Back to Journals » Breast Cancer: Targets and Therapy » Volume 17
Dexmedetomidine Suppresses Mitochondrial Autophagy and Apoptosis While Promoting Proliferation in Breast Cancer Cells in vitro via PI3K/AKT Signaling
Authors Gu M, Xia Y, Qian J, Shao C, Li Y, Lu X, Qin X
Received 27 May 2025
Accepted for publication 21 November 2025
Published 20 December 2025 Volume 2025:17 Pages 1265—1278
DOI https://doi.org/10.2147/BCTT.S543090
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Pranela Rameshwar
Mengting Gu,* Yanfei Xia,* Jiang Qian, Caiqun Shao, Yujia Li, Xing Lu, Xiarong Qin
Anesthesiology Department, Zhejiang Hospital, Hangzhou, Zhejiang, 310013, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiarong Qin, Anesthesiology Department, Zhejiang Hospital, No. 12 Lingyin Road, Xihu District, Hangzhou, Zhejiang, 310013, People’s Republic of China, Tel +571-87987373, Email [email protected]
Research Purpose: To investigate how dexmedetomidine (DEX) controls the proliferation and death of breast cancer cells.
Methods: Human breast cancer cells were cultured in vitro with DEX at different concentrations (25, 50, 100 ng/mL) or 30 μM LY294002. Cancer cell viability, proliferation, apoptosis and the expression of Microtubule-associated protein light chain 3 (LC3)-II/LC3-I protein were separately analyzed using cell counting kit 8 (CCK-8), colony formation, flow cytometry and Western blot assays after DEX treatment. The effect of DEX on mitochondrial membrane potential (MMP) level in cancer cells was determined using immunofluorescence. The expressions of B cell lymphoma-2 (Bcl-2), Bcl-2 associated X (Bax), phosphatidylinositol 3-kinase (PI3K), phosphorylated (p)-PI3K, protein kinase B (AKT) and p-AKT in DEX-treated cancer cells were measured by Western blot.
Results: DEX promoted cell growth activity and proliferation, inhibited cell autophagy and apoptosis and down-regulated the ratio of LC3-II/LC3-I to reverse the effect of LY294002 on breast cancer cells. DEX also abrogated LY294002-induced down-regulation of MMP, p-PI3K/PI3K, p-AKT/AKT and Bcl-2 and up-regulation of Bax in breast cancer cells.
Conclusion: DEX may promote the development of breast cancer cells while preventing cancer cell autophagy and apoptosis in vitro via PI3K/AKT signaling.
Keywords: dexmedetomidine, breast cancer, mitochondrial autophagy, apoptosis, PI3K/AKT signaling
Introduction
Breast cancer is one of the most common cancers in women, with local recurrence and a tendency to metastasize as primary contributors for its high mortality.1 According to the Global Cancer Statistics Report, an estimated 2.3 million new breast cancer cases were diagnosed worldwide in 2022, accounting for 11.6% of all malignancies in women.2 Breast cancer imposes substantial physical, emotional and socio-economic burdens on patients, their families, and society.3 Nowadays, the treatment methods include surgery, hormone therapy, immunotherapy, chemotherapy and radiotherapy, but there are still major challenges such as high mortality, poor prognosis after metastasis and chemotherapy resistance.4 Triple-negative breast cancer (TNBC), the most malignant subtype, is a highly heterogeneous cancer characterized by specific mutations and abnormal activation of signaling pathways, and has a poor prognosis. Current treatment options are limited to surgery, adjuvant chemotherapy, and radiotherapy; however, targeted therapy is a promising avenue for TNBC.1
Dexmedetomidine (DEX) is a kind of α2-adrenergic receptor agonist, which has strong specificity, good sedative and analgesic effects, and high safety.5 DEX is commonly used as a sedative and perioperative anesthetic adjuvant, particularly in patients undergoing surgery for breast cancer.6 Apoptosis, proliferation, and autophagy are involved in local anesthetic-induced cytotoxicity of human breast cancer cells.7 However, emerging preclinical evidence has revealed a concerning paradox: despite its beneficial anesthetic profile, DEX may exhibit tumor-promoting effects. A previous study showed that DEX can increase the proliferation, migration or invasion of breast cancer cells.8,9 These findings raise critical questions regarding the drug’s potential impact on long-term oncological outcomes and pose a pressing dilemma for the anesthesiologist: how to balance optimal perioperative care with potential oncological safety. This highlights an urgent need to investigate DEX’s potential impact on cancer progression and the underlying mechanisms.
One key factor for the development of drug resistance is cellular plasticity. The plasticity of cancer cells arises from the interaction between genetic factors and programs, leading to tumor progression and metastasis to distant organs, and conferring therapy resistance in breast cancer cells.10 The phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) pathway is one of the important intracellular signaling pathways that regulate basic cellular functions such as cell proliferation, metabolism, autophagy, motility, and apoptosis.11 PI3K/Akt pathway plays important roles in breast tumorigenesis and is related to worse prognosis in patients.12 Moreover, PI3K/AKT pathway promotes tumor growth by suppressing apoptosis and modulating autophagy.13 The major intrinsic cellular signaling pathways such as MAPK, PI3K/AKT/mammalian target of rapamycin (mTOR) have been found to be dysregulated in cancer and have the potential to induce cancer cell plasticity.10 Therefore, plasticity may modulate the feedback loop in our study, enabling cells to evade pathway inhibition via alternative transcriptional programs.
Further, it has been discovered that mitochondrial autophagy is connected to the survival of breast cancer cells.14 As the main energy supplier of cells, mitochondria mediate basic biological functions such as cell survival or energy metabolism.15 According to recent research, mitochondrial autophagy contributes to the development of breast cancer in two ways. On one hand, proper mitochondrial autophagy helps clear damaged or excess mitochondria, maintain the homeostasis and function of tumor cells, and is conducive to their survival and adaptation to the invasion environment. On the other hand, excessive mitochondrial autophagy can destroy normal mitochondria, while insufficient autophagy can lead to accumulation of damaged mitochondria, triggering cell death or apoptosis.16,17 Maintaining this balance is crucial for the growth or apoptosis of breast cancer cells.
Reportedly, DEX can alleviate mitochondrial dysfunction in diabetic peripheral neuropathy.18 Furthermore, evidence suggests that DEX’s effects in other contexts (cerebral ischemia-reperfusion injury) are mediated through the activation of the PI3K/AKT pathway.19 Given that (1) DEX promotes breast cancer cell behaviors, (2) DEX activates PI3K/AKT signaling, and (3) the PI3K/AKT pathway regulates mitophagy and apoptosis, we hypothesized a novel mechanistic link: DEX promotes breast cancer cell survival and growth by activating the PI3K/AKT pathway, which subsequently inhibits mitophagy and apoptosis.
However, few studies have elucidated the role of DEX in breast cancer. Whether DEX affects the occurrence and development of breast cancer through mitochondrial regulation, and what mechanisms underlie the development of cancerous breast cells, remain important questions. Given its widespread use as an anesthetic adjuvant in breast cancer surgery, we sought to determine how DEX affects mitochondrial autophagy and apoptosis in cancer cells. To this end, we carried out a series of experiments designed to elucidate the underlying mechanisms, with the ultimate goal of applying these findings to mitigate any adverse effects of DEX on patient prognosis.
Materials and Methods
Cell Experimental Grouping
Human breast cancer cells (MDA-MB-231) were obtained from the Wuhan Pricella Biotechnology Co., Ltd. Cells were identified by short tandem repeat (STR). 10% fetal bovine serum (A5256701, Thermofisher, USA) and RPMI 1640 medium (11875093, Thermofisher, USA) were used to cultivate the cells at 37°C under 5% CO2. The culture medium was changed 24 h later, and refreshed every two days.
The experiment was divided into two parts. In the first part, the cells were divided into DEX intervention and control groups. In the DEX intervention groups, 25, 50, or 100 ng/mL of DEX was added during cell culture for 24, 48, and 72 h, respectively. After drug treatment, follow-up cell tests such as cell counting kit 8 (CCK8), colony formation, Western blot, flow cytometry and immunofluorescence were performed.
In the second part, cells were assigned into four groups: control group, LY294002 group (cells were treated with 30 μM LY294002 for 48 h),20 DEX group (cells were treated with 100 ng/mL DEX for 48 h), and LY294002+DEX group (cells were treated with 30 μM LY294002 and 100 ng/mL DEX for 48 h). After drug treatment, CCK8, colony formation, Western blot, flow cytometry and immunofluorescence assays were carried out.
CCK8 Assay
Using the CCK8 kit (HY-K0301, MCE, USA), cell activity in each group was tested. Cells (5×103 cells/well) were inoculated in 96-well plates, and received treatment according to the grouping. The cells were cultivated for 24, 48, and 72h at 37°C with 5% CO2, and later cultured with 10 μL of CCK8 solution (HY-K0301, MCE, USA) in the incubator (Forma Steri-Cult, Thermofisher, USA) at 37°C for 2 h. Using an enzyme-labeled device (Varioskan LUX, Thermofisher, USA), the absorbance at 450 nm was monitored, and the cell viability was examined.
Colony Formation Experiment
The suspended cells were precipitated, counted, and diluted to 1×103 cells/mL. 6-well plates were used with a gradient density of 200 cells/well, and cell culture was conducted in the incubator. After cell adhesion the next day, drug treatment was performed according to the experimental group. 2–3 weeks later, when clones were visible in the petri dish, the culture medium was discarded, cells were carefully washed with PBS (ST477, Beyotime, China) 2–3 times. Next, cells were fixed with 1 mL methanol (C06901102, Nanjing Reagent, China) for 15 min, and then stained with 1 mL Giemsa dyeing solution (C0133, Beyotime, China) for 10–30 min. The dyeing solution was slowly washed away with running water. Ultimately, clones with more than 50 cells were counted using a microscope (THUNDER Imager Tissue, Leica, Germany) and photographed.
Western Blot Test
To accurately evaluate the expression levels of key proteins in the ACE2/Ang(1–7)/Mas axis and autophagy-related proteins (LC3, p62), Western blotting assay was carried out. Western blotting is currently the “gold standard” method for quantitative analysis of specific protein expressions in the field of life sciences, which has been widely used to verify the targets of bioactive molecules and downstream signaling pathways.21,22 Total protein was extracted from cells using RI-PA lysate (R0010, Solarbio, China). After quantification, electrophoresis separation, membrane transfer and blockage, primary antibodies were added for incubation at 4°C overnight. The film was washed three times with Tris-HCI buffered salt solution (including Tween) (T9039, Sigma-Aldrich, Germany). Secondary antibodies (ab6721, Abcam, UK, 1:2000) were later added for 2-h culture at room temperature. Gel imaging equipment was used to expose these bands, and Image J software was applied to calculate grayscale values. The primary antibodies used in Western blot were shown in Table 1.
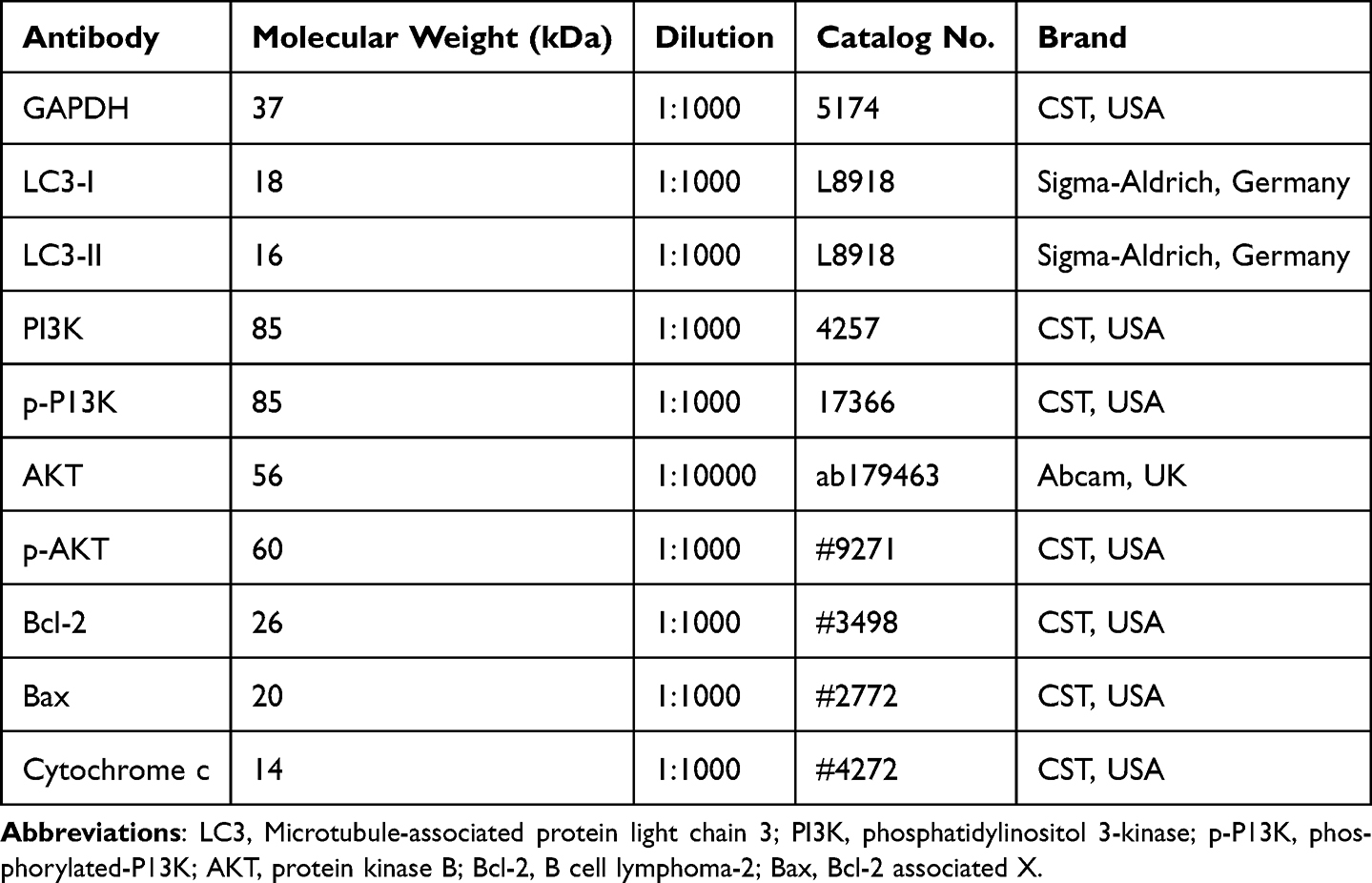 |
Table 1 The Primary Antibodies Used in Western Blot |
Flow Cytometry
The digested cells were centrifuged with EDTA-free pancreatic enzyme (HY-Y0682, MedChemExpress, USA) for 5 min at 1000 rpm, and then washed twice with pre-cooled PBS (ST477, Beyotime, China). The Annexin V-FITC/PI Apoptosis detection Kit (E-CK-A211, Elabscience, China) was utilized for the detection of cell apoptosis. Using a flow cytometer (CytoFLEX, Beckmancoulter, USA), apoptosis was observed. Three wells were provided for each sample, and the experiment was carried out three times or more.
Mitochondrial Membrane Potential (MMP) Measurement
A total of 6×105 cells were re-suspended in 1 mL cell culture solution. The measurement was carried out in accordance with the MMP detection kit’s instructions (JC-1) (C2006, Beyotime, China). Cells were incubated with 1 mL JC-1 dyeing solution, inverted and mixed, followed by 20-min incubation at 37°C and 3-min centrifugation at 4°C for 2000 rpm. After two washes with 1 mL of JC-1 staining buffer (1×), the cells were centrifuged at 4°C for 3 min and the supernatant was discarded. This wash cycle was repeated once. 2 mL of cell culture medium was added, and the fluorescence microscope was utilized for observation and photography. JC-1 polymer emitted red fluorescence, which represents the increase of MMP. JC-1 monomer emitted green fluorescence, indicating a low MMP.
Caspase 3/7 Activity Assay
Caspase 3/7 Activity Assay Kit (E-CK-A383, Elabscience, Wuhan, China) was used to detect Caspase 3/7 activity. After the cells were lysed and centrifuged, the supernatant was collected for protein quantification using Bradford method (P0006, Beyotime, Shanghai, China). Subsequently, 45 µL Reaction buffer, 50 µL samples, and 5 µL Ac-DEVD-pNA were sequentially mixed. The absorbance was recorded at 405 nm after incubation at 37°C for 1–2 h. The percentage of Caspase3/7 activity = [(OD sample − OD blank control) /Cpr sample] ÷ [(OD negative − OD blank control) /Cpr negative]×100%.
Statistical Analysis
GraphPad Prism 8.0 was used for statistical analysis. Mean ± standard deviation was used to express the measurement data. One- or two-way analysis of variance (ANOVA) and Tukey’s test were used for multi-group comparison. Normality was tested using the Shapiro–Wilk test, and homogeneity of variance was tested using the Levene test. The data met both normality and homogeneity of variance. The power analysis uses the effect size in the F-test. P<0.05 was regarded to be statistically significant.
 |
Figure 1 Effects of dexmedetomidine (DEX) at different concentrations (25, 50, 100 ng/mL) on proliferation, viability, autophagy and apoptosis of breast cancer cells. (A) CCK-8 was performed to detect the viability of breast cancer cells treated with different concentrations of DEX for 24, 48, 72 h. (B and C) Colony formation assay was used to detect the proliferation ability of cancer cells treated with DEX at different concentrations. (40×) (D and H) The expression of LC3-II/LC3-I and PINK1/Parkin protein in breast cancer cells treated with DEX at different concentrations was measured by Western blot. (I and J) The apoptosis of breast cancer cells treated with DEX at different concentrations was determined by flow cytometry. *P<0.05, **P<0.01, ***P<0.001 vs Control. n=3 biological replicates. |
 |
Figure 2 Effects of DEX at different concentrations (25, 50, 100 ng/mL) on mitochondrial membrane potential (MMP) level and expression levels of Bcl-2, Bax, p-PI3K, PI3K, p-AKT and AKT proteins in breast cancer cells. (A, B) The MMP levels of cancer cells treated with DEX at different concentrations were detected by immunofluorescence. (200×) (C-G) The expression levels of Bcl-2, Bax, p-PI3K, PI3K, p-AKT and AKT in breast cancer cells treated with DEX at different concentrations were measured by western blot. *P<0.05, **P<0.01, ***P<0.001 vs. Control. n=3 biological replicates. |
Results
To Explore the Effect of DEX on Mitochondrial Autophagy and Apoptosis of Breast Cancer Cells
The Viability of Cancer Cells Treated with Various Concentrations of DEX Was Measured by CCK-8
CCK-8 was used to test the viability of cancer cells treated with DEX at varying concentrations for 24, 48, and 72 h. The results demonstrated a progressive rise of cell vitality in a time-dependent manner. At 24/48/72 h, cell viability was considerably higher in DEX groups than control group in a dose-dependent way, suggesting that the higher the drug concentration, the higher the cell viability (P<0.05, Figure 1A). Overall, 100 ng/mL DEX yielded the optimal effect on enhancing the viability of breast cancer cells.
The Proliferation of Cancer Cells Treated with Different Concentrations of DEX Was Detected by Colony Formation Assay
Compared to control group, DEX groups exhibited stronger cell proliferation in a concentration-dependent manner. The number of cell colonies in the DEX groups presented the most significant changes after treatment with DEX at 100 ng/mL (P<0.01, Figure 1B and C).
The Expression of the LC3-II/LC3-I Protein in Cancer Cells Treated with DEX at Various Concentrations Was Examined by Western Blot
Relative to control group, LC3-II/LC3-I and PINK1/Parkin protein expression was decreased in DEX groups, and the decreasing trend was more apparent in the presence of 50/100 ng/mL DEX treatment (P<0.05, Figure 1D-H). These data suggested that DEX can prevent breast cancer cells from autophagy, with the strongest impact at 100 ng/mL.
The Apoptosis of Cancer Cells Treated with Various Concentrations of DEX Was Identified by Flow Cytometry
The apoptosis rate of breast cancer cells was significantly lower in DEX groups than in control group and showed a DEX-concentration-dependent decrease (P<0.01, Figure 1Iand J). The outcomes demonstrated that DEX might prevent breast cancer cells from apoptosis, with 100 ng/mL DEX having the strongest impact.
The MMP Levels of Cancer Cells Treated with Various Concentrations of DEX Was Tested by Immunofluorescence
Following DEX therapy, the MMP level in breast cancer cells was elevated in a dose-dependent manner, and 50/100 ng/mL DEX produced the most pronounced effects. (P<0.05, Figure 2A and B). These results revealed that DEX can raise the MMP level in breast cancer cells, reaching a peak effect at a concentration of 100 ng/mL.
 |
Figure 3 Effects of different drug treatments on PI3K, PI3K, p-AKT and AKT proteins, Cytochrome c and Caspase-3/7 activity in btreast cancer cells. (A-C) The expression of p-PI3K, PI3K, p-AKT, AKT in breast cancer treated with DEX at different time were measured by western blot. (D-E) The expresison level of Cytochrome c in breast cancer cells treated with DEX at different time were measured by western blot. (F) The acitivity of Caspase-3/7 was detected by Caspase -3/7 Activity Assay Kit. ###P<0.001 vs. 0h; *P<0.05, **P<0.01, ***P<0.001 vs Control. n=3 biological replicates. |
The Protein Expression of Cancer Cells Treated with Various Concentrations of DEX Was Quantified by Western Blot
The levels of p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 were significantly greater, whereas Bax level was lower in the DEX (20, 50, and 100 ng/mL) groups than those in the control group. As the concentration of DEX was increased, the expression levels of p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 protein in breast cancer cells were steadily augmented, while the level of Bax showed a downward trend (P<0.001, Figure 2C-G). Specifically, 50/100 ng/mL DEX significantly increased the levels of Bcl-2 and p-AKT/AKT in cancer cells (P<0.05, Figure 2C-G), while 20, 50, or 100 ng/mL DEX reduced Bax protein (P<0.05, Figure 2C-G). These findings showed that DEX can down-regulate the Bax protein and up-regulate p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 proteins levels in breast cancer cells, with DEX at 100 ng/mL exhibiting the greatest effects.
Further, effect of different treatment time of DEX (100 ng/mL) on p-PI3K, PI3K, p-AKT and AKT protein expression was detected by western blot. The result showed that DEX treatment has no significant effect on the phosphorylation of the PI3K/AKT signaling pathway within a short period (5 min/12 h), but it can significantly activate this pathway after a longer period (24 h) (P<0.001, Figure 3A-C), which is manifested by the continuous upregulation of the expression levels of p-PI3K and p-AKT proteins (P<0.001, Figure 3A-C).
Moreover, compared with the Control group, as the DEX concentration increased (25, 50, 100 ng/mL), the protein expression level of Cytochrome c in the cytoplasm showed a significant concentration-dependent decrease (P<0.05, Figure 3D-E), the activity of Caspase-3/7 decreased (P<0.05, Figure 3F).
Mitochondrial Autophagy and Apoptosis of Breast Cancer Cells Were Regulated via PI3K/AKT
Cancer Cell Viability Was Detected by CCK-8
CCK-8 was applied to measure the viability of cancer cells, and the results demonstrated that, at 24/48/72 h, following LY294002 treatment, the cancer cell viability was lowered compared to the control group, but was signally augmented after treatment with DEX (P<0.001, Figure 4A). LY294002 and DEX in combination resulted in enhanced cancer cell viability as compared to LY294002 monotherapy (P<0.001, Figure 4A). This suggested that LY294002 repressed cancer cell viability, while DEX reversed the effect of LY294002 and enhanced cell viability.
The Proliferative Ability Was Measured by Colony Formation Assay
When compared to the control group, there was a substantial increase in the number of cancer cell colonies in DEX groups (P<0.001, Figure 4B and C), yet a statistically insignificant decrease in the number of cell colonies in LY294002 group. The cancer cell colony number in the LY294002+DEX group was considerably higher than that in the LY294002 group (P<0.01, Figure 4B and C), but signally lower than that in the DEX group (P<0.05, Figure 4B and C). These data implied that LY294002 dampened breast cancer cell proliferation, which was reversed by DEX.
The Expression of LC3-II/LC3-I Protein Was Detected by Western Blot
As compared to control group, LY294002 therapy elevated but DEX diminished LC3-II/LC3-I ratio and PINK1/Parkin in cancer cells (P<0.05, Figure 4D-H). Moreover, the LC3-II/LC3-I ratio and PINK1/Parkin in LY294002+DEX group was lower than that in LY294002 group (P<0.01, Figure 4D-H), while being higher than that in DEX group (P<0.05, Figure 4D-H). These findings demonstrated that LY294002 raised the LC3-II/LC3-I ratio and PINK1/Parkin and boosted autophagy in breast cancer cells, which was offset by DEX intervention.
The Apoptosis of Cancer Cells Was Examined by Flow Cytometry
Breast cancer cells treated with LY294002 had a much higher apoptosis rate than the control cells. Conversely, DEX significantly lowered apoptosis rate when compared to the control cells (P<0.001, Figure 4I-J). The LY294002+DEX therapy caused lower apoptosis rate relative to the LY294002 monotherapy (P<0.001, Figure 4I-J), but higher apoptosis rate compared to the DEX monotherapy (P<0.001, Figure 4I-J). These data proved that LY294002 stimulated breast cancer cell apoptosis, which was counteracted by DEX.
The MMP Levels of Cancer Cells Were Quantitated by Immunofluorescence
In contrast with the control group, the LY294002 group exhibited a modest, albeit statistically insignificant, reduction in MMP level, whereas the DEX group presented remarkably elevated MMP level (P<0.05, Figure 5A and B). Further, the MMP level in the LY294002+DEX group was higher than that in the LY294002 group (P<0.05, Figure 5A and B), but lower than that in the DEX group (P<0.05, Figure 5A and B). These findings suggested that LY294002 can lower the MMP level of breast cancer cells, which was neutralized by DEX.
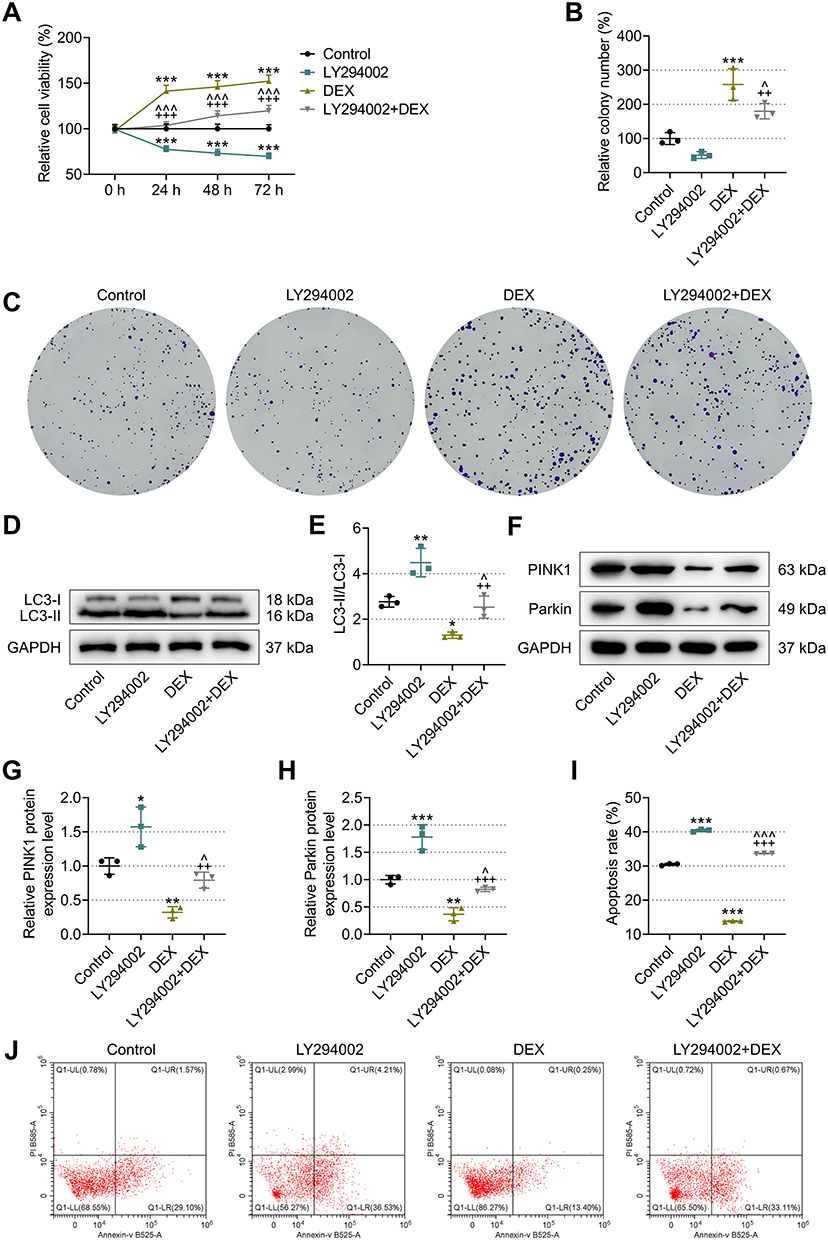 |
Figure 4 Effects of different drug treatments on proliferation, cell viability, autophagy and apoptosis of breast cancer cells. (A) CCK-8 was used to detect breast cancer cell viability after treatment with different drugs. (B, C) Colony formation assays were conducted to examine the proliferative ability of breast cancer cells. (40×) (D-H) The expression of LC3-II/LC3-I and PINK1/Parkin protein in breast cancer cells was detected by western blot. (I, J) The apoptosis of breast cancer cells was tested by flow cytometry. *P<0.05, **P<0.01, ***P<0.001 vs. Control; ^P<0.05, ^^^P<0.001 vs. DEX. ++P<0.01, +++P<0.001 vs. LY294002. n=3 biological replicates. |
Cancer Cell-Associated Proteins Were Quantified by Western Blot
The cancer cells in the LY294002 group had markedly lower p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 protein levels, but significantly higher Bax and Cytochrome c protein levels and Caspase-3/7 activity (P<0.05, Figure 5C-J). The DEX group showed a significant rise in p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 protein levels and a drop in Bax and Cytochrome c protein levels and Caspase-3/7 (P<0.01, Figure 5C-J). When comparison was made with LY294002 treatment, the up-regulation of p-PI3K/PI3K, p-AKT/AKT and Bcl-2 protein levels, as well as a down-regulation of Bax and Cytochrome c protein level and Caspase-3/7 was detected following LY294002+DEX treatment (P<0.01, Figure 5C-J). Relative to DEX group, LY294002+DEX group displayed down-regulation of p-PI3K/PI3K, p-AKT/AKT and Bcl-2 protein levels, but up-regulation of Bax and Cytochrome c protein level and Caspase-3/7 activity (P<0.05, Figure 5C-J). This implied that in breast cancer cells, LY294002 can up-regulate Bax, Cytochrome c, Caspase-3/7 activity and down-regulate p-PI3K/PI3K, p-AKT/AKT, and Bcl-2 protein levels, while DEX can counteract the effects of LY2940
 |
Figure 5 Effects of different drug treatments on MMP level and expression levels of Bcl-2, Bax, p-PI3K, PI3K, p-AKT and AKT proteins in breast cancer cells. (A, B) The MMP levels of cancer cells were assessed by immunofluorescence. (200×) (C-I) The expression levels of Bcl-2, Bax, p-PI3K, PI3K, p-AKT and AKT and Cytochrome c in breast cancer cells were detected by western blot. (J) The acitivity of Caspase-3/7 was detected by Caspase -3/7 Activity Assay Kit. *P<0.05, **P<0.01, ***P<0.001 vs. Control; ^P<0.05 vs. DEX. +P<0.05, ++P<0.01, +++P<0.001 vs. LY294002. n=3 biological replicates. |
Discussion
Breast cancer, one of the most frequent malignancies in women, is characterized by a high incidence, a dismal prognosis, and a high death rate.23 Molecular-level interventions for breast cancer have aroused considerable attention, such as the search for potential biomarkers and gene therapy, which may be novel strategies to enhance therapy efficacy.23 DEX is widely used for perioperative analgesia and sedation of breast cancer.24 Reports indicated that DEX activates hepatic stellate cells to stimulate IL-6 secretion and accelerates progression of hepatocellular carcinoma via STAT3 activation.25 DEX can also promote hypoxia-induced lung and colorectal cancer cell growth and metastasis.26 These observations underscore a close relation between DEX and tumourigenesis. Notably, in vitro studies revealed that DEX can also promote breast cancer cell migration and regulate its malignant behaviours.8 A clinical study also provided corresponding evidence that postoperative serum of DEX-treated patients is associated with breast cancer cell proliferation, invasion, and migration, which indirectly suggests that perioperative DEX may adversely affect the prognosis of breast cancer patients.27
Elucidating the mechanisms by which DEX fosters breast cancer cell growth is therefore essential for mitigating its adverse effects. Mitochondria, the principal ATP-generating organelles in both plant and animal cells, also play a crucial role in cell death and energy conversion.28 Maintaining normal MMP is necessary for adenosine triphosphate production and mitochondrial oxidative phosphorylation, and MMP stability supports the preservation of regular cellular physiological processes.29,30 In addition, mitochondria are a key part of apoptosis. When the cell receives the death signal, apoptotic proteins will undergo conformational changes on the mitochondrial membrane and oligomerize, forming perforating proteins that allow apoptotic molecules to enter the cytoplasm, and triggering a series of apoptosis procedures.31 Bax is a proapoptotic protein mainly located in the outer membrane of mitochondria,32 and is a single-stranded membrane protein involved in the regulation of apoptosis.33 Activation and aggregation of Bax can mediate the permeability of the mitochondrial membrane and the release of chemicals linked to apoptosis, like cytochrome c.33 Our study found that DEX not only up-regulated MMP in breast cancer cells, but also inhibited the apoptosis of cancer cells by up-regulating Bax protein. The content of autophagy-related molecular markers, LC3-I and LC3-II, varies with the degree of autophagy, and the ratio of LC3-II to LC3-I can be used to determine autophagy.34 This study found that DEX intervention inhibited autophagy by down-regulating LC3-II/LC3-I ratio in breast cancer cells.
The PI3K/AKT pathway is considered one of the key signaling pathways most closely associated with breast cancer, and is involved in glucose metabolism, differentiation, proliferation, DNA repair, apoptosis and migration.12 The activated PI3K/AKT signaling promotes breast cancer cell growth, survival, and metastasis.12 PI3K, a phospholipase family member composed of the catalytic subunit P110 and the regulatory subunit P85, functions as a proto-oncogene. Under the influence of cytokines, physical radiation, chemical stimulation and other adverse factors, PI3K is phosphorylated and activates the downstream effector molecule AKT. Activated AKT triggers downstream targeted molecules, leading to the spread and motility of cancer cells.35,36 To investigate the potential role of DEX in breast cancer development, we used the PI3K inhibitor LY294002 to treat cancer cells in vitro.37 Comparison with DEX intervention revealed that DEX could reverse the influence of LY294002 and promote breast cancer cell growth by up-regulate p-PI3K/PI3K and p-AKT/AKT proteins. However, whether the relationship between DEX and PI3K is direct or indirect remains unknown. A previous study showed that DEX activates the PI3K pathway through glucocorticoid receptor.38 Besides, the combination of a pan PI3K p110 inhibitor and DEX strengthens the DEX-induced up- or down-regulation of several genes involved in apoptosis.39 Therefore, more experiments are needed for further verification.
There are some limitations in this study. First, receptor blockade (involving α2-AR) or functional rescue assays have not been conducted. While our in vitro findings reveal a potent oncogenic signaling pathway activated by DEX, it is important to note that the concentrations used are supra-clinical (Perioperative DEX infusions achieve ≤1 ng/mL plasma levels40). Translating these results to the clinical context requires further investigation in models that employ clinically relevant dosing, such as patient-derived cells in vitro studies, long-term low-dose exposure studies or in vivo xenograft studies with controlled drug delivery to mimic surgical conditions.41 Despite these limitations, our study provides a foundational mechanistic insight. To build upon this work, future research should pursue several promising directions. Firstly, exploring combination strategies, such as co-administering DEX with PI3K/AKT pathway inhibitors, could assess whether the potential pro-tumor effects of DEX can be mitigated, thereby enhancing its safety profile. Secondly, given the complexity of cancer signaling networks, it is plausible that DEX engages additional pathways beyond PI3K/AKT. Investigating crosstalk with other oncogenic cascades or its influence on the tumor immune microenvironment would provide a more holistic understanding of its biological impact.
Conclusion
In conclusion, we demonstrate in vitro that DEX may promote the growth of breast cancer cells, while inhibiting mitochondrial autophagy and apoptosis by stimulating PI3K/AKT pathway. In subsequent studies, it is crucial to explore how to mitigate the promoting effect of DEX on the malignant biological behavior of breast cancer cells, and this will be a key direction for ongoing investigation in this study, which will provide new methods for the diagnosis and treatment of breast cancer.
Funding
This work was supported by the 2023 Zhejiang Province Medical and Health Science and Technology Plan Project [grant number 2023KY435].
Disclosure
The authors declare that there is no conflict of interest regarding the publication of this article.
References
1. Li Y, Zhang H, Merkher Y, et al. Recent advances in therapeutic strategies for triple-negative breast cancer. J Hematol Oncol. 2022;15(1):121. doi:10.1186/s13045-022-01341-0
2. Hao W, Qing L. Epidemiological trends and implications of breast cancer prevention and control in China and the world: interpretation of the Global Cancer Statistics Report 2018–2022. Chin J Basic Clin Stud Gen Surg. 2024;31(07):796–802.
3. Trayes KP, Cokenakes SEH. Breast Cancer Treatment. Am Fam Physician. 2021;104(2):171–178.
4. Wong GL, Manore SG, Doheny DL, Lo H-W. STAT family of transcription factors in breast cancer: pathogenesis and therapeutic opportunities and challenges. Semi Cancer Biol. 2022;86:84–106. doi:10.1016/j.semcancer.2022.08.003
5. Chen Z, Liu Z, Feng C, Jin Y, Zhao X. Dexmedetomidine as an adjuvant in peripheral nerve block. Drug Des Devel Ther. 2023;17:1463–1484. doi:10.2147/DDDT.S405294
6. Persson NDÅ, Uusalo P, Nedergaard M, Lohela TJ, Lilius TO. Could dexmedetomidine be repurposed as a glymphatic enhancer? Trends Pharmacol Sci. 2022;43(12):1030–1040. doi:10.1016/j.tips.2022.09.007
7. Chen JL, Liu ST, Huang SM, Wu ZF. Apoptosis, proliferation, and autophagy are involved in local anesthetic-induced cytotoxicity of human breast cancer cells. Int J Mol Sci. 2022;23(24):15455. doi:10.3390/ijms232415455
8. Waloejo CS, Musalim DAP, Budi DS, Pratama NR, Sulistiawan SS, Wungu CDK. Dexmedetomidine as an adjuvant to nerve block for cancer surgery: a systematic review and meta-analysis. J Clin Med. 2024;13(11):3166. doi:10.3390/jcm13113166
9. Nair AS, Saifuddin MS, Naik V, Rayani BK. Dexmedetomidine in cancer surgeries: present status and consequences with its use. Indian J Cancer. 2020;57(3):234–238. doi:10.4103/ijc.IJC_376_19
10. Chatterjee N, Pulipaka B, Subbalakshmi AR, Jolly MK, Nair R. Unraveling the dangerous duet between cancer cell plasticity and drug resistance. Comput Syst Oncol. 2023;3(3):e1051. doi:10.1002/cso2.1051
11. Zhang X, Zhao S, Yuan Q, et al. TXNIP, a novel key factor to cause Schwann cell dysfunction in diabetic peripheral neuropathy, under the regulation of PI3K/Akt pathway inhibition-induced DNMT1 and DNMT3a overexpression. Cell Death Dis. 2021;12(7):642. doi:10.1038/s41419-021-03930-2
12. Zhu K, Wu Y, He P, et al. PI3K/AKT/mTOR-targeted therapy for breast cancer. Cells. 2022;11(16):2508. doi:10.3390/cells11162508
13. Das N, Mukherjee S, Das A, Gupta P, Bandyopadhyay A, Chattopadhyay S. Intra-tumor ROS amplification by melatonin interferes in the apoptosis-autophagy-inflammation-EMT collusion in the breast tumor microenvironment. Heliyon. 2024;10(1):e23870. doi:10.1016/j.heliyon.2023.e23870
14. Mao L, Liu H, Zhang R, et al. PINK1/Parkin-mediated mitophagy inhibits warangalone-induced mitochondrial apoptosis in breast cancer cells. Aging. 2021;13(9):12955–12972. doi:10.18632/aging.202965
15. Andrieux P, Chevillard C, Cunha-Neto E, Nunes JPS. Mitochondria as a cellular hub in infection and inflammation. Int J Mol Sci. 2021;22(21):11338. doi:10.3390/ijms222111338
16. Xia J, Chu C, Li W, et al. Mitochondrial protein UCP1 inhibits the malignant behaviors of triple-negative breast cancer through activation of mitophagy and pyroptosis. Int J Bio Sci. 2022;18(7):2949–2961. doi:10.7150/ijbs.68438
17. Deng R, Zhang HL, Huang JH, et al. MAPK1/3 kinase-dependent ULK1 degradation attenuates mitophagy and promotes breast cancer bone metastasis. Autophagy. 2021;17(10):3011–3029. doi:10.1080/15548627.2020.1850609
18. Lin Y, Wei Y, Wei Y, et al. Dexmedetomidine alleviates oxidative stress and mitochondrial dysfunction in diabetic peripheral neuropathy via the microRNA-34a/SIRT2/S1PR1 axis. Int Immunopharmacol. 2023;117:109910. doi:10.1016/j.intimp.2023.109910
19. Li J, Wang K, Liu M, He J, Zhang H, Liu H. Dexmedetomidine alleviates cerebral ischemia-reperfusion injury via inhibiting autophagy through PI3K/Akt/mTOR pathway. J Mol Histol. 2023;54(3):173–181. doi:10.1007/s10735-023-10120-1
20. Qu J, Li J, Zhang Y, et al. AKR1B10 promotes breast cancer cell proliferation and migration via the PI3K/AKT/NF-κB signaling pathway. Cell Biosci. 2021;11(1):163. doi:10.1186/s13578-021-00677-3
21. Ou L, Zhu Z, Hao Y, et al. 1,3,6-trigalloylglucose: a novel potent Anti-Helicobacter pylori adhesion agent derived from aqueous extracts of Terminalia chebula Retz. Molecules. 2024;29(5):1161. doi:10.3390/molecules29051161
22. Ou L, Hao Y, Liu H, et al. Chebulinic acid isolated from aqueous extracts of Terminalia chebula Retz inhibits Helicobacter pylori infection by potential binding to Cag A protein and regulating adhesion. Front Microbiol. 2024;15:1416794. doi:10.3389/fmicb.2024.1416794
23. Emens LA. Breast cancer immunotherapy: facts and hopes. Clin Cancer Res. 2018;24(3):511–520. doi:10.1158/1078-0432.CCR-16-3001
24. Huang Z, Liu N, Hu S, Ju X, Xu S, Wang S. Effect of dexmedetomidine and two different doses of esketamine combined infusion on the quality of recovery in patients undergoing modified radical mastectomy for breast cancer - a randomised controlled study. Drug Des Devel Ther. 2023;17:2613–2621. doi:10.2147/DDDT.S422896
25. Chen P, Luo X, Dai G, et al. Dexmedetomidine promotes the progression of hepatocellular carcinoma through hepatic stellate cell activation. Exp Mol Med. 2020;52(7):1062–1074. doi:10.1038/s12276-020-0461-6
26. Chen H, Li G, Tan G, et al. Dexmedetomidine enhances hypoxia‑induced cancer cell progression. Exp Ther Med. 2019;18(6):4820–4828. doi:10.3892/etm.2019.8136
27. Liu Y, Sun J, Wu T, et al. Effects of serum from breast cancer surgery patients receiving perioperative dexmedetomidine on breast cancer cell malignancy: a prospective randomized controlled trial. Cancer Med. 2019;8(18):7603–7612. doi:10.1002/cam4.2654
28. Annesley SJ, Fisher PR. Mitochondria in health and disease. Cells. 2019;8(7):680. doi:10.3390/cells8070680
29. Begum HM, Shen K. Intracellular and microenvironmental regulation of mitochondrial membrane potential in cancer cells. WIREs Mech Dis. 2023;15(3):e1595. doi:10.1002/wsbm.1595
30. Sakamuru S, Zhao J, Attene-Ramos MS, Xia M. Mitochondrial membrane potential assay. In: High-Throughput Screening Assays in Toxicology. 2022:11–19.
31. Vringer E, Tait SWG. Mitochondria and cell death-associated inflammation. Cell Death Differ. 2022;30(2):304–312. doi:10.1038/s41418-022-01094-w
32. Uchime O, Dai Z, Biris N, et al. Synthetic antibodies inhibit Bcl-2-associated X Protein (BAX) through blockade of the N-terminal activation site. J Biol Chem. 2016;291(1):89–102. doi:10.1074/jbc.M115.680918
33. Spitz AZ, Gavathiotis E. Physiological and pharmacological modulation of BAX. Trends Pharmacol Sci. 2022;43(3):206–220. doi:10.1016/j.tips.2021.11.001
34. Cocco S, Leone A, Roca MS, et al. Inhibition of autophagy by chloroquine prevents resistance to PI3K/AKT inhibitors and potentiates their antitumor effect in combination with paclitaxel in triple negative breast cancer models. J Transl Med. 2022;20(1):290. doi:10.1186/s12967-022-03462-z
35. Yuan Y, Long H, Zhou Z, Fu Y, Jiang B. PI3K–AKT-targeting breast cancer treatments: natural products and synthetic compounds. Biomolecules. 2023;13(1):93. doi:10.3390/biom13010093
36. Alves CL, Ditzel HJ. Drugging the PI3K/AKT/mTOR pathway in ER+ breast cancer. Int J Mol Sci. 2023;24(5):4522. doi:10.3390/ijms24054522
37. Fang S, Wan X, Zou X, et al. Arsenic trioxide induces macrophage autophagy and atheroprotection by regulating ROS-dependent TFEB nuclear translocation and AKT/mTOR pathway. Cell Death Dis. 2021;12(1):88. doi:10.1038/s41419-020-03357-1
38. Zhang Y, Shi G, Zhang H, et al. Dexamethasone enhances the lung metastasis of breast cancer via a PI3K-SGK1-CTGF pathway. Oncogene. 2021;40(35):5367–5378. doi:10.1038/s41388-021-01944-w
39. Evangelisti C, Cappellini A, Oliveira M, et al. Phosphatidylinositol 3-kinase inhibition potentiates glucocorticoid response in B-cell acute lymphoblastic leukemia. J Cell Physiol. 2018;233(3):1796–1811. doi:10.1002/jcp.26135
40. Weerink MAS, Struys M, Hannivoort LN, Barends CRM, Absalom AR, Colin P. Clinical pharmacokinetics and pharmacodynamics of dexmedetomidine. Clin Pharmacokinet. 2017;56(8):893–913. doi:10.1007/s40262-017-0507-7
41. Liu H, Dilger JP, Lin J. Effects of local anesthetics on cancer cells. Pharmacol Ther. 2020;212:107558. doi:10.1016/j.pharmthera.2020.107558
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.