Back to Journals » Journal of Pain Research » Volume 15
Differences and Similarities in Spontaneous Activity Between Animal Models of Cancer-Induced Pain and Neuropathic Pain
Authors Zhu YF ![]() , Kan P
, Kan P ![]() , Singh G
, Singh G ![]()
Received 29 July 2022
Accepted for publication 29 September 2022
Published 12 October 2022 Volume 2022:15 Pages 3179—3187
DOI https://doi.org/10.2147/JPR.S383373
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Yong Fang Zhu,1,2 Peter Kan,3 Gurmit Singh1,2
1Michael G. DeGroote Institute for Pain Research and Care, McMaster University, Hamilton, ON, Canada; 2Department of Pathology & Molecular Medicine, McMaster University, Hamilton, ON, Canada; 3Health Sciences, McMaster University, Hamilton, ON, Canada
Correspondence: Gurmit Singh, Email [email protected]
Background: Clinical data on cancer-induced pain (CIP) demonstrate widespread changes in sensory function. It is characterized in humans not only by stimulus-invoked pain, but also by spontaneous pain. In our previous studies in an animal model of CIP, we observed changes in intrinsic membrane properties and excitability of dorsal root ganglion (DRG) sensory neurons corresponding to mechanical allodynia and hyperalgesia, of which abnormal activities of Aβ-fiber sensory neurons are consistent in a rat model of peripheral neuropathic pain (NEP).
Objective: To investigate whether there are related peripheral neural mechanisms between the CIP and NEP models of spontaneous pain, we compared the electrophysiological properties of DRG sensory neurons at 2– 3 weeks after CIP and NEP model induction.
Methods: CIP models were induced with metastasis tumour-1 rat breast cancer cells implanted into the distal epiphysis of the femur. NEP models were induced with a polyethylene cuff implanted around the sciatic nerve. Spontaneous pain in animals is measured by spontaneous foot lifting (SFL). After measurement of SFL, the animals were prepared for electrophysiological recordings of spontaneous activity (SA) in DRG neurons in vivo.
Results: Our data showed that SFL and SA occurred in both models. The proportion of SFL and SA of C-fiber sensory neurons in CIP was more significantly increased than in NEP models. There was no difference in duration of SFL and the rate of SA between the two models. The duration of SFL is related to the rate of SA in C-fiber in both models.
Conclusion: Thus, SFL may result from SA activity in C-fiber neurons in CIP and NEP rats. The differences and similarities in spontaneous pain between CIP and NEP rats is related to the proportion and rate of SA in C-fibers, respectively.
Keywords: electrophysiology recording, spontaneous pain, sensory neuron, cancer-induced pain, neuropathic pain
Introduction
Cancer-induced pain (CIP), especially from primary cancer metastases to the bone, is a common and highly debilitating complication of many cancer patients.1 Overall, 75–90% of patients with metastatic or advanced cancer will experience severe pain that significantly affects their quality of life.1,2 CIP typically involves background pain, spontaneous pain, and incidence or movement-evoked pain.1–3 While background pain is relatively stable and adequately controlled, spontaneous and movement-evoked pain, often referred to as breakthrough pain, involves episodes of extreme pain that breach the underlying level of background pain.1,3 Therapeutically, the spontaneous and movement-evoked pains are more difficult to control, and the treatment is often accompanied by intolerable adverse effects.1,4
One of the main reasons for the poor pharmacological targeting of CIP is the complexity of the pain state, which may involve inflammatory, neuropathic, and cancer-specific pain mechanisms.1 The biological mechanisms of cancer-related pain are multifaceted. Depending on the type and anatomical location of the cancer, different pain processes may be responsible.5–8 In our previous investigation of an animal model of CIP, we observed changes in intrinsic membrane properties and excitability of dorsal root ganglion (DRG) C-fiber and Aβ-fiber neurons corresponding to mechanical allodynia and hyperalgesia, of which abnormal activities of Aβ-fiber sensory neurons are consistent in a rat model of peripheral neuropathic pain (NEP).9,10 This has prompted an investigation and comparison of the intracellular electrophysiological characteristics of DRG sensory neurons corresponding to spontaneous pain in rat models of NEP and CIP.
Clinical symptoms of spontaneous pain include pinching, shooting, burning, stabbing, and paroxysmal pain, sometimes with dysesthesia and/or paresthesia.11,12 In animal models, spontaneous foot lifting (SFL) behavior is considered a manifestation of spontaneous pain.12–14 In this study, we compared the spontaneous activity (SA) properties of DRG sensory neurons at 2–3 weeks after CIP model induction with properties in NEP animals. Our finding indicates that SA in C-fiber neurons play an important role in spontaneous pain. The differences and similarities in spontaneous pain between CIP and NEP rats is related to the proportion and rate of SA in C-fibers, respectively.
Methods
All experimental procedures were in accordance with the Guide for the Care and Use of Laboratory Animals, Vols. 1 and 2, of the Canadian Council on Animal Care. All protocols were reviewed and approved by the McMaster University Animal Research Ethics Board.
CIP Model Rats and Tumor Induction
Immunocompetent female Sprague-Dawley (SD) rats (Charles River Inc., St. Constant, QC) weighing 170–200 g were randomly assigned to the CIP surgery group (n = 18). CIP animals were induced as described in our previous study, following a method established by others.15,16 The rats were anesthetized with inhaled isoflurane (3–5% in O2) and injected with 3.0×104 MRMT-1 cells resuspended in 20 μL of HBSS into the femur.15,16
NEP Model Rats and Peripheral Neuropathic Surgery
Immunocompetent female Sprague-Dawley (SD) rats (Charles River Inc., St. Constant, QC) weighing 170–200 g were also randomly assigned to the NEP surgery group (n = 18). A peripheral neuropathy was induced according to the method previously described in detail.17,18 Under anesthesia, the right sciatic nerve was exposed in the mid-thigh. Two 0.5mm polyethylene (PE 90) tubing cuffs (Intramedic PE-90, Fisher Scientific Ltd., Whitby, Ontario, Canada) were inserted around the exposed nerve approximately 1 mm apart. The wound was then sutured.
Control Rats
In our previous study, CIP-control rats (sham injection rats) only received an injection of 20 μL of HBSS by the same procedures. NEP-control rats were induced using the same procedure except that no cuff was inserted around the sciatic nerve. Our previous studies showed that DRG neuronal membrane properties in CIP-sham and NRP-sham rats were similar to unoperated control rats.16,19 Therefore, in this study, we did not use a separate sham group.
Behavioral Tests
Our previous studies showed that both the NEP model and the CIP model were well developed with mechanical allodynia within 14–21 days.9,10,20,21 For comparing mechanistic allodynia and spontaneous neuronal activity in the same time frame, behavior tests were performed immediately before the anesthesia required for electrophysiological recordings (14–21 days).
We used spontaneous foot-lifting (SFL) as a marker of spontaneous pain. SFL was measured according to the method previously described in detail.12 Rat lifting of the ipsilateral hindpaw accompanied by aversive behaviors (eg, foot shaking/licking) was measured during the 30-minute observation. The contralateral hindfoot was also observed during the same period. Foot lifts associated with exploratory behavior, locomotion, body repositioning, and grooming were excluded. The duration of persistent paw elevation was recorded with a stopwatch. Rats with SFL of >1 sec were considered to have spontaneous pain. SFL duration was calculated as an average of the duration the rats lifted their ipsilateral hindfoot during the observation time.
One form of evoked pain behavior (mechanical allodynia) was measured by Von Frey paw withdrawal threshold test, as described in our previous study.13,16 Briefly, rats were placed in a clear plexiglass box which can allow full paw access. Rats were habituated to the box until cage exploration and primary grooming activities ceased. Von Frey filaments (Stoelting Co., Wood Dale, IL) were applied to the plantar surface of the ipsilateral hind paw to determine the mechanical retraction threshold using the up–down method.16,22
In vivo Intracellular DRG Recordings
Details of acute intracellular electrophysiological recording techniques in our study have been reported previously.9,10,18,21 Briefly, each rat was anesthetized and fixed in a stereotaxic frame with the vertebral column rigidly clamped at lumbar L2 and L6. L4 DRGs containing large number of hind leg afferent somatic cells were selected and exposed for somata intracellular recordings. The L4 dorsal root was sectioned for stimulation. The distance from the stimulation site to the recording site (centre of the DRG) was measured at the end of the experiment to determine conduction distance. This value was used to calculate the conduction velocity (CV) of the dorsal root axon associated with each neuron. DRG sensory neurons were classified according to their conduction velocity (C-fiber neurons [≤0.8 mm/ms], and Aβ-fiber neurons [>6.5 mm/ms]).10,16,18
Spontaneous Activity
Neurons are classified as spontaneous activity (SA), which refers to sustained firing that does not respond to known stimuli. SA was identified according to the method previously described in detail.12 C-cooling units, which usually fire at room temperature, and muscle spindle afferents, which have high stretch-evoked firing rates affected by muscle manipulation, were excluded in this study.
Statistical Analysis of Behavioral and Electrophysiological Data
Comparisons of the proportion of animals with and without SFL between CIP and NEP in Figure 1A were made with Fisher’s exact test in contingency table comparisons (2 × 2). Comparisons of SFL duration in Figure 1B were made between CIP and NEP with the Mann–Whitney U test. Linear regression analyses were performed on data in Figure 2. Multiple pairwise comparisons of proportions of neurons with and without SA between control, CIP and NEP in Figure 3 were also made with Fisher’s exact test. Comparisons of SA median firing rates in Figure 4 were made between CIP and NEP with the Mann–Whitney U test. Linear regression analyses were performed on data Figure 5. GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA) was used for statistical analyses and graphing.
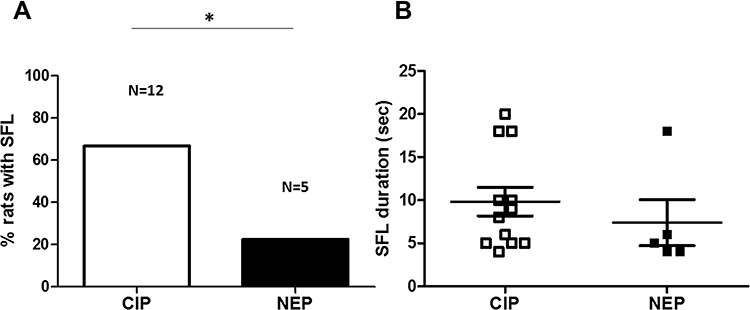 |
Figure 1 SFL in CIP (n = 18) and NEP (n = 18) rats. Numbers above histogram columns in (A) indicate number of rats with >1s SFL. (A) shows that the percentage of CIP rats with SFL is significantly greater than that of NEP rats (p = 0.043, Fisher’s exact test). (B) shows SFL duration in CIP and NEP rats (p = 0.003, Mann–Whitney test). *p <0.05. |
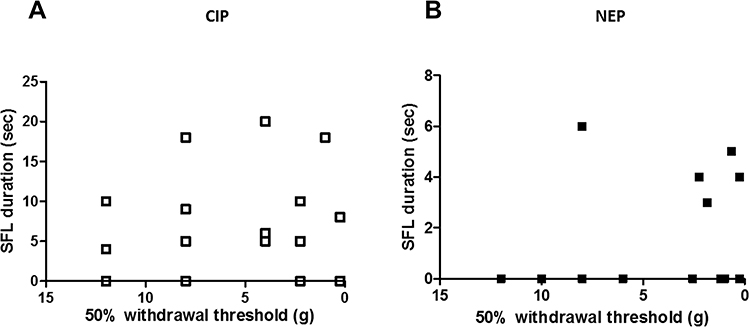 |
Figure 2 Relationship between spontaneous pain and mechanical allodynia from CIP rats (A) and NEP rats (B). The 50% withdrawal threshold was determined using von Frey filaments as a measure of mechanical allodynia. The direction of the axes shows more extreme pain-related behaviors toward the top and right of the graphs. Linear regression analysis was performed on CIP and NEP separately. Allodynia was not significantly correlated with SFL in CIP and in NEP rats. |
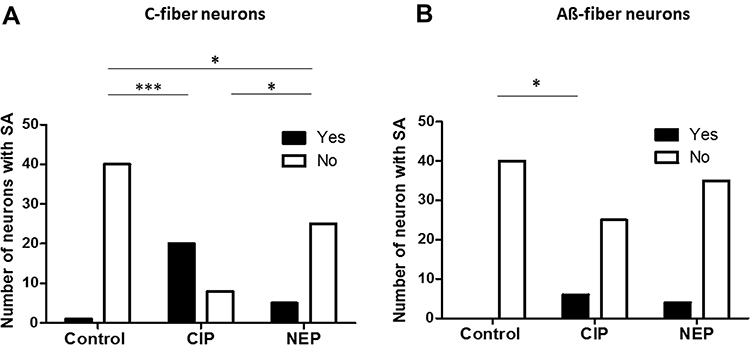 |
Figure 3 Number of (A) C-fiber neurons and (B) Aß-fiber neurons with SA in CIP and NEP rats. The columns show neurons from control, CIP, and NEP rats. The filled columns are number of neurons with SA and the open columns are number of neurons with no SA. The horizontal lines above the graphs link columns between which there is a significant difference using Fisher’s exact test for 2 × 2 contingency tables. Levels of significance are indicated as follows: *p <0.05; ***p <0.001. |
 |
Figure 4 The median rate of spontaneous firing in (A) C-fiber neurons and (B) Aß-fiber neurons in CIP and NEP rats. (C) and (D) show example records of two typical C-fiber (C) and Aß-fiber (D) neurons firing spontaneously in CIP rats. |
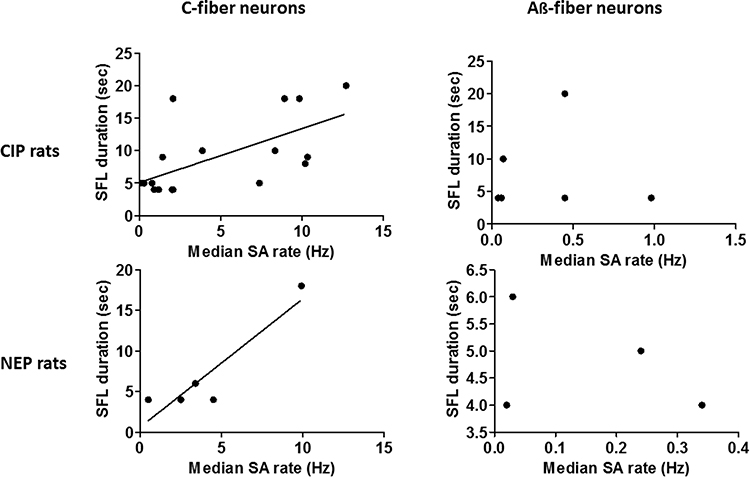 |
Figure 5 Plot of SFL duration for CIP and NEP rats (from data plotted in Figure 1B) against median firing rates (from data plotted in Figures 4A and B) of C-fiber and Aß-fiber nociceptor-type neurons with SA for that group. The lines indicate significant correlation. |
In this study, a sensory neuron firing rate was assessed based on a measure of the median SA rate (Hz) in one trial. We wrote the code in MATLAB (The MathWorks Inc., Natick, Massachusetts) to calculate the SA median rate described above.
Results
Behavior Study
Behavioral Signs of Spontaneous Pain
No SFL was observed in control rats (n = 18) and none of the rats exhibited SFL on the contralateral foot. SFL occurred in both CIP and NEP rats. A higher percentage of CIP rats showed SFL compared to NEP rats (66.67% [12 of 18] vs 27.78% [5 of 18]; p = 0.043, Fisher’s exact test) (Figure 1A). Figure 1B shows no difference in SFL duration between CIP and NEP rats (p = 0.219).
Relationships Between Mechanical Allodynia and SFL
To ensure that SFL is not an evoked response from mechanical allodynia, we performed linear regression analysis between the two behaviors (Figure 2). There was no significant correlation between SFL and mechanical allodynia in CIP rats (p = 0.436) (Figure 2A) and in NEP rats (p = 0.940) (Figure 2B).
Electrophysiological Study
Electrophysiological recordings were made in 98 C-fiber neurons (40 in control, 28 in CIP, 30 in NEP) and 110 Aβ-fiber neurons (40 in control, 31 in CIP, 39 in NEP), respectively.
Proportion of SA
SA in DRG sensory neurons of different groups are shown in Figure 3.
In C-fiber neurons, one of 40 neurons in control rats showed SA. However, 20 of 28 neurons (71.4%) in CIP rats and 5 of 30 neurons (16.67%) in NEP rats showed SA; both significantly increased in proportion compared to control rats (p < 0.0001 and p = 0.033, respectively) (Figure 3A). Notably, the proportion of C-fiber with SA was also significantly increased in CIP rats compared to NEP rats (p < 0.0001) (Figure 3A).
In Aβ-fiber neurons, none of the 40 in CIP rats showed SA. The proportion of Aβ-fiber neurons showing SA was slightly increased in NEP rats compared to control rats (4 of 39 neurons, 10.25%), but not significantly (p = 0.054). The proportion of Aβ-fiber neurons (6 out of 31 neurons, 19.35%) in CIP rats showed a significant increase in the percentage of SA compared to control rats (p = 0.038). There was no significant difference between CIP and NEP rats (p = 0.320) (Figure 3B).
Rate of SA
In C-fiber neurons, the median SA firing rate in CIP rats (4.163±4.34 Hz; range 0.18–12.71Hz) was not significantly (p = 0.563) different in NEP rats (4.164±3.54 Hz; range, 0.50–9.92 Hz) (Figure 4A).
In Aβ-fiber neurons, the median SA rate for CIP rats (0.342±0.150 Hz; range 0.04–0.98 Hz) was also not significantly different (p = 0.249) compared to NEP rats (0.158±0.16 Hz; range 0.02–0.34 Hz) (Figure 4B). Figures 4C and D show recordings of two typical C-fiber spontaneous firings at 10.2 Hz and Aß fiber firings at 0.21 Hz in CIP rats.
Link Between Behavior and Electrophysiological Study
Link Between Proportion of SFL and SA
As described above, in CIP rats we have SFL recordings from 12 out of 18 rats (66.67%), in which 20 out of 28 C-fiber neurons (71.4%) and 6 out of 31 Aβ-fiber neurons (19.35%) have SA. In NEP rats, we also recorded SFL recordings from 5 out of 18 rats (27.78), in which 5 out of 30 C-fiber neurons (16.67) and 4 out of 39 Aβ-fiber neurons (10.25%) have SA. The proportions of SFL and SA of C-fiber neurons in CIP rats were significantly higher than those in NEP rats. The relationship between proportion of SFL and SA of C-fiber neurons is positively correlated.
Link Between SFL Duration and SA Rate
In Figure 5, the median SA rate in Figures 4A and B is plotted against the SFL duration in Figure 1B. A highly significant linear correlation indicated that SFL duration was associated with the median SA rate of C-fiber neurons in CIP rats (p = 0.002; r2 = 0.43) and NEP rats (p = 0.028; r2 = 0.84). However, there was no relationship between SFL duration and the median SA of Aβ-fiber neurons in CIP rats (p = 0.98; r2 = 7.484e-005) and NEP rats (p = 0.62; r2 = 0.14).
Discussion
This is the first report comparing spontaneous pain (indicated by SFL) and the characteristics of spontaneous firing (indicated by SA) in sensory neurons in cancer-induced pain and neuropathic pain models.
Functional Significance of Spontaneous Activity in CIP and NEP Models
Our findings on SFL activity and SA from L4 C-fiber neurons and Aß-fiber neurons in CIP and NEP models has been reported in models of cutaneous and joint inflammation, as well as peripheral nerve injury.12,23–37 In particular, animal models of partial and complete nerve injury, chronic peripheral nerve constriction, dorsal root ganglion compression, nerve inflammation, diabetes, neurotoxin exposure such as chemotherapy, viral infections, and spinal cord injury increased primary afferent spontaneous activity.38 Electrophysiological recordings on dissociated primary neurons, intact dorsal root ganglions, and peripheral projections have all been used to demonstrate the associations between primary afferent SA and neuropathic pain.38 The different values in our NEP model study (female SD rats; SFL: 26.32%; C-fiber SA: 16,0.67%; Day 14–21) from those of Wu et al (female SD rats; ligation and transection of the L5 spinal nerve; SA in L4 C-fiber >50%) and Djouhri et al (female Wistar rats: spinal nerve L5 axotomy model; SFL: 17%; SA in L4 C-fiber 35%; Day 7) may relate to differing strain, induced model, recording time period, and DRG site.
In our study, we used the same strain, recording period (2–3 weeks), and recording site (L4 DRG) between NEP and CIP models. The differences and similarities between these two models are based on the following observations: SFL occurred in both NEP and CIP models; SA was observed in C and Aβ-fiber sensory neurons, in both NEP and CIP models; the proportion of SFL was more significantly increased in the CIP model (66.67%) than in the NEP model (26.32%). The proportion of C-fiber neurons showing SA in CIP model (71.4%) was also more significantly increased than in NEP model (16.67%). The SFL duration and the firing rate of SA showed the same range in both NEP and CIP models.
Comparing CIP and NEP models, the relationship between the proportion of SFL and SA in C-fiber neurons is positive. The linear correlation between SA firing rate of C-fiber neurons and SFL duration in both CIP and NEP models is consistent with a previous study using neuropathic and inflammatory models.12
Possible Mechanism of SA in C-Fiber Sensory Neurons
Causes of SA may include changes in afferent properties and/or environment of the fibers. SA in neuropathic models is considered to be caused by damage to the axon, soma, or neural sheath.38 This damage can arise from a variety of pathologies: trauma, compression or entrapment, inflammation (sterile, infectious, or autoimmune), direct infection, metabolic derangements, nutritional deficiencies, radiation, neurotoxin exposure, vascular insult, or genetic mutations.38,39 Although the etiology of cancer-induced pain remains unclear, animal models of cancer-induced pain have unraveled some of the cancer-induced neuropathologic processes.9 Previous studies revealed that cancer and immune cells produce and secrete mediators into the cancer microenvironment. Such mediators include protons40,41, ET-1,42–45 TNFα,46,47 NGF29–31,35,38, trypsin,41 opioids41 etc., which can modulate nociception.41
Previous studies also proposed that spontaneous pain may result from cumulative neuroinflammatory influences.12 Djouhri et al found that CFA treatment rats (intradermal complete Freund’s adjuvant-induced hind limb inflammation) showed a high rate of SA in C-fiber sensory neurons (35%).12 In another neuropathic pain model, application of the proinflammatory cytokine TNFα (tumor necrosis factor α) to the DRG resulted in increased dorsal root SA in C-fiber sensory neurons.48
These studies suggest that inflammation causes relatively rapid spontaneous firing of C-fiber sensory neurons. Thus, in our CIP model, the proportion of SA in C-fiber neurons is significantly higher than that in our NEP model, which may be due to the increased and ongoing inflammation influences provided by cancer-induced peripheral inflammation.
SA in C-Fiber Neurons May Act as a Peripheral Drive for SFL
Our previous study showed that there is no significant difference between CIP and NEP rats in mechanical allodynia.16,20,21 The linear regression analysis in this study indicates the lack of correlation between SFL and evoked pain behaviors (mechanical allodynia) in both CIP and NEP rats. Notably, our previous data showed that Aß-fiber but not C-fiber sensory neurons contribute to mechanical allodynia in NEP mode.10,18 in this study, we found that C-fiber neurons mainly play a role in SFL/spontaneous pain. All this evidence suggests that SFL in these rats may not be attributable to allodynia.
The primary afferent neuron is often cited as a site for the origin of SA in neuropathic pain.28,49–53 A previous study showed that activation of C- and Aß-fiber sensory neurons produces unpleasant/unbearable burning and sharp pain, respectively. Thus, SA in these two sensory groups may be responsible for the burning and stabbing spontaneous pain reported in human patients, respectively.12,54 In this study, the correlation between C-fiber SA proportion /SFL proportion and C-fiber SA firing rate/SFL duration in CIP and NEP models suggests that C-fiber SA occurrence and firing rate is an important drive for SFL. Therefore, SA in C-fiber neurons may cause the burning spontaneous pain which describes SFL in this study.
Conclusion
In summary, we present the differences and similarities of spontaneous pain between CIP and NEP models. We suggest there is an underlying neuronal mechanism linking inflammation, proportion and rate of spontaneous firing in C-fiber neurons and spontaneous pain. Elimination of SA in these C-fiber sensory neurons may therefore relieve cancer-induced and neuropathic spontaneous pain. Additional studies are needed to elucidate the altered electrophysiological properties and/or chemical influences causing this C-fiber SA in CIP and NEP models.
Acknowledgments
This study was supported by Michael G. DeGroote Institute for Pain Research and Care Seed Grant.
Author Contributions
All authors made a significant contribution to the work reported. Yong Fang Zhu designed the experiment, induced models and conducted the experiments. Peter Kan wrote MATLAB code to analyze the rate of spontaneous spiking. Gurmit Singh, the PI, supervised the overall project. All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests.
References
1. Falk S, Bannister K, Dickenson AH. Cancer pain physiology. Br J Pain. 2014;8(4):154–162. doi:10.1177/2049463714545136
2. Svendsen KB, Andersen S, Arnason S, et al. Breakthrough pain in malignant and non-malignant diseases: a review of prevalence, characteristics and mechanisms. Eur J Pain. 2005;9(2):195–206. doi:10.1016/j.ejpain.2004.06.001
3. Zeppetella G. Impact and management of breakthrough pain in cancer. Curr Opin Support Palliat Care. 2009;3(1):1–6. doi:10.1097/SPC.0b013e3283260658
4. Deandrea S, Montanari M, Moja L, Apolone G. Prevalence of undertreatment in cancer pain. A review of published literature. Ann Oncol. 2008;19(12):1985–1991. doi:10.1093/annonc/mdn419
5. Viet CT, Schmidt BL. Biologic mechanisms of oral cancer pain and implications for clinical therapy. J Dent Res. 2012;91(5):447–453. doi:10.1177/0022034511424156
6. Currie GL, Delaney A, Bennett MI, et al. Animal models of bone cancer pain: systematic review and meta-analyses. Pain. 2013;154(6):917–926. doi:10.1016/j.pain.2013.02.033
7. Epstein JB, Miaskowski C. Oral pain in the cancer patient. J Natl Cancer Inst Monogr. 2019;2019(53):lgz003. doi:10.1093/jncimonographs/lgz003
8. Schmidt BL. The neurobiology of cancer pain. J Oral Maxillofac Surg. 2015;73(12 Suppl):S132–135. doi:10.1016/j.joms.2015.04.045
9. Zhu YF, Ungard R, Zacal N, Huizinga JD, Henry JL, Singh G. Rat model of cancer-induced bone pain: changes in nonnociceptive sensory neurons in vivo. Pain Rep. 2017;2(4):e603. doi:10.1097/PR9.0000000000000603
10. Zhu YF, Wu Q, Henry JL. Changes in functional properties of A-type but not C-type sensory neurons in vivo in a rat model of peripheral neuropathy. J Pain Res. 2012;5:175–192. doi:10.2147/JPR.S26367
11. Attal N, Bouhassira D. Neuropathic pain: experimental advances and clinical applications. Rev Neurol. 2004;160(2):199–203. doi:10.1016/S0035-3787(04)70891-7
12. Djouhri L, Koutsikou S, Fang X, McMullan S, Lawson SN. Spontaneous Pain, Both Neuropathic and Inflammatory, Is Related to Frequency of Spontaneous Firing in Intact C-Fiber Nociceptors. J Neurosci. 2006;26(4):1281–1292. doi:10.1523/JNEUROSCI.3388-05.2006
13. Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33(1):87–107. doi:10.1016/0304-3959(88)90209-6
14. Yoon C, Wook YY, Sik NH, Ho KS, Mo CJ. Behavioral signs of ongoing pain and cold allodynia in a rat model of neuropathic pain. Pain. 1994;59(3):369–376. doi:10.1016/0304-3959(94)90023-X
15. Carlton SM, Zhou S, Coggeshall RE. Evidence for the interaction of glutamate and NK1 receptors in the periphery. Brain Res. 1998;790(1–2):160–169. doi:10.1016/S0006-8993(97)01471-6
16. Zhu YF, Linher-Melville K, Wu J, et al. Bone cancer-induced pain is associated with glutamate signalling in peripheral sensory neurons. Mol Pain. 2020;1:16.
17. Mosconi T, Kruger L. Fixed-diameter polyethylene cuffs applied to the rat sciatic nerve induce a painful neuropathy: ultrastructural morphometric analysis of axonal alterations. Pain. 1996;64(1):37–57. doi:10.1016/0304-3959(95)00077-1
18. Zhu YF, Henry JL. Excitability of Aβ sensory neurons is altered in an animal model of peripheral neuropathy. BMC Neurosci. 2012;13:15. doi:10.1186/1471-2202-13-15
19. Zhu YF, Linher‐Melville K, Niazmand MJ, et al. An evaluation of the anti-hyperalgesic effects of cannabidiolic acid-methyl ester in a preclinical model of peripheral neuropathic pain. Br J Pharmacol. 2020;177(12):2712–2725. doi:10.1111/bph.14997
20. Zhu YF, Kwiecien JM, Dabrowski W, et al. Cancer pain and neuropathic pain are associated with A β sensory neuronal plasticity in dorsal root ganglia and abnormal sprouting in lumbar spinal cord. Mol Pain. 2018;14:1744806918810099. doi:10.1177/1744806918810099
21. Zhu YF, Ungard R, Seidlitz E, et al. Differences in electrophysiological properties of functionally identified nociceptive sensory neurons in an animal model of cancer-induced bone pain. Mol Pain. 2016;12:54.
22. Dixon WJ. Efficient analysis of experimental observations. Annu Rev Pharmacol Toxicol. 1980;20:441–462. doi:10.1146/annurev.pa.20.040180.002301
23. Schaible HG, Schmidt RF. Discharge characteristics of receptors with fine afferents from normal and inflamed joints: influence of analgesics and prostaglandins. Agents Actions Suppl. 1986;19:99–117.
24. Kocher L, Anton F, Reeh PW, Handwerker HO. The effect of carrageenan-induced inflammation on the sensitivity of unmyelinated skin nociceptors in the rat. Pain. 1987;29(3):363–373. doi:10.1016/0304-3959(87)90051-0
25. Xiao WH, Bennett GJ. Persistent low-frequency spontaneous discharge in A-fiber and C-fiber primary afferent neurons during an inflammatory pain condition. Anesthesiology. 2007;107(5):813–821. doi:10.1097/01.anes.0000286983.33184.9c
26. Wu G, Ringkamp M, Hartke TV, et al. Early Onset of Spontaneous Activity in Uninjured C-Fiber Nociceptors after Injury to Neighboring Nerve Fibers. J Neurosci. 2001;21(8):RC140–RC140. doi:10.1523/JNEUROSCI.21-08-j0002.2001
27. Ma C, LaMotte RH. Multiple sites for generation of ectopic spontaneous activity in neurons of the chronically compressed dorsal root ganglion. J Neurosci. 2007;27(51):14059–14068. doi:10.1523/JNEUROSCI.3699-07.2007
28. Xie W, Strong JA, Meij JTA, Zhang JM, Yu L. Neuropathic pain: early spontaneous afferent activity is the trigger. Pain. 2005;116(3):243–256. doi:10.1016/j.pain.2005.04.017
29. Burchiel KJ, Russell LC, Lee RP, Sima AA. Spontaneous activity of primary afferent neurons in diabetic BB/Wistar rats. A possible mechanism of chronic diabetic neuropathic pain. Diabetes. 1985;34(11):1210–1213. doi:10.2337/diab.34.11.1210
30. Mayer ML, James MH, Russell RJ, Kelly JS, Wise JC, Pasternak CA. Spontaneous electrical activity induced by herpes virus infection in rat sensory neuron cultures. Brain Res. 1985;341(2):360–364. doi:10.1016/0006-8993(85)91075-3
31. Study RE, Kral MG. Spontaneous action potential activity in isolated dorsal root ganglion neurons from rats with a painful neuropathy. Pain. 1996;65(2–3):235–242. doi:10.1016/0304-3959(95)00216-2
32. Liu CN, Wall PD, Ben-Dor E, Michaelis M, Amir R, Devor M. Tactile allodynia in the absence of C-fiber activation: altered firing properties of DRG neurons following spinal nerve injury. Pain. 2000;85(3):503–521. doi:10.1016/S0304-3959(00)00251-7
33. Kajander KC, Wakisaka S, Bennett GJ. Spontaneous discharge originates in the dorsal root ganglion at the onset of a painful peripheral neuropathy in the rat. Neurosci Lett. 1992;138(2):225–228. doi:10.1016/0304-3940(92)90920-3
34. Djouhri L, Fang X, Koutsikou S, Lawson SN. Partial nerve injury induces electrophysiological changes in conducting (uninjured) nociceptive and nonnociceptive DRG neurons: possible relationships to aspects of peripheral neuropathic pain and paresthesias. Pain. 2012;153(9):1824–1836. doi:10.1016/j.pain.2012.04.019
35. Liu X, Eschenfelder S, Blenk KH, Jänig W, Häbler H. Spontaneous activity of axotomized afferent neurons after L5 spinal nerve injury in rats. Pain. 2000;84(2–3):309–318. doi:10.1016/S0304-3959(99)00211-0
36. Ma C, LaMotte RH. Enhanced excitability of dissociated primary sensory neurons after chronic compression of the dorsal root ganglion in the rat. Pain. 2005;113(1–2):106–112. doi:10.1016/j.pain.2004.10.001
37. Zhang H, Dougherty PM. Enhanced excitability of primary sensory neurons and altered gene expression of neuronal ion channels in dorsal root ganglion in paclitaxel-induced peripheral neuropathy. Anesthesiology. 2014;120(6):1463–1475. doi:10.1097/ALN.0000000000000176
38. North RY, Lazaro TT, Dougherty PM. Ectopic Spontaneous Afferent Activity and Neuropathic Pain. Neurosurgery. 2018;65(CN_suppl_1):49–54. doi:10.1093/neuros/nyy119
39. Devor M. Ectopic discharge in Abeta afferents as a source of neuropathic pain. Exp Brain Res. 2009;196(1):115–128. doi:10.1007/s00221-009-1724-6
40. Lozano-Ondoua AN, Symons-Liguori AM, Vanderah TW. Cancer-induced bone pain: mechanisms and models. Neurosci Lett. 2013;557(Pt A):52–59. doi:10.1016/j.neulet.2013.08.003
41. Schmidt BL, Hamamoto DT, Simone DA, Wilcox GL. Mechanism of Cancer Pain. Mol Interv. 2010;10(3):164–178. doi:10.1124/mi.10.3.7
42. Gokin AP, Fareed MU, Pan HL, Hans G, Strichartz GR, Davar G. Local injection of endothelin-1 produces pain-like behavior and excitation of nociceptors in rats. J Neurosci. 2001;21(14):5358–5366. doi:10.1523/JNEUROSCI.21-14-05358.2001
43. Hans G, Deseure K, Robert D, De Hert S. Neurosensory changes in a human model of endothelin-1 induced pain: a behavioral study. Neurosci Lett. 2007;418(2):117–121. doi:10.1016/j.neulet.2007.03.008
44. Khodorova A, Navarro B, Jouaville LS, et al. Endothelin-B receptor activation triggers an endogenous analgesic cascade at sites of peripheral injury. Nat Med. 2003;9(8):1055–1061. doi:10.1038/nm885
45. Davar G. Endothelin-1 and metastatic cancer pain. Pain Med. 2001;2(1):24–27. doi:10.1046/j.1526-4637.2001.002001024.x
46. Woolf CJ, Allchorne A, Safieh-Garabedian B, Poole S. Cytokines, nerve growth factor and inflammatory hyperalgesia: the contribution of tumour necrosis factor alpha. Br J Pharmacol. 1997;121(3):417–424. doi:10.1038/sj.bjp.0701148
47. Wacnik PW, Eikmeier LJ, Simone DA, Wilcox GL, Beitz AJ. Nociceptive characteristics of tumor necrosis factor-alpha in naive and tumor-bearing mice. Neuroscience. 2005;132(2):479–491. doi:10.1016/j.neuroscience.2004.12.035
48. Schäfers M, Lee DH, Brors D, Yaksh TL, Sorkin LS. Increased Sensitivity of Injured and Adjacent Uninjured Rat Primary Sensory Neurons to Exogenous Tumor Necrosis Factor-α after Spinal Nerve Ligation. J Neurosci. 2003;23(7):3028–3038. doi:10.1523/JNEUROSCI.23-07-03028.2003
49. Yang Q, Wu Z, Hadden JK, et al. Persistent pain after spinal cord injury is maintained by primary afferent activity. J Neurosci. 2014;34(32):10765–10769. doi:10.1523/JNEUROSCI.5316-13.2014
50. Baron R, Hans G, Dickenson AH. Peripheral input and its importance for central sensitization. Ann Neurol. 2013;74(5):630–636. doi:10.1002/ana.24017
51. Haroutounian S, Nikolajsen L, Bendtsen TF, et al. Primary afferent input critical for maintaining spontaneous pain in peripheral neuropathy. Pain. 2014;155(7):1272–1279. doi:10.1016/j.pain.2014.03.022
52. Pitcher GM, Henry JL. Governing role of primary afferent drive in increased excitation of spinal nociceptive neurons in a model of sciatic neuropathy. Exp Neurol. 2008;214(2):219–228. doi:10.1016/j.expneurol.2008.08.003
53. Nordin M, Nyström B, Wallin U, Hagbarth KE. Ectopic sensory discharges and paresthesiae in patients with disorders of peripheral nerves, dorsal roots and dorsal columns. Pain. 1984;20(3):231–245. doi:10.1016/0304-3959(84)90013-7
54. Djouhri L, Lawson SN. Abeta-fiber nociceptive primary afferent neurons: a review of incidence and properties in relation to other afferent A-fiber neurons in mammals. Brain Res Brain Res Rev. 2004;46(2):131–145. doi:10.1016/j.brainresrev.2004.07.015
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.