Back to Journals » Neuropsychiatric Disease and Treatment » Volume 18
Differences in mGluR5 Availability Depending on the Level of Social Avoidance in Drug-Naïve Young Patients with Major Depressive Disorder
Authors Kim JH ![]() , Joo YH, Son YD
, Joo YH, Son YD ![]() , Kim HK, Kim JH
, Kim HK, Kim JH ![]()
Received 21 June 2022
Accepted for publication 3 September 2022
Published 12 September 2022 Volume 2022:18 Pages 2041—2053
DOI https://doi.org/10.2147/NDT.S379395
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Taro Kishi
Jeong-Hee Kim,1,2 Yo-Han Joo,1 Young-Don Son,1– 3 Hang-Keun Kim,1– 3 Jong-Hoon Kim1,3,4
1Neuroscience Research Institute, Gachon University, Incheon, Republic of Korea; 2Department of Biomedical Engineering, College of Health Science, Gachon University, Incheon, Republic of Korea; 3Gachon Advanced Institute for Health Sciences & Technology, Gachon University, Incheon, Republic of Korea; 4Department of Psychiatry, Gachon University College of Medicine, Gil Medical Center, Incheon, Republic of Korea
Correspondence: Jong-Hoon Kim, Department of Psychiatry, Gachon University College of Medicine, Gil Medical Center, 1198 Guwol-dong, Namdong-gu, Incheon, 21565, Republic of Korea, Tel +82 32 460 2696, Fax +82 32 472 8813, Email [email protected] Young-Don Son, Department of Biomedical Engineering, College of Health Science, Gachon University, 191 Hambakmoe-ro, Yeonsu-gu, Incheon, 21936, Republic of Korea, Tel +82 32 820 4416, Email [email protected]
Background: Previous research has shown that metabotropic glutamate receptor-5 (mGluR5) signaling is significantly involved in social avoidance. We investigated the relationship between levels of social avoidance and mGluR5 availability in drug-naïve young patients with major depressive disorder (MDD).
Methods: Twenty non-smoking patients and eighteen matched non-smoking healthy controls underwent [11C]ABP688 positron emission tomography (PET) and magnetic resonance imaging scans. The binding potential (BPND) of [11C]ABP688 was obtained using the simplified reference tissue model. Patients’ level of social avoidance was assessed using the Social Avoidance and Distress Scale (SADS). For [11C]ABP688 BPND, the region-of-interest (ROI)-based between-group comparisons and correlations with SADS scores were investigated. The frontal cortices were chosen as a priori ROIs based on previous PET investigations in MDD, and on literature underscoring the importance of the frontal cortex in social avoidance.
Results: Independent samples t-tests revealed no significant differences in [11C]ABP688 BPND in the frontal cortices between the MDD patient group as a whole and healthy controls. One-way analysis of variance with post-hoc tests revealed significantly lower BPND in the bilateral superior frontal cortex (SFC) and left middle frontal cortex (MFC) in MDD patients with low levels of social avoidance (L-SADS) than in healthy controls. The L-SADS patients also had significantly lower BPND in the medial part of the right SFC than both MDD patients with high levels of social avoidance (H-SADS) and healthy controls. The L-SADS patients also showed significantly lower BPND in the orbital parts of the SFC, MFC, and inferior frontal cortex than H-SADS patients. No significant group differences were found between H-SADS patients and healthy controls. The ROI-based correlation analysis revealed significant positive correlations between social avoidance levels and frontal [11C]ABP688 BPND in the entire patients.
Conclusion: Our exploratory study shows significant differences in frontal mGluR5 availability depending on the level of social avoidance in drug-naïve non-smoking MDD patients, suggesting that social avoidance should be considered as one of the clinical factors involved in mGluR5 signaling changes in depression.
Keywords: social avoidance, metabotropic glutamate receptor-5, positron emission tomography, [11C]ABP688, major depressive disorder
Introduction
Rodent studies have reported that metabotropic glutamate receptor-5 (mGluR5) antagonists or negative allosteric modulators exhibit antidepressant effects.1–3 Moreover, reduced mGluR5 densities were found in the postmortem brains of patients with major depressive disorder (MDD)4 and in a rodent model of depression.5 These findings suggest that mGluR5-mediated neurotransmission plays a role in the neurobiology of MDD.
However, the results of in vivo mGluR5 positron emission tomography (PET) studies in patients with MDD have been inconclusive. Two studies have demonstrated significantly lower levels of mGluR5 in patients than in controls in the frontal cortex or in the clusters within the frontal, temporal, and parietal cortices,4,6 while two other studies have revealed no significant changes in mGluR5 availability in patients with MDD compared to control subjects.7,8 These inconsistent results may reflect the clinical heterogeneity of MDD.9
One of the important clinical features associated with the heterogeneity of MDD is social avoidance or fear.10 Patients with the co-occurrence of MDD and social phobia are more likely to report agitation, sleep disturbance, and increased functional impairment.10,11 Other studies have shown that the clinical features of MDD and comorbid social avoidance or fear had similarities with those of atypical depression.12,13
Notably, preclinical studies have shown that mGluR5 signaling is significantly involved in fear conditioning and avoidance. The allosteric mGluR5 antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP) has been shown to impair social fear extinction in a mouse model.14 MPEP also prevents the acquisition and expression of conditioned fear in rodents15 and impairs the acquisition of auditory and contextual fear conditioning in a dose-dependent manner.16 Xu et al17 also reported that the acquisition of fear conditioning and the extinction of both contextual and auditory fear were impaired in mGluR5 knock-out mice. Fenobam, another negative allosteric modulator of mGluR5, was reported to reduce the freezing response in contextual fear conditioning.18 Although the mechanisms and molecular underpinnings involved in social fear conditioning are unclear, preclinical studies have suggested that mGluR5 plays an important role in these learning processes, which are particularly relevant in human conditions of social avoidance or fear.9,17 In addition, cerebral regions that play a crucial role in fear extinction and reversal tasks involve the frontal cortex,19 where mGluR5 is highly expressed.20
Based on these preclinical studies, we hypothesized that in vivo mGluR5 availability would differ in patients with MDD according to the levels of social avoidance and classified the patients into two groups with low and high levels of social avoidance. In the present study, we quantified mGluR5 availability using PET with [11C]ABP688, a specific radioligand that binds to mGluR5, in drug-naïve young patients with MDD and investigated the differences in mGluR5 availability between the groups. We also examined the correlations between the levels of social avoidance and mGluR5 availability in MDD patients. The frontal cortices were chosen as a priori regions-of-interest (ROIs) based on previous in vivo PET investigations in patients with MDD,4,6 in which significantly low levels of mGluR5 were observed; and based on the literature, which underscores the importance of the frontal cortex in the mechanisms underlying the expression of social avoidance.21
Materials and Methods
Participants
The study was approved by the Institutional Review Board of the Gachon University Gil Medical Center, and all study procedures were performed in accordance with international ethical standards and the Declaration of Helsinki. All participants provided written informed consent after a full explanation of the study proposes and procedures before study participation. Patients were recruited from outpatient clinics, and through advertisements in local posters, and included if they (i) were aged from 19 (legal adult age in South Korea) to 29 years; (ii) met the diagnostic criteria for MDD by the Diagnostic and Statistical Manual of Mental Disorders 4th edition (DSM-IV),22 which was established using the Structured Clinical Interview for DSM-IV (SCID-IV),23 with no other current Axis I diagnosis except for social phobia; (iii) had no past or current substance use/dependence; (iv) had no history of medical or neurological disorders; and (v) had no past or current use of psychotropic medications such as antidepressants, benzodiazepines/anxiolytics, hypnotics, antipsychotics, or mood stabilizers. Patients requiring immediate pharmacotherapy or urgent inpatient treatment were excluded. Twenty patients (7 males and 13 females) who met these criteria were enrolled in the study and all enrolled patients were East Asians (Koreans). The patients’ mean age was 23.2 ± 2.6 years, mean duration of current episode of depression was 9.8 ± 12.5 months, and mean age of onset was 16.6 ± 1.1 years (Table 1). Three patients with comorbid social phobia were included in patient group. Eighteen age- and gender-matched healthy controls (9 males and 9 females), who met the criteria of no past or current psychiatric, neurological, or medical disorders, and no past or current use of medications/substances known to affect the central nervous system, were also recruited through advertisements in local posters and enrolled in the study (Table 1). All patients and healthy control subjects were non-smokers. None of the participants showed any structural abnormalities on brain magnetic resonance imaging (MRI), which was confirmed by a board-certified radiologist.
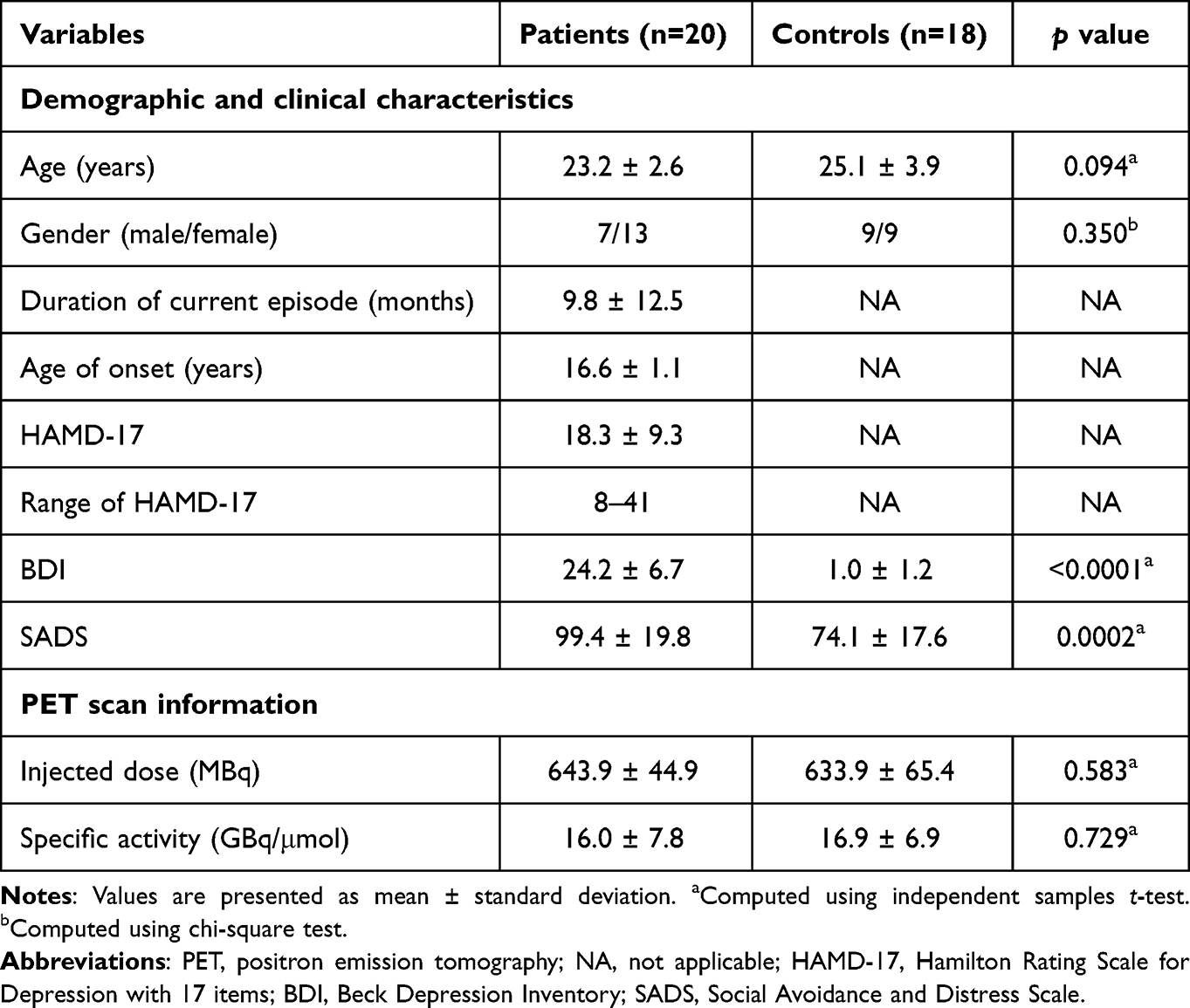 |
Table 1 Demographic/Clinical Characteristics and PET Scan Information |
Clinical Assessments
The severity of depressive symptoms was assessed using the Hamilton Rating Scale for Depression with 17 items (HAMD-17)24 and Beck Depression Inventory (BDI).25 The patients’ mean HAMD-17 score was 18.3 ± 9.3 and mean BDI score was 24.2 ± 6.7 (Table 1). For the HAMD-17 and BDI, higher scores indicate more severe depressive symptoms. The aspects of social avoidance, including avoidance, discomfort, and fear, were evaluated using the Social Avoidance and Distress Scale (SADS).26,27 The SADS is a 28-item questionnaire with each item rated on a 5‐point scale from 1 to 5.27 A higher score indicates greater social avoidance. The standardized Korean version of the SADS27 was used in this study.
Patient Group Classification Based on Levels of Social Avoidance
The highest SADS score in the healthy control group was selected as the cut-off score for classifying patients into two groups according to the levels of social avoidance. Accordingly, 11 out of the 20 patients had low SADS scores and 9 had high SADS scores. Notably, three patients with comorbid social phobia were included in the patient group with high SADS scores. In this study, the two patient groups were classified as MDD patients with low levels of social avoidance (L-SADS) and those with high levels of social avoidance (H-SADS). Mean SADS scores of the L-SADS patient group and the H-SADS patient group were 84.6 ± 13.5 (range: 61–97) and 117.3 ± 6.1 (range: 106–126), respectively (Table 2).
 |
Table 2 Demographic/Clinical Characteristics and PET Scan Information for L-SADS Patients, H-SADS Patients, and Healthy Controls |
Scan Protocol
PET scans were performed using a Biograph 6 positron emission tomography/computed tomography (PET/CT) scanner (Siemens Medical Imaging Systems, Knoxville, USA) with [11C]ABP688 at the same time of day (10:00 a.m.) to minimize possible diurnal variations in mGluR5.28,29 For attenuation correction, the CT-based transmission scan was conducted before [11C]ABP688 injection. Following bolus injection of a mean dose of 639.2 ± 55.0 MBq [11C]ABP688 with a mean specific activity of 16.4 ± 7.3 GBq/μmol, dynamic emission data were acquired in list mode for 60 min. These emission data were reconstructed using the 2-dimensional ordered-subset expectation maximization (OSEM-2D) algorithm into 21 frames of the following duration: 2 × 15 s, 3 × 30 s, 3 × 60 s, 2 × 90 s, 3 × 120 s, 2 × 180 s, 4 × 300 s, and 2 × 600 s. The reconstructed PET frames had a matrix size of 256 × 256 × 109 and a voxel size of 1.33 × 1.33 × 1.50 mm3. Corrections for attenuation, random and scatter coincidences, detector dead-time, decay, and detector normalization were applied to these frames.
All participants were scanned using a 3-Tesla MRI scanner (Magnetom Verio; Siemens, Erlangen, Germany) with a 12-channel transmit/receive phased array head coil (iPAT, Siemens, Erlangen, Germany). The structural MRI data were obtained using a 3-dimensional T1-weighted magnetization-prepared rapid gradient echo (3D T1MPRAGE) sequence with the following scan parameters: repetition time = 1900 ms, echo time = 3.3 ms, inversion time = 900 ms, flip angle = 9°, voxel size = 0.5 × 0.5 × 1.0 mm3, matrix size = 416 × 512, and number of slices = 160. To minimize head movement during the MRI and PET scans, all participants’ heads were fixed to the sponges as comfortably as possible.
Image Analysis
The spatial preprocessing step of [11C]ABP688 PET was performed using Statistical Parametric Mapping 12 (SPM12; The Wellcome Centre for Human Neuroimaging, London, UK; www.fil.ion.ucl.ac.uk). Realignment was performed to correct the motion of all reconstructed PET frames. For each subject, the structural MRI image was coregistered to the mean PET image obtained from the realignment step. The coregistered structural MRI images were spatially normalized to the standard anatomical space using the Montreal Neurological Institute (MNI) template with the nonlinear deformation field, and the estimated transform was applied to the corresponding PET frames.
Based on the parameter estimation implemented in the PMOD software v3.8 (PMOD Technologies Ltd., Zürich, Switzerland), the non-displaceable binding potential (BPND) of [11C]ABP688 was derived from kinetic modeling using the simplified reference tissue model 2 (SRTM2)30 with cerebellar gray matter as a reference region as previously proposed for this tracer.6,7,28,31–34 During the [11C]ABP688 BPND calculation process, regional time-activity curves were extracted from spatially normalized PET frames by averaging all the voxels within each ROI defined based on structural T1 MRI image. [11C]ABP688 BPND values were obtained from the 26 predefined ROIs in the frontal cortex using the automated anatomical labeling (AAL) atlas,35 which included the superior frontal cortex (SFC), middle frontal cortex (MFC), opercular part of the inferior frontal cortex (IFCoperc), triangular part of the inferior frontal cortex (IFCtriang), medial part of the superior frontal cortex (SFCmedial), supplementary motor area (SMA), paracentral lobule (PCL), orbital part of the superior frontal cortex (SFCorb), medial orbital part of the superior frontal cortex (SFCventmed), orbital part of the middle frontal cortex (MFCorb), orbital part of the inferior frontal cortex (IFCorb), gyrus rectus (REC), and olfactory cortex (OLF). The left and right brain regions were analyzed separately, given their structural and functional asymmetry and deficits in MDD.36,37 Representative mean images of [11C]ABP688 BPND, PET, and 3-Tesla MRI in controls are shown in Figure 1.
 |
Figure 1 Representative mean images of [11C]ABP688 BPND, PET, and 3-Tesla MRI for 18 healthy controls. The [11C]ABP688 PET image is generated by averaging the reconstructed 21 frames of dynamic image. Abbreviations: BPND, non-displaceable binding potential; PET, positron emission tomography; MRI, magnetic resonance imaging. |
Statistical Analysis
Statistical analyses were performed using the Statistical Package for the Social Sciences (SPSS) v23.0 (IBM Corp., Armonk, NY, USA). Levene’s test was performed to assess the equality of variances for [11C]ABP688 BPND in each ROI, and p values less than 0.05 were considered significant. For [11C]ABP688 BPND in the frontal regions, independent samples t-tests were used to compare the mean values between whole MDD patients and healthy controls. Either one-way analysis of variance (ANOVA) with Bonferroni post-hoc test or Welch’s ANOVA with Games-Howell post-hoc test was used when three groups (ie, L-SADS patients, H-SADS patients, and healthy controls) were compared. Considering the exploratory perspective of the ROI-based between-group comparison analyses, when no significant group differences were found at the Bonferroni-corrected p < 0.0019 (0.05/26), significant results were identified at the threshold of the two-tailed uncorrected p < 0.05. Effect size was also calculated for one-way ANOVA or Welch’s ANOVA.
In the entire patient group, the ROI-based Pearson’s correlation analyses were performed to investigate the relationship between the SADS scores and [11C]ABP688 BPND values in the frontal regions with significant group differences. In the analyses, a two-tailed p < 0.05 was considered significant.
Results
The demographic/clinical characteristics and PET scan information for the patient and healthy control groups are summarized in Table 1. There were no significant group differences in age, gender distribution, or PET scan parameters (Table 1). Patient subgroups (ie, L-SADS patients and H-SADS patients) did not differ significantly in terms of the aforementioned variables, duration of the current episode, age of onset, HAMD-17 score, or BDI score (Table 2). The SADS score was significantly higher in the H-SADS patient group than in both the L-SADS patient and healthy control groups (Table 2). There was no significant difference in the SADS score between the L-SADS patient group and healthy control group (Table 2).
Between-Group Comparisons for in vivo mGluR5 Availability
An independent samples t-test revealed that there were no significant differences between entire MDD patients and healthy controls in all subregions of the frontal cortex (uncorrected p > 0.05).
Between-group comparisons with one-way ANOVA or Welch’s ANOVA revealed significant group differences in mGluR5 availability in the bilateral SFC (right: effect size = 0.49, uncorrected p = 0.031, left: effect size = 0.50, uncorrected p = 0.021), bilateral MFC (right: effect size = 0.45, uncorrected p = 0.041, left: effect size = 0.49, uncorrected p = 0.024), right IFCoperc (effect size = 0.44, uncorrected p = 0.043), bilateral SFCmedial (right: effect size = 0.52, uncorrected p = 0.014, left: effect size = 0.44, uncorrected p = 0.045), left SMA (effect size = 0.44, uncorrected p = 0.045), bilateral SFCorb (right: effect size = 0.47, uncorrected p = 0.029, left: effect size = 0.45, uncorrected p = 0.038), bilateral SFCventmed (right: effect size = 0.43, uncorrected p = 0.049, left: effect size = 0.44, uncorrected p = 0.045), bilateral MFCorb (right: effect size = 0.55, uncorrected p = 0.009, left: effect size = 0.49, uncorrected p = 0.023), and bilateral IFCorb (right: effect size = 0.47, uncorrected p = 0.032, left: effect size = 0.48, uncorrected p = 0.027) (Table 3). Pairwise group comparisons with Bonferroni or Games-Howell post-hoc test showed that the L-SADS patients had significantly lower [11C]ABP688 BPND in the bilateral SFC (right: uncorrected p = 0.024, left: uncorrected p = 0.029) and left MFC (uncorrected p = 0.032) compared to healthy controls (Table 3; Figure 2). The L-SADS patients also showed significantly lower [11C]ABP688 BPND in the right SFCmedial than both the H-SADS patients (uncorrected p = 0.042) and healthy controls (uncorrected p = 0.024) (Table 3; Figure 2). The L-SADS patients also had significantly lower [11C]ABP688 BPND in the bilateral SFCorb (right: uncorrected p = 0.040, left: uncorrected p = 0.048), bilateral MFCorb (right: uncorrected p = 0.008, left: uncorrected p = 0.037), and bilateral IFCorb (right: uncorrected p = 0.034, left: uncorrected p = 0.032) compared to the H-SADS patients (Table 3; Figure 2). No significant group differences were found between H-SADS patients and healthy controls (Table 3; Figure 2).
 |
Table 3 The ROI-Based Between-Group Comparisons of [11C]ABP688 BPND |
 |
Figure 2 Box plots of the ROI-based between-group comparisons in [11C]ABP688 BPND values. (A–C) The L-SADS patient group had significantly lower [11C]ABP688 BPND in the bilateral SFC (right: uncorrected p = 0.024, left: uncorrected p = 0.029) and left MFC (uncorrected p = 0.032) compared to control group. (D) In addition, the L-SADS patient group showed significantly lower [11C]ABP688 BPND in the right SFCmedial relative to both the H-SADS patient group (uncorrected p = 0.042) and control group (uncorrected p = 0.024). (E–J) Moreover, the L-SADS patient group had significantly lower [11C]ABP688 BPND in the bilateral SFCorb (right: uncorrected p = 0.040, left: uncorrected p = 0.048), bilateral MFCorb (right: uncorrected p = 0.008, left: uncorrected p = 0.037), and bilateral IFCorb (right: uncorrected p = 0.034, left: uncorrected p = 0.032) compared with the H-SADS patient group. In box plots, the statistical significance is indicated as *Uncorrected p < 0.05 and **Uncorrected p < 0.01. Abbreviations: ROI, region of interest; BPND, non-displaceable binding potential; L-SADS, depressed patients with low levels of social avoidance; SFC, superior frontal cortex; MFC, middle frontal cortex; SFCmedial, superior frontal cortex (medial part); H-SADS, depressed patients with high levels of social avoidance; SFCorb, superior frontal cortex (orbital part); MFCorb, middle frontal cortex (orbital part); IFCorb, inferior frontal cortex (orbital part). |
Relationship Between Levels of Social Avoidance and in vivo mGluR5 Availability
In the entire patients, the ROI-based Pearson’s correlation analyses showed significant positive correlations between the SADS score and the [11C]ABP688 BPND in the right SFCmedial (r = 0.450, p = 0.046), bilateral SFCorb (right: r = 0.458, p = 0.042, left: r = 0.463, p = 0.040), bilateral MFCorb (right: r = 0.499, p = 0.025, left: r = 0.451, p = 0.046), and bilateral IFCorb (right: r = 0.450, p = 0.046, left: r = 0.503, p = 0.024) (Figure 3). There was a tendency of positive correlations in the bilateral SFC (right: r = 0.389, p = 0.090, left: r = 0.396, p = 0.084) and left MFC (r = 0.401, p = 0.080).
 |
Figure 3 Scatter plots of the correlations between the SADS score and [11C]ABP688 BPND in the entire MDD patient group. The SADS score had significant positive correlations with [11C]ABP688 BPND in the (A) right SFCmedial (r = 0.450, p = 0.046), (B and C) bilateral SFCorb (right: r = 0.458, p = 0.042, left: r = 0.463, p = 0.040), (D and E) bilateral MFCorb (right: r = 0.499, p = 0.025, left: r = 0.451, p = 0.046), and (F and G) bilateral IFCorb (right: r = 0.450, p = 0.046, left: r = 0.503, p = 0.024). Yellow and blue dots indicate the L-SADS patient group and the H-SADS patient group, respectively. †Cross marks indicate MDD patients with comorbid social phobia. The solid and dotted lines indicate the regression lines and 95% confidence intervals, respectively. Abbreviations: SADS, Social Avoidance and Distress Scale; BPND, non-displaceable binding potential; MDD, major depressive disorder; SFCmedial, superior frontal cortex (medial part); SFCorb, superior frontal cortex (orbital part); MFCorb, middle frontal cortex (orbital part); IFCorb, inferior frontal cortex (orbital part); L-SADS, depressed patients with low levels of social avoidance; H-SADS, depressed patients with high levels of social avoidance. |
Discussion
In this study, we found significantly lower mGluR5 availability in the frontal regions in the L-SADS patients than in both H-SADS patients and healthy controls. We also observed that the H-SADS patient group had no significant differences in mGluR5 availability in the frontal regions compared to the healthy control group. Furthermore, in the entire patient group, significant positive correlations were found between the levels of social avoidance and mGluR5 availability in certain frontal regions. To the best of our knowledge, this is the first PET study to explore the relationship between the levels of social avoidance and in vivo mGluR5 availability in patients with MDD.
As mentioned earlier, we classified the patient group into the L-SADS and H-SADS groups based on the SADS score. Compared to the healthy control group, the L-SADS patient group had similar levels of social avoidance but significantly lower mGluR5 availability in the frontal cortex, whereas the H-SADS patient group showed no significant differences in mGluR5 availability in the frontal cortex compared to the healthy control group. These results indicate that mGluR5 availability increases in the frontal cortex as levels of social avoidance increase within patient groups, which is consistent with our hypothesis that mGluR5 availability in the frontal regions would differ in patients with MDD according to the levels of social avoidance. In addition, these results suggest that the level of social avoidance affects changes in mGluR5 availability in the frontal cortical regions in MDD.
The molecular underpinnings and mechanisms involved in social avoidance in MDD remain unclear; however, there is a growing body of evidence that group I mGlu receptors, such as mGluR5, are important in the physiological and behavioral sequelae associated with stressful social stimuli.9,38,39 Several preclinical studies have reported that antagonism14–16,40 and genetic deletion17 of mGluR5 block or reduce conditioned fear. Systemic administration of MPEP also blocks both the acquisition and expression of conditioned fear in a rodent model.15 The mGluR5 blockade may attenuate ionotropic glutamate-mediated postsynaptic excitability, which leads to reduced fear and avoidance.38 These preclinical studies suggest that mGluR5 plays a crucial role in the acquisition and expression of social fear-related memory.38 In addition, other studies have shown that mGluR5 antagonism has anxiolytic effects in animal models41–44 and that mGluR5 suppression normalizes social interactions in mouse models of social deficits.45,46 Furthermore, down-regulation of mGluR5 has been suggested to be associated with lower anxiety symptoms in patients with MDD.9 Based on these studies, we may explain our results as an association between high levels of social avoidance and increased mGluR5 availability in the frontal cortical regions that are critically involved in the pathophysiology of both MDD and social avoidance/fear.39,47
We observed no significant differences in frontal mGluR5 availability between the MDD patient group as a whole and healthy controls. This is likely because of the increased mGluR5 availability in the H-SADS patient subgroup. This result suggests that high levels of social avoidance may be associated with a phenotype that masks a general deficit in mGluR5 availability in MDD and that social avoidance or fear should be considered a clinical factor influencing mGluR5 signal changes in MDD. Our results are in line with those of two previous PET investigations,7,8 in which no significant differences in mGluR5 availability were observed between patients with MDD and healthy control subjects, which may reflect the clinical heterogeneity of MDD. More mGluR5 PET studies on various MDD subgroups according to clinical characteristics are needed for further analyses.
In this study, the frontal cortex was selected as the a priori ROI based on previous in vivo PET investigations in MDD patients,4,6 in which significantly low mGluR5 levels were observed. In addition, this region is a crucial cerebral region in rodent studies of fear-related tasks that examine the role of glutamatergic signaling.19 We found that mGluR5 availability in the right SFCmedial, bilateral SFCorb, bilateral MFCorb, and bilateral IFCorb showed significant differences between the groups and had positive correlations with levels of social avoidance in the entire patient group. These regions are subregions of the prefrontal cortex (PFC), which plays a crucial role in fear conditioning21 and social–emotional processing.48 Previous studies have also suggested that prefrontal dysfunction is associated with social deficits in psychiatric disorders.49–51 Thus, our results are consistent with the role of the PFC in the mechanisms underlying the expression of social avoidance/fear21 and with previous neuroimaging findings reporting prefrontal dysfunction in MDD.52–58 Our PET findings are also in line with a previous report that the PFC is critically involved in top-down neural mechanisms related to social avoidance in the human brain.59 Future multimodal imaging studies with a combination of [11C]ABP688 PET and resting-state or task-related functional MRI may shed light on the downstream effects of altered frontal mGluR5 availability on social avoidance and related functional abnormalities in MDD.
The interpretation of our results should be considered in light of some limitations. First, compared with previous studies,4,7,8,60 the age of the patients in our study was significantly different, but the disease severity and sex distribution were not. In our study, all patients were drug-naïve young adults, with a mean age of 23.2 years. This minimized the confounding effects of age and psychotropic medications on in vivo mGluR5 availability but may not allow generalization of our findings to other groups of patients with MDD. Future studies in an MDD cohort with a wider age range are needed. Second, the relatively small cohort sizes for each group (L-SADS patient group: n = 11, H-SADS patient group: n = 9, healthy control group: n = 18) reduced the statistical power to identify significance in the analyses. In particular, the correlations between the SADS score and [11C]ABP688 BPND in the MDD patient group as a whole might have been driven by differences in mGluR5 availability between the L-SADS and H-SADS patient subgroups. Future studies on a larger cohort of patients with a greater range of SADS scores are necessary to confirm our results. Third, quantification of the [11C]ABP688 BPND was performed using the simplified reference tissue model, rather than the gold standard two-tissue compartment model (2TCM) with the arterial input function.28,61 The arterial input function for 2TCM is measured using invasive methods such as radial artery cannulation, and the resulting discomfort can be a confounding factor in data acquisition. Moreover, inaccuracies in the measured plasma metabolite fractions could be a source of variance in the parameter estimation using 2TCM. In our study, the cerebellar gray matter was used as the reference region based on postmortem studies reporting that specific mGluR5 binding is negligible in this region.4,62,63 However, an in vitro binding study with [18F]F-PEB, another PET radioligand for mGluR5, reported small but measurable mGluR5 signals in the cerebellar gray matter.64 Therefore, our analysis method might have underestimated the mGluR5 BPND values.
Conclusion
Our in vivo PET molecular imaging study using [11C]ABP688 showed significant differences in frontal mGluR5 availability depending on the level of social avoidance in drug-naïve non-smoking MDD patients. These results suggest that social avoidance should be considered as one of the clinical factors involved in mGluR5 signaling changes in depression. Future PET molecular imaging studies are required in various MDD subgroups with specific clinical characteristics to clarify further the role of mGluR5-mediated neurotransmission in MDD.
Acknowledgments
The authors are grateful to all the participants who took part in this study. The authors thank the staff of the cyclotron facility and PET/MRI technologists at Gachon University Neuroscience Research Institute.
Funding
This research was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (2022R1A2C2009580, 2020R1A4A1019623, and 2020R1C1C1012612).
Disclosure
The authors declare no conflicts of interest or financial disclosures for this work.
References
1. Tatarczyńska E, Klodzińska A, Chojnacka-Wójcik E, et al. Potential anxiolytic- and antidepressant-like effects of MPEP, a potent, selective and systemically active mGlu5 receptor antagonist. Br J Pharmacol. 2001;132(7):1423–1430. doi:10.1038/sj.bjp.0703923
2. Li X, Need AB, Baez M, Witkin JM. Metabotropic glutamate 5 receptor antagonism is associated with antidepressant-like effects in mice. J Pharmacol Exp Ther. 2006;319(1):254–259. doi:10.1124/jpet.106.103143
3. Belozertseva IV, Kos T, Popik P, Danysz W, Bespalov AY. Antidepressant-like effects of mGluR1 and mGluR5 antagonists in the rat forced swim and the mouse tail suspension tests. Eur Neuropsychopharmacol. 2007;17(3):172–179. doi:10.1016/j.euroneuro.2006.03.002
4. Deschwanden A, Karolewicz B, Feyissa A, et al. Reduced metabotropic glutamate receptor 5 density in major depression determined by [C-11]ABP688 PET and postmortem study. Am J Psychiatry. 2011;168:727–734. doi:10.1176/appi.ajp.2011.09111607
5. Kovačević T, Skelin I, Minuzzi L, Rosa-Neto P, Diksic M. Reduced metabotropic glutamate receptor 5 in the Flinders Sensitive Line of rats, an animal model of depression: an autoradiographic study. Brain Res Bull. 2012;87(4–5):406–412. doi:10.1016/j.brainresbull.2012.01.010
6. Kim JH, Joo YH, Son YD, et al. In vivo metabotropic glutamate receptor 5 availability-associated functional connectivity alterations in drug-naïve young adults with major depression. Eur Neuropsychopharmacol. 2019;29(2):278–290. doi:10.1016/j.euroneuro.2018.12.001
7. DeLorenzo C, Sovago J, Gardus J, et al. Characterization of brain mGluR5 binding in a pilot study of late-life major depressive disorder using positron emission tomography and [11C]ABP688. Transl Psychiatry. 2015;5:e693. doi:10.1038/tp.2015.189
8. Abdallah CG, Hannestad J, Mason GF, et al. Metabotropic glutamate receptor 5 and glutamate involvement in major depressive disorder: a multimodal imaging study. Biol Psychiatry Cogn Neurosci Neuroimaging. 2017;2(5):449–456. doi:10.1016/j.bpsc.2017.03.019
9. Esterlis I, Holmes SE, Sharma P, Krystal JH, DeLorenzo C. mGluR5 and stress disorders: knowledge gained from receptor imaging studies. Biol Psychiatry. 2018;84(2):95–105. doi:10.1016/j.biopsych.2017.08.025
10. Ohayon MM, Schatzberg AF. Social phobia and depression: prevalence and comorbidity. J Psychosom Res. 2010;68(3):235–243. doi:10.1016/j.jpsychores.2009.07.018
11. Stein MB, Chavira DA. Subtypes of social phobia and comorbidity with depression and other anxiety disorders. J Affect Disord. 1998;50(Suppl 1):S11–16. doi:10.1016/s0165-0327(98)00092-5
12. Alpert JE, Uebelacker LA, McLean NE, et al. Social phobia, avoidant personality disorder and atypical depression: co-occurrence and clinical implications. Psychol Med. 1997;27(3):627–633. doi:10.1017/s0033291797004765
13. Liebowitz MR, Gorman JM, Fyer AJ, Klein DF. Social phobia. Review of a neglected anxiety disorder. Arch Gen Psychiatry. 1985;42(7):729–736. doi:10.1001/archpsyc.1985.01790300097013
14. Slattery DA, Neumann ID, Flor PJ, Zoicas I. Pharmacological modulation of metabotropic glutamate receptor subtype 5 and 7 impairs extinction of social fear in a time-point-dependent manner. Behav Brain Res. 2017;328:57–61. doi:10.1016/j.bbr.2017.04.010
15. Schulz B, Fendt M, Gasparini F, Lingenhöhl K, Kuhn R, Koch M. The metabotropic glutamate receptor antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP) blocks fear conditioning in rats. Neuropharmacology. 2001;41(1):1–7. doi:10.1016/s0028-3908(01)00036-3
16. Rodrigues SM, Bauer EP, Farb CR, Schafe GE, LeDoux JE. The group I metabotropic glutamate receptor mGluR5 is required for fear memory formation and long-term potentiation in the lateral amygdala. J Neurosci. 2002;22(12):5219–5229. doi:10.1523/JNEUROSCI.22-12-05219.2002
17. Xu J, Zhu Y, Contractor A, Heinemann SF. mGluR5 has a critical role in inhibitory learning. J Neurosci. 2009;29(12):3676–3684. doi:10.1523/JNEUROSCI.5716-08.2009
18. Jacob W, Gravius A, Pietraszek M, et al. The anxiolytic and analgesic properties of fenobam, a potent mGlu5 receptor antagonist, in relation to the impairment of learning. Neuropharmacology. 2009;57(2):97–108. doi:10.1016/j.neuropharm.2009.04.011
19. Burgos-Robles A, Vidal-Gonzalez I, Santini E, Quirk GJ. Consolidation of fear extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex. Neuron. 2007;53(6):871–880. doi:10.1016/j.neuron.2007.02.021
20. Ametamey SM, Treyer V, Streffer J, et al. Human PET studies of metabotropic glutamate receptor subtype 5 with 11C-ABP688. J Nucl Med. 2007;48(2):247–252.
21. Wang J, Tian Y, Zeng LH, Xu H. Prefrontal disinhibition in social fear: a vital action of somatostatin interneurons. Front Cell Neurosci. 2020;14:611732. doi:10.3389/fncel.2020.611732
22. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV.
23. First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV Axis I Disorders Research Version (SCID-I). New York: Biometrics Research. New York State Psychiatric Institute; 1996.
24. Hamilton M. A rating scale for depression. J Neurol Neurosurg Psychiatry. 1960;23:56–62. doi:10.1136/jnnp.23.1.56
25. Beck AT. Depression. Clinical, Experimental and Theoretical Aspects. New York (Hoeber). 1967.
26. Watson D, Friend R. Measurement of social-evaluative anxiety. J Consult Clin Psychol. 1969;33(4):448–457. doi:10.1037/h0027806
27. Lee JY, Choi CH. A study of the reliability and the validity of the Korean versions of Social Phobia Scales (K-SAD, K-FNE). Korean J Clin Psychol. 1997;16(2):251–264.
28. DeLorenzo C, Kumar JSD, Mann JJ, Parsey RV. In vivo variation in metabotropic glutamate receptor subtype 5 binding using positron emission tomography and [11C]ABP688. J Cereb Blood Flow Metab. 2011;31(11):2169–2180. doi:10.1038/jcbfm.2011.105
29. DeLorenzo C, Gallezot JD, Gardus J, et al. In vivo variation in same-day estimates of metabotropic glutamate receptor subtype 5 binding using [11C]ABP688 and [18F]FPEB. J Cereb Blood Flow Metab. 2017;37(8):2716–2727. doi:10.1177/0271678X16673646
30. Wu Y, Carson RE. Noise reduction in the simplified reference tissue model for neuroreceptor functional imaging. J Cereb Blood Flow Metab. 2002;22(12):1440–1452. doi:10.1097/01.WCB.0000033967.83623.34
31. DuBois JM, Rousset OG, Rowley J, et al. Characterization of age/sex and the regional distribution of mGluR5 availability in the healthy human brain measured by high-resolution [(11)C]ABP688 PET. Eur J Nucl Med Mol Imaging. 2016;43(1):152–162. doi:10.1007/s00259-015-3167-6
32. Akkus F, Treyer V, Ametamey SM, Johayem A, Buck A, Hasler G. Metabotropic glutamate receptor 5 neuroimaging in schizophrenia. Schizophr Res. 2017;183:95–101. doi:10.1016/j.schres.2016.11.008
33. Akkus F, Mihov Y, Treyer V, et al. Metabotropic glutamate receptor 5 binding in male patients with alcohol use disorder. Transl Psychiatry. 2018;8(1):17. doi:10.1038/s41398-017-0066-6
34. Smart K, Cox SML, Nagano-Saito A, Rosa-Neto P, Leyton M, Benkelfat C. Test-retest variability of [11 C]ABP688 estimates of metabotropic glutamate receptor subtype 5 availability in humans. Synapse. 2018;72(9):e22041. doi:10.1002/syn.22041
35. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage. 2002;15(1):273–289. doi:10.1006/nimg.2001.0978
36. Bruder GE, Stewart JW, McGrath PJ. Right brain, left brain in depressive disorders: clinical and theoretical implications of behavioral, electrophysiological and neuroimaging findings. Neurosci Biobehav Rev. 2017;78:178–191. doi:10.1016/j.neubiorev.2017.04.021
37. de Aguiar Neto FS, Rosa JLG. Depression biomarkers using non-invasive EEG: a review. Neurosci Biobehav Rev. 2019;105:83–93. doi:10.1016/j.neubiorev.2019.07.021
38. Swanson CJ, Bures M, Johnson MP, Linden AM, Monn JA, Schoepp DD. Metabotropic glutamate receptors as novel targets for anxiety and stress disorders. Nat Rev Drug Discov. 2005;4(2):131–144. doi:10.1038/nrd1630
39. Terbeck S, Akkus F, Chesterman LP, Hasler G. The role of metabotropic glutamate receptor 5 in the pathogenesis of mood disorders and addiction: combining preclinical evidence with human Positron Emission Tomography (PET) studies. Front Neurosci. 2015;9:86. doi:10.3389/fnins.2015.00086
40. Gravius A, Pietraszek M, Schmidt WJ, Danysz W. Functional interaction of NMDA and group I metabotropic glutamate receptors in negatively reinforced learning in rats. Psychopharmacology. 2006;185(1):58–65. doi:10.1007/s00213-005-0249-3
41. Molina-Hernández M, Tellez-Alcántara NP, Pérez-García J, Olivera-Lopez JI, Jaramillo MT. Antidepressant-like and anxiolytic-like actions of the mGlu5 receptor antagonist MTEP, microinjected into lateral septal nuclei of male Wistar rats. Prog Neuropsychopharmacol Biol Psychiatry. 2006;30(6):1129–1135. doi:10.1016/j.pnpbp.2006.04.022
42. Chen L, Liu J, Ali U, et al. Chronic, systemic treatment with a metabotropic glutamate receptor 5 antagonist produces anxiolytic-like effects and reverses abnormal firing activity of projection neurons in the basolateral nucleus of the amygdala in rats with bilateral 6-OHDA lesions. Brain Res Bull. 2011;84(3):215–223. doi:10.1016/j.brainresbull.2011.01.005
43. Fontanez-Nuin DE, Santini E, Quirk GJ, Porter JT. Memory for fear extinction requires mGluR5-mediated activation of infralimbic neurons. Cereb Cortex. 2011;21(3):727–735. doi:10.1093/cercor/bhq147
44. Iijima M, Fukumoto K, Chaki S. Acute and sustained effects of a metabotropic glutamate 5 receptor antagonist in the novelty-suppressed feeding test. Behav Brain Res. 2012;235(2):287–292. doi:10.1016/j.bbr.2012.08.016
45. Chung W, Choi SY, Lee E, et al. Social deficits in IRSp53 mutant mice improved by NMDAR and mGluR5 suppression. Nat Neurosci. 2015;18(3):435–443. doi:10.1038/nn.3927
46. Wagner KV, Hartmann J, Labermaier C, et al. Homer1/mGluR5 activity moderates vulnerability to chronic social stress. Neuropsychopharmacology. 2015;40(5):1222–1233. doi:10.1038/npp.2014.308
47. Xu H, Liu L, Tian Y, et al. A disinhibitory microcircuit mediates conditioned social fear in the prefrontal cortex. Neuron. 2019;102(3):668–682.e5. doi:10.1016/j.neuron.2019.02.026
48. Bittar TP, Labonté B. Functional contribution of the medial prefrontal circuitry in major depressive disorder and stress-induced depressive-like behaviors. Front Behav Neurosci. 2021;15. doi:10.3389/fnbeh.2021.699592
49. Pinkham AE, Hopfinger JB, Pelphrey KA, Piven J, Penn DL. Neural bases for impaired social cognition in schizophrenia and autism spectrum disorders. Schizophr Res. 2008;99(1–3):164–175. doi:10.1016/j.schres.2007.10.024
50. Rassovsky Y, Dunn W, Wynn J, et al. The effect of transcranial direct current stimulation on social cognition in schizophrenia: a preliminary study. Schizophr Res. 2015;165(2–3):171–174. doi:10.1016/j.schres.2015.04.016
51. Shin JE, Choi SH, Lee H, Shin YS, Jang DP, Kim JJ. Involvement of the dorsolateral prefrontal cortex and superior temporal sulcus in impaired social perception in schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2015;58:81–88. doi:10.1016/j.pnpbp.2014.12.006
52. Greicius MD, Krasnow B, Reiss AL, Menon V. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proc Natl Acad Sci U S A. 2003;100(1):253–258. doi:10.1073/pnas.0135058100
53. Fitzgerald PB, Oxley TJ, Laird AR, Kulkarni J, Egan GF, Daskalakis ZJ. An analysis of functional neuroimaging studies of dorsolateral prefrontal cortical activity in depression. Psychiatry Res. 2006;148(1):33–45. doi:10.1016/j.pscychresns.2006.04.006
54. Hasler G, van der Veen JW, Tumonis T, Meyers N, Shen J, Drevets WC. Reduced prefrontal glutamate/glutamine and gamma-aminobutyric acid levels in major depression determined using proton magnetic resonance spectroscopy. Arch Gen Psychiatry. 2007;64(2):193–200. doi:10.1001/archpsyc.64.2.193
55. Johnstone T, van Reekum CM, Urry HL, Kalin NH, Davidson RJ. Failure to regulate: counterproductive recruitment of top-down prefrontal-subcortical circuitry in major depression. J Neurosci. 2007;27(33):8877–8884. doi:10.1523/JNEUROSCI.2063-07.2007
56. Siegle GJ, Thompson W, Carter CS, Steinhauer SR, Thase ME. Increased amygdala and decreased dorsolateral prefrontal BOLD responses in unipolar depression: related and independent features. Biol Psychiatry. 2007;61(2):198–209. doi:10.1016/j.biopsych.2006.05.048
57. Sheline YI, Barch DM, Price JL, et al. The default mode network and self-referential processes in depression. PNAS. 2009;106(6):1942–1947. doi:10.1073/pnas.0812686106
58. Sheline YI, Price JL, Yan Z, Mintun MA. Resting-state functional MRI in depression unmasks increased connectivity between networks via the dorsal nexus. Proc Natl Acad Sci U S A. 2010;107(24):11020–11025. doi:10.1073/pnas.1000446107
59. Gellner AK, Voelter J, Schmidt U, et al. Molecular and neurocircuitry mechanisms of social avoidance. Cell Mol Life Sci. 2021;78(4):1163–1189. doi:10.1007/s00018-020-03649-x
60. Davis MT, Hillmer A, Holmes SE, et al. In vivo evidence for dysregulation of mGluR5 as a biomarker of suicidal ideation. Proc Natl Acad Sci U S A. 2019;116(23):11490–11495. doi:10.1073/pnas.1818871116
61. DeLorenzo C, Milak MS, Brennan KG, Kumar JSD, Mann JJ, Parsey RV. In vivo positron emission tomography imaging with [11C]ABP688: binding variability and specificity for the metabotropic glutamate receptor subtype 5 in baboons. Eur J Nucl Med Mol Imaging. 2011;38(6):1083–1094. doi:10.1007/s00259-010-1723-7
62. Daggett LP, Sacaan AI, Akong M, et al. Molecular and functional characterization of recombinant human metabotropic glutamate receptor subtype 5. Neuropharmacology. 1995;34(8):871–886. doi:10.1016/0028-3908(95)00085-k
63. Berthele A, Platzer S, Laurie DJ, et al. Expression of metabotropic glutamate receptor subtype mRNA (mGluR1-8) in human cerebellum. Neuroreport. 1999;10(18):3861–3867. doi:10.1097/00001756-199912160-00026
64. Patel S, Hamill TG, Connolly B, Jagoda E, Li W, Gibson RE. Species differences in mGluR5 binding sites in mammalian central nervous system determined using in vitro binding with [18F]F-PEB. Nucl Med Biol. 2007;34(8):1009–1017. doi:10.1016/j.nucmedbio.2007.07.009
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.