clinical Characteristics of Patients with Pseudoexfoliation of Different Stages Namely Pseudoexfoliation Syndrome, PXF with Ocular Hypertension (OHT) and Pseudoexfoliation Glaucoma (PXG) |
Table 1 gives the clinical details of the patients included. The total RNA obtained from each patient ranged from 398 ng/µL to 684 ng/µL which was not statistically different between the different groups, p=0.6.
miRNA Profile in PBMC in Pseudoexfoliation Stages
In the discovery phase, we first evaluated novel miRNA differentially expressed in the different stages of pseudoexfoliation in 3 patients of each group including controls, PXF, OHT (moderate stage) and pseudoexfoliation glaucoma (advanced stage). We also compared the expression in controls and different phenotypes of PXF (classical and pigmentary) to see how these phenotypes differ from controls.
It was observed that 12 of 84 miRNA were significantly downregulated in OHT compared to PXG stage where these were significantly upregulated in PXG, Figure 1A, Table 2 (see https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE135661 for list of all miRNA).
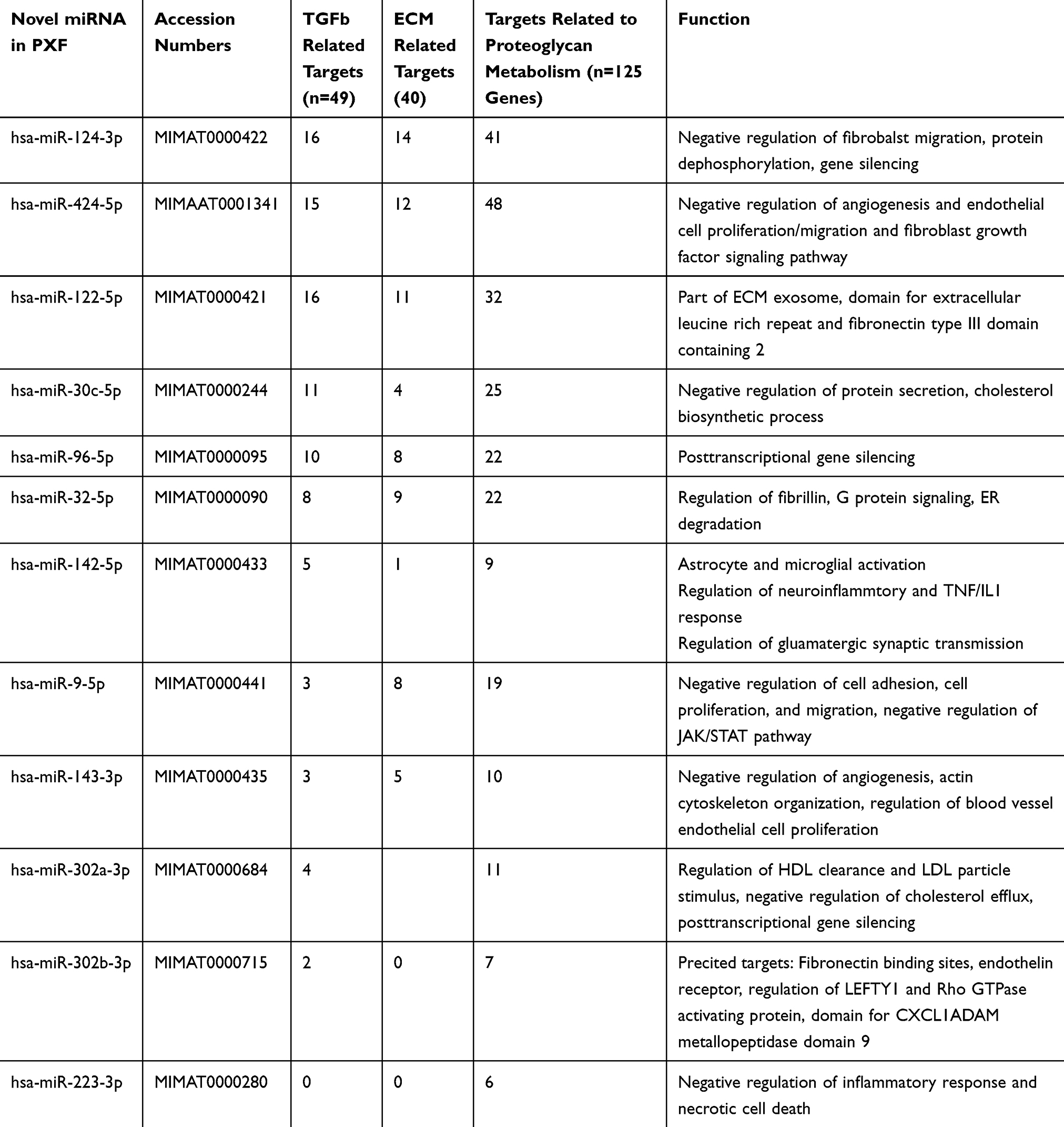 |
Table 2 Novel miRNA Identified with >2.5 Fold Differential Expression Between Different Pseudoexfoliation Stages with Their Predicted Targets and Functions Using DIANA mirPath, See Text for Detailed Description) |
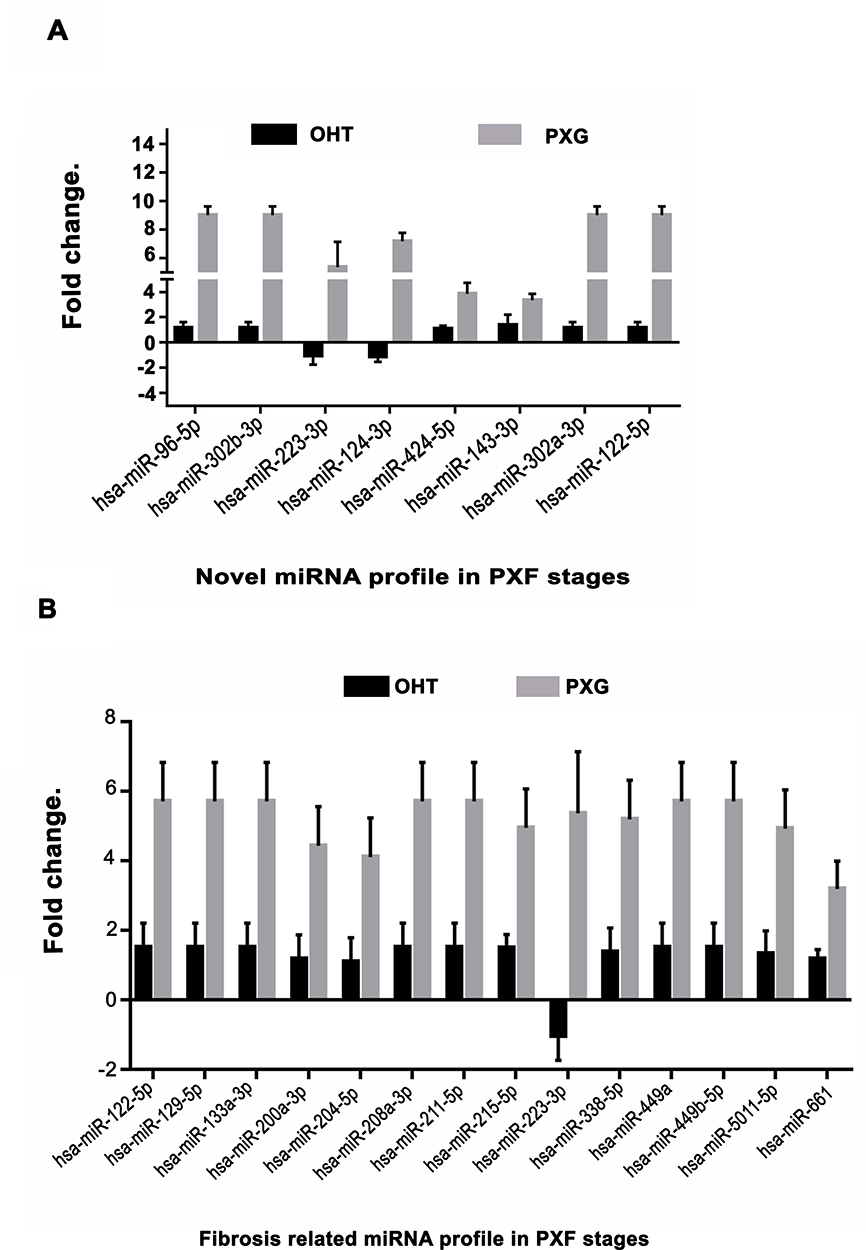 |
Figure 1 (A) Novel miRNA identified in discovery stage showing >2.5 fold differential expression (out of a total 12 of 84 significantly different between groups, see text for detailed description) in PXF stages. (B) Fibrosis related miRNA identified to be >2.5 fold differentially expressed (out of 20 miRNA identified by array, see text for detailed description) in pseudoexfoliation stages (see (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE135661 for list of all miRNA). |
Comparing phenotypes of PXF (classical and pigmentary) with controls, hsa-miR-l44-3p showed differential down-regulation in pigmentary phenotype of PXF, Table S1 and Figure S1.
We analyzed which of the KEGG pathways were enriched with the selected 12 significantly differentially expressed miRNA which revealed maximal involvement of proteoglycan metabolism, ECM interaction and TGF related pathways, Table 2. Of these, 3 miRNA including hsa-miR-124-3p, hsa-miR-424-5p and hsa-miR-122-5p were involved in all three pathways with maximum targeted interaction with fibrosis, MMP9 and TGF related genes including TGFR1 and SMAD2/1. It was also seen that hsa-miR-424-5p was closely linked to TGF pathway with targeted action on SMAD, activin (ACVR) receptors apart from interaction with integrins, Table 2. Among phenotypes, all 3 miRNA identified to have different expression among phenotypes were closely related to TGF pathway with targeted interaction with TGFR1, TGFR2, LTBP1 and SMAD2, Table S1. Of these, hsa-let-7e-5p and hsa-miR-26b-5p had direct targeted interaction with TGFR1 and TGR2 part from SMAD2.
Since most miRNA significantly differentially expressed between stages and phenotypes belonged to TGF-β1 and fibrosis/proteoglycan metabolism, we next evaluated miRNA expression known to be involved in fibrosis on a separate cohort and also evaluated TGF/fibrosis related genes in other cohorts.
Validation of PBMC miRNA Related to Fibrosis/TGF Pathway
The miRNAs related to fibrosis were validated in various disease stages in triplicate experiments which confirmed the role of hsa-miR-122-5p which was upregulated in PXG, Figure 1B, Table 3. We further found a total of 20 other miRNAs related to fibrosis or TGF-β1pathway, which were significantly upregulated in later stages of PXG (see https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE135661 for list of all miRNA). Phenotype comparisons with fibrosis related miRNA gave similar results with hsa-miR-19a-3p and hsa-miR-30a-5p related to proteoglycans being significantly downregulated in OHT compared to PXG, Figure S1B,C and Table S2.
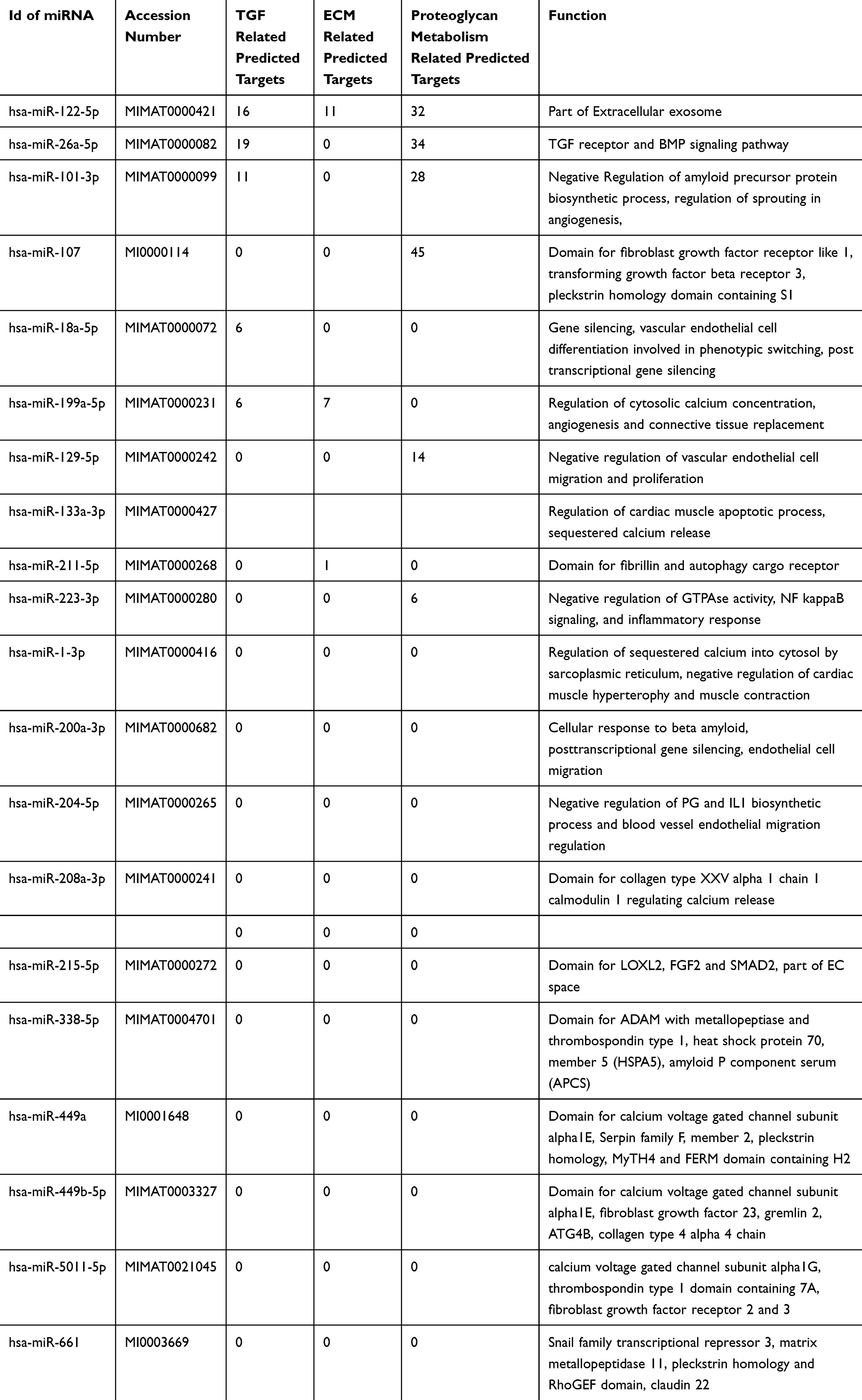 |
Table 3 Novel miRNA Related to Fibrosis Identified with >2.5 Fold Differential Expression Between Different Pseudoexfoliation Stages with Their Predicted Targets Using mirPath (See Text for Detailed Description) |
In summary, miRNA hsa-miR-122-5p was found to be significantly upregulated as PXF progressed into later stages in both discovery and validation phases.
Transforming Growth Factor in Different Pseudoexfoliation Stages and Glaucoma
Levels of TGF-β1 in serum measured by ELISA showed significant up-regulation in PXG eyes with maximal differences between PXG and controls or PXF, Figure 2A. The earlier forms like classical and pigmentary PXF had expression of TGF-β1 comparable to that of controls.
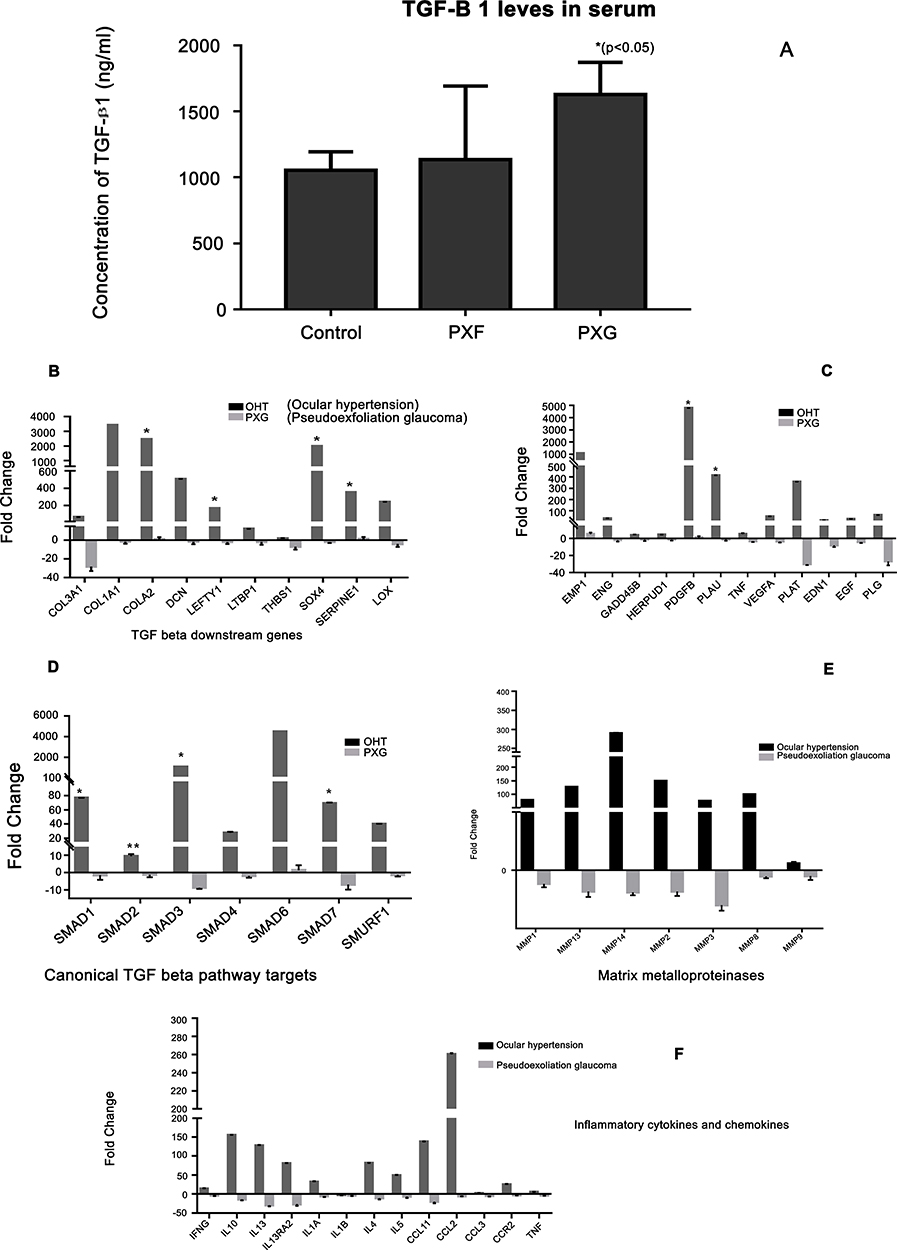 |
Figure 2 (A) Serum TGF-B1 levels measured by ELISA in eyes with pseudoexfoliation without (PXF) and with glaucoma (PXG) compared to controls (asterix indicates the groups which were significantly different from other groups). (B and C) show downstream effectors of the TGF pathway differentially expressed among ocular hypertension (OHT) and PXG compared to PXF, canonical pathway targets like SMADs (D), levels of matrix metalloproteinase (E) in serum and inflammatory cytokines/chemokines (F) detected in different pseudoexfoliation stages. |
Comparing the TGF related downstream gene expression in different disease stages, 71 of 84 genes seemed to be up-regulated in OHT compared to PXG where they were down-regulated with statistically differential expression of COL1A1, COL1A2 which play an important role in collagen synthesis, Figure 2B and C. These also included genes like HERPUD1, ENG and ENDN which were downregulated in PXG. Notably, LOXL1 was found to be down-regulated in advanced stages of the diseases in this study. This pattern of upregulation of genes in OHT stages and down-regulation in PXG was also followed by genes regulating ECM remodeling and fiber assembly like ACTA2 (α-smooth muscle actin), ECM degradation like MMPs, inflammatory cytokine and chemokines associated with TGF and pro-fibrotic molecules, Figure 2(B-F).
Since TGF-β1 regulates downstream processes through both canonical and non-canonical pathways, it was important to know the predominant pathway mediating the downstream effects which are differentially involved in different PXF stages. Most prominent was recruitment of the canonical pathways including SMAD1, SMAD3 and SMAD 6 which were up-regulated in OHT and downregulated in PXG correlating with downstream effects suggesting the role of these molecules in causing downstream effects, Figure 2D. This also correlated with significant upregulation of inflammatory cytokines and chemokines in OHT stage possibly mediated by TGF-β1, Figure 2F. The MMP’s downregulation in PXG compared to OHT stage contrasted with increasing TGF levels in serum in PXG compared to OHT, Figure 2E. Surprisingly, TIMP levels, regulating the MMP activity with the TIMP/MMP ratio, were also up-regulated suggesting alternate regulation of MMPs in PXF. Also LTBP1 and LOXL1, believed to be involved in fiber assembly in the ECM, were clearly downregulated in PXG while being significantly up-regulated in OHT stage. Comparing UPR response elements, most genes were downregulated in all stages of PXF clearly indicating the down-regulation of the protein unfolding response related machinery as the key mechanism of protein aggregate formation in PXF, Figure 3. This included members of the ERAD family like VIMP, ELF2AK3 and members of the ER folding quality control like PDIA3 and GANAB. Also seen was that the most of the ERAD response elements were downregulated significantly in the PXG stage compared to OHT stage reflecting again that these regulatory controls over protein folding also get compromised as disease progresses.
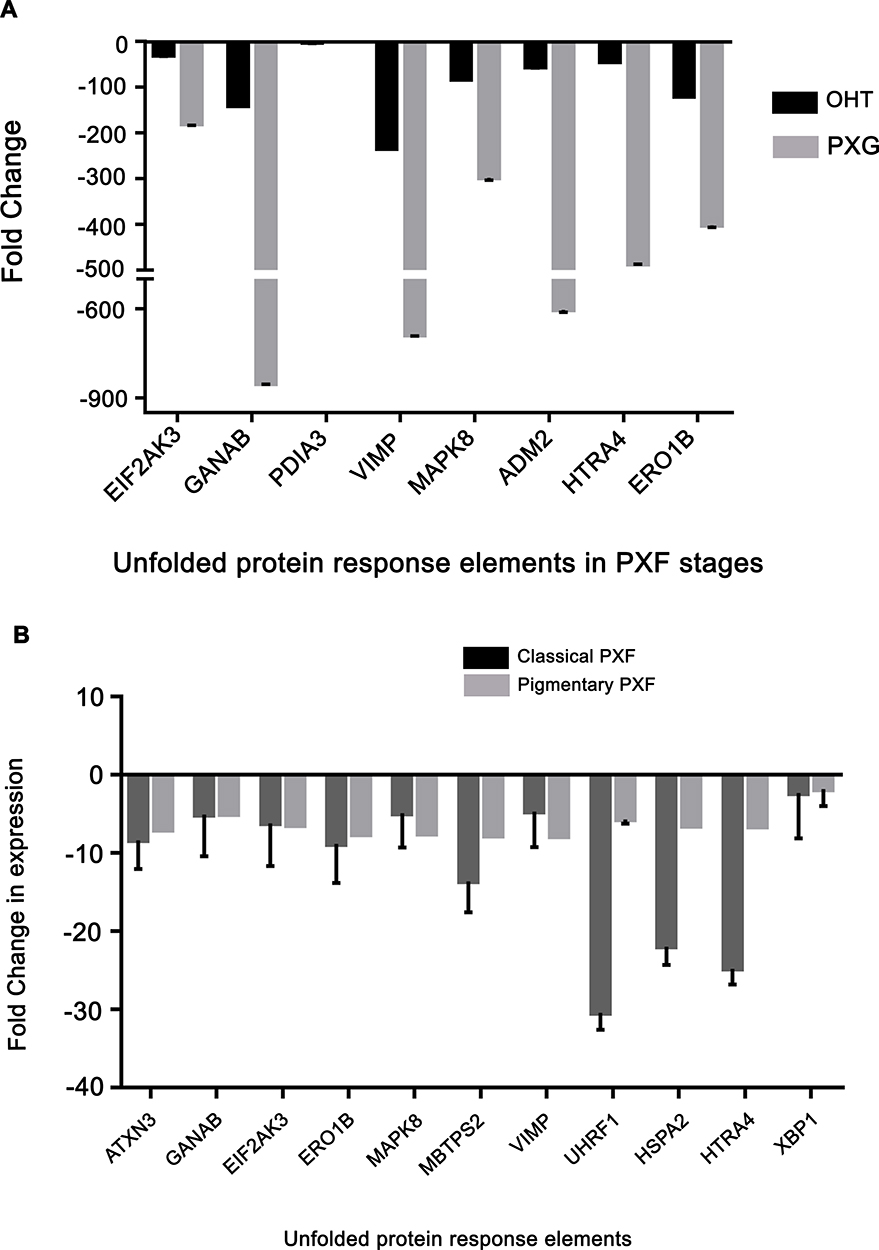 |
Figure 3 Genes related to unfolded protein response (UPR) detected to be differentially expressed in pseudoexfoliation between stages like eyes with ocular hypertension (OHT) and those with glaucoma, PXG (A) and between phenotypes like classical and pigmentary forms (B). |
Comparing differences in disease phenotypes, TGF related molecules including COL1A1, COL1A2 and DCN were significantly up-regulated in classical form while comparable to levels of controls in pigmentary form predominantly through SMAD3, a canonical pathway marker, Figure S2. Comparing fibrotic genes, we found MMPs being significantly up-regulated in classical PXF form while being comparable to that of controls in the pigmentary form. The UPR response genes were universally downregulated suggesting the predominant mechanisms of TGF mediated protein aggregate formation in PXF, Figure 3B.
Discussion
In the present study, we identified TGF-β1 to be significantly up-regulated as disease progressed to later stages. This elevation in advanced stages was associated with differential expression of downstream genes in OHT compared to PXG predominantly through the SMAD3, a canonical pathway marker. These suggest that downstream effectors are differentially regulated by TGF-β1 as the disease progresses from OHT to PXG with a switch from canonical SMAD3 mediated effects in OHT to other alternative pathways in PXG. The structural or functional disturbance in the TM causing PXG reflects an end result (not necessarily a different pathophysiology in PXG compared to OHT) suggesting that interventions directed toward reversal of the changes at the OHT stage may help prevent onset of PXG.
We found several miRNA specific to 3 pathways namely TGF- β1, fibrosis/ECM, and proteoglycan metabolism with common effectors like SMAD/3/2 which are key for TGF mediated regulation of fibrotic genes in PXF and regulating genes of the unfolded protein response in PXF. Of all miRNA identified in PXF, hsa-miR-124-3p, hsa-miR-424-5p, and hsa-miR-122-5p were seen to be controlling TGF-β1, protein binding and ECM related processes all of which were downregulated in PXG and therefore may be biomarkers of disease progression to glaucoma in any eye with unilateral cases or clinically unmanifest disease. The UPR response genes were significantly downregulated in all stages and phenotypes of PXF suggesting this as the key mechanism for protein aggregates in PXF syndrome.
miRNAs are small circulatory RNA sequences which regulate several cell processes and tissue functions.8,10–15 We found several novel miRNA, notably, hsa-miR-122-5p, controlling TGF and ECM related cell processes differentially expressed in PXF stages which has not been reported in ocular disorders like glaucoma earlier. These miRNA like hsa-miR-122-5p may act like switches that trigger differential downstream processes despite elevated TGF levels by post-transcriptional gene modifications in the tissues like TM marking the onset of glaucoma or disease progression. There is only one study which implicates hsa-miR-122-5p in glaucoma in regulation of TGF and Optineurin pathway, both of which are genes associated with glaucoma.15 Yet the authors in this study only identified miRNA expression in exfoliation and primary glaucoma while not studying expression in different stages of PXF. Further, miRNA-124 is known to be expressed highly in the brain and is thought to have a role in neural development.16–18 It is now believed to have a potential role in Alzheimer’s and Parkinson’s disease with a possible therapeutic potential in these diseases. Though miRNA-124 and miRNA-424 have been reported to be repressed in brain tumors or other tumors, their role in the human eye remains unexplored.18 The specific association of miRNA-124 with PXF or PXG has not been reported earlier. The hsa-miR-124-3p is also known to be a regulator of autophagy related genes like Beclin-1.19 Peripheral blood hsa-miR-122-5p correlated with altered liver enzyme levels in HCV infected patients and has also been reported to be involved in lipoprotein metabolism in fatty liver disease.18,20 This study found that these specific miRNA were found to be upregulated in OHT stage of PXF compared to glaucoma through common target being SMAD 2/3 and TGFR1 and TGFR2, which suggests these miRNAs regulate TGF in PXF differently via possible double feedback loops acting differently in different disease stages.21,22 Since these miRNA regulate TGF/ECM interactions, EMT as well as autophagy related debris clearing directly, these can serve as therapeutic targets for reversing/preventing the fibrotic changes in the TM and possibly clear off the complex aggregates blocking the inter-trabecular spaces. We chose to focus on PBMC related signatures since TGF related signature controlling cell processes would logically be expected to be more active in cells in the peripheral blood than in serum as seen in this study. Further credence to this theory was lent by our preliminary experiments in miRNA expression in serum and PBMC which showed more consistent results in the latter, Figure S3. Yet, TM changes may not mirror the peripheral blood changes in a similar way in all eyes which needs to be explored with longitudinal studies.
The exfoliative material in PXF is comprised of complex protein and carbohydrate aggregates which accumulate owing to defective cell machinery or response elements. The UPR response genes like XBP were significantly downregulated in all stages and phenotypes of the disease. The UPR response is activated when the endoplasmic reticulum (ER) is overwhelmed by unfolded/misfolded proteins in the cells causing ER stress activating a chain of complex cascade of reactions, in general called the unfolded protein response.23,24 The IRE1α-XBP-1s pathway targets genes that increase ER protein-folding capacity and facilitate degradation of misfolded proteins through ER-associated protein degradation, ERAD.23 Downregulation of the UPR genes causes misfolded proteins to aggregate in the cells rather than being cleared off which may cause cell death.
Previous studies have shown that microRNA (miR)-214 can negatively regulate XBP1 expression and is down-regulated during ER stress.24 The UPR response is believed to play a crucial role in angiogenesis and endothelial cell activity through pro-angiogenic factors like VEGF. The role of miRNA in regulating TGF is known in cancer.25,26 This study highlighted the possible role of specific miRNA and TGF in regulating fibrosis and UPR response elements in PXF stages or glaucoma pathogenesis. It is possible that altered regulation of the UPR response related genes by TGF with parallel cross talk with VEGF may cause vascular disturbances and ischemic associations seen in PXF. This may also explain the association of PXF with vascular disease like hypertension or retinal vaso-occlusive disorders, with common risk factors of ischemia and altered endothelial permeability, known to be regulated by TGF-β1.2 Studies evaluating how miRNA can rescue the regulation of the TGF mediated UPR response may be a potential therapeutic target in PXF. The double feedback loops between specific miRNAs identified in this study and TGF in mediating downstream functions including angiogenesis, UPR and fibrosis may be key in reversing changes before onset of glaucoma in PXF.
PXF is an age related fibrillopathy characterized by abnormal ECM production, aberrant ECM degradation and collagen homeostasis.2,5,9 So it is not surprising that related genes like COL1A1, COL1A2, DCN were found to be up-regulated in PXF despite significant upregulation of serum TGF levels in PXG stage. Genes related to plasminogen activity and endothelial cell permeability like HERPUD, ENDN, EGN and VEGF were significantly upregulated in OHT stages in PXF which may explain the association of vein occlusive disease in the eye and ischemic associations like transient ischemic attacks and ischemic heart disease seen in patients with PXF. T HERPUD is a homocysteine responsive element which increases the levels of beta-amyloid protein by both activating arms of the unfolded protein response and ERAD suggesting that this may be involved in the formation of complex protein aggregates found in PXG. This suggests an underlying refractoriness to elevated TGF serum levels which lead to the changes typical of PXG. It also may be possible that changes in PXG may actually be as a result of exhaustion of compensatory mechanisms that are upregulated in the OHT stage. However, parallel downregulation of genes like Noggin which interfere with receptor activation in the TGF pathway in the advanced stages hints at a possible excessive receptor activation or saturation which may lead to a refractoriness of downstream targets as seen in this study. This is partly called as the TGF paradox which explains the difference between benign and malignant neoplasm and is also believed to play a role in increased cell survival and spread of cancer cells.27 It is likely that the TGF paradox drives differential downstream effects causing onset of raised IOP or later glaucoma in later disease stages despite elevated TGF levels between stages compared to controls. In our earlier study, we found decreased MMP9 activity in advanced stages compared to earlier stages of adult glaucoma which is similar to the results of this study.6 The reduced levels of MMPs reflect decrease in ECM degradation mechanisms as the disease progresses which explains formation of larger protein complex aggregates or gradual structural/functional disturbances in tissue where aggregates form. This was associated with no parallel changes in TIMP levels which reflect alternate control of MMP activity by TGF by other molecules. The results are again explained by the TGF paradox which also may be overlaid by exhaustion of ECM degradation by MMP in very advanced stages. We believe that this paradox is driven by the specific peripheral blood miRNAs identified in this study that trigger differential responses of TGF pathway leading to irreversible changes like TM fibrosis, raised IOP in OHT and glaucoma.
Of a large number screened, we selected patients with strict inclusion criteria and took medically naïve patients with no systemic or ocular associations to avoid confounding effects of other disease and medications on gene expression patterns. This however does not reflect routine clinical scenarios where several associations may cause difficulty in generalizing these genes or miRNA as markers for disease or stage transition. The sample size selected for each set of experiments in this study was small owing to stringent strictly defined criteria. Therefore despite statistical significance for several miRNAs in disease stages, caution needs to be exercised while generalizing results or before direct translation into clinical practice. Also this was a cross sectional study capturing different patients in different stages; we understand that a longitudinal study of patients transitioning from one stage or phenotype to the other may truly reflect the differential involvement of these miRNA beyond doubt. We also acknowledge that all patients were of Indian Asian origin. Ethnic differences in gene expression in PXF cause difficulty in direct application of miRNA profiles reported in this study to other ethnicities.
Abbreviations
Refer to appendix for full names of all genes studied.
Data Sharing Statement
Data deposited in NCBI GEO website with accession id GSE135661 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE135661).
Funding
The study was funded by the DBT wellcome Trust India alliance grant: ref no IA/CPHI/15/1/502031.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ekström C, Alm A. Pseudoexfoliation as a risk factor for prevalent open-angle glaucoma. Acta Ophthalmol. 2008;86:741–746. doi:10.1111/j.1755-3768.2008.01248.x
2. Aboobakar IF, Johnson WM, Stamer WD, Hauser MA, Allingham RR. Exfoliation syndrome; advances in disease genetics, molecular biology, and epidemiology. Exp Eye Res. 2017;154:88–103. doi:10.1016/j.exer.2016.11.011
3. Jelodari-Mamaghani S, Haji-Seyed-Javadi R, Suri F, et al. Contribution of the latent transforming growth factor-β binding protein 2 gene to etiology of primary open angle glaucoma and pseudoexfoliation syndrome. Mol Vis. 2013;19:333–347.
4. Ghavami S, Yeganeh B, Zeki AA, et al. Autophagy and the unfolded protein response promote profibrotic effects of TGF-β1 in human lung fibroblasts. Am J Physiol Lung Cell Mol Physiol. 2018;314:L493–L504. doi:10.1152/ajplung.00372.2017
5. Vranka JA, Kelley MJ, Acott TS, Keller KE. Extracellular matrix in the trabecular meshwork: intraocular pressure regulation and dysregulation in glaucoma. Exp Eye Res. 2015;133:112–125. doi:10.1016/j.exer.2014.07.014
6. Sahay P, Rao A, Padhy D, et al. Functional activity of matrix metalloproteinases 2 and 9 in tears of patients with glaucoma. Invest Ophthalmol Vis Sci. 2017;58:BIO106–BIO113. doi:10.1167/iovs.17-21723
7. Budi EH, Duan D, Derynck R. Transforming growth factor-β receptors and smads: regulatory complexity and functional versatility. Trends Cell Biol. 2017;27:658–672. doi:10.1016/j.tcb.2017.04.005
8. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi:10.1016/S0092-8674(04)00045-5
9. Jayaram H, Cepurna WO, Johnson EC, Morrison JC. MicroRNA expression in the glaucomatous retina. Invest Ophthalmol Vis Sci. 2015;56:7971–7982. doi:10.1167/iovs.15-18088
10. Rao A, Padhy D, Sahay P, et al. Clinical spectrum of pseudoexfoliation syndrome-an electronic records audit. PLoS One. 2017;12(10):e0185373. doi:doi:10.1371/journal.pone.0185373.
11. Ghanbari M, Iglesias AI, Springelkamp H, et al. International Glaucoma Genetics Consortium (IGGC). A genome-wide scan for MicroRNA-related genetic variants associated with primary open-angle glaucoma. Invest Ophthalmol Vis Sci. 2017;58:5368–5377. doi:10.1167/iovs.17-22410
12. Hindle AG, Thoonen R, Jasien JV, et al. Identification of candidate miRNA biomarkers for glaucoma. Invest Ophthalmol Vis Sci. 2019;60:134–146. doi:10.1167/iovs.18-24878
13. Molasy M, Walczak A, Szaflik J, Szaflik JP, Majsterek I. MicroRNAs in glaucoma and neurodegenerative diseases. J Hum Genet. 2017;62:105–112. doi:10.1038/jhg.2016.91
14. Liu Y, Chen Y, Wang Y, et al. microRNA profiling in glaucoma eyes with varying degrees of optic neuropathy by using next-generation sequencing. Invest Ophthalmol Vis Sci. 2018;59:2955–2966. doi:10.1167/iovs.17-23599
15. Drewry MD, Challa P, Kuchtey JG, et al. Differentially expressed microRNAs in the aqueous humor of patients with exfoliation glaucoma or primary open-angle glaucoma. Hum Mol Genet. 2018;27:1263–1275. doi:10.1093/hmg/ddy040
16. Jiao S, Liu Y, Yao Y, Teng J. miR-124 promotes proliferation and neural differentiation of neural stem cells through targeting DACT1 and activating Wnt/β-catenin pathways. Mol Cell Biochem. 2018;449:305–314. doi:10.1007/s11010-018-3367-z
17. He Y, Li HB, Li X, Zhou Y, Xia XB, Song WT. MiR-124 promotes the growth of retinal ganglion cells derived from müller cells. Cell Physiol Biochem. 2018;45:973–983.
18. Recabarren D, Alarcón M. Gene networks in neurodegenerative disorders. Life Sci. 2017;183:83–97. doi:10.1016/j.lfs.2017.06.009
19. Gozuacik D, Akkoc Y, Ozturk DG, Kocak M. Autophagy regulating microRNAs and cancer. FrontOncol. 2017;7:65. doi:10.3389/fonc.2017.00065.
20. Raitoharju E, Seppala I, Lyytikainen LP, et al. Blood hsa-miR-122-5p and hsa-miR-885-5p levels associate with fatty liver and related lipoprotein metabolism—The Young Finns Study. Sci Rep. 2016;6:38262. doi:10.1038/srep38262
21. Deng M, Hou SY, Tong BD, Yin JY, Xiong W. The Smad2/3/4 complex binds miR-139 promoter to modulate TGFβ-induced proliferation and activation of human Tenon’s capsule fibroblasts through the Wnt pathway. J Cell Physiol. 2019;2019:1–11. doi:doi.10.1002/jcp.28011.
22. Baptista R, Marques C, Catarino S, et al. MicroRNA-424(322) as a new marker of disease progression in pulmonary arterial hypertension and its role in right ventricular hypertrophy by targeting SMURF1. Cardiovasc Res. 2018;114:53–64. doi:10.1093/cvr/cvx187
23. Adams CJ, Kopp MC, Larburu N, Nowak PR, Ali MMU. Structure and molecular mechanism of ER stress signaling by the unfolded protein response signal activator IRE1. Front Mol Biosci. 2019;6:11.
24. Duan Q, Wang X, Gong W, et al. ER stress negatively modulates the expression of the miR-199a/214 cluster to regulates tumor survival and progression in human hepatocellular cancer. PLoS One. 2012;7(2):e31518. doi:10.1371/journal.pone.0031518.
25. Suzuki HI. MicroRNA Control of TGF-β Signaling. Int J Mol Sci. 2018;19:1901. doi:10.3390/ijms19071901
26. Moradi-Marjaneh R, Khazaei M, Ferns GA, Aghaee-Bakhtiari SH. The role of TGF-β signaling regulatory MicroRNAs in the pathogenesis of colorectal cancer. Curr Pharm Des. 2018;24:4611–4618. doi:10.2174/1381612825666190110150705
27. Zhang Q, Yu N, Lee C. Mysteries of TGF-β paradox in benign and malignant cells. Front Oncol. 2014;4:94. doi:10.3389/fonc.2014.00094
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.