Back to Journals » Drug Design, Development and Therapy » Volume 11
Differential effect of platelet-rich plasma fractions on β1-integrin signaling, collagen biosynthesis, and prolidase activity in human skin fibroblasts
Authors Guszczyn T ![]() , Surażyński A, Zaręba I
, Surażyński A, Zaręba I ![]() , Rysiak E, Popko J, Pałka J
, Rysiak E, Popko J, Pałka J
Received 2 March 2017
Accepted for publication 30 April 2017
Published 22 June 2017 Volume 2017:11 Pages 1849—1857
DOI https://doi.org/10.2147/DDDT.S135949
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sukesh Voruganti
Tomasz Guszczyn,1 Arkadiusz Surażyński,2 Ilona Zaręba,2 Edyta Rysiak,2 Janusz Popko,1 Jerzy Pałka2
1Department of Pediatric Orthopedics and Traumatology, 2Department of Medicinal Chemistry, Medical University of Białystok, Białystok, Poland
Abstract: The study was conducted to evaluate the effects of platelet-rich plasma (PRP), supernatant of PRP (SPRP) obtained by centrifugation, and supernatant of activated PRP (SActi-PRP) obtained by Ca2+ solution-treated PRP on collagen biosynthesis, prolidase activity, and β1-integrin signaling in cultured human skin fibroblasts. Incubation of fibroblasts with 5% PRP for 24 h contributed to ~5-fold increase in collagen biosynthesis compared to the control. In the cells treated with 5% of SPRP or SActi-PRP, collagen biosynthesis showed a 3-fold increase of the control. PRP, SPRP, and SActi-PRP stimulated prolidase activity similar to collagen biosynthesis. Collagen biosynthesis and prolidase activity are regulated by β1-integrin receptor signaling. Incubation of fibroblasts with PRP for 24 h contributed to a dose-dependent increase in the expression of β1-integrin receptor, while SActi-PRP increased the process to a much lower extent. SPRP had no effect on the β1-integrin receptor expression. All the studied fractions of blood increased the expression of FAK as well as the expression of phosphorylated MAP-kinases. However, PRP was found to be the most effective stimulator of expression of these particular kinases. These studies suggest that a complex of factors, including growth factors, adhesion molecules, and prolidase contained in PRP, all evoke growth and collagen-promoting activities in human dermal fibroblasts.
Keywords: fibroblasts, FAK, MAPK
Introduction
Platelet-rich plasma (PRP) is defined as a sample of blood with platelet (PLT) concentration above baseline values. PRP sample is obtained by centrifugation of blood and separation of appropriate blood fraction.1 Depending on the method used, it can contain 1.3–8 times higher concentration of PLTs in plasma than in whole blood. Moreover, PRP consists of plasma proteins, leukocytes, and erythrocytes. PRP is also a rich source of many cytokines and growth factors (GFs).2–4 It has been hypothesized that the PLTs, and particularly platelet-derived growth factors (PDGFs) contained in their alpha granules, are responsible for the biologic effects of PRP.5,6
The GFs released from the alpha granules of PLTs during thrombosis include PDGF, transforming growth factor-beta, platelet-derived epidermal growth factor, platelet-derived angiogenesis factor, insulin-like growth factor-I (IGF-I), and platelet factor 4. They are the best known GFs. Other GFs present in PLT granules are the vascular endothelial growth factors and the endothelial growth factors. The interaction between these GFs and the surface receptors on the target cells activates intracellular signaling pathways, which induces the expression of genes required for regenerative processes such as cellular proliferation and extracellular matrix formation.2–4,7
The most abundant component of the connective tissue responsible for the maintenance of its architecture and integrity is collagen. Collagen also plays an important role in the interaction with cell surface integrin receptors, through which it may participate in the regulation of gene expression, differentiation, growth, and other cellular functions. The interaction is involved in cytoskeleton reorganization, intracellular ion transport, lipid metabolism, kinase activation, gene expression, cell cycle regulation, and cancer metastasis.8–11 Stimulated β1-integrin receptor by the ligand, such as collagen, induce phosphorylation of non-receptor focal adhesion kinase (FAK) which is then capable of interaction with the region’s SH2 kinase Src. This phenomenon allows interaction with FAK adapter growth factor receptor bound protein 2 (Grb-2) and Shc through phosphorylation of the kinase. Thus, activated kinase Shc binds to protein Grb-2. Adapter protein Grb-2, the SH2 domain-rich protein which determines the binding to proline-rich domain of SOS protein (which is a protein exchanging the guanidine nucleotides), causes translocation of SOS protein to the plasma membrane where it activates Ras protein. Activated Ras protein interacts with Raf protein that phosphorylates MAP-kinase.12,13 MAP-kinases are translocated into the nucleus, where they activate several transcription factors such as NFkB, STAT, and p53 and regulate the expression of genes for many proteins involved in the regulation of cell growth, differentiation, and metabolism.14 One of the enzymes involved in regulation of extracellular matrix metabolism is prolidase (E.C. 3.4.13.9). Previously, it has been shown that β1-integrin receptor is involved in signaling that regulates collagen biosynthesis15 and prolidase activity.16
Prolidase is the enzyme that catalyzes the final step in collagen degradation by releasing proline or hydroxyproline from imidodipeptides.17 On the other hand, prolidase plays an important role in recycling proline for protein biosynthesis, especially collagen, which, among all known proteins, contains the highest amount of proline. Therefore, prolidase may represent an important factor involved in the regulation of collagen metabolism. All the above-mentioned factors and processes may be involved in the mechanism of PRP-dependent tissue regeneration.
PRP has been successfully used as an effective treatment in maxillofacial, periodontal and oral surgery, orthopedic surgery, trauma, gastrointestinal surgery, soft tissue injuries, burns, cosmetic and plastic surgery for about 20 years.18,19 Numerous reports suggest the use of PRP in delayed bone healing and nonunions, chondropathy, osteoarthritis, tendinopathy, acute and chronic soft-tissue injuries, enhancement of healing after ligament reconstruction, muscle strains, and in skin rejuvenation.2–4,20 The specific role of collagen in bones, tendons, ligaments and skin repair is well established.21,22 Besides the role of being a structural protein, collagen is a ligand of integrin receptors that transmit signals for cell growth, prolidase activity upregulation, and stimulation of collagen biosynthesis. All these processes are required for proper tissue regeneration during wound healing. PRP effectively assists in tissue repair by stimulation of collagen metabolism. Little is known about the mechanism of PRP on collagen biosynthesis. Studies of tissue engineering have shown that PRP increases collagen gene expression through GF receptor signaling.23,24
The interest in PRP is focused on its application in wound healing of injured tissues. However, the effectiveness of this process depends on the manner of PRP preparation. Although some data on optimization of PRP preparation for the purpose of regenerative medicine were reported,7 the mechanism for effectiveness of wound healing is not known.
The aim of this study was to evaluate the effects of PRP, plasma from PRP obtained by centrifugation – supernatant of PRP (SPRP), and supernatant of activated PRP obtained by Ca2+ solution-treated PRP (SActi-PRP) on collagen biosynthesis, prolidase activity, and β1-integrin signaling in cultured human skin fibroblasts. The PRP sample contains a high concentration of PLTs with white blood cells (WBCs) and small amounts of red blood cells (RBCs). The samples of SPRP and SActi-PRP showed no cellular elements. SActi-PRP contains cytokines and GFs released from the alpha granules during the Ca2+-mediated process of PLT activation.6,7,25
Materials and methods
The ethics committee of the Medical University of Bialystok approved the study, and all participants provided written informed consent prior to enrollment.
PRP production
PRP was obtained by a commercial system: SmartPReP®2 Autologous Platelet Concentrate+ (Harvest Technologies Corp., Plymouth, MA, USA). With the donor’s consent, 20 mL of venous blood (VB) was collected in a syringe containing 2 mL of Anticoagulant Citrate Dextrose Solution A. Another 1 mL of the Anticoagulant Citrate Dextrose Solution A was transferred into the white port on the process disposable. The content of the syringe was transferred into the red port on the process disposable and centrifuged for 14 min at two different speeds: at 2,000× g for the first 10 min and at 1,800× g for the remaining time, at room temperature. Using the plasma syringe with blunt cannula and spacer in the white port on the process disposable, PLT-poor plasma was withdrawn. Approximately 3 mL of PRP remained in the process disposable. PRP was then extracted into an empty sterile syringe.
To prepare SPRP, 1 mL of PRP was used and centrifuged for 10 min at 2,500× g. The SPRP was represented by the supernatant. To prepare SActi-PRP, 1 mL of PRP was incubated with 100 μL of Ca2+ solution (9 mg/mL, Teva, Petach Tikva, Israel) for 30 min at 37°C. After incubation, the sample was centrifuged (10 min at 2,500× g). The SActi-PRP was represented by the supernatant.
The concentrations of the cellular constituents of the blood, WBCs, RBCs, and PLTs, in each sample of VB, PRP, SPRP, and SActi-PRP were analyzed with an automated hematology system XT 4000i Sysmex.
Tissue culture
All studies were performed on normal human dermal fibroblasts (CRL-1474), which were purchased from the American Type Culture Collection (Manassas, VA, USA). The cells were maintained in DMEM supplemented with 5% fetal bovine serum, 2 mM glutamine, 50 U/mL penicillin, and 50 μg/mL streptomycin at 37°C in a 5% CO2 incubator. Cells were counted in a hemocytometer and cultured at 105 cells per well in 2 mL of growth medium in six-well plates (Costar, Waltham, MA, USA). Cells reached confluence at day 6 and were subsequently used for assays. Cells were used in the 8th–14th passages. Fibroblasts were incubated with reagents for 24 h.
Determination of prolidase activity
The activity of prolidase was determined according to the method of Myara et al,26 which involves colorimetric determination of proline applying Chinard’s reagent. Protein concentration was measured by the method of Lowry et al.27
Briefly, the harvested cells were centrifuged at 2,000× g for 15 min. The supernatant was discarded and the cells were suspended in 1 mL of 50 mM HEPES, pH 7.8, and sonicated for 3×10 s at 0°C. The degraded cells were centrifuged at 12,000× g for 30 min at 4°C. To activate prolidase, the enzyme was incubated with Mn(II) in the following mixture: 100 μL of cell extract supernatant with 100 μL of 50 mM HEPES, pH 7.8, in the presence of MnCl2 to achieve a final concentration of 1 mM in the mixture, and incubated for 24 h at 37°C. After this, the prolidase reaction was initiated by adding 100 μL of the activated mixture to 100 μL of 94 mM glycylproline (Gly-Pro) for a final concentration of 47 mM. The samples were then incubated for 1 h at 37°C. To terminate the reaction, 1 mL of 0.45 M trichloroacetic acid was added and the reaction mixture was centrifuged at 10,000× g for 15 min. The released proline was determined as follows: 0.5 mL of the reaction mixture was added to 2 mL of a 1:1 mixture of glacial acetic acid:Chinard’s reagent (25 g of ninhydrin dissolved at 70°C in 600 mL of glacial acetic acid and 400 mL of 6 M orthophosphoric acid) and incubated for 10 min at 90°C. The amount of released proline was determined by measurement of absorbance at 515 nm and reported as nanomoles per minute per milligram of protein.
Collagen production and total protein synthesis
Cells were counted in a hemocytometer and cultured at 105 cells per well in 2 mL of growth medium in six-well plates, (Costar) to confluency. Incorporation of a radioactive precursor into proteins was measured after labeling of confluent cells in the growth medium with varying concentrations of PRP, SPRP, and SActi-PRP for 24 h with 5[3H]proline (5 μCi/mL, 28 Ci/mmol) as described previously.28 Incorporation of 5[3H]proline into collagen was determined by digesting proteins with purified Clostridium histolyticum collagenase, according to the method of Peterkofsky et al.29 Collagen biosynthesis was reported as disintegrations per minute of proline incorporated to proteins susceptible to the action of bacterial collagenase, per 106 cells. Total protein synthesis was calculated from the sum of radioactivity of collagenase-resistant proteins and collagen digest. Results are shown as combined values for cell plus medium fractions, and relative differences in collagen biosynthesis between the studied groups were calculated as a per percent of the control value.
DNA biosynthesis
Cells were cultured in six-well plates (105 cells per well) as described above and were incubated with varying concentrations of PRP, SPRP, and SActi-PRP for 24 h with 0.5 Ci/mL of [3H]thymidine for 4 h at 37°C. After this, the cell surface was rinsed twice with 1 mL of 0.05 M Tris-HCl (pH 7.4) containing 0.1 M NaCl, and twice with 1 mL of 5% trichloroacetic acid. The cells were then lysed in 1 mL of 0.1 M NaOH containing 1% sodium dodecyl sulfate. The cell lysate was added to 4 mL of scintillation liquid, and incorporation of radioactivity into DNA was measured in a scintillation counter.
Immunofluorescence staining and bioimaging
Cells were seeded in BD Falcon™ 96-well black/clear bottom tissue culture plates optimized for imaging applications at 10,000 cells per well. After incubation, the cells were rinsed with PBS and fixed with 3.7% formaldehyde solution at room temperature for 10 min. After fixation, the cells were washed three times with PBS and permeabilized with 0.1% Triton X-100 solution at room temperature for 5 min. The cells were then washed twice with PBS, and nonspecific binding was blocked by adding 3% fetal bovine serum solution and then cells were incubated at room temperature for 30 min. After this, the cells were rinsed and incubated with mouse monoclonal β1-integrin receptor antibody, rabbit polyclonal phospho-ERK1/2 antibody, and rabbit polyclonal FAK antibody for 1 h at room temperature, washed three times with PBS, and incubated with fluorescent (fluorescein isothiocyanate) anti-mouse or anti-rabbit secondary antibody (BD Pharmingen, San Diego, CA, USA) for 60 min in the dark. After washing, the nuclei were stained with Hoechst 33342 (2 μg/mL) and analyzed using fluorescent microscopy imaging. Cells were imaged with a BD Pathway 855 confocal system using a 40× (0.75 NA) objective. Cell populations were analyzed for cytoplasmic/nuclear fluorescence intensity. Images of fluorescein isothiocyanate-labeled cells were acquired using a 488/10 excitation laser and a 515LP emission laser.
Statistical analysis
In all experiments, the mean values for three assays ± SD were calculated. The results were submitted to statistical analysis using the unpaired, one-sided Student’s t-test, considering P<0.05 as significant.
Results
The median number of PLTs per cubic millimeter was 990.2±329 and 224.7±72.6 for PRP and VB, respectively. The median concentration of WBCs per cubic millimeter was 12.73±5.4 and 5.74±1.0 for PRP and VB, respectively. The median number of RBCs per cubic millimeter was 1,204±960 and 4,500±860 for PRP and VB, respectively. The samples of SPRP and SActi-PRP did not show the presence of cellular elements.
Collagen biosynthesis was measured in confluent human dermal fibroblasts treated with 1% and 5% of PRP, SPRP, and a SActi-PRP. As can be seen in Figure 1, 24 h incubation of the fibroblasts with 1% and 5% PRP contributed to increase collagen biosynthesis by about 350% and 550%, respectively, compared to control. In cells treated with 1% and 5% of SPRP, collagen biosynthesis was increased to about 125% and 200% of the control, respectively. Addition of SActi-PRP to fibroblasts also increased collagen biosynthesis to about 220% (at 1%) and 290% (at 5%), compared to control.
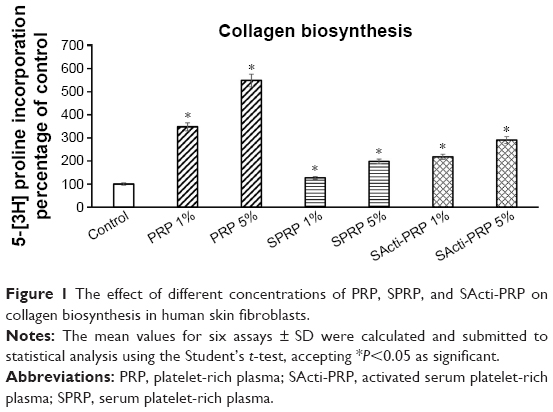 | Figure 1 The effect of different concentrations of PRP, SPRP, and SActi-PRP on collagen biosynthesis in human skin fibroblasts. |
PRP, SPRP, and SActi-PRP were found to stimulate prolidase activity similarly to collagen biosynthesis. As can be seen in Figure 2, PRP at concentrations of 1% and 5% increased the enzyme activity by about 300% and 500%, respectively. In cells treated with 1% and 5% of SPRP, prolidase activity was increased to about 125% and 150% of the control, respectively. Addition of SActi-PRP to fibroblasts also increased prolidase activity to about 150% (at 1%) and 175% (at 5%), compared to control.
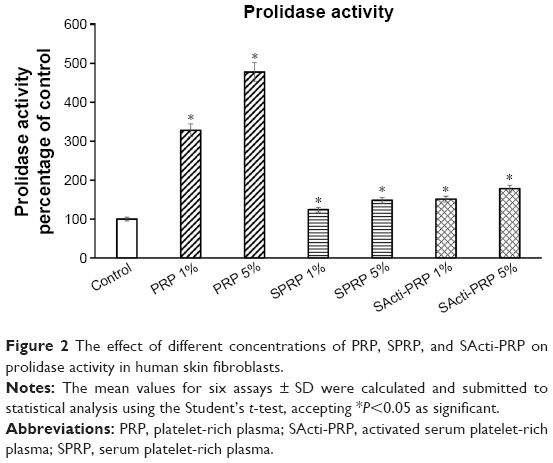 | Figure 2 The effect of different concentrations of PRP, SPRP, and SActi-PRP on prolidase activity in human skin fibroblasts. |
Collagen biosynthesis and prolidase activity are regulated by β1-integrin receptor signaling. In the cells incubated with PRP for 24 h, the expression of β1-integrin receptor was increased in a dose-dependent manner, compared to control (Figure 3). Especially, the effect was pronounced at 5% PRP. However, 5% SActi-PRP also increased the expression of this receptor, but to a much lower extent than PRP. Of interest is the finding that SPRP had no effect on the β1-integrin receptor expression.
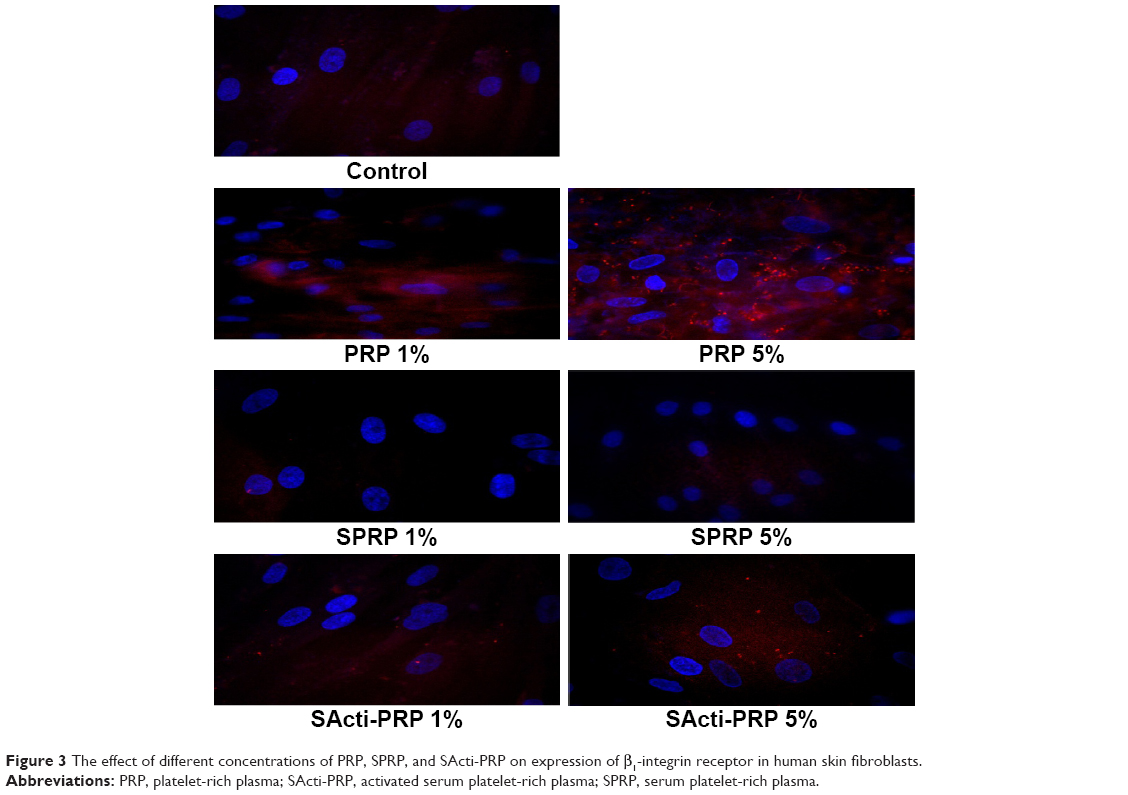 | Figure 3 The effect of different concentrations of PRP, SPRP, and SActi-PRP on expression of β1-integrin receptor in human skin fibroblasts. |
All fractions of blood affected the expression of FAK and phosphorylated MAP-kinases. We found that PRP, SPRP, and SActi-PRP increased the expression of FAK and its translocation to nucleus, (Figure 4) as well as the expression of phosphorylated MAP-kinases (Figure 5). PRP was found to be the most effective stimulator of the expression of tested kinases.
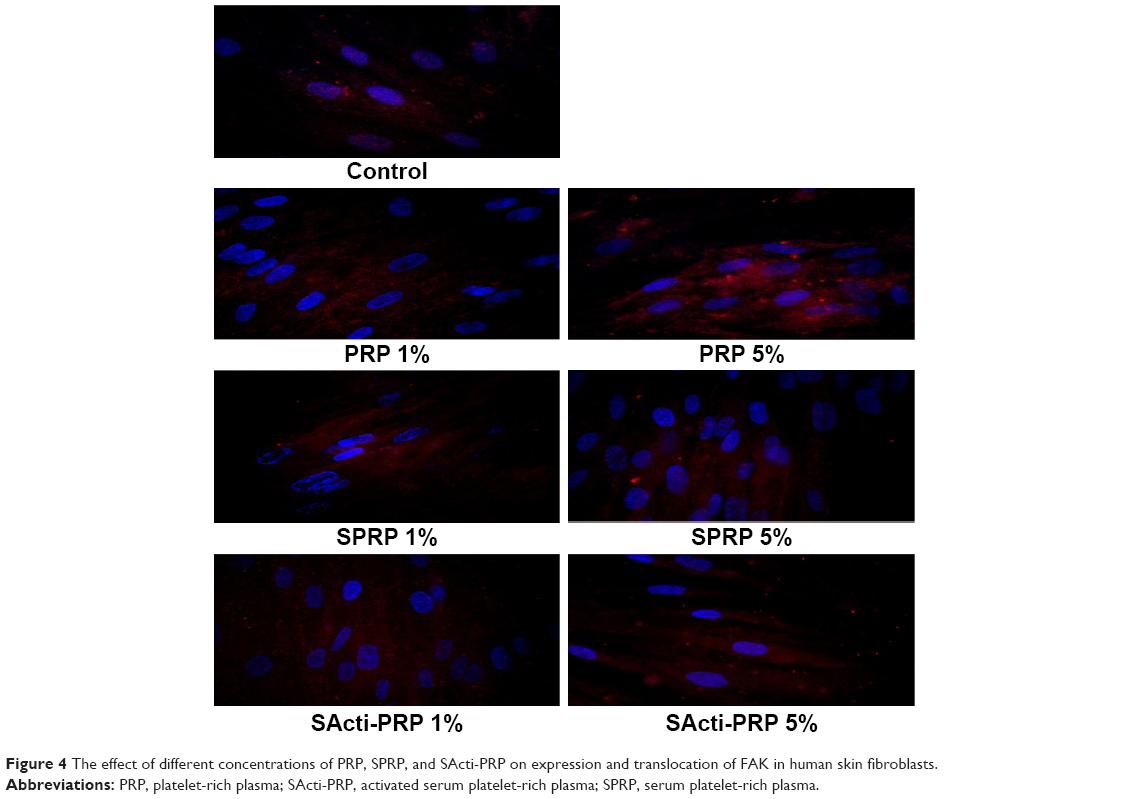 | Figure 4 The effect of different concentrations of PRP, SPRP, and SActi-PRP on expression and translocation of FAK in human skin fibroblasts. |
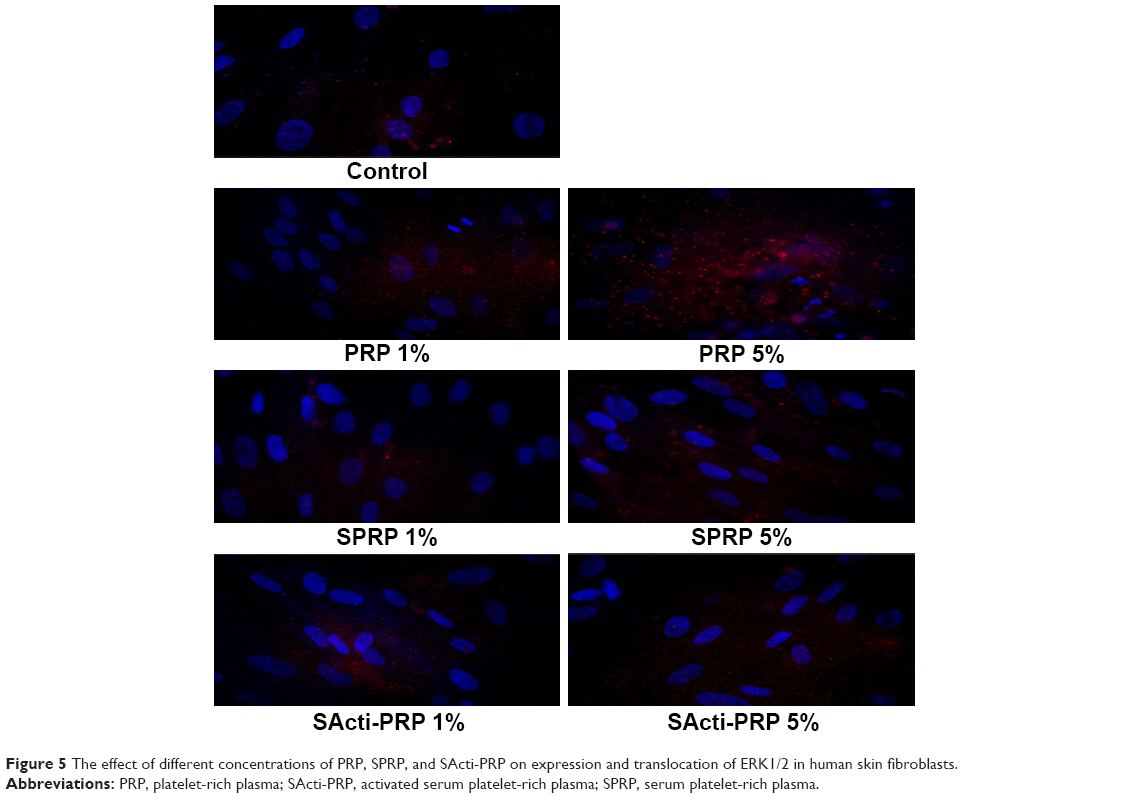 | Figure 5 The effect of different concentrations of PRP, SPRP, and SActi-PRP on expression and translocation of ERK1/2 in human skin fibroblasts. |
DNA biosynthesis was measured in confluent cells treated with 1% and 5% of PRP, SPRP, and SActi-PRP. As can be seen in Figure 6, 24 h incubation of the fibroblasts with 1% and 5% PRP contributed to increase DNA biosynthesis by about 125% and 140%, respectively, compared to control. In cells treated with 1% and 5% of SPRP, DNA biosynthesis was at the same level as in the control. Addition of 5% SActi-PRP to fibroblasts increased DNA biosynthesis to about 135%, compared to control.
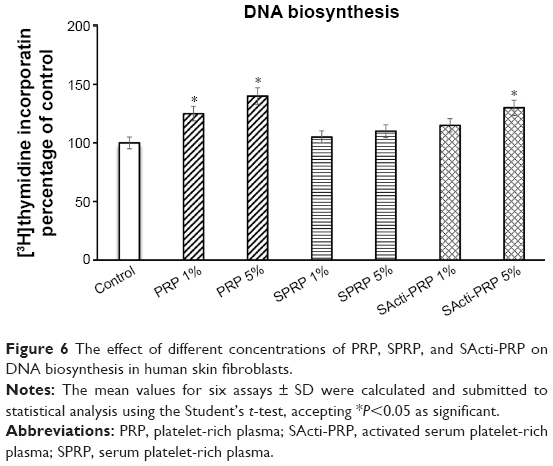 | Figure 6 The effect of different concentrations of PRP, SPRP, and SActi-PRP on DNA biosynthesis in human skin fibroblasts. |
Discussion
In this report, we provide evidence that PRP is the most effective blood fraction (among PRP, SPRP, and SActi-PRP) in stimulation of growth and collagen biosynthesis in cultured fibroblasts. It also represents an in vitro model of tissue response to the blood constituents. This property of PRP is of great interest to regenerative medicine. The first application of PRP is dated to the 1990s in maxillofacial surgery.1 At present, PRP is extensively employed in sport medicine, orthopedics, dentistry, and plastic surgery. The benefit and safety of PRP were confirmed in >5,000 studies on the subjects of wound, bone, tendon, and cartilage healing and skin rejuvenation.30 There are at least three reasons for such broad utilization of the PRP; it is an autogenic material, cheaply acquired, and safe in application. Despite the facts, and apparent widespread use, there is a lack of high-level evidence regarding randomized clinical trials assessing the efficacy of PRP in treating musculoskeletal disorders. There are some aspects of PRP that need to be determined, namely, methods of obtaining and activation. Depending on the preparation protocols, the PRP may contain different concentrations of PLTs and other cell types which can participate in tissue healing processes. Previous studies in humans have reported that the ideal PRP product should have anywhere from a 4- to 7-fold increase in PLTs.18,31 However, PLT concentration is not the only important component of a PRP product. Inclusion or exclusion of mononuclear cells, neutrophils, and RBCs not only define an autologous PLT product, but also have been reported to affect the clinical efficacy of the product and play a major role in influencing the inflammatory responses after PRP injection.32–36 In general, it is believed that RBCs and neutrophils should be reduced as they produce an inflammatory effect, while the effect of mononuclear cells remains largely unknown.35–39
The question we raised in this study was: which constituent of PRP is responsible for the effectiveness in growth and collagen-promoting activity? PRP can potentially regenerate tissue by multiple mechanisms. The most self-evident and most frequently described mechanism of PRP application is its influence on tissue metabolism through the GFs. GFs are released from the α-granules during PLTs’ activation process. However, despite the evident impact of the GFs on tissue regenerating processes, they appear not to be the only factors that activate cell anabolism. Presently, attention is focused on PLTs. It is well known that the life of a PLT is only about 5–7 days, but >95% of the presynthesized GFs are released within 1 h from the beginning of thrombosis. Since the protein release could last an hour, the half-life of any GF would be a few minutes. If GFs are not immediately used upon release from PLTs, they might be degraded before they reach the cell surface receptors. For this reason, we suggest that other PRP constituents may play a significant role in the processes of regeneration.
PRP also contains structural constituents of extracellular matrix (ECM), such as fibronectin, vitronectin, and thrombospondin, and also some fibrin and proinflammatory cytokines (eg, interleukin (IL)-1, IL-6, tumor necrosis factor alpha). All of them are known to act as cell adhesion molecules which are important for the migration of connective tissue cells. PRP sample also consists of serotonin, histamine, dopamine, adenine, and thrombin. Thrombin itself is not only a mediator of coagulation and fibrinolysis, but also primarily acts as a signaling molecule for PLTs, epithelium cells, macrophages, leukocytes, fibroblasts, and myocytes.
Our samples of PRP, unlike SPRP and SActi-PRP, also contain WBCs and RBCs. The importance of leukocytes in immunology is commonly known, but in recent literature, no difference in antibacterial activity is seen between PRP preparations with high leukocyte concentration and those with no leukocytes.39 Furthermore, it has not been proven that leukocytes play a key role in the activation of PLTs by releasing GFs and cytokines; however, they do drastically increase the level of proinflammatory IL-1β.40 Proinflammatory cytokines released from WBCs are the source of an inflammatory process necessary to start wound healing. Release of ECM-degrading enzymes occurs during the inflammatory process, and prolidase is one among them. Prolidase activity is mainly detected in RBCs, WBCs, and in various organs such as liver, brain, heart, uterus, and thymus.41 The prolidase enzyme has been implicated in delayed wound healing because it plays a vital role in inflammatory and angiogenic signaling pathways that regulate matrix degradation and collagen turnover.42 Increased prolidase activity in PRP, as compared to SPRP and SActi-PRP, may result from the release of the enzyme from RBCs and WBCs and from the inflammation mitigated by proinflammatory cytokines. The analysis of DNA biosynthesis, collagen biosynthesis, and prolidase activity suggests that PRP-dependent increase in cell proliferation is not responsible for increase in collagen biosynthesis and prolidase activity. The effect of PRPs on cell proliferation was much lower than on collagen biosynthesis and prolidase activity. Moreover, the data on collagen biosynthesis were normalized to the cell number, and on prolidase activity to the protein concentration.
The controlled clinical level I study, examining the efficiency of PRP containing WBCs in the treatment of chronic tendinopathy, demonstrated a significant improvement in clinical outcomes, as compared to corticosteroids.43 This proves that the anti-inflammatory treatment in tissue repair is less effective than the anabolic effects caused by the PRP, with a proinflammatory effect resulting from the presence of WBCs.
Analyzing the impact of RBCs on tissue healing, there is evidence that the synoviocytes treated with RBC concentrate demonstrate significant increase of cell death.35 In contrast, there is no data relating to small amounts of RBCs in the PRP preparation. Erythrocytes can not only be a potential source of prolidase, but also play an essential role in thrombin generation;44 also, it is well known that thrombin is the agonist of β1-integrin receptor. These results underline the complexity of GF and cell interactions in PLT concentrates and the importance of understanding the mechanisms governing the cellular process during wound healing.
In this report, we suggest that the complex of factors contained in PRP, including GFs and integrin receptor ligands, is required for maximal growth and collagen biosynthesis-promoting activity in fibroblasts. Fibroblasts are involved in the development and function of virtually all organs since they synthesize and maintain the connective tissue. It was established that the regulatory mechanism of experimental wound healing observed in tissue culture was also functioning in vivo. In the repair phase of wound healing, fibroblasts migrate toward inflammatory sites, where they proliferate and synthesize the structural elements of the ECM of connective tissues (eg, fibronectin, collagens) as well as certain cytokines and GFs (eg, PDGF, endothelial growth factor, IGF-I). Both the structural elements of ECM, such as ligands for adhesion receptors as well as GFs, are required to coordinate signaling for proper wound healing processes. In fact, it has been documented previously that coupling of insulin-like growth factor receptor and thrombin-activated β1-integrin receptor signaling augmented proliferation and collagen biosynthesis in different cell types.45 In contrast, disintegrins were found to suppress both processes.46,47 Therefore, we suggest that in contrast to SPRP and SActi-PRP, the PRP contains ligands for integrin receptors, for example, thrombin. However, the identification of specific protein in PRP that is involved in the upregulation of integrin signaling is of particular interest and requires further studies. In addition, the presence of WBCs and RBCs in PRP may account for the higher activity of prolidase, thereby increasing the potential of wound healing.
Acknowledgment
Linguistic help by Mr Mark A Nicholson is greatly appreciated. The study was conducted with the use of equipment purchased by Medical University of Bialystok as part of the OP DEP 2007-2013, Priority Axis I.3, contract No POPW.01.03.00-20-008/09 and financially supported by the Medical University of Bialystok, Poland (KNOW Project No.119/KNOW/15), MUB grant number N/ST/ZB/15/001/2214 and N/ST/ZB/17/005/2214.
Disclosure
The authors report no conflicts of interest in this work.
References
Marx RE. Platelet-rich plasma (PRP): what is PRP and what is not PRP? Implant Dent. 2001;10(4):225–228. | ||
Piccin A, Di Pierro AM, Canzian L, et al. Platelet gel: a new therapeutic tool with great potential. Blood Transfus. 2016;25:1–8. | ||
Zhang N, Wu YP, Qian SJ, Teng C, Chen S, Li H. Research progress in the mechanism of effect of PRP in bone deficiency healing. ScientificWorldJournal. 2013;2013:134582. | ||
Zhou Y, Wang JH. PRP treatment efficacy for tendinopathy: a review of basic science studies. Biomed Res Int. 2016;2016:9103792. | ||
Molloy T, Wang Y, Murrell G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003;33(5):381–394. | ||
Blair P, Flaumenhaft R. Platelet alpha-granules: basic biology and clinical correlates. Blood Rev. 2009;23(4):177–189. | ||
Amable PR, Carias RB, Teixeira MV, et al. Platelet-rich plasma preparation for regenerative medicine: optimization and quantification of cytokines and growth factors. Stem Cell Res Ther. 2013;4(3):67. | ||
Akiyama SK, Nagata K, Yamada K. Cell surface receptors for extracellular matrix components. Biochim Biophys Acta. 1990;1031(1):91–110. | ||
Bissel M. How does extracellular matrix direct gene expression? J Theor Biol. 1981;99(1):31–68. | ||
Carey DJ. Control of growth and differentiation of vascular cells by extracellular matrix. Ann Rev Physiol. 1991;53:161–177. | ||
Donjacour AA, Cunha GR. Stromal regulation of epithelial function. Cancer Treat Res. 1991;53:335–364. | ||
Juliano RL, Haskill S. Signal transduction from the extracellular matrix. J Cell Biol. 1993;120(3):577–585. | ||
Seger R, Krebs EG. The MAPK signaling cascade. FEBS J. 1995;9(9):726–735. | ||
Labat-Robert J, Robert L. Interaction between cells and extracellular matrix: signaling by integrins and the elastin-laminin receptor. Prog Mol Subcell Biol. 2000;25:57–70. | ||
Ivaska J, Reunanen H, Westermarck J, Koivisto L, Kahari VM, Heino J. Integrin α2β1 mediates isoform-specific activation of p38 and up-regulation of collagen gene transcription by a mechanism involving the alpha2 cytoplasmic tail. J Cell Biol. 1999;147(2):401–416. | ||
Palka JA, Phang JM. Prolidase activity in fibroblasts is regulated by interaction of extracellular matrix with cell surface integrin receptor. J Cell Biochem. 1997;67(2):166–175. | ||
Mock WL, Green PC, Boyer KD. Specificity and pH dependence acylproline cleavage by prolidase. J Biol Chem. 1990;265(32):19600–19605. | ||
Hsu WK, Mishra A, Rodeo SR, et al. Platelet-rich plasma in orthopaedic applications: evidence-based recommendations for treatment. J Am Acad Orthop Surg. 2013;21(12):739–748. | ||
Marques LF, Stessuk T, Camargo IC, Sabeh Junior N, dos Santos L, Ribeiro-Paes JT. Platelet-rich plasma (PRP): methodological aspects and clinical applications. Platelets. 2015;26(2):101–113. | ||
Abuaf OK, Yildiz H, Baloglu H, Bilgili ME, Simsek HA, Dogan B. Histologic evidence of new collagen formulation using platelet rich plasma in skin rejuvenation: a prospective controlled clinical study. Ann Dermatol. 2016;28(6):718–724. | ||
Liu SH, Yang RS, al-Shaikh R, Lane JM. Collagen in tendon, ligament, and bone healing a current review. Clin Orthop Relat Res. 1995;(318):265–278. | ||
Abou Neel EA, Bozec L, Knowles JC, et al. Collagen – emerging collagen based therapies hit the patient. Adv Drug Deliv Rev. 2013;65(4):429–456. | ||
Vavken P, Saad FA, Murray MM. Age dependence of expression of growth factor receptors in porcine ACL fibroblasts. J Orthop Res. 2010;28(8):1107–1112. | ||
Cheng MY, Johnson VM, Murray MM. The Effects of age and platelet-rich plasma on ACL cell viability and collagen gene expression. J Orthop Res. 2012;30(1):79–85. | ||
Rodriguez IA, Growney Kalaf EA, Bowlin GL, Sell SA. Platelet-rich plasma in bone regeneration: engineering the delivery for improved clinical efficacy. Biomed Res Int. 2014;2014:392398. | ||
Myara I, Charpentier C, Lemonnier A. Optimal conditions for prolidase assay by proline colorimetric determination: application to imidopeptiduria. Clin Chim Acta. 1982;125(2):193–205. | ||
Lowry OH, Rosebrough NI, Farr AL, Randall IR. Protein measurement with the Folin reagent. J Biol Chem. 1951;193(1):265–275. | ||
Oyamada I, Pałka J, Schalk EM, Takeda K, Peterkofsky B. Scorbutic and fasted guinea pig sera contain an insulin-like growth factor I reversible inhibitor of proteoglycan and collagen synthesis in chick embryo chondrocytes and adult human skin fibroblasts. Arch Biochem Biophys. 1990;276(1):85–93. | ||
Peterkofsky B, Chojkier M, Bateman J. Determination of collagen synthesis in tissue and cell culture system. In: Furthmay M, editor. Immunochemistry of the Extracellular Matrix. Boca Raton, FL: CRC Press; 1982:19–47. | ||
Rajabi H, Sheikhani Shahin H, Norouzian M, Mehrabani D, Dehghani Nazhvani S. The healing effects of aquatic activities and allogenic injection of Platelet-Rich Plasma (PRP) on injuries of achilles tendon in experimental rat. World J Plast Surg. 2015;4(1):66–73. | ||
Pelletier MH, Malhotra A, Brighton T, Walsh WR, Linderman R. Platelet function and constituents of platelet rich plasma. Int J Sports Med. 2013;34(1):74–80. | ||
McLellan J, Plevin S. Does it matter which platelet-rich plasma we use? Equine Vet Educ. 2011;23(2):101–104. | ||
Dragoo JL, Braun HJ, Durham JL, et al. Comparison of the acute inflammatory response of two commercial platelet-rich plasma systems in healthy rabbit tendons. Am J Sports Med. 2012;40(6):1274–1281. | ||
McCarrel TM, Minas T, Fortier LA. Optimization of leukocyte concentration in platelet-rich plasma for the treatment of tendinopathy. J Bone Joint Surg Am. 2012;94(19):1–8. | ||
Braun HJ, Kim HJ, Chu CR, Dragoo JL. The effect of platelet-rich plasma formulations and blood products on human synoviocytes. Am J Sports Med. 2014;42(5):1204–1210. | ||
Sundman EA, Cole BJ, Fortier LA. Growth factor and catabolic cytokine concentrations are influenced by the cellular composition of platelet-rich plasma. Am J Sports Med. 2013;39(10):2135–2140. | ||
Cavallo C, Filardo G, Mariani E, et al. Comparison of platelet-rich plasma formulations for cartilage healing. J Bone Joint Surg Am. 2014;96(5):423–429. | ||
Boswell SG, Schnabel LV, Mohammed HO, Sundman EA, Minas T, Fortier LA. Increasing platelet concentrations in leukocyte-reduced platelet-rich plasma decrease collagen gene synthesis in tendons. Am J Sports Med. 2013;42(1):42–49. | ||
Intravia J, Allen DA, Durant TJ, et al. In vitro evaluation of the anti-bacterial effect of two preparations of platelet rich plasma compared with cefazolin and whole blood. Muscles Ligaments Tendons J. 2014;4(1):79–84. | ||
Anitua E, Zalduendo MM, Alkhraisat MH, Orive G. Release kinetics of platelet-derived and plasma-derived growth factors from autologous plasma rich in growth factors. Ann Anat. 2013;195(5):461–466. | ||
Kaleli S, Kotan D, Akdogan M, Ceylan M, Yalcin A. Serum prolidase activity as a biomarker for choroid plexus calcification. Tohoku J Exp Med. 2016;238(3):255–259. | ||
Dunn R, Varigos G, Winship I. A photographic essay of prolidase deficiency. Clin Dysmorphol. 2011;20(4):194–199. | ||
Gosens T, Peerbooms JC, van Laar W, den Oudsten BL. Ongoing positive effect of platelet-rich plasma versus corticosteroid injection in lateral epicondylitis: a double-blind randomized controlled trial with 2-year follow-up. Am J Sports Med. 2011;39(6):1200–1208. | ||
Whelihan MF, Zachary V, Orfeo T, Kenneth G, Mann KG. Prothrombin activation in blood coagulation: the erythrocyte contribution to thrombin generation. Blood. 2012;120(18):3837–3845. | ||
Surazyński A, Sienkiewicz P, Wołczyński S, Pałka J. Differential effects of echistatin and thrombin on collagen production and prolidase activity in human dermal fibroblasts and their possible implication in beta1-integrin-mediated signaling. Pharmacol Res. 2005;51(3):217–221. | ||
Miltyk W, Surazyński A, Wołczyński S, Pałka JA. Combined therapy with disintegrin and melphalan as a new strategy in inhibition of endometrial cancer cell line (Ishikawa) growth. Folia Histochem Cytobiol. 2009;47(5):121–125. | ||
Kononczuk J, Surazynski A, Czyzewska U, et al. αIIbβ3-integrin ligands: abciximab and eptifibatide as proapoptotic factors in MCF-7 human breast cancer cells. Curr Drug Targets. 2015;16(13):1429–1437. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.