Back to Journals » Journal of Inflammation Research » Volume 13
Differential Immune Activating, Anti-Inflammatory, and Regenerative Properties of the Aqueous, Ethanol, and Solid Fractions of a Medicinal Mushroom Blend
Authors Davis R ![]() , Taylor A
, Taylor A ![]() , Nally R
, Nally R ![]() , Benson KF
, Benson KF ![]() , Stamets P
, Stamets P ![]() , Jensen GS
, Jensen GS ![]()
Received 1 September 2019
Accepted for publication 13 February 2020
Published 25 February 2020 Volume 2020:13 Pages 117—131
DOI https://doi.org/10.2147/JIR.S229446
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Renee Davis,1 Alex Taylor,1 Regan Nally,1 Kathleen F Benson,2 Paul Stamets,1 Gitte S Jensen2
1Fungi Perfecti, Olympia, WA 98507, USA; 2NIS Labs, Klamath Falls, OR 97601, USA
Correspondence: Gitte S Jensen
NIS Labs, 1437 Esplanade, Klamath Falls, OR 97601, USA
Tel +1 541 884-0112
Fax +1403 441-5236
Email [email protected]
Purpose: To compare three fractions of a medicinal mushroom blend (MMB), MyCommunity, on immune-activation, inflammation-regulation, and induction of biomarkers involved in regenerative functions.
Methods: A seventeen-species MMB was sequentially extracted: first, saline solution at ambient temperature, followed by re-extraction of the solids in ethanol, and finally resuspension of the homogenized ethanol-insoluble solids in cell-culture media. Fractions were tested on peripheral blood mononuclear cells from three healthy donors. Immunostaining, flow-cytometry, and Luminex protein-arrays measured immune-cell activation and cytokine response. Dose-responses for induction of the CD69 early activation marker and individual cytokine and growth-factor responses for each donor were evaluated. The CD69 and the combined cytokine and growth-factor results were subjected to Non-metric Multidimensional Scaling (NMDS) and multivariate ordination to aid interpretation of the aggregate immune response and pairwise permutational MANOVA on a distance-matrix to evaluate statistical differences between treatments on pooled data from all donors.
Results: Differential effects were induced by water-soluble, ethanol-soluble, and insoluble immunomodulatory compounds of the MMB. The aqueous and ethanol fractions upregulated expression of CD69 on all tested cell types. Monocyte-activation was correlated with the ethanol fraction, while NKT and non-NK non-T cell-activation was more closely correlated with the aqueous fraction. The solid fraction was the most potent inducer of Tumor Necrosis Factor-α, as well as the anti-viral cytokines interferon-γ, MCP-1 (CCL-2), MIP-1α (CCL-3), and MIP-1β (CCL-4), and induced G-CSF and b-FGF—growth-factors involved in regenerative functions—and the anti-inflammatory cytokine IL-1ra.
Conclusion: The aqueous, ethanol, and insoluble compounds within MMB induced differential immune-activating, anti-inflammatory, and regenerative effects. This in vitro data suggests that, upon consumption, MMB may induce a concerted series of immunomodulatory events based on the differential solubility and bioavailability of the active constituents. These differential responses support both immune-activation and resolution of the host defense-induced inflammatory reactions, thus assisting a post-response return to homeostasis.
Keywords: CD69, cytokines, growth factors, anti-viral peptides, medicinal mushrooms, mycelium, immune support, redundancy analysis, rda, multivariate ordination
Introduction
Mushrooms have been embraced for centuries due to their nutritional and medicinal properties. They have been historically used in the treatment of infectious disease, gastrointestinal disorders and asthmatic conditions, as well as to support overall wellbeing.1 Fungi now occupy their own kingdom, but they were once considered plants due to their resemblance and root-like structures. One of many characteristics that separate fungal from plant organisms is the cell wall structure. The cell walls of fungi contain chitin,2 a modified form of the polysaccharide cellulose. Chitin is comprised of β-(1→4)-linked N-acetylglucosamine monomers, whereas cellulose is comprised of β-(1→4)-linked glucose units. Chitin degrades into a mixture of shorter-chained polysaccharides along with monosaccharide products.3,4 This degradation can occur with a variety of processing techniques that implement heat and drying.5
Mushroom polysaccharides possess documented immunomodulatory properties, specifically through the activation of natural killer cells, macrophages, and neutrophils, as well as induction of innate immune cytokines and interleukins.5 β-glucans are another class of polymers present in the cell walls of fungi. The generic term β-glucan refers to the polymeric form of glucose residues connected by β-(1→3), β-(1→4), and β-(1→6)-linkages. The type of β-glucans isolated from fungi consist mainly of a linear backbone of β-(1→3) glucose monomers and side branches comprised β-(1→3) and β-(1→6)-linked oligosaccharides.6
The most widely studied β-glucans are comprised of (1→3)-β, and (1,6)-β linkages, which exhibit immunostimulatory and antitumor properties.7,8 These polysaccharides are ligands for the dectin-1 and toll-like receptor 2 (TLR-2) receptor systems expressed on macrophages and dendritic cells, inducing NK cells, neutrophils, T-cells, B-cells, as well as TNF-a, IL-4, and IL-6 signaling.9 The Complement Receptor-3 (CD11b/CD18) in context of extracellular matrix is also involved in immune responses to fungal β-glucans.10 Research by Quayle et al demonstrates that other structural components in the fungal cell wall matrix affect pattern recognition receptor activity.11 The TLR-2 activity of polysaccharide-K (PSK) was reduced by 81% upon treatment with lipoprotein lipase, revealing the biological activity of a previously unreported lipid in the PSK complex. Mushroom β-glucans may contain other functional groups that contribute additional biological effects, particularly regarding antioxidant activity.12 Mushroom polysaccharide and β-glucan products are common dietary supplements, often available as whole mushrooms, mycelial powders, dried or liquid extracts from fruitbodies or from mycelium.
Specialized protein-bound polysaccharide products have been developed as adjuvant immunotherapies in oncological clinical settings. Polysaccharide-K (PSK, Krestin®, Kureha) and polysaccharide peptide (PSP) are produced from the cultured mycelium of the turkey tail mushroom (Trametes versicolor) and are used as antineoplastic agents and immunostimulants in China and Japan.13–18 Lentinan is a polysaccharide extracted from the mycelium and fruiting body of the shiitake mushroom (Lentinula edodes),19 and has demonstrated efficacy as a biological response modifier and chemotherapy adjuvant in gastrointestinal and lung cancers.20 Grifolan is another polysaccharide extracted from maitake (Grifola frondosa) which enhances production of IL-6, IL-1, and TNF-a.21
Another class of immunologically active compounds from mushrooms, which are typically much smaller in size compared to cell-wall polymers, are secondary metabolites. These are compounds that may not be absolutely required for the growth of the organism but assist in its survival by offering protection and communication among other important but not metabolically essential functions. When consumed, secondary metabolites are associated with myriad biological effects, including antioxidant, anti-viral, anti-inflammatory, neuroregenerative, and hepatoprotective effects. Classes of fungal metabolites with medical significance include sterols, terpenes, and phenols, notable examples of which include ganoderic acid in Ganoderma lucidum,22 erinacines in Hericium erinaceus,23–25 and betulin in Inonotus obliquus,26 and cordycepin in Cordyceps militaris.27
A growing body of international scientific and medical research continues to help define the precise biochemical pathways leading to improved physiological outcomes. Cordycepin, 3ʹ-deoxyadenosine, has shown very potent anti-inflammatory effects in a spectrum of in vitro and animal models, specifically via effects on adipose-derived mesenchymal stem cells, where higher doses helped maintain the stem-ness of the cells and lower doses supported osteogenic differentiation.28 Isolated compounds from reishi (Ganoderma lucidum) were tested on the MCF-7 breast cancer cell line and its non-transformed counterpart MCF10A and showed selective killing of the transformed cells, both actively growing, and quiescent slow-cycling cancer stem cells.29 Polysaccharides from reishi30 and maitake31 have also been shown to promote and enhance the survival/renewal abilities of primitive hematopoietic stem/progenitor cells. For maitake, this was specifically linked to β-glucan-mediated increases in the production of granulocyte-colony stimulating factor.32 Notably, this research demonstrated that this effect supported stem cell transplantation in a NOD/SCID mouse model. To the best of our knowledge, non-β-glucan fractions of maitake were not previously evaluated in stem cell related studies.
While much prior research on medicinal mushrooms has focused on the solid, β-glucan-rich fraction, and β-glucan-mediated responses are clearly important, focusing on this compound class in isolation clearly does not reflect the overall bioactivity of a complex blend when consumed for immune support. Emerging evidence suggests that a blend of mushrooms may provide additive or synergistic effects on the host immune response. Preclinical work on a 7-mushroom blend (Ganoderma resinaceum, Cordyceps sinensis, Trametes versicolor, Hericium erinaceus, Inonotus obliquus, Grifola frondosa, Agaricus brasiliensis f. blazei, Phellinus linteus) discovered greater NK cell upregulation with a blend over any one isolated species.33 Findings from another study suggest synergistic radical scavenging activity with a combination of Boletus edulis and Marasmius oreades mushroom extracts.34
The purpose of this study was to investigate the differential immunological effects of aqueous, ethanol, and solid fractions of MMB in vitro, to determine the differences in biological activity between the soluble and insoluble fractions of a complex medicinal mushroom blend (MMB), a blend of 17 mushroom species (Table 1) that is used by consumers for seasonal immune wellness, and by clinicians to enhance innate immunity. The study of a complex blend rather than isolated single compounds was of importance. The experimental model included effects on multiple immune cell types in vitro, because of parallels to events in the gut mucosal tissue where dendritic cells and macrophages sample antigens in the gut lumen, and present to lymphocytes in the gut-associated mucosal tissue, leading to local immune cell activation, and cytokine secretion that has systemic effects.
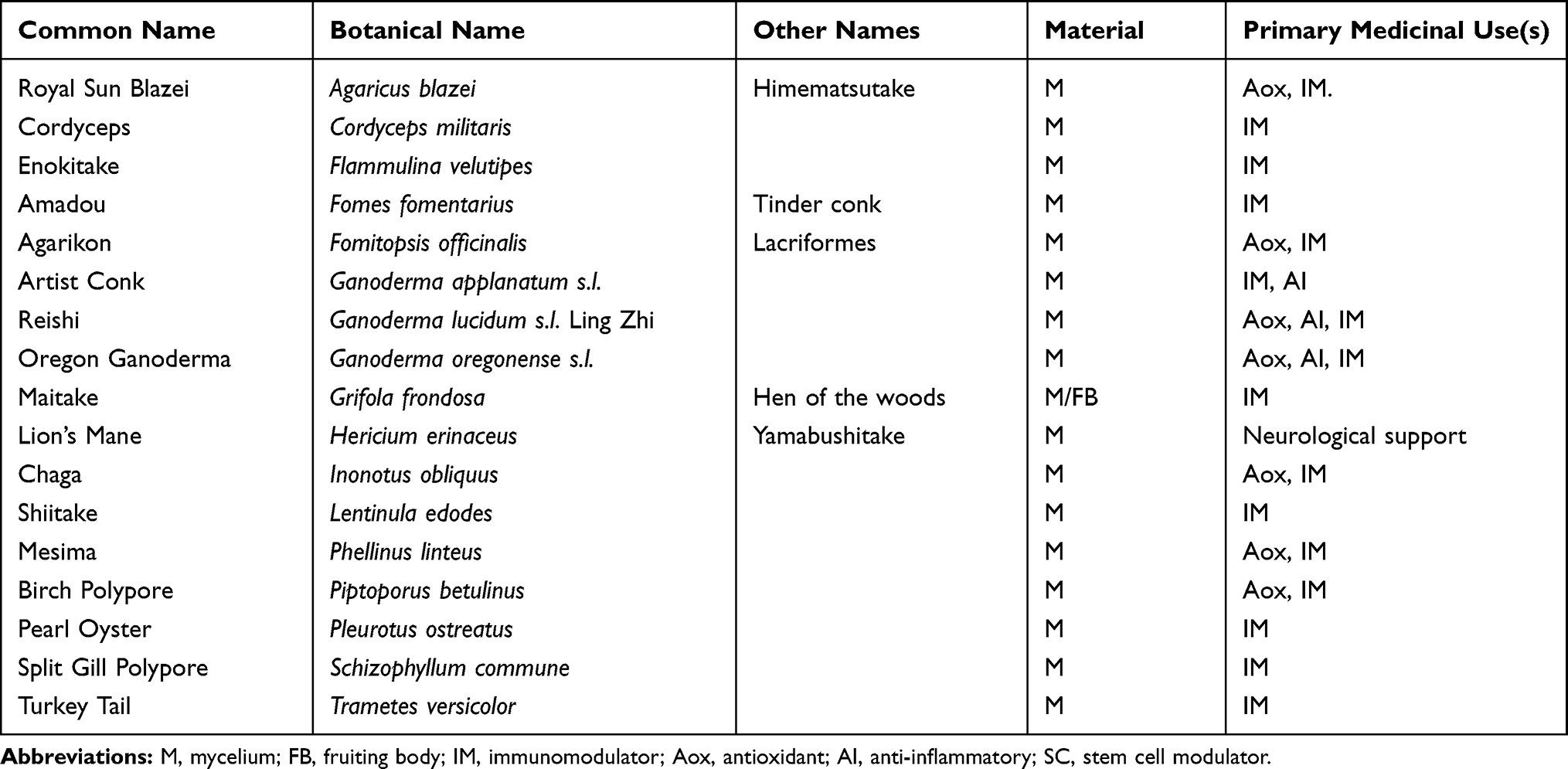 |
Table 1 Mushroom Species in the Medicinal Mushroom Blend |
Materials and Methods
Reagents
Phosphate-buffered saline, Roswell Park Memorial Institute 1640 medium, penicillin–streptomycin 100×, interleukin-2 (IL-2), and lipopolysaccharide (LPS) from Salmonella enterica were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Lympholyte-Poly was obtained from Thermo-Fisher Scientific (Waltham, MA, USA). CD69 fluorescein isothiocyanate, CD56 phycoerythrin, CD3 peridinin chlorophyll protein, and heparin Vacutainer tubes were purchased from Becton-Dickinson (Franklin Lakes, NJ, USA). Bio-Plex Pro™ human cytokine arrays were purchased from Bio-Rad Laboratories Inc. (Hercules, CA, USA).
Medicinal Mushroom Blend
The medicinal mushroom blend MyCommunity is sold internationally under the brand Host Defense® MushroomsTM and was obtained from the manufacturer—Fungi Perfecti, LLC, Olympia, WA. It is a certified organic freeze-dried blend of 17 medicinal mushroom species (mycelium/fruiting bodies) for immune system health: Royal sun blazei (Agaricus brasiliensis f. blazei), cordyceps (Cordyceps militaris), enokitake (Flammulina velutipes), amadou (Fomes fomentarius), agarikon (Fomitopsis officinalis), artist conk (Ganoderma applanatum), reishi (Ganoderma lucidum s.l.), Oregon ganoderma (Ganoderma oregonense s.l.), maitake (Grifola frondosa), lion’s mane (Hericium erinaceus), chaga (Inonotus obliquus), shiitake (Lentinula edodes), mesima (Phellinus linteus), birch polypore (Piptoporus betulinus), pearl oyster (Pleurotus ostreatus), split gill polypore (Schizophyllum commune), and turkey tail (Trametes versicolor; Table 1).
Testing for Endotoxins
The MMB powder was tested for endotoxin levels at Associates of Cape Cod Inc., East Falmouth MA, using the quantitative kinetic turbidimetric method for the detection of Gram-negative bacterial endotoxin, and reported in Endotoxin Units (EU).
Testing for Beta-Glucans
The whole MMB powder, as well as the freeze-dried solids of the aqueous and ethanol MMB fractions, were each tested for beta-glucan content by the Megazyme® assay at Venture Laboratories (Lexington, KY). This analysis involves total glucan hydrolysis by sulfuric acid and alpha-glucan hydrolysis by various enzymes. Total glucan and α-glucan content are measured spectrophotometrically, and β-glucans are estimated mathematically by the difference of these two values. All materials submitted for analysis were from the same lot number of material used in the immunological testing.
Preparation of Mushroom Fractions for Immune Bioassays
The powder was handled in the following manner, using pyrogen-free disposables: A 100 mg/mL suspension was prepared in phosphate-buffered saline (PBS) and allowed to rehydrate and extract aqueous compounds for 1 hr at 20°C under gentle agitation. The suspension was centrifuged at 400 g for 10 mins, and the aqueous fraction harvested. Ethanol (95%) was added to the pellet and vortexed, and extraction of non-aqueous, ethanol-soluble compounds allowed for 1 hr at 20°C under gentle agitation. The suspension was centrifuged at 400 g for 10 mins, and the ethanol fraction harvested. The remaining solid pellet was resuspended in PBS. The aqueous and ethanol fractions were filtered through a 0.22-micron syringe filter before adding to cell cultures. The solid fraction was passed through homogenization spin columns (QIAshredder, Qiagen, Hercules, CA), but were not filtered through a 0.22-micron filter. From each fraction, serial dilutions were made in pyrogen-free physiological saline. See also Figure 1.
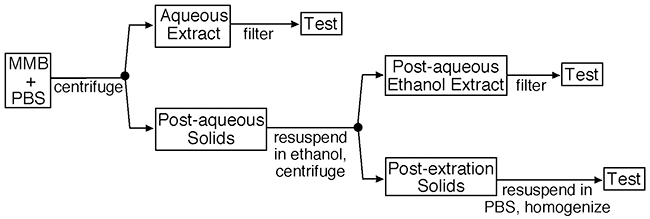 |
Figure 1 Production of test fractions. Notes: The medicinal mushroom blend was supplied as a powder, and suspended in PBS at a concentration of 100 g/L. The aqueous extraction was performed over 1 hr at ambient temperature under constant agitation. The post-aqueous solid fraction was extracted using ethanol under similar conditions. Both the aqueous and post-aqueous ethanol extracts were filtered through 0.22-micron cellulose acetate filters before adding to bioassays. The remaining post-extraction solids were homogenized but not filtered prior to testing in bioassays. |
Immune Cell Activation
Peripheral venous blood was drawn from three healthy human donors upon written informed consent, as approved by the Sky Lakes Medical Center Institutional Review Board, Federalwide Assurance 2603. The blood was drawn into heparin vacutainer vials, and the peripheral blood mononuclear cells (PBMC) isolated using Lympholyte Poly by centrifugation for 35 mins at 400 g. The PBMC were washed twice in PBS, counted, and the density adjusted to establish cultures with a cell density at 106/mL, using Roswell Park Memorial Institute 1640 medium containing penicillin–streptomycin and fetal bovine serum.
The highly inflammatory lipopolysaccharide (LPS) from Salmonella enterica was used as a positive control for immune-cell activation. Serial dilutions of products or LPS (10 ng/mL) were added to cultures at a volume of 20 μL, and cultures were then incubated at 37°C, 5% CO2 for 24 hrs. In parallel, IL-2 was used as a positive control for natural killer (NK)-cell activation, at a concentration of 100 IU/mL. Untreated negative control cultures consisted of PBMC exposed to phosphate-buffered saline in the absence of test products. All treatments, including each dose of test product and each positive and negative control, were tested in triplicate. After 24 hrs, cells were transferred to V-bottom microtiter plates, washed in PBS containing bovine serum albumin and sodium azide, and stained for 10 mins with fluorochrome-labeled anti-CD3, anti-CD56, and anti-CD69 monoclonal antibodies at the recommended concentration. PBMC were then fixed in formalin. The fluorescence intensities for CD3, CD56, and CD69 were measured by flow cytometry, using an Attune acoustic-focusing flow cytometer (Thermo Fisher Scientific, Waltham, MA, USA). Data analysis utilized gating on forward and side scatter to evaluate CD69 expression on lymphocyte versus monocyte/macrophage subsets. The lymphocyte subpopulation was further analyzed for CD69 expression on CD3-CD56+ NK cells, CD3+CD56+ NKT cells, CD3+CD56- T cells, and non-NK non-T lymphocytes.
Production of Cytokines, Chemokines, and Growth Factors
After 24 hrs of incubation, after transfer to V-bottom microtiter plates and before cells were stained for flow-cytometry analysis, the culture supernatants were harvested from the PBMC cultures described above. Levels of cytokines, chemokines, and growth factors were quantified using Bio-Plex protein arrays (Bio-Rad Laboratories Inc., Hercules CA, USA) and utilizing xMAP technology (Luminex, Austin, TX, USA).
Statistical Analysis
Data organization, exploration, and analysis were conducted using the statistical computing language R (version 3.5.2) implemented in the RStudio (version 1.0.143) software environment.35,36 Most data handling was conducted using the base package, with additional functions from the dplyr package (version 0.7.8) used in the reorganization of the raw data.37 Statistical analysis was implemented using the R ‘stats’ (version 3.5.2) and “car” (version 3.0–2) packages,38 as well as the packages “vegan” (version 2.5–4), and “vegan3d” (version 1.1–2) for multivariate ordination plots and Non-metric Multidimensional Scaling (NMDS).39,40 Multiple comparison test after Kruskal–Wallis used the “pgirmess” package (version 1.6.9), and the “RVAideMemoire” package (version 0.9–72) was used for pairwise permutational MANOVA with Bonferroni correction.41,42
Results
Immune Activation
The aqueous and the ethanol fractions of the MMB blend were tested for immune cell activation in cultures of peripheral blood mononuclear cells (PBMC) from healthy donors (Figures 2–4). Whereas the aqueous fraction triggered up-regulation of CD69 expression on lymphocytes (Figures 2A and 4A), the ethanol fraction triggered up-regulation of CD69 on monocytes (Figures 2B and 4B). Within the lymphocyte subset treated with the aqueous extract of MMB, only minor increases in CD69 expression were observed on NK cells (Figures 3A and 4C), NKT (Figures 3B and 4D), and T cells (Figures 3D and 4F). The most robust increase in CD69 expression was seen for the CD3- CD56- non-NK non-T lymphocytes where the increase was statistically significant across all doses tested (Figure 3C) and across all donors (Figure 4E). This CD3- CD56- population contains dendritic cells, B lymphocytes, and stem cells, all which may express CD69.
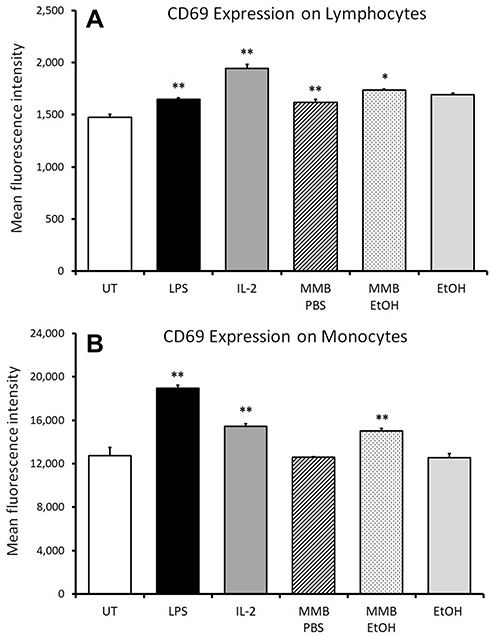 |
Figure 2 Expression of the early activation marker CD69 on lymphocytes and monocytes. Notes: *P<0.05; **P<0.01. CD69 expression on lymphocytes (A) and monocytes (B) in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS) and MMB ethanol extract (MMB EtOH). Results are shown for the 2 g/L dose. Mean fluorescence intensity for CD69 expression is shown. Data presented as mean ± standard from triplicate cultures and represents data from PBMC cell cultures from one of the three different healthy human donors. |
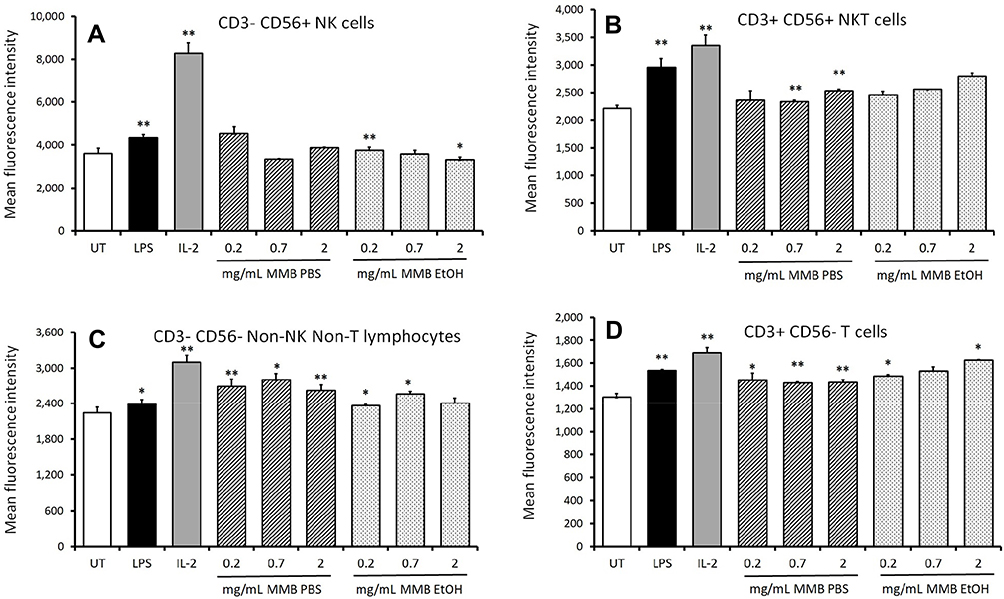 |
Figure 3 Expression of the early activation marker CD69 on lymphocyte subpopulations. Notes: *P<0.05; **P<0.01. CD69 expression on NK cells (A), NKT cells (B), non-NK non-T lymphocytes (C), and T cells (D) in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS) and MMB ethanol extract (MMB EtOH). Mean fluorescence intensity for CD69 expression is shown. Data presented as mean ± standard deviation from triplicate cultures and represents one of three separate experiments using PBMC cells from three different healthy human donors. |
The CD69 expression across all subpopulations of peripheral blood mononuclear cells was analyzed for the untreated cultures from all three donors and compared to CD69 expression in cultures treated with the two highest doses of MMB (2 mg/mL and 0.4 mg/mL). The analysis demonstrated the difference in induction of CD69 expression between the aqueous and the post-aqueous ethanol-based extracts (Figure 4). In these univariate box plots, the implicit statistical assumption is that the response of each cell type is independent of the response by each other cell type.
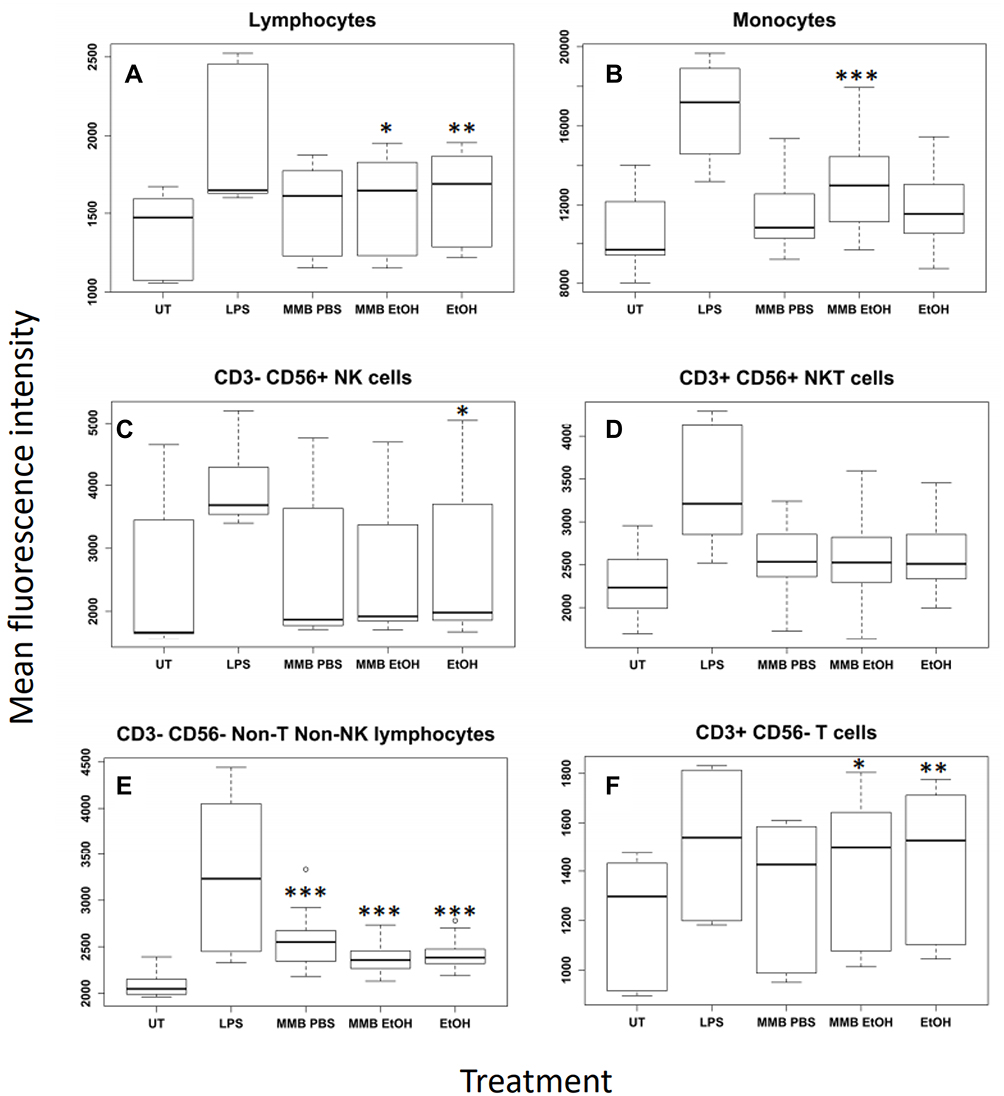 |
Figure 4 Expression levels of the CD69 early marker across white blood cell types. Notes: *P<0.05; **P<0.01, ***P<0.001. Mean fluorescence intensity resulting from immunostaining for the CD69 early activation marker for lymphocytes (A), monocytes (B), NK cells (C), NKT cells (D), non-T non-NK cells (E), and T cells (F). Data were plotted with respect to cell type and treatment group for the two highest concentrations pooled across all donors. Fluorescence responses that followed a normal distribution (B, D, E) were statistically evaluated with ANOVA followed by a Tukey Honest Significant Difference post hoc test. Non-normal fluorescence responses that could not be analyzed by ANOVA were evaluated using a Kruskal–Wallis Rank Sum Test followed by a multiple comparison post hoc test with Bonferroni correction. Statistical significance is indicated as the difference between untreated cells and treatments with a family-wise error rate indicated by p<0.05 (*); p<0.01 (**); p<0.001 (***). |
In addition to evaluation of cellular activation and CD69 expression on different cell types by standard univariate methodology, we also performed non-metric multidimensional scaling (NMDS)—a statistical technique borrowed from quantitative ecology—and permutational MANOVA. This is based on the rationale that the PBMC cultures contain many cell types that interact and affect each other dynamically. These interactions affect the transcriptional landscape within each culture. This can be analyzed in the same way as ecologists study changes in flora and fauna abundance across diverse landscapes. The method was also applied to enable comparison of responses across the PBMC cultures from all three donors. In this analogy, each donor is like an ecological research site, each cytokine, and growth factor is like a plant or animal type, and each MMB extract treatment is like a different type of land management practice (e.g. controlled burn forest vs untended forest vs open range forest). In the multivariate ordination plots, an assumption of independence among cell types (in the case of CD69 expression) or assumed statistical independence among co-induced cytokines is no longer necessary or appropriate. This is because the contemporaneously collected data is a snapshot of the aggregate PBMC response and it can be analyzed to account for covariance of responses among cell types or cytokines. The differences between the treatment groups is apparent when the covariance structure of the data set is represented using NMDS (Figure 5).
In the NMDS ordination plot in Figure 5, each dot represents the “Cartesian coordinate” location of each PBMC treatment well in a 5-dimensional data space (one dimension for each cell type), where the distance between points is proportional to the dissimilarity between each measurement. The dissimilarity between measurements was calculated using Bray-Curtis method to create a statistical distance matrix.43 This 5-dimensional data space is then projected into a plot where each axis is an eigenvector of the distance matrix. An eigenvector is a transect through the data space that explains the greatest proportion of variance in the data. Figure 5 is a 3-dimensional projection along the three eigenvectors (NMDS1, NMDS2, and NMDS3) that explain the greatest proportion of the variances. The overall “goodness of fit” statistic for this multivariate representation, known as stress, indicates that the projection in 3-dimensions provides an excellent representation of the underlying relationships (stress = 0.043).
Permutational MANOVA and post hoc pairwise comparisons with Bonferroni correction revealed that when considered collectively across cell types, the CD69 expression induced by MMB PBS was significantly different from the Untreated PBS (p<0.1), the MMB EtOH was different from the Untreated EtOH (p<0.05), and the MMB PBS extract was different from the MMB EtOH extract (p<0.05). Notably, the proportion of the variance in CD69 expression levels that can be explained by each treatment type was distinct for different white blood cell types (Table 2).
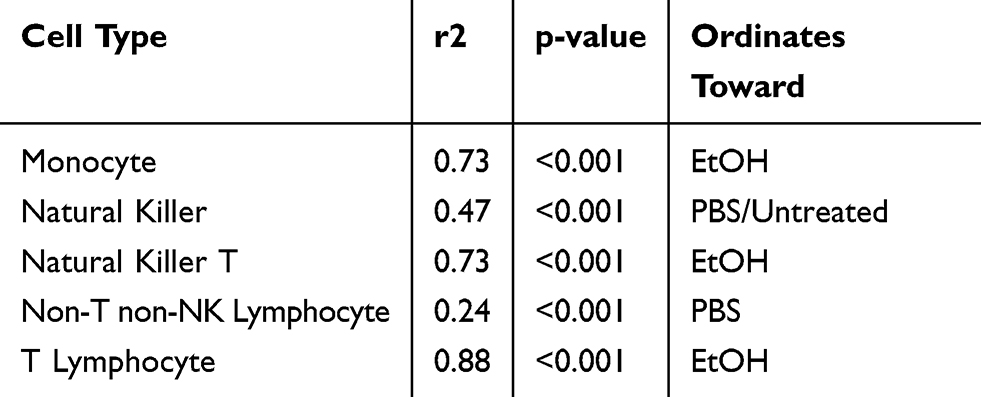 |
Table 2 Redundancy Analysis: Summary Statistics for Correlation Between Cell Type and CD69 Induction |
The NMDS ordination also confirmed the general trends identified in the univariate analysis: monocyte activation was strongly associated with the ethanol extract; non-T non-NK lymphocyte activation strongly correlated with the aqueous extract; and NKT cell, and T lymphocyte cell activation having a weaker but detectable activation by both the aqueous and ethanol extracts.
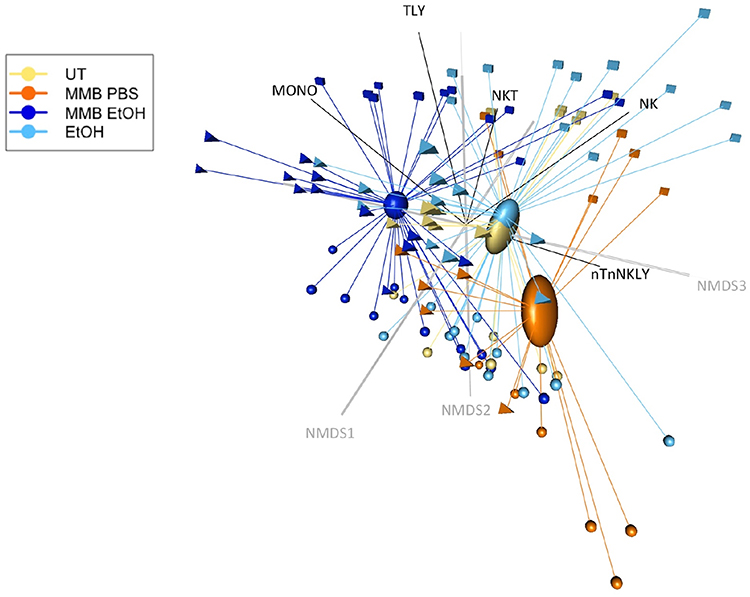 |
Figure 5 NMDS ordination plot of CD69 expression by treatment and cell type. Notes: Ordination plot of NMDS site scores representing Bray-Curtis distance dissimilarities. The three-dimensions of the plot provide an excellent representation of the data in reduced dimensions (stress < 0.05). The CD69 activation of each cell type is displayed as NMDS species vectors, with the vector direction and length indicating the strength of the correlation. Ovals represent the standard error of each treatment groups’ data centroid within the multivariate data space. |
Immune-Activating Cytokines
Supernatants from the PBMC cultures exposed to various doses of the MMB fractions for 24 hrs were assayed for the levels of cytokines, chemokines, and growth factors, using a magnetic bead-based array and Luminex xMAP technology. Increases in the levels of immune-activating cytokines included robust upregulation of specific pro-inflammatory cytokines, including IL-1β and TNF-α (Figure 6). The solid fraction of MMB (MMB solids) showed the most robust immune-activating properties, but the aqueous extract (MMB PBS), free of insoluble β-glucans, also showed induction of immune-activating cytokines. The post-aqueous ethanol extract (MMB EtOH) of the solids remaining after aqueous extraction showed only very minor effects on these cytokines.
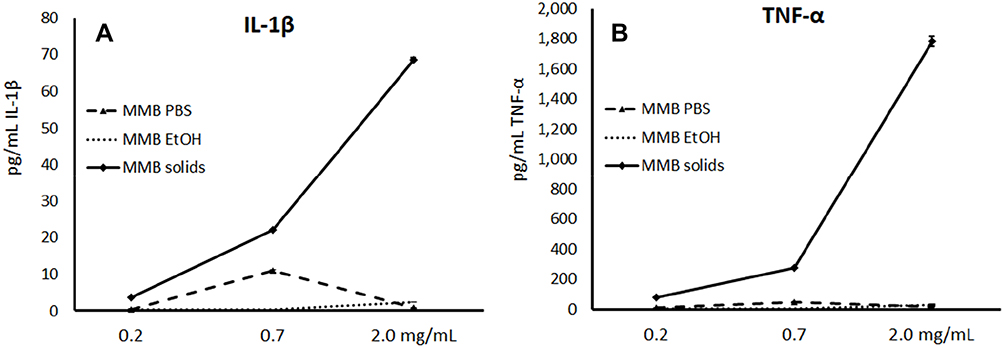 |
Figure 6 Changes in immune-activating pro-inflammatory cytokine levels in human PBMC cultures. Notes: Changes in Interleukin-1β (IL-1β), (A) and Tumor Necrosis Factor-α (TNF-α), (B) cytokine levels in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS), MMB post-aqueous ethanol extract (MMB EtOH), and the solid fraction (MMB solids). Data are presented as picogram per milliliter (pg/mL) based on the mean ± standard deviation from triplicate cultures and represents one of three experiments using PBMC from three different healthy human donors. Cytokine levels for LPS-treated control cultures were as follows: Il-1β: 772±28 pg/mL; TNF-α: 7073±185 pg/mL. |
Anti-Viral Cytokines
The MMB solid fraction triggered increases in the production of four specific cytokines and chemokines that are specifically associated with anti-viral activities and cellular recruitment (Figure 7). Interferon-γ (IFN-γ) was moderately induced by the MMB solid fraction (Figure 7A). The monocyte chemoattractant protein-1 (MCP-1) and macrophage inflammatory proteins 1α and 1β were strongly increased by MMB solid fraction (Figure 7B–D). No major changes to the biomarkers were apparent in the univariate analysis of the aqueous or post-aqueous ethanol extracts.
 |
Figure 7 Changes in anti-viral cytokine levels in human PBMC cultures. Notes: Changes in Interferon-γ (IFN-γ), (A), Monocyte Chemoattractant Protein-1 (MCP-1), (B), Macrophage Inflammatory Protein-α (MIP-1α), (C), and Macrophage Inflammatory Protein-β (MIP-1β), (D) cytokine levels in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS), MMB post-aqueous ethanol extract (MMB EtOH), and the solid fraction (MMB solids). Data are presented as picogram per milliliter (pg/mL) based on the mean ± standard deviation from triplicate cultures and represents one of three experiments using PBMC from three different healthy human donors. Cytokine levels for LPS-treated control cultures were as follows: IFN-γ: 71±2 pg/mL; MCP-1: 2052±35 pg/mL; MIP-1α: 2116±0 pg/mL; MIP-1β: 6497±814 pg/mL. |
Anti-Inflammatory Cytokines
The MMB solid fraction was a very strong inducer of the anti-inflammatory cytokine IL-1ra (Figure 8A), which functions as an IL-1 receptor antagonist, a strong anti-inflammatory protein due to its ability to prevent IL-1 in engaging in receptor binding and cellular signaling.44 To a much lesser extent, MMB solids triggered increased levels of IL-10 (Figure 8B); however, this was not statistically significant in the multivariate data analysis. The aqueous fraction triggered a mild induction of both anti-inflammatory cytokines. The post-aqueous ethanol extract of MMB did not trigger changes to either cytokine.
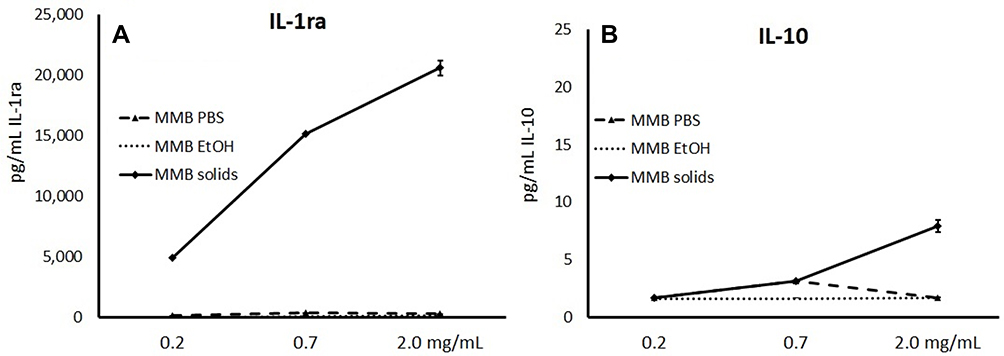 |
Figure 8 Changes in anti-inflammatory cytokines. Notes: Changes in interleukin-1 receptor antagonist (IL-1ra), (A) and interleukin-10 (IL-10), (B) cytokine levels in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS), MMB post-aqueous ethanol extract (MMB EtOH), and the solid fraction (MMB Solids). Data are presented as picogram per milliliter (pg/mL) based on the mean ± standard deviation from triplicate cultures and represents one of three experiments using PBMC from three different healthy human donors. Please note the 1000-fold difference in the scales on the Y-axes. Cytokine levels for LPS-treated control cultures were as follows: IL-1ra: 8134±473 pg/mL; IL-10: 301±15 pg/mL. |
Cytokines with Effects on Regenerative Functions
The effects of the MMB fractions on three cytokines with regenerative functions were clearly apparent in the univariate analysis of a representative donor, namely Granulocyte-Colony stimulating Factor (G-CSF), basic Fibroblast Growth Factor (bFGF), and Vascular Endothelial Growth Factor (VEGF) (Figure 9). Both the aqueous and the solid fractions induced increases in these three cytokines (Figure 9), with the MMB solid fraction showing the stronger effect.
 |
Figure 9 Changes in cytokines with effects on regenerative functions. Notes: Changes in Granulocyte-colony stimulating factor (G-CSF), (A), basic Fibroblast Growth Factor (Basic FGF), (B), and Vascular Endothelial Growth Factor (VEGF), (C) growth factor levels in human PBMC cultures treated for 24 hrs with serial dilutions of MMB aqueous extract in PBS (MMB PBS), MMB post-aqueous ethanol extract (MMB EtOH), and the solid fraction (MMB Solids). Data are presented as picogram per milliliter (pg/mL) based on the mean ± standard deviation from triplicate cultures and represents one of three experiments using PBMC from three different healthy human donors. Cytokine levels for LPS-treated control cultures were as follows: G-CSF: 1458±58 pg/mL; bFGF: 221±8 pg/mL; VEGF: 421±17 pg/mL. |
Multivariate Analysis of Cytokine Activation
Because the levels of cytokines, chemokines, and growth factors were all quantified simultaneously using bead-based protein arrays, the meaning of expression levels of an individual analyte are more appropriately analyzed in the context of the other contemporaneously induced signaling molecules. Mathematically, this means the variance-covariance structure of the immune analytes must be taken into account. In the cytokine NMDS, the concentrations (pg/mL) of all 27 monitored cytokines and growth factors from the two highest MMB doses for all three fractions and including all three donors were analyzed simultaneously. The NMDS ordination visualization (Figure 10) and summary statistics (Table 3) illustrate the strength of the solid fraction in inducing an immune response. Permutational MANOVA and post hoc pairwise comparisons with Bonferroni correction revealed that when considered collectively across cytokines and growth factor expression, the MMB PBS was different from the Untreated PBS (p=0.02), and the MMB Solids were different from the Untreated PBS (p=0.02), the MMB PBS (p=0.02), and MMB EtOH (p=0.03). The net effect of the MMB EtOH on cytokines and growth factors was not significantly different from the Untreated PBS, the Untreated EtOH, or the MMB PBS.
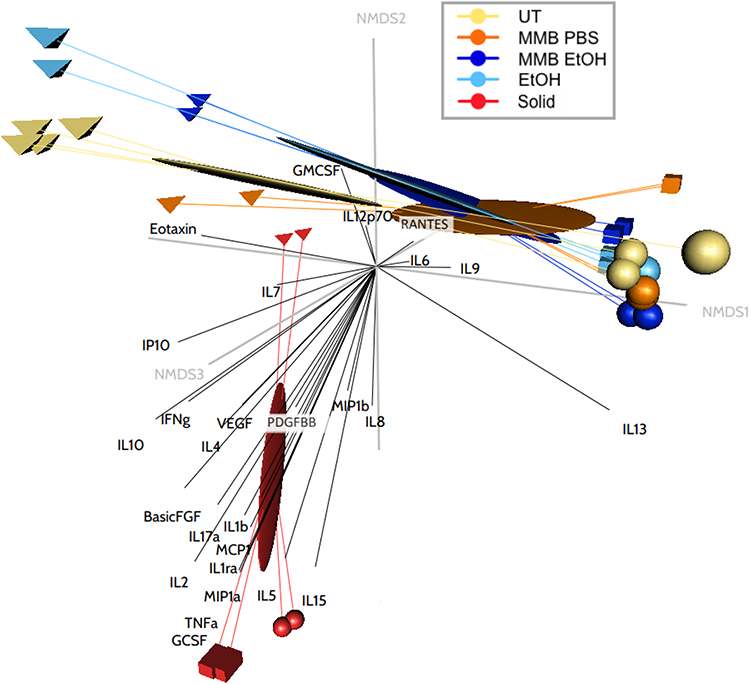 |
Figure 10 NMDS ordination plot of changes to cytokine and growth factor expression in PBMC cultures. Notes: Ordination plot of NMDS site scores representing Bray-Curtis distance dissimilarities between each treatment. The three-dimensions of the plot provide an excellent representation of the data in reduced dimensions (stress < 0.05). |
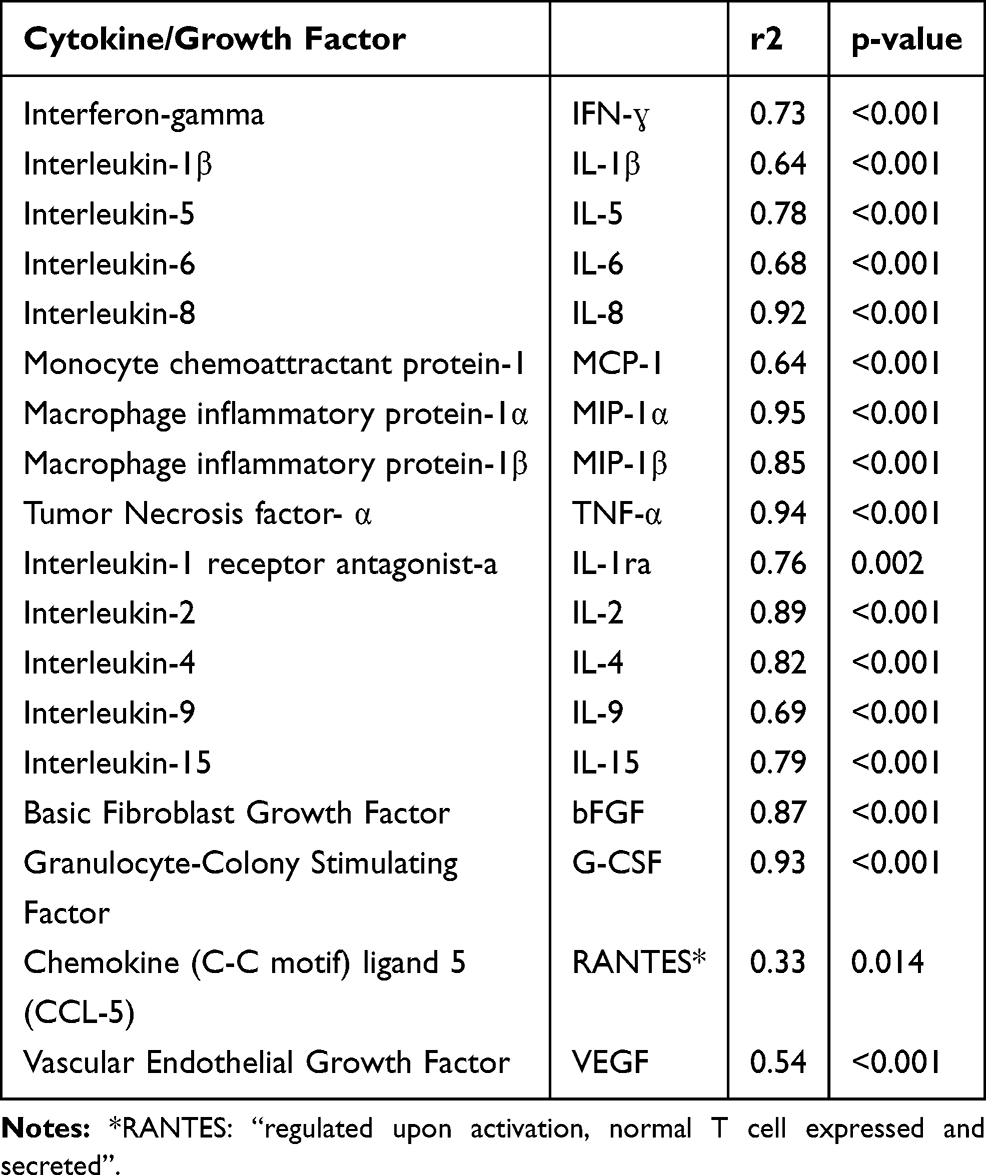 |
Table 3 Multivariate Redundancy Analysis: Summary Statistics by Cytokine and Growth Factor |
Endotoxin Testing
The level of endotoxin was 3.97 Endotoxin Units (EU)/mg MMB powder. Using the same assay, purified LPS results in 10 EU/ng, equivalent to 10,000,000 EU/mg purified LPS. This 2.5 million-fold difference shows a much lower content of endotoxin in MMB compared to LPS and serves as a foundation upon which the immune activation data can be interpreted, since LPS was used as a positive control in the immune cell cultures. In several data sets below (Figures 4C and 8A), the immune-activating properties of MMB fractions exceeded that of a 10 ng/mL dose of LPS, suggesting that endotoxins in MMB would not be a major contributing factor to the induction of immune cell activation and cytokine production.
Beta-Glucan Testing
The whole MMB powder was tested for beta-glucan content by the Megazyme® assay and showed 38% (w/w) beta-glucan. The water and ethanol fractions used in the immune assays were measured by the same assay at 2.0% w/w and 1.1% w/w, respectively. The solid fraction gave inconclusive results when tested in the Megazyme® assay, though it assumed that the difference between the whole MMB powder (38% beta-glucan) and the liquid extract beta-glucan yields the remaining insoluble beta-glucan in the solid fraction at approximately 35% w/w.
Discussion
Inflammation is a complex biological response to harmful stimuli, such as pathogens, as well as trauma or disease leading to tissue damage. It is a protective response involving immune cells, endothelial cells, and various types of stem cells, where the various cell types communicate via secreted biomarkers. The function of inflammation is to 1) increase blood and fluid flow to an area to promote the circulation of immune and stem cells, 2) eliminate the initial cause of cell injury, 3) clear out dead cells and tissues damaged from the original insult and from the inflammatory process, and 4) initiate tissue repair. Acute inflammation favors a healing process and return to homeostasis, whereas chronic inflammation does not support a resolution of the inflammation associated with healing and regeneration. Efforts to reduce inflammation, increase stem cell homing, and accelerate repair is thus an active area of research.45
While the immune-activating, pro-inflammatory properties of fungal water-insoluble β-glucans are well documented, the immune-modulating effects of fungal non-β-glucan-fractions are less recognized. This study was performed to examine three non-overlapping fractions of a medicinal mushroom blend, to compare the effects of the aqueous fraction to the post-aqueous ethanol fraction, and to compare both to the solid fraction. The solid fraction contained 38% insoluble beta-glucans, while the aqueous and post-aqueous ethanol fractions contained small amounts of soluble beta-glucans accounting for very little content on a mass basis. The comparison showed that while the aqueous fraction was able to induce CD69 expression on lymphocytes, the post-aqueous ethanol fraction induced CD69 on monocytes, thus exhibiting complementary immune-modulating activities. The solid fraction provided a highly robust effect on cytokine and growth factor production.
The post-aqueous ethanol fraction showed a more selective immune regulating activity. It supported a stronger monocyte activation than the aqueous extract and comparable induction of IL-6, IL-9, and MIP-1β. This suggests that hydrophobic ethanol-soluble compounds in this fraction have immune activating and anti-viral properties, while also supporting selective aspects of regenerative functions related to IL-9.
This selective effect of the three MMB fractions is of interest when discussing the potential differential immune regulating events after consumption. When a medicinal mushroom powder is ingested, it is assumed that aqueous compounds easily dissolve and are absorbed into the gastrointestinal mucosa. Compounds that are not water soluble either remain solids or get broken down by digestive enzymes and pH changes. The experimental post-aqueous ethanol fraction used for this project represents a method to test some compounds that may be released during the digestive process. The remaining solid fraction represents material that may get into direct contact with mucosal immune cells such as dendritic cells, known to extend trans-mucosal dendrites into the gut lumen, as well as absorption via transmucosal transport mechanisms to engage with tissue-residing gut mucosal immune cells.46
The strong induction of IL-1β, TNF-α, and IFN-γ, in combination with induction of G-CSF, suggests that a cascade of events is triggered by exposure of cells to MMB fractions, potentially involving mesenchymal stem cells as a pivotal regulating cell type. Mesenchymal stem cells can cross the blood–brain barrier and contribute to repair of brain injuries such as stroke. Mesenchymal stem cells respond to inflammation in a manner that leads to events to counteract inflammation and promote homeostasis. When mesenchymal stem cells from healthy human donors were treated with the inflammatory cytokines IL-1, TNF-α, and IFN-γ, these cells responded with a strong increase in G-CSF production.47 This response was able to reprogram highly inflamed LPS-activated microglial cells to reduce the production of inflammatory mediators.
Further work should include evaluation of temporal events in cell culture during the cascade when human immune cells respond to MMB fraction exposure. Future testing should also include examination of immune activation when cells are treated with a combination of all three fractions, thereby simulating the exposure of gut mucosal cells to the compounds in the MMB. Additional work may also include cell cultures of immune cells isolated from gut mucosal tissue such as lamina propria and Peyer’s patches, in the presence of bacterial and viral challenges, as well as characterization of the different mushroom fractions and respective contributions of each of the 17 mushroom species in the MMB. Clinical evaluation of immune support, for example in acute and chronic viral infections, is also warranted and is currently in the planning stage.
Conclusion
The exposure of human immune cells to fractions of the medicinal mushroom blend MMB triggered different and distinct immune responses by the hydrophilic, hydrophobic, and insoluble fractions. The data have documented unique and potentially synergistic effects, where the aqueous extract activated cells within the non-NK non-T cell fraction, which contains the dendritic cell types involved in immune recognition and antigen presentation. In contrast, the post-aqueous ethanol extract contained compounds able to activate monocytes, T-lymphocytes, and Natural Killer T cells. The solid fraction was a potent but highly selective inducer of multiple cytokines with both pro-and anti-inflammatory activity and was also supportive of induction of growth factors involved in regenerative functions. The results show that the MMB extract fractions selectively induced immune cell activation and increased host-defense activity, including anti-viral effects. These data also provide a mechanistic explanation for the post-inflammatory induction of G-CSF and multiple other cytokines and growth factors involved in tissue repair. Subsequently, induction of anti-inflammatory cytokines, such as IL-1ra, help moderate and resolve the immune defense-induced pro-inflammatory reactions.
Acknowledgments
The study was performed at NIS Labs, an independent research laboratory focusing on natural products research. The work was sponsored by Fungi Perfecti, LLC, a grower and producer of commercially available mushrooms, and finished consumable products.
Author Contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
RD, RN, and AT are employed by the sponsor of the study, Fungi Perfecti, LLC. PS holds several patents on topics related to the presented work and is the founder and owner of Fungi Perfecti, LLC. PS also conducted this research on a formulation that he has devised which is sold as a nutritional supplement. The authors report no other conflicts of interest in this work.
References
1. Wasser SP. Medicinal mushroom science: current perspectives, advances, evidences, and challenges. Biomed J. 2014;37(6):345–356. doi:10.4103/2319-4170.138318
2. Wu T, Zivanovic S, Draughon F, Conway W, Sams C. Physicochemical properties and bioactivity of fungal chitin and chitosan. J Agric Food Chem. 2005;53(10):3888–3894. doi:10.1021/jf048202s
3. Kozarski M, Klaus A, Niksic M, Jakovljevic D, Helsper JPFG, Van Griensven LJLD. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phellinus linteus. Food Chem. 2011;129(4):1667–1675. doi:10.1016/j.foodchem.2011.06.029
4. Wang Q, Wang F, Xu Z, Ding Z. Bioactive mushroom polysaccharides: a review on monosaccharide composition, biosynthesis and regulation. Molecules. 2017;22(6):E955.
5. Ruthes AC, Smiderle FR, Iacomini M. Mushroom heteropolysaccharides: A review on their sources, structure and biological effects. Carbohydr Polym. 2016;136:358–375. doi:10.1016/j.carbpol.2015.08.061
6. He X, Wang X, Fang J, et al. Polysaccharides in Grifola frondosa mushroom and their health promoting properties: A review. Int J Biol Macromol. 2017;101:910–921. doi:10.1016/j.ijbiomac.2017.03.177
7. Yoon TJ, Koppula S, Lee KH. The effects of beta-glucans on cancer metastasis. Anticancer Agents Med Chem. 2013;13(5):699–708. doi:10.2174/1871520611313050004
8. Rop O, Mlcek J, Jurikova T. Beta-glucans in higher fungi and their health effects. Nutr Rev. 2009;67(11):624–631. doi:10.1111/nure.2009.67.issue-11
9. Lee DH, Kim HW. Innate immunity induced by fungal beta-glucans via dectin-1 signaling pathway. Int J Med Mushrooms. 2014;16(1):1–16. doi:10.1615/IntJMedMushr.v16.i1.10
10. Johnson CM, O’Brien XM, Byrd AS, et al. Integrin cross-talk regulates the human neutrophil response to fungal β-glucan in the context of the extracellular matrix: a prominent role for VLA3 in the antifungal response. J Immunol. 2017;198(1):318–334. doi:10.4049/jimmunol.1502381
11. Quayle K, Coy C, Standish L, Lu H. The TLR2 agonist in polysaccharide-K is a structurally distinct lipid which acts synergistically with the protein-bound β-glucan. J Nat Med. 2015;69(2):198–208. doi:10.1007/s11418-014-0879-z
12. Ashraf Khan A, Gani A, Masoodi FA, Mushtaq U, Silotry Naik A. Structural, rheological, antioxidant, and functional properties of β–glucan extracted from edible mushrooms Agaricus bisporus, Pleurotus ostreatus and Coprinus atramentarius. Bioact Carbohydr Diet Fibre. 2017;11:67–74. doi:10.1016/j.bcdf.2017.07.006
13. Fritz H, Kennedy DA, Ishii M, et al. Polysaccharide K and Coriolus versicolor extracts for lung cancer: a systematic review. Integr Cancer Ther. 2015;14(3):201–211. doi:10.1177/1534735415572883
14. Rosendahl AH, Sun C, Wu D, Andersson R. Polysaccharide-K (PSK) increases p21(WAF/Cip1) and promotes apoptosis in pancreatic cancer cells. Pancreatology. 2012;12(6):467–474. doi:10.1016/j.pan.2012.09.004
15. Zhang H, Morisaki T, Nakahara C, et al. PSK-mediated NF-kappaB inhibition augments docetaxel-induced apoptosis in human pancreatic cancer cells NOR-P1. Oncogene. 2003;22(14):2088–2096. doi:10.1038/sj.onc.1206310
16. Mitomi T, Tsuchiya S, Iijima N, et al. Randomized, controlled study on adjuvant immunochemotherapy with PSK in curatively resected colorectal cancer. The Cooperative Study Group of Surgical Adjuvant Immunochemotherapy for Cancer of Colon and Rectum (Kanagawa). Dis Colon Rectum. 1992;35(2):123–130. doi:10.1007/BF02050666
17. Ooi VE, Liu F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr Med Chem. 2000;7(7):715–729. doi:10.2174/0929867003374705
18. Nakazato H, Koike A, Saji S, Ogawa N, Sakamoto J, Nakazato H. Efficacy of immunochemotherapy as adjuvant treatment after curative resection of gastric cancer. Study Group of Immunochemotherapy with PSK for Gastric Cancer. Lancet. 1994;343(8906):1122–1126. doi:10.1016/S0140-6736(94)90233-X
19. Turlo J, Lubinski O, Gutkowska B. Isolation of lentinan, an immunomodulating (1-3)-b-D-glucan from submerged cultivated mycelium of Lentinus edodes and culture medium. Acta Pol Pharm. 2004;61:40–42.
20. Wang H, Cai Y, Zheng Y, Bai Q, Xie D, Yu J. Efficacy of biological response modifier lentinan with chemotherapy for advanced cancer: a meta-analysis. Cancer Med. 2017;6(10):2222–2233. doi:10.1002/cam4.2017.6.issue-10
21. Adachi Y, Okazaki M, Ohno N, Yadomae T. Enhancement of cytokine production by macrophages stimulated with (1–>3)-beta-D-glucan, grifolan (GRN), isolated from Grifola frondosa. Biol Pharm Bull. 1994;17(12):1554–1560. doi:10.1248/bpb.17.1554
22. You BJ, Tien N, Lee MH, et al. Induction of apoptosis and ganoderic acid biosynthesis by cAMP signaling in Ganoderma lucidum. Sci Rep. 2017;7(1):318. doi:10.1038/s41598-017-00281-x
23. Lee EW, Shizuki K, Hosokawa S, et al. Two novel diterpenoids, erinacines H and I from the mycelia of Hericium erinaceum. Biosci Biotechnol Biochem. 2000;64(11):2402–2405. doi:10.1271/bbb.64.2402
24. Ma B-J, Shen J-W, Yu H-Y, Ruan Y, Wu -T-T, Zhao X. Hericenones and erinacines: stimulators of nerve growth factor (NGF) biosynthesis in Hericium erinaceus. Mycology. 2010;1(2):92–98. doi:10.1080/21501201003735556
25. Kawagishi H, Shimada A, Shirai R, et al. Erinacines A, B and C, strong stimulators of nerve growth factor (NGF)-synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1994;35(10):1569–1572. doi:10.1016/S0040-4039(00)76760-8
26. Kim YJ, Park J, Min BS, Shim SH. Chemical constituents from the sclerotia of Inonotus obliquus. J Korean Soc Appl Biol Chem. 2011;54(2):287–294.
27. Zhou X, Luo L, Dressel W, et al. Cordycepin is an immunoregulatory active ingredient of Cordyceps sinensis. Am J Chin Med. 2008;36(5):967–980. doi:10.1142/S0192415X08006387
28. Yang J, Cao Y, Lv Z, Jiang T, Wang L, Li Z. Cordycepin protected against the TNF-α-induced inhibition of osteogenic differentiation of human adipose-derived mesenchymal stem cells. Int J Immunopathol Pharmacol. 2015;28(3):296–307. doi:10.1177/0394632015592160
29. Dai J, Miller MA, Everetts NJ, et al. Elimination of quiescent slow-cycling cells via reducing quiescence depth by natural compounds purified from Ganoderma lucidum. Oncotarget. 2017;8(8):13770–13781. doi:10.18632/oncotarget.14634
30. Chen WY, Yang WB, Wong CH, Shih DT. Effect of Reishi polysaccharides on human stem/progenitor cells. Bioorg Med Chem. 2010;18(24):8583–8591. doi:10.1016/j.bmc.2010.10.016
31. Lin H, She YH, Cassileth BR, Sirotnak F, Cunningham Rundles S. Maitake beta-glucan MD-fraction enhances bone marrow colony formation and reduces doxorubicin toxicity in vitro. Int Immunopharmacol. 2004;4(1):91–99. doi:10.1016/j.intimp.2003.10.012
32. Lin H, Cheung SW, Nesin M, Cassileth BR, Cunningham-Rundles S. Enhancement of umbilical cord blood cell hematopoiesis by maitake beta-glucan is mediated by granulocyte colony-stimulating factor production. Clin Vaccine Immunol. 2007;14(1):21–27. doi:10.1128/CVI.00284-06.
33. Stamets PE. Potentiation of cell-mediated host defense using fruitbodies and mycelia of medicinal mushrooms. Int J Med Mushrooms. 2003;5(2). doi:10.1615/InterJMedicMush.v5.i2.70
34. Hip S, Fook Y, Swee K, Chun W. Phenolic profiles of selected edible wild mushrooms as affected by extraction solvent, time and temperature. As J Food Ag-Ind. 2009;2(3):392–401.
35. R Core Team. R: A Language and Environment for Statistical Computing; Version 3.4.0. Vienna, Austria: R Foundation for Statistical Computing; 2017. Available from: https://www.R-project.org.
36. RStudio Team. RStudio: Integrated Development for R. Version 1.0.143. Boston, MA, USA: RStudio, Inc.; 2016. Available from: http://www.rstudio.com.
37. Wickham H, Francois R, Henry L, Müller K, Wickham H. Dplyr: A grammar of data manipulation. Available from: https://CRAN.R-project.org/package=dplyr.
38. Fox J, Weisberg S. An {R} Companion to Applied Regression.
39. Oksanen J, Blanchet FG, Friendly M, et al. Vegan: community ecology package. Available from: https://CRAN.R-project.org/package=vegan.
40. Oksanen J, Kindt R, Simpson GL (2018). vegan3d: static and dynamic 3D plots for the ‘vegan’ package. R package version 1.1-2. Available from: https://CRAN.R-project.org/package=vegan3d.
41. Giraudoux P (2018) pgirmess: spatial analysis and data mining for field ecologists. R package version 1.6.9. Available from: https://CRAN.R-project.org/package=pgirmess.
42. Hervé M (2019). RVAideMemoire: testing and plotting procedures for biostatistics. R package version 0.9-72. Available from: https://CRAN.R-project.org/package=RVAideMemoire.
43. Ricotta C, Podani J. On some properties of the Bray-Curtis dissimilarity and their ecological meaning. Ecol Complex. 2017;31:201–205. doi:10.1016/j.ecocom.2017.07.003
44. Arend WP, Malyak M, Guthridge CJ, Gabay C. Interleukin-1 receptor antagonist: role in biology. Annu Rev Immunol. 1998;16:27–55. doi:10.1146/annurev.immunol.16.1.27
45. Giordano S, Zhao X, Chen YF, et al. Induced pluripotent stem cell-derived endothelial cells overexpressing interleukin-8 receptors A/B and/or C-C chemokine receptors 2/5 inhibit vascular injury response. Stem Cells Transl Med. 2017. doi:10.1002/sctm.16-0316
46. Sun M, He C, Cong Y, Liu Z. Regulatory immune cells in regulation of intestinal inflammatory response to microbiota. Mucosal Immunol. 2015;8(5):969–978. doi:10.1038/mi.2015.49
47. Redondo-Castro E, Cunningham C, Miller J, et al. Interleukin-1 primes human mesenchymal stem cells towards an anti-inflammatory and pro-trophic phenotype in vitro. Stem Cell Res Ther. 2017;8(1):79. doi:10.1186/s13287-017-0531-4
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.