Back to Journals » Drug Design, Development and Therapy » Volume 20
Dendrophthoe pentandra (L.) Miq. as a Source of Anticancer Agents: A Review of Phytochemical Constituents and Preclinical Efficacy
Authors Anggraini L, Febriyanti RM ![]() , Muhaimin M
, Muhaimin M ![]() , Endrini S
, Endrini S
Received 8 August 2025
Accepted for publication 18 January 2026
Published 3 February 2026 Volume 2026:20 559286
DOI https://doi.org/10.2147/DDDT.S559286
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Leonidas Panos
Lovera Anggraini,1,2 Raden Maya Febriyanti,3 Muhaimin Muhaimin,3 Susi Endrini2
1Doctoral Program in Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, Indonesia; 2Department of Pharmacy, Faculty of Pharmacy and Health Sciences, Abdurrab University, Pekanbaru, Riau, Indonesia; 3Department of Biological Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, Indonesia
Correspondence: Raden Maya Febriyanti, Department of Biological Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, Indonesia, Tel +6222-84288888 Ext 3510, Email [email protected] Muhaimin Muhaimin, Department of Biological Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, Indonesia, Tel +6222-84288888 Ext 3510, Email [email protected]
Abstract: The emergence of drug-resistant cancers has led to the discovery of novel therapeutic agents. Medicinal plants are a promising source, with Dendrophthoe pentandra (L.) Miq. a parasitic plant used in ethnomedicine, showing promising anticancer potential. This study reviews preclinical evidence of the anticancer activity of D. pentandra, focusing on its phytochemical constituents and molecular mechanisms of action. A systematic search of Scopus and PubMed databases was conducted for preclinical studies published between 2015 and 2025. After screening, 13 articles met the eligibility criteria and were included in narrative synthesis. The 13 included studies demonstrated that D. pentandra extracts exhibit potent cytotoxic and antiproliferative effects against multiple cancer cell lines. The primary anticancer mechanism identified was the induction of apoptosis, which is frequently mediated by the upregulation of p53 and Bax proteins, downregulation of Bcl-2, and induction of cell cycle arrest at the G1/S or G2/M phase. Flavonoids, particularly quercetin, have been identified as the key bioactive phytoconstituents that contribute to these effects. In vivo studies further support these findings, showing that D. pentandra extract can inhibit tumor progression in colitis-associated cancer models by reducing inflammatory markers such as myeloperoxidase (MPO).
Keywords: anticancer activity, Dendrophtoe petandra, preclinical studies, parasitic plants phytoconstituent
Introduction
Over the past three decades, cancer mortality has shown a significant decline, dropping by approximately 34% since 1991, primarily due to decreased smoking rates, advances in the early detection of certain cancers, and the emergence of more effective treatments including targeted therapies and immunotherapy.1 Despite this progress, efforts to prevent cancer have remained insufficient. However, the incidence rates of six of the ten most prevalent cancers in individuals under the age of 65 continue to rise in LMICs- and middle-income countries (LMICs).2 By 2025, the United States is projected to record more than 2 million new cancer cases and more than 618 thousand cancer deaths.1 Although mortality rates have declined due to better prevention, detection, and treatment, significant disparities persist.3,4
Plant-derived bioactive compounds have gained increasing attention as potential alternatives or complementary therapies for cancer treatment, particularly in developing countries, where they offer low-cost, accessible solutions with fewer adverse effects.5–7 Between 1981 and 2014, approximately 35% of the anticancer drugs were derived from plant-based compounds, underscoring their importance in cancer care.8 Compounds from Taxus brevifolia have become the cornerstone of cancer therapy.9,10
These therapeutic effects are often attributed to secondary metabolites or phytoconstituents such as alkaloids, terpenoids, and flavonoids. Unlike primary metabolites, which are essential for basic cell function, these compounds often serve defensive roles in plants and possess potent bioactivity. Research has demonstrated that many phytoconstituents can selectively target cancer cells by interfering with tumor growth, reducing inflammation, and mitigating oxidative stress. Flavonoids, for example, have been shown to modulate reactive oxygen species (ROS), arrest the cell cycle, induce apoptosis (programmed cell death), and suppress cancer cell proliferation and invasion.11
Among the promising botanical sources, Dendrophthoe pentandra (L.) Miq., (Loranthaceae) shows a broad range of bioactivity.12 This semi-parasitic plant that can grow on variety of host plants with woody shrub branches and can grow to a height ranging from 0.5–2.0 m.11,12 Depending on its host plant, D. pentandra is known by different names. In Brazil, it is referred to as Psittacanthus robustus Mart; in the Samaru region of Niger, it is called Tapinanthus globiferus; and in Central Mexico, it is known as Archetobium globosum.13–15 The bioactivities and phytochemicals of D. pentandra can vary depending on its host plant, as mistletoe absorbs nutrients and self-defense compounds from its host to support its survival and evade detection by herbivorous animals.16 Phytochemical analysis revealed that D. pentandra contains a variety of secondary metabolites including alkaloids, flavonoids, phenolics, saponins, and terpenoids.17 These compounds contribute to the plant’s potential medical benefits, including antioxidant,11 immunomodulatory,17 anti-inflammatory,18 and anticancer activities, as well as anti-proliferative and cytotoxic activities.19,20
While a previous review summarized the general pharmacology of D. pentandra,16 systematic synthesis of mechanistic evidence for its anticancer properties is currently lacking.17 Such rigorous evaluation is essential to identify both the most promising therapeutic pathways and the critical gaps in existing research, thereby guiding future studies toward potential clinical translation. Therefore, this systematic review aimed to identify and synthesize preclinical studies investigating the anticancer effects of D. pentandra, summarize its known phytochemical constituents associated with anticancer activity, and analyze the proposed molecular mechanisms of action.
Methods
Search Strategy
A systematic search was performed for articles published between 2015 and 2025 using PubMed and Scopus databases. Keywords such as Dendrophthoe pentandra, anticancer activity, and phytochemical properties also incorporated phrases relevant to the topic. Synonyms and related terms were used interchangeably throughout this process. Boolean operators such as “AND” and “OR” were employed to refine the search and include or exclude specific terms. Specifically, the PubMed database was queried with the keywords (“Dendrophthoe pentandra” OR “Loranthus pentandrus” OR “D. pentandra”) AND (phytochemical OR “secondary metabolite” OR flavonoid OR phenolic OR alkaloid OR terpenoid) AND (anticancer OR antitumor OR cytotoxic OR antiproliferative OR apoptosis) within the Abstract and free full-text sections from the last 10 years, resulting in 19 articles. In the Scopus database, the exact keywords used in the textual content resulted in a total of 74 articles. All identified publications were assessed and those meeting the inclusion criteria focusing on the anticancer properties of D. pentandra were reviewed.
Data Selection and Collection Process
Study selection was conducted through a systematic two-phase process after duplicate articles were removed from the initial search results. In the first phase, two reviewers (LA and RMF) independently screened the titles and abstracts of all identified records using the web-based tool Rayyan.ai. Articles that met the initial criteria were advanced to the second phase of comprehensive full-text assessment. The included studies had to be preclinical investigations, either in vitro or in vivo, published in English between January 2015 and August 2025. This study evaluated extracts and isolated compounds from D. pentandra and reported outcomes related to anticancer activity, such as cytotoxicity, apoptosis, or cell cycle arrest. Accordingly, articles were excluded if they were reviews, commentaries, clinical trials, or if their primary focus was not on cancer or D. pentandra. Key data extracted from each article included medicinal plants (family), collection location and year, plant part used, extraction method and solvent, class of metabolites, phytochemical constituents, quantification of phytoconstituents, cell line used, and mechanisms of anticancer activity. Thus, 13 articles were selected for in-depth analysis (Figure 1). Further search was conducted to gather additional sources to support this analysis.
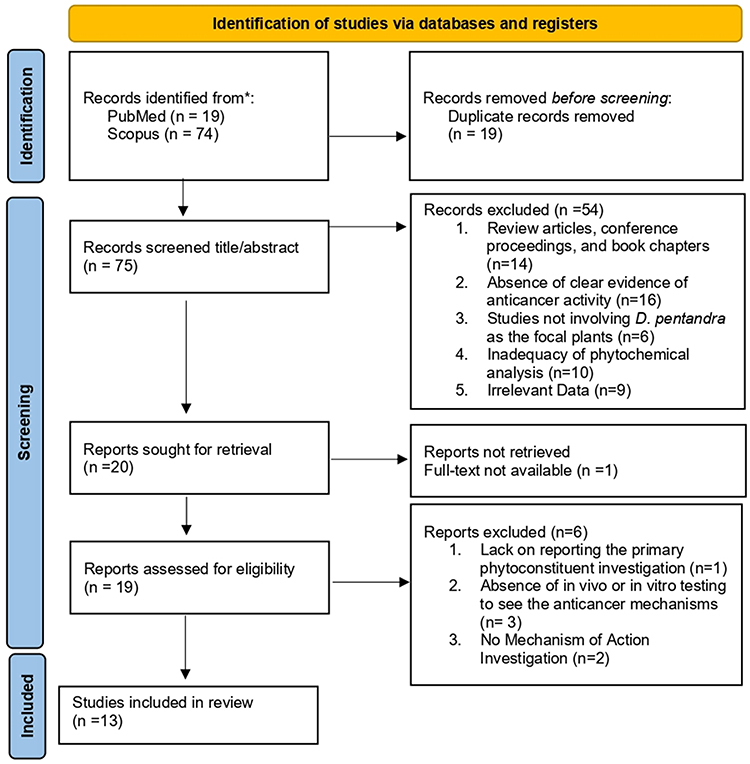 |
Figure 1 PRISMA Flow Diagram. Adapted from Page MJ, McKenzie JE, Bossuyt PM, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021:372:n71. Creative commons.21 |
Data Synthesis
Owing to significant heterogeneity across the included studies in terms of host plants, extraction methods, solvents, cancer models, and outcome measures, a quantitative meta-analysis was not feasible. Therefore, a narrative synthesis of the evidence was conducted. The findings were qualitatively summarized and grouped thematically based on the reported phytochemical constituents and primary anticancer mechanisms investigated. Additionally, to provide a more holistic context and to fill the gap for a comprehensive review, this synthesis includes dedicated sections on the morphological overview and traditional uses of D. pentandra. This broader approach provides a complete resource for researchers by integrating the plant’s ethnobotanical and biological backgrounds with the systematic synthesis of modern preclinical evidence.
Characteristics of Included Studies
Most studies described Southeast Asia as the origin location of the plants, including Indonesia (n = 9), Malaysia (n = 3), and Thailand (n = 1). Of the 13 articles, five discussed phytoconstituents in D. pentandra in any host plant, and eight articles described anticancer activity in preclinical studies, including in vitro and in vivo studies. These studies confirm that the metabolites in D. pentandra have anticancer mechanisms and activities.
In preclinical studies, various types of cancer cell lines and inducers were used, especially MCF-7 cells, which were selected more frequently (70%) than the CML Cell line (20%) and other cell lines (10%). This cell line has been extensively used because of its ideal characteristics for in vitro studies, particularly its ability to retain estrogen receptor expression. This allows the study of estrogen response and hormone resistance, making it an ideal model for hormone-responsive breast cancer.22
Results
Morphological Overview
Dendrophthoe pentandra (L.) Miq. (Loranthaceae) is a parasitic shrub found on various tropical and subtropical trees and is commonly found in urban parks and residential areas where it is used as an ethnomedicinal plant.23 This parasitic plant damages its host by stunting growth, severing branches, and ultimately, killing the host.24,25 Morphological variations in D. pentandra depend on the location of the host plant, specifically in different areas of the wood and bark of the stem.26,27 The plant attaches to the host through sucker roots modified into haustoria (Figure 2).
 |
Figure 2 Dendrophthoe pentandra plant attaches itself to a host plant [Private Documentation, 2025]. |
The morphological characteristics of D. pentandra include a shrubby form with ligneous stems that penetrate host tissue. The plant comprises roots, stems, leaves, fruits, and seeds.28,29 The roots are designed and known as haustoria and are pricked into host tissues and connected to the xylem or phloem of the host. The haustoria are spherical, and their size varies depending on the host species, bark hardness, and light intensity.30 Furthermore, it appears to be similar to a ball attached to a branch.31 From the closest part of the branch, nutrients and photosynthates migrated primarily to D. pentandra as opposed to the distal portion. The stems are shrubs with woody branches that grow upright and have drooping branches arranged in an axillary pattern. They were grayish-brown in color and featured scattered lenticels.30,32
The leaves of D. pentandra, a highly adaptable species, exhibit significant size variation depending on the infested host plant. In Indonesia, D. pentandra leaves range in size from 7.1 to 16 cm in length, 3–7 cm in width, and 0.3–0.7 mm in thickness.30 In contrast, in Singapore, the leaves are narrower and have a sub-orbicular shape, measuring 8–10 cm in length and 5–15 cm in width, and 5–20 mm thickness.29
Furthermore, the leaves varied from dark to light green and are arranged alternately along the branches. The young shoots were covered with pubescent (tiny pubescent hairs), and the young leaves were purple-red. The mature leaves are green, with thick, leathery leaf blades and a glossy surface, have 2–4 pairs of lateral veins, base cuneate or obtuse, apex acute or rounded, and glabrous. They have opposite alternate phyllotaxes, with acute, acuminate, or obtuse tips, and acute, attenuated, or truncated bases.28–30,32
The inflorescences of D. pentandra were cylindrical and were categorized as hermaphrodite flowers.28,32 These flowers contain both male and female reproductive organs, specifically the stamen (male) and pistil (female), within the same inflorescence.33 The plants consisted of 4–13 flowers.29 In Indonesia, inflorescences range in size with a 3–6 cm length. The pistil length varies from 1.1–2.1 cm, the stamens are 1–2.1 cm long, and the crown tube measures 0.7–1.2 cm; the corolla lobe length varies from 4 to 9 cm.30 Moreover, in Singapore, each flower is approximately 1.5–2 cm long, and fully developed flowers usually form clusters of 3–10 flowers.29 Each cluster can occur individually, in pairs, or in threes. Furthermore, the flowers initially develop from orange buds into blossoms characterized by reddish, grayish, or white stellate hairs. Upon blooming, the flowers exhibit five narrow petals that retract, revealing an interior orange, an exterior greenish hue, and a greenish exterior.28,29
The fruits of D. pentandra are small, egg-shaped, and resemble fleshy red berries, exhibiting a size of 8.4 to 11.7 mm in length and 6.1 to 8.4 mm in width. The fruit was either minutely pilose or glabrous.32 The young fruits were yellowish-green to pink, whereas the ripened fruits were red in color.30 These fruits were single-seeded drupes without stony endocarps. Each fruit contained one seed covered with the sticky part of the fruit. This sticky part, called viscin, is comprised of cellulosic strands confined by mucilaginous pectic materials. Viscin plays a crucial role in seed dispersal by helping seeds adhere to the host plant.34 The seeds’ size is estimated within 0.6 to 1 cm in length and 0.2 to 0.5 cm in width, and they are greenish or yellowish-white in color.30 These seeds are dispersed by birds that consume D. pentandra fruit.31
Traditional Uses
Almost all parts of D. pentandra have been traditionally used for medicinal and alternative therapeutic purposes.35 These uses have been documented in Ayurvedic and other ethnomedical systems because of the broad therapeutic effects of the plant on human health.36 The plant is often crushed and mixed with other ingredients to produce edible teas, decoctions, or plasters to treat various diseases.23,37,38 Raw materials are also fed to livestock to improve growth performance. In folklore, D. pentandra is given various trivial names reflecting cultural differences. Additionally, their therapeutic effects vary across different ethnomedicinal systems.23
However, D. pentandra is relatively rare, and its medicinal use has primarily been reported in Southeast Asia, especially in Indonesia. It is commonly known as “Benalu” in Indonesia and is used to treat skin diseases, minor sores, coughs, post-childbirth ailments, and hypertension.30,34,39 In Sulawesi, D. pentandra is used to treat mumps and cancer,11,35 whereas in Central Kalimantan, it is used to treat dyspnea, diabetes, and tonsillitis.40 In some regions of Thailand, plants are used to treat high blood pressure, chronic diseases, and immunological disorders.41 Additionally, D. pentandra leaves have antioxidant and antidiabetic properties.35,42–44
Phytochemical Constituents
Phytochemical analysis of D. pentandra has revealed a variety of bioactive compounds that contribute to its medicinal properties. Initial qualitative tests, including phytochemical screening and thin-layer chromatography (TLC) analysis, confirmed the presence of key secondary metabolites such as alkaloids, flavonoids, saponins, and tannins. Quantitative quantification using GC-MS, HPLC, and 1H-NMR also identified specific phytoconstituents such as quercetin, quercitrin, and morin.45–47 These compounds were consistently detected across the various solvent fractions, indicating their abundance and stability in plants (Table 1).
 |
Table 1 Phytochemical Content of D. Pentandra |
Flavonoids are the primary phytochemical constituents, with the specific key compounds identified as quercetin and quercitrin (quercetin-3-O-rhamnoside). Additionally, other flavonoid compounds such as genistein, 5,7,3′,4′-pentamethoxyflavone, pelargonidin 3-glucoside, morin, 7-hydroxy-1-methoxy-2-methoxy xanthone, kaempferol-3-O-β-D-glucopyranoside, and flavonoid glycosides contribute to D. pentandra bioactivity.48–50 Quercetin, in particular, was present in significant amounts (8062 mg), highlighting its potential as a therapeutic agent.46
While the plant contains diverse phytochemical constituents such as flavonoids, phenolics, alkaloids, terpenoids, and saponins that may contribute to anticancer and antioxidant activity, the precise molecular mechanisms underlying these effects remain inadequately understood. Reports suggest involvement of apoptosis and cell cycle arrest pathways in vitro, but detailed mechanistic studies in mammalian systems are lacking. D. pentandra also exhibits antioxidant properties in several solvent fractions, indicating potential modulation of oxidative stress pathways, but these results are primarily based on in vitro DPPH assays and phytochemical screenings.
Anticancer Activities
Diverse mechanisms of anticancer action in plants, such as cytotoxicity, apoptosis, and anti-proliferation, have paved the way for investigation D. pentandra’s anticancer activities of D. pentandra. Through rigorous in vitro and in vivo assays, D. pentandra emerged as a promising therapeutic agent against a spectrum of cancer types. This plant has demonstrated potent anticancer activity via mechanisms such as apoptosis induction, cell cycle arrest, and inhibition of proliferation and metastasis. A detailed overview of the anticancer mechanisms of the bioactive compounds extracted from D. pentandra against various cancer types is presented in Table 2.
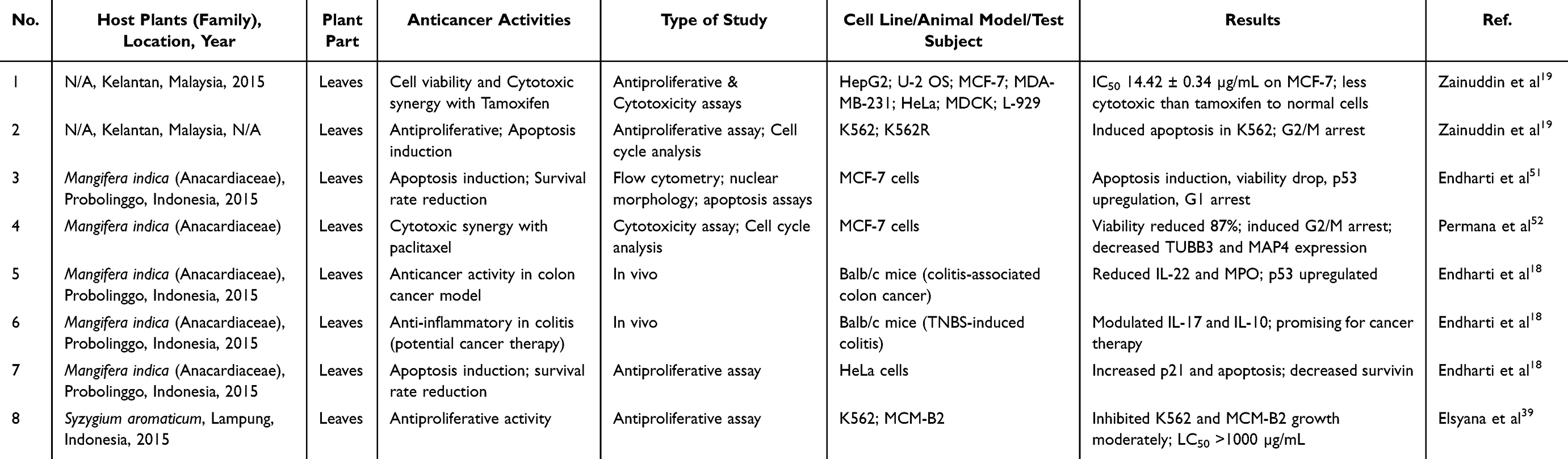 |
Table 2 Anticancer Properties of Plant’s Extract |
Although numerous studies have highlighted the bioactive potential of D. pentandra as a source of anticancer agents, several important limitations should be considered. Most of the current evidence is derived from in vitro assays and early preclinical screenings, such as cytotoxicity tests and antiproliferative evaluations against cancer cell lines. For example, cytotoxicity and antiproliferative activity of various fractions of D. pentandra extracts have shown significant effects in vitro with LC50 values as low as ~55 µg/mL in some fractions, yet these results are not directly translatable to human clinical contexts without further validation.12
Another significant gap relates to toxicity and bioavailability: few studies have systematically evaluated safety profiles, pharmacokinetics, or systemic toxicological effects. Acute and subchronic toxicity studies are limited, and data on in vivo pharmacodynamic parameters remain sparse.26 Furthermore, environmental factors, such as variation due to host plant species, can influence secondary metabolite profiles, potentially affecting reproducibility and pharmacological outcomes.9
Importantly, there is a lack of controlled clinical trials assessing anticancer efficacy or safety in humans. Most existing studies do not provide structured methodological quality assessments, including bias evaluation, sample size adequacy, or reproducibility across independent experiments. Therefore, while D. pentandra shows promising anticancer and related biological activities in preclinical settings, rigorous in vivo studies and clinical trials are urgently needed to establish safety, efficacy, and mechanisms of action before considering therapeutic applications.
Induction of Cell Cycle Arrest
Dysregulation of the cell cycle is a hallmark of cancer and is characterized by unnatural disruption of cell cycle progression through chemical or genetic manipulation, resulting in uncontrolled proliferation.53,54 This arrest often coincides with changes in cyclin-dependent kinases and cell cycle regulatory proteins, as well as the inhibition of Fatty Acid Synthase (FAS) and aryl hydrocarbon receptor (AHR).55
Studies have demonstrated that D. pentandra exhibits anti-proliferative activity in MCF-7 breast cancer cells, inducing G2/M phase arrest with a significant increase in cell cycle inhibition from 36% to 65%.52 In the K562 cell line, this plant also showed G2/M phase cell cycle arrest.56 Previous studies indicated that cancer cells become more susceptible to DNA-damaging agents when MAP4 expression is uncontrolled, suggesting that MAP4 plays a role in promoting breast cancer cell proliferation by regulating microtubule dynamics.57,58
D. pentandra is reported to contain quercetin, genistein, morin, and kaempferol-3-O-β-D-glucopyranoside.59 These polyphenols have documented actions that converge on mechanisms including G2/M checkpoint control and microtubule regulation. Quercetin and genistein suppressed cyclin B1–CDK1 activity and activated the p53/p21 axis. Both compounds can interfere with tubulin polymerization and sensitize cells to microtubule targeting or DNA-damaging agents. Furthermore, Morin and kaempferol derivatives similarly downregulated survivin and modulated mitogen-activated protein kinase/phosphoinositide 3-kinase (MAPK/PI3K) signaling, culminating in G2/M arrest and apoptosis.60 These activities were consistent with the observed G2/M blockade in MCF-7 and K562 cells and the reduced MAP4/TUBB3 expression reported after combination treatment.
Inhibition of Cell Proliferation
Cancer cells have the abnormal ability to survive beyond their typical lifespan and proliferate uncontrollably. This mechanism inhibits cell proliferation and is a critical strategy for cancer treatment. Targeting the high proliferation rate of cancer cells through specific interventions has proven to be effective in slowing or halting cancer progression.61 These interventions can work through various mechanisms, naturally in healthy cells, and through therapeutic strategies, such as cancer treatment.
Bioactive flavonoids, such as quercetin, have been shown to reduce cancer cell viability in vitro by downregulating pro-survival signaling pathways (eg, Akt and NF-kB) and upregulating cell cycle inhibitors.62,63 D. pentandra, which contains bioactive flavonoids, has demonstrated antiproliferative activity with an IC50 value of 1.25 µg/mL, and induces apoptosis in T47D cells (a breast cancer cell line). However, the methanol extract of D. pentandra showed an IC50 value of 14.42 ± 0.34 µg/mL against MCF-7 breast cancer cells.64 Notably, there was no significant effect on the proliferation of non-malignant cells such as L-929 and MDCK.19
Induction of Apoptosis
Apoptosis, also known as programmed cell death, is a highly regulated process essential for maintaining cellular balance, facilitating tissue development, and removing damaged or unwanted cells. Unlike necrosis, which is an uncontrolled and inflammatory process, apoptosis occurs in a well-organized manner, minimizing disruption to surrounding tissues. This process involves two primary pathways: the intrinsic (mitochondrial) and extrinsic (death receptor) pathways. Both pathways activate caspases, a family of proteases that causes cell death.65
The intrinsic pathway is regulated by the BCL-2 family of proteins, which includes both anti-apoptotic proteins (such as BCL-2 and BCL-xL) and pro-apoptotic proteins (such as BAX and BAK), as well as BH3-only proteins (such as BIM and BID). When cells experience stress, these proteins maintain the integrity of the mitochondrial membrane and release cytochrome c. This release forms an apoptosome, which activates caspase-9. The subsequent activation of executioner caspases such as caspases-3 and −7 results in the breakdown of cellular components.54,65
Recent studies have identified flavonoids such as quercetin and kaempferol as potent modulators of the apoptosis pathway. These bioactive compounds promote intrinsic apoptosis by upregulating pro-apoptotic proteins, such as Bax and Bad, activating caspases-3 and −7, and downregulating anti-apoptotic proteins, such as Bcl-2 and Bcl-xL.66 Additionally, quercetin and kaempferol inhibit proliferative signaling pathways, including the PI3K/Akt and MAPK/ERK pathways, thereby reducing tumor growth and promoting cancer cell death.67 These findings highlight the potential of flavonoids as adjuncts to conventional cancer therapies, particularly because of their ability to target key apoptotic regulators and inhibit cancer cell survival.
The anticancer potential of D. pentandra has also been explored and yielded promising results. The methanol extract of D. pentandra demonstrated significant antiproliferative effects on MCF-7 breast cancer cells, with an IC50 value of 14.42 ± 0.34 µg/mL, and induced apoptosis by modulating key apoptotic proteins. Notably, the expression of p53, a critical tumor suppressor protein, was significantly elevated in MCF-7 cells treated with D. pentandra methanol extract of D. pentandra (p < 0.0001).51 Additionally, the pro-apoptotic protein Bax was upregulated, whereas the anti-apoptotic protein Bcl-2 was downregulated in a time-dependent manner, further supporting the induction of apoptosis. These findings suggest that p53 activation may play an essential role in the anticancer effects of D. pentandra by promoting Bax expression and inhibiting Bcl-2 expression, leading to mitochondrial dysfunction and cell death.68 These results align with studies on other anticancer agents such as tamoxifen, which also modulates the expression of Bcl-2 family proteins to induce apoptosis in breast cancer cells.69
Effect on Oxidative Stress
ROSs are partially reduced oxygen metabolites with strong oxidizing properties. At high concentrations, ROS can cause significant damage to the cellular components. However, at moderate levels, they are essential for cellular signaling and maintaining homeostasis. In cancer cells, elevated ROS levels can induce cell cycle arrest, senescence, and apoptosis through both intrinsic (mitochondrial) and extrinsic (death receptor) apoptotic pathways.54,70
The mechanisms by which plants scavenge ROS have been extensively studied, revealing a complex and multifaceted defence system. Key antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) play crucial roles in neutralizing ROS.71 D. pentandra demonstrated notable antioxidant activity, inhibited DPPH radicals, and showed positive results in the FRP assay and total antioxidant content analysis. Phytochemical analysis of D. pentandra extracts identified important phenolic compounds, including quercetin and rutin, which act as free-radical scavengers. These compounds neutralize ROS by donating hydrogen atoms or electrons.72
Most ROS are generated in the mitochondrial electron transport chain (METC), where the electrons transferred through the chain create a force that moves protons across the inner mitochondrial membrane.73 While electrons are typically safely transferred through redox pathways, some may leak prematurely from complexes I, II, and III, leading to the one-electron reduction of oxygen to the superoxide anion (O2·−), which is subsequently converted to hydrogen peroxide (H2O2). When the permeability of the mitochondrial membrane increases, ROS produced in the METC can leak into the cytosol, triggering inflammation through various signaling pathways.74 ROS can activate inflammatory mediators such as nuclear factor kappa B (NF-κB) and mitogen-activated protein kinases (MAPKs), which in turn regulate the expression of pro-inflammatory cytokines and enzymes.75 A key enzyme in this inflammatory process is myeloperoxidase (MPO), which is produced by activated neutrophils and plays a central role in ROS-mediated inflammation. MPO catalyzes the conversion of hydrogen peroxide into hypochlorous acid (HOCl), a potent oxidant that contributes to tissue damage and propagation of inflammation.76
In vivo studies reinforced these in vitro findings, showing that D. pentandra extract administration significantly reduced the levels of IL-22 and MPO.18 Mice treated with D. pentandra extract at doses of 250 mg/kg and 500 mg/kg D. pentandra extract showed significantly lower MPO levels than the untreated group (p = 0.01; p = 0.00), suggesting that D. pentandra extract offers protection against inflammation. Neutrophils, which amplify inflammation by secreting MPO and releasing inflammatory mediators, are likely inhibited by DPE treatment.
Flavonoids, which are polyphenolic compounds with antioxidant properties, significantly modulate ROS levels and protect cells from oxidative stress.77 Among flavonoids, quercetin has notable chemopreventive properties in various cancer types. It reduces the proliferation of hepatocellular carcinoma (HepG2) cells by lowering intracellular ROS levels and paradoxically increasing ROS levels and apoptosis in gastric (AGS) and breast (MCF-7) cancer cells.78
Collectively, D. pentandra, which contains quercetin, is known to inhibit inflammatory pathways by blocking IκB phosphorylation, underscoring the therapeutic potential of D. pentandra in modulating inflammation and cancer progression.79 Additionally, D. pentandra showed promise in improving the histopathological features of animal models of colon cancer.80,81 These findings suggest that D. pentandra is a promising natural source for the development of novel anti-cancer therapies. However, further studies are needed to confirm its efficacy and establish its potential as an adjunct or alternative to conventional chemotherapy, which requires additional clinical validation.
Although the study demonstrated significant effects of 250 and 500 mg/kg doses in mice, the safety profile and potential toxicity of D. pentandra at higher or prolonged doses were not explored. Determining the optimal therapeutic dose and identifying long-term adverse effects are crucial for future studies.
Comparison with Other Mistletoe Species
Mistletoe derived plants have long attracted scientific interest for their anticancer potential, with Viscum album (European mistletoe) being the most extensively studied and clinically utilized species, particularly in complementary oncology in Europe. Extracts of V. album are characterized by the presence of mistletoe lectins (ML I–III), viscotoxins, polysaccharides, and triterpenes, which exhibit immunomodulatory, cytotoxic, and pro-apoptotic effects.82 These bioactive proteins are known to stimulate immune responses, including activation of natural killer cells, macrophages, and T lymphocytes, thereby contributing to antitumor immunity in addition to direct cytotoxicity.83
In contrast, Dendrophthoe pentandra demonstrates a distinct phytochemical and mechanistic profile compared to V. album. Rather than being dominated by protein-based toxins, D. pentandra is rich in polyphenolic compounds, particularly flavonoids such as quercetin, quercitrin, morin, and kaempferol derivatives, which are primarily responsible for its anticancer activity. These compounds exert anticancer effects mainly through oxidative stress modulation, mitochondrial-mediated apoptosis, and cell cycle arrest, rather than immune stimulation alone.23,28
Another key distinction lies in the host-dependent variability of phytochemical composition. While V. album is typically cultivated under controlled conditions to ensure relatively consistent extract composition for pharmaceutical use, D. pentandra exhibits substantial variation in metabolite content depending on the host plant species. This host-specific dependency has been shown to influence flavonoid concentration, antioxidant capacity, and biological activity, presenting challenges for extract standardization but also offering opportunities for identifying host-optimized bioactive profiles.27
From a translational perspective, V. album extracts have advanced further into clinical application, with multiple studies reporting improvements in quality of life and supportive outcomes in cancer patients, although definitive survival benefits remain debated.84,85 In contrast, research on D. pentandra remains largely at the preclinical stage, with evidence primarily derived from in vitro and limited in vivo models. Nevertheless, the strong flavonoid-driven anticancer mechanisms observed in D. pentandra suggest that it may complement existing mistletoe-based therapies by targeting tumor cell proliferation and apoptosis through non-protein, low molecular weight compounds, which may offer advantages in terms of stability and safety.
Overall, while V. album represents a clinically established mistletoe species with predominant immunomodulatory activity, D. pentandra occupies a distinct mechanistic niche as a flavonoid-rich anticancer candidate. These differences underscore the importance of further systematic and comparative investigations to determine whether D. pentandra can serve as a complementary or alternative source of mistletoe-derived anticancer agents.
Limitations of The Study
Despite providing a comprehensive synthesis of the anticancer potential of Dendrophthoe pentandra, several important limitations should be acknowledged, particularly regarding the quality and consistency of the included preclinical studies. First, most of the included studies relied on in vitro cancer cell line models, predominantly MCF-7 breast cancer cells, with limited representation of other cancer types. Although in vitro assays are valuable for mechanistic exploration, they do not fully replicate the complexity of tumor microenvironments in vivo. Moreover, only a small proportion of studies employed in vivo animal models, and these used heterogeneous dosing regimens and outcome measures, limiting cross-study comparability.
Second, there was a lack of standardized methodological quality assessment across studies. Variability was observed in extraction methods, solvent systems, phytochemical characterization, and reporting of extract standardization. Many studies did not clearly report sample sizes, biological replicates, randomization procedures, or blinding, which may introduce experimental bias and limit reproducibility. In addition, few studies included appropriate positive controls or benchmark anticancer agents for comparison.
Third, although apoptosis induction and cell cycle arrest were frequently reported, the depth of mechanistic validation varied substantially. Most studies relied on protein expression analysis without complementary genetic or pharmacological pathway inhibition, making it difficult to confirm causal mechanisms. Furthermore, synergistic or antagonistic interactions among phytochemical constituents particularly flavonoids such as quercetin and kaempferol remain largely unexplored.
Fourth, toxicity, bioavailability, and pharmacokinetic data were insufficiently addressed. While some studies reported selective cytotoxicity toward cancer cells, comprehensive safety evaluations, including long-term toxicity and organ-specific effects, were largely absent. The bioavailability of key compounds, especially quercetin, which is known to exhibit limited oral bioavailability, was not systematically evaluated in the reviewed studies.
Finally, the absence of clinical or translational studies represents a major limitation. Although the preclinical evidence is promising, differences in metabolism, immune response, and tumor biology between animal models and humans necessitate cautious interpretation. Future research should prioritize standardized experimental designs, rigorous quality control, dose–response and toxicity profiling, and well-designed clinical studies to support the translational development of D. pentandra derived anticancer agents.
Conclusion
D. pentandra has been noted for contains a variety of bioactive compounds, including alkaloids, flavonoids, and saponins, which exhibit diverse biological properties, such as anti-inflammatory, antioxidant, and anticancer activities. In terms of anticancer potential, bioactive plant compounds have demonstrated promising effects in inhibiting cancer cell growth through various mechanisms.
Among the phytoconstituents, quercetin, the primary phytoconstituent of D. pentandra, showed diverse anticancer properties by modulating ROS, influencing cell cycle progression by triggering G1 and G2 arrest, and regulating the proliferation of cancer cells. The proposed anticancer mechanisms of D. pentandra phytoconstituents are primarily associated with their ability to disrupt proliferative cellular processes, induce cell cycle arrest, and activate apoptosis in malignant cells. Overall, the current evidence highlights D. pentandra as a promising source of anticancer agents, supported by its diverse phytochemical profile and consistent antiproliferative, pro-apoptotic, and immunomodulatory activities demonstrated across in vitro and in vivo studies. Beyond establishing its therapeutic potential, these findings provide important scientific justification for advancing D. pentandra into more rigorous pharmacological evaluations. In the context of rising cancer prevalence and the limitations of existing chemotherapy including toxicity, financial burden, and resistance the bioactive constituents of D. pentandra offer a valuable foundation for the development of safer, more accessible plant-based therapeutics. However, further research is necessary to fully elucidate the underlying mechanisms and specific roles of the individual phytochemicals in this process.
Acknowledgments
This publication charge is funded by Unpad through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science and Technology and managed under the EQUITY Program (Contract No. 4303/ B3/DT.03.08/2025 and 3927/UN6. RKT/HK.07.00/2025)
Disclosure
The authors declare no potential conflicts of interest regarding the research, authorship, or publication of this manuscript.
References
1. Siegel RL, Kratzer TB, Giaquinto AN, Sung H, Jemal A. Cancer statistics, 2025. CA Cancer J Clin. 2025;75(1):10–15. doi:10.3322/caac.21871
2. Francies FZ, Hull R, Khanyile R, Dlamini Z. Breast cancer in low-middle income countries: abnormality in splicing and lack of targeted treatment options. Am J Cancer Res. 2020;10(5):1568–1591.
3. Wilson J, Sule AA. Disparity in early detection of breast cancer. 2020.
4. Zonderman AB, Ejiogu N, Norbeck J, Evans MK. The influence of health disparities on targeting cancer prevention efforts. Am J Prev Med. 2014;46(3):S87–S97. doi:10.1016/j.amepre.2013.10.026
5. Situmorang PC, Ilyas S, Nugraha SE, Syahputra RA, Nik Abd Rahman NMA. Prospects of compounds of herbal plants as anticancer agents: a comprehensive review from molecular pathways. Front Pharmacol. 2024;15:1387866. doi:10.3389/fphar.2024.1387866
6. Hashim GM, Shahgolzari M, Hefferon K, Yavari A, Venkataraman S. Plant-derived anti-cancer therapeutics and biopharmaceuticals. Bioengineering. 2024;12(1):7. doi:10.3390/bioengineering12010007
7. Chandra S, Gahlot M, Choudhary AN, et al. Scientific evidences of anticancer potential of medicinal plants. Food Chem Adv. 2023;2:100239. doi:10.1016/j.focha.2023.100239
8. Newman DJ, Cragg GM. Natural products as sources of new drugs from 1981 to 2014. J Nat Prod. 2016;79(3):629–661. doi:10.1021/acs.jnatprod.5b01055
9. Wani MC, Taylor HL, Wall ME, Coggon P, McPhail AT. Plant antitumor agents. VI. Isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J Am Chem Soc. 1971;93(9):2325–2327. doi:10.1021/ja00738a045
10. Xiao H, Wang H, Zhang X, et al. Structural evidence for cooperative microtubule stabilization by taxol and the endogenous dynamics regulator MAP4. ACS Chem Biol. 2012;7(4):744–752. doi:10.1021/cb200403x
11. Yismairai E, Hemelda NM, Yasman, Handayani W. Antioxidant activity of extract of Mistletoe, Dendrophthoe pentandra (L.) Miq. lived in three different host plants, collected from Kampus UI, Depok. AIP Conf Proc. 2019;2168:020100. doi:10.1063/1.5132527
12. Artanti N, Firmansyah T, Darmawan A, Firmansyah H, Darmawan A. Bioactivities evaluation of indonesian mistletoes (Dendrophthoe pentandra (L.) Miq.) leaves extracts. J Appl Pharm Sci. 2012;2(1):24–27.
13. Teodoro GS, van den Berg E, Arruda R. Metapopulation dynamics of the mistletoe and its host in savanna areas with different fire occurrence. PLoS One. 2013;8(6):e65836. doi:10.1371/journal.pone.0065836
14. Tizhe TD, Alonge SO, Aliyu RE. Mistletoe presence on five tree species of Samaru area, Nigeria. Afr J Plant Sci. 2016;10(1):16–22. doi:10.5897/AJPS2015.1335
15. Queijeiro‐Bolaños ME, González EJ, Martorell C, Cano‐Santana Z. Competition and facilitation determine dwarf mistletoe infection dynamics. J Ecol. 2017;105(3):775–785. doi:10.1111/1365-2745.12699
16. Muche M, Muasya AM, Tsegay BA. Biology and resource acquisition of mistletoes, and the defense responses of host plants. Ecol Process. 2022;11(1):24. doi:10.1186/s13717-021-00355-9
17. Kristiningrum N, Ridlo M, Pratoko DK. Phytochemical screening and determination of total phenolic content of Dendophthoe pentandra L. leaves ethanolic extract on Mango host. Ann Trop Med Public Health. 2020;23(03):23–32. doi:10.36295/ASRO.2020.2334
18. Endharti AT, Permana S. Extract from Mango mistletoes Dendrophthoe pentandra ameliorates TNBS-induced colitis by regulating CD4+ T cells in mesenteric lymph nodes. BMC Complement Altern Med. 2017;17(1):468. doi:10.1186/s12906-017-1973-z
19. Zainuddin N, Zakaria Y, Sul’ain MD. Dendrophthoe pentandra Induced Apoptosis and Cell Cycle Arrest at G1/S in Human Breast Adenocarcinoma Cells, MCF-7 via Up- Regulation of p53. J Appl Pharm Sci. 2018;8(9):130–141. doi:10.7324/JAPS.2018.8919
20. Yee LS, Fatihah N, Fauzi M, et al. Study of Dendrophthoe pentandra ethyl acetate extract as potential anticancer candidate on safety and toxicity aspects. J Anal Pharm Res. 2017;6(1):00167. doi:10.15406/japlr.2017.06.00167
21. Page MJ, McKenzie JE, Bossuyt PM, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021;372:n71. doi:10.1136/bmj.n71
22. Czarnecka-Czapczyńska M, Aebisher D, Oleś P, et al. The role of photodynamic therapy in breast cancer – a review of in vitro research. Biomed Pharmacother. 2021;144:112342. doi:10.1016/j.biopha.2021.112342
23. Kong D, Wang L, Niu Y, et al. Dendrophthoe falcata (L.f.) Ettingsh. and Dendrophthoe pentandra (L.) Miq.: a review of traditional medical uses, phytochemistry, pharmacology, toxicity, and applications. Front Pharmacol. 2023;14:1096379. doi:10.3389/fphar.2023.1096379
24. Haryanta D, Susilo A, Kusuma WH. Effect of Mango’s mistletoe (Dendrophthoe pentandra (L.) miq) leaf extract on the biology of Spodoptera litura F. Ecol EnvironConserv. 2020;26(2):2020–2471.
25. Solikin S. Population dynamics of mistletoes species on Cassia fistula in Purwodadi Botanic Garden, Indonesia. Biodiversitas. 2021;22(4). doi:10.13057/biodiv/d220404
26. Ishiwu CN, Obiegbuna JE, Aniagolu NM. Evaluation of chemical properties of mistletoe leaves from three different trees (Avocado, African Oil Bean and Kola). Niger Food J. 2013;31(2):1–7. doi:10.1016/S0189-7241(15)30070-9
27. Ajithkumar TG, Thomas S, Mathew L. Influence of hosts on the production of bioactive compounds in the hemiparasitic plant Helicanthes elasticus. Environ Exp Biol. 2021;19(3):161–171. doi:10.22364/eeb.19.16
28. Awang MA, Nik Mat Daud NNN, Mohd Ismail NI, Abdullah FI, Benjamin MAZ. A review of Dendrophthoe pentandra (Mistletoe): phytomorphology, extraction techniques, phytochemicals, and biological activities. Processes. 2023;11(8):2348. doi:10.3390/pr11082348
29. Yong JWH, Wei JW, Khew JYT, Rong SC, San WW. A Guide to The: Common Epiphytes and Mistletoes of Singapore.
30. Adiansyah NP, Islami PD. Variation and phenetic relationships of Dendrophthoe pentandra (L.) Miq. from various host trees based on morphological characters. Biogenesis. 2023;11(1):48–58.
31. Nisa RN, Nasrullah N. Evaluation of roadside green belt trees damaged by mistletoes parasite plant in Medan Merdeka Road, Central Jakarta, Indonesia. IOP Conf Ser Earth Environ Sci. 2018;203(1):012031. doi:10.1088/1755-1315/203/1/012031
32. Jayanti ED. Uji Aktivitas Antibakteri Ekstrak Etanol Dan Fraksi Daun Benalu Mangga Gadung (Dendrophthoe pentandra (L.) Miq.) Terhadap Staphylococcus aureus ATCC 6538 Dan Escherichia Coli ATCC 25922. 2018.
33. Jabbour F, Espinosa F, Dejonghe Q, Le Péchon T. Development and Evolution of Unisexual Flowers: a Review. Plants. 2022;11(2):155. doi:10.3390/plants11020155
34. Pramestya CD, Yasman Y, Hemelda NM, Handayani W. Organo-specific distribution of untargeted metabolomics of mistletoe’s extract, Dendrophthoe pentandra (L.) Miq. from FKM UI, Kampus UI Depok. AIP Conf Proc. 2019;2168:020089. doi:10.1063/1.5132516
35. Hasan M, Ali MT, Khan R, et al. Hepatoprotective, antihyperglycemic and antidiabetic effects of Dendrophthoe pentandra leaf extract in rats. Clin Phytosci. 2018;4(1):16. doi:10.1186/s40816-018-0076-9
36. Bhagat VC, Kondawar MS. Antitubercular potential of Dendrophthoe falcate (L.) and Tridax procumbens (L.) plants extracts against H37Rv stain of Mycobacteria tuberculosis. Int J Pharm Sci Res. 2019;10(1):251–259.
37. Reddy PS, Sowmya B, Sravani N, Sandhya P, Mohan CK. Evaluation of diuretic potential of petroleum ether extract of Dendrophthoe falcata leaves in wistar rats. Asian J Pharm Pharmacol. 2019;5(6):1086–1090. doi:10.31024/ajpp.2019.5.6.2
38. Nigam YP. The bornean mistletoes as versatile parasites-A systematic review. Syst Rev Pharm. 2022;13(1):42–47. doi:10.33545/27067009.2022.v4.i1a.33
39. Elsyana V, Bintang M, Priosoeryanto BP. Cytotoxicity and antiproliferative activity assay of clove mistletoe (Dendrophthoe pentandra (L.) Miq.) leaves extracts. Adv Pharmacol Sci. 2016;2016:1–6. doi:10.1155/2016/3242698
40. Alharits L, Handayani W, Yasman, Hemelda NM. Phytochemical analysis and antioxidant activity of leaves and flowers extracts of mistletoe (Dendrophthoe pentandra (L.) Miq.), collected from UI Campus, Depok. AIP Conf Proc. 2019;2168:020101. doi:10.1063/1.5132528
41. Kwanda N, Noikotr K, Sudmoon R, Tanee T, Chaveerach A. Medicinal parasitic plants on diverse hosts with their usages and barcodes. J Nat Med. 2013;67(3):438–445. doi:10.1007/s11418-012-0695-2
42. Hardiyanti R, Marpaung L, Adnyana IK, Simanjuntak P. Antioxidant and antibacterial activities of various extracts of Duku’s mistletoe leaf (Dendrophthoe pentandra (L.) Miq) collected from Medan, Indonesia. Asian J Pharm Clin Res. 2018;11(12):526. doi:10.22159/ajpcr.2018.v11i12.29725
43. Widowati W, Mozef T, Risdian C, Ratnawati H, Tjahjani S, Sandra F. The comparison of antioxidative and proliferation inhibitor properties of Piper betle L. Catharanthus roseus [L] G. Don, Dendrophtoe petandra L. Curcuma mangga Val. Extracts on T47D cancer cell line. Int Res J Biochem Bioinform. 2011;1:22–28.
44. Artanti N, Ma’arifa Y, Hanafi M. Isolation and identification of active antioxidant compound from star fruit (Averrhoa carambola) mistletoe (Dendrophthoe pentandra (L.) Miq.) ethanol extract. J Appl Sci. 2006;6(8):1659–1663. doi:10.3923/jas.2006.1659.1663
45. Sembiring HB, Marpaung L, Basyuni M, Simanjuntak P. Glycosylated flavonoid compound from cocoa parasite leaves (Dendrophthoe pentandra (L.) Mig.). Rasayan J Chem. 2022;15(01):700–704. doi:10.31788/RJC.2022.1516382
46. Mochamad L, Malarvili S, Jasmine K, Lim V. In vitro analysis of quercetin-like compounds from mistletoe Dendrophthoe pentandra (L.) Miq as a potential antiviral agent for Newcastle disease. F1000Res. 2024;12:1214. doi:10.12688/f1000research.133489.8
47. Kristiningrum N, Wulandari L, Zuhriyah A. Phytochemical screening, total phenolic content, and antioxidant activity of water, ethyl acetate, and n-hexane fractions from mistletoe Moringa oleifera Lam. (Dendrophthoe pentandra (L.) Miq.). Asian J Pharm Clin Res. 2018;11(10):104. doi:10.22159/ajpcr.2018.v11i10.25271
48. Bunchalee P, Monthakantirat O, Luecha P, Srichai T, Maneechai S. GC-MS analysis and antioxidant potential of the stem and leaf extracts of Dendrophthoe pentandra (L.) Miq. Hemiparasite on Shorea roxburghii G. Don and Cassia fistula L. Trop J Nat Prod Res. 2022;6(10):1638–1643. doi:10.26538/tjnpr/v6i10.13
49. Lazuardi M, Anjani QK, Budiatin AS, Restiadi TI. Efficacy of quercetin-like compounds from the mistletoe plant of Dendrophthoe pentandra L. Miq, as oral random blood sugar lowering treatment in diabetic rats. Vet Q. 2024;44(1):1–14. doi:10.1080/01652176.2024.2372090
50. Dewi RT, Ekapratiwi Y, Sundowo A, Ariani N, Yolanda T, Filaila E. Bioconversion of quercetin glucosides from Dendrophthoe pentandra leaf using Aspergillus acueletus LS04-3. AIP Conf Proc. 2019;2175:020048. doi:10.1063/1.5134612
51. Endharti AT, Wahyuningtyas TE, Handono K, Widjajanto E, Permana S. Dendrophthoe pentandra leaves extract promotes apoptotic effects of doxorubicin in human breast cancer cell via modulation of intracellular calcium and survivin. J Appl Pharm Sci. 2018;8(8):39–43. doi:10.7324/JAPS.2018.8806
52. Permana S, Lukman H, Norahmawati E, et al. The combination therapy of targeting both paclitaxel and Dendrophthoe pentandra leaves extract nanoparticles for improvement breast cancer treatment efficacy by reducing TUBB3 and MAP4 expressions. Acta Biochim Pol. 2021;68(4):679–686. doi:10.18388/abp.2020_5563
53. Khorasanchi Z, Shafiee M, Kermanshahi F, et al. Crocus sativus a natural food coloring and flavoring has potent anti-tumor properties. Phytomedicine. 2018;43:21–27. doi:10.1016/j.phymed.2018.03.041
54. Febriyanti RM, Rafif S, Mikdar N, et al. Anticancer potential of bioactive compounds in Premna serratifolia, Premna odorata, and Premna tomentosa: a review of in vitro evidence. Cancer Manag Res. 2025;17:1029–1045. doi:10.2147/CMAR.S516204
55. Wang Z. Cell cycle progression and synchronization: an overview. Methods Mol Biol. 2022;2579:3–23. doi:10.1007/978-1-0716-2736-5_1
56. Zamani A, Mat Jusoh SA, Al-Jamal HAN, Sul’ain MD, Johan MF. Anti-proliferative effects of Dendrophthoe pentandra methanol extract on BCR/ABL-positive and imatinib-resistant Leukemia cell lines. Asian Pac J Cancer Prev. 2016;17(11):4857–4861. doi:10.22034/APJCP.2016.17.11.4857
57. Li SZ, Qiao SF, Zhang JH, Li K. Quercetin increase the chemosensitivity of breast cancer cells to Doxorubicin via PTEN/Akt pathway. Anticancer Agents Med Chem. 2015;15(9):1185–1189. doi:10.2174/1871520615999150121121708
58. Xia X, He C, Wu A, Zhou J, Wu J. Microtubule-associated protein 4 is a prognostic factor and promotes tumor progression in lung adenocarcinoma. Dis Markers. 2018;2018:1–8. doi:10.1155/2018/8956072
59. Mochamad L, Malarvili S, Jasmine K, Lim V. In vitro analysis of quercetin-like compounds from mistletoe Dendrophthoe pentandra (L.) Miq as a potential antiviral agent for Newcastle disease. F1000Research. 2023;12:1214. doi:10.12688/f1000research.133489.2
60. Lomphithak T, Jaikla P, Sae-Fung A, Sonkaew S, Jitkaew S. Natural flavonoids quercetin and kaempferol targeting G2/M cell cycle-related genes and synergize with smac mimetic LCL-161 to induce necroptosis in cholangiocarcinoma cells. Nutrients. 2023;15(14):3090. doi:10.3390/nu15143090
61. Pavel M, Renna M, Park SJ, et al. Contact inhibition controls cell survival and proliferation via YAP/TAZ-autophagy axis. Nat Commun. 2018;9(1):2961. doi:10.1038/s41467-018-05388-x
62. Zhen ZG, Ren SH, Ji HM, et al. Linarin suppresses glioma through inhibition of NF-κB/p65 and up-regulating p53 expression in vitro and in vivo. Biomed Pharmacother. 2017;95:363–374. doi:10.1016/j.biopha.2017.08.023
63. Ward AB, Mir H, Kapur N, Gales DN, Carriere PP, Singh S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J Surg Oncol. 2018;16(1):108. doi:10.1186/s12957-018-1400-z
64. Zainuddin, Nik Aina Syazana Nik, Sul‘ain, Mohd Dasuki Antiproliferative effect of Dendrophthoe pentandra extract towards human breast adenocarcinoma cells (MCF-7). Jurnal Teknologi. 2015;2:35–39 doi:10.11113/jt.v77.5983.
65. Baig S, Seevasant I, Mohamad J, Mukheem A, Huri HZ, Kamarul T. Potential of apoptotic pathway-targeted cancer therapeutic research: where do we stand? Cell Death Dis. 2016;7(1):e2058–e2058. doi:10.1038/cddis.2015.275
66. Khorsandi L, Orazizadeh M, Niazvand F, Abbaspour MR, Mansouri E, Khodadadi A. Quercetin induces apoptosis and necroptosis in MCF-7 breast cancer cells. Bratisl Med J. 2017;118(02):123–128. doi:10.4149/BLL_2017_025
67. Silva-Pinto PA, de Pontes JTC, Aguilar-Morón B, Canales CSC, Pavan FR, Roque-Borda CA. Phytochemical insights into flavonoids in cancer: mechanisms, therapeutic potential, and the case of quercetin. Heliyon. 2025;11(4):e42682. doi:10.1016/j.heliyon.2025.e42682
68. Zazali KE, Abdullah H, Jamil NIN. Methanol extract of oroxylum indicum leaves induces G 1 /S cell cycle arrest in HeLa Cells via P53-mediated pathway. Int J Med Plant Res. 2013;2(7):225–237.
69. Harmalkar M, Upraity S, Kazi S, Shirsat NV. Tamoxifen-induced cell death of malignant glioma cells is brought about by oxidative-stress-mediated alterations in the expression of BCL2 family members and is enhanced on miR-21 inhibition. J Mol Neurosci. 2015;57(2):197–202. doi:10.1007/s12031-015-0602-x
70. Kallio A, Zheng A, Dahllund J, Heiskanen KM, Härkönen P. Role of mitochondria in tamoxifen-induced rapid death of MCF-7 breast cancer cells. Apoptosis. 2005;10(6):1395–1410. doi:10.1007/s10495-005-2137-z
71. Kesawat MS, Satheesh N, Kherawat BS, et al. Regulation of reactive oxygen species during salt stress in plants and their crosstalk with other signaling molecules-current perspectives and future directions. Plants. 2023;12(4):864. doi:10.3390/plants12040864
72. Taesuk N, Boonpen S, Manee K, et al. Hemiparasitic features and antioxidant activity of Dendrophthoe pentandra and Scurrula atropurpurea on Diospyros rhodocalyx. Biodiversitas. 2025;26(5):2531–2539. doi:10.13057/biodiv/d260548
73. Zhao R, Jiang S, Zhang L, Yu Z. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). IntJ Mol Med. 2019;44(1):3–15. doi:10.3892/ijmm.2019.4188
74. He L, He T, Farrar S, Ji L, Liu T, Ma X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell Physiol Biochem. 2017;44(2):532–553. doi:10.1159/000485089
75. Checa J, Aran JM. Reactive oxygen species: drivers of physiological and pathological processes. J Inflamm Res. 2020;13:1057–1073. doi:10.2147/JIR.S275595
76. Hawkins CL, Davies MJ. Role of myeloperoxidase and oxidant formation in the extracellular environment in inflammation-induced tissue damage. Free Radic Biol Med. 2021;172:633–651. doi:10.1016/j.freeradbiomed.2021.07.007
77. Slika H, Mansour H, Wehbe N, et al. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed Pharmacother. 2022;146:112442. doi:10.1016/j.biopha.2021.112442
78. Jeon JS, Kwon S, Ban K, et al. Regulation of the intracellular ROS level is critical for the antiproliferative effect of quercetin in the hepatocellular carcinoma cell line HepG2. Nutr Cancer. 2019;71(5):861–869. doi:10.1080/01635581.2018.1559929
79. Min K, Ebeler SE. Quercetin inhibits hydrogen peroxide-induced DNA damage and enhances DNA repair in Caco-2 cells. Food Chem Toxicol. 2009;47(11):2716–2722. doi:10.1016/j.fct.2009.07.033
80. Kiesler P, Fuss IJ, Strober W. Experimental models of inflammatory bowel diseases. Cell Mol Gastroenterol Hepatol. 2015;1(2):154–170. doi:10.1016/j.jcmgh.2015.01.006
81. Zou Y, Li WY, Wan Z, et al. Huangqin-Tang Ameliorates TNBS-induced colitis by regulating effector and regulatory CD4 + T cells. Biomed Res Int. 2015;2015:1–13. doi:10.1155/2015/102021
82. Urech K, Schaller G, Jäggy C, et al. Mistletoe lectins and their role in cancer therapy. Curr Pharm Des. 2014;20(10):1415–1422. doi:10.2174/13816128113199990363
83. Kienle GS, Kiene H. Influence of Viscum album L. (European mistletoe) extracts on quality of life in cancer patients: a systematic review of controlled clinical studies. Integr Cancer Ther. 2010;9(2):142–157. doi:10.1177/1534735410369673
84. Melzer J, Iten F, Hostanska K, Saller R. Efficacy and safety of mistletoe preparations (Viscum album) for cancer therapy. Phytomedicine. 2009;16(4):251–257. doi:10.1016/j.phymed.2008.11.004
85. Horneber M, Bueschel G, Huber R, Linde K, Rostock M. Mistletoe therapy in oncology. Cochrane Database Syst Rev. 2008;2008(2):CD003297. doi:10.1002/14651858.CD003297.pub2
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.