")
Back to Journals » Drug Design, Development and Therapy » Volume 9
CXCR4 in breast cancer: oncogenic role and therapeutic targeting
Authors Xu C, Zhao H, Chen H, Yao Q
Received 18 March 2015
Accepted for publication 13 April 2015
Published 28 August 2015 Volume 2015:9 Pages 4953—4964
DOI https://doi.org/10.2147/DDDT.S84932
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Shu-Feng Zhou
Chao Xu,1,* Hong Zhao,1,* Haitao Chen,1 Qinghua Yao2,3
1First Clinical College of Zhejiang Chinese Medical University, 2Department of Integrated Traditional Chinese and Western Medicine, Zhejiang Cancer Hospital, 3Key Laboratory of Integrated Traditional Chinese and Western Medicine, Zhejiang Cancer Hospital, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Abstract: Chemokines are 8–12 kDa peptides that function as chemoattractant cytokines and are involved in cell activation, differentiation, and trafficking. Chemokines bind to specific G-protein-coupled seven-span transmembrane receptors. Chemokines play a fundamental role in the regulation of a variety of cellular, physiological, and developmental processes. Their aberrant expression can lead to a variety of human diseases including cancer. C-X-C chemokine receptor type 4 (CXCR4), also known as fusin or CD184, is an alpha-chemokine receptor specific for stromal-derived-factor-1 (SDF-1 also called CXCL12). CXCR4 belongs to the superfamily of the seven transmembrane domain heterotrimeric G protein-coupled receptors and is functionally expressed on the cell surface of various types of cancer cells. CXCR4 also plays a role in the cell proliferation and migration of these cells. Recently, CXCR4 has been reported to play an important role in cell survival, proliferation, migration, as well as metastasis of several cancers including breast cancer. This review is mainly focused on the current knowledge of the oncogenic role and potential drugs that target CXCR4 in breast cancer. Additionally, CXCR4 proangiogenic molecular mechanisms will be reviewed. Strict biunivocal binding affinity and activation of CXCR4/CXCL12 complex make CXCR4 a unique molecular target for prevention and treatment of breast cancer.
Keywords: breast cancer, CXCR4, drug target, chemokine, angiogenesis
CXCR4 structure, functions, and signaling
Chemokines are a superfamily of small molecule chemoattractive cytokines that regulate many cellular functions.1 The CXC chemokine stromal-derived-factor-1 (SDF-1 or CXCL12) is expressed in a variety of cells, including stromal cells (fibroblasts and endothelial cells).2,3 Chemokine receptors are a family of G protein-coupled cell surface receptors (GPCRs) with seven transmembrane-spanning domains. Chemokines are small, structurally related proteins that play a significant role in leukocyte trafficking and are divided into four groups (CXC, CC, C, and CX3C) based on the position of the first two conserved cysteines.4,5 The chemokine receptor CXCR4 is a 352-amino acid rhodopsin-like GPCR that selectively binds to the CXC chemokine SDF-1 or CXCL12. CXCR4 is the physiological receptor for SDF-1, in addition to CXCR7.6 The SDF-1/CXCR4 axis plays an important role in many physiological processes, including trafficking and homeostasis of immune cells such as T-lymphocytes7 and homing and retention of hematopoietic stem cells within the bone marrow.8 The interaction between SDF-1 and CXCR4 leads to the activation of various intracellular signaling transduction pathways and downstream effectors that mediate cell survival, proliferation, chemotaxis, migration, and adhesion.1 CXCR4 is highly expressed in various types of cancers, including oral cancer,9,10 esophageal cancer,11–13 gastric cancer,14 colon cancer,15 liver cancer,16 pancreatic cancer,17 thyroid cancer,18,19 breast cancer,1 ovarian cancer,20 prostate cancer,21,22 lung cancer,23,24 kidney cancer,25,26 brain cancer,27,28 melanoma,29,30 and leukemia,31,32 and is associated with chemotaxis, invasion, angiogenesis, and cell proliferation contributing tumorigenesis and cancer progression. A potential mechanism of CXCR4’s involvement in tumor dissemination and metastasis is through promoting its transendothelial migration at the primary site.
Regulation of CXCR4 expression
In a typical model, CXCR4 mediates virtually all signaling pathways via its coupled G proteins which triggers a highly conserved DRY (Asp-Arg-Tyr) motif located in the second intracellular loop. This concept has changed after the finding of activation of downstream MAPK (mitogen-activated protein kinase) pathways through β-arrestins in the absence of detectable G protein activities. The latter delivers signals through the C-terminal (CT) tail of GPCRs which is rich in serines and threonines.33 CXCR4 is expressed constitutively by lymphatic tissues, thymus, brain, spleen, stomach, and small intestine.34 With regard to breast tissues, CXCR4 protein has no or low-level expression in normal breast epithelium and is highly expressed in ductal carcinoma, progressively increasing from atypical hyperplasia to carcinoma.35
Transcriptional mechanism
Hypoxia inducible factor 1 alpha (HIF-1α) and HIF-2: CXCR4 promoter has a functional hypoxia response element, so HIF can activate CXCR4 transcription. In normoxia, Von Hippel Lindau (VHL) protein, a negative regulator of HIF, degrades CXCR4 protein through HIF downregulation. In hypoxia, due to lack of VHL protein in a variety of cancer cell lines, HIF-1 and HIF-2 are stabilized and then induce CXCR4 transcription, expression, and, subsequently, CXCR4/CXCL10 pathway activation.36–38
Nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB): Similar to HIF, there is an NF-κB binding site within in the CXCR4 gene promoter. Upon stimulation of some ligands such as hepatocyte growth factor (HGF), NF-κB subunits p65 and p50 bind at −66 to +7 of the CXCR4 promoter, transcriptionally activate CXCR4, and regulate tumor invasion.39–44
Other factors: Vesnarinone-responsive molecule Kruppel-like factor 2 and histone deacetylase 3-interacting protein CREB3 have also been reported to inactivate the transcription of CXCR4 and, therefore, regulate cell migration.45,46 Transcription factor YY1, a 65 kDa DNA-binding protein, which can either repress or activate transcription depending on the context, is phosphorylated by carboxyl-terminal Src kinase homologous kinase (CHK), and activated YY1 represses expression of CXCR4 in breast cancer cells.47,48
Posttranscriptional and translational mechanism
CXCR4-mediated feedback loop of multiple signaling pathways
CXCR4 expression is positively regulated by the developmental signaling pathways Wnt/β-catenin49 (while knockdown of CXCR4 could obviously reduce proliferation and invasion of ovarian cancer cell and inhibit Wnt target genes50–52), SHH-GLI-NANOG53 (while a functional CXCR4 pathway is necessary to maintain the SHH-GLI1-NANOG network and self-renewal properties54), and Notch as well as oncogenic pathways phosphoinositide 3-kinase (PI3K)/AKT,55,56 NF-κB,44,57 and Janus kinase (JAK)/signal transducer and activator of transcription (STAT).58,59 In turn, the activation of the SDF-1/CXCR4 signaling activates MAPKs signaling, which promotes chemotaxis and proliferation,55,60 induces phospholipase C (PLC)/protein kinase C (PKC)-Ca2+ signaling promoting cell migration, and affects PI3K/AKT promoting cell survival.55,56 This indicates a positive feedback loop between CXCR4 and the signaling pathways mediating tumorigenicity of cancer cells (Figure 1). In addition, SDF-1/CXCR4 signaling pathway may transactivate epidermal growth factor receptor (EGFR)/HER2-neu signaling to promote invasive signals and metastatic growth of breast,61 prostate,62 and ovarian cancers.63
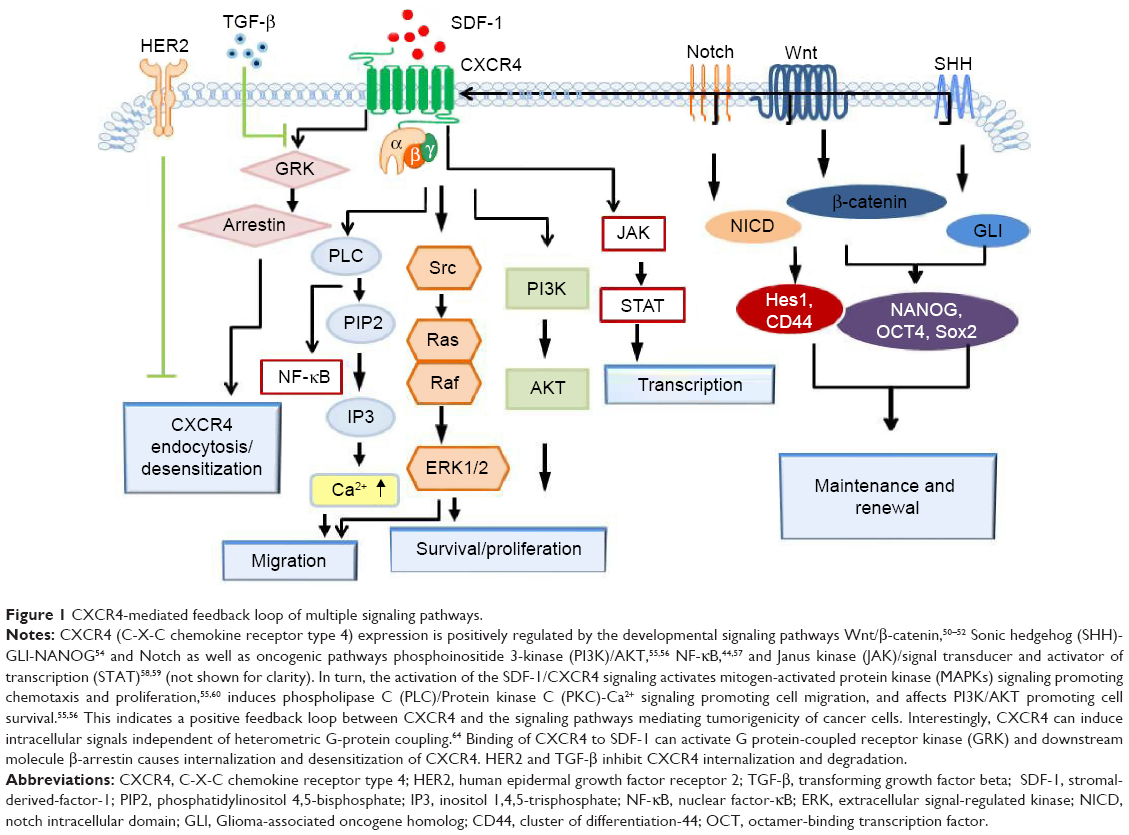 | Figure 1 CXCR4-mediated feedback loop of multiple signaling pathways. |
Interestingly, CXCR4 can induce intracellular signals independent of heterometric G-protein coupling.64 Binding of CXCR4 to SDF-1 can activate G protein-coupled receptor kinase (GRK) and the downstream molecule β-arrestin. β-arrestin can cause internalization and desensitization of CXCR4. However, if β-arrestin is recruited and activated by the CXCR4/CXCR7 heterodimeric complex, the downstream signaling cascades such as PI3K/Akt, extracellular signal-regulated kinase (ERK)1/2, PLC/MAP, p38 MAPK, and stress-activated protein kinase (SAPK)/c-Jun N-terminal kinase (JNK) are enhanced, and thus tumor growth, cell migration, dissemination, and neovascularization are facilitated.65–69
Vascular endothelial growth factor (VEGF) level is elevated in both breast cancer cell lines and clinical breast cancer samples. VEGF potentiates the metastasis of breast cancer cells in two ways:55 in a paracrine manner, VEGF increases vascular permeability by mediating actin rearrangement and affecting gap junction, thus enhancing angiogenesis and tumor cell extravasation;70 in an autorcrine manner, VEGF promotes breast cancer cell migration and metastasis by inducing CXCR4 expression.
Levels of CXCR4 protein are also regulated via posttranscriptional mechanisms. Prolonged exposure of cells to SDF-1 under physiological situations desensitizes them to SDF-1 through endocytosis of CXCR4 receptor. The receptor is ubiquitinated by the E3 ligase atrophin-1-interacting protein 4 (AIP4), which targets the CXCR4 receptor to the endosome and degrades it in lysosomes.71 However, Luker and Luker35 and Basu and Broxmeyer72 report that the hematopoietic stem cells and myeloid progenitor cells retain responsiveness to SDF-1 even in the presence of sustained exposure to SDF-1. EGFR seems to be important for the CXCR4 internalization. The oncogene Her2 not only increases the translation of CXCR4 mRNA by 2.5-fold over controls, but it also blocks the ubiquitination and the degradation of CXCR4 after the binding of CXCL12 in breast cancer cells.35,73 Similar to Her2, another EGFR variant, EGFvIII affects CXCR4 expression at multiple levels. For instance, it increases CXCR4 expression transcriptionally through the upregulation of HIF-1α. It can also enhance CXCR4 expression posttranslationally by activating p38 MAPK and inactivating AIP4 and β-arrestin1/2 to reduce CXCR4 internalization, cellular trafficking, and lysosomal degradation.74
CXCR4 in primary breast cancer
The role of CXCR4 in promoting breast cancer cell proliferation
Due to the similarity between the migration and invasion of cancer cells and chemotaxis of immune cells, initial studies of CXCR4 focused on its functions in metastatic breast cancer. In addition to metastasis, the SDF-1/CXCR4 signaling pathway plays a role in growth and proliferation of primary breast cancer. The first evidence for this is that Michigan Cancer Foundation-7 (MCF-7) breast cancer cell line with low expression of CXCR4 formed much smaller tumor mass at inoculated sites when the cells were inoculated subcutaneously in the SCID (severe combined immunodeficiency) mice compared to those generated by MD Anderson-metastatic breast (MDA-MB)-231 cell line with high expression of CXCR4.1 In addition, tumors cells with downregulated expression of CXCR4 by RNA interference and stable RNA interference grew more slowly than cells with upregulated or normal levels of CXCR4 when the cells were implanted into syngeneic, immunocompetent mice.75,76
The mechanism by which CXCR4 promotes breast cancer growth
The molecular mechanisms by which CXCR4 facilitates the growth of primary breast cancers are widely investigated, and can be summarized as follows:
- Angiogenesis: it has been found that disruption of the interaction between SDF-1 and CXCR4 with neutralizing antibodies dampens the growth of human breast cancer xenografts in vivo, partly through reducing numbers of blood vessels.77 Stromal cells present in primary breast cancers are the important sources of SDF-1. SDF-1 produced in the tumor microenvironment recruits circulating endothelial progenitors to sites of primary breast tumors and generates more microvessels thus increasing density of microvessels in cancer.78
- Typical signaling pathways related to cell proliferation: a number of downstream effectors of CXCR4 (PI3K/Akt, Src/ERK1-2, NF-κB, and STAT3) which are well known for their growth-promoting ability have been identified to contribute to the growth of primary breast tumors (Figure 1). The cross talk between CXCR4 and other stem cell-related pathways such as Notch, Wnt, and SHH are also likely to be involved in breast cancer cell proliferation (Figure 1).
- Recruitment of immune cells: in addition to endothelial progenitors, dendritic cells are also recruited to tumor sites through SDF-1. Dendritic cells suppress antitumor immunity by inactivation of cytotoxic T-cells, promoting tumor growth.79 The schematic effects of CXCR4 on primary and metastatic breast cancer are shown in Figure 2.
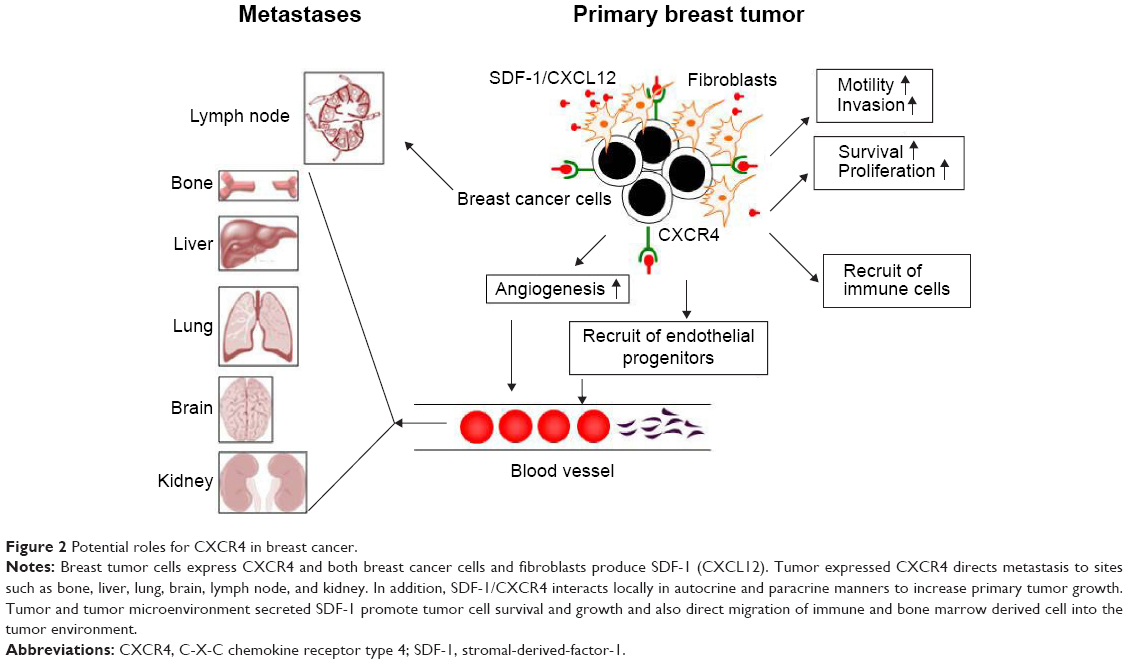 | Figure 2 Potential roles for CXCR4 in breast cancer. |
Recently, some studies have successfully shown the relationship between hormone-dependent breast cancer cell proliferation and CXCR4 signaling. Stimulation of estrogen E2 on the two receptors of CXCL12, CXCR4, and CXCR7 promotes E2-mediated breast cancer growth. Estrogen E2 was found to stimulate CXCR4 transcription via estrogen receptor (ER)-binding sites at the CXCR4 promoter enhancing E2-mediated cellular growth.80 In contrast, another receptor for the chemokine SDF-1 (CXCL12), CXCR7 expression was repressed by estrogen through inhibition of the recruitment of transcription factor NF-κB at the CXCR7 promoter. CXCR7 overexpression decreases the growth effect of E2, although it can increase the basal MCF-7 cell growth rate.80,81 In addition, a nuclear receptor COUP-TF is overexpressed in MCF-7 breast cancer cells and inhibits the expression of the chemokine CXCL12, while enhancing the expression of its receptor CXCR4. Through the activation of epithelial growth factor (EGF), COUP-TFI increases the invasive potential of ER-positive breast cancer.82
CXCR4 in metastatic breast cancer
Metastasis of cancer cells to distant locations needs several sequential steps, such as intravasation from the primary tumor sites into blood vessels, survival in the circulation, migration to secondary organs, adhesion, and proliferation of cancer cells in targeting organs and tissues.83 CXCR4 promotes breast cancer metastasis to organs (bone, liver, and lung) where its ligand, SDF-1, is generated in large quantity. The cytokine SDF-1 is released in large amounts by lung, bone, and liver, which are the sites commonly affected by metastatic breast cancer. The interaction between SDF-1 and CXCR4 makes breast cancer cells move out of the circulation and traffic into organs with high amounts of chemokines, and thus forming metastatic tumors.2 SCID mice inoculated with CXCR4-low-expressing MCF-7 did not demonstrate organ metastasis, while SCID mice inoculated with CXR4-high-expressing MDA-MB-231 cells exhibited spontaneous metastasis in lung.1 Moreover, the metastatic lung cancer cells express more CXCR4 than primary lung cancer cells.44
Bone
Increased CXCR4 expression is present in breast cancer cells metastasized to bone.84 Some bone cells such as osteoblasts produce SDF-1 (CXCL12), and this is the reason that breast cancer cells can easily metastasize to bone. The same mechanisms apply to liver and lungs, the other two target sites of breast cancer metastasis where large amount of SDF-1 are produced. Binding of SDF-1 (CXCL12) to CXCR4 activates the tyrosine kinase Src, and downstream effector AKT enhances the survival of breast cancer cells residing in the bone.85
Brain
Tumor cells have the ability of breaching the blood–brain barrier, a tightly sealed area around the brain with high electrical resistance produce by tight junctions, to generate a metastatic cerebral tumor.55 An in vitro study demonstrated that SDF-1 introduces the breast cancer cells into the brain via increased vascular permeability. Breast cancer metastasizing to the brain involves several signaling pathways, particularly those involving growth factors and chemokines. Several cofactors of CXCR4 are involved in breast cancer cell invasion. The CXCR4/SDF-1-mediated activation of PI3K/Akt and phosphorylation of focal adhesion kinase (FAK) and the frokhead transcription factor like 1 (FKHRL1) were necessary for breast cancer cell migration through the blood–brain barrier.70 The mutual interplay between the CXCR4-VEGF,70 CXCR4-NFκB,84 and CXCR4-EGFR-ERK1/244,86 pathways have been observed to promote breast cancer cell migration and metastasis.
Lung
As stated previously, CXCR4 signaling activated by its ligand SDF-1 causes chemotaxis and migration of breast cancer cells. Breast cancer cells that metastasized to the lungs exhibited high levels of CXCR4 expression compared to their parental cells, supporting the role of CXCR4 in organ directed-metastasis and invasion of breast cancer. Neutralizing antibodies to CXCR4 remarkably inhibited metastases of breast cancers to lymph nodes and lung.35
Lymph node, liver, and kidney
Local lymph node involvement in breast cancer patients has been widely investigated; a recent meta-analysis showed that breast cancer with CXCR4 expression were associated with lymph node status (RR =0.99, 95% CI: 1.01–1.43). Systemic studies of the role of CXCR4 on liver and kidney metastasis of breast cancer are not available at present.
CXCR4 and prognosis of breast cancer patients
A number of studies have assessed the effects of CXCR4 expression on breast cancer prognosis. Kato et al87 systematically analyzed the expression levels of CXCR4 in 79 surgically resected invasive ductal carcinomas and the relation between the immunohistochemical staining pattern and clinicopathological outcomes. They found that the patients with high levels of CXCR4 had more extensive metastasis to lymph nodes compared to those with low levels of CXCR4.87 Recently, more powerful meta-analyses were performed by two different groups quantitatively analyzing the impact of CXCR4 expression on breast cancer prognosis. Zhang et al88 included 13 eligible studies consisting of 3,865 participants from USA, Canada, China, Japan, Korea, and Taiwan. Their meta-analysis showed that CXCR4 overexpression was associated with lymph node infiltration (risk ratio RR =1.20, 95% CI [confidence interval]: 1.01–1.43, P<0.001) and distant metastasis (RR =1.52, 95% CI: 1.17–1.98). CXCR4 overexpression significantly reduced disease free survival (DFS) (RR =0.7, 95% CI: 0.70–0.86) and overall survival (OS), with the risk ratio RR being 0.77 (95% CI: 0.70–0.86).88
Xu et al6 investigated 15 published studies that included 3,104 patients which are from USA, Canada, Germany, France, China, and Japan. Their results are consistent with Zhang et al’s88 results and showed that the OS and DFS were found to be significantly reduced in breast cancer patients with high levels of CXCCR4 expression relative to those with low levels of CXCR4 expression, with the hazard ratio being 1.65 (95% CI: 1.34–2.03, P<0.00001) and 1.94 (95% CI: 1.42–2.65, P<0.00001), respectively. Stratified analysis indicated that high expression of CXCR4 in whole cells, cytoplasm, and nucleus predicts reduced OS, while only elevated levels of CXCR4 in whole cells and cytoplasm predicts a poor DFS.6 Therefore, CXCR4 is an efficient prognostic factor for breast cancer.
CXCR4 as a potential therapeutic target for breast cancer
Beyond potential applications for diagnosis of breast cancer, a variety of studies have shown that peptides or small molecule inhibitors of CXCR4 attenuate the growth of breast cancer in vivo and in vitro. Several novel CXCR4 antagonists have shown promising results in both in vitro and in vivo anticancer activity in several tumor cell types (using animal tumor models), including those derived from breast cancer. We summarize the effects of various CXCR4 inhibitors on breast cancer in Table 1.
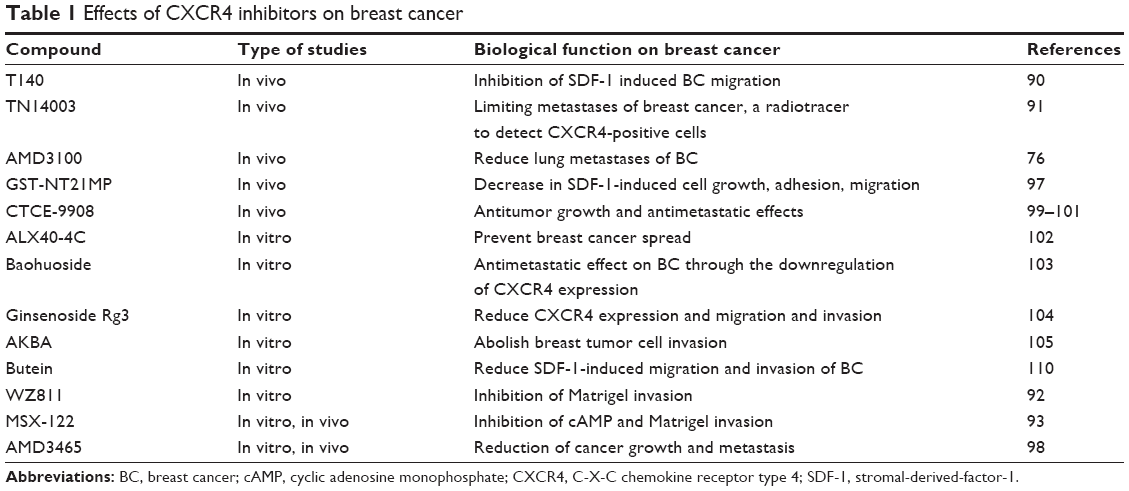 | Table 1 Effects of CXCR4 inhibitors on breast cancer |
T140 and its analogs as CXCR4 antagonists
Fourteen-mer peptides, T140 and its analogs, were originally developed as specific CXCR4 antagonists, as HIV-entry inhibitors, and as antirheumatoid arthritis agents.89 CXCR4 antagonist T140 and its analogs actually block the CXCR4 receptor binding to its ligand SDF-1.89 In 2003, Tamamura et al90 first reported that T140 analogs, peptidic CXCR4 antagonists composed of 14 amino acid residues which were previously developed as anti-HIV agents, effectively inhibited SDF-1-induced migration, proliferation, and metastasis in human breast cancer MDA-MB-231. In the following year, Liang et al91 improved the compound T140 by substituting basics residue of the COOH-terminal with nonbasic polar amino acids to compromise the total positive charges of the molecule to generate a synthetic antagonist 14-mer peptide compound, TN14003. TN14003 (BKT140) is far less cytotoxic and more stable in serum relative to T140, and TN14003 not only proved to be effective in reducing metastases of breast cancer by inhibiting migration, but also is useful as a diagnostic tool for detecting CXCR4 receptor-positive tumor cells in cultured cells, living animals, and probably also patients, using positron emission tomography.35,91 However, lack of oral availability prompted Zhan et al92 to further look for alternatives that possess better pharmacologic profiles. Using TN14003 as their benchmark, a first nitrogen atom substitution of the terminal aromatic rings (ie, pyridyl instead of phenyl) was designated as WZ811,92 while a second nitrogen atom substitution in the terminal aromatic rings (ie, pyrimidyl) was designated as MSX-122. Compared to WZ811, MSX-122, which was prepared in a single chemical step using a reductive amination reaction, further improved the pharmacokinetic profile.93 Both WZ811 and MSX-122 block lung metastasis of breast cancer. However, the advantage of MSX-122 is its ability to intervene the Gai-signaling pathway (cAMP modulation), but not the Gq-pathway (calcium flux).94 Consequently, MSX-122 blocks chemotaxis and the homing of the CXCR4-positive cells (including cancer cells) to distant organ sites when enriched with CXCL12 in the stroma without disturbing the retention of hematopoietic progenitor cells in the bone marrow. Therefore, MSX-122 could have a unique therapeutic application for cancer metastasis or inflammation.
Cyclic pentapeptides, such as FC131 [cyclo(D-Tyr-Arg-Arg-L-3-(2-naphthyl)alanine-Gly)], were also previously developed as CXCR4 antagonists based on pharmacophores of T140.89,95,96 Binding models for FC131 in CXCR4 have previously been suggested based on molecular docking guided by structure–activity relationship data; however, none of these have been verified by in vitro experiments.89
Non-T140-related CXCR4 antagonists
To date, several non-T140-related CXCR4 antagonists have been reported. Inhibition of CXCR4 with ABD3100 (plerixafor [Mozobil, Sanofi, Bridgewater, NJ, USA]), a chemokine receptor antagonist and a small molecule inhibitor of CXCR4 receptor has significantly limited lung metastases from orthotopically transplanted breast cancer cells.76 Yang et al’s97 data also suggest that GST-NT21MP could be a potential anticancer agent for the treatment of breast cancer. A highly purified recombination polypeptide (GST-NT21MP), which is a synthetic 21-mer peptide antagonist of CXCR4 (NT21MP) derived from the viral macrophage inflammatory protein II, and was found to block the CXCR4 pathway, thus decreasing SDF-1-induced cell growth, adhesion, and migration capacities in breast cancer cells. Furthermore, GST-NT21MP-mediated antitumor activity was found to be associated with reduced phosphorylated Src, AKT, FAK, and ERK1/2 as well as decreased Bcl-2. A small molecule antagonist of CXCR4, AMD3465, can inhibit breast cancer growth and metastasis and demonstrate the biologically relevant modulation of oncogenic signaling and tumor microenvironment.98 Ling et al reported that AMD3465 triggers a reduction in breast cancer cell invasiveness both in vitro, and in vivo in murine syngeneic immunocompetent breast cancer models, and they found that AMD3465 inhibited breast tumor initiation and metastases to the lung and liver. Not only negatively regulating breast tumor cells, AMD3465 also reduces the infiltration of immune cells at the metastatic sites as well as at the spleen. CTCE-9908 (Chemokine Therapeutics Corp, Vancouver, BC, Canada) is a peptide analog of SDF-1 consisting of a dimer of the first eight amino acids of SDF-1, which competitively binds to CXCR4 and serves as a competitive inhibitor to SDF-1. To test the effects of CTCE-9908 in breast cancer, a transgenic mouse mammary tumor virus-driven Polyoma Middle T Antigen (PyMT) model has been widely used based on the overexpression of HER2/neu and stepwise progression from hyperplasia to preinvasive, invasive, and distant metastasis in this model. Increasing doses of CTCE-9908 alone slowed the rate of tumor growth. Once combined with docetaxel or DC101 (an anti-VEGFR2 monoclonal antibody) or an antiangiogenic agent, the effects were induced much more than that observed with docetaxel alone. The markedly increased antitumor and antimetastatic effects of CTCE-9908, which indicated that it is a potentially new effective combinatorial therapeutic strategies in the treatment of breast cancer, is involved in targeting the SDF-1/CXCR4 ligand/receptor pair.99,100 Another study to evaluate the efficacy of CTCE-9908 in a xenograft mouse model did not show inhibition of primary tumor growth with or without paclitaxel (a chemotherapeutic) relative to control, but it still inhibited organ-specific metastasis to leg.101 Another CXCR4-neutralizing peptide ALX40-4C (N-alpha-acetyl-nona-D-arginine amide; American Petptide Company, Sunnyvale, CA, USA) blocks the binding of SDF-1 to CXCR4 without influencing the binding of other chemokines to their respective GPCRs.17,18 To establish a role for CXCR4 in invasion, the effect of ALX40-4C was tested in MDA-MB-231.102 This peptide significantly inhibited Matrigel invasion by 75% but did not reduce cell viability, which suggest a distinct contribution of CXCR4 to the invasion but not the survival of breast carcinoma cells.
Natural products inhibiting CXCR4
A few of the new CXCR4 inhibitors from natural product have also been reported. Kim and Park103 investigated baohuoside I, a component of Epimedium koreanum, as a regulator of CXCR4 expression and function in cervical cancer and breast cancer cells. They observed that baohuoside I downregulated CXCR4 expression in a dose- and time-dependent manner. The decrease in the level of CXCR4 expression caused by baohuoside I correlated with inhibition of the CXCL12-induced invasion of both cervical and breast cancer cells, indicating that it plays a role in the suppression of cancer metastasis.103 Ginsenoside Rg3 is a component extracted from puffed ginseng (Panax ginseng C.A. Meyer). In Chen et al’s104 study, at a dosage without obvious cytotoxicity, Rg3 treatment reduces CXCR4 expression, decreases the ability of migration and invasion of breast cancer MDA-MB-231cells induced by CXCL12 suggesting that Rg3 is a new CXCR4 inhibitor from a natural product. Acetyl-11-keto-b-boswellic acid (AKBA) is a derivative of boswellic acid, which is the main component of a gum resin from Boswellia serrata. AKBA has been used traditionally to treat a number of inflammatory diseases, including osteoarthritis, chronic colitis, ulcerative colitis, Crohn disease, and bronchial asthma. AKBA abolished breast tumor cell invasion, and this effect correlated to the downregulation of both the CXCR4 mRNA and CXCR4 protein.105 Butein (3, 4, 20, 40-tetrahydroxychalcone), a novel regulator of CXCR4 expression and function, which is derived from numerous plants, including the stembark of cashews (Semecarpus anacardium) and the heartwood of Dalbergia odorifera, has substantial antitumor activities, as indicated by inhibition of proliferation of a wide variety of tumor cells,106,107 suppression of phorbol ester-induced skin tumor formation,108 and inhibition of carrageenan-induced rat paw edema.109 The decrease in CXCR4 expression induced by butein was not cell type-specific, and the downregulation of CXCR4 was due to transcriptional regulation. Suppression of CXCR4 expression by butein correlated to the inhibition of CXCL12-induced migration and invasion of breast cancer cells, suggesting that butein is a novel inhibitor of CXCR4 expression and thus has a potential in suppressing metastasis of cancer.110
Recombinant chimeric protein
CXCL12/54R
In a transgenic mouse with mutant CXCL12, obtained by deleting the 55th to 67th residues of its COOH terminus (CXCL12/54R), SDF-1 was unable to bind to CXCR4. CXCR4 was quickly internalized, subsequently downstream signals mediated by CXCR4 were inactivated, resulting in the inhibition of tumor cell migration.111 However, the inhibitory function of CXCL12/54R tends to be temporary and reversible, and TAT/54R/KDEL can produce a longer or more permanent inhibition of CXCR4 expression on the cellular surface.
TAT/54R/KDEL
A novel recombinant chimeric protein, TAT/54R/KDEL was developed, in which TAT and KDEL were linked to the NH2-terminal and COOH-terminal of CXCL12/54R, respectively. TAT, which is from HIV-1 TAT (47–57, YGRKKRRQRRR), is able to permeate the plasma membrane of cells either alone or fused with full-length proteins or peptides112,113 can deliver proteins ranging from 10 to 120 kDa into the cells without any damage to cells.114–116 Four-peptide KDEL or DDEL is a site-specific signal which detained the soluble endoplasmic reticulum-resident proteins in ER for degradation.117,118 The systemic treatment of TAT/54R/KDEL could impair lung metastasis of a highly metastatic, triple-negative mammary cancer cell line, 4T1, with decrease of CXCR4 on their membrane, suggesting that the phenotypic knockout strategy of CXCR4 using a novel recombinant protein TAT/54R/KDEL could potentially be a possible approach for inhibiting relative tumor metastasis mediated by CXCR4/CXCL12 interaction. Taken together, CXCR4 may be an effective therapeutic in preventing breast cancer spread.
In addition to breast cancer, some studies have successfully demonstrated that blockade of CXCR4 or SDF-1/CXCR4 interaction by small molecule inhibitor of CXCR4 suppresses prostate cancer (eg, CTCE-9908)119 and lung cancer (eg, TN14003).23 At present, clinical trials involving CXCR4 inhibition are tested in hematological malignancies.24 Administration of a CXCR4 antagonist would probably not be used alone; combinations with established chemotherapy would be likely. Clinical trials of CXCR4 antagonists in breast cancer patients are rarely available; the likely reason might be as a result of intervention failures and high attrition rates of candidate drugs that show success in animal models but fail in human clinical trials.
Conclusion
In the past 10 years, numerous investigations have been conducted on the role of SDF-1/CXCR4 signaling pathway in solid tumors, including breast cancer. The antagonists of CXCR4 could be promising agents for prevention and treatment of breast cancer metastasis. However, we must keep in mind that CXCR4 plays a critical role in embryogenesis, homeostasis, and inflammation in the fetus, especially in the embryonic development of hemopoietic, cardiovascular, and central nervous systems. Therefore, caution should be taken when inhibition of the SDF-1-CXCR4 signaling pathway is applied in human subjects.89 Inhibition of CXCR4 signaling attenuates the immune responses, therefore moderate activation of CXCR4 pathway contributes to depression of inflammation and is beneficial for the cancer patients. However, excessive activation of CXCR4 pathway might dampen the hosts’ immune responses and decrease anticancer ability. In addition, preclinical data have shown that blockade of CXCR4 may increase osteoclastic bone resorption therefore promoting tumor cell growth in bone.120 Caution should be taken when the utility of blockade of the SDF-1/CXCR4 axis is evaluated.
Acknowledgment
This work was supported by Zhejiang Provincial Natural Science Foundation of China (Grant number LQ12H16006) and Zhejiang Provincial Natural Science Foundation of China (LQ12H16006).
Disclosure
The authors have no financial involvement with any organization or entity with a financial interest in the subject matter or materials discussed in the manuscript. The authors report no conflicts of interest in this work.
References
Dewan MZ, Ahmed S, Iwasaki Y, Ohba K, Toi M, Yamamoto N. Stromal cell-derived factor-1 and CXCR4 receptor interaction in tumor growth and metastasis of breast cancer. Biomed Pharmacother. 2006;60:273–276. | ||
Muller A, Homey B, Soto H, et al. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410:50–56. | ||
Salvucci O, Yao L, Villalba S, Sajewicz A, Pittaluga S, Tosato G. Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1. Blood. 2002;99:2703–2711. | ||
Zlotnik A, Yoshie O. Chemokines: a new classification system and their role in immunity. Immunity. 2000;12:121–127. | ||
Murphy PM, Baggiolini M, Charo IF, et al. International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol Rev. 2000;52:145–176. | ||
Xu TP, Shen H, Liu LX, Shu YQ. The impact of chemokine receptor CXCR4 on breast cancer prognosis: a meta-analysis. Cancer Epidemiol. 2013;37:725–731. | ||
Busillo JM, Benovic JL. Regulation of CXCR4 signaling. Biochim Biophys Acta. 2007;1768:952–963. | ||
Kucia M, Reca R, Miekus K, et al. Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4 axis. Stem Cells. 2005;23:879–894. | ||
Ishikawa T, Nakashiro K, Hara S, et al. CXCR4 expression is associated with lymph-node metastasis of oral squamous cell carcinoma. Int J Oncol. 2006;28:61–66. | ||
Onoue T, Uchida D, Begum NM, Tomizuka Y, Yoshida H, Sato M. Epithelial-mesenchymal transition induced by the stromal cell-derived factor-1/CXCR4 system in oral squamous cell carcinoma cells. Int J Oncol. 2006;29:1133–1138. | ||
Kodama J, Hasengaowa, Kusumoto T, et al. Association of CXCR4 and CCR7 chemokine receptor expression and lymph node metastasis in human cervical cancer. Ann Oncol. 2007;18:70–76. | ||
Yang YC, Lee ZY, Wu CC, Chen TC, Chang CL, Chen CP. CXCR4 expression is associated with pelvic lymph node metastasis in cervical adenocarcinoma. Int J Gynecol Cancer. 2007;17:676–686. | ||
Wu J, Wu X, Liang W, Chen C, Zheng L, An H. Clinicopathological and prognostic significance of chemokine receptor CXCR4 overexpression in patients with esophageal cancer: a meta-analysis. Tumour Biol. 2014;35:3709–3715. | ||
Han M, Lv S, Zhang Y, et al. The prognosis and clinicopathology of CXCR4 in gastric cancer patients: a meta-analysis. Tumour Biol. 2014;35:4589–4597. | ||
Lv S, Yang Y, Kwon S, et al. The association of CXCR4 expression with prognosis and clinicopathological indicators in colorectal carcinoma patients: a meta-analysis. Histopathology. 2014;64:701–712. | ||
Schimanski CC, Bahre R, Gockel I, et al. Dissemination of hepatocellular carcinoma is mediated via chemokine receptor CXCR4. Br J Cancer. 2006;95:210–217. | ||
Billadeau DD, Chatterjee S, Bramati P, et al. Characterization of the CXCR4 signaling in pancreatic cancer cells. Int J Gastrointest Cancer. 2006;37:110–119. | ||
De Falco V, Guarino V, Avilla E, et al. Biological role and potential therapeutic targeting of the chemokine receptor CXCR4 in undifferentiated thyroid cancer. Cancer Res. 2007;67:11821–11829. | ||
Borrello MG, Alberti L, Fischer A, et al. Induction of a proinflammatory program in normal human thyrocytes by the RET/PTC1 oncogene. Proc Natl Acad Sci U S A. 2005;102:14825–14830. | ||
Kajiyama H, Shibata K, Terauchi M, Ino K, Nawa A, Kikkawa F. Involvement of SDF-1alpha/CXCR4 axis in the enhanced peritoneal metastasis of epithelial ovarian carcinoma. Int J Cancer. 2008;122:91–99. | ||
Hirata H, Hinoda Y, Kikuno N, et al. CXCL12 G801A polymorphism is a risk factor for sporadic prostate cancer susceptibility. Clin Cancer Res. 2007;13:5056–5062. | ||
Miki J, Furusato B, Li H, et al. Identification of putative stem cell markers, CD133 and CXCR4, in hTERT-immortalized primary nonmalignant and malignant tumor-derived human prostate epithelial cell lines and in prostate cancer specimens. Cancer Res. 2007;67:3153–3161. | ||
Cavallaro S. CXCR4/CXCL12 in non-small-cell lung cancer metastasis to the brain. Int J Mol Sci. 2013;14:1713–1727. | ||
Gangadhar T, Nandi S, Salgia R. The role of chemokine receptor CXCR4 in lung cancer. Cancer Biol Ther. 2010;9:409–416. | ||
Jones J, Marian D, Weich E, et al. CXCR4 chemokine receptor engagement modifies integrin dependent adhesion of renal carcinoma cells. Exp Cell Res. 2007;313:4051–4065. | ||
Pan J, Mestas J, Burdick MD, et al. Stromal derived factor-1 (SDF-1/CXCL12) and CXCR4 in renal cell carcinoma metastasis. Mol Cancer. 2006;5:56. | ||
Bian XW, Yang SX, Chen JH, et al. Preferential expression of chemokine receptor CXCR4 by highly malignant human gliomas and its association with poor patient survival. Neurosurgery. 2007;61:570–578; discussion 8–9. | ||
Ping YF, Yao XH, Chen JH, et al. The anti-cancer compound Nordy inhibits CXCR4-mediated production of IL-8 and VEGF by malignant human glioma cells. J Neurooncol. 2007;84:21–29. | ||
Murakami T, Maki W, Cardones AR, et al. Expression of CXC chemokine receptor-4 enhances the pulmonary metastatic potential of murine B16 melanoma cells. Cancer Res. 2002;62:7328–7334. | ||
Scala S, Ottaiano A, Ascierto PA, et al. Expression of CXCR4 predicts poor prognosis in patients with malignant melanoma. Clin Cancer Res. 2005;11:1835–1841. | ||
Kalinkovich A, Tavor S, Avigdor A, et al. Functional CXCR4-expressing microparticles and SDF-1 correlate with circulating acute myelogenous leukemia cells. Cancer Res. 2006;66:11013–11020. | ||
Konoplev S, Rassidakis GZ, Estey E, et al. Overexpression of CXCR4 predicts adverse overall and event-free survival in patients with unmutated FLT3 acute myeloid leukemia with normal karyotype. Cancer. 2007;109:1152–1156. | ||
Cronshaw DG, Nie Y, Waite J, Zou YR. An essential role of the cytoplasmic tail of CXCR4 in G-protein signaling and organogenesis. PLoS One. 2010;5:e15397. | ||
Nagasawa T, Kikutani H, Kishimoto T. Molecular cloning and structure of a pre-B-cell growth-stimulating factor. Proc Natl Acad Sci U S A. 1994;91:2305–2309. | ||
Luker KE, Luker GD. Functions of CXCL12 and CXCR4 in breast cancer. Cancer Lett. 2006;238:30–41. | ||
Phillips RJ, Mestas J, Gharaee-Kermani M, et al. Epidermal growth factor and hypoxia-induced expression of CXC chemokine receptor 4 on non-small cell lung cancer cells is regulated by the phosphatidylinositol 3-kinase/PTEN/AKT/mammalian target of rapamycin signaling pathway and activation of hypoxia inducible factor-1α. J Biol Chem. 2005;280:22473–22481. | ||
Schioppa T, Uranchimeg B, Saccani A, et al. Regulation of the chemokine receptor CXCR4 by hypoxia. J Exp Med. 2003;198:1391–1402. | ||
Staller P, Sulitkova J, Lisztwan J, Moch H, Oakeley EJ, Krek W. Chemokine receptor CXCR4 downregulated by von Hippel-Lindau tumour suppressor pVHL. Nature. 2003;425:307–311. | ||
Esencay M, Newcomb EW, Zagzag D. HGF upregulates CXCR4 expression in gliomas via NF-κB: implications for glioma cell migration. J Neurooncol. 2010;99:33–40. | ||
Maroni P, Bendinelli P, Matteucci E, Desiderio MA. HGF induces CXCR4 and CXCL12-mediated tumor invasion through Ets1 and NF-κB. Carcinogenesis. 2007;28:267–279. | ||
Matteucci E, Ridolfi E, Maroni P, Bendinelli P, Desiderio MA. c-Src/histone deacetylase 3 interaction is crucial for hepatocyte growth factor dependent decrease of CXCR4 expression in highly invasive breast tumor cells. Mol Cancer Res. 2007;5:833–845. | ||
Ridolfi E, Matteucci E, Maroni P, Desiderio MA. Inhibitory effect of HGF on invasiveness of aggressive MDA-MB231 breast carcinoma cells, and role of HDACs. Br J Cancer. 2008;99:1623–1634. | ||
Cao Y, Karin M. NF-κB in mammary gland development and breast cancer. J Mammary Gland Biol Neoplasia. 2003;8:215–223. | ||
Helbig G, Christopherson KW 2nd, Bhat-Nakshatri P, et al. NF-κB promotes breast cancer cell migration and metastasis by inducing the expression of the chemokine receptor CXCR4. J Biol Chem. 2003;278:21631–21638. | ||
Kim HC, Choi KC, Choi HK, et al. HDAC3 selectively represses CREB3-mediated transcription and migration of metastatic breast cancer cells. Cell Mol Life Sci. 2010;67:3499–3510. | ||
Uchida D, Onoue T, Begum NM, et al. Vesnarinone downregulates CXCR4 expression via upregulation of Kruppel-like factor 2 in oral cancer cells. Mol Cancer. 2009;8:62. | ||
Begon DY, Delacroix L, Vernimmen D, Jackers P, Winkler R. Yin Yang 1 cooperates with activator protein 2 to stimulate ERBB2 gene expression in mammary cancer cells. J Biol Chem. 2005;280:24428–24434. | ||
Lee BC, Lee TH, Zagozdzon R, Avraham S, Usheva A, Avraham HK. Carboxyl-terminal Src kinase homologous kinase negatively regulates the chemokine receptor CXCR4 through YY1 and impairs CXCR4/CXCL12 (SDF-1α)-mediated breast cancer cell migration. Cancer Res. 2005;65:2840–2845. | ||
Choe Y, Pleasure SJ. Wnt signaling regulates intermediate precursor production in the postnatal dentate gyrus by regulating CXCR4 expression. Dev Neurosci. 2012;34:502–514. | ||
Wang J, Cai J, Han F, et al. Silencing of CXCR4 blocks progression of ovarian cancer and depresses canonical Wnt signaling pathway. Int J Gynecol Cancer. 2011;21:981–987. | ||
Tamura M, Sato MM, Nashimoto M. Regulation of CXCL12 expression by canonical Wnt signaling in bone marrow stromal cells. Int J Biochem Cell Biol. 2011;43:760–767. | ||
Jin Z, Zhao C, Han X, Han Y. Wnt5a promotes Ewing sarcoma cell migration through upregulating CXCR4 expression. BMC Cancer. 2012;12:480. | ||
Bai R, Zhao H, Zhang X, Du S. Characterization of sonic hedgehog inhibition in gastric carcinoma cells. Oncol Lett. 2014;7:1381–1384. | ||
Fareh M, Turchi L, Virolle V, et al. The miR 302-367 cluster drastically affects self-renewal and infiltration properties of glioma-initiating cells through CXCR4 repression and consequent disruption of the SHH-GLI-NANOG network. Cell Death Differ. 2012;19:232–244. | ||
Hinton CV, Avraham S, Avraham HK. Role of the CXCR4/CXCL12 signaling axis in breast cancer metastasis to the brain. Clin Exp Metastasis. 2010;27:97–105. | ||
Peng SB, Peek V, Zhai Y, et al. Akt activation, but not extracellular signal-regulated kinase activation, is required for SDF-1α/CXCR4-mediated migration of epitheloid carcinoma cells. Mol Cancer Res. 2005;3:227–236. | ||
Han Y, He T, Huang DR, Pardo CA, Ransohoff RM. TNF-α mediates SDF-1α-induced NF-κB activation and cytotoxic effects in primary astrocytes. J Clin Invest. 2001;108:425–435. | ||
Vila-Coro AJ, Rodriguez-Frade JM, Martin De Ana A, Moreno-Ortiz MC, Martinez AC, Mellado M. The chemokine SDF-1α triggers CXCR4 receptor dimerization and activates the JAK/STAT pathway. FASEB J. 1999;13:1699–1710. | ||
Soldevila G, Licona I, Salgado A, Ramirez M, Chavez R, Garcia-Zepeda E. Impaired chemokine-induced migration during T-cell development in the absence of Jak 3. Immunology. 2004;112:191–200. | ||
Kukreja P, Abdel-Mageed AB, Mondal D, Liu K, Agrawal KC. Up-regulation of CXCR4 expression in PC-3 cells by stromal-derived factor-1α (CXCL12) increases endothelial adhesion and transendothelial migration: role of MEK/ERK signaling pathway-dependent NF-κB activation. Cancer Res. 2005;65:9891–9898. | ||
Cabioglu N, Summy J, Miller C, et al. CXCL-12/stromal cell-derived factor-1α transactivates HER2-neu in breast cancer cells by a novel pathway involving Src kinase activation. Cancer Res. 2005;65:6493–6497. | ||
Chinni SR, Yamamoto H, Dong Z, Sabbota A, Bonfil RD, Cher ML. CXCL12/CXCR4 transactivates HER2 in lipid rafts of prostate cancer cells and promotes growth of metastatic deposits in bone. Mol Cancer Res. 2008;6:446–457. | ||
Porcile C, Bajetto A, Barbieri F, et al. Stromal cell-derived factor-1α (SDF-1α/CXCL12) stimulates ovarian cancer cell growth through the EGF receptor transactivation. Exp Cell Res. 2005;308:241–253. | ||
Cojoc M, Peitzsch C, Trautmann F, Polishchuk L, Telegeev GD, Dubrovska A. Emerging targets in cancer management: role of the CXCL12/CXCR4 axis. Onco Targets Ther. 2013;6:1347–1361. | ||
Decaillot FM, Kazmi MA, Lin Y, Ray-Saha S, Sakmar TP, Sachdev P. CXCR7/CXCR4 heterodimer constitutively recruits β-arrestin to enhance cell migration. J Biol Chem. 2011;286:32188–32197. | ||
Singh AK, Arya RK, Trivedi AK, et al. Chemokine receptor trio: CXCR3, CXCR4 and CXCR7 crosstalk via CXCL11 and CXCL12. Cytokine Growth Factor Rev. 2013;24:41–49. | ||
Sun X, Cheng G, Hao M, et al. CXCL12/CXCR4/CXCR7 chemokine axis and cancer progression. Cancer Metastasis Rev. 2010;29:709–722. | ||
Cheng ZJ, Zhao J, Sun Y, et al. β-arrestin differentially regulates the chemokine receptor CXCR4-mediated signaling and receptor internalization, and this implicates multiple interaction sites between β-arrestin and CXCR4. J Biol Chem. 2000;275:2479–2485. | ||
Sun Y, Cheng Z, Ma L, Pei G. β-arrestin2 is critically involved in CXCR4-mediated chemotaxis, and this is mediated by its enhancement of p38 MAPK activation. J Biol Chem. 2002;277:49212–49219. | ||
Berkman RA, Merrill MJ, Reinhold WC, et al. Expression of the vascular permeability factor/vascular endothelial growth factor gene in central nervous system neoplasms. J Clin Invest. 1993;91:153–159. | ||
Marchese A, Raiborg C, Santini F, Keen JH, Stenmark H, Benovic JL. The E3 ubiquitin ligase AIP4 mediates ubiquitination and sorting of the G protein-coupled receptor CXCR4. Dev Cell. 2003;5:709–722. | ||
Basu S, Broxmeyer HE. Transforming growth factor-β1 modulates responses of CD34+ cord blood cells to stromal cell-derived factor-1/CXCL12. Blood. 2005;106:485–493. | ||
Li YM, Pan Y, Wei Y, et al. Upregulation of CXCR4 is essential for HER2-mediated tumor metastasis. Cancer Cell. 2004;6:459–469. | ||
Rahimi M, George J, Tang C. EGFR variant-mediated invasion by enhanced CXCR4 expression through transcriptional and post-translational mechanisms. Int J Cancer. 2010;126:1850–1860. | ||
Lapteva N, Yang AG, Sanders DE, Strube RW, Chen SY. CXCR4 knockdown by small interfering RNA abrogates breast tumor growth in vivo. Cancer Gene Ther. 2005;12:84–89. | ||
Smith MC, Luker KE, Garbow JR, et al. CXCR4 regulates growth of both primary and metastatic breast cancer. Cancer Res. 2004;64:8604–8612. | ||
Darash-Yahana M, Pikarsky E, Abramovitch R, et al. Role of high expression levels of CXCR4 in tumor growth, vascularization, and metastasis. FASEB J. 2004;18:1240–1242. | ||
Orimo A, Gupta PB, Sgroi DC, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121:335–348. | ||
Zou W, Machelon V, Coulomb-L’Hermin A, et al. Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat Med. 2001;7:1339–1346. | ||
Boudot A, Kerdivel G, Habauzit D, et al. Differential estrogen-regulation of CXCL12 chemokine receptors, CXCR4 and CXCR7, contributes to the growth effect of estrogens in breast cancer cells. PLoS One. 2011;6:e20898. | ||
Kerdivel G, Boudot A, Pakdel F. Estrogen represses CXCR7 gene expression by inhibiting the recruitment of NFκB transcription factor at the CXCR7 promoter in breast cancer cells. Biochem Biophys Res Commun. 2013;431:729–733. | ||
Boudot A, Kerdivel G, Lecomte S, et al. COUP-TFI modifies CXCL12 and CXCR4 expression by activating EGF signaling and stimulates breast cancer cell migration. BMC Cancer. 2014;14:407. | ||
Chambers AF, Groom AC, MacDonald IC. Dissemination and growth of cancer cells in metastatic sites. Nat Rev Cancer. 2002;2:563–572. | ||
Allinen M, Beroukhim R, Cai L, et al. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell. 2004;6:17–32. | ||
Zhang XH, Wang Q, Gerald W, et al. Latent bone metastasis in breast cancer tied to Src-dependent survival signals. Cancer Cell. 2009;16:67–78. | ||
Rubin JB, Kung AL, Klein RS, et al. A small-molecule antagonist of CXCR4 inhibits intracranial growth of primary brain tumors. Proc Natl Acad Sci U S A. 2003;100:13513–13518. | ||
Kato M, Kitayama J, Kazama S, Nagawa H. Expression pattern of CXC chemokine receptor-4 is correlated with lymph node metastasis in human invasive ductal carcinoma. Breast Cancer Res. 2003;5:R144–R150. | ||
Zhang Z, Ni C, Chen W, et al. Expression of CXCR4 and breast cancer prognosis: a systematic review and meta-analysis. BMC Cancer. 2014;14:49. | ||
Tsutsumi H, Tanaka T, Ohashi N, et al. Therapeutic potential of the chemokine receptor CXCR4 antagonists as multifunctional agents. Biopolymers. 2007;88:279–289. | ||
Tamamura H, Hori A, Kanzaki N, et al. T140 analogs as CXCR4 antagonists identified as anti-metastatic agents in the treatment of breast cancer. FEBS Lett. 2003;550:79–83. | ||
Liang Z, Wu T, Lou H, et al. Inhibition of breast cancer metastasis by selective synthetic polypeptide against CXCR4. Cancer Res. 2004;64:4302–4308. | ||
Zhan W, Liang Z, Zhu A, et al. Discovery of small molecule CXCR4 antagonists. J Med Chem. 2007;50:5655–5664. | ||
Liang Z, Zhan W, Zhu A, et al. Development of a unique small molecule modulator of CXCR4. PLoS One. 2012;7:e34038. | ||
Dorsam RT, Gutkind JS. G-protein-coupled receptors and cancer. Nat Rev Cancer. 2007;7:79–94. | ||
Kobayashi K, Oishi S, Hayashi R, et al. Structure-activity relationship study of a CXC chemokine receptor type 4 antagonist, FC131, using a series of alkene dipeptide isosteres. J Med Chem. 2012;55:2746–2757. | ||
Yoshikawa Y, Kobayashi K, Oishi S, Fujii N, Furuya T. Molecular modeling study of cyclic pentapeptide CXCR4 antagonists: new insight into CXCR4-FC131 interactions. Bioorg Med Chem Lett. 2012;22:2146–2150. | ||
Yang Q, Zhang F, Ding Y, et al. Antitumour activity of the recombination polypeptide GST-NT21MP is mediated by inhibition of CXCR4 pathway in breast cancer. Br J Cancer. 2014;110:1288–1297. | ||
Ling X, Spaeth E, Chen Y, et al. The CXCR4 antagonist AMD3465 regulates oncogenic signaling and invasiveness in vitro and prevents breast cancer growth and metastasis in vivo. PLoS One. 2013;8:e58426. | ||
Hassan S, Buchanan M, Jahan K, et al. CXCR4 peptide antagonist inhibits primary breast tumor growth, metastasis and enhances the efficacy of anti-VEGF treatment or docetaxel in a transgenic mouse model. Int J Cancer. 2011;129:225–232. | ||
Huang EH, Singh B, Cristofanilli M, et al. A CXCR4 antagonist CTCE-9908 inhibits primary tumor growth and metastasis of breast cancer. J Surg Res. 2009;155:231–236. | ||
Singh B, Cook KR, Martin C, et al. Evaluation of a CXCR4 antagonist in a xenograft mouse model of inflammatory breast cancer. Clin Exp Metastasis. 2010;27:233–240. | ||
Bachelder RE, Wendt MA, Mercurio AM. Vascular endothelial growth factor promotes breast carcinoma invasion in an autocrine manner by regulating the chemokine receptor CXCR4. Cancer Res. 2002;62:7203–7206. | ||
Kim B, Park B. Baohuoside I suppresses invasion of cervical and breast cancer cells through the downregulation of CXCR4 chemokine receptor expression. Biochemistry. 2014;53:7562–7569. | ||
Chen XP, Qian LL, Jiang H, Chen JH. Ginsenoside Rg3 inhibits CXCR4 expression and related migrations in a breast cancer cell line. Int J Clin Oncol. 2011;16:519–523. | ||
Park B, Sung B, Yadav VR, Cho SG, Liu M, Aggarwal BB. Acetyl-11-keto-beta-boswellic acid suppresses invasion of pancreatic cancer cells through the downregulation of CXCR4 chemokine receptor expression. Int J Cancer. 2011;129:23–33. | ||
Pandey MK, Sandur SK, Sung B, Sethi G, Kunnumakkara AB, Aggarwal BB. Butein, a tetrahydroxychalcone, inhibits nuclear factor (NF)-κB and NF-κB-regulated gene expression through direct inhibition of IκBα kinase beta on cysteine 179 residue. J Biol Chem. 2007;282:17340–173450. | ||
Pandey MK, Sung B, Ahn KS, Aggarwal BB. Butein suppresses constitutive and inducible signal transducer and activator of transcription (STAT) 3 activation and STAT3-regulated gene products through the induction of a protein tyrosine phosphatase SHP-1. Mol Pharmacol. 2009;75:525–533. | ||
Aizu E, Nakadate T, Yamamoto S, Kato R. Inhibition of 12-O-tetradecanoylphorbol-13-acetate-mediated epidermal ornithine decarboxylase induction and skin tumor promotion by new lipoxygenase inhibitors lacking protein kinase C inhibitory effects. Carcinogenesis. 1986;7:1809–1812. | ||
Selvam C, Jachak SM, Bhutani KK. Cyclooxygenase inhibitory flavonoids from the stem bark of Semecarpus anacardium Linn. Phytother Res. 2004;18:582–584. | ||
Chua AW, Hay HS, Rajendran P, et al. Butein downregulates chemokine receptor CXCR4 expression and function through suppression of NF-κB activation in breast and pancreatic tumor cells. Biocheml Pharmacol. 2010;80:1553–1562. | ||
Cai SH, Tan Y, Ren XD, Li XH, Cai SX, Du J. Loss of C-terminal alpha-helix decreased SDF-1α-mediated signaling and chemotaxis without influencing CXCR4 internalization. Acta Pharmacol Sin. 2004;25:152–160. | ||
Schwarze SR, Dowdy SF. In vivo protein transduction: intracellular delivery of biologically active proteins, compounds and DNA. Trends Pharmacol Sci. 2000;21:45–48. | ||
Watson K, Edwards RJ. HIV-1-trans-activating (Tat) protein: both a target and a tool in therapeutic approaches. Biochem Pharmacol. 1999;58:1521–1528. | ||
Becker-Hapak M, McAllister SS, Dowdy SF. TAT-mediated protein transduction into mammalian cells. Methods. 2001;24:247–256. | ||
Jin LH, Bahn JH, Eum WS, et al. Transduction of human catalase mediated by an HIV-1 TAT protein basic domain and arginine-rich peptides into mammalian cells. Free Radical Biol Med. 2001;31:1509–1519. | ||
Wadia JS, Dowdy SF. Transmembrane delivery of protein and peptide drugs by TAT-mediated transduction in the treatment of cancer. Adv Drug Deliv Rev. 2005;57:579–596. | ||
Jackson MR, Nilsson T, Peterson PA. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990;9:3153–3162. | ||
Munro S, Pelham HR. A C-terminal signal prevents secretion of luminal ER proteins. Cell. 1987;48:899–907. | ||
Furusato B, Mohamed A, Uhlen M, Rhim JS. CXCR4 and cancer. Pathol Int. 2010;60:497–505. | ||
Hirbe AC, Rubin J, Uluckan O, et al. Disruption of CXCR4 enhances osteoclastogenesis and tumor growth in bone. Proc Natl Acad Sci U S A. 2007;104:14062–14067. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.